
First record of sabellid and serpulid polychaetes from the Permian of Sicily
ROSSANA SANFILIPPO, ANTONIETTA ROSSO, AGATINO REITANO, and GIANNI INSACCO
Sanfilippo, R., Rosso, A., Reitano, A., and Insacco, G. 2017. First record of sabellid and serpulid polychaetes from the Permian of Sicily. Acta Palaeontologica Polonica 62 (1): 25–38.
Some tubular fossils attributable to sabellids and serpulids, and a serpulid operculum are described for the first time from the Wordian to upper Permian “Pietra di Salomone”, “Rupe di San Calogero”, and “Rupe di San Benedetto” limestones (Sosio Valley, western Sicily, Italy). Tubes are attached to skeletons of sponges, crinoids, brachiopods and stromatoporoids. The material consists of about twenty incomplete tubes (attached and free portions) and includes some small tubes, ca. 0.2 mm wide, belonging to the sabellid Glomerula (with the species G. testatrix comb. nov. and G. gemmellaroi sp. nov.), the serpulid Filograna sp., and large-sized tubes, up to 3 mm wide, belonging to three newly described serpulids: Propomatoceros permianus sp. nov., “Serpula” distefanoi sp. nov., and Serpulidae sp. indet. They are triangular or circular in cross-sections, and display growth lines and/or ornamentations (keels or ribs) on their outer surfaces. Furthermore, an opercular cup referable to those assigned to the genus Pyrgopolon, is also described under the name P. gaiae sp. nov. All specimens show large secondary calcite crystals in their tubes, the original structure being biased by diagenesis. Material represents the first evidence of genuine calcareous tube-dwelling polychaetes from the Palaeozoic, and possibly so far unknown ancestral representatives of the families Sabellidae and Serpulidae. The tubeworm association together with the invertebrates upon which they settled, presumably lived in an exposed shallow-water palaeoenvironment, at the outer edge of a carbonate platform.
Key words: Polychaeta, Sabellidae, Serpulidae, operculum, tube, Palaeozoic, Sicily.
Rossana Sanfilippo [sanfiros@unict.it] and Antonietta Rosso [rosso@unict.it], Department of Biological, Geological and Environmental Sciences, University of Catania, Corso Italia 57, 95129 Catania, Italy.
Agatino Reitano [tinohawk@yahoo.it] and Gianni Insacco [g.insacco@comune.comiso.rg.it], Museo di Storia Naturale di Comiso, Via degli Studi 9, 97013 Comiso, Italy.
Received 28 June 2016, accepted 7 September 2016, available online 30 September 2016.
Copyright © 2017 R. Sanfilippo et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Sabellidae Latreille, 1825, with Glomerula Brünnich Nielsen, 1931 as the sole genus of this family possessing a calcareous tube, and Serpulidae Rafinesque, 1815 are polychaetes with biomineralized skeletons and represent the most important encrusting worms in the Mesozoic, Cenozoic, and today (Hove and van den Hurk 1993; Ippolitov et al. 2014). Only a minor share of all known fossil and extant serpulid species possess calcified opercula. Hitherto, serpulid opercula were known only from the Late Cretaceous onward (e.g., Wrigler 1951; Lommerzheim 1979; Cupedo 1980a, b; Radwańska 1994), and it is not surprising that the fossil record of opercula is much poorer than that of tubes, given their small sizes and the low preservation potential of non-calcified opercula (Gatto and Radwańska 2000).
From the current literature, first appearance of calcareous sabellids (e.g., Glomerula) was during the Late Carboniferous (late Palaeozoic) or Early Jurassic (Parsch 1956; Vinn et al. 2008a, b; Ippolitov et al. 2014). Then they flourished during the Mesozoic times (Jäger 2005) and decreased in abundance during the Cenozoic. Calcareous sabellids are known from tropical and subtropical seas (Perkins 1991; Hove and van den Hurk 1993; Fischer et al. 2000).
The earliest unequivocal representatives of serpulids appeared in the Middle Triassic (see Ippolitov et al. 2014 for a review) and remained rare for the entire Triassic, in most species maintaining minute tube diameters, smooth tube surfaces, and circular cross sections (Vinn and Mutvei 2009). They presumably lived in sheltered shallow platform settings of marginal areas of the Tethys Ocean, associated with fine-grained sediments (Flügel et al. 1984; Braga and López-López 1989; Hove and van den Hurk 1993; Berra and Jadoul 1996; Cirilli et al. 1999; Stiller 2000). The diversity of serpulids rapidly increased since the Early Jurassic up to the Late Jurassic and Cretaceous, when they became a common component of marine invertebrate communities, also colonizing reefs and shallow water exposed setting in temperate and tropical areas all over the world (Parsch 1961; Regenhardt 1961, 1964; Lommerzheim 1979; Jäger 1983, 2005; Palma and Angeleri 1992; Hove and van den Hurk 1993, Kiessling et al. 2006; Vinn et al. 2012), displaying larger and variously shaped tubes. They became even more common worldwide during the Cenozoic (e.g., Rovereto 1899; 1904; Schmidt 1950, 1951, 1955; Radwańska 1994; Di Geronimo et al. 1997b) until today, colonizing a wide range of marine environments, from shallow waters to bathyal (Sanfilippo 1998; Sanfilippo et al. 2013; D’Onghia et al. 2015), as well as abyssal (Zibrowius 1973; Hove and Zibrowius 1986; Kupriyanova 1993; Sanfilippo 2001; Kupriyanova et al. 2011) and hadal (Kupriyanova et al. 2014) zones. They also live in submarine semi-dark and dark cave habitats (Zibrowius 1968; Di Geronimo et al. 1997a; Sanfilippo and Móllica 2000; Sanfilippo et al. 2014) as well as in cold seeps (Vinn et al. 2012). A few species were able to access brackish environments (Fauvel 1923; Hove and Weerdenburg 1978; Aliani et al. 1995) and a single species to thrive in fresh waters of continental karstic caves (Absolon and Hrabe 1930; Bianchi and Sanfilippo 2003; Kupriyanova et al. 2009).
Mentions of putative calcified tube-dwelling organisms go back into the Palaeozoic. A true serpulid is possibly Palaeotubus sosiensis Sanfilippo, Reitano, Insacco, and Rosso, 2016 recently described from the Permian “Pietra di Salomone” Limestone. Further tubeworms, Late Carboniferous to late Permian in age, which show minute coiled tubes (Etheridge 1892; Stuckenberg 1905; Hoare et al. 2002), are attributable to the sabellid Glomerula. Late Permian attached coiled tubes, namely some specimens figured as Serpula pusilla Geinitz, 1848, Vermilia obscura King, 1850, and possibly also Serpulites from Australia, have been claimed as potential serpulids (Guppy et al. 1951). Nevertheless, their minute sizes and unclear morphologies (as deducible from old descriptions and figures) seem to preclude any relationship with veritable serpulids. Indeed, this species has been now moved to the Foraminifera and attributed to the genus Agathammina (Nestell and Nestell 2006: pl. 3: 1–5) because morphologies first misinterpreted as sections of a small coiled serpulid tube actually correspond to chambers of a foraminifer. A comparable misinterpretation would involve Vermilia obscura King, 1850.
As for all Palaeozoic spirorbids later attributed to microconchids (Weedon 1994; Taylor and Vinn 2006; Wilson et al. 2011; Zatoń et al. 2014), most Palaeozoic tubicolous problematica (see Ippolitov et al. 2014 for a review) have been reassigned to trypanoporids, cornulitids, tentaculitids, hederelloids or microconchids (Hove 1994; Vinn and Mutvei 2009) based on their skeletal microstructure.
