

A novel form of postcranial skeletal pneumaticity in a sauropod dinosaur: Implications for the paleobiology of Rebbachisauridae
LUCIO M. IBIRICU, MATTHEW C. LAMANNA, RUBÉN D.F. MARTÍNEZ, GABRIEL A. CASAL, IGNACIO A. CERDA, GASTÓN MARTÍNEZ, and LEONARDO SALGADO
Ibiricu, L.M., Lamanna, M.C., Martínez, R.D.F., Casal, G.A., Cerda, I.A., Martínez, G., and Salgado, L. 2017. A novel form of postcranial skeletal pneumaticity in a sauropod dinosaur: Implications for the paleobiology of Rebbachisauridae. Acta Palaeontologica Polonica 62 (2): 221–236.
In dinosaurs and other archosaurs, the presence of foramina connected with internal chambers in axial and appendicular bones is regarded as a robust indicator of postcranial skeletal pneumaticity (PSP). Here we analyze PSP and its paleobiological implications in rebbachisaurid diplodocoid sauropod dinosaurs based primarily on the dorsal vertebrae of Katepensaurus goicoecheai, a rebbachisaurid from the Cenomanian–Turonian (Upper Cretaceous) Bajo Barreal Formation of Patagonia, Argentina. We document a complex of interconnected pneumatic foramina and internal chambers within the dorsal vertebral transverse processes of Katepensaurus. Collectively, these structures constitute a form of PSP that has not previously been observed in sauropods, though it is closely comparable to morphologies seen in selected birds and non-avian theropods. Parts of the skeletons of Katepensaurus and other rebbachisaurid taxa such as Amazonsaurus maranhensis and Tataouinea hannibalis exhibit an elevated degree of pneumaticity relative to the conditions in many other sauropods. We interpret this extensive PSP as an adaptation for lowering the density of the skeleton, and tentatively propose that this reduced skeletal density may also have decreased the muscle energy required to move the body and the heat generated in so doing. Given that several rebbachisaurids inhabited tropical to subtropical paleolatitudes during the extreme warmth of the mid-Cretaceous, increased PSP may have better enabled these sauropods to cope with extraordinarily high temperatures. Extensive skeletal pneumaticity may have been an important innovation in Rebbachisauridae, and perhaps also in saltasaurine titanosaurs, which evolved an even greater degree of PSP. This may in turn have contributed to the evolutionary success of rebbachisaurids, which were the only diplodocoids to survive into the Late Cretaceous.
Key words: Dinosauria, Rebbachisauridae, Katepensaurus, air sac system, postcranial skeletal pneumaticity, pulmonary system, Cretaceous, Bajo Barreal Formation, Argentina.
Lucio M. Ibiricu [ibiricu@cenpat-conicet.gob.ar], Instituto Patagónico de Geología y Paleontología (CCT CONICET-CENPAT), Boulevard Almirante Brown 2915, 9120 Puerto Madryn, Chubut, Argentina.
Matthew C. Lamanna [lamannam@carnegiemnh.org], Section of Vertebrate Paleontology, Carnegie Museum of Natural History, 4400 Forbes Avenue, Pittsburgh, Pennsylvania, 15213 USA.
Rubén D. F. Martínez [rudaframartinez@gmail.com] and Gabriel A. Casal [paleogac@yahoo.com.ar], Laboratorio de Paleovertebrados, Universidad Nacional de la Patagonia San Juan Bosco, C.C. 360, 9000 Comodoro Rivadavia, Chubut, Argentina.
Ignacio A. Cerda [nachocerda6@yahoo.com.ar] and Leonardo Salgado [lsalgado@unrn.edu.ar], CONICET, Instituto de Investigación en Paleobiología y Geología, Universidad Nacional de Río Negro, Avenida General Roca 1242, 8332 General Roca, Río Negro, Argentina.
Gastón Martínez [gmartinezpsf@gmail.com], Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Avenida Vélez Sarsfield 299, 5000 Córdoba, Argentina.
Received 13 October 2016, accepted 14 February 2017, available online 7 April 2017.
Copyright © 2017 L.M. Ibiricu et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Studies of archosaurian anatomy demonstrate that the presence of foramina connected with internal chambers in axial and appendicular bones is a consistent indicator of postcranial skeletal pneumaticity (PSP) (Janensch 1947; Britt 1997; Wedel et al. 2000; Wedel 2003, 2009; O’Connor 2004, 2006; O’Connor and Claessens 2005; Sereno et al. 2008; Cerda et al. 2012; Fanti et al. 2013; Fig. 1). PSP occurs in several ornithodiran archosaur clades (i.e., Pterosauria and some groups within Dinosauria, such as Theropoda and Sauropoda; see Claessens et al. 2009; Butler et al. 2012); therefore, the tendency to pneumatize postcranial bones may constitute a synapomorphy of Ornithodira (O’Connor and Claessens 2005). Birds are the only extant vertebrates to display PSP, which originates from the pulmonary air sac system and lungs (O’Connor 2006; Wedel 2009). Birds are also unique among modern vertebrates in the heterogeneous architecture of their respiratory system (Duncker 1972; O’Connor 2004; Perry and Sander 2004). The lungs, which are fixed on the dorsal portion of the body cavity and attached to the thoracic vertebrae, are comparatively rigid structures that are united to a series of thin-walled air sacs by intrapulmonary bronchi (O’Connor 2004). One single (the clavicular) and four paired (the cervical, anterior and posterior thoracic, and abdominal) air sacs are recognized in most birds, although a minority of avian species deviate from this pattern (Duncker 1972; Wedel 2003; O’Connor 2004). The pneumatic diverticula (a network of epithelial “connector structures” that originate from the lungs and air sacs and insert in the walls of soft tissues and bones [O’Connor 2006]) have been divided into four categories: visceral, intermuscular, subcutaneous, and intraosseous (O’Connor 2004). Intraosseous diverticula arising from the cervical air sacs pneumatize the cervical vertebrae and ribs and the anterior thoracic vertebrae, whereas diverticula from the abdominal air sacs pneumatize the posterior thoracic, sacral, and caudal vertebrae plus the pelvic girdle and hind limbs (O’Connor and Claessens 2005; O’Connor 2006). The pectoral girdle and forelimbs are pneumatized by diverticula from the clavicular air sac, whereas some thoracic vertebrae and ribs are pneumatized by the lung itself. Diverticula from the anterior thoracic air sacs often pneumatize the sternum and sternal ribs, but the posterior thoracic air sacs do not pneumatize bones (Duncker 1972; Perry et al. 2011). Functionally, PSP has frequently been regarded as a weight-reducing specialization in large-bodied and/or flying avian taxa (e.g., O’Connor 2004, 2009; Benson et al. 2012). Air sacs and their respective diverticula are also generally associated with the following functions in birds: ventilation, buoyancy, vocalization, and display (Wedel 2003, 2006; Henderson 2004; Perry et al. 2009).
Rebbachisauridae is a clade of small to medium-sized, morphologically peculiar diplodocoid sauropod dinosaurs known from the Early and mid-Cretaceous of Europe, South America, and Africa. Previous works have attempted to elucidate the paleobiology of rebbachisaurids based on the skeletal anatomy of these unusual sauropods (e.g., Apesteguía 2005; Sereno and Wilson 2005; Sereno et al. 2007; Apesteguía et al. 2010; Fanti et al. 2013, 2015; Wilson and Allain 2015; Paulina Carabajal et al. 2016). Nevertheless, many aspects of rebbachisaurid paleobiology remain poorly understood, a circumstance that is due in large part to the highly incomplete nature of most fossils of these dinosaurs (Ibiricu et al. 2012). Although PSP has been described in members of various sauropod lineages, particularly by Wedel (2003), rebbachisaurids have received comparatively little attention in this regard (but see Fanti et al. 2013, 2015).
Katepensaurus goicoecheai (Ibiricu et al. 2013a, 2015) is a recently-named rebbachisaurid that was recovered from an exposure of the Cenomanian–Turonian (Upper Cretaceous) Bajo Barreal Formation in the Cañadón de Las Horquetas on the Estancia Laguna Palacios in Chubut Province, central Patagonia, Argentina. Katepensaurus, the southernmost-known rebbachisaurid taxon, was recently assigned to the subclade Limaysaurinae (Ibiricu et al. 2015); however, Fanti et al.’s (2015) phylogenetic analysis recovered this taxon as a member of Rebbachisaurinae (= Nigersaurinae; see Wilson and Allain 2015). Here, we analyze rebbachisaurid PSP and its paleobiological implications, based primarily on the dorsal vertebrae of Katepensaurus. Most notably, we document and describe a form of PSP—pneumatic chambers connected to foramina within the dorsal vertebral transverse processes—that has not previously been observed in sauropods.

Fig. 1. A. Simplified phylogeny of Diapsida showing position of Rebbachisauridae and both components of the rebbachisaurid Extant Phylogenetic Bracket sensu Witmer 1995 (Crocodylia and Aves). Rectangle indicates absence of postcranial pneumaticity; triangles indicate presence of postcranial pneumaticity. B. Hypothesized osteological correlates of pneumatic and other soft tissue structures in archosaurian vertebrae (based on O’Connor 2006).
Institutional abbreviations.—MMCH, Museo Municipal “Ernesto Bachmann”, Villa El Chocón, Neuquén Province, Argentina; MOZ-Pv, Museo Provincial de Ciencias Naturales “Prof. Dr. Juan A. Olsacher”, Colección de Paleovertebrados, Zapala, Neuquén Province, Argentina; UNPSJB-PV, Universidad Nacional de la Patagonia San Juan Bosco Laboratorio de Paleovertebrados, Comodoro Rivadavia, Chubut Province, Argentina.
Other abbreviations.—Mh, maximum height of the fossa; Ml, maximum length of the fossa; PP, Pneumaticity Profile; PSP, postcranial skeletal pneumaticity.
Material and methods
We studied the holotypic partial skeleton of Katepensaurus goicoecheai (UNPSJB-PV 1007) from the Cenomanian–Turonian (early Late Cretaceous) of central Patagonia (Lower Member of the Bajo Barreal Formation, Golfo San Jorge Basin, Chubut Province, Argentina) (Ibiricu et al. 2013a, 2015; Figs. 2–8). We examined the specimen firsthand. Furthermore, we subjected three dorsal vertebrae of the Katepensaurus holotype to computed tomographic (CT) scanning to evaluate their internal structures. The scans were performed at the Instituto del Diagnóstico del Este del Chubut, in Puerto Madryn, Chubut Province, Argentina, and at the Sanatorio Asociación Española S.M. in Comodoro Rivadavia, also in Chubut. Because of their relatively good preservation, the vertebrae selected for scanning were UNPSJB-PV 1007/4, /5, and /12 (which are anterior to middle [UNPSJB-PV 1007/12] and middle to posterior [UNPSJB-PV 1007/4, /5] dorsal vertebrae, respectively). Images in DICOM format were imported in 3D Slicer (visualization software; www.slicer.org). Soft tissues such as ligaments, muscles, cartilage, and pneumatic diverticula are almost never directly preserved in the fossil record; however, such tissues often leave traces on bones. We utilized these osteological correlates as evidence to infer the presence of these soft tissues (following, for example, Witmer 1995).
We focused on the dorsal vertebrae of Katepensaurus that exhibit the hypothesized pneumatic morphologies that are most relevant to this study (see Ibiricu et al. 2013a, 2015 for detailed descriptions and comparisons of these and other bones). The nomenclature of vertebral fossae employed here is that of Wilson et al. (2011), whereas that of vertebral laminae is from Wilson (1999).
Histological sections of a dorsal rib of Katepensaurus (UNPSJB-PV 1007/28) were prepared, based on the methodology outlined by Chinsamy and Raath (1992), in the petrographic laboratory of the Universidad Nacional de San Luis, in San Luis, San Luis Province, Argentina. The sections were studied using a Nikon E400 petrographic polarizing microscope. Nomenclature and definitions of osteohistological structures are derived from Francillon-Vieillot et al. (1990) and Chinsamy-Turan (2005).
