
A new tribe of castoroidine beavers from the late Arikareean to Hemphillian (Oligocene–Miocene) of western North America
WILLIAM W. KORTH
Korth, W.W. 2017. A new tribe of castoroidine beavers from the late Arikareean to Hemphillian (Oligocene–Miocene) of western North America. Acta Palaeontologica Polonica 62 (2): 249–258.
Two genera of Miocene beavers (Castoridae), Hystricops and Priusaulax are referred to a new tribe Hystricopini of the subfamily of giant beavers, Castoroidinae based on cranial and dental morphologies. Previously undescribed and more complete specimens of Hystricops demonstrate that it is referable to the Castoroidinae rather than the Castorinae. Along with Priusaulax, it is believed to represent a group of basal castoroidines, distinct from later Castoroidinae by the retention of the primitive morphology of the contact of the lacrimal bone with the jugal on the cranium, as well as more primitive dental morphologies (lower crown-height and simple occlusal pattern of cheek teeth). A partial cranium and several isolated teeth from the Hemingfordian of Colorado, originally referred to Anchitheriomys?, are referred here to a new species, Priusaulax wilsoni sp. nov. Monosaulax senrudi from the Barstovian of Montana is also referred to Priusaulax. Hystricops is limited to the type species H. venustus from the Clarendonian and Barstovian North American Land Mammal Age (NALMA), and H. browni Shotwell, 1963, from the Hemphillian NALMA.
Key words: Mammalia, Rodentia, Castoridae, Castoroidinae, cranial anatomy, dental terminology, North American Land Mammal Ages, Colorado.
William W. Korth [wwkorth@frontiernet.net], Rochester Institute of Vertebrate Paleontology, 265 Carling Road, Rochester, New York 14610, USA.
Received 10 January 2017, accepted 9 February 2017, available online 11 April 2017.
Copyright © 2017 W.W. Korth. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Hystricops was named by Leidy (1858) as a subgenus of the Recent Old World porcupine Hystrix Linneaus, 1758, based on two isolated cheek teeth from the “Niobrara River beds” in northern Nebraska. This horizon was later interpreted as the Ogallala Group and divided into the Valentine and Ash Hollow formations, representing the middle to early–late Miocene: Barstovian to Clarendonian, repectively (see Voorhies 1990, for historical review). Matthew (1902) was the first to suggest that the teeth of Hystricops were more likely those of a beaver (= Castoridae) because of the implausibility of having a genus of an Old World family occurring in Tertiary rocks of North America. Stirton (1935) definitely referred the genus to the Castoridae based on the morphology of the cheek teeth, even though no additional specimens had been recovered at the time of his publication. Additional isolated teeth referred to Hystricops were later reported from the Clarendonian and Hemphillian (late Miocene) of Oregon (Shotwell and Russell 1963; Shotwell 1963), including a new species from the Hemphillian, H. browni Shotwell, 1963. The first larger sample (more than ten specimens) of Hystricops was reported from the late Clarendonian Ash Hollow Formation of Nebraska (Korth 1997, 1998b). This collection included the first specimens other than isolated teeth. It was suggested that these specimens were likely from the same horizon as the holotype material of H. venustus, and referred to that species (Korth 1998b).
Two other species have been suggested as referable to Hystricops by some authors: Monosaulax senrudi Wood, 1945, from the Barstovian of Montana (see Stout and Stone 1971) and Anchitheriomys? sp. (Wilson 1960) from the Hemingfordian of Colorado (also see Korth 1998b).
Due to the sparse record of Hystricops (predominantly isolated cheek teeth), its systematic position within the family Castoridae has been in question (Stirton 1935; Korth 1994). McKenna and Bell (1998) placed it in the clade along with the Recent beavers in the subfamily Castorinae. Korth (2001b) included it in the Agnotocastorinae along with the most primitive genera of the family. More recently, Flynn and Jacobs (2008) included the genus in “Basal Castoroids”, again suggesting inclusion among the more primitive genera. However, in the cladogram presented by the latter (Flynn and Jacobs 2008: fig. 23.2), Hystricops was figured as the sister-taxon of the Recent Castor Linnaeus, 1758, similar to the position suggested by McKenna and Bell (1998). Rybczynski (2007: fig. 3) placed Hystricops within the subfamily Castorinae.