In the present paper, genuine calcareous tube-dwelling serpulid polychaetes, associated with sabellids, are firstly reported from Permian sediments of the Sosio Valley, western Sicily (Italy). Species described represent the earliest evidence of representatives of this group to which a further calcified tubeworm (P. sosiensis) found in the same rock (Sanfilippo et al. 2016) could seemingly add.
This discovery sheds light on the origin of both taxonomic groups. The present work aims to: (i) describe the morphology of these new sabellids and serpulids; (ii) discuss the status of some early fossil records of both groups; (iii) elucidate the importance of the present finding to reconstruct the early history of these polychaete taxa; and (iv) give information on the habitats they colonized.
Institutional abbreviations.—MSNC, Museo Civico di Storia Naturale di Comiso, Ragusa, Italy.
Material and methods
Specimens examined in the present paper originate from the Permian limestone of the Sosio Valley, cropping out south of the village of Palazzo Adriano in the Sicani Mountains, western Sicily (Fig. 1A, B). Permian outcrops from this area became famous worldwide for the exceptional abundance and diversity of fossils since the discovery by Gemmellaro (1887–1889) who described several invertebrates from this locality in the late 19th century. Today, more than 500 invertebrate species have been described including foraminifera, sponges, corals, brachiopods, molluscs, bryozoans, trilobites, crustaceans, echinoderms, and other fossils of uncertain taxonomic assignment (see Flügel et al. 1991 for a general review; also Ernst 2000; Jenny-Deshusses et al. 2000; Jones et al. 2015). As a whole, the benthic association testifies to a reef habitat (Senowbari and Di Stefano 1988; Flügel et al. 1991) whose marginal sectors partly collapsed as debris flow and turbidite sediments in neighbouring shallow shelf to open sea settings (see Di Stefano and Gullo 1996; Di Stefano et al. 2013). The age of these limestone blocks is still debated. Their formation has been referred to the Wordian (Murghabian) by Flügel et al. (1991), but considered at least partly Capitanian (Midian) or Wuchiapingian (Dzhulfian), and even Changhsingian (Dorashamanian) by Jenny-Deshusses et al. (2000).
The specimens described in this paper were collected from three limestone megablocks exposed along the Torrente San Calogero valley (Fig. 1): the “Pietra di Salomone” upper part, the “Rupe di San Calogero”, and the “Rupe di San Benedetto”. The “Pietra di Salomone” megablock, which delivered most of the studied specimens, consists of nearly 70 m thick poorly defined beds of coarse calcareous breccia and is visible for about 200 m in length, 100 m in width and 30 m in height at the nucleus of an asymmetric anticline (Flügel et al. 1984). The “Rupe di San Calogero” and Rupe di San Benedetto” megablocks are smaller and less fossiliferous than the “Pietra di Salomone” one but stratigraphically correspond to it and were possibly deposited in comparable palaeoenvironments.
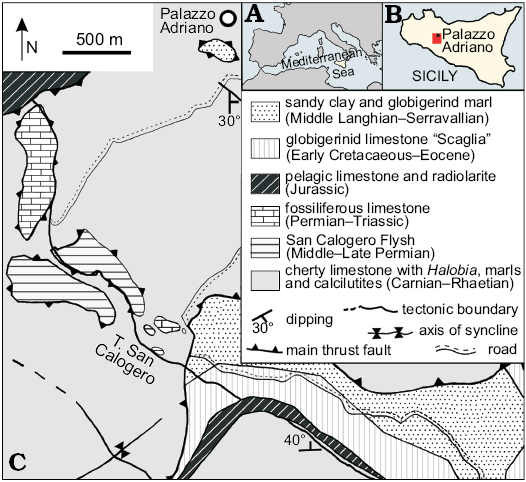 |
Fig. 1. A. Location of the study area in the Mediterranean and Sicily. B. Geological map of the Palazzo Adriano area (Sicani Mountains) showing the location of the fossiliferous limestone blocks. Holocene deposits not shown. Material was collected in blocks cropping out in areas indicated as “fossiliferous limestone”. |
Sampling was performed by one of us (AR) during a field survey within the Riserva Naturale Orientata Monti di Palazzo Adriano e Valle del Sosio, as part of the activities of the Natural History Museum of Comiso.
The Permian limestone is very rich in fossils among which are the crinoids Palermocrinus jaekeli Gislén, 1924 and Heterobrachiocrinus fabianii Yakovlev, 1938, the gastropods Bellerophon sosiensis Gemmellaro, 1889 and Plocostoma piazzii (Gemmellaro, 1889) and the brachiopods Coscinaria communis (Gemmellaro, 1894) and Martinia sp. Almost all examined tubes were found attached to biogenic substrata, mostly sponges but also crinoids, stromatoporoids and brachiopods.
All tubes were accurately cleaned to examine their external surfaces, the free portions separated mechanically from the embedding calcilutite. Low magnification photos were acquired with a Zeiss Discovery V8A stereomicroscope equipped with an Axiovision acquisition system, in order to document the general tube morphologies. Specimens were also examined uncoated under a Tescan Vega 2 LMU Scanning Electron Microscope in Low Vacuum modality to investigate tube micromorphology. All images were acquired at the Department of Biological, Geological and Environmental Sciences (University of Catania).
Systematic palaeontology
Class Polychaeta Grübe, 1850
Subclass Sedentaria Lamarck, 1818
Infraclass Canalipalpata Rouse and Fauchald, 1997
Order Sabellida Fauchald, 1977
Family Sabellidae Latreille, 1825
Subfamily Sabellinae Chamberlin, 1919
Genus Glomerula Brünnich Nielsen, 1931
Type species: Serpulites gordialis von Schlotheim, 1820, Middle or Late Oxfordian (161–158 Ma) of Mordovia region, Russia.
Emended diagnosis.—Tube calcareous, solitary or pseudocolonial, straight to variably coiled or meandering, typically disposed as knots, spirals and tangles; an erect portion may be present. Tube occasionally showing crescentic cavities within the tube wall, and specimens from the Cretaceous and Cenozoic may occasionally show trilobate narrowings of the lumen. Tubes smooth, without ornamentation, peristomes and basal seam (compiled from Regenhardt 1961; Jäger 2005; Ippolitov 2007a).
Stratigraphic and geographic range.—Upper Permian of Australia; Wordian to upper Permian of Italy; Mesozoic and Cenozoic of Europe, Asia, South America; Recent (Lizard Island, Great Barrier Reef, Australia).
Glomerula testatrix (Etheridge, 1892) comb. nov.
Fig. 2A–C.
1892 Serpula testatrix sp. nov.; Etheridge 1892: 120, pl. 18: 4, 5.
Material.—Two glomerate coiling tubes on sponges (MSNC 4515, MSNC 4516) and one incomplete glomerate tube on a crinoid stalk (MSNC 4517). One aggregate of tubes nearly aligned and irregularly coiled, attached on a sponge (MSNC 4518). All specimens come from the “Pietra di Salomone” megablock , the Sosio Valley, western Sicily (Italy); Wordian to upper Permian.
Emended diagnosis.—Tube long in respect to its width, circular in cross-section and smooth, increasing in diameter slowly. Base of the tube not widened by a basal cementing flange. Tubes more or less irregularly curved, often forming irregular spirals, intermingled meanders or chaotic knots.
Description.—Tubes are small-sized (outer diameter of 0.2–0.25 mm) and weakly attached to the substrate, their outer surface being smooth, only marked by feeble growth lines at places. Tubes form intermingled overlaid whorls growing on previous tube portions. The original tube wall structure is biased by recrystallization showing large diagenetic crystals which locally expand inwards, also infilling the tube lumen.
