
Tarsal morphology of the pleuraspidotheriid mammal Hilalia from the middle Eocene of Turkey
GREGOIRE METAIS, OZAN ERDAL, KORHAN ERTURAÇ, and K. CHRISTOPHER BEARD
Métais, G., Erdal, O., Erturaç, K., and Beard, K.C. 2017. Tarsal morphology of the pleuraspidotheriid mammal Hilalia from the middle Eocene of Turkey. Acta Palaeontologica Polonica 62 (1): 173–179.
Pleuraspidotheriids are a group of primitive ungulate mammals that, until recently, were thought to be restricted to the late Paleocene of Western Europe. It has been hypothesized that this family actually survived in Central Anatolia until at least the middle Eocene. However, these anachronistically young Anatolian “survivors”, including the genus Hilalia, were previously documented mainly by dental remains. Here, we describe the previously unknown astragalus of Hilalia saribeya, which confirms the pleuraspidotheriid affinities of the genus, and supports phylogenetic reconstructions that place Hilalia as the sister group of Pleuraspidotherium. The morphology of the astragalus suggests sub-cursorial plantigrade locomotion for H. saribeya, although its tarsal morphology remains generalized enough that scansorial capabilities cannot be ruled out. The evolution of Hilalia is addressed in the context of the apparent geographic isolation of Central Anatolia during the Eocene. The endemic character of the mammalian fauna of Central Anatolia during the middle Eocene emphasizes how the complex paleogeography of the northern margin of Neotethys impacted local biotas in a region situated at the crossroads of very distinctive biogeographic zones.
Key words: Mammalia, Pleuraspidotheriidae, paleogeography, Eocene, Turkey, Anatolia.
Grégoire Métais [metais@mnhn.fr], CR2P, Paléobiodiversité et Paléoenvironnements, UMR 7207 (CNRS, MNHN, UPMC), Sorbonne Université, Muséum National d’Histoire Naturelle, 8 rue Buffon, 75005 Paris, France; and Department of Ecology & Evolutionary Biology, University of Kansas, Lawrence, Kansas 66045, USA.
Ozan Erdal [erdalo@itu.edu.tr], Eurasia Institute of Earth Sciences, İstanbul Technical University, Istanbul, Turkey.
Korhan Erturaç [erturac@gmail.com], Department of Geography, Sakarya University, 54187 Esentepe, Sakarya, Turkey.
K. Christopher Beard [chris.beard@ku.edu], Biodiversity Institute, University of Kansas, Lawrence, Kansas 66045, USA; and Department of Ecology & Evolutionary Biology, University of Kansas, Lawrence, Kansas 66045, USA.
Received 6 October 2016, accepted 22 November 2016, available online 21 December 2016.
Copyright © 2017 G. Métais et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The early Cenozoic mammalian group “Condylarthra” was originally described by Cope (1881, 1884) on the basis of several postcranial features, including a distally convex astragalar head and the nature and extent of its contact with the navicular. It is now widely recognized that Cope’s conception of “Condylarthra” includes a highly diverse and paraphyletic or even polyphyletic assemblage of early eutherian herbivores and omnivores (e.g., Archibald 1998). Nevertheless, tarsal morphology has often been used to interpret the systematics and locomotor behavior of primitive ungulate mammals, including many of the taxa that have conventionally been considered as “condylarths” (Matthew 1937; Schaeffer 1947; Cifelli 1983; Rose 1985, 1996).
During the late Paleocene in Western Europe, the family Pleuraspidotheriidae, represented by the genera Orthaspidotherium and Pleuraspidotherium, was a particularly abundant group of medium-sized basal ungulates or “condylarths” (e.g., Russell 1964). The youngest known record of pleuraspidotheriids in Western Europe occurs in the latest Paleocene Rivecourt local fauna from the north-central part of the Paris Basin (Smith et al. 2014). While pleuraspidotheriids apparently became extinct in Western Europe at or near the Paleocene–Eocene boundary, the family persisted at least until the middle Eocene in Central Turkey, where it is documented by several species of the genus Hilalia. Maas et al. (2001) first described Hilalia as an indeterminate “condylarth”, but Hilalia has more recently been interpreted as a pleuraspidotheriid on the basis of its dental morphology (Ladevèze et al. 2010). Likewise, Parabunodon, from the ?early Eocene Çeltek Formation, Amasya Province, Central Anatolia, was first described as a choeropotamid artiodactyl (Ducrocq and Sen 1991), but has since been re-interpreted as a bunodont pleuraspidotheriid by Métais et al. (2012).
In addition to the relatively abundant teeth and jaws of Hilalia reported by Maas et al. (2001), these workers also described a partial calcaneum as pertaining to Hilalia saribeya, which is the genotypic (and most common) species. Maas et al. (2001) found that this calcaneum lacks derived features of artiodactyl, perissodactyl or hyracoid calcanea. In contrast, they noted several features in which the partial calcaneum of Hilalia resembles those of the “condylarths” Meniscotherium and (especially) Pleuraspidotherium. However, with the notable exception of the transverse orientation of the cuboid facet, which Hilalia shares with Pleuraspidotherium but not with Meniscotherium, these calcaneal features were interpreted as symplesiomorphies, and thus of little significance for systematics.
