
The diploporite blastozoan Lepidocalix pulcher from the Middle Ordovician of northern Algeria: Taxonomic revision and palaeoecological implications
YAMOUNA MAKHLOUF, BERTRAND LEFEBVRE, ELISE NARDIN, AHMED NEDJARI, and CHRISTOPHER R.C. PAUL
Makhlouf, Y., Lefebvre, B., Nardin, E., Nedjari, A., and Paul, C.R.C. 2017. The diploporite blastozoan Lepidocalix pulcher from the Middle Ordovician of northern Algeria: Taxonomic revision and palaeoecological implications. Acta Palaeontologica Polonica 62 (2): 299–310.
We present revision of the taxonomy and palaeoecology of the Ordovician aristocystitid Lepidocalix pulcher from the Zaouïa of Stita (Great Kabylia, Algeria). An emended diagnosis is proposed, highlighting the four-fold ambulacral system and the typical thecal plating organised in circlets. Lepidocalix is here assigned to the subfamily Calicinae of the family Aristocystitidae. The latex casts show fitted sutures between plates, slightly abraded spines, and well-preserved oral surface. The thecal plates possess up to three dipores, each, included into the spines. The presence of such covered diplopores would have reduced the respiration rate, by restricting their exchange surface area. The spines covering the dipores are not articulated and they could have a protective role. Lepidocalix is interpreted as stationary epifauna, probably using iceberg strategy to be stabilized into the soft substrate.
Key words: Echinodermata, Diploporita, taxonomy, palaeoecology, Ordovician, Algeria, Stita.
Yamouna Makhlouf [yamouna.makhlouf@gmail.com],Université Frères Mentouri Constantine, FSTGAT, Campus Zouaghi Slimane, Route Aïn el Bey, 25000 Constantine, Algérie-Laboratoire Géodynamique des Bassins Sédimentaires et des Orogènes (LGBSO), Université des Sciences et de la Technologie Houari Boumediene, FSTGAT, BP 32, El Alia, Bab Ezzouar, 16000 Alger, Algeria.
Bertrand Lefebvre [bertrand.lefebvre@univ-lyon1.fr], UMR CNRS 5276 LGLTPE, Université Lyon 1, bâtiment Géode, 2 rue Raphaël Dubois, 69622 Villeurbanne, France.
Elise Nardin [elise.nardin@get.omp.eu], Géosciences Environnement Toulouse, Observatoire Midi-Pyrénées, CNRS, IRD, Université de Toulouse, 14 avenue Edouard Belin, 31400 Toulouse, France.
Ahmed Nedjari [nedjaria@gmail.com], Laboratoire Géodynamique des Bassins Sédimentaires et des Orogènes (LGBSO), Université des Sciences et de la Technologie Houari Boumediene, FSTGAT, BP 32, El Alia, Bab Ezzouar, 16000 Alger, Algeria.
Christopher R.C. Paul [glcrcp@bristol.ac.uk], The Beeches, Rhode Lane, Uplyme, Lyme Regis, Dorset, DT7 3TU, UK.
Received 21 June 2016, accepted 6 March 2017, available online 26 April 2017.
Copyright © 2017 Y. Makhlouf et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Echinoderms represent a major component of Ordovician faunas, largely contributing to the Great Ordovician Biodiversification Event (e.g., Servais et al. 2008). Among them, diploporite and rhombiferan blastozoans, the so-called “cystoids”, constitute emblematic clades of Ordovician benthic communities (Sprinkle and Guensburg 2004; Lefebvre et al. 2013). In mid to high latitude peri-Gondwanan regions of the southern hemisphere, aristocystitid diploporites were one of the dominant groups of Middle to Late Ordovician invertebrates, e.g., in Algeria (Termier and Termier 1950a), Burma (Bather 1906), China (Sun 1948), Czech Republic (Parsley 1990), France (Chauvel 1941, 1977, 1980; Chauvel and Nion 1969), Iran (Lefebvre et al. 2005), Morocco (Chauvel 1966, 1978; Álvaro et al. 2007), Portugal (Couto and Gutiérrez-Marco 1999), and Spain (Chauvel and Meléndez 1986; Gutiérrez-Marco and Baeza 1996; Gutiérrez-Marco and Aceñolaza 1999; Gutiérrez-Marco 2000; Gil Cid and García Rincón 2012).
Ordovician echinoderm faunas have been superficially investigated in Algeria. Termier and Termier (1950a) reported the presence of diploporite blastozoans in the Middle Ordovician of northern Algeria (Great Kabylia). Since this pioneer but incomplete study, no further palaeontological investigation has been conducted on the Ordovician Algerian “cystoids”. The aim of this study is to describe and revise taxonomically the poorly known aristocystitid species Lepidocalix pulcher Termier and Termier, 1950a, from the Middle Ordovician of northern Algeria. Its particular morphology could contribute to improve the discussion about the palaeoecology of the diploporites.
Institutional abbreviations.—MUA, Musée de l’Université d’Alger, Algeria.
Other abbreviations.—BB, basals; CO, circum-oral; COO, circum-orals; F, facetals; ILL, infralaterals; LL, laterals; OO, orals; PO, peri-oral; POO, peri-orals; RR, radials; UEH, Universal Elemental Homology.
Historical background
The presence of Palaeozoic strata in Great Kabylia was first acknowledged by Lambert (1939), based on the discovery of Orthoceras and Tentaculites in limestone beds previously assigned to the Infralias by Ficheur (1888).
On the northern edge of the Kabylian mountains, the occurrence of Ordovician echinoderms was first reported by Barbier et al. (1948), based on material collected in the “schistes X” (Darriwilian) at three distinct outcrops. The main locality is situated near the confluence of Wadi Stita and Wadi Sebaou, on the southern side of a hillock called the Zaouia. This outcrop yielded Protocrinites cf. oviformis Eichwald, 1840, large thecae of Sphaeronites sp., Sinocystis sp., and isolated plates of Sinocystis cf. yunnanensis (Reed 1917). The second fossiliferous site is located about 1500 m southeast of the right bank of Wadi Sebaou. The fauna is similar to that of the first locality, but with a higher abundance of Sphaeronites sp. The last outcrop (Akaoudj village) yielded small individuals of Sphaeronites. All Darriwilian echinoderm assemblages from Kabylia consist only of diploporites.
The presence of a younger (Katian) echinoderm fauna was also reported by Lambert et al. (1948) in calcareous and micaceous shale, on the northern flank of the Djurdjura (Fig. 1). This Late Ordovician echinoderm assemblage is composed exclusively of rhombiferans: Heliocrinites rouvillei von Koenen, 1886, Corylocrinus cf. crassus von Koenen, 1886, Echinosphaerites sp., and Caryocystites sp.
In the Darriwilian schists of the Zaouïa Formation (Fig. 1), the occurrence of two new diploporite “cystoids” characterized by several small and imbricate thecal plates was briefly mentioned by Termier and Termier (1948). The first one was described as an “amphorid”. It shows an oral zone with four brachiole facets covered by ambulacral plates (Termier and Termier 1948). The second one was interpreted as an asteroblastid. It is characterized by a pentameric peristome connected to ladder-shaped ambulacra (Termier and Termier 1948).
The whole Ordovician echinoderm fauna from Kabylia was reinvestigated and described in more detail by Termier and Termier (1950a). The two Darriwilian “cystoids” from Stita were named: Lepidocalix pulchrum (aristocystitid diploporite) and Barbieria stitensis (sphaeronitid diploporite). The associated echinoderm fauna from the same outcrop was also reported. It includes Sphaeronites sp., Sinocystis sp., and undescribed crinoids. The Katian “cystoid” fauna from the Djurdjura was also revised by Termier and Termier (1950a), who identified Caryocrinites cf. elongatus (Jaekel, 1899), Caryocystites sp., Echinosphaerites sp., Heliocrinites rouvillei von Koenen, 1886, and Sphaeronites sp.
Recently, Botquelen et al. (2006) briefly described crinoid columnals (Squameocrinus sp. and Ristnacrinus cf. cirrifer Le Menn in Chauvel and Le Menn, 1973) and several isolated rhombiferan thecal plates from younger deposits (Late Katian) at Oued Isser (on the edge of the Kabylian Massif).