Remarks.—This species was originally described as a serpulid under the name Serpula testatrix by Etheridge (1892), based on late Permian specimens from Australia. No further specimen has been found since then and no modern images or descriptions are available. Nevertheless, Etheridge’s description and figures (1892: pl. 18: 4, 5) of the holotype are sufficiently diagnostic and unequivocally indicate that the species lacks serpulid characters. In contrast, its small-sized and typically glomerate coiled tube fits well in the genus Glomerula (Fig. 2A), as recently suspected by Ippolitov et al. (2014). Jäger (1983, 2005) considered Omasaria Regenhardt, 1961 as a synonym of Glomerula, mainly on the basis of its typical irregularly interweaving tubes. Such tubes seem to have a very wide stratigraphic distribution. Following Ippolitov et al. (2014), the oldest Palaeozoic representatives likely attributable to Glomerula could be those reported as “Serpula” spp. by Stuckenberg (1905) and as “tubeworms” by Hoare et al. (2002: fig. 1.1–1.7) from the Late Carboniferous. Indeed, these specimens show slowly widening tubes and a characteristic glomerate coiling.
Owing to their morphological characters, specimens found in the “Pietra di Salomone” megablock can be identified as G. testatrix. Furthermore, Australian and Sicilian specimens share the same substratum, and possibly the same habitat, because pl 18: 5 in Etheridge (1892) depicts the species encrusting crinoids. Interestingly, the stratigraphic distribution of the new G. testatrix findings is consistent with the age of the holotype material.
Like other Palaeozoic congeners, G. testatrix, with a tube diameter of only 0.25 mm, is minute in respect to Mesozoic species of Glomerula, which possess quite large tubes, from slightly less than 1 mm (G. lombricus Defrance, 1827) to ca. 4 mm in diameter at maximum (e.g., Jäger 1983). Also the only known Recent species G. piloseta (Perkins, 1991) is larger, having a diameter of about 0.5 mm.
The presence of trilobate lumina has been reported from some Cretaceous species assigned to Glomerula and used as a diagnostic feature to distinguish them from some more ancient, Jurassic congeners (Jäger 1983, 1993, 2005). This character is also absent in G. testatrix and in other old species, as well as in Cenozoic to Modern Glomerula representatives. Consequently, it seems to be restricted only to a few Cretaceous species. We suggest here to place such species in a separate genus, pending examination of the type materials.
Stratigraphic and geographic range.—Upper Permian of Australia; Wordian to upper Permian of Italy.
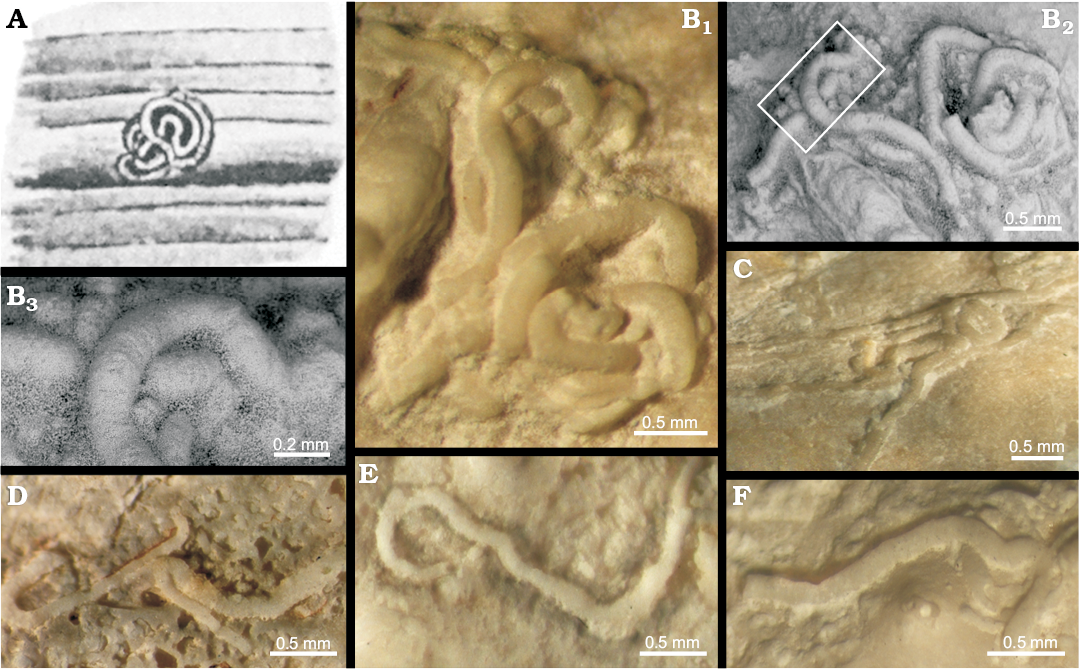
Fig. 2. Sabellids from literature (A) and from study material (B–F). A. Holotype of Serpula testatrix Etheridge, 1892 (pl. 18: 5) on a crinoid stem from East Maitland (South Wales), Permo-Carboniferous, showing typical loose loops and spirals. B, C. Glomerula testatrix (Etheridge, 1892) comb. nov., from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian. B. MSNC 4515, irregular glomerate interweaving tubes; photograph (B1), SEM image (B2); B3, detail of B2, showing the tube outer surface with feeble growth lines. C. MSNC 4518, set of sub-parallel tubes slightly meandering and locally forming a sharp bend encrusting a sponge. D–F. Glomerula gemmellaroi sp. nov. from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian. D. MSNC 4519, holotype, specimen forming meanders and bending at angles of 90° and 180°. E. MSNC 4520, tube initially loop-like curved and then bent at right angle. F. MSNC 4521, meandering tube on a sponge.
Glomerula gemmellaroi sp. nov.
Fig. 2D–F.
Etymology: Named after Professor Gaetano Giorgio Gemmellaro (1832–1904), who discovered and first studied the Permian fossiliferous limestone rocks of the Sosio Valley.
Type material: Holotype: single tube curved at different angles attached for all of its length on a sponge (MSNC4519). Paratypes: single slightly curved incomplete tubes: two on sponges (MSNC 4520, MSNC 4521), one on a stromatoporoid (MSNC 4522), and one on a brachiopod shell (MSNC 4523). All specimens come from the “Pietra di Salomone” megablock, except for specimen MSNC 4522 originating from the “Rupe di San Calogero” megablock.
Type locality: Sosio Valley, western Sicily (Italy).
Type horizon: “Pietra di Salomone” Limestone, Wordian to upper Permian.
Diagnosis.—Tube long in respect to its width, circular in cross-section and smooth, increasing in diameter slowly. Base of the tube not widened by a basal cementing flange. Tubes more or less irregularly curved, but never forming spirals or knots.
Description.—Tubes small-sized (outer diameter of 0.2–0.25 mm), the outer surface smooth, only at places marked by feeble growth lines. Tubes are weakly attached to the substrate for all their length, isolated and irregularly curved, sometimes forming close meanders angled more typically at 90° and 180°. Aggregates of aligned to irregularly coiled tubes also occur. An obvious recrystallization affects the specimens, their original tube wall structure being totally biased.
Remarks.—The present species has been assigned to Glomerula because of its small-sized smooth tubes, circular in cross-section and slowly increasing in diameter, and the general development included in the generic diagnosis. The particular meandering and the bending angles clearly differentiate these tubes from those of G. testatrix in the studied collection, and from other species in the genus.
Stratigraphic and geographic range.—Wordian to upper Permian of Italy.
Family Serpulidae Rafinesque, 1815
Genus Filograna Berkeley, 1835
Type species: Serpula filograna Linnaeus, 1767; Recent of Europe.
Filograna sp.
Fig. 3.
Material.—Two loose aggregates of few sub-parallel tubes, encrusting sponges (MSNC 4524, MSNC 4525). All specimens come from “Pietra di Salomone” Limestone, the Sosio Valley, western Sicily (Italy); Wordian to upper Permian.