Priusaulax was named by Korth and Bailey (2006) from the latest Arikareean and early Hemingfordian (latest Oligocene–early Miocene) of Nebraska. It was regarded as basal castoroidine based on dental and cranial features as well as age of occurrence. Rybczynski (2007: fig. 3), in a cladistic analysis of the Castoridae, had a similar result, placing Priusaulax as the sister-taxon to the clade that included both the Castoroidinae and Castorinae. Similarities between Priusaulax and Hystricops appear to ally the two genera, and suggest a systematic position near the Castoroidinae.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; F:AM, Frick Collections, American Museum of Natural History, New York, USA; KU, University of Kansas Museum of Natural History, Lawrence, Kansas, USA; UNSM, University of Nebraska State Museum, Lincoln, Nebraska, USA; UOMNH, University of Oregon Museum of Natural History, Eugene, Oregon, USA; UW, University of Wyoming, Geological Museum, Laramie, Wyoming, USA.
Material and methods
The majority of the fossil material discussed herein has been previously cited with the exception of three specimens of Hystricops venustus from Nebraska, housed in the Frick Collections of the American Museum of Natural History and the University of Nebraska State Museum, the bulk of which were collected in the 1940s and 1950s. The holotype and topotypic specimens of the new species of Priusaulax from Colorado are housed at the University of Kansas Museum of Natural History, originally described by Wilson (1960). Those of P. browni are at the University of Nebraska State Museum (Korth and Bailey 2006). The holotype and referred specimens of H. browni are at the University of Oregon Museum of Natural History, collected in the 1950s and early 1960s (Shotwell 1963).
Dental terminology follows that of Stirton (1935), and cranial terminology follows that of Wahlert (1977). Division of North American Land Mammal Ages follows that of Janis et al. (2008; e.g., Ar1, Cl2, etc.). All measurements in mm, taken to the nearest 0.01 mm.
Systematic palaeontology
Family Castoridae Hemprich, 1820
Subfamily Castoroidinae Allen, 1877
Tribe Hystricopini nov.
Type genus: Hystricops Leidy, 1958.
Included genera: Hystricops Leidy, 1958; Priusaulax Korth and Bailey, 2006.
Diagnosis.—Derived features shared with castorines, castoroidines, and palaeocastorines: P3 lacking; P4/p4 larger than molars; upper tooth rows slightly diverge posteriorly; upper diastema arched; posterior palatine foramina intersected by maxillary-palatine suture; palatal surface distinctly grooved; length of incisive foramen less than 30% length of upper diastema; loss of stapedial foramen in auditory bulla. Derived characters shared with castoroidines: larger size (compared to contemporaneous species); angle of dentary laterally expanded with flattened ventral surface, mandibular condyle medial to angle and ascending ramus (= zig-zag pattern). Retained primitive features of castorids: cheek teeth mesodont in crown-height; occlusal morphology of cheek teeth with single, persistent re-entrant (lingual hypoflexus on upper; buccal hypoflexid on lowers) and three persistent, transversely oriented fossettes (-ids); jugal bone contacts lacrimal at its anterior end; enamel surface of incisors smooth, and unornamented; nasal bones extend more posteriorly than premaxillaries on dorsal surface of cranium; rostrum elongated; upper incisors slightly procumbent.