Finally, an Ordovician echinoderm Lagerstätte (of ?latest Darriwilian to Sandbian age) was recently discovered in screes close to Ougarta village in western Algeria during geological field work performed for the Ceramines Agency, providing abundant specimens of the eocrinoid Ascocystites Barrande, 1887 (Makhlouf et al. 2010). Additional field work in the Bou M’Haoud Formation of the Ougarta Range (Foum Ezzeïdya, Kerzaz area, western Algeria) resulted in several fully articulated individuals of the eocrinoid Ascocystites Barrande, 1887. All specimens are preserved as delicate imprints in Upper Ordovician sandstone (Makhlouf et al. 2015). The same formation had previously delivered (Ghienne et al. 2007) indeterminate echinoderm remains in several sections: Foum Ezzeïdya, Sebkhet el Melah, Djebel Bou Kbaissat and Kheneg Tlaïa.
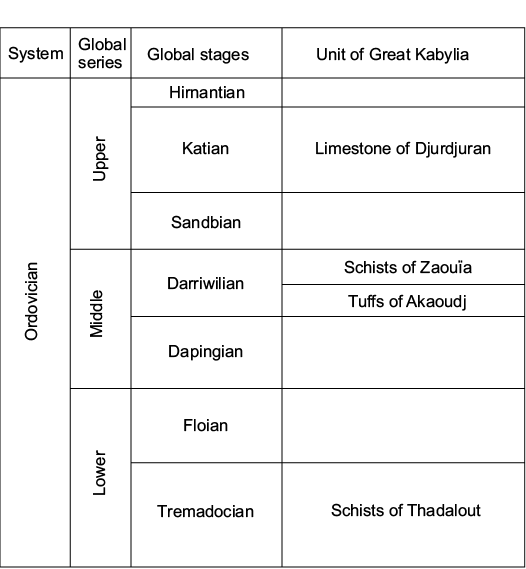
Fig. 1. Lithostratigraphic units defined in Great Kabylia (after Termier and Termier 1950a; Baudelot and Géry 1979).
Geological setting
In Tellian areas, the extent of Palaeozoic outcrops is restricted to a few occurrences in a limited number of regions: Ghar-Rouban Mountains, Tiffrit, Traras, Chenoua, Great and Lesser Kabylia.
Great Kabylia belongs to the Maghrebides, and is a part of the Alpine peri-Mediterranean orogeny (Durand-Delga 1969). This chain is composed of orogenic segments straddling over 2000 km, from the Strait of Gibraltar to the north of Calabria (Italy), and comprising the Moroccan Rif, the Atlas coastline of Algeria (Kabylia and Tell), the Atlas of Tunisia, Sicily, and Calabria (Aubouin 1977; Durand-Delga and Fontboté 1980). It probably results from the convergence and collision of an element of the southern European margin with the northern African margin of the Maghreb Tethys basin (Wildi 1983).
The two historical Darriwilian outcrops yielding echinoderm remains are located in Great Kabylia: Zaouïa of Stita and Akaoudj (Fig. 2). These two localities belong to the Aïssa Mimoun Massif, and they are about 29 km away from the Katian outcrops of northern Djurdjura.
The original type locality of Lepidocalix is located in the Zaouïa of Stita (Fig. 2) in the north-western part of Great Kabylia, at the confluence of Wadi Stita and Wadi Sebaou at about 5 km northeast of Tizi-Ouzou, in the municipality of Makouda.
In addition to the aristocystitid Lepidocalix pulcher Termier and Termier, 1950a, this locality yielded a diverse associated fauna comprising brachiopods (Strophomena barbieri Termier and Termier, 1950a; Leptaena cf. trigonalis Schmidt, 1908, and Strophomena (Actinomena) orta Öpik, 1930), graptolites (Inocaulis? gigas Termier and Termier, 1950a; Airograptus? betieri Termier and Termier, 1950a, Thallograptus barbieri Termier and Termier, 1950a, and Dendrograptus sp.), and trilobites (Pharostoma pulchra [Barrande, 1846], and Colpocoryphe arago [Rouault, 1849]). The Zaouïa of Stita assemblage is preserved in slightly metamorphosed green or grey micaceous mudstone with many intercalated tuff beds similar to those of Akaoudj. The associated acritarch fauna (Baudelot and Géry 1979) confirms the Darriwilian age proposed by Termier and Termier (1950a) based on the macrofauna.
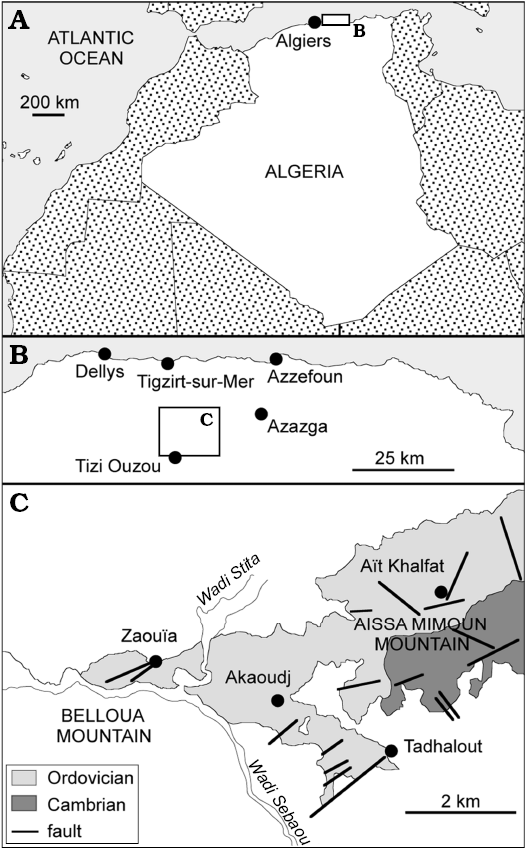
Fig. 2. General map of Algeria (A) and northern Algeria (B) showing location of the fossiliferous sites (solid circles). C. Geological map of Aissa Mimoun Mountain (Great Kabylia) and location of the studied section (Zaouïa of Stita).
Material and methods
All specimens of Lepidocalix studied here were originally briefly described and illustrated (as rough line drawings) by Termier and Termier (1950a). None of them is complete, and all lack ambulacral appendages and are preserved as partial thecae, which is a frequent case in organisms with multi-element skeletons (Brett et al. 1997). Where part and counterpart moulds are available, both external and internal surface of the same part of the theca can be studied (MUA.1024020 and MUA.1023001). The rest of the material only reveals external surface, probably crushed before complete burial. The tectonic distortion of the study material prevented any attempt at providing a detailed reconstruction of all aspects of its original morphology.
The six examined specimens are deposited in the collections of the Museum of Algiers University. Their preservation as internal or external empty moulds made it necessary to prepare latex casts, whitened with ammonium chloride. These casts were observed, drawn and photographed using a Zeiss SteREO Discovery V8 stereomicroscope equipped with a camera lucida and a Zeiss Axio Cam MRc5 digital camera.
Terminology.—Until recently, the very high morphological disparity displayed by blastozoans (“cystoids”) made it particularly difficult to identify thecal plate homologies, and thus to define a common terminology for all classes. The system of plate designation proposed by Jaekel (1899) for glyptocystitoid rhombiferans was further elaborated by Regnéll (1945), Kesling (1963, 1968), and Paul (1967). In this model, the theca can be subdivided into five circlets of plates: BB, ILL, LL, RR, and OO. In glyptocystitoids, the seven oral plates are consistently located in interradial position: in the AB (O4), BC (O5), CD (O1, O6, and O7), DE (O2), and AE (O3) interrays. Thecal plate homologies in diploporite cystoids were first investigated by Paul (1973), who proposed two distinct terminologies for skeletal elements framing the peristome: circum-oral plates in sphaeronitids, and peri-oral plates in holocystitids (Paul and Bockelie 1983). Paul (1973) did not use the same plate designation as in glyptocystitoids, so as to avoid any implication in terms of homologies: for example, the radial position of several COO in sphaeronitids makes it difficult to consider them as equivalent to the interradial OO of glyptocystitoids (Paul 2017).