Description.—The examined specimens are aggregates composed of a low number of slightly curved sub-parallel tubes attached for all of their length and growing more or less close to each other. Tubes are circular in cross-section and small, with outer diameter of 0.25 to 0.40 mm, increasing very slowly. The outer surface is smooth, with only feeble growth lines visible at places. A Y-shaped ramification of a single tube due to asexual reproduction has been observed. Diagenetic recrystallization biased the original structure of the tube, and its wall is not always distinguishable from the inner filling, since large crystals occur from the outer surface towards the lumen.
Remarks.—Examined specimens can be assigned to the genus Filograna mostly based on the occurrence of bifurcating tubes. Nevertheless, the scarcity of specimens and diagnostic characters do not allow a specific determination. Tubes of Filograna, very simple and smooth, resemble those of Glomerula, found at the same studied locality, but have a slightly larger diameter and are not characteristically coiled to form aggregates.

Fig. 3. Serpulid Filograna sp. from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian. A. MSNC 4524, set of subparallel tubes on a sponge. Photograph (A1), SEM image showing a Y bifurcation of a tube (A2). B. MSNC 4525, slightly curved tubes on a sponge.
Genus Propomatoceros Ware, 1975
Type species: Propomatoceros sulcicarinatus Ware, 1975; Aptian of Southern England.
Propomatoceros permianus sp. nov.
Fig. 4.
Etymology: Named after the Permian period.
Holotype: A unique tube (MSNC4526) attached to a sponge, incomplete at anterior end.
Type locality: Sosio Valley, western Sicily (Italy).
Type horizon: “Pietra di Salomone” Limestone, Wordian to upper Permian.
Diagnosis.—Tube medium-sized, rapidly increasing in diameter, attached for all its length or seemingly rising above the substrate at its terminal end. Tube triangular in cross-section, with a median slightly denticulate keel. Denticles spaced ca. 0.30 mm from each other, directed towards the aperture. Lateral surfaces with transverse ribs, each developing from a denticle of the keel and directed towards the basal part. Basal lateral rounded thickenings enlarging the contact with the substrate.
Description.—The only available specimen incomplete at anterior end, lacking a portion of the median keel and likely its erect part, consists of a medium-sized tube, 1.6 cm long, which rapidly increases in diameter from 0.2 mm at its posteriormost preserved part to 1.4 mm at the anterior end. The tube is attached for all its length, the intermediate portion curved to form a loop. It shows a triangular cross-section and a median keel bearing barely visible denticles. The lateral surfaces are wrinkled by delicate growth striae and transverse ornamentations consisting of ribs slightly concave towards the aperture, which follow each other at short regular distances. Tube wall thickness about 0.20 mm. Basal part of attachment large and irregularly contoured due to the presence of lateral rounded thickenings in correspondence of transverse ribs. Tube strongly recrystallized.
Remarks.—Serpulid tubes showing a more or less triangular cross-section and a well-developed median keel resembling the genus Pomatoceros Philippi, 1844 (which nowadays is considered a subjective synonym of Spirobranchus Blainville, 1818) have a long range in the fossil record, reaching from the Triassic or Early Jurassic (Ippolitov 2007b; Ippolitov et al. 2014) to the Recent. The genus Propomatoceros was introduced by Ware (1975) for several Cretaceous species resembling the genus Pomatoceros, but differing at least in the anterior tube portion of adult specimens, “in having a further convex and sometimes a free cylindrical stage of variable length” (Ware 1975: 99). Moreover, Propomatoceros often has a more developed lateral ornamentation. Later, more and more fossil forms have been assigned to Propomatoceros, so that now Propomatoceros includes a plethora of species, most of questionable status or uncertain generic assignment (see Ippolitov 2007b; Vinn and Wilson 2010 for further comments and the general status of this genus).
The rapidly widening tube, the feeble denticulate keel, the conspicuous regularly spaced lateral ribs, and the irregularly thickened and contoured basal flange are the distinctive characters of P. permianus. It shows affinities to Propomatoceros semicostatus Regenhardt, 1961 and P. sulcicarinatus Ware, 1975 from the Lower Cretaceous of Germany, England, and Argentina (see Luci et al. 2013: 218, fig. 3 in part). Like the new species, P. semicostatus has a slender median keel and conspicuous growth striae, but lacks the typical irregularly contoured basal expansions which characterize the tube wall of P. permianus. P. sulcicarinatus is also similar to the present species, but possesses a thicker keel and peristome-like thickenings separated by constrictions.
Placostegus Philippi, 1844 and the attached portions of some species of Filogranula Langerhans, 1884 are similar to Propomatoceros for the keeled tube, triangular in cross-section. The genus Filogranula, documented from the late Early Jurassic to the Recent (Toarcian, ~180 Ma; Ippolitov et al. 2014), has peristomes in the attached portions and more prominent denticulated keels, especially in some one-keeled species. Placostegus known from the Upper Jurassic (Late Oxfordian, ~158 Ma) until today, also differs in the tube growth having a conspicuous erect part, and a more pronounced denticulated keel.
The name Dorsoserpula was introduced by Parsch (1956), who grouped Jurassic serpulid species according to the number of angles in the cross-section of the anterior tube portion respectively by the number of keels, for many different species possessing a median keel or comb and a tube more or less triangular in cross-section. Ware (1975) started to disentangle the Dorsoserpula lot by introducing and defining the genus Propomatoceros; see above. Jäger (2005) considerably diminished the species content of Dorsoserpula by reducing it to its type-species Serpula delphinula Goldfuss, 1831, which has a much more circular than triangular cross-section and only a weak keel, and a few other species which are morphologically very similar to the type species and also are more or less circular, although some of them have no keel at all. However, more important than keel or cross-section, the revised Dorsoserpula is characterized by the facultative presence of special morphological features which, unfortunately, in most populations are developed only in some of the individuals: a small longitudinal tube embedded laterally or along the expanded attachment flange, and a tendency to grow forming a spiral case around cylindrical or filiform substrata. To sum it up, after reducing the content of Dorsoserpula to its type species and a few similar species, the general morphological meaning of Dorsoserpula has shifted considerably away from Parsch’s (1956) original concept of systematics of Jurassic serpulids, and also the hitherto known stratigraphical range of Dorsoserpula has shifted from Lower–Upper Jurassic (Parsch 1956) to Late Jurassic–Late Cretaceous (Middle Oxfordian to latest Maastrichtian, 160–166 Ma; Jäger 2005; Ippolitov et al. 2014).
Stratigraphic and geographic range.—Wordian to upper Permian of Italy.
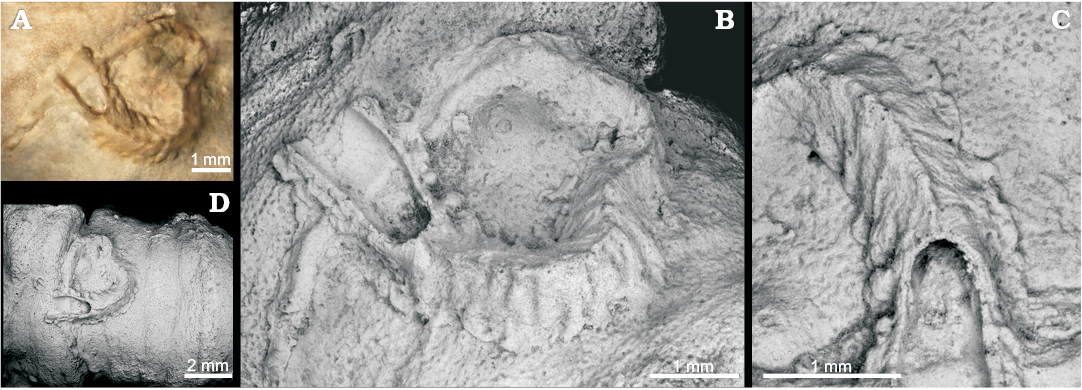
Fig. 4. Serpulid Propomatoceros permianus sp. nov. from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian, holotype MSNC 4526. A. General view of the tube encrusting a sponge. B, C. SEM images, general view (B) and detail of the tube (C) showing denticles of the median keel and the equally spaced transverse ribs. D. Detail of the median keel bearing inconspicuous denticles.