Discussion.—The Hystricopini share nearly all of the derived cranial characters of the Castoroidinae (Korth 2001b: 289, fig. 4). The only major difference is in the contact of the lacrimal bone with the jugal. In more derived castoroidines the jugal ends anteriorly well before joining the lacrimal. This feature in hystricopines is clearly primitive among castorids. In the original description of Priusaulax, it was suggested that the presence of most of the castoroidine features and presence of the lacrimal-jugal contact suggested a basal position of the genus within the castoroidines. In addition, the cheek teeth of both Hystricops and Priusaulax are similar to other early castoroidines (e.g., Monosaulax Stirton, 1935) being mesodont in crown-height and retaining a single re-entrant and three transversely elongated enamel fossettes after minimal to moderate wear.
Hystricopini differ from the nothodipoidine castoroidines in being larger and lacking the fossorial adaptions of the skull of the latter (smaller size; higher-crowned cheek teeth; low, wide cranium; more procumbent incisors; greater postorbital constriction; diastema more strongly arched; Korth 2007: 1536), as well as retaining a more primitive occlusal pattern of the cheek teeth (never attaining the S-pattern of nothodipoidines; Korth 2002: 21). The Hystriocopini also differ from the Castoroidini in being smaller in size, having lower-crowned cheek teeth, and lacking the specialized pattern of the cheek teeth (S-pattern) and ridged incisors of the most derived genera (Korth 2001b: 289). As previously suggested for Priusaulax (Korth and Bailey 2006), the Hystricopini appear to be basal castoroidines.
Stratigraphic and geographic range.—Late Arikareean (latest Oligocene) to Hemphillian (late Miocene) of western North America.
Genus Priusaulax Korth and Bailey, 2006
Type species: P. browni Korth and Bailey, 2006, UNSM locality Sf-105E/N, Nebraska, USA, Upper Arikaree Group, latest Arikareean (early Miocene).
Emended diagnosis.—Hystricopine; markedly smaller and less robust than species of Hystricops; anterior enamel surface on i1 gently convex, cross-section longer than wide; cheek teeth slightly lower-crowned.
Stratigraphic and geographic range.—Late Arikareean (late Oligocene: Ar4) to early Barstovian (middle Miocene: Ba1) of western North America.
Priusaulax wilsoni sp. nov.
Figs. 1–3; Table 1.
1960 Anchitheriomys? sp.; Wilson 1960: 66, figs. 68–70.
1980 Hystricops senrudi (Wood, 1945); Cassilliano 1980: 36, fig. 14 (in part).
1998 Hystricops sp.; Korth 1998b: 313, fig. 6.
2006 Priusaulax sp.; Korth and Bailey 2006: 240.
Holotype: KU 10173, partial cranium with right side of rostrum, maxilla and orbital wall containing right P4–M3.
Type locality: Quarry A, Logan County, Colorado, USA (Wilson 1960).
Type horizon: Pawnee Creek Formation, Early Hemingfordian (early Miocene), He1of Janis et al. (2008).
Material.—KU 10175, left m1 or m2; KU 10174, right M1 or M2; from the type locality. UW 10010, right m3 from Horse Creek Quarry (“…about 2.5 m above the contact with the Harrison Formation”) Laramie County Wyoming, USA (Cassiliano 1980: 36). UNSM 26599, right dentary with dp4 (p4 in crypt)–m3; UNSM 11984, m3; from Runningwater Formation, Dawes and Box Butte counties, Nebraska, USA (Korth and Bailey 2006: 240).
Diagnosis.—Slightly larger than Priusaulax browni and smaller than P. senrudi; no anterior re-entrant valley or accessory cuspule (anterostylid) on p4 as in P. browni.
Description.—Wilson (1960: 66) only briefly described the cranium of P. wilsoni (as Anchitheriomys? sp.), KU 10173, but provided a table of measurements (Wilson 1960: 67). The following description will focus on the details not noted by Wilson (1960). In dorsal view, the rostrum is long and parallel-sided (Fig. 1A). The nasals taper in width posteriorly, and end well posterior of the frontal-premaxillary suture in a point (approximately 9.2 mm). A small piece of the lacrimal bone is present at the anteromedial corner of the orbit. Although the jugal bone is missing, the jugal-maxillary suture is preserved on the dorsal surface of the zygomatic arch, and it is evident that the most-anterior extent of the jugal did contact the lacrimal bone. Well-developed temporal crests are present on the frontal bones. There is a slight swelling at the anterior end of the crests that converge posteriorly in a slight curve. However, the two crests do not meet at the posterior end of the frontals. The parietals are lacking on the cranium, so it cannot be determined whether the temporal crests are united more posteriorly.