Recently, the terminology originally defined for the plates framing the peristome in glyptocystitoids was applied to blastoids, coronates, diploporites, eocrinoids, and paracrinoids (Sumrall 2008, 2010, 2015; Sumrall and Waters 2012; Kammer et al. 2013). The generalization of this model, coined UEH by Sumrall (2008), represents a major advance for blastozoan phylogeny. However, the very distinctive plate patterns displayed by some diploporites question the universality of this model (Paul 2017). In holocystitids and protocrinitids, the plate pattern of the oral area is compatible with the UEH (Sumrall 2008, 2010, 2015; Sheffield and Sumrall 2015): all skeletal elements framing the peristome are in interradial position, and the possibility that POO are equivalent to OO cannot be ruled out. In sphaeronitids, only two mouth frame elements are in interradial position (CO6 and CO7, in the CD interray) and can be tentatively compared with topologically equivalent plates in holocystitids (PO1 and F1) and/or other blastozoans (O6 and O7). The other elements forming the mouth frame in sphaeronitids are in radial position (CO1–5) and possibly correspond to enlarged facetals by comparison with other diploporites (Paul 1973, 2017; Paul and Bockelie 1983; but see Sumrall 2015 for a different interpretation).
The identification of the skeletal elements forming the mouth frame in aristocystitid diploporites has been so far largely neglected in all systematic descriptions (e.g., Chauvel 1966, 1977, 1978, 1980; Parsley 1990; Gutiérrez-Marco and Aceñolaza 1999; Gutiérrez-Marco 2000; Gil Cid and Garcia Rincon 2012), as well as in all recent surveys dealing with plate homology in blastozoans (e.g., Paul 1973; Sumrall 2008, 2010, 2015; Sumrall and Waters 2012; Kammer et al. 2013). This situation probably results from the limited number of specimens showing well-preserved oral areas. The peristome of aristocystitids is delimited by a variable number of plates: four to six elements are in interradial position and represent putative homologues of POO in holocystitids and/or OO in other blastozoans; two (Aristocystites), three (Triamara), four (e.g., Calix, Lepidocalix), or five (Oretanocalix) other plates are in radial position. These radial mouth frame elements are very likely equivalent to COO in sphaeronitids and have no equivalent in other blastozoans.
Consequently, the UEH terminology is not followed herein. This model does not provide any clues for the identification of radial mouth frame elements in both aristocystitids and sphaeronitids. In Lepidocalix, only four plates involved in the peristomial margin could be putatively compared with topologically equivalent elements in other blastozoans (O3 and O4 in the BE interray, O1 and O6 in the CD interray). On the other hand, the plate designation proposed by Paul (1971) for mouth frame elements in diploporite cystoids makes it possible to identify all plates in Lepidocalix, and to compare its plate pattern with those of both holocystitids and sphaeronitids. Discussion about putative homologies of interradial mouth frame elements in diploporites and other blastozoans is beyond the scope of this paper, as it would require the detailed reexamination of the oral surfaces of all (most) aristocystitid genera.
Systematic palaeontology
Subphylum Blastozoa Sprinkle, 1973
Class Diploporita Müller, 1854, Paul (1972)
Superfamily Aristocystitoida Neumayr, 1889
Family Aristocystitidae Neumayr, 1889
Subfamily Calicinae Termier and Termier, 1950b
Diagnosis (translated from Chauvel 1966).—Aristocystitids with an elongate to pyrifom theca, composed of numerous plates regularly (juvenile stages) to irregularly organised. Plate surface flattened to convex and bearing central umbo to well-developed spine. Roundish to elliptical peripores including one large diplopore, sometimes covered by a tubercle. Elongate peristome central to a four-fold ambulacral system.
Remarks.—The subfamily Calixinae (sic!) was erected by Chauvel (1966) after the revision of the family Calicidae Termier and Termier, 1950b to emphasize the singularity of this taxonomic group. Kesling (1968) did not acknowledge the different subfamilies (Aristocystitinae, Calicinae, and Hippocystinae) identified by Chauvel (1966) within the family Aristocystitidae. Until a detailed phylogenetic analysis of aristocystitids is performed, Chauvel’s (1966) systematic scheme is followed here, and his original diagnosis of the subfamily Calicinae is here simply translated from French to English. The subfamily Calicinae differs from the two other ones by the shape of the diplopores, the presence of a strong ornamentation, the small size of thecal plates, and the ambulacral pattern.
Genus Lepidocalix Termier and Termier, 1950a
Type species: Lepidocalix pulchrum Termier and Termier, 1950a, monotypic, see below.
Diagnosis.—As for type species by monotypy.
Remarks.—The original description of the species did not contain a diagnosis (see Termier and Termier 1950a). The diagnosis proposed here has been emended from the only previously published one by Kesling (1968; Fig. 3).
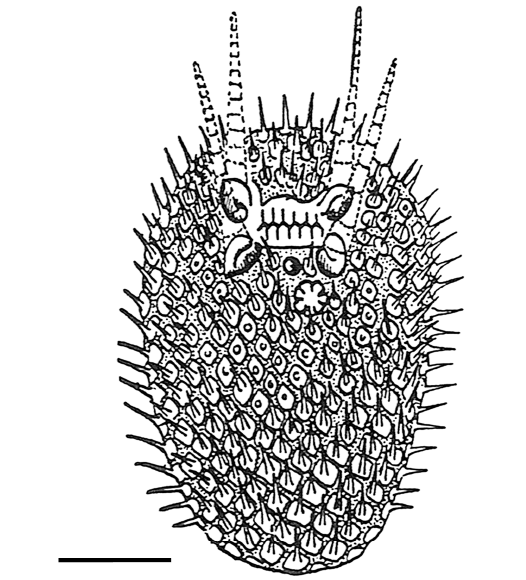
Fig. 3. Original reconstruction of the diploporite blastozoan Lepidocalix pulcher Termier and Termier, 1950a (see Termier and Termier 1959; Kesling 1968). Scale bar 10 mm.
Lepidocalix pulcher Termier and Termier, 1950a
Figs. 4–6.
1950 Lepidocalix pulchrum sp. nov.; Termier and Termier 1950a: 26, pl. 8: 2, 3.
1959 Lepidocalix pulchrum Termier H. & G., 1950; Termier and Termier 1959: 89, 498.
1968 ?Lepidocalix pulchrus [sic] Termier and Termier, 1950a; Kesling 1968: 254, 143.
2011 Calix pulchra (Termier and Termier, 1950a); Gutiérrez-Marco and Colmenar 2011: 190.
Type material: Lectotype (MUA. 1024020), a flattened theca showing the oral zone. Paralectotypes (MUA. 1023001, MUA. 1023002, MUA. 1024016, MUA. 1024021 and MUA. 1024022), by fragmentary samples.
Type locality: Zaouïa of Stita, Great Kabylia, northern Algeria.
Type horizon: Darriwilian, Middle Ordovician.
Emended diagnosis.—Calicinae with an elongate theca composed of abundant, thin, and small thecal plates organised in circlets. Primary plates bear up to three dipores, while secondary and tertiary plates, one tubercular diplopore. Relatively small and elongate peristome connected to two main ambulacral rays subdivided once laterally. Ambulacral rays and peristome covered by two series of polygonal cover plates. Periproct restricted to an anal pyramid composed of triangular plates. Slit-like hydropore located between the peristome and the periproct in the CD interray.

Fig. 4. Diploporite blastozoan Lepidocalix pulcher Termier and Termier, 1950a (MUA. 1023001); Middle Ordovician, Stita, Algeria. A. Photograph of latex cast of external mould showing spines, tubercles, and the appearance of imbricate plates. B. Camera lucida drawing, note three generations of plates. Scale bars 2 mm.