Here, we describe an isolated astragalus of Hilalia from the Lülük Member of the Uzunçarşıdere Formation or UCF (Fig. 1), the same lithological unit that has yielded all other known specimens of Hilalia (Maas et al. 2001). More than half of all fossil mammal specimens collected so far from the UCF belong to Hilalia, and the size and morphology of the new astragalus are fully consistent with an attribution to this taxon. Likewise, no other placental mammal taxon currently documented from the UCF fauna is appropriate in size to be a potential candidate for this astragalus (Kappelman et al. 1996; Maas et al. 1998). The specimen was collected by surface prospecting during the 2011 field season.
Institutional abbreviations.—AÜJM, Ankara University Science Faculty Geology Museum, Ankara, Turkey; EOU, Eskişehir Osmangazi University, Eskişehir, Turkey
Other abbreviations.—IAES, İzmir-Ankara-Erzincan Suture; UCF, Uzunçarşıdere Formation.
Geological setting
The Neotethyan realm within the extent of the modern eastern Mediterranean region can be considered as a flexible collage of interconnected small ocean basins, microcontinents and carbonate platforms (Şengör and Yılmaz 1981; Robertson and Dixon 1984). Within this collage, five different tectonic zones are recognized for modern Turkey. These include the Pontides, Sakarya continent, Kırşehir Block, Taurides (sometimes Anatolides-Taurides) and Menderes Block (Okay and Tüysüz 1999; Bozkurt and Mittwede 2001). Three separate ocean basins are recognized within the Eastern Mediterranean: Intra-Pontide, northern Neotethys, and southern Neotethys. A portion of the southern Neotethys still survives as the modern Eastern Mediterranean, in contrast to the two former oceanic basins, which are no longer in existence. The northern branch of Neotethys was closed by a terminal collision and sutured during the Late Cretaceous–early Paleogene between the Pontides and the Tauride-Anatolides platform along the İzmir-Ankara-Erzincan Suture (IAES). The Orhaniye Basin, within which fossiliferous outcrops of the UCF occur, is a retroarc foreland basin that developed immediately north of the IAES zone during the early Paleogene (Licht et al. in press). A thick sequence of marine and continental rocks in the Orhaniye Basin including the fluviolacustrine UCF records the subaerial and marine episodes that occurred locally during the Paleogene (Fig. 1). The best outcrops and the type section of the UCF occur in the N-S trending valley excavated by the Uzunçarşı stream, originating from Dedeçam Hill (Fig. 1). The thickness of the UCF approximates 500 m, but the thickness varies laterally. The basal Lülük Member of the UCF consists of braided river deposits alternating with fossiliferous reddish paleosols developed on abandoned fluvial bars. The Lülük Member is conformably overlain by off-white lacustrine limestone and green/gray mudstone that have been dated as Lutetian (~43 Ma; Licht et al. in press).
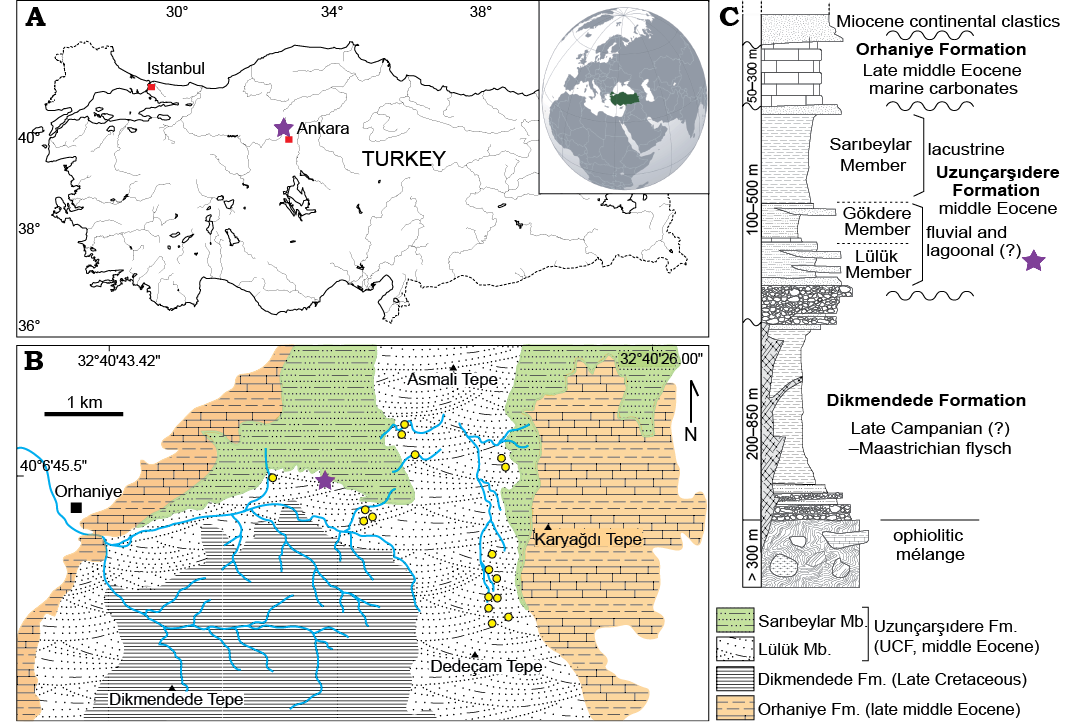
Fig. 1. Map of Turkey with the location of the Orhaniye Basin in Central Anatolia (A), geological map of the Orhaniye Basin (B) showing the fossil locality that yielded the astragalus described here, and a synthetic log (C) showing the succession of lithological units exposed in the Orhaniye Basin. Purple stars indicate the location of the fossil site that yielded EOU-UCF-1.