Results
Pneumatic dorsal vertebral structures in Katepensaurus.—The external osteological evidence for the presence of soft tissues in extinct animals such as sauropod dinosaurs includes vertebral fossae (including blind [= non-communicating] and communicating fossae), cortical foramina, grooves, sulci, rugosities, and bone excavations in general (O’Connor 2006; Organ 2006; Schwarz-Wings 2009). Recently, Ibiricu et al. (2015) described new fossils of Katepensaurus goicoecheai; among these are one anterior and two anterior to middle dorsal vertebrae (specimen numbers UNPSJB-PV 1007/13, /12, and /31, respectively; Figs. 2A, B, 3). In the anterior dorsal vertebra UNPSJB-PV 1007/13, the left lateral side of the centrum, which is the best preserved, is excavated by a well-defined fossa (i.e., a “pleurocoel” of many previous works). This fossa is ovate in outline and extends anteroposteriorly for approximately three-fourths the preserved length of the centrum (Ml, 87.9 mm; Mh, 50 mm). Closely comparable fossae are associated with pneumatic invasion by the pulmonary system in many extant birds (O’Connor 2004).
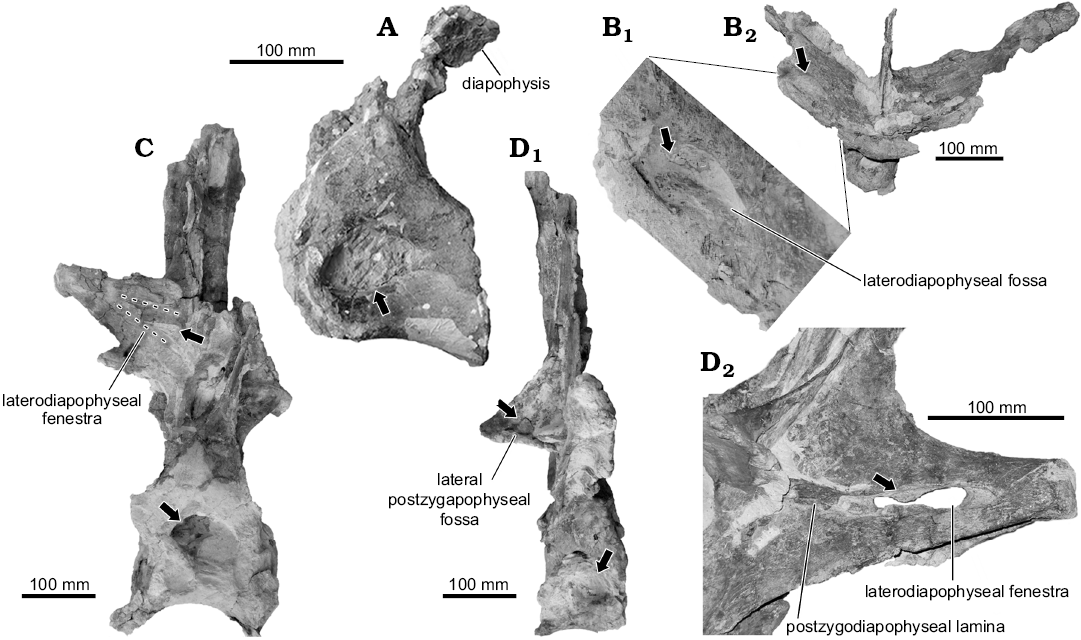
Fig. 2. Dorsal vertebrae of the rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. UNPSJB-PV 1007/13, partial anterior dorsal vertebra in left lateral view. B. UNPSJB-PV 1007/12, partial anterior to middle dorsal vertebra in dorsal view (B2), detail of right laterodiapophyseal fossa in dorsal view (B1). C. UNPSJB-PV 1007/4, middle or posterior dorsal vertebra in right lateral view (dashed lines indicate approximate margins of laterodiapophyseal fenestra). D. UNPSJB-PV 1007/5, middle or posterior dorsal vertebra in right lateral view (D1), detail of right laterodiapophyseal fenestra in right posterolateral view (D2). Arrows indicate hypothesized pneumatic structures.
Both transverse processes of the anterior to middle dorsal vertebra UNPSJB-PV 1007/12 exhibit a deep, well-defined ovoid fossa that penetrates to the ventral floor of the bone (the laterodiapophyseal fossa of Ibiricu et al. 2015). The right laterodiapophyseal fossa of this vertebra has a Ml of 52.1 mm and a Mh of 24.8 mm. Furthermore, as shown by CT imagery, much of the interior of the left diapophysis of vertebra UNPSJB-PV 1007/12 is hollow (Fig. 3B). A laterodiapophyseal fossa is also present on the preserved left transverse process of another of the recently described Katepensaurus anterior to middle dorsal vertebrae, UNPSJB-PV 1007/31. Nevertheless, in this latter vertebra, the orientation and shape of the fossa differ from those in UNPSJB-PV 1007/12. These differences may be related to the distinct positions of these vertebrae along the dorsal column, since UNPSJB-PV 1007/12 is thought to be situated slightly more posteriorly in the sequence. However, these distinctions might also be due to variation in the distribution and/or morphology of pneumatic diverticula and their associated osteological correlates independent of serial position. For instance, in the holotypic partial skeleton of the Tunisian rebbachisaurid Tataouinea hannibalis, the sacrum, caudal vertebrae, and ischia exhibit asymmetrical pneumatization, with the development of probable pneumatic features being more extensive on the left side of the animal than the right (Fanti et al. 2013, 2015). Moreover, in another sauropod group, saltasaurine titanosaurs, variation in the presence of fossae and foramina between specimens, vertebral sequences, and the left and right halves of individual vertebrae also appears to be related to the distribution of pneumatic diverticula and does not always vary systematically according to serial position (Zurriaguz and Álvarez 2014; Zurriaguz and Powell 2015). For example, the anterior caudal vertebrae of the saltasaurine Neuquensaurus australis exhibit well-defined but often asymmetrical lateral fossae that were probably associated with pneumatic diverticula in this derived titanosaur (note, however, that blind fossae such as these do not constitute unambiguous evidence for the presence of such diverticula; O’Connor 2006; Ibiricu et al. 2013b; LMI personal observations). Comparable bilateral variation in probable pneumatic structures has also been documented in the caudal vertebrae of other neosauropods, such as apatosaurine diplodocids and the brachiosaurid Giraffatitan brancai (Wedel and Taylor 2013).
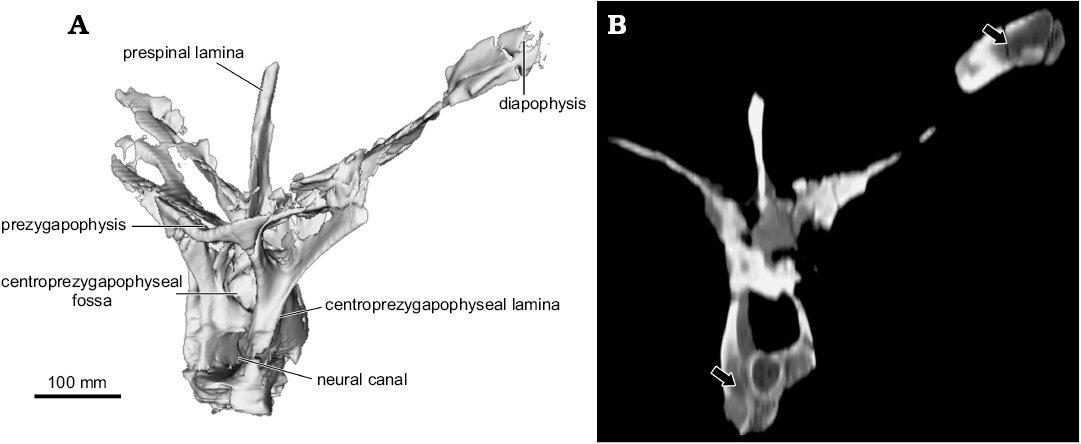
Fig. 3. Partial anterior to middle dorsal vertebra (UNPSJB-PV 1007/12) of the rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. Computed tomography (CT)-based digital reconstruction in left anterolateral view. B. Axial CT “slice” in anterior view, with hypothesized pneumatic structures indicated by arrows. Note internal cavity in left diapophysis.
The Katepensaurus holotype also includes three middle to posterior dorsal vertebrae (UNPSJB-PV 1007/4–6; Ibiricu et al. 2013a, 2015; Figs. 2C, D, 4, 5). In UNPSJB-PV 1007/4, the lateral sides of the centrum are invaded by large, well-defined, ovate fossae (“pleurocoels”) that extend anteroposteriorly for approximately half its length (Ml of the right fossa: 75.4 mm; Mh: 54.2 mm). The right fossa is relatively deep and internally partitioned by a thin, dorsoventrally-oriented lamina. Similarly, although it is smaller and shallower, an ovate lateral fossa is present in the centrum of UNPSJB-PV 1007/5 (see Ibiricu et al. 2013a).
Perhaps the most distinctive features of the middle to posterior dorsal vertebrae of Katepensaurus (UNPSJB-PV 1007/4–6) are the well-defined fenestrae in their transverse processes (the laterodiapophyseal fenestrae of Ibiricu et al. 2013a). These fenestrae pierce completely through the transverse processes. Although, at first glance, these structures might appear to be taphonomic artifacts, they certainly are not, as evidenced by their smooth, undamaged margins as well as their occurrence in all known middle to posterior dorsal vertebrae of the Katepensaurus holotype (see Ibiricu et al. 2013a). Neural arch perforations have occasionally been documented in other sauropod presacral vertebrae as well; for example, in the neural arch pedicels of dorsal vertebrae of the Spanish rebbachisaurid Demandasaurus darwini (Torcida Fernández-Baldor et al. 2011: fig. 9a, c), in the anterior centroparapophyseal laminae of dorsal vertebrae of Giraffatitan (Taylor 2009), and in cervical vertebral neural spines of the giant titanosauriform Sauroposeidon proteles (Wedel et al. 2000: figs. 6, 9). Nevertheless, among sauropods, fenestrae that penetrate through the dorsal vertebral transverse processes are presently unique to Katepensaurus. The right laterodiapophyseal fenestra of UNPSJB-PV 1007/5 has a Ml of 84.8 mm and a Mh of 24.4 mm. Interestingly, and as revealed by CT images, this fenestra is united directly to a pneumatic chamber within the distal (i.e., lateral) end of the transverse process, which is here termed the intradiapophyseal chamber (Fig. 4; see also Discussion). Most of the intradiapophyseal chamber is surrounded by solid but relatively thin cortical bone; however, trabecular bone is present at the distal end of the chamber. As observed in and reconstructed from CT images, the intradiapophyseal chamber extends to the distal end of the transverse process, which is eroded. This pneumatic structure is dorsoventrally taller laterally than it is medially, and exhibits a total internal Ml of 58.8 mm, a Mh of 23.7 mm, and a maximum anteroposterior width of 18.5 mm (Fig. 4). The estimated volume (from the distal [= lateral] end of the laterodiapophyseal fenestra to that of the transverse process, which in turn, represents the reconstructed air space) of the chamber is 20 534 mm³ (= 20.534 cm3).
In the vertebra UNPSJB-PV 1007/4, the posterior centroparapophyseal and posterior centrodiapophyseal laminae delimit the deep parapophyseal centrodiapophyseal fossa. This fossa is subdivided by two accessory laminae, the dorsal and ventral parapophyseal centrodiapophyseal fossa laminae of Ibiricu et al. (2013a). The lateral surfaces of the neural arch also exhibit deep, ovoid cavities, the postzygapophyseal centrodiapophyseal fossa and the parapophyseal centroprezygapophyseal fossa. In UNPSJB-PV 1007/5, there are also well-defined, rounded fossae placed lateral to the articular facets of the postzygapophyses (the lateral postzygapophyseal fossae of Ibiricu et al. 2013a). The right lateral postzygapophyseal fossa of UNPSJB-PV 1007/5 has a Ml of 31.2 mm and a Mh of 19.5 mm.