Description.—The theca is composed of small, numerous (hundreds), adjacent plates of various sizes, revealing three generations of plates (Fig. 4). The largest preserved theca reaches 62 mm in diameter (MUA.1023002). Primary plates are pentagonal in shape, sometimes hexagonal with rounded corners. Each primary plate carries one central spine and sometimes one or two tubercles. Secondary and tertiary plates are smaller, hexagonal to irregularly polygonal. They sometimes bear one small tubercle (Fig. 5A2). In larger specimens, primary plates are up to 3 mm wide and 4.5 mm long, while secondary and tertiary elements are up to 2 mm in diameter (MUA. 1023001). All plates are thin at their edges, less than 0.25 mm in thickness. Plates appear to be aligned to form continuous circlets reflecting three generations of circlets. Spines are narrow and elongated (up to 5 mm long). They have a large base, up to the width of the plate, and taper distally, becoming elliptical (no articulation of spines was observed). They are adorally orientated with their base located at the aboral side of the plate. Perpendicular canals can be observed within abraded tubercles and spines. Each secondary and tertiary plate bears one tubercle thinly covering one superficial diplopore. The pattern in the primary plates is more complex. Their spines shelter one internal diplopore, while their tubercles can cover one haplopore or two perpendicular but sinuous canals possibly connected to form a diplopore. Externally the pores can only be seen where the spines have been damaged. Specimens MUA.1024016 and MUA.1023002 confirm the presence of fine spiny ornament on the facetal plates around the ambulacral facets (Fig. 5B, C). The latter reach about 3 × 3 mm, are slightly concave and appear to have supported a single, robust ambulacral appendage, none of which is preserved.
The dipores consist of narrow paired canals about 0.2 mm in diameter (MUA.1023001B). Internally some openings increase in diameter as they reach the internal surface. In the lectotype, pores open externally in an oval tubercle (width 0.65–0.7 mm; length 0.8–0.9 mm) with individual pores about 0.35 mm in diameter. The tubercles apparently represent damaged spines. Specimen MUA.1024016 shows lateral views of some damaged spines that appear to have canals within them (Fig. 5B1). The variability of the pores apparently depends on where the original spines were damaged (MUA.1024022; Fig. 5D).
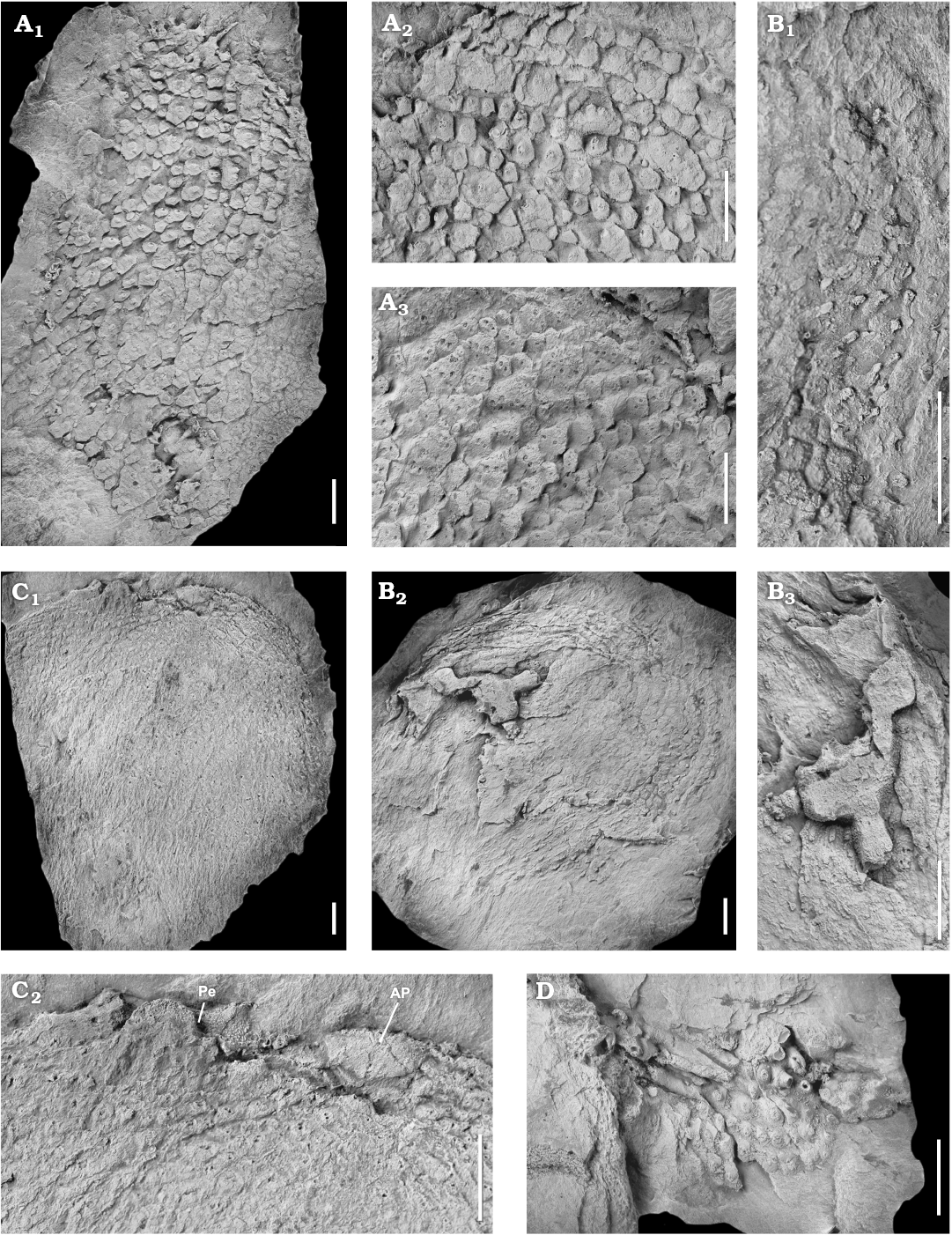
Fig. 5. Photographs of latex casts of the diploporite blastozoan Lepidocalix pulcher Termier and Termier, 1950a; Middle Ordovician, Stita, Algeria. A. MUA. 1023001; general aspect of the theca, in lateral view (A1); enlargement of external surface (A2, turned 90° counter clock-wise), showing the central tubercles on the plates; enlargement of inner surface (A3), showing the distribution of the diplopores and apparent imbrication of plates. B. MUA. 1024016; summit view of the theca (B2), showing circlets of tessellate thecal plates; enlargement of the summit (B3, turned 90° clockwise), composed of four ambulacral rays recumbent on ornamented facetal plates; enlargement of the right thecal periphery (B1), showing tessellate plates ornamented with tubercles and long spines. C. MUA. 1023002; general outline of the theca (C1); enlargement of the top of C1 (C2), showing the organisation of the peristome (Pe) and the periproct (AP). D. MUA. 1024022, showing some abraded spines revealing the diplopores. Scale bars 5 mm.
The oral surface is slightly flattened at the top of the theca (MUA.1023002). Peristome is a wide, transversally elongated slit (5 mm wide), with two short ambulacra diverging from each extremity and ending in large obovate ambulacral facets (Fig. 6A). It is approximately 20 mm wide by 13 mm in the presumed anterior-posterior direction (MUA. 1024020A). Peristome is framed by eight plates (Fig. 6A): four circum-orals in radial position (CO1, CO2, CO4, and CO5) and four peri-orals in interradial position (PO1, PO3, PO4, and PO6). The A ambulacrum is absent, thus leading to a four-fold ambulacral system. Ambulacral facets are deep and large (0.75 mm) recumbent on larger oral plates. Peristome is covered by two series of triangular to polygonal plates, and ambulacra, by one series of polygonal plates. The internal surface shows no trace of a central oral opening (mouth), while the oral plates show many dipores with no clear corresponding openings on the external surface (MUA. 1024020B; Fig. 6B).
The periproct corresponds to a roundish anal pyramid composed of 7 smooth triangular plates (MUA.1023002A). It is about 6 × 3 mm, despite a slight crushing. Hydropore is a slit-like fold located in the CD interray across the PO1–PO6 suture. The gonopore and the aboral region are unknown.