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Order “Condylarthra” Cope, 1881
Family Pleuraspidotheriidae Zittel, 1892
Genus Hilalia Maas, Thewissen, Sen, Kazanci, and Kappelman, 2001
Hilalia saribeya Maas, Thewissen, Sen, Kazanci, and Kappelman, 2001
Material.—EOU-UCF-1, right astragalus from late middle Eocene East Orhaniye locality 1, Lülük Member of the Uzunçarşıdere Formation, central Anatolia (Fig. 1B).
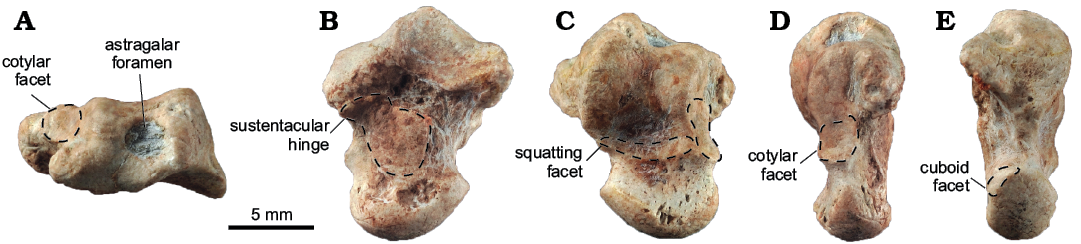
Fig. 2. Hilalia saribeya Maas, Thewissen, Sen, Kazanci, and Kappelman, 2001 from the middle Eocene Uzunçarşıdere Formation, Central Turkey; right astragalus (EOU-UCF-1) in proximal (A), plantar (B), dorsal (C), medial (D), and lateral (E) views. Each homologous articular facet is marked by dashed line.
Table 1. Measurements (in mm) for the EOU-UCF-1 astragalus of Hilalia saribeya from the middle Eocene Uzunçarşıdere Formation, Central Turkey.
|
Proximal width of astragalar body |
7.1 |
|
Lateral length of astragalar body |
8.9 |
|
Maximum width of astragalar body |
11.4 |
|
Proximal width of astragalar trochlea |
6.6 |
|
Length of neck parallel to long axis |
1.4 |
|
Maximum width of neck |
6.1 |
|
Maximum length of astragalar parallel to long axis |
14.0 |
|
Long axis of astragalar head |
3.2 |
|
Long axis of astragalar head (transverse) |
7.2 |
Description.—EOU-UCF-1 is a right astragalus (Fig. 2) that is comparable in size (Table 1) to that of Pleuraspidotherium aumonieri from Cernay (France, MP6). The astragalar neck is proximodistally short and mediolaterally broad compared to the astragali of Orthaspidotherium and Pleuraspidotherium (Ladevèze et al. 2010: figs. 9, 10). The relatively deep astragalar body bears a moderately grooved tibial trochlea with well-defined and asymmetrical medial and lateral crests. The tibial trochlea is shallow, without sharp crests, and it forms an angle of only 12° with the neck, roughly half the value found in Pleuraspidotherium and Orthaspidotherium. The astragalar foramen superior is present, suggesting the presence of a large astragalar canal for passage of a branch of the peroneal artery. However, the astragalar foramen inferior, if present, is obscured by diagenetic micrite.
In dorsal view, the distal edge of the tibial trochlea displays a transversely elongated squatting facet that extends slightly onto the astragalar neck as in Pleuraspidotherium and Orthaspidotherium. The astragalar head is mediolaterally broader than in Pleuraspidotherium and (especially) Orthaspidotherium, and it is moderately rounded and much wider transversely than dorsoplantarly. The navicular facet occupies most of the distal side of the astragalar head, and it extends far onto the medial side of the neck to approximate the cotylar fossa. A small articular facet for the cuboid occurs on the planto-lateral side of the astragalar head in EOU-UCF-1, thus resembling the astragali of Pleuraspidotherium (Thewissen, 1991) and apheliscine “condylarths” such as Apheliscus and Haplomylus (Zack et al. 2005: fig. 2). On the distomedial side of the bone, a marked facet is present for the medial collateral ligament like in Pleuraspidotherium.
In medial view, the articular surface for the medial malleolus of the tibia extends distally to form a distinct cotylar fossa. This concave surface is delimited by a sharp bony ridge and is dorsoventrally taller than its homologue in Pleuraspidotherium. The cotylar facet is present but even smaller in Orthaspidotherium. The distal calcaneal facet is not visible.