Genus Serpula Linnaeus, 1758
Type species: Serpula vermicularis Linnaeus, 1767; Recent of England.
“Serpula” distefanoi sp. nov.
Figs. 5, 6.
Etymology: Named after Pietro Di Stefano from the University of Palermo, an expert on geology and palaeontology of the Sosio Valley area.
Type material: Holotype: MSNC4527, one incomplete tube partly damaged on its upper side, attached to a sponge, partially filled by lithified sediment. Paratypes: MSNC 4528, one incomplete tube attached on a sponge; MSNC 4529, one tube fragment broken at both ends representing a part of the anterior free portion. All specimens come from type locality, except for specimen MSNC 4528 originating from the “Rupe di San Benedetto” megablock.
Type locality: Sosio Valley, western Sicily (Italy).
Type horizon: “Pietra di Salomone” Limestone, Wordian to upper Permian.
Diagnosis.—Tube quite long, straight to slightly meandering or curved. Anterior free tube portion well developed. Transverse tube section circular in the free tube portion, somewhat flattened on the upper side in the attached part. Tube relatively thick, outer surface rough.
Description.—The holotype is an adult, 5.5 cm long tube slowly increasing in diameter up to 2.4 mm. Encrusting part initially forming a loop and then nearly straight. Anterior end free, circular in cross-section, 3 mm in outer diameter, without peristomes. Tube sub-circular in cross-section and somewhat flattened on the upper side in the encrusting adult part. Outer surface quite irregular and rough, with lateral transverse arc-shaped ridges on the encrusting part. Growth lines feeble, curved like the transverse ridges, annular and straight in the erect part. Tube wall relatively thick, ca. 0.2 mm. Internal structure of the tube wall biased by recrystallization, causing the formation of densely arranged crystals, the larger crystals situated in the central part of the wall. Paratypes: incomplete tube 11 mm long and tube fragment 5 mm long broken at both ends.
Remarks.—We consider the attached tube and the fragments of free anterior portion as belonging to the same species, due to corresponding tube diameters and wall thicknesses and to corresponding characteristic rough ornamentation of the external surfaces. One additional incomplete tube attached on a sponge, lacking its upper part, is tentatively assigned to this species.
The combination of features characteristic of the tube of this species, including the general morphology and dimensions, shape of cross-section and the distinctive rough ornamentation, are not shared with any other known tube and are considered sufficient to introduce a new species. In contrast, the generic attribution of this serpulid is difficult, because neither the unaltered original tube wall structure nor the operculum is known. Consequently, we provisionally assign the new species to “Serpula”, a genus name often misused as a waste-bin to allocate species of the family Serpulidae whose characters are insufficiently known and/or do not allow an unequivocal generic attribution (see Jäger 1993). A more reliable generic attribution could be possible following a hopeful future finding of specimens preserving the unaltered tube wall structure.
Stratigraphic and geographic range.—Wordian to upper Permian of Italy.

Fig. 5. Serpulid “Serpula” distefanoi sp. nov. from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian. A. MSNC 4527, general view of the holotype. B. MSNC 4529, two opposite views (B1, B2) of a tube fragment of the anterior erect part, showing the rough surface; cross section of the erect tube portion (B3).

Fig. 6. Serpulid “Serpula” distefanoi sp. nov., from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian. A. Holotype MSNC 4527, SEM image of the adult portion of the tube evidencing the flattened upper part and thickenings concave towards the aperture (A1), arrow indicates the growth direction; detail of the initial part of the tube which is loop-like curved (A2); detail of the tube showing lateral growth lines and irregular thickening and roughness (A3). B. Paratype MSNC 4529, tube fragment of the anterior erect part, broken at both ends (B1), straight growth lines and rough surface are obvious on the outer tube surface. The internal mould of lithified sediment occurs within the lumen. Detail of the tube wall (B2), showing heavy recrystallization with densely patched crystals, smaller in the periphery.
Genus Pyrgopolon de Montfort, 1808
Type species: Pyrgopolon mosae de Montfort, 1808; Latest Maastrichtian of The Netherlands.
Subgenus Pyrgolophon (Turbinia) Michelin, 1845
Pyrgopolon (Turbinia?) gaiae sp. nov.
Fig. 7.
Etymology: Named after Gaia, the daughter of RS.
Holotype: One opercular cup (MSNC4530), with exposed upper surface, the remaining parts hidden by embedding lithified calcilutite.
Type locality: Sosio Valley, western Sicily (Italy).
Type horizon: “Pietra di Salomone” Limestone, Wordian to upper Permian.
Diagnosis.—Cup conical, upper side concave with numerous (34), straight radial ridges. Radial ridges relatively low, inversely U-shaped in cross-section, not reaching the centre of the cup. Outer margin of the cup forming a characteristic thick band, sculptured by numerous, regularly spaced teeth that give it the aspect of a cogwheel.
Description.—The cup is conical, 2.3 mm in diameter. The upper side is sculptured by 34 relatively low and straight radial ridges, occupying about 3/4 of the cup radius, around a smooth central depression. They are of similar length, a few ones being shorter, starting from about mid length. Ridges are inversely U-shaped in cross-section becoming thicker and more prominent towards the edge. Edge of the cup thick, sculptured like a cogwheel, with regularly spaced (0.15 mm) conical teeth each corresponding to a ridge’s termination, forming a nearly continuous ring with their bases.
Remarks.—Serpulid opercula are modified radioles of the branchial crown very variable in shape and structure, thus extensively used as taxonomic characters, both at generic and specific level. Only some serpulids possess a calcified operculum, the mineralization being normally restricted to the terminal plate. In other serpulids the terminal plate remains chitinous. In a few genera, as in Pyrgopolon, the operculum is entirely calcareous (Hove 1973; Hove and Nishi 1996; Vinn and ten Hove 2011) and thus its fossil record is diagnostically important for systematics of fossil serpulids.
The operculum described as P. (T.) gaiae has been found disconnected from its tube. It has a diameter larger than the lumina of any serpulid known from the investigated association. Nevertheless, the concave upper side of the cup and the strait radial ridges are typical of the opercula known for species of the genus Pyrgopolon de Montfort, 1808 and are sufficient to establish a separate species. In addition, the thick denticulate band on the margin of the cup and the inversely U-shaped radial ridges allow to include the new operculum in Turbinia Michelin, 1845 which Jäger (1993) considers to be a subgenus of Pyrgopolon. Particularly, the new species is similar to P. (Turbinia?) cunialense Gatto and Radwańska, 2000, but differs for its larger size (diameter 2.3 mm instead of 0.7–1.36 mm), more concave shape and the lower number of radial ridges (34 instead of 36–46) (Gatto and Radwańska 2000: 350–351, pls. 7, 8).
The strong similarity between the new Permian species and those described by Gatto and Radwańska (2000) from the upper Eocene on NE Italy, was unexpected and points to a certain persistence of such morphological features during very long geological time. Furthermore, the new species P. (T.) gaiae largely predates the first appearance of Pyrgopolon, hitherto known from the Barremian, Early Cretaceous (Jäger 2011). This finding thus highlights a wide geographic and temporal hiatus with previous known occurrences.
Stratigraphic and geographic range.—Wordian to upper Permian of Italy.

Fig. 7. Operculum of serpulid Pyrgopolon (Turbinia?) gaiae sp. nov. from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian, MSNC 4530, holotype. Frontal (A) and lateral (B) views of the cup. C. Detail of upper side of the cup showing radial ridges endings with cone-shaped teeth regularly spaced (0.15 mm). D. SEM image of the upper side of the cup, the denticulated outer edge forming a nearly continuous ring is obvious.