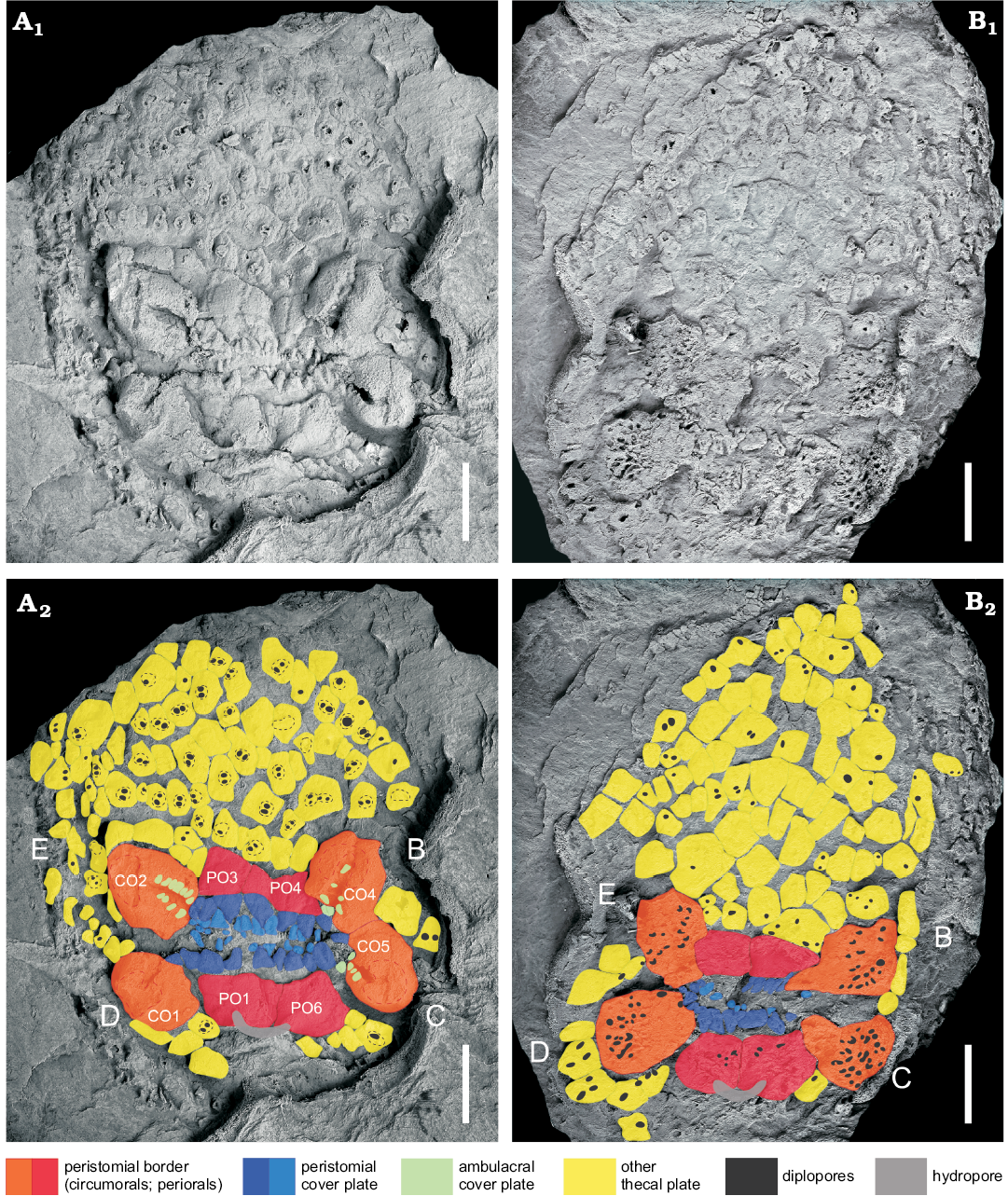
Fig. 6. Oral surface of the diploporite blastozoan Lepidocalix pulcher Termier and Termier, 1950a (MUA. 1024020); Middle Ordovician, Stita, Algeria. A. Four peri-orals (PO1, PO3, PO4, and PO6) between four circum-orals (CO1, CO2, CO4, and CO5), peristome partially covered by small subtriangular oral cover plates biseries, diplopore-bearing polygonal thecal plates immediately surrounding the peristome, hydropore over PO1 and PO6; in external view. B. Surface showing numerous diplopores. Photographs (A1, B1), coloured interpretations (A2, B2) of both surfaces illustrating names using circumorals and periorals (Paul 1973, 2017); in internal view. B–E ambulacral designations in Carpenter’s system. Scale bars 5 mm.
Remarks.—The masculine of the Latin word for beautiful is pulcher not pulchrus. Calix is masculine, so we assume Lepidocalix will be too. Thus the correct form would be Lepidocalix pulcher Termier and Temier, 1950a. The subfamily Calicinae is characterized by a four-fold ambulacral system, as shown in Calix sedgwicki Rouault, 1851; Calix inornatus (Meléndez, 1958); Glaphocystis globulus Chauvel, 1966; Sinocystis loczyi Reed, 1917; Sinocystis gigas (Termier and Termier, 1950a), and Phlyctocystis sp. (Chauvel, 1978). The ambulacral facets of Lepidocalix pulcher appear to have given rise to a single appendage or brachiole each. In contrast, in Calix sedgwicki Rouault, 1851, each of the four ambulacral facets bears four brachioles. The presence of this four-fold ambulacral pattern combined with the possession of the typical thecal plating of Calicinae, both support the assignment of Lepidocalix to this subfamily.
The plates of Lepidocalix are very thin (0.25–0.3 mm thick), whereas most aristocystitids have plates over 1 mm thick (e.g., Calix) and many over 2–3 mm (e.g., Pachycalix). Their plates are very small and only have a maximum of 3 diplopores per plate, while most aristocystitids have several tens (20–30 at least) of diplopores in their largest plates. The plates bear very prominent spines over the diplopores. These spines reach 3 times as long as the plates are wide. Sinocystis has short tubercles over individual diplopores, but these are a fraction of one mm high and the plates are covered with the tubercles. “Spiny” plates in Calix are much thicker, the plates bear numerous diplopores often distributed all over the “spines”.
In general, these aristocystitids are characterized by a more elongate, ellipsoidal theca, composed of generational circlets of plates showing an ornamentation strongly linked to the development of the respiratory structures (Chauvel 1966, 1980; Chauvel and Meléndez 1978; Kesling 1968). The general thecal shape of Lepidocalix is difficult to reconstruct, because of its preservation. The larger specimen (MUA. 1023002) shows a circular outline with a slightly flattened oral region. The specific plating organised in circlets leads us to suppose an original cylindrical shape of the theca (Paul 1971; Frest et al. 2011). It could therefore be similar in thecal shape and plating to Calix and Phlyctocystis, both possessing a conical theca, composed of numerous circlets of three generations of plates (Chauvel 1978). Thecal plating in circlets is a diagnostic characteristic of the subfamily (Chauvel 1966). However, it is more strongly expressed in all Calix species, and only in the aboral regions of the species of Sinocystis and Glaphocystis (Chauvel 1966). Ornamentation as spines and/or tubercles is also known in several species of Calix. However, these ornamentation features are usually restricted to the primary plates in the aboral part of the theca, as in Calix sedgwicki and Calix segaudi Termier and Termier, 1950b. In contrast, the outer surface of Calix inornatus is smooth, and without tubercles. All specimens of Lepidocalix studied here show spines and tubercles distributed all over the theca. A final major singularity of Lepidocalix among its subfamily is in grade, based on the presence of an extremely reduced number of dipores per thecal plate.
Termier and Termier (1950a) described imbricate thecal plating in Lepidocalix. However, new series of latex casts of their original material show tessellate sutures between primary and secondary/tertiary plates and no hollow on the external surface of the plates suggesting any plate overlapping (Fig. 5A). When buried, the spines caused the plates near the edge of the specimens to rotate slightly producing the apparently imbricate appearance of some plates, because these spines were initially at an oblique angle to the sediment surface (Fig. 7). In the centre of the specimen, plates would not have rotated but simply been forced down onto the sediment surface by compaction. Nevertheless, the spines were not completely buried, so that in the centre of specimens they are truncated and often reveal the pore canals inside (similar hypothesis has been proposed for sphaeronitid diploporites; Bockelie 1984). So, the plates are usually still articulated and do not show any apparent imbrication. This appearance is considered here as a preservational artefact, due to the crushing of the theca and the subsequent slight rotation of the spiny plates. Therefore, we reject the hypothesis of imbricate plating in Lepidocalix.
Stratigrapic and geographic range.—Type locality and horizon only.