In plantar view, the sustentacular facet is proximodistally longer than its transverse dimension, and its outline is similar to that of Pleuraspidotherium. The sustentacular facet is distally connected to the navicular facet by two very thin bony bridges. A wide and deep interarticular sulcus intervenes between the ectal facet and a proximomedial extension of the sustentacular facet. The latter structure, which also occurs in Pleuraspidotherium and Orthaspidotherium, was designated as the sustentacular hinge by Ladevèze et al. (2010). The sustentacular hinge is gently concave and has a rounded proximomedial edge. The sustentacular facet, which occupies the inferior surface of the astragalar neck, is slightly concave along an oblique axis parallel to the long axis of the ectal facet. The latter occupies about half of the plantar side of the body, is concave, and obliquely oriented with respect to the long axis. The long axis of the ectal facet forms an angle of ~58° with the neck; this angle is 52° in Pleuraspidotherium, and 42° in Orthaspidotherium (Ladevèze et al. 2010). The navicular facet occupies the entire width of the astragalar head, but there is a small trapezoidal facet for the cuboid plantarly.
Remarks.—According to Ladevèze et al. (2010), diagnostic tarsal features of pleuraspidotheriids include: a transversely oriented cuboid facet on the calcaneum and the presence of a squatting facet (sensu Szalay and Decker 1974), a cotylar fossa, and a sustentacular hinge on the astragalus. EOU-UCF-1 displays all three of these astragalar features and is thus regarded as an astragalus of Hilalia, the only “condylarth” known so far from the UCF. The partial calcaneum (AÜJM99-5), described previously by Maas et al. (2001) from the UCF as pertaining to Hilalia, likewise bears a transversely oriented cuboid facet. Accordingly, Hilalia appears to retain all of the known diagnostic pleuraspidotheriid tarsal features. Assignment of EOU-UCF-1 to H. saribeya is based on size. With regard to its overall morphology and proportions, EOU-UCF-1 more closely resembles astragali of Pleuraspidotherium than those of Orthaspidotherium (Lemoine 1891; Ladevèze et al. 2010). The astragalus of Orthaspidotherium is much smaller, the neck is more gracile, and the ectal facet is oriented more proximodistally (Fig. 3), suggesting a different locomotor adaptation than in Hilalia and Pleuraspidotherium. EOU-UCF-1 differs from astragali of Pleuraspidotherium in having a more robust astragalar neck and head that are less medially deviated, and a dorsoventrally wider and more strongly delimited cotylar fossa that articulates with the tibial malleolus on the medial aspect of the astragalar body. In both of the latter respects, EOU-UCF-1 diverges even more from the astragalar morphology of Orthaspidotherium. Although we have not performed a formal phylogenetic analysis that includes the new tarsal data for Hilalia, the strong phenetic similarities between EOU-UCF-1 and astragali of Pleuraspidotherium observed here are consistent with the phylogenetic results reported by Ladevèze et al. (2010), who recovered Hilalia and Pleuraspidotherium as sister taxa, with Orthaspidotherium as the sister group of the former clade.
The pleuraspidotheriids are currently known by four genera, and their origin remains unknown. Ladevèze et al. (2010) provided an historical overview of the proposed systematic affiliations of pleuraspidotheriids as well as a comprehensive analysis of the family and its phylogenetic position with respect to other “condylarths” and some modern ungulates. The strict consensus trees yielded by their analyses do not definitively resolve the affinities of pleuraspidotheriids, although a possible relationship with Chriacus is supported by two postcranial features, including the presence of a squatting facet on the astragalus of Chriacus. Afrotheres were not included in the analysis of Ladevèze et al. (2010). Louisinids, including Paschatherium and Teilhardimys (formerly Microhyus; Hooker and Russell 2012), are another group of “condylarths” restricted to the Paleocene and early Eocene of Europe. Paschatherium and Teilhardimys are considerably smaller than known pleuraspidotheriids. The astragalus of Paschatherium bears a well-developed cotylar fossa (Godinot et al. 1996), but this structure is much weaker in Teilhardimys (Tabuce et al. 2006). Both of these louisinid taxa lack the squatting facet and sustentacular hinge on the astragalus that are characteristic features of pleuraspidotheriids. Moreover, unlike the condition seen in pleuraspidotheriids, the calcanei of Paschatherium and Teilhardimys display a proximodistally elongated ectal facet (it is more tranversely oriented in Pleuraspidotherium and Orthaspidotherium).