In lateral view (Fig. 1A), the diastema is long (38.5 mm); approximately 1.6 times longer than the length of the tooth row. Its ventral surface is gently arched dorsally. The infraorbital foramen is just above the ventral surface of the diastema at its posterior end (below the anterodorsal end of the zygomatic arch). The foramen opens anteriorly and is laterally compressed. A flange of bone is present lateral to the foramen (= attachment of masseter lateralis). The orbital wall is badly broken so little can be determined. The nasolacrimal foramen is apparently large and in the anterodorsal corner of the orbital wall. The only other observable foramen is the sphenopalatine which is within the maxillary bone, dorsal to M1. All other sutures and foramina are unobservable.
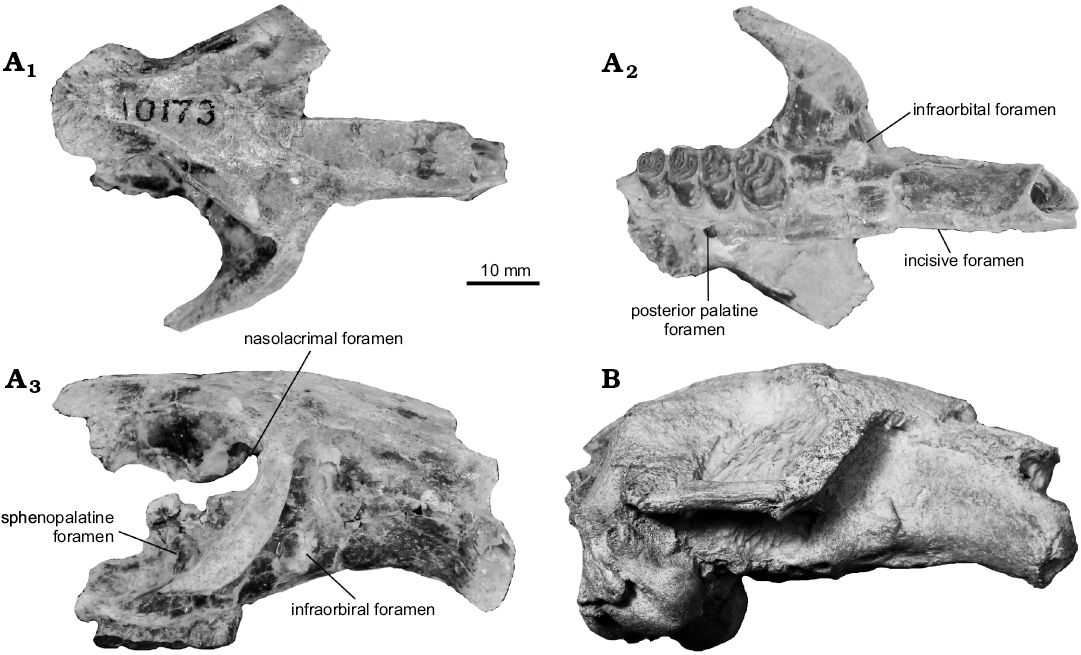
Fig. 1. Crania of castoroidine beavers Priusaulax spp. A. Priusaulax wilsoni sp. nov., KU 10173 (holotype) from the early Hemingfordian (early Miocene) of Colorado, USA, in dorsal (A1), ventral (A2), and right lateral (A3) views. B. Priusaulax browni Korth and Bailey, 2006, UNSM 119707 from the late Arikareean (earliest Miocene) of Nebraska, USA, in right lateral view.