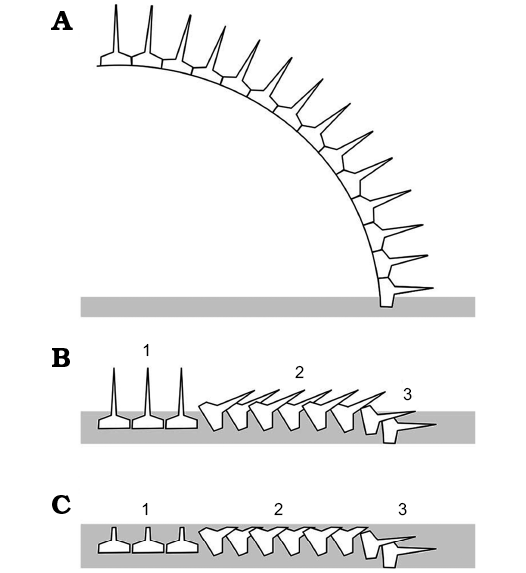 |
Fig. 7. Interpretation of taphonomy of the diploporite blastozoan Lepidocalix pulcher. A. Original disposition of partial theca still articulated. B. Positions of isolated plates after decay of ligaments and possible accumulation of a little more sediment (grey). Three areas can be distinguished. 1, areas where the spines were directed upwards in which the plate orientation has not changed significantly; 2, areas where the plates have rotated slightly so the spines rest on adjacent plates; 3, peripheral areas where the plates have again not rotated significantly, but the spines are buried in the sediment. C. Final stage of preservation in which parts of the plates not buried in sediment have been eroded, leaving plates in areas 1 and 2 lacking complete spines, those in area 2 apparently imbricate and spines only preserved in area 3. |
Discussion
Respiratory structures.—Some thecal plates of Lepidocalix clearly show the presence of diplopores on the internal surface, but no sign of them on the external surface. Indeed, external pores are only seen, where the spines have been damaged. This suggests that the diplopores of Lepidocalix were very likely sealed externally by permanent periporal roof forming the spine, like many representatives of Aristocystitidae but less developed (Bockelie 1984; Parsley 1990). Such covered diplopores are not uncommon in Aristocystites and Sinocystis and probably other aristocystitid genera. Parsley (1990) illustrated covered diplopores in Aristocystites bohemicus Barrande, 1887, and no trace of diplopores can be observed externally in Aristocystites metroi Parsley and Prokop in Parsley, 1990. This would reduce their respiratory capacity by up to about 50% (Paul 1976), by reducing their effective exchange surface area while maintaining their role in the respiratory exchange (Paul 1972). Sealed diplopores of Lepidocalix are within the range of variation seen in other species from the family Aristocystitidae. Bather (1919) illustrated some diplopores of Sinocystis which were at the tips of calcite tubercles and similar to those of Lepidocalix. Bockelie (1984) suggested that the diplopore calcitic sealing could have protected from predation of soft tissue or prevented toxic gas poisoning. However, the latter is unlikely: if the diplopores could still exchange oxygen, they could still exchange other (toxic) gases. The presence of numerous long spines might favour the first hypothesis.
The number of dipores increases with the size of plates. There is evidence that the number of dipores could increase during plate growth. The appearance of new diplopores at the plate sutures could occur without resorption of the plate epistereom in Lepidocalix (Paul 1971, 1973; Bockelie 1984; Parsley 1990), where, nascent formation of dipores has not been observed.
Mode of life.—Aristocystitids appear to have developed three strategies to stabilize the theca on the substrate. Aristocystites Barrande, 1887, has an obvious attachment area developed aborally allowing its settlement on to hard substrates at least early in ontogeny. Aristocystites has a recumbent mode of life (Parsley 1990). It often has very thick plates aborally as in Magrebocystis Chauvel, 1966, which was proposed to live either attached to shell fragments or resting upward into a loose sediment, stabilized by their thick base (Chauvel 1978). In contrast, Calix Rouault, 1851, often has an aboral, stem-like extension to the theca (Chauvel 1966; Gutierrez-Marco and Colmenar 2011). With Phlyctocystis, it has an erect mode of life with aboral structure of attachment (Gil Cid and Domínguez-Alonso 2001). We have seen no evidence of any attachment structures; although the thinness of the thecal plates would conflict with thickened basal plates. Large preserved thecal fragments encompass two-thirds of the reconstructed thecal length. The specimens of Lepidocalix are preserved in a slightly metamorphic mudstone with few complete wide strophomenid shells, in addition to graptolites and leaf-shaped bryozoans. The specimens of Lepidocalix might have lived on a soft substrate among a low-level suspension feeder community, as stationary epifauna possibly with the basal theca partly implanted into the soft sediment, using either an iceberg strategy or as a shallow sediment sticker or rester (Thayer 1975; Chauvel 1941, 1966, 1978; Bottjer and Ausich 1986, Brett 1991; Gil Cid and Domínguez-Alonso 2001; Dornbos et al. 2005).
Conclusion
Lepidocalix pulcher Termier and Termier, 1950a is characterized by a large theca organised in circlets of plates, few diplopores per plate, and a four-fold ambulacral system. The genus is now assigned to the subfamily Calicinae Termier and Termier, 1950b. Our description of the species enables us to propose a diagnosis emended from Kesling (1968). The major singularity is the presence of a reduced number of dipores per thecal plate, with frequently one dipore per plate. Larger specimens show three generations of thecal plates; the primary plates bearing up to three dipores each. Diplopore addition during growth could therefore have occurred at the suture of plates without resorption (Paul 1971, 1973; Bockelie 1984; Parsley 1990). The dipores are covered, as in many representatives of the Aristocystitidae, which would result in a reduction of the respiratory capacity, by decreasing the exchange surface area (Paul 1972). The thecal plates are ornamented by tubercles and spines; the latter hosting the diplopores. Both ornaments constitute the extension of plate epistereom. They might favour the hypothesis of protection from predation on soft tissue (Bockelie 1984). During fossilization, some spine-bearing plates rotated, thus producing their apparently imbricate appearance. The imbricate thecal plating in Lepidocalix described by Termier and Termier (1950a), is therefore considered here as preservational artefact. All specimens of Lepidocalix are preserved in a slightly metamorphic mudstone. Despite the absence of preserved aboral region, we suggest that Lepidocalix probably lived on the substrate as stationary epifauna (Bottjer and Ausich 1986; Brett 1991).
Acknowledgements
We would like to acknowledge the Museum of Algiers University for access to the type specimens of the André Barbier collection. We are particularly grateful to people struggling to preserve the Museum of Algiers University, especially Fettouma Chikhi-Aouimeur, Anissa Dahoumane, Wassila Bourouiba, and Malika Larara (all University of Sciences and Technology Houari Boumediene, Algiers, Algeria), with special thanks to Yasmina Chaid-Saoudi (Algiers University, Algeria) and Daho Djerbal (Director of NAQD revue, Algiers, Algeria). The authors appreciate the comments of the two reviewers, Jim Sprinkle (University of Texas, Austin, USA) and Samuel Zamora (Instituto Geológico y Minero de España, Madrid, Spain), that greatly improved the manuscript.
References
Álvaro, J.J., Vennin, E., Villas, E., Destombes, J., and Viscaïno, D. 2007. Pre-Hirnantian (latest Ordovician) benthic community assemblages: Controls and replacements in a siliciclastic-dominated platform of the eastern Anti-Atlas, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 245: 20–36. Crossref
Aubouin, J. 1977. Méditerranée orientale et Méditerranée occidentale: esquisse d’une comparaison du cadre alpin. Bulletin de la Société Géologique de France 7: 486–495. Crossref
Barbier, A., Termier, H., and Termier, G. 1948. Présence du Llanvirn en Grande Kabylie (Algérie). Comptes Rendus de l’Académie des Sciences, Paris 226: 1385–1386.
Barrande, J. 1846. Notice préliminaire sur le Système Silurien et les trilobites de Bohême. 96 pp. Hirschfeld, Stockholm.
Barrande, J. 1887. Système Silurien du Centre de la Bohême: Volume VII. Classe des Echinodermes, Ordre des Cystidées. 233 pp. Rivnac, Prague.