The presence of a cotylar fossa has been proposed as a key character linking various afrotheres with some Holarctic apheliscine “condylarths” (Zack et al. 2005) and South American “native ungulates” (Agnolin and Chimento 2011), but this structure also occurs in pleuraspidotheriids, casting some doubt on the phylogenetic significance of this feature. Comparisons between the astragalus of Hilalia and those of other “condylarths” such as Protungulatum, Arctocyon, or Hyopsodus reveal that the proximal tarsus of Hilalia is distinctive in having the following combination of features: astragalar neck relatively short, transversely wide, and weakly constricted, and the presence of a squatting facet and a sustentacular hinge. During our comparisons with astragali of other “condylarths”, we noticed some similarities between EOU-UCF-1 and the astragalus of Meniscotherium (Fig. 3): the tibial trochlea is shallow and the sustentacular facet tends to extend proximomedially. However, the interarticular sulcus for the interosseous ligament remains continuous in Meniscotherium, unlike the condition in Hilalia and other pleuraspidotheriids, in which the sustentacular hinge obliterates the interarticular sulcus. Moreover, in dorsal view, the slight development of a distal extension of the tibial trochlea on the astragalus of Meniscotherium tends to mimic the squatting facet found in pleuraspidotheriids, although the structure is much less extensive in Meniscotherium.
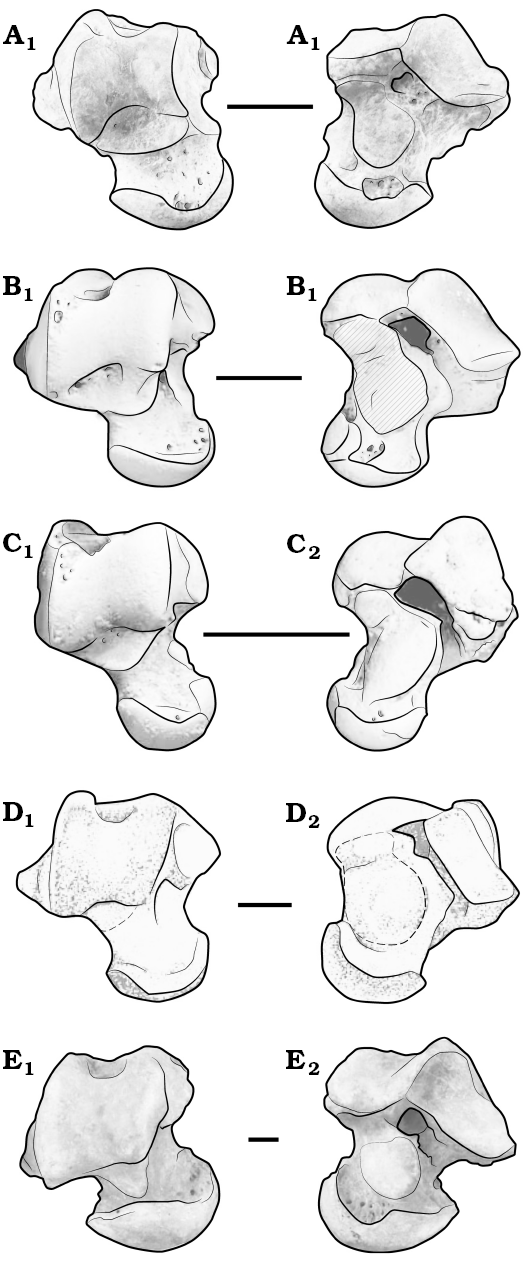
Fig. 3. Comparison of right astragali of Paleocene and Eocene “condylarths”: Hilaliasaribeya (A), Pleuraspidotherium aumonieri (B), Orthaspidotherium edwardsi (C), Meniscotherium sp. (D), Arctocyon primaevus (E), in dorsal (A1–E1) and plantar (A2–E2) views. Adapted from Argot (2013), Ladevèze et al. (2010), Williamson and Lucas (1992). Scale bars 5 mm.
Stratigraphic and geographic range.—Middle Eocene, Lülük Member of the Uzunçarşıdere Formation, Orhaniye Basin, central Turkey.
Discussion
Locomotion of Hilalia.—The EOU-UCF-1 astragalus and the fragmentary calcaneum referred to Hilalia by Maas et al. (2001) illuminate some functional aspects of the locomotion of Hilalia. The functional implications of pleuraspidotheriid tarsal morphology have been debated. Thewissen (1991) concluded that Pleuraspidotherium was a generalist terrestrial taxon with plantigrade feet, lacking the supination ability and the wide range of mobility at the ankle joint that has been described in the arctocyonid Chriacus (Rose 1987, 1996). By contrast, Ladevèze et al. (2010) noted a number of similarities between the tarsus of Pleuraspidotheriidae and that of Chriacus. Body mass estimates for Pleuraspidotherium vary from 3 to 5 kg depending on the anatomical element and metric (e.g., femoral length versus femoral midshaft diameter) employed in the analysis (Thewissen 1991). Assuming the absence of unusual allometric scaling, a similar body mass estimate can be proposed for Hilalia. The astragalar morphology of Hilalia differs from that of Pleuraspidotherium in having a wider and less medially deviated astragalar neck and head, yielding a reduced angle between the tibial trochlea and the long axis of the astragalus. These characters favor parasagittal movements and may indicate a reduced capacity for inversion and eversion in Hilalia, consistent with a terrestrial and sub-cursorial mode of locomotion. This is consistent with the moderately grooved tibial trochlea, which suggests enhanced stability at the proximal tarsal joint. The tibial trochlea is almost flat transversely in the arctocyonid Chriacus, which is interpreted to have had arboreal capabilities (Rose 1987). Likewise, the relatively transverse orientation of the ectal facet of the calcaneum in Pleuraspidotherium and Orthaspidotherium (this feature is not observable on the damaged calcaneum of Hilalia reported by Maas et al. 2001) suggests terrestrial locomotion. The ectal facet is proximodistally elongated in more scansorial/arboreal taxa such as the extant hyracoid Dendrohyrax or the arctocyonid Chriacus (Szalay and Lucas 1996: fig. 17), and the European louisinid “condylarths” Paschatherium and Teilhardimys (Godinot et al. 1996). The distal extension of the trochlear articular surface onto the dorsal side of the astragalar neck (or “squatting facet”) in Hilalia and other pleuraspidotheriids suggests that these animals frequently maintained dorsiflexed postures at the upper ankle joint, which may indicate a generalized “ambling” type of terrestrial locomotion (probably more adapted for walking on variable substrates than for running). The ranges of movement interpreted for a joint are based upon the osteological structure of the joints themselves and do not include limitations to movement imposed by soft tissue. Further data from the appendicular skeleton of Hilalia are needed in order to elucidate its locomotor adaptations and movement capabilities. It is worth noting that the posture and locomotion of primitive ungulates are often difficult to infer from skeletal morphology, mostly because there are no modern analogues, and thus biomechanical and functional hypotheses are difficult to test.