Serpulidae gen. et sp. indet.
Fig. 8.
Material.—A fragment, broken at both ends, of a presumably adult, straight free anterior tube portion (MSNC 4531) originating from “Pietra di Salomone” Limestone, the Sosio Valley, western Sicily (Italy); Wordian to upper Permian.
Description.—The sole collected specimen is 9.5 mm long and 2.9 mm in cross section and appears entirely recrystallized. It consists of a tube fragment corresponding to an anterior tube portion. It is rather massive and solid, circular in cross-section with a constant diameter of 2.9 mm. Lumen 2.4 mm wide. Tube wall thick (0.3 mm). Outer surface smooth, except for densely situated thin but distinct circular growth lines, equally spaced. Some more marked growth lines may be interpreted as weak annular peristomes, although there is no obvious difference between growth lines and peristomes. Posterior encrusting tube portion unknown. Large secondary calcite crystals occur in the tube wall, whose original structure is totally biased by diagenetic recrystallization.
Remarks.—The specimen exhibits unequivocal serpulid characters, which are otherwise too poor for specific and generic assignment.
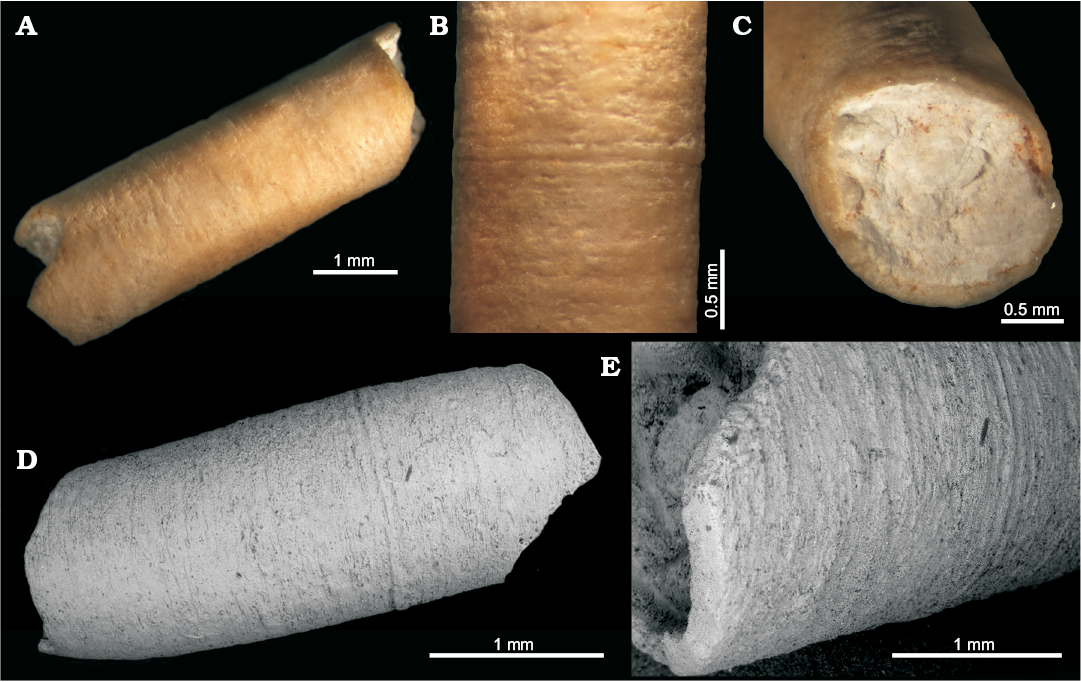
Fig. 8. Serpulidae sp. indet. from the Sosio Valley, western Sicily (Italy), Wordian to upper Permian, MSNC 4531. A. Tube fragment of the anterior erect part, broken at both ends. B. Detail of the outer surface showing thin and straight growth lines. C. Circular cross-section of the tube displaying the wall thickness; the lumen of the tube is filled by lithified sediment. D. Micromorphology on the outer surface under SEM image magnification: a straight peristome extremely feeble and thin growth lines equally spaced are obvious. E. Detail of one tube end showing the wall thickness (ca. 0.3 mm).
Discussion
Systematics.—The main characters normally used in the taxonomy of fossil serpulids, and also calcareous sabellids, are dimension and shape of the tubes (including coiling/curving and expansion rate), ornamentation, structures for attachment to the substrate and type of aggregation (see Ippolitov et al. 2014), and, since a few decades, tube structure and ultrastructure which, however, is often obliterated in the fossil material, especially when derived from relatively old epochs and/or termally maturated rocks.
Owing to this limited number of characters, taxonomy of fossil serpulids (exclusively represented by their tubes and, in only a few species, by their opercula) may be sometimes difficult, especially in some genera displaying simple morphologies. Furthermore, reliable generic attribution based exclusively on tubes may be problematic, especially in cases of high intrageneric variability (Ippolitov et al. 2014). Therefore, the taxonomic attribution of fossil tubes and their inclusion in genera represented by extant species is often problematic, and attributions mainly relate to some more easily recognizable genera.
Despite these taxonomic difficulties, the morphological characters of the described fossil tubes allowed us to allocate them within the families Sabellidae and Serpulidae. The herein described species include two sabellid and five serpulid species, four of them being new species and one new combination of already existing genus and species names. The remaining two forms belong also to typical serpulids, but the insufficient preservation state and the marked incompleteness of their tubes allow only a determination at genus (Filograna sp.) or even higher taxonomic rank (Serpulidae sp. indet.). Both sabellid species (belonging to the genus Glomerula) and one serpulid (Filograna sp.) share smooth and tiny tubes, as in their primitive congeneric forms (see below). Conversely, further associated serpulids are relatively large-sized and already diversified, displaying different shapes and ornamentations. Indeed, tubes of P. permianus sp. nov. and “S.” distefanoi sp. nov. are 1.4 and 2.4 mm wide respectively, the latter species being more than 5 cm long. The tube of P. permianus is triangular and bears a denticulate keel, lateral ribs and basal structures, while “S.” distefanoi has a subcircular flattened tube, with an extremely rough surface. Thin but distinct incremental lines, the strongest of them may be interpreted as weak peristomes, are present in Serpulidae sp. which is also larger (tube diameter 2.9 mm) than the previous two species. The finding of a calcified operculum (P. gaiae) points to the first presence of a further genus Pyrgopolon, with tubes typically stout and marked by strong keels.
The presence of a differentiate serpulid association also has implications in the taxonomic attribution of P. sosiensis. This species, recorded from the same rock and first prudently described as incertae sedis although with a probable serpulid affinity (Sanfilippo et al. 2016) can be now attributed to Serpulidae with more confidence.
Evolutionary history.—Veritable first records of serpulids so far date back to the early Middle Triassic (Anisian, ca. 245–237 Ma) and refer to very simple forms from China with the two genera Weixiserpula and Ruxigella described by Stiller (2000), from Turkey with the single species Filograna minor Senowbari-Daryan, Link, and Işintek, 2007, and from southwestern Poland with Serpula schimischowensis Assmann, 1937 and Salmacina incerta Assmann, 1937. These serpulids consist of primitive minute (up to 0.25 mm in diameter) and unornamented tubes. During the late Middle Triassic (Ladinian, 237–228 Ma) serpulids likely became widely distributed along the northern and southern margins of the Tethys Ocean (Ippolitov et al. 2014), as documented by numerous records from Turkey (Senowbari-Daryan and Link 2005), Spain (Schmidt and von Pia 1935; Flügel et al. 1984; Braga and Lopez-Lopez 1989) and Italy (Iannace and Zamparelli 1996; Cirilli et al. 1999). These serpulids still possessed simple unornamented tubes, but were larger than the Anisian ones, ranging in diameter from 0.3 to 0.9 mm and were attributed to Sarcinella (Filograna) socialis (Goldfuss, 1831) and S. schimischowensis by Flügel et al. (1984). Further larger tubes (up to 1.3 mm in diameter) with a single median keel were attributed to Dorsoserpula (Flügel et al. 1984).