Bather, F.A. 1906. Ordovican Cystidea from Burma. In: F.R.C. Reed (ed.), The Lower Palaeozoic fossils from the northern Shan States, Burma. Memoirs of the Geological Survey of India, Palaeontologia Indica, New Series 2: 1–154. Crossref
Bather, F.A. 1919. Notes on Yunnan Cystidea. III. Sinocystis compared with similar genera. Geological Magazine 6: 71–77.
Baudelot, S. and Géry, B. 1979. Découverte d’Acritarches du Cambrien supérieur et du Trémadoc dans le Massif ancien de Grande Kabylie (Algérie). Comptes Rendus de l’Académie des Sciences, Paris 288: 1513–1516.
Bockelie, J.F. 1984. The Diploporita of the Oslo region, Norway. Palaeontology 27: 1–68.
Botquelen, A., Le Menn, J., and Loi, A. 2006. Echinodermes de l’Ordovicien supérieur (Ashgill) de Sardaigne et d’Algérie. Geobios 39: 13– 23. Crossref
Bottjer, D.J. and Ausich, W.I. 1986. Phanerozoic development of tiering in soft substrate suspension-feeding communities. Paleobiology 12: 400–420. Crossref
Brett, C.E. 1991. Organism-sediment relationships in Silurian marine environment. Palaeontology 44: 301–344.
Brett, C.E., Moffat, H.A., and Taylor, W.L. 1997. Echinoderm taphonomy, taphofacies, and Lagerstätten. Paleontological Society Papers 3: 147–190.
Chauvel, J. 1941. Recherches sur les Cystoïdes et les Carpoïdes armoricains. Mémoires de la Société Géologique et Minéralogique de Bretagne 5: 1–286.
Chauvel, J. 1966. Echinodermes de l’Ordovicien du Maroc. 117 pp. Editions du CNRS-Cahiers de Paléontologie, Paris.
Chauvel, J. 1977. Calix sedgwicki Rouault (Echinoderme Cystoïde, Ordovicien du Massif armoricain) et l’appareil ambulacraire des Diploporites. Comptes Rendus sommaires de la Société Géologique de France 6: 314–317.
Chauvel, J. 1978. Compléments sur les Echinodermes du Paléozoïque marocain (Diploporites, Eocrinoïdes, Edrioasteroïdes). Note du Service Géologique du Maroc 39: 27–78.
Chauvel, J. 1980. Données nouvelles sur quelques Cystoïdes Diploporites (Echinodermes) du Paléozoïque armoricain. Bulletin de la Société géologique et minéralogique de Bretagne C 12: 1–28.
Chauvel, J. and Le Menn, J. 1973. Echinodermes de l’Ordovicien supérieur de Coat-Carrec, Argol (Finistère). Bulletin de la Société géologique et minéralogique de Bretagne C 4: 39–61.
Chauvel, J. and Meléndez, B. 1978. Les Echinodermes (Cystoïdes, Astérozoaires, Homalozoaires) de l’Ordovicien moyen des Monts de Tolède (Espagne). Estudios geológico 34: 75–87.
Chauvel, J. and Meléndez, B. 1986. Note complémentaire sur les échinodermes ordoviciens de Sierra Morena. Estudios Geológicos 42: 451–459. Crossref
Chauvel, J. and Nion, J. 1969. Sur la présence de Phlyctocystis Chauvel, 1966, échinoderme cystoïde, dans l’Ordovicien de la presqu’île de Crozon (Finistère). Bulletin de la Société Géologique de France 7: 443–446. Crossref
Couto, H.M. and Gutiérrez-Marco, J.C. 1999. Nota sobre algunos Diploporita (Echinodermata) de las pizarras de la Formación Valongo (Ordovícico Medio, Portugal). Temas Geológico-Mineros ITGE, Madrid 26: 541–545.
Dornbos, S.Q., Bottjer, D.J., and Chen, J.-Y. 2005. Paleoecology of benthic metazoans in the Early Cambrian Maotianshan Shale biota and the Middle Cambrian Burgess Shale biota: evidence for the Cambrian substrate revolution. Palaeogeography, Palaeoclimatology, Palaeoecology 220: 47–67. Crossref
Durand-Delga, M. 1969. Mise au point sur la structure du Nord-Est de la Berbérie. Bulletin du Service de la Carte géologique d’Algérie. Nouvelle série, Alger 39: 89–131.
Durand-Delga, M. and Fontboté, J.M. 1980. Le cadre structural de la Méditerranée occidentale. In: J. Auboin, J. Debelmas, and M. Latreille (eds.), Géologie des chaînes alpines issues de la Téthys. Mémoires du Bureau de Recherches Géologiques et Minières 115: 67–85.
Eichwald, E.V. 1840. Sur le système silurien de l’Esthonie. Journal de Médecine et d’Histoire Naturelle de l’Académie de Médecine de Saint-Pétersbourg 1 (2): 1–222.
Ficheur, F. 1888. Esquisse géologique de la chaîne du Djurdjura. Association Française pour l’Avancement des Sciences, Compte-rendu de la 17ème session (Oran, 1888), 248–262. Edition Masson, Paris.
Frest, T.J., Strimple, H.L., and Paul, C.R.C. 2011. The North American Holocystites fauna (Echinodermata: Blastozoa: Diploporita): palaeobiology and systematics. Bulletins of American Paleontology 380: 1–141.
Ghienne, J.-F., Boumendjel, K., Paris, F., Videt, B., Rachebœuf, P., and Ait Salem, H. 2007. The Cambrian–Ordovician succession in the Ougarta Range (western Algeria, North Africa) and interference of the Late Ordovician glaciation on the development of the Lower Paleozoic transgession on northern Gondwana. Bulletin of Geosciences 82: 183– 214. Crossref
Gil Cid, M.D. and Domínguez-Alonso, P.D. 2001. Attachment strategies in Diploporita inhabiting soft-substratum communities. In: F.K. Barker (ed.), Echinoderms 2000. Proceedings of the 10th International Echinoderm Conference, 83–86. A.A. Balkema, Rotterdam.
Gil Cid, M.D. and García Rincón, J.M. 2012. Thecal (oral zone) elements in cystoids from Spain. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 264: 181–190. Crossref
Gutiérrez-Marco, J.C. 2000. Revisión taxonómica de “Echinosphaerites” murchisoni Verneuil y Barrande, 1855 (Echinodermata, Diploporita) del Ordovícico Medio centroibérico (España). Geogaceta 27: 83–86.
Gutiérrez-Marco, J.C. and Aceñolaza, G.F. 1999. Calix inornatus (Meléndez, 1958) (Echinodermata, Diploporita): morfología de la región oral de la teca y revisión bioestratigráfica. Temas Geológico-Mineros ITGE, Madrid 26: 557–565.
Gutiérrez-Marco, J.C. and Baeza, E. 1996. Descubrimiento de Aristocystites metroi Parsley y Prokop, 1990 (Echinodermata, Diploporita) en el Ordovícico medio centroibérico (España). Geogaceta 20: 225–227.
Gutiérrez-Marco, J.C. and Colmenar, J. 2011. Biostratigraphy of the genus Calix (Echinodermata, Diploporita) in the Middle Ordovician of the Southern Central Iberian Zone (Spain). In: J.C. Gutiérrez-Marco, I. Rábano, and D. Garcia-Bellido (eds.), Ordovician of the World. Cuadernos del Museo Geominero 14: 189–197.
Jaekel, O. 1899. Stammesgeschichte der Pelmatozoen. 1, Thecoidea und Cystoidea. 422 pp. Springer, Berlin.
Kammer, T.W., Sumrall, C.D., Zamora, S., Ausich, W.I., and Deline, B. 2013. Oral region homologies in Paleozoic crinoids and other plesiomorphic pentaradial echinoderms. Plos One 8 (11): 1–16. Crossref
Kesling, R.V. 1963. Key for classification of cystoids. Contribution from the Museum of Paleontology, The University of Michigan 18: 101–116.
Kesling, R.V. 1968. Cystoids. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part S. Echinodermata 1, S85–S267. Geological Society of America, Boulder and University of Kansas, Lawrence.
von Koenen, A. 1886. Ueber neue Cystideen aus den Caradoc-Schichten der Gegend von Montpellier. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 2: 246–254.