Paleobiogeography and paleoecology.—The close similarities in tarsal and dental morphology shared by Hilalia and Paleocene pleuraspidotheriids known from Western Europe make it clear that this clade persisted at least until the middle Eocene on the Pontide terrane in what is now Central Anatolia. The apparently relictual occurrence of pleuraspidotheriids in Central Anatolia highlights the endemic nature of the mammalian fauna of this region during the middle Eocene. This high level of faunal endemism presumably reflects the strong paleogeographic isolation of the Pontide terrane from Africa to the south, Europe to the north and west, and Asia to the north and east. Nevertheless, the apparent proximity of the Pontide terrane to adjacent tectonic blocks, and the high potential for emergent Tethyan island arcs due to N-S convergence in the region, made the Pontides a potential locus for mammalian dispersal from the Late Cretaceous through the Eocene. Pleuraspidotheriids must have dispersed onto the Pontide terrane sometime prior to the Paleocene–Eocene boundary, based on their documented stratigraphic distribution in Western Europe. However, the paleogeographic linkages that allowed pleuraspidotheriids to colonize the Pontide terrane must have been severed by the earliest Eocene, because there is no evidence of such common Eocene Laurasian mammal taxa as artiodactyls, perissodactyls, and rodents in the UCF fauna.
The persistence of pleuraspidotheriids on the Pontide terrane for roughly 15 Ma after their extinction in Western Europe raises many questions as to the date of their extinction in the region. The spotty Paleogene fossil record of Central Anatolia precludes attempts to track their persistence locally later in the Eocene. The late Eocene Süngülü fauna of easternmost Anatolia, although documented exclusively by small mammals, clearly displays typical Eurasian taxa (de Bruijn et al. 2003). However, paleogeographic continuity between the Süngülü fauna, from what is now the Caucasus area near Turkey’s border with the Republic of Georgia, and the Pontide terrane bearing the UCF fauna has not yet been demonstrated. In any case, the late Oligocene faunas of the Kızılırmak Formation in the Çankırı-Çorum Basin of Central Anatolia clearly show that the endemic UCF fauna was replaced by a typical Eurasian fauna by the end of the Paleogene (Antoine et al. 2008; Métais et al. 2016).
Four, and perhaps five, species of Hilalia (mostly differentiated by size) have been reported from the UCF (Maas et al. 2001), suggesting a local radiation of the genus in ecological niches occupied elsewhere in Eurasia by artiodactyls and perissodactyls. Although H. saribeya was probably a terrestrial mammal, the locomotion and ecological preferences of the other species remain unknown. Smaller species such as H. sezerorum and another unnamed tiny species mentioned by Maas et al. (2001) possibly retained some climbing capabilities, as Ladevèze et al. (2010) hypothesized for the European pleuraspidotheriid Orthaspidotherium. In any case, these closely related Turkish pleuraspidotheriids must have practiced some form of ecological niche partitioning in order to coexist, even if they maintained a very similar dental morphology.
Conclusions
The astragalar morphology of the middle Eocene “condylarth” Hilalia saribeya from central Turkey shows clear affinities with that of the late Paleocene pleuraspidotheriid Pleuraspidotherium aumonieri, a species known only from the Paris Basin. This is consistent with dental evidence suggesting that Pleuraspidotherium is more closely related to Hilalia than it is to the contemporary and sympatric genus Orthaspidotherium (Ladevèze et al. 2010). The tarsal morphology of Hilalia suggests a generalist terrestrial locomotion, although its tarsal morphology remains generalized enough that some scansorial capabilities cannot be ruled out. The endemic character of the UCF mammalian fauna emphasizes how the complex paleogeography of the northern margin of Neotethys impacted local biotas, in a region situated at the crossroads of very distinctive biogeographic zones during the Eocene.