The middle to posterior dorsal vertebral neural spines display wide, bilateral fossae, the prezygapophyseal spinodiapophyseal and spinopostzygapophyseal lamina fossae. These shallow fossae exhibit flat “floors”. The lateral spinopostzygapophyseal lamina separates the comparatively mediolaterally narrow, dorsomedially positioned spinopostzygapophyseal lamina fossa from the wider, ventrolaterally situated postzygapophyseal spinodiapophyseal fossa (see Ibiricu et al. 2013a).
In sum, the dorsal vertebral sequence of Katepensaurus includes anterior to middle dorsals with laterodiapophyseal fossae (i.e., UNPSJB-PV 1007/12, 31) and middle to posterior dorsals with laterodiapophyseal fenestrae (UNPSJB-PV 1007/4–6). Stated another way, the anterior to middle dorsal vertebral transverse processes of Katepensaurus are excavated by fossae, whereas the more posterior dorsal transverse processes are pierced by fenestrae (see Ibiricu et al. 2015). Moreover, the system of fossae in the dorsal vertebral neural arches is well-developed.
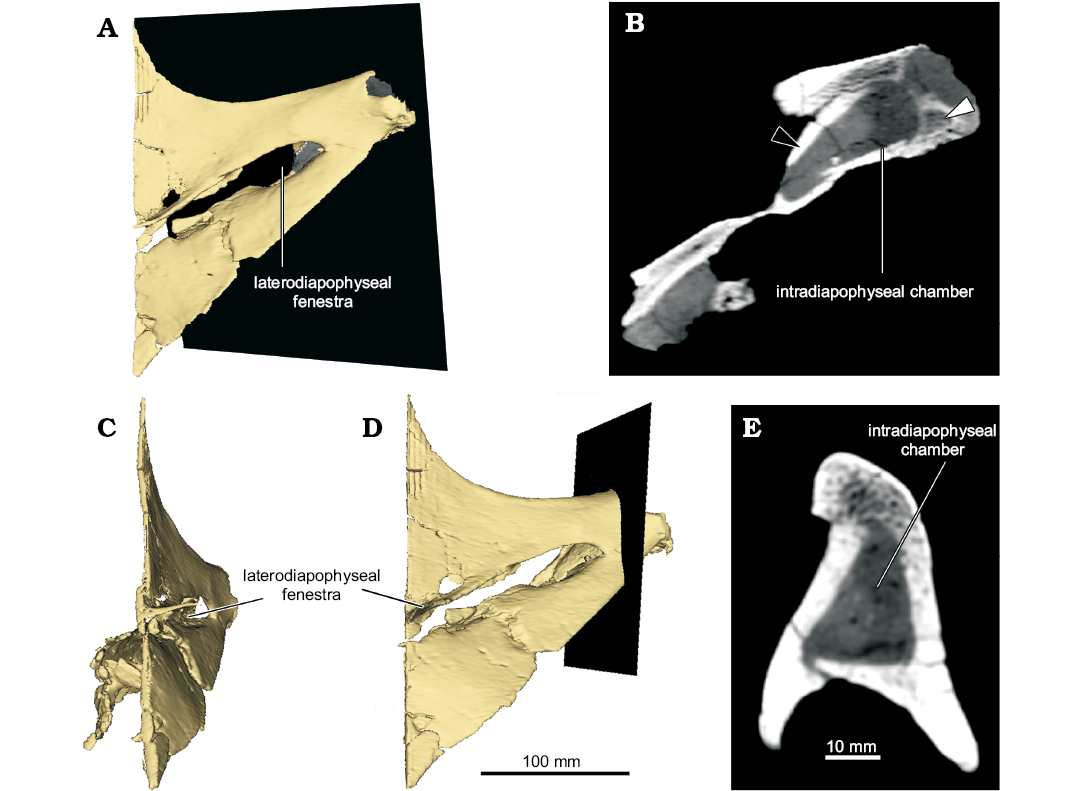
Fig. 4. Detail of the right transverse process of a middle or posterior dorsal vertebra (UNPSJB-PV 1007/5) of the rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. Computed tomography (CT)-based reconstruction of the transverse process and diapophysis in posterior view, with black rectangle indicating location of axial CT “slice” shown in B. B. Axial CT “slice” showing intradiapophyseal chamber, with black and white arrowheads indicating cortical and trabecular tissues, respectively. Reconstruction of the transverse process and diapophysis in medial (C) and posterior (D) views. Black rectangle indicates location of sagittal CT “slice” shown in E. E. Sagittal CT “slice” through intradiapophyseal chamber.
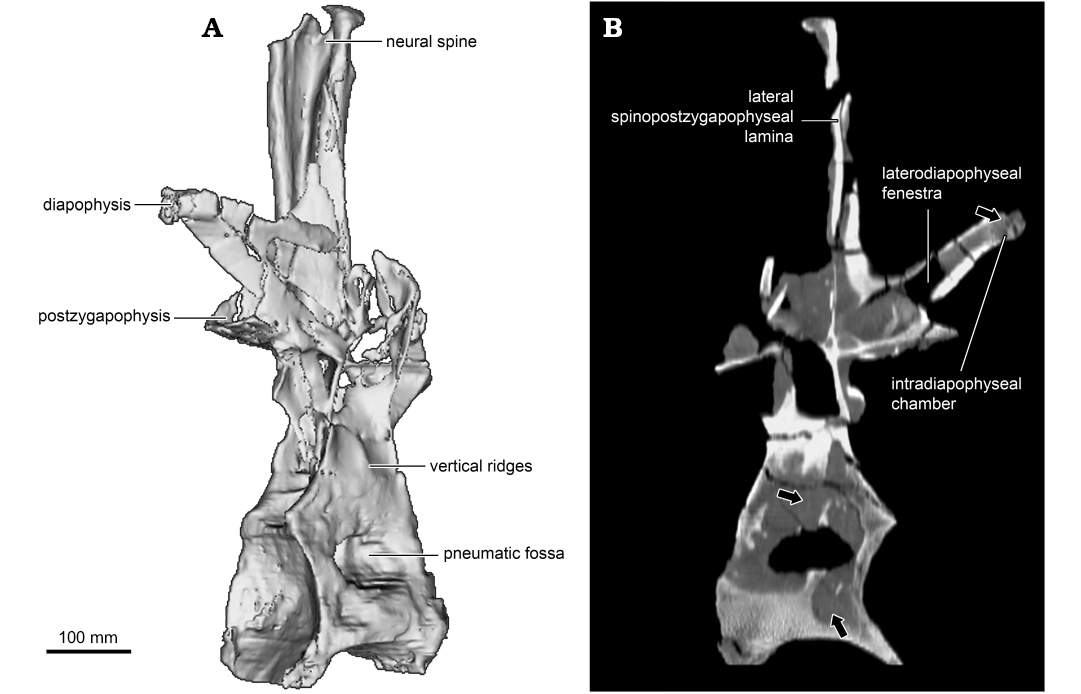
Fig. 5. Computed tomography-based visualizations of middle to posterior dorsal vertebra (UNPSJB-PV 1007/4) of the rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. Three-dimensional digital model in right posterolateral view. B. Sagittal section (anterior to left) with arrows indicating internal cavities hypothesized as pneumatic structures.
Histological context and ontogenetic assessment.—To assess the minimum ontogenetic age of the Katepensaurus type specimen, a transverse histological thin section was taken from a dorsal rib (UNPSJB-PV 1007/28). Although the ontogenetic stages of sauropod specimens have usually been deduced from long bone histology (e.g., Klein and Sander 2008), dorsal ribs have recently been demonstrated to be useful for at least minimum age estimation (Waskow and Sander 2014).
The rib has a relatively thick cortex that surrounds a small marrow cavity that is filled with cancellous bone (Fig. 6A). There is no evidence of pneumatic cavities. The perimedullary cortex gradually transforms into cancellous bone. The inner cortex is highly remodeled, exhibiting several generations of secondary osteons (Fig. 6B). This remodeling is clearly more pronounced on one side of the rib, in which secondary osteons reach the outer cortex. Primary bone consists of well-vascularized fibrolamellar bone tissue (Fig. 6C). Primary osteons are mostly longitudinally oriented (Fig. 6D). Osteocyte lacunae are rounded or slightly elongate. Sharpey’s fibers are mostly observed on both the anterior and posterior regions of the rib. Fibrolamellar bone is interrupted by growth marks (annuli and lines of arrested growth; i.e., LAGs) (Fig. 6E). Seven growth marks are evident from the innermost cortex to the periosteal surface. Spacing between concentric growth marks diminishes moderately towards the periphery. There are no significant changes in the intrinsic fiber orientation and the density of vascular spaces in the outer cortex.
Based on the number of growth marks, we interpret a minimum age of seven years for the Katepensaurus holotype. Furthermore, judging from the absence of a strong decrease in the spacing of the peripheral growth marks, we infer that the individual represented by this specimen died during an active phase of growth; in other words, that the Katepensaurus holotype was not fully grown when it died.
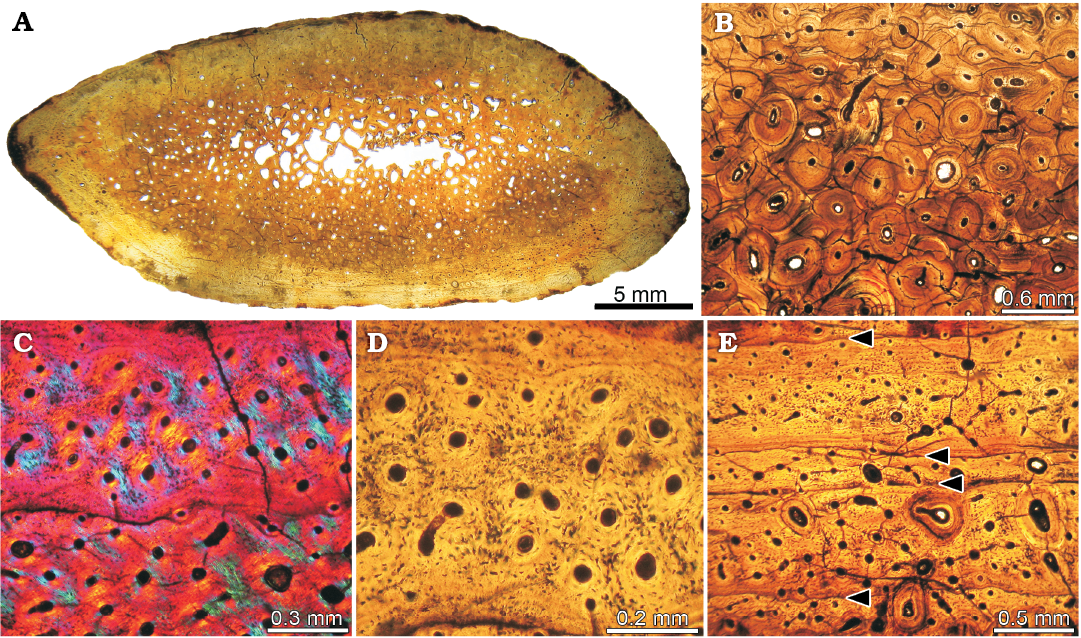
Fig. 6. Histology of a dorsal rib (UNPSJB-PV 1007/28) of the holotype of the rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. Complete cross section of the rib sampled for histology. B. Detail of the dense Haversian bone in the cortex. C. General view of fibrolamellar bone tissue. D. Detail of fibrolamellar bone tissue. Note the abundance of longitudinally oriented vascular spaces in C and D. E. Four of the seven growth marks preserved in the primary bone tissue (indicated by arrowheads).