Lambert, A. 1939. Sur les terrains primaires et infra-liasiques du Djurdjura (Algérie). Comptes-Rendus de l’Académie des Sciences, Paris 209: 414.
Lambert, A., Termier, H., and Termier, G. 1948. Découverte de l’Ordovicien en Algérie atlasienne. Comptes Rendus de l’Académie des Sciences, Paris 226: 824– 825.
Lefebvre, B., Ghobadipour, M., and Nardin, E. 2005. Ordovician echinoderms from the Tabas and Damghan regions, Iran: palaeobiogeographical implications. Bulletin de la Société géologique de France 176: 231–242. Crossref
Lefebvre, B., Sumrall, C.D., Shroat-Lewis, R.A., Reich, M., Webster, G.D., Hunter, A.W., Nardin, E., Rozhnov, S.V., Guensburg, T.E., Touzeau, A., Noailles, F., and Sprinkle, J. 2013. Palaeobiogeography of Ordovician echinoderms. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society of London, Memoirs 38: 173–198.
Makhlouf, Y., Lefebvre, B., Régnault, S., and Hunter, A.W., 2010. First report of an echinoderm Lagerstätte in the Ordovician of the Ougarta Range, western Algeria. In: M. Reich, J. Reitner, V. Roden, and B. Thuy (eds.), Echinoderm Research 2010, 67–68. Universität Verlag Göttingen, Göttingen.
Makhlouf, Y., Nedjari, A., Lefebvre, B., Nardin, E., and Dahoumane, A. 2015. The eocrinoid Ascocystites Barrande (Echinodermata, Blastozoa) in the Upper Ordovician of Kerzaz (Ougarta, western Algeria). Abstracts, The Rise of Animal Life-Promoting Geological Heritage: Challenges and Issues, Marrakesh, 65. Cadi Ayyad University, Marrakesh.
Meléndez, B. 1958. Nuevo Cistideo del Ordoviciense de los Montes de Toledo. Notas y Communicaciones del Instituto Geologico y Minero de España 50: 321–328, 405–406.
Müller, J. 1854. Über den Bau der Echinodermen. Abhandlungen Königlichen Akademie der Wissenschaften, Berlin 1853: 125–220.
Neumayr, M. 1889. Die Stämme der Thierreiches. 1. Wirbellose Thiere. 603 pp. Tempsky, Wien.
Öpik, A.A. 1930. Beiträge zur kenntnis der Kukruse-(C2–C3-) stufe in Eesti: IV. 34 pp. Geoloogia Instituut, Tartu.
Parsley, R.L. 1990. Aristocystites, a recumbent diploporid (Echinodermata) from the Middle and Late Ordovician of Bohemia, ČSSR. Journal of Paleontology 64: 278–293. Crossref
Paul, C.R.C. 1967. The British Silurian cystoids. Bulletin of the British Museum (Natural History) Geology 13: 299–355.
Paul, C.R.C. 1971. Revision of the Holocystites fauna (Diploporita) of North America. Fieldiana: Geology 24: 1–166.
Paul, C.R.C. 1972. Morphology and function of exothecal pore-structures in cystoids. Palaeontology 15: 1–28.
Paul, C.R.C. 1973. British Ordovician cystoids. Palaeontographical Society Monographs, London: 1–64.
Paul, C.R.C. 1976. Respiration rates in primitive (fossil) echinoderms. Thalassia Jugoslavica 12: 277–286.
Paul, C.R.C. 2017. Testing for homologies in the axial skeleton of primitive echinoderms. Journal of Paleontology [published online]. Crossref
Paul, C.R.C. and Bockelie, J.F. 1983.Evolution and functional morphology of the cystoid Sphaeronites in Britain and Scandinavia. Palaeontology 26: 687–734.
Reed, F.R.C. 1917. Ordovician and Silurian fossils from Yunnan. Memoirs of the Geological Survey of India 6: 1–69.
Regnéll, G. 1945. Non-crinoid Pelmatozoa from the Paleozoic of Sweden. A taxonomic study. Meddelanden Lunds geologisk-mineralogisk Institution 108: 1–225.
Rouault, M. 1849. Note sur de nouvelles espèces de fossiles découvertes en Bretagne. Bulletin de la Société Géologique de France 2: 67–83.
Rouault, M. 1951. Mémoire sur le terrain paléozoïque des environs de Rennes. Bulletin de la Société géologique de France 2 (8): 358–399.
Schmidt, F. 1908. Beitrag zur Kenntniss der ostbaltischen, vorzüglich untersilurischen, Brachiopoden der Gattungen Plectambonites Pand., Leptaena Dalm. und Strophomena Blainv. Bulletin de l’Académie Impériale des Sciences de St.-Pétersbourg 2 (9): 717–726.
Servais, T., Lehnert, O., Li, J., Mullins, G.L., Munnecke, A., Nützel, A., and Vecoli, M. 2008. The Ordovician biodiversification: Revolution in the oceanic trophic chain. Lethaia 41: 99–109. Crossref
Sheffield, S.L. and Sumrall, C.D. 2015. A new interpretation of the oral plating patterns of the Holocystites fauna (Diploporita, Echinodermata). In: S. Zamora and I. Rábano (eds.), Progress in Echinoderm Palaeobiology. Cuadernos del Museo Geominero 19: 159–162.
Sprinkle, J. 1973. Morphology and Evolution of Blastozoan Echinoderms. 283 pp. Museum of Comparative Zoology Harvard University, Cambridge.
Sprinkle, J. and Guensburg, T.E. 2004. Crinozoan, blastozoan, echinozoan, asterozoan, and homalozoan echinoderms. In: B.D. Webby, F. Paris, M.L. Droserand, and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, 266–280. Columbia University Press, New York. Crossref
Sumrall, C.D. 2008. The origin of Lovén’s Law in glyptocystitoid rhombiferans and its bearing on the plate homology and the heterochronic evolution of the hemicosmitid peristomal border. In: W.I. Ausich and G.D. Webster (eds.), Echinoderm Paleobiology, 228–241. University of Indiana Press, Bloomington.
Sumrall, C.D. 2010. A model for elemental homology for the peristome and ambulacra in blastozoan echinoderms. In: L.G. Harris, S.A. Böttger, C.W. Walker, and M.P. Lesser (eds.), Echinoderms: Durham, 269–276. CRC Press, London.
Sumrall, C.D. 2015. Understanding the oral area of derived stemmed echinoderms. In: S. Zamora and I. Rábano (eds.), Progress in Echinoderm Palaeobiology. Cuadernos del Museo Geominero 19: 169–173.
Sumrall, C.D. and Waters, J.A. 2012. Universal elemental homology in Glyptocystitoids, Hemicosmitoids, Coronoids and Blastoids: Steps towards echinoderm phylogenetic reconstruction in derived Blastozoa. Journal of Paleontology 86: 956–972. Crossref
Sun, Y.C. 1948. The early occurrence of some Ordovician and Silurian cystoids from western Yunnan and its significance. Palaeontological Novitates 1: 1–9.
Termier, H. and Termier, G. 1948. Suggestions à propos de deux cystoïdes algériens. Comptes Rendus de l’Académie des Sciences, Paris 226: 1915–1917.
Termier, H. and Termier, G. 1950a. Contribution à l’étude des faunes paléozoïques de l’Algérie. Bulletin du Service de la Carte Géologique de l’Algérie:1–83.
Termier, H. and Termier, G. 1950b. Paléontologie Marocaine. 2 (4). Invertébrés de l’Ère Primaire. Annélides, arthropodes, échinodermes, conularides et graptolithes. Notes et Mémoires du Service Géologique du Maroc 79: 1–279.
Termier, H. and Termier, G. 1959. Paléontologie stratigraphique. Premier fascicule. 201 pp. Masson, Paris.
Thayer, C.W. 1975. Morphologic adaptations of benthic invertebrates to soft substrata. Journal of Marine Research 33: 177–189.
Wildi, W. 1983. La chaîne tello-rifaine (Algérie, Maroc, Tunisie): Structure, stratigraphie et évolution du Trias au Miocène. Revue de Géologie Dynamique et Géographie physique 24: 201–297.
Acta Palaeontol. Pol. 62 (2):
299–310, 2017
https://doi.org/10.4202/app.00286.2016