Acknowledgements
We thank Philippe Loubry (Muséum nationale d’Histoire naturelle, Paris, France) for the photos, and Oscar Sanisidro (Biodiversity Institute, University of Kansas, Lawrence, USA) for the Fig. 3. We thank Alexis Licht (University of Washington, Seattle, USA), Faruk Ocakoğlu (Eskişehir Osmangazi University, Eskişehir, Turkey), Mike Taylor (Geology Department, University of Kansas, Lawrence, USA), and Pauline Coster (Biodiversity Institute, University of Kansas, Lawrence, USA) for helpful discussions regarding the geology and age of the UCF, John Kappelman and Mary Maas (both University of Texas, Austin, USA) for information on the UCF fauna, and Sevket Sen (Muséum nationale d’Histoire naturelle, Paris), Thierry Smith (Royal Belgian Institute of Natural Science, Bruxelles, Belgium), and Hans Thewissen (Northeast Ohio Medical University, USA) for their constructive comments on a previous version of the manuscript. This work has been supported by the INSU-2011 CT4 Interrvie, the Action Transversale Muséum (2012), the National Geographic Society CRE GRANT #9215-12, and U.S. National Science Foundation grant EAR-1543684.
References
Agnolin, F.L. and Chimento, N.R. 2011. Afrotherian affinities for endemic South American “ungulates”. Mammalian Biology 76: 101–108. Crossref
Antoine, P.-O., Karadenizli, L., Saraç, G., and Sen, S. 2008. A giant rhinocerotoid (Mammalia, Perissodactyla) from the late Oligocene of north-central Anatolia (Turkey). Zoological Journal of the Linnean Society 152: 581–592. Crossref
Archibald, J.D. 1998. Archaic ungulates (“Condylarthra”). In: C.M. Janis, K.M. Scott, and L.L. Jacobs (eds.), Evolution of Tertiary Mammals of North America, 292–331. Cambridge University Press, Cambridge.
Argot, C. 2013. Postcranial analysis of a carnivoran-like archaic ungulate: The case of Arctocyon primaevus (Arctocyonidae, Mammalia) from the late Paleocene of France. Journal of Mammalian Evolution 20: 83–114. Crossref
Bozkurt, E. and Mittwede, S.K. 2001. Introduction to the geology of Turkey—a synthesis. International Geology Review 43: 578–594. Crossref
Cifelli, R.L. 1983. The origin and affinities of the South American Condylarthra and early Tertiary Litopterna (Mammalia). American Museum Novitates 2772: 1–49.
Cope, E.D. 1881. On some Mammalia of the lower beds of New Mexico. Proceedings of the American Philosophical Society 19: 484–495.
Cope, E.D. 1884. The Condylarthra. American Naturalist 18: 790–805, 892–905. Crossref
de Bruijn, H., Ünay, E., Saraç, G., and Yïlmaz, A. 2003. A rodent assemblage from the Eo/Oligocene boundary interval near Süngülü, Lesser Caucasus, Turkey. Coloquios de Paleontología, Volumen Extraordinario 1: 47–76.
Ducrocq, S. and Sen, S. 1991. A new Haplobunodontidae (Mammalia, Artiodactyla) from the Eocene of Turkey. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1991 (1): 12–20.
Godinot, M., Smith, T., and Smith, R. 1996. Mode de vie et affinités de Paschatherium (Condylarthra, Hyopsodontidae) d’après ses os du tarse. In: M. Godinot and P.D. Gingerich (eds.), Paléobiologie et évolution des mammifères paléogènes, 225–242. Palaeovertebrata, Montpellier.
Hooker, J.J. and Russell, D.E. 2012. Early Palaeogene Louisinidae (Macroscelidea, Mammalia), their relationships and north European diversity. Zoological Journal of the Linnean Society 164: 856–936. Crossref
Kappelman, J.W., Maas, M.C., Sen, S., Alpagut, B., Fortelius, M., and Lunkka, J.-P. 1996. A new early Tertiary mammalian fauna from Turkey and its paleobiogeographic significance. Journal of Vertebrate Paleontology 16: 592–595. Crossref
Ladevèze, S., Missiaen, P., and Smith, T. 2010. First skull of Orthaspidotherium edwardsi (Mammalia, “Condylarthra”) from the late Paleocene of Berru (France) and phylogenetic affinities of the enigmatic European family Pleuraspidotheriidae. Journal of Vertebrate Paleontology 30: 1559–1578. Crossref
Lemoine, V. 1891. Etude d’ensemble sur les dents de mammifères fossiles des environs de Reims. Bulletin de la Société géologique de France 19: 263–290.
Licht, A., Coster, P., Ocakoğlu, F., Campbell, C., Métais, G., Mulch, A., Taylor, M., Kappelman, J., and Beard, K.C. (in press). Tectonostratigraphy of the Orhaniye Basin, Turkey: Implications for collision chronology and Paleogene biogeography of central Anatolia. Journal of Asian Eath Sciences.