Although the upper incisor is lacking, it is evident from the alveolus that the anterior enamel surface was slightly convex (Fig. 1A), and in cross-section, the tooth was slightly longer than wide (alveolus: length = 7.6 mm; width = 6.3 mm). The anterior and posterior ends of the incisive foramen are preserved so that it can be measured (8.1 mm). The ratio of incisive foramen length to the length of the diastema is 0.21. The premaxillary-maxillary suture crosses the incisive foramen at its posterior end. There is a distinct groove running anteroposteriorly along the maxilla medial to the tooth row that ends posteriorly at the level of M2, at the posterior palatine foramen. The palatine bone is not preserved, but the maxillary-palatine suture is evident and joins the posterior palatine foramen.
P4 is the largest of the cheek teeth which decrease in size toward M3. The tooth row diverges slightly posteriorly (Fig. 1, 2A; Wilson 1960: fig. 70c). The width of palate can be estimated (at P4 approximately 8 mm; at M3 approximately 15 mm). The occlusal morphology of the cheek teeth have been well described by Wilson (1960). On P4, M2, and M3, there is a smaller accessory enamel lake posterior to the mesofossette or mesoflexus. These are smaller than the other fossettes and will likely be eliminated with additional wear, thus producing the basic three-fossette pattern. There is also an additional enamel fossette on the anterior half of M3 that appears to be a split of the parafossette.
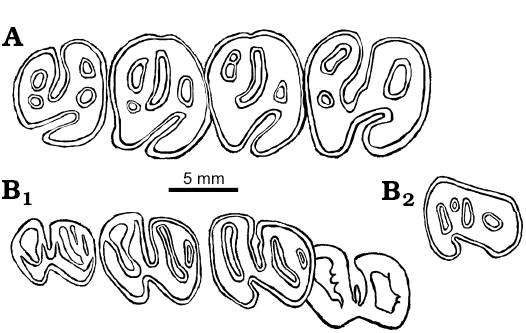
Fig. 2. Cheek teeth of castoroidine beaver Priusaulax wilsoni sp. nov. A. KU 10173 (holotype) from early Hemingfordian (early Miocene) Pawnee Creek Formation, Colorado, USA, right P4–M3 in occlusal view. B. UNSM 26599 from Hemingfordian (early Miocene) Upper Arikaree Group, Nebraska, USA, p4 (in crypt)–m3 (B1) and dp4 (B2) in occlusal views.
The dentary referred to Priusaulax sp. by Korth and Bailey (2006), UNSM 26599, is slightly larger than those of P. browni, but very similar in morphology (Fig. 3; also see Korth and Bailey 2006: 241–243 for complete description). This specimen is comparable in size to the type of P. wilsoni, and is here referred to the latter. The age of occurrence of UNSM 26599 is early Hemingfordian, also compatible with that of the type of P. wilsoni. The greatest difference in the morphology of the cheek teeth of UNSM 26599 with those of P. browni is that of p4. In P. browni, there is a distinct anterostylid (or anteroconid) and a narrow valley along the anterior margin of the tooth. In the unerupted p4 of UNSM 26599, there is no distinct anterostylid and the anterior lophid is continuous for its entire width (Fig. 2B).
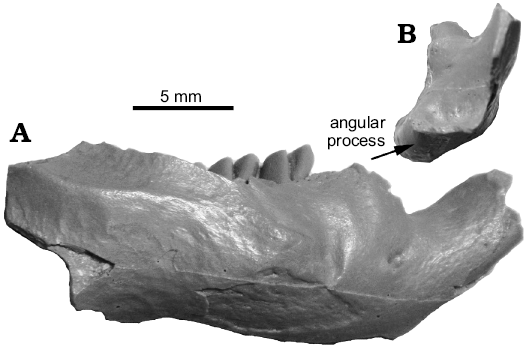
Fig. 3. Dentary of Priusaulax wilsoni sp. nov., UNSM 26599 from Hemingfordian (early Miocene) Upper Arikaree Group, Nebraska, USA, in right lateral (A) and posterior (B) views.