Discussion
Reconstruction of rebbachisaurid pulmonary anatomy.—The axial skeleton of Katepensaurus displays a suite of hypothesized pneumatic features that are, in most respects, comparable to those seen in other rebbachisaurid taxa. For instance, the dorsal vertebral centra of this Patagonian form are excavated by well-defined lateral fossae (“pleurocoels”), a feature that is shared with many other sauropods, including all other rebbachisaurids for which this part of the skeleton has been described (i.e., Amazonsaurus maranhensis, Carvalho et al. 2003, Comahuesaurus windhauseni, Carballido et al. 2012, Demandasaurus darwini, Torcida Fernández-Baldor et al. 2011, Histriasaurus boscarollii, Dalla Vecchia 1999, Limaysaurus tessonei, Calvo and Salgado 1995, Nigersaurus taqueti, Sereno et al. 2007, Nopcsaspondylus alarconensis, Nopcsa 1902; Apesteguía 2007, Rebbachisaurus garasbae, Wilson and Allain 2015, and several unidentified or unnamed forms, e.g., MMCH-Pv 49, Haluza et al. 2012, MOZ-Pv 1240, Salgado et al. 2012). Moreover, in Katepensaurus, the general morphology of the dorsal neural arches and their associated complex of laminae and fossae closely resembles those of other rebbachisaurids such as Limaysaurus, Nopcsaspondylus, Rebbachisaurus, and especially MMCH-Pv 49 (Apesteguía et al. 2010; Haluza et al. 2012). These neural arch fossae are probably related to the presence of pneumatic diverticula associated with the dorsal column. Evidence consistent with this hypothesis is present in the form of numerous, irregular foramina and fossae in the middle to posterior dorsal neural arches of Rebbachisaurus that Wilson and Allain (2015) regarded as pneumatic in origin. Additionally, Torcida Fernández-Baldor et al. (2011) documented the occurrence of foramina that penetrate entirely through the dorsal neural arch pedicels of the Spanish rebbachisaurid Demandasaurus, and interpreted these as pneumatic structures as well. Torcida Fernández-Baldor et al. (2011) also reported evidence of pneumaticity in the area of the capitulotubercular web in dorsal ribs of Demandasaurus.
Katepensaurus offers additional, compelling evidence that the dorsal series of at least some rebbachisaurids was pneumatized by diverticula from the pulmonary system. As detailed above, the middle to posterior dorsal neural arches of this Patagonian taxon display features that have not been documented elsewhere within Sauropoda, most notably the laterodiapophyseal fenestrae and their associated intradiapophyseal chambers. As demonstrated by CT images, the right laterodiapophyseal fenestra of the middle to posterior dorsal vertebra UNPSJB-PV 1007/5 is united to the intradiapophyseal chamber, an internal cavity that extends to the distal (= lateral) end of the transverse process. The intradiapophyseal chamber is surrounded by solid cortical bone that terminates distally in trabecular bone (Fig. 4B). Based on the PP concept, O’Connor (2006) concluded that the only unambiguous indicators of PSP (i.e., pneumatic invasion of postcranial bone) in fossil taxa are fossae or foramina that are connected to large internal chambers, such as those seen in the middle to posterior dorsal vertebral transverse processes of Katepensaurus. We therefore interpret the laterodiapophyseal fenestra–intradiapophyseal chamber complex as an unambiguous indicator that the dorsal transverse processes of this rebbachisaurid were pneumatized. Selected non-avian theropods (e.g., the abelisaurid Majungasaurus crenatissimus; see O’Connor 2007: fig. 9b) and extant birds (e.g., the wood duck Aix sponsa; see Fajardo et al. 2007) exhibit a comparable condition, i.e., internal pneumatic chambers connected to foramina in the transverse processes of presacral vertebrae. Following from the patterns of PSP observed in birds (O’Connor and Claessens 2005; O’Connor 2006), in Katepensaurus, the laterodiapophyseal fenestrae and associated intradiapophyseal chambers of the middle to posterior dorsal vertebrae would have accommodated diverticula from the lungs and/or abdominal air sacs. Pneumatic invasion of the middle to posterior dorsal vertebrae by these pulmonary structures has also been hypothesized for other sauropods, such as the enigmatic neosauropod Haplocanthosaurus priscus (Wedel 2009) and the saltasaurine titanosaur Saltasaurus loricatus (Cerda et al. 2012).
Similarly, following from the PP concept, the laterodiapophyseal fossae present in the anterior to middle dorsal vertebral transverse processes of Katepensaurus may also be an indicator of pneumaticity. As demonstrated by CT imagery, much of the interior of the left diapophysis of the anterior to middle dorsal vertebra UNPSJB-PV 1007/12 is hollow, indicating the presence of an intradiapophyseal chamber (Fig. 3B). Unfortunately, the area that would have linked the left laterodiapophyseal fossa with the remainder of the transverse process is broken in this vertebra; therefore, it is not possible to definitively establish an internal connection between the fossa and chamber. However, assuming that, as seems likely, these fossae and chambers are indeed indicators of pneumatic invasion of the anterior to middle dorsal vertebrae of Katepensaurus, these vertebrae may have been pneumatized by diverticula from the lungs and/or cervical air sacs (as has also been hypothesized for this part of the skeleton in other sauropods, e.g., Haplocanthosaurus, Saltasaurus).
Although the focus of this study is the dorsal vertebrae, the holotype of Katepensaurus also includes cervical and caudal vertebrae. The cervical centra and neural arches of this rebbachisaurid are excavated by several deep, well-defined fossae (Fig. 7; see also Ibiricu et al. 2013a: figs. 3–5), as is also the case in all other rebbachisaurids for which cervical vertebrae have been described (i.e., Cathartesaura anaerobica, Gallina and Apesteguía 2005, Demandasaurus, Torcida Fernández-Baldor et al. 2011, Limaysaurus, Calvo and Salgado 1995, Nigersaurus, Sereno et al. 2007, Zapalasaurus bonapartei, Salgado et al. 2006, and unidentified or unnamed forms, e.g., MMCH-Pv 49, Haluza et al. 2012, MOZ-Pv 1248, Salgado et al. 2012, UNPSJB-PV 1005, Ibiricu et al. 2012). These fossae almost certainly represent pneumatic structures associated with the presence of diverticula from the cervical air sacs, as has been reconstructed for many other saurischians, including other sauropods and basal sauropodomorphs (e.g., Wedel 2006, 2007, 2009; Cerda et al. 2012; Yates et al. 2012).
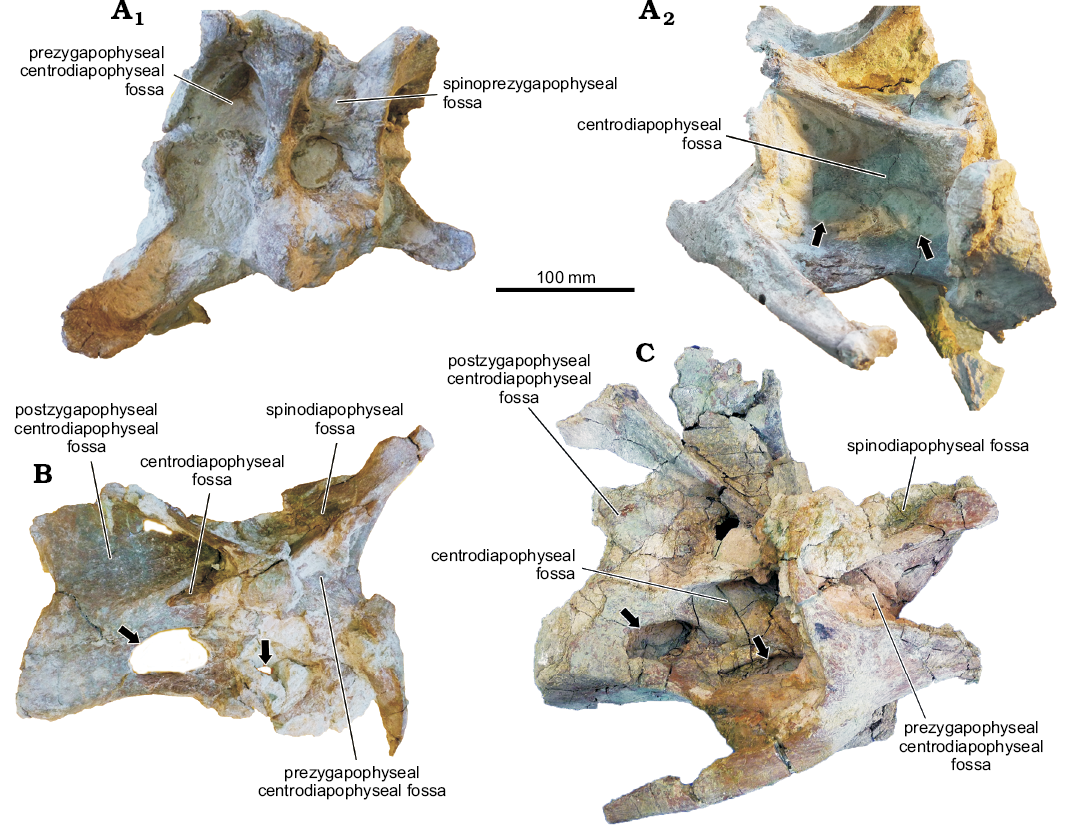
Fig. 7. Cervical vertebrae of rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. UNPSJB-PV 1007/1, anterior cervical vertebra in anterior (A1) and right ventrolateral (A2) views. B. UNPSJB-PV 1007/2, anterior cervical vertebra in right lateral view. C. UNPSJB-PV 1007/3, middle cervical vertebra in right lateral view. Lateral fossae of the centrum (hypothesized as pneumatic in origin) indicated by arrows.
The most complete anterior caudal vertebra of Katepensaurus (UNPSJB-PV 1007/7) exhibits a wing-like right transverse process, the anterior surface of which is excavated by a large prezygapophyseal centrodiapophyseal fossa. This fossa is divided by a well-developed lamina, the prezygapophyseal centrodiapophyseal fossa lamina (Fig. 8A; see also Ibiricu et al. 2013a). The caudal vertebral neural arches are also excavated by several fossae that differ in their degrees of development (e.g., the spinodiapophyseal and spinoprezygapophyseal fossae). Unfortunately, however, the caudal centra of this rebbachisaurid are all missing or incomplete; therefore, it is not possible to observe any potentially pneumatic features in these elements. Consequently, the evidence for pneumaticity in the caudal series of Katepensaurus is not as robust as it is for the dorsal vertebrae.
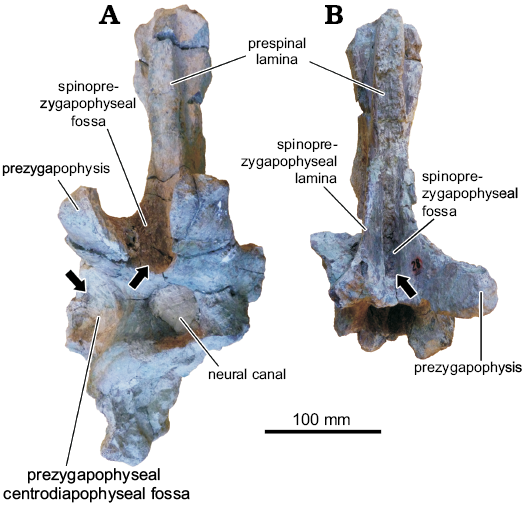
Fig. 8. Anterior view of anterior caudal vertebrae of rebbachisaurid sauropod Katepensaurus goicoecheai Ibiricu, Casal, Martínez, Lamanna, Luna, and Salgado, 2013a from the Cenomanian–Turonian Bajo Barreal Formation of Chubut Province, Argentina. A. UNPSJB-PV 1007/7. B. UNPSJB-PV 1007/8. Hypothesized pneumatic fossae indicated by arrows.