By the Jurassic, serpulids evolved and diversified into new genera, including Serpula and Propomatoceros, and a total of about 100 species due to the high rate of speciation and ecological radiation (see Ippolitov et al. 2014 for a review). The new taxa were characterized by larger tubes showing a wide range of cross sections, ornamentations and peristomes (Parsch 1956; Jäger 1983, 1993, 2005; Ippolitov 2007a, b, 2010; Jäger and Schubert 2008; Vinn and Wilson 2010). From the Cretaceous onwards, serpulids are relatively well known and recorded in a series of papers spanning from the beginning of the 20th century to nowadays (e.g., Götz 1931; Wrigley 1951; Parsch 1956; Regenhardt 1961; Lommerzheim 1979; Jäger 1983, 1993, 2005; Ippolitov 2010; Ziegler 1984). Further records relate to Plio-Pleistocene times and the Recent, with a high number of genera and species (e.g., Zibrowius 1973, 1979; Hove and van den Hurk 1993; Di Geronimo et al. 1995, 1997a, b, 2002; Sanfilippo 1999, 2001, 2003, 2009a, b; Rosso et al. 2010, 2013, 2015; Kupriyanova et al. 2011).
In this historical context the finding of the described species from the Sosio Valley in Sicily is relevant because it further confirms the occurrence of Sabellidae already in the Palaeozoic. More interestingly, these findings predate the first appearance of Serpulidae from the Middle Triassic back to the Permian, at least for about 5 My (if a Changhsingian age is confirmed) or even for some tens of My (if an Artinskian age is confirmed). At the same time, the new data alter the order of appearance of some groups (see Ippolitov et al. 2014), thus questioning phylogenetic relationships between these groups. The finding of the genera Serpula, Propomatoceros, and Pyrgopolon is a proof of the existence of some representatives of the Clade A of Ippolitov et al. (2014) since the Palaeozoic, thus largely pre-dating the Early Jurassic appearance of this more advanced serpulid group (Ippolitov et al. 2014).
Palaeoecology.—According to the associated faunas, recorded species must have thrived in a shallow-marine environment, corresponding to the outer edge of a carbonate platform (see Sanfilippo et al. 2016). Calcareous breccia and limestone, which constitute the blocks including the examined taxa, represent broken(?) marginal reef portions displaced at the base of the reef slope within deeper palaeoenvironments. We suggest that the stout and massive tubes of some of the described species, i.e., “S.” distefanoi sp. nov., P. permianus sp. nov., Serpulidae sp., and P. sosiensis may represent an adaptive response to exposed and shallow-reef environment subject to waves. In such environment, the minute-sized taxa Glomerula and Filograna could have thrived in sheltered reef micro-habitats.
Stratigraphic and palaeogeographic range.—Up to now all the species described here except for G. testatrix are known exclusively from limestone beds of Wordian to upper Permian age cropping out along the Sosio Valley. More precise stratigraphic specification would require an accurate dating of sediment including the tubes, which is beyond the scope of this paper.
As far as known at present, the palaeogeographic distribution is restricted to the western margin of the Tethys. Like for other species first discovered in the Sosio Valley outcrops and subsequently found in other sectors of the Permian Tethys, it can be hypothesized that these sabellid and serpulid species were seemingly more widespread than presently known. The report of some figured “tube worms, resembling serpulids” recently found in uppermost Permian limestone of Slovenia (Sremac et al. 2016), corroborates this hypothesis.
Conclusions
The morphological characters of fossil tubes found from the Permian limestone of western Sicily allowed to unequivocally allocate them within the families Sabellidae and Serpulidae. A total of seven taxa was recorded, confirming the presence of sabellids in the Palaeozoic, and ascertaining the earliest records of veritable serpulids in the Permian period.
These species, together with the recently described P. sosiensis (Sanfilippo et al. 2016), indicate a calcified tube-dwelling polychaete community surprisingly diversified, thriving in a Permian reef paleoenvironment. The presence of large sized serpulid species with relatively complex shapes considered as advanced characters, suggests an even earlier evolution for this group.
Permian appearance of serpulids before the Permian–Triassic mass extinction points to their persistence through this catastrophic event and to the survival of the entire group.
Further study about the taxonomic significance of tube features in serpulid Polychaeta especially if supported by further findings, are needed to shed light on the phylogeny of this group and its earliest history.
Acknowledgements
The authors thank the Dipartimento Azienda Foreste Demaniali of Sicily for authorisation to collect Permian material from the outcrop and the staff of researchers, naturalists, and geologists collaborators: Filippo Spadola, Francesco Cavallaro (both Natural History Museum of Comiso, Italy) and Mario Dipasquale, Rosario Occhipinti, and Lorenzo Zurla (all Associazione Onlus Ulixes, Ragusa, Italy) for collecting the material. Alfio Viola (Department of Biological, Geological, and Environmental Sciences, University of Catania, Italy) and Ruggero Sciuto (University of Oxford, UK) are acknowledged for assistance with the SEM acquisition of images and helping in bibliography research, respectively. Alexei Ippolitov (University of Moscow, Russia) suggested some taxonomic refinements. The two reviewers Manfred Jäger (Rosenfeld, Germany) and Olev Vinn (University of Tartu, Estonia) greatly improved the paper with their invaluable comments and suggestions. This paper was financially supported by University of Catania Grants (Catania Palaeontological Research Group: contribution n. 417) to RS and AR.
References
Berra, F. and Jadoul, F. 1996. Norian serpulid and microbial bioconstructions: implications for the platform evolution in the Lombardy Basin (Southern Alps, Italy). Facies 35: 143–162. Crossref
Braga, J.C. and López-López, J.R. 1989. Serpulid bioconstructions at the Triassic–Liassic boundary in southern Spain. Facies 21: 1–10. Crossref
Cirilli, S., Iannace, A., Jadoul, F., and Zamparelli, V. 1999. Microbial serpulid build-ups in the Norian–Rhaetian of the Western Mediterranean area: ecological response of shelf margin communities to stressed environments. Terra Nova 11: 195–202. Crossref
Di Stefano, P. and Gullo, M. 1996. Permian deposits of Sicily. A review. Geodiversitas 19: 193–202.