Maas, M.C.,
Thewissen, J.G.M., and Kappelman, J.W. 1998. Hypsamasia seni
(Mammalia: Embrithopoda) and other mammals from the Eocene Kartal
Formation of Turkey. In: K.C. Beard, and M.R. Dawson (eds.), Dawn of
the age of mammals in Asia, 286-297. Bulletin of Carnegie Museum of
Natural History, Pittsburgh.
Maas, M.C., Thewissen, J.G.M., Sen, S., Kazanci, N., and Kappelman, J.W. 2001. Enigmatic new ungulates from the early middle Eocene of Central Anatolia, Turkey. Journal of Vertebrate Paleontology 21: 578–590. Crossref
Matthew, W.D. 1937. Paleocene fauna of the San Juan Basin, New Mexico. Transactions of the American Philosophical Society, New Series 30: 1–510.
Métais, G., Gheerbrant, E., and Sen, S. 2012. Re-interpretation of the genus Parabunodon (Ypresian, Turkey): implications for the evolution and distribution of pleuraspidotheriid mammals. Palaeobiodiversity and Palaeoenvironments 92: 477–486.
Métais, G., Albayrak, E., Antoine, P.-O., Erdal, O., Karadenızlı, L., Oyal, N., Saraç, G., İslamoğlu, Y., and Sen, S. 2016. Oligocene ruminants from the Kızılırmak Formation, Çankırı-Çorum Basin, Central Anatolia, Turkey. Palaeontologia Electronica 19.3.37A: 1–23.
Okay, A.I. and Tüysüz, O. 1999. Tethyan Sutures of northern Turkey. In: B. Durand, L. Jolivet, F. Hovarth, and M. Séranne (eds.), The Mediterranean Basins: Tertiary extension within the Alpine Orogen. Geological Society of London, Special Publication 156: 475–515. Crossref
Robertson, A. and Dixon, J. 1984. Introduction: aspects of the geological evolution of the Eastern Mediterranean. Geological Society of London, Special Publication 17: 1–74. Crossref
Rose, K.D. 1985. Comparative osteology of North American dichobunid artiodactyls. Journal of Paleontology 59: 1203–1226.
Rose, K.D. 1987. Climbing adaptations in the early Eocene mammal Chriacus and the origin of Artiodactyla. Science 236: 314–316. Crossref
Rose, K.D. 1996. On the origin of the order Artiodactyla. Proceedings of National Academy of Science, USA 93: 1705–1709. Crossref
Russell, D.E. 1964. Les mammifères Paléocènes d’Europe. Mémoires du Muséum National d’Histoire Naturelle, série C, Sciences de la Terre 13: 1–324.
Schaeffer, B. 1947. Notes on the origin and function of the artiodactyl tarsus. American Museum Novitates 1356: 1–24.
Şengör, A.M.C. and Yilmaz, Y. 1981. Tethyan evolution of Turkey: a plate tectonic approach. Tectonophysics 75: 181–241. Crossref
Smith, T., Quesnel, F., De Plöeg, G., De Franceschi, D., Métais, G., De Bast, E., Solé, F., Folie, A., Boura, A., Claude, J., Dupuis, C., Gagnaison, C., Iakovleva, A.I., Martin, J., Maubert, F., Prieur, J., Roche, E., Storme, J.-Y., Thomas, R., Tong, H., Yans, J., and Buffetaut, E. 2014. First Clarkforkian equivalent land mammal age in the latest Paleocene basal Sparnacian facies of Europe: fauna, flora, paleoenvironment and (bio)stratigraphy. PlosOne 9 (1): 1–19. Crossref
Szalay, F.S. and Decker, R.L. 1974. Origin, evolution, and function of the tarsus in Late Cretaceous Eutheria and Paleocene Primates. In: F.A. Jenkins, Jr. (ed.), Primate Locomotion, 223–259. Academic Press, London. Crossref
Szalay, F.S. and Lucas, S.G. 1996. The postcranial morphology of Paleocene Chriacus and Mixodectes and the phylogenetic relationships of archontan mammals. Bulletin of the New Mexico Museum of Natural History and Science 7: 1–47.
Tabuce, R., Antunes, M.T., Smith, R., and Smith, T. 2006. Dental and tarsal morphology of the European Paleocene/Eocene “condylarth” mammal Microhyus. Acta Paleontologica Polonica 51: 37–52.
Thewissen, J.G.M. 1991. Limb osteology and function of the primitive Paleocene ungulate Pleuraspidotherium with notes on Tricuspiodon and Dissacus. Géobios 24: 483–495. Crossref
Williamson, T.E. and Lucas, S.G. 1992. Meniscotherium (Mammalia, “Condylarthra”) from the Paleocene–Eocene of Western North America. Bulletin of the New Mexico Museum of Natural History and Science 1: 1–75.
Zack, S.P., Penkrot, T.A., Bloch, J.I., and Rose, K.D. 2005. Affinities of “hyopsodontids” to elephant shrews and a Holarctic origin of Afrotheria. Nature 434: 497–501. Crossref
Acta Palaeontol. Pol. 62 (1): 173–179,
2017 https://doi.org/10.4202/app.00314.2016