Nevertheless, other rebbachisaurid taxa provide more convincing evidence for the invasion of the tail skeleton by pneumatic diverticula. In the holotypic specimen of Tataouinea, for example, the first 17 caudal vertebrae were preserved in articulation (Fanti et al. 2015). Of these, the centra of caudals 1–6 exhibit lateral foramina (i.e., “pleurocoels”), whereas those of vertebrae 14–16 possess well-defined lateral fossae. Interestingly, the centra of vertebrae 7–13 lack lateral foramina or fossae (see Fanti et al. 2013, 2015). Fanti et al. (2015) interpreted this suite of features as evidence of pneumatic invasion of the most anterior and several of the middle caudal centra, interrupted by a pneumatic hiatus in the centra of vertebrae 7–13. Comparable lateral foramina are present in anterior caudal centra of Comahuesaurus (Carballido et al. 2012: fig. 7a, b) and in UNPSJB-PV 580, a rebbachisaurid caudal vertebra from the Bajo Barreal Formation that probably represents the first or second in the sequence (Ibiricu et al. 2012); given its nearly identical provenance, it is possible that this latter element may be referable to Katepensaurus, though this is presently unclear (Ibiricu et al. 2013a, 2015). Probable pneumatic excavations have been reported in the caudal vertebrae of other rebbachisaurid specimens as well; for instance, Torcida Fernández-Baldor et al. (2011: fig. 11a) identified the prezygapophyseal centrodiapophyseal fossa of an anterior caudal of Demandasaurus as a “pneumatic depression”, whereas Medeiros and Schultz (2004) and Mannion and Barrett (2013) noted lateral fossae in the centra of isolated rebbachisaurid anterior caudals from the mid-Cretaceous of Brazil and Morocco, respectively. As such, and based on the morphological patterns observed in extant birds, the caudal vertebrae of rebbachisaurids may well have been pneumatized by the posterior diverticula of the abdominal air sacs, as has also been proposed for Haplocanthosaurus (Wedel 2009), diplodocids and brachiosaurids (Wedel and Taylor 2013), and saltasaurine titanosaurs (Cerda et al. 2012), among other sauropods. Within Rebbachisauridae, at least Rebbachisaurus and Tataouinea display evidence of sacral pneumaticity as well (see Fanti et al. 2013, 2015; Wilson and Allain 2015).
Compelling evidence for the presence of abdominal air sacs in Rebbachisauridae occurs in the form of probable pneumatic structures in the pelves of selected members of the clade. Fanti et al. (2013, 2015) documented the existence of internal cavities in the pelvic elements of Tataouinea, specifically in several regions of the ilium (the preacetabular process, iliac blade, and pubic peduncle) and in most or all of the preserved parts of the ischium. These ischial cavities constitute a particularly persuasive indicator of PSP in that they are clearly connected to an external foramen (on the lateral side of the iliac peduncle of the ischium), thus satisfying all of the criteria specified in the Pneumaticity Profile of O’Connor (2006). Fanti et al. (2015: 39) also reported ischial pneumaticity in Rebbachisaurus, but this was not mentioned by Wilson and Allain (2015) in their detailed redescription of this taxon, so the condition in this animal remains unclear. Lastly, Carvalho et al. (2003) reported a pneumatic chamber in the region of the ischial peduncle of the ilium of Amazonsaurus. Significantly, most recent phylogenetic analyses that have included this taxon (e.g., Carballido et al. 2012; Mannion et al. 2012; Fanti et al. 2015; Ibiricu et al. 2015; Wilson and Allain 2015) have recovered it as the earliest-diverging rebbachisaurid, suggesting that pelvic pneumaticity may be synapomorphic for, and therefore probably widespread within, this sauropod clade. In extant birds, only the abdominal air sacs pneumatize pelvic elements; consequently, the occurrence of pneumatic ilia and ischia in at least some rebbachisaurids strongly suggests that abdominal air sacs were present in these dinosaurs.
Taking all of the above into account, it is possible to generate a hypothetical reconstruction of the rebbachisaurid pulmonary system (Fig. 9A). In addition to lungs, there is convincing evidence of the existence of cervical and abdominal air sacs in these sauropods, with the most compelling indicators of the latter being the pneumatized caudal and pelvic elements of selected genera (e.g., Tataouinea, probably Amazonsaurus). Cervical and abdominal air sacs have also been hypothesized in a variety of other neosauropods, such as Haplocanthosaurus (Wedel 2009), diplodocids and brachiosaurids (Wedel and Taylor 2013), and saltasaurine titanosaurs (Cerda et al. 2012). The existence of these air sacs may therefore represent the plesiomorphic condition for Neosauropoda, and indeed (given the probable occurrence of these structures in theropod dinosaurs) possibly a much more inclusive clade (e.g., Saurischia) as well. There is, however, no evidence of clavicular air sacs in rebbachisaurids, in contrast to what has been observed in saltasaurines (Cerda et al. 2012).
It is interesting to note that, in several rebbachisaurid taxa, there are indications of an extraordinary degree of invasion of postcranial skeletal elements by pneumatic diverticula relative to the conditions in many other sauropods. This appears particularly pronounced in the thoracic and abdominal regions: for instance, uniquely among sauropods, Katepensaurus has laterodiapophyseal fossae/fenestrae and intradiapophyseal chambers in the dorsal vertebral transverse processes, Demandasaurus has presumably pneumatic foramina that pass through the middle to posterior dorsal neural arch pedicels (Torcida Fernández-Baldor et al. 2011), and Rebbachisaurus has an abundance of pneumatic foramina in the laminae of the middle to posterior dorsal neural spines (Wilson and Allain 2015). Further, Amazonsaurus, Tataouinea, and possibly Rebbachisaurus show evidence of pneumatized pelvic elements, a condition that, in sauropods, is otherwise seen only in a few somphospondylan titanosauriforms (see Cerda et al. 2012; Lamanna and Hasegawa 2014); pneumatic ischia are not definitively known elsewhere within Sauropoda. The potential implications of this extensive PSP are discussed below.
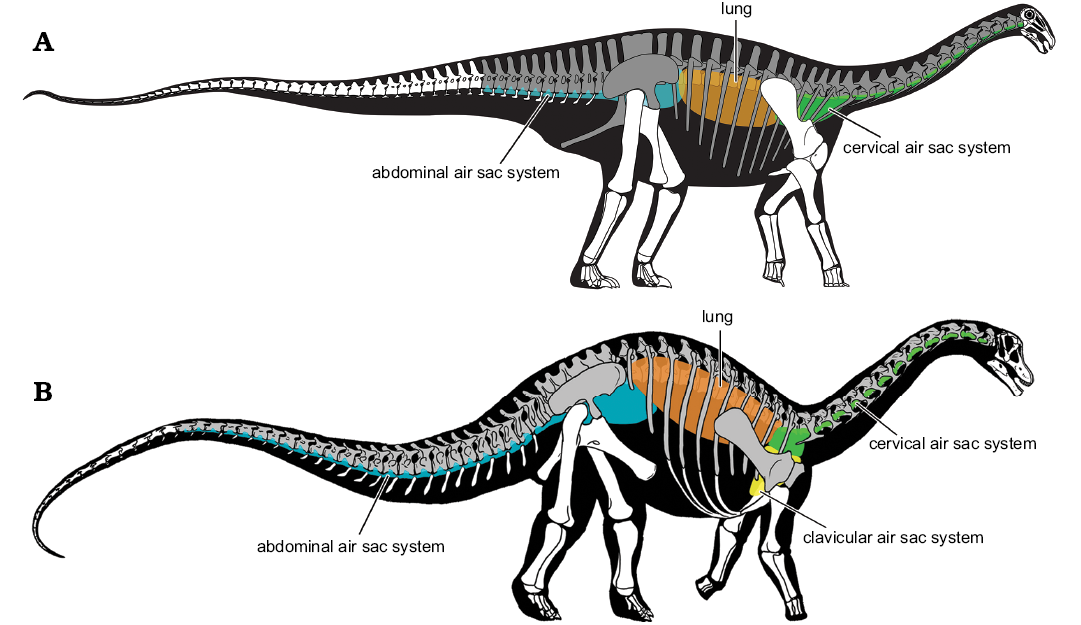
Fig. 9. Comparison of the reconstructed pulmonary systems of the sauropod clades Rebbachisauridae (Diplodocoidea) and Saltasaurinae (Titanosauria) in right lateral view. A. Reconstructed pulmonary anatomy of a generalized rebbachisaurid. Skeletal reconstruction based largely on Nigersaurus taqueti (Sereno et al. 2007: fig. 3a); pulmonary anatomy based on data presented herein, i.e., hypothesized osteological correlates of pneumaticity described in multiple rebbachisaurids, but primarily Katepensaurus goicoecheai (cervical and dorsal pneumaticity) and Tataouinea hannibalis (sacral, caudal, and pelvic pneumaticity). B. Reconstructed pulmonary anatomy of a saltasaurine (after Cerda et al. 2012: fig. 4a). Color coding of pulmonary structures is as follows: orange, lung; green, cervical air sac system; yellow, clavicular air sac system; blue, abdominal air sac system. Postcranial skeletal elements that are known to have been pneumatized are shown in gray; bones that were either apneumatic or for which this condition is uncertain are in white.
Implications for rebbachisaurid paleobiology.—Although deciphering the paleobiological implications of the extensive PSP seen in rebbachisaurids is necessarily speculative, some tentative inferences may be made by examining the significance of postcranial pneumaticity in the closest extant relatives of sauropods, modern birds. In birds, PSP lessens the density of bones by filling their internal cavities with air rather than marrow. By lightening the skeleton in this way, PSP reduces the muscle energy required to power the body, thereby decreasing metabolic energy consumption, which may in turn be allocated to other purposes such as locomotion and foraging (O’Connor 2009). It therefore seems reasonable to infer that the extensive PSP seen in Katepensaurus and other rebbachisaurids may have provided similar energetic benefits to these sauropods. Since sauropods were obviously nonvolant, the selective pressure to reduce skeletal density may have been minimal in comparison to this same pressure in flying birds. From an energetics standpoint, however, the capacity to pneumatize parts of the skeleton may have been beneficial even to large-bodied, ground-dwelling animals such as rebbachisaurids (O’Connor 2009). Furthermore, as it is well-known that the contraction of vertebrate skeletal muscle produces heat (e.g., Huxley 1974), decreasing the amount of muscle energy required to support and move the body may have also reduced the amount of heat generated during these activities. This may in turn have provided additional benefit to rebbachisaurids, in that many, if not all, of these animals inhabited exceedingly warm paleoenvironments. For example, the northern South American rebbachisaurid Amazonsaurus and the North African Rebbachisaurus and Tataouinea lived at tropical to subtropical paleolatitudes during the Albian–Cenomanian (see paleogeographic reconstructions of these areas in Blakey 2008 and Seton et al. 2012), when global climates were exceptionally warm (Huber et al. 2002; Holz 2015); consequently, these sauropods would presumably have been routinely subjected to extraordinarily high temperatures (Fig. 10). Katepensaurus, though it comes from a higher paleolatitude, lived at approximately the Cenomanian–Turonian boundary—the time of the Cretaceous Thermal Maximum, one of the hottest intervals of the entire Phanerozoic (Clarke and Jenkyns 1999; Huber et al. 2002; Poulsen et al. 2003)—and therefore would likely have experienced greatly elevated temperatures as well.