D’Onghia, G., Capezzuto, F., Cardone, F., Carlucci, R., Carluccio, A., Chimienti, G., Corriero, G., Longo, C., Maiorano, P., Mastrototaro, F., Panetta, P., Rosso, A., Sanfilippo, R., Sion, L., and Tursi, A. 2015. Macro- and megafauna recorded in the submarine Bari Canyon (southern Adriatic, Mediterranean Sea) using different tools. Mediterranean Marine Science 16: 180–196. Crossref
Ernst, A.K. 2000. Permian Bryozoans of the NW-Tethys. Facies 43: 79–102. Crossref
Fischer, R., Pernet, B., and Reitner, J. 2000. Organomineralization of cirratulid annelid tubes—fossil and Recent examples. Facies 42: 35–50. Crossref
Flügel, E., Di Stefano, P., and Senowbari-Daryan, B. 1991. Microfacies and depositional structure of allochthonous carbonate base-of-slope deposits: The Late Permian Pietra di Salomone megablock, Sosio Valley (western Sicily). Facies 25: 147–186. Crossref
Flügel, E., Flügel-Kahler, E., Martin, J.M., and Martin-Algarra, A. 1984. Middle Triassic reefs from Southern Spain. Facies 11: 173–218. Crossref
Hove, H.A. ten and Weerdenburg, J.C.A. 1978. A generic revision of the brackish-water serpulid Ficopomatus Southern, 1921 (Polychaeta: Serpulinae), including Mercierella Fauvel 1923, Sphaeropomatus Treadwell 1934, Mercierellopsis Rioja 1945 and Neopomatus Pillai 1960. Biological Bulletin 154: 96–120. Crossref
Hove, H.A. ten and Zibrowius, H. 1986. Laminatubus alvini gen. et sp. n. and Protis hydrothermica sp. n. (Polychaeta, Serpulidae) from the bathyal hydrothermal vent communities in the eastern Pacific. Zoologica Scripta 15: 21–31. Crossref
Ippolitov, A.P. 2007a. Contribution to the revision of some late Callovian serpulids (Annelida, Polychaeta) of central Russia: Part 1. Paleontological Journal 41: 260–267. Crossref
Ippolitov, A.P. 2007b. Contribution to the revision of some late Callovian serpulids (Annelida, Polychaeta) of central Russia: Part 2. Paleontological Journal 41: 429–436. Crossref
Jones, W.T, Feldmann, R.M., Schweitzer, C.E., Reitano, A., and Insacco, G. 2015. A new pygocephalomorph (Peracarida) from the Permian of the Sosio Valley (Sicily, Italy). Journal of Crustacean Biology 35: 627–632. Crossref
Kiessling, W., Scasso, R., Aberhan, M., Ruiz, L., and Weidemeyer, S. 2006. A Maastrichtian microbial reef and associated limestones in the Roca Formation of Patagonia (Neuquén Province, Argentina). Fossil Record —Mitteilungen aus dem Museum für Naturkunde 9: 183–197. Crossref
Kupriyanova, E.K., Hove, H.A. ten, Sket, B., Trontelj, P., Zakšek, V., and Rouse, G.W. 2009. Evolution of a unique freshwater cave dwelling serpulid polychaete Marifugia cavatica Absolon and Hrabĕ, 1930. Systematics and Biodiversity 7: 389–401. Crossref
Kupriyanova, E.K., Vinn, O., Taylor, P.D., Schopf, J.W., Kudryavtsev, A., and Bailey-Brock, J. 2014. Serpulids living deep: calcareous tubeworms beyond the abyss. Deep-Sea Research Part I 90, 91–104. Crossref
Luci, L., Garberoglio, R.M., and Lazo, R.G. 2013. Serpulids and other calcareous tube-dwelling encrusting polychaetes from the Early Cretaceous Agrio Formation (Neuquén Basin, Argentina). Géobios 46: 213–224. Crossref
Nestell, G.P. and Nestell, M.K. 2006. Middle Permian (Late Guadalupian) foraminifers from Dark Canyon, Guadalupe Mountains, New Mexico. Micropaleontology 52: 1–50. Crossref
Palma, R.M. and Angeleri, M.P. 1992. Early Cretaceous serpulid limestones: Chachao Formation, Neuquén Basin, Argentina. Facies 27: 175–178. Crossref
Rosso, A., Sanfilippo, R., Ruggieri, R., Maniscalco, R., and Vertino, A. 2015. Exceptional record of submarine cave communities from the Pleistocene of Sicily (Italy). Lethaia 48: 133–144. Crossref
Rosso, A., Vertino, A., Di Geronimo, I., Sanfilippo, R., Sciuto, F., Di Geronimo, R., Violanti, D., Corselli, C. Taviani, M., Mastrototaro, F., and Tursi, A. 2010. Hard and soft-bottom thanatofacies from the Santa Maria di Leuca deep-water coral province, Mediterranean. Deep Sea Research II. Topical Studies in Oceanography 57: 360–379. Crossref
Sanfilippo, R. 2001. Bathyvermilia islandica (Polychaeta, Serpulidae): new species from the southern Icelandic Sea. Sarsia 86: 177–182. Crossref
Sanfilippo, R. 2009a. New species of Hyalopomatus Marenzeller, 1878 (Annelida, Polychaeta, Serpulidae) from Recent Mediterranean deep-water coral mounds and comments on some congeners. Zoosystema 31: 147–161. Crossref
Sanfilippo, R. 2009b. Systematics and life habit in Serpula israelitica Amoureux, 1976 (Polychaeta Serpulidae) from the Mediterranean with remarks on other soft-bottom serpulids. Journal of Natural History 43: 2009–2025. Crossref
Sanfilippo, R., Vertino, A., Rosso, A., Beuck, L., Freiwald, A., and Taviani, M. 2013. Serpula aggregates and their role in deep-sea coral communities in the southern Adriatic Sea. Facies 59: 663–677. Crossref
Senowbari-Daryan, B. and Link, M. 2005. Filograna (colonial serpulid worm tubes) from Upper Triassic (Norian) reef boulders of Taurus Mts. (southern Turkey). Facies 51: 454–459. Crossref
Sremac, J., Jurkovšek, B., Aljinović, D., and Kolar-Jurkovšek, T. 2016. Equatorial Palaeotethys as the last sanctuary for late Permian metazoan reef-builders: New evidence from the Bellerophon Formation of Slovenia. Palaeogeography, Palaeoclimatology, Palaeoecology 454: 91–100. Crossref
Taylor, P.D. and Vinn, O. 2006. Convergent morphology in small spiral worm tubes (“Spirorbis”) and its palaeoenvironmental implications. Journal of the Geological Society, London 163: 225–228. Crossref
Vinn, O. and Mutvei, H. 2009. Calcareous tubeworms of the Phanerozoic. Estonian Journal of Earth Sciences 58: 286–296. Crossref
Vinn, O. and Wilson, M.A. 2010. Sabellid-dominated shallow water calcareous polychaete tubeworm association from the equatorial Tethys Ocean (Matmor Formation, Middle Jurassic, Israel). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 258: 31–38. Crossref
Vinn, O. and Hove, H.A. ten 2011. Microstructure and formation of the calcareous operculum in Pyrgopolon ctenactis and Spirobranchus giganteus (Annelida, Serpulidae). Zoomorphology 130: 181–188. Crossref
Vinn, O., Hove, H.A. ten, and Mutvei, H. 2008a. On the tube ultrastructure and origin of calcification in sabellids (Annelida Polychaeta). Palaeontology 51: 295–301. Crossref
Vinn, O., Hove, H.A. ten, Mutvei, H., and Kirsimäe, K. 2008b. Ultrastructure and mineral composition of serpulid tubes (Polychaeta, Annelida). Zoological Journal of the Linnean Society, London 154: 633–650. Crossref
Vinn, O., Kupriyanova, E.K., and Kiel, S. 2012. Systematics of serpulid tubeworms (Annelida, Polychaeta) from Cretaceous and Cenozoic hydrocarbon-seep deposits in North America and Europe. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 265: 315–325. Crossref
Vinn, O., Kupriyanova, E.K., and Kiel, S. 2013. Serpulids (Annelida, Polychaeta) at Cretaceous to modern hydrocarbon seeps: ecologic and evolutionary patterns. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 35–41. Crossref
Ware, S. 1975. British Lower Greensand Serpulidae. Palaeontology 18: 93–116.
Wilson, M.A., Yancey, T.E., and Vinn, O. 2011. A new microconchid tubeworm from the Lower Permian (Artinskian) of central Texas, USA. Acta Palaeontologica Polonica 56: 785–791. Crossref
Wrigley, A. 1951. Some Eocene serpulids. Proceedings of the Geologists Association 62: 177–202. Crossref
Zatoń, M., Grey, M., and Vinn, O. 2014. Microconchid tubeworms (class Tentaculita) from the Joggins Formation (Pennsylvanian), Nova Scotia, Canada. Canadian Journal of Earth Sciences 51: 669–676. Crossref