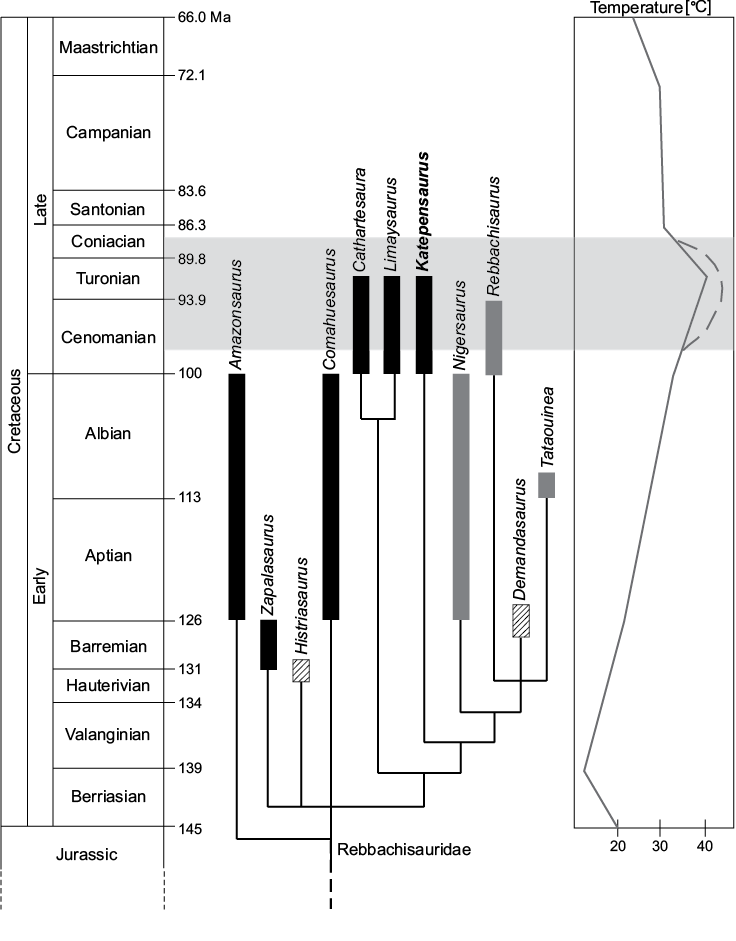
Fig. 10. Time-calibrated phylogeny of Rebbachisauridae and the relationship of rebbachisaurid evolution to Mesozoic geography and temperature. Genera with hatched stratigraphic ranges are European (although Histriasaurus, from Croatia, is more appropriately regarded as an African taxon; see Ibiricu et al. 2015), genera in solid black are South American, and genera in gray are African. Light gray band indicates the Cretaceous Thermal Maximum (KTR); note the existence of multiple rebbachisaurid genera during this interval. Geologic time scale after Walker et al. (2012); phylogenetic topology and stratigraphic ranges of taxa after Fanti et al. (2015: fig. 21b); paleotemperature curve based on and simplified from Holz (2015: appendix 1); curved, dashed line at KTR based on Zhou et al. 2012.
The hypothesis that Katepensaurus frequently had to contend with extreme warmth is supported by reconstructions of its paleoenvironment. The holotype was recovered from an exposure of the Cenomanian–Turonian Bajo Barreal Formation on the Estancia Laguna Palacios in the Golfo San Jorge Basin of Chubut Province, Argentina. The Bajo Barreal Formation is interpreted as having been deposited by a low-sinuosity braided fluvial system (Rodríguez 1992; Figari et al. 1999; Umazano et al. 2008, 2012; Casal et al. 2013) and is notable for its abundance of paleosols. Since arid climatic conditions inhibit the formation of paleosols (Retallack 1988), these data suggest the existence of a humid paleoclimate in this region during the early Late Cretaceous (see Casal et al. 2016). Plant fossils provide additional evidence that the Bajo Barreal Formation was deposited under a warm, humid paleoclimatic setting. Based on their study of fossil wood (pertaining to Araucariaceae, probably Agathoxylon sp.) from this unit, Pujana et al. (2007) proposed that the paleoclimate of central Patagonia during the early Late Cretaceous was characterized by an absence of seasonality. This inference is supported by the weakly defined boundaries of the growth rings in this wood, a feature that is associated with warm temperate to subtropical climates. Moreover, based on their studies of palynological assemblages, Archangelsky et al. (1994, 2009) and Barreda and Archangelsky (2006) also suggested the existence of a warm, humid paleoclimate in the Golfo San Jorge Basin during the Cenomanian.
Clearly, at least some rebbachisaurids habitually had to endure unusually hot temperatures. Although even the larger-bodied members of the clade were small compared to most other sauropods (e.g., the holotype of Rebbachisaurus is estimated at 7915–12 015 kg in body mass, Wilson and Allain 2015; that of Tataouinea was roughly 12 m long in life, Fanti et al. 2015), these were still very large (approx. elephant-sized) animals that had to function in what were presumably extraordinarily warm climates, and that therefore may have been susceptible to heat stress. Within this context, the extensive PSP of Katepensaurus and other rebbachisaurids may be plausibly, if cautiously, interpreted, at least in part, as an adaptive response to the exceptional warmth of the mid-Cretaceous. Minimizing the density of the skeleton through increased invasion of postcranial bones by air sac diverticula may have reduced the muscle power required to move the body and the heat energy produced in so doing. This may in turn have enabled these dinosaurs to better cope with exceptionally hot conditions. Bearing this in mind, it is perhaps not coincidental that several of the rebbachisaurids that exhibit the most extensive PSP yet documented within the clade—Amazonsaurus, Rebbachisaurus, and Tataouinea—inhabited equatorial to near-equatorial habitats during the extreme warmth of the mid-Cretaceous.
Nevertheless, it is important to note that other sauropod taxa—that often exhibit greater overall body sizes and possibly lower “grades” of PSP—lived in the same paleoenvironments. For example, Katepensaurus shared its paleoecosystem with comparably-sized titanosaurs such as Sarmientosaurus musacchioi (which was recovered from a near-equivalent horizon of the Bajo Barreal Formation at the same locality, the Estancia Laguna Palacios; see Martínez et al. 2016) and possibly Epachthosaurus sciuttoi, which comes from the same member of the same formation but from a different locality approximately 40 km to the southwest (Martínez et al. 2004). Similarly, isolated material that indicates the presence of very large-bodied titanosauriforms has been reported from the “Kem Kem beds” of Morocco (e.g., Lamanna and Hasegawa 2014; Ibrahim et al. 2016), the stratigraphic unit that yielded Rebbachisaurus. To conclude, although this hypothesis is weakly supported, we tentatively suggest that the extensive PSP seen in Katepensaurus and other rebbachisaurids may represent an adaptation for lightening the skeleton and lowering the body mass in response to the extremely warm climates of the mid-Cretaceous.
Extensive pneumaticity in rebbachisaurids: convergence with saltasaurine titanosaurs.—As documented above, rebbachisaurid diplodocoids such as Katepensaurus and Tataouinea exhibit manifestations of postcranial skeletal pneumaticity that are otherwise unknown within Sauropoda. Similarly, Cerda et al. (2012) and Zurriaguz and Cerda (2017) described the occurrence of elevated (“extreme”) PSP in another neosauropod clade, saltasaurine titanosaurs (i.e., Neuquensaurus australis, Rocasaurus muniozi, and Saltasaurus loricatus; Fig. 9B), although the patterns of pneumaticity in rebbachisaurids and saltasaurines differ significantly. For instance, pneumatic pectoral girdle elements are presently known in saltasaurines but not rebbachisaurids, whereas a pneumatized ischium is known in at least one rebbachisaurid (Tataouinea) but not in saltasaurines. Furthermore, like other titanosaurs, saltasaurines primarily exhibit camellate and semicamellate pneumatic bone tissue rather than the camerate tissue that predominates in rebbachisaurids and other diplodocoids.
Since, as noted above, PSP is widespread in Sauropodomorpha and other ornithodiran archosaur clades, it is clear that the common ancestor of Rebbachisauridae and Saltasaurinae would have possessed a pneumatized postcranial skeleton; in other words, that PSP is a plesiomorphic condition shared by these diplodocoid and titanosaurian groups. Nevertheless, both rebbachisaurids and saltasaurines exhibit a “grade” or degree of PSP that appears to be elevated in comparison to that of their closest relatives (Fig. 9). For example, as detailed above, the dorsal vertebrae of Katepensaurus possess laterodiapophyseal fossae/fenestrae and intradiapophyseal chambers, and Tataouinea and probably Amazonsaurus have pneumatic pelvic elements, characters that are otherwise unknown in Diplodocoidea. Similarly, saltasaurines are the only titanosauriforms known to possess pneumatic posterior caudal vertebrae and pectoral elements. Evidently, then, the elevated degree of PSP seen in rebbachisaurids and saltasaurines (relative to other diplodocoids and titanosaurs, respectively) was convergently acquired.
Rebbachisaurids are the latest-surviving representatives of Diplodocoidea, having persisted until the mid-Cretaceous (Cenomanian–Turonian; Ibiricu et al. 2013a). Saltasaurines are also among the latest-surviving titanosaurs, with fossils having been recovered from deposits as recent as latest Cretaceous (Campanian–Maastrichtian) in age (Cerda et al. 2012; Zurriaguz and Cerda 2017), though other titanosaurian lineages persisted into the latest Cretaceous as well (e.g., Curry Rogers 2009; Lacovara et al. 2014). As such, the unusually well-developed postcranial skeletal pneumaticity of rebbachisaurids and saltasaurines—which in turn implies the presence of an extensive system of air sacs and pneumatic diverticula—may have provided some form of competitive advantage to these sauropods over many of their relatives. Elevated PSP may therefore have played an important role in the course of rebbachisaurid and saltasaurine evolution.
Conclusions
The type specimen of the Patagonian rebbachisaurid sauropod dinosaur Katepensaurus goicoecheai pertained to an individual with a minimum ontogenetic age of seven years at the time of its death. The dorsal vertebrae of Katepensaurus display a pneumatic configuration that has not been documented previously among sauropods, the laterodiapophyseal fenestrae and their associated intradiapophyseal chambers. This character complex constitutes an unambiguous indicator that, in this rebbachisaurid, the middle to posterior dorsal vertebral transverse processes were internally pneumatized. Within Sauropoda, this manifestation of postcranial skeletal pneumaticity is presently unique to Katepensaurus, and it is herein interpreted as an adaptation for minimizing the density of the skeleton.
Katepensaurus inhabited a warm, humid paleoenvironment at or near the Cretaceous Thermal Maximum, which was one of the hottest temporal intervals of the Phanerozoic. Interestingly, several other rebbachisaurids that also exhibit extensive PSP (e.g., Amazonsaurus maranhensis, Rebbachisaurus garasbae, Tataouinea hannibalis) lived at tropical to subtropical paleolatitudes during the mid-Cretaceous, and therefore would also have had to contend with extraordinarily warm conditions. We therefore tentatively propose that the decreased skeletal density afforded by the well-developed PSP of rebbachisaurids would have lowered the amount of energy required to move the bodies of these animals and the heat generated in so doing. As such, increased PSP may have enabled rebbachisaurids to better cope with extremely warm climatic conditions. Though PSP was undoubtedly widespread among sauropods and other ornithodirans, rebbachisaurid diplodocoids and saltasaurine titanosaurs independently evolved an elevated degree of PSP relative to other representatives of Diplodocoidea and Macronaria, respectively. Extensive PSP may therefore have contributed to the evolutionary success of rebbachisaurids and saltasaurines, both of which were among the latest-surviving representatives of their respective neosauropod clades.
Acknowledgments
We thank Marcelo Luna and Edmundo Ivany (both UNPSJB-PV) for their invaluable assistance in the field, particularly with the collection of the Katepensaurus type specimen. We also thank Marcelo Luna for photographing the fossils. We are grateful to the Goicoechea family (owners of the Estancia Laguna Palacios) for their more than two decades of support to the UNPSJB-PV. We also thank Juan Iturra, Juan Vargas, Guillermo McDonald, Lucas Penayo, Ezequiel Gonzalez, Vanesa Mansilla, Muriel Vilardo, and Marcelo Krause (all UNPSJB-PV), Mandi Lyon (Carnegie Museum of Natural History, Pittsburgh, USA), Matt Herne (The University of Queensland, Brisbane, Australia), Phil Fraley (Phil Fraley Productions, Pittsburgh, USA), Craig Cutler (Craig Cutler Studio, New York, USA), Daniel Zvereff (New York, USA), and residents of the village of Buen Pasto (especially Humberto Hourcade) for support in various field seasons. We also thank Martin Digorado, Marisol Aguirre, and Marcelo Fellner (all Sanatorio Asociación Española S.M., Comodoro Rivadavia) and Alejandro Panes (Instituto del Diagnóstico del Este del Chubut, Puerto Madryn, Argentina) for access to CT scanners. Andrew McAfee (Carnegie Museum of Natural History, Pittsburgh, USA) provided considerable assistance with the figures, especially Fig. 9A. We acknowledge Patrick O’Connor (Ohio University, Athens, USA) and Andrew Smith (The Natural History Museum, London, UK) for informative discussions that significantly improved an early version of the manuscript. A later version of the manuscript was substantially improved by reviews from José Carballido (Museo Paleontológico Egidio Feruglio, Trelew, Argentina) and Mathew Wedel (Western University of Health Sciences, Pomona, USA) and by editorial comments from Stephen Brusatte (University of Edinburgh, UK). IAC thanks J. Bar, Alexandra Elbakyan, and the Wikipaleo group for access to relevant literature. This work was supported by The Jurassic Foundation (grant to LMI in 2011, which supported several collecting visits to the Katepensaurus quarry), the Agencia Nacional de Promoción Científica y Tecnológica (PICT 2012-2884 to LMI; PICT 2015-1021 to IAC), and Carnegie Museum of Natural History (to MCL).
References
Apesteguía, S. 2005. Evolution of the hyposphene-hypantrum complex within Sauropoda. In: V. Tidwell and K. Carpenter (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs, 248–267. Indiana University Press, Bloomington.
Apesteguía, S. 2007. The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). Gondwana Research 12: 533–546. Crossref
Apesteguía, S., Gallina, P.A., and Haluza, A. 2010. Not just a pretty face: anatomical peculiarities in the postcranium of rebbachisaurids (Sauropoda: Diplodocoidea). Historical Biology 22: 165–174. Crossref
Archangelsky, S., Barreda, V., Passalia, M.G., Gandolfo, M., Prámparo, M., Romero, E., Cúneo, R., Zamuner, A., Iglesias, A., Llorens, M., Puebla, G.G., Quattrocchio, M., and Volkheimer, W. 2009. Early angiosperm diversification: evidence from southern South America. Cretaceous Research 30: 1073–1082. Crossref
Archangelsky, S., Bellosi, E.S., Jalfin, G.A., and Perrot, C. 1994. Palynology and alluvial facies from the mid-Cretaceous of Patagonia, subsurface of San Jorge Basin, Argentina. Cretaceous Research 15: 127–142. Crossref
Barreda, V., and Archangelsky, S. 2006. The southernmost record of tropical pollen grains in the mid-Cretaceous of Patagonia, Argentina. Cretaceous Research 27: 778–787. Crossref
Benson, R.B.J., Butler, R.J., Carrano, M.T., and O’Connor, P.M. 2012. Air-filled postcranial bones in theropod dinosaurs: physiological implications and the “reptile”–bird transition. Biological Reviews 87: 168–193. Crossref
Blakey, R.C. 2008. Gondwana paleogeography from assembly to breakup—a 500 my odyssey. Geological Society of America Special Paper 441: 1–28.
Britt, B.B. 1997. Postcranial pneumaticity. In: P.J. Currie and K. Padian (eds.), Encyclopedia of Dinosaurs, 590–593. Academic Press, San Diego.
Butler, R.J., Barrett, P.M., and Gower, D.J. 2012. Reassessment of the evidence for postcranial skeletal pneumaticity in Triassic archosaurs, and the early evolution of the avian respiratory system. PLoS ONE 7: e34094. Crossref
Calvo, J.O., and Salgado, L. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian–Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia 11: 13–33.
Carballido, J.L., Salgado, L., Pol, D., Canudo, J.I., and Garrido, A.C. 2012. A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin: evolution and biogeography of the group. Historical Biology 24: 631–654. Crossref
Carvalho, I.S., Avilla, L.S., and Salgado, L. 2003. Amazonsaurus maranhensis gen. et sp. nov. (Sauropoda, Diplodocoidea) from the Lower Cretaceous (Aptian–Albian) of Brazil. Cretaceous Research 24: 697–713. Crossref
Casal, G.A., Martínez, R.D., Ibiricu, L.M., González Riga, B., and Foix, N. 2013. Tafonomía del dinosaurio terópodo Aniksosaurus darwini, Formación Bajo Barreal, Cretácico Tardío de Patagonia (Argentina). Ameghiniana 50: 571–592. Crossref
Casal, G.A., Martínez, R.D., Luna, M., and Ibiricu, L.M. 2016. Ordenamiento y caracterización faunística del Cretácico Superior del Grupo Chubut, Cuenca del Golfo San Jorge, Argentina. Revista Brasileira de Paleontologia 19: 53–70. Crossref
Cerda, I.A., Salgado, L., and Powell, J.E. 2012. Extreme postcranial pneumaticity in sauropod dinosaurs from South America. Paläontologische Zeitschrift 86: 441–449. Crossref
Chinsamy, A., and Raath, M.A. 1992. Preparation of fossil bone for histological examination. Palaeontologia Africana 29: 39–44.
Chinsamy-Turan, A. 2005. The Microstructure of Dinosaur Bone: Deciphering Biology with Fine-Scale Techniques. 216 pp. Johns Hopkins University Press, Baltimore.
Claessens, L.P.A.M., O’Connor, P.M., and Unwin, D.M. 2009. Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism. PLoS ONE 4: e4497. Crossref
Clarke, L.J., and Jenkyns, H.C. 1999. New oxygen isotope evidence for long-term Cretaceous climatic change in the Southern Hemisphere. Geology 27: 699–702. Crossref
Curry Rogers, K. 2009. The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 29: 1046–1086. Crossref
Dalla Vecchia, F.M. 1999. Atlas of the sauropod bones from the upper Hauterivian-lower Barremian of Bale/Valle (SW Istria, Croatia). Natura Nacosta 18: 6–41.
Duncker, H.R. 1972. Structure of avian lungs. Respiration Physiology 14: 44–63. Crossref
Fajardo, R.J., Hernandez, E., and O’Connor, P.M. 2007. Postcranial skeletal pneumaticity: a case study in the use of quantitative microCT to assess vertebral structure in birds. Journal of Anatomy 211: 138–147. Crossref
Fanti, F., Cau, A., Cantelli, L., Hassine, M., and Auditore, M. 2015. New information on Tataouinea hannibalis from the Early Cretaceous of Tunisia and implications for the tempo and mode of rebbachisaurid sauropod evolution. PLoS ONE 10: e0123475. Crossref
Fanti, F., Cau, A., Hassine, M., and Contessi, M. 2013. A new sauropod dinosaur from the Early Cretaceous of Tunisia with extreme avian-like pneumatization. Nature Communications 4: 2080. Crossref
Figari, E.G., Strelkov, E., Laffitte, G., Cid de La Paz, M.S., Courtade, S.F., Celaya, J., Vottero, A., Lafourcade, P., Martínez, R., and Villar, H.J. 1999. Los sistemas petroleros de la Cuenca del Golfo San Jorge: síntesis estructural, estratigráfica y geoquímica. In: G.A. Chebli (ed.), 4º Congreso de Exploración y Desarrollo de Hidrocarburos, Actas, 197–237. Mar del Plata.
Francillon-Vieillot, H., De Buffrénil, V., Castanet, J., Géraudie, J., Meunier, F.J., Sire, J.Y., Zylberberg, L., and De Ricqlès, A. 1990. Microstructure and mineralization of vertebrate skeletal tissues. In: J.G. Carter (ed.), Skeletal Biomineralization: Patterns, Processes, and Evolutionary Trends, 175–234. American Geophysical Union, Washington.
Gallina, P.A. and Apesteguía, S. 2005. Cathartesaura anaerobica gen. et sp. nov., a new rebbachisaurid (Dinosauria, Sauropoda) from the Huincul Formation (Upper Cretaceous), Río Negro, Argentina. Revista del Museo Argentino de Ciencias Naturales 7: 153–166. Crossref
Haluza, A., Canale, J.I., Otero, A., Pérez, L.M., and Scanferla, C.A. 2012. Changes in vertebral laminae across the cervicodorsal transition of a well-preserved rebbachisaurid (Dinosauria, Sauropoda) from the Cenomanian of Patagonia. Journal of Vertebrate Paleontology 32: 219–224. Crossref
Henderson, D.M. 2004. Tipsy punters: sauropod dinosaur pneumaticity, buoyancy and aquatic habits. Proceedings of the Royal Society of London B 271: S180–S183. Crossref
Holz, M. 2015. Mesozoic paleogeography and paleoclimates—a discussion of the diverse greenhouse and hothouse conditions of an alien world. Journal of South American Earth Sciences 61: 91–107. Crossref
Huber, B.T., Norris, R.D., and MacLeod, K.G. 2002. Deep-sea paleotemperature record of extreme warmth during the Cretaceous. Geology 30: 123–126. Crossref
Huxley, A.F. 1974. Muscular contraction. The Journal of Physiology 243: 1–43. Crosref
Ibiricu, L.M., Casal, G.A., Lamanna, M.C., Martínez, R.D., Harris, J.D., and Lacovara, K.J. 2012. The southernmost records of Rebbachisauridae (Sauropoda: Diplodocoidea), from early Late Cretaceous deposits in central Patagonia. Cretaceous Research 34: 220–232. Crossref
Ibiricu, L.M., Casal G.A., Martínez, R.D., Lamanna, M.C., Luna, M., and Salgado, L. 2013a. Katepensaurus goicoecheai gen. et sp. nov., a Late Cretaceous rebbachisaurid (Sauropoda, Diplodocoidea) from central Patagonia, Argentina. Journal of Vertebrate Paleontology 33: 1351–1366. Crossref
Ibiricu, L.M., Casal, G.A., Martínez, R.D., Lamanna, M.C., Luna, M., and Salgado, L. 2015. New material of Katepensaurus goicoecheai (Sauropoda: Diplodocoidea) and its significance for the morphology and evolution of Rebbachisauridae. Ameghiniana 52: 430–446. Crossref
Ibiricu, L.M., Lamanna, M.C., and Lacovara, K.J. 2013b. The influence of caudofemoral musculature on the titanosaurian (Saurischia: Sauropoda) tail skeleton: morphological and phylogenetic implications. Historical Biology 26: 454–471. Crossref
Ibrahim, N., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Gorscak, E., and Lamanna, M.C. 2016. Evidence of a derived titanosaurian (Dinosauria, Sauropoda) in the “Kem Kem beds” of Morocco, with comments on sauropod paleoecology in the Cretaceous of Africa. In: A. Khosla and S.G. Lucas (eds.), Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin 71: 149–159.
Janensch, W. 1947. Pneumatizitat bei Wirbeln von sauropoden und anderen saurischien. Palaeontographica 3: 1–25.
Klein, N., and Sander, M. 2008. Ontogenetic stages in the long bone histology of sauropod dinosaurs. Paleobiology 34: 247–263. Crossref
Lacovara, K.J., Lamanna, M.C., Ibiricu, L.M., Poole, J.C., Schroeter, E.R., Ullmann, P.V., Voegele, K.K., Boles, Z.M., Carter, A.M., Fowler, E.K., Egerton, V.M., Moyer, A.E., Coughenour, C.L., Schein, J.P., Harris, J.D., Martínez, R.D., and Novas, F.E. 2014. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Scientific Reports 4: 6196. Crossref
Lamanna, M.C., and Hasegawa, Y. 2014. New titanosauriform sauropod dinosaur material from the Cenomanian of Morocco: implications for paleoecology and sauropod diversity in the Late Cretaceous of North Africa. Bulletin of Gunma Museum of Natural History 18: 1–19.
Mannion, P.D. and Barrett, P.M. 2013. Additions to the sauropod dinosaur fauna of the Cenomanian (early Late Cretaceous) Kem Kem beds of Morocco: palaeobiogeographical implications of the mid-Cretaceous African sauropod fossil record. Cretaceous Research 45: 49–59. Crossref
Mannion, P.D., Upchurch, P., Mateus, O., Barnes, R.N., and Jones, M.E.H. 2012. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology 10: 521–551. Crossref
Martínez, R.D., Giménez, O., Rodríguez, J.F., Luna, M., and Lamanna, M.C. 2004. An articulated specimen of the basal titanosaurian (Dinosauria: Sauropoda) Epachthosaurus sciuttoi from the early Late Cretaceous Bajo Barreal Formation of Chubut Province, Argentina. Journal of Vertebrate Paleontology 24: 107–120. Crossref
Martínez, R.D.F., Lamanna, M.C., Novas, F.E., Ridgely, R.C., Casal, G.A., Martínez, J.E., Vita, J.R., and Witmer, L.M. 2016. A basal lithostrotian titanosaur (Dinosauria: Sauropoda) with a complete skull: implications for the evolution and paleobiology of Titanosauria. PLoS ONE 11: e0151661. Crossref
Medeiros, M.A. and Schultz, C.L. 2004. Rayososaurus (Sauropoda, Diplodocoidea) no meso-Cretáceo do norte-nordeste Brasileiro. Revista Brasileira de Paleontologia 7: 275–279. Crossref
Nopcsa, F. 1902. Notizen über cretacische Dinosaurier. 3. Wirbel eines südamerikanischen Sauropoden. Sitzungsberichte der Mathematisch-Naturwissenschaftliche Classe der Kaiserlichen Akademie der Wissenschaften, Wien 111: 108–114.
O’Connor, P.M. 2004. Pulmonary pneumaticity in the postcranial skeleton of extant Aves: a case study examining Anseriformes. Journal of Morphology 261: 141–161. Crossref
O’Connor, P.M. 2006. Postcranial pneumaticity: an evaluation of soft-tissue influences on the postcranial skeleton and the reconstruction of pulmonary anatomy in archosaurs. Journal of Morphology 267: 1199–1226. Crossref
O’Connor, P.M. 2007. The postcranial axial skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar. Society of Vertebrate Paleontology Memoir 8: 127–162. Crossref
O’Connor, P.M. 2009. Evolution of archosaurian body plans: skeletal adaptations of an air-sac-based breathing apparatus in birds and other archosaurs. Journal of Experimental Zoology Part A 311: 629–646. Crossref
O’Connor, P.M. and Claessens, L.P. 2005. Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs. Nature 436: 253–256. Crossref
Organ, C.L. 2006. Thoracic epaxial muscles in living archosaurs and ornithopod dinosaurs. The Anatomical Record Part A 288: 782–793. Crossref
Paulina Carabajal, A., Canale, J.I., and Haluza, A. 2016. New rebbachisaurid (Sauropoda, Diplodocoidea) cranial remains from the Cretaceous of Patagonia, Argentina, and the first endocranial description for a South American representative of the clade. Journal of Vertebrate Paleontology 36: e1167067. Crossref
Perry, S.F. and Sander, M. 2004. Reconstructing the evolution of the respiratory apparatus in tetrapods. Respiratory Physiology and Neurobiology 144: 125–139. Crossref
Perry, S.F., Breuer, T., and Pajor, N. 2011. Structure and function of the sauropod respiratory system. In: N. Klein, K. Remes, C.T. Gee, and P.M. Sander (eds.), Biology of the Sauropod Dinosaurs: Understanding the Life of Giants, 83–93. Indiana University Press, Bloomington.
Perry, S.F., Christian, A., Breuer, T., Pajor, N., and Codd, J.R. 2009. Implications of an avian-style respiratory system for gigantism in sauropod dinosaurs. Journal of Experimental Zoology Part A 311: 600–610. Crossref
Poulsen, C.J., Gendaszek, A.S., and Jacob, R.L. 2003. Did the rifting of the Atlantic Ocean cause the Cretaceous thermal maximum? Geology 31: 115–118. Crossref
Pujana, R., Umazano, A., and Bellosi, E. 2007. Maderas fósiles afines a Araucariaceae de la Formación Bajo Barreal, Cretácico Tardío de Patagonia central (Argentina). Revista del Museo Argentino de Ciencias Naturales, nueva serie 9: 161–167.
Retallack, G.J. 1988. Field recognition of paleosols. Geological Society of America Special Paper 216: 1–20. Crossref
Rodríguez, J.F.R. 1992. Interpretación paleoambiental de la Formación Bajo Barreal (Cretácico Tardío) en Estancia Ocho Hermanos, Chubut. In: G.A. Chebli (ed.), 4º Reunión Argentina de Sedimentología, Actas, 81–88. La Plata.
Salgado, L., Canudo, J.I., Garrido, A.C., and Carballido, J.L. 2012. Evidence of gregariousness in rebbachisaurids (Dinosauria, Sauropoda, Diplodocoidea) from the Early Cretaceous of Neuquén (Rayoso Formation), Patagonia, Argentina. Journal of Vertebrate Paleontology 32: 603–613. Crossref
Salgado, L., Carvalho, I.S., and Garrido, A.C. 2006. Zapalasaurus bonapartei, un nuevo dinosaurio saurópodo de la Formación La Amarga (Cretácico Inferior), noroeste de Patagonia, Provincia de Neuquén, Argentina. Geobios 39: 695–707. Crossref
Schwarz-Wings, D. 2009. Reconstruction of the thoracic epaxial musculature of diplodocid and dicraeosaurid sauropods. Journal of Vertebrate Paleontology 29: 517–534. Crossref
Sereno, P.C. and Wilson, J.A. 2005. Structure and evolution of a sauropod tooth battery. In: K.A. Curry Rogers and J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, 157–177. University of California Press, Berkeley. Crossref
Sereno, P.C., Martinez, R.N., Wilson, J.A., Varricchio, D.J., Alcober, O.A., and Larsson, H.C.E. 2008. Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina. PLoS ONE 3: e3303. Crossref
Sereno, P.C., Wilson, J.A., Witmer, L.M., Whitlock, J.A., Maga, A., Ide, O., and Rowe, T.A. 2007. Structural extremes in a Cretaceous dinosaur. PLoS ONE 2: e1230. Crossref
Seton, M., Müller, R.D., Zahirovic, S., Gaina, C., Torsvik, T., Shephard, G., Talsma, A., Gurnis, M., Turner, M., Maus, S., and Chandler, M. 2012. Global continental and ocean basin reconstructions since 200 Ma. Earth-Science Reviews 113: 212–270.
Taylor, M.P. 2009. A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914). Journal of Vertebrate Paleontology 29: 787–806. Crossref
Torcida Fernández-Baldor, F., Canudo, J.I., Huerta, P., Montero, D., Pereda Suberbiola, X., and Salgado, L. 2011. Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica 56: 535–552. Crossref
Umazano, A.M., Bellosi, E.S., Visconti, G., and Melchor, R.N. 2008. Mechanisms of aggradation in fluvial systems influenced by explosive volcanism: an example from the Upper Cretaceous Bajo Barreal Formation, San Jorge Basin, Argentina. Sedimentary Geology 203: 213–228. Crossref
Umazano, A.M., Bellosi, E.S., Visconti, G., and Melchor, R.N. 2012. Detecting allocyclic signals in volcaniclastic fluvial successions: Facies, architecture and stacking pattern from the Cretaceous of central Patagonia, Argentina. Journal of South American Earth Sciences 40: 94–115. Crossref
Walker, J.D., Geissman, J.W., Bowring, S.A., and Babcock, L.E. 2012. Geologic Time Scale v. 4.0. Geological Society of America, Boulder.
Waskow, K., and Sander, P.M. 2014. Growth record and histological variation in the dorsal ribs of Camarasaurus sp. (Sauropoda). Journal of Vertebrate Paleontology 34: 852–869. Crossref
Wedel, M.J. 2003. The evolution of vertebral pneumaticity in sauropod dinosaurs. Journal of Vertebrate Paleontology 23: 344–357. Crossref
Wedel, M.J. 2005. Postcranial skeletal pneumaticity in sauropods and its implications for mass estimates. In: K. Curry Rogers and J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, 201–228. University of California Press, Berkeley. Crossref
Wedel, M.J. 2006. Origin of postcranial skeletal pneumaticity in dinosaurs. Integrative Zoology 1: 80–85.
Wedel, M.J. 2007. What pneumaticity tells us about “prosauropods”, and vice versa. Special Papers in Palaeontology 77: 207–222.
Wedel, M.J. 2009. Evidence for bird-like air sacs in saurischian dinosaurs. Journal of Experimental Zoology Part A 311: 611–628. Crossref
Wedel, M.J. and Taylor, M.P. 2013. Caudal pneumaticity and pneumatic hiatuses in the sauropod dinosaurs Giraffatitan and Apatosaurus. PLoS ONE 8: e78213. Crossref
Wedel, M.J., Cifelli, R.L., and Sanders, R.K. 2000. Osteology, paleobiology, and relationships of the sauropod dinosaur Sauroposeidon. Acta Palaeontologica Polonica 45: 343–388.
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Wilson, J.A. and Allain, R. 2015. Osteology of Rebbachisaurus garasbae Lavocat, a diplodocoid
(Dinosauria: Sauropoda) from the early Late Cretaceous-aged Kem Kem beds
of southeastern Morocco. Journal of Vertebrate
Paleontology 35: e1000701. Crossref
Wilson, J.A., D’Emic, M.D., Ikejiri, T., Moacdieh, E.M., and Whitlock, J.A. 2011. A nomenclature for vertebral fossae in sauropods and other saurischian dinosaurs. PLoS ONE 6: e17114. Crossref
Witmer, L.M. 1995. The extant phylogenetic bracket and the importance of reconstructing soft tissues in fossils. In: J. Thomason (ed.), Functional Morphology in Vertebrate Paleontology, 19–33. Cambridge University Press, Cambridge.
Yates, A.M., Wedel, M.J., and Bonnan, M.F. 2012. The early evolution of postcranial skeletal pneumaticity in sauropodomorph dinosaurs. Acta Palaeontologica Polonica 57: 85–100. Crossref
Zhou, J., Poulsen, C.J., Rosenbloom, N., Shields, C., and Briegleb, B. 2012. Vegetation-climate interactions in the warm mid-Cretaceous. Climate of the Past 8: 565–576. Crossref
Zurriaguz, V. and Álvarez, A. 2014. Shape variation in presacral vertebrae of saltasaurine titanosaurs (Dinosauria, Sauropoda). Historical Biology 26: 801–809. Crossref
Zurriaguz, V.L. and Cerda, I.A. 2017. Caudal pneumaticity in derived titanosaurs (Dinosauria: Sauropoda). Cretaceous Research 73: 14–24. Crossref
Zurriaguz, V. and Powell, J. 2015. New contributions to the presacral osteology of Saltasaurus loricatus (Sauropoda, Titanosauria) from the Upper Cretaceous of northern Argentina. Cretaceous Research 54: 283–300. Crossref
Acta Palaeontol. Pol. 62 (2):
221–236, 2017
https://doi.org/10.4202/app.00316.2016