Discussion.—Korth (1998a: 317) suggested that the cheek teeth of KU 10173, originally referred to Anchitheriomys? sp. by Wilson (1960), were more similar to those of H. venustus than any other castorid (large size, mesodonty, hypoflexus with three fossettes, P4 larger than M1). The cheek teeth differ from contemporaneous and similarly sized castoroidines (e.g., Monosaulax) in being lower crowned, even though the occlusal pattern is similar, and having the tooth rows not diverge quite as strongly (palatal width of M3 much greater in castoroidines; see Stirton 1935: fig. 92; Korth 2008: fig. 2C). Several other cranial features separate KU 10173 from castoroidines: jugal bone contacts the lacrimal in KU 10173 (no contact in castoroidines), and the infraorbital foramen is slightly lower on the rostrum in KU 10173, just above the surface of the diastema, whereas it is higher in castoroidines, approximately one-third to one-half the height of the rostrum.
However, all of these features of KU 10173 are similar to those of the reported cranium of Priusaulax browni (Fig. 1B) from the late Arikareean of western Nebraska (Korth and Bailey 2006). P. wilsoni differs from P. browni in being slightly larger (Table 1). The best comparison is in the length of the upper and lower tooth rows. For the type of P. wilsoni the length of P4–M2 is 18.1 mm and that of P. browni is 16.5–16.7 mm (mistakenly listed as P4–M3 in Korth and Bailey 2006: table 1). Similarly, the length of m1–m3 in the referred specimen of P. wilsoni is 17.8 mm, and that of P. browni ranges from 15.5–16.0 mm.
A single isolated m3 from Wyoming originally referred to H. senrudi by Casilliano (1980) is transfered to H. wilsoni based on the occlusal morphology and age of occurrence (see below discussion of H. senrudi).
Stratigraphic and geographic range.—The Hemingfordian (early Miocene) of Colorado, Nebraska, and Wyoming (USA).
Table 1. Comparative dental measurements (in mm) of species of Hystricops and Priusaulax. Abbreviations: L, anteroposterior length; W, transverse width; *, dp4–m3 for P. wilsoni. Measurements for H. browni from Shotwell (1963: table 19), for P. wilsoni from Wilson (1960: 67), Cassilliano (1980: table 1), and Korth and Bailey (2006: table 3).
| |
P4L |
P4W |
M1L |
M1W |
M2W |
M2W |
M3L |
M3W |
P4–M3 |
|
Priusaulax wilsoni |
7.04 |
7.95 |
5.50 |
7.15 |
5.50 |
6.09 |
5.00 |
5.54 |
23.21 |
|
Hystricops venustus (mean) |
8.94 |
9.58 |
6.55 |
7.06 |
7.20 |
6.96 |
5.70 |
6.04 |
28.50 |
|
Hystricops browni |
9.60 |
8.70 |
6.50 |
7.00 |
– |
– |
– |
– |
– |
| |
p4L |
p4W |
m1L |
m1W |
m2L |
m2W |
m3L |
m3W |
p4–m2 |
|
Priusaulax wilsoni (mean) |
– |
– |
5.60 |
6.13 |
5.55 |
5.60 |
5.67 |
5.08 |
17.83* |
|
Priusaulax senrudi |
8.06 |
6.56 |
5.37 |
6.91 |
6.27 |
6.54 |
5.95 |
5.44 |
19.63 |
|
Hystricops venustus (mean) |
10.54 |
8.42 |
7.23 |
7.79 |
7.00 |
7.41 |
6.46 |
6.05 |
26.09 |
Priusaulax browni Korth and Bailey, 2006
Fig. 1B.
Material.—UNSM 119711, dentary with p4–m3 (holotype); UNSM 119707, nearly complete skull lacking teeth; UNSM 119708 dentary with incisor and p4–m3; UNSM 119709, dentary fragment with i1 and p4; UNSM 119710 fragment of dentary with m2–m3; late Arikareean (earliest Miocene) Stage Hill I local fauna, Scott’s Bluff County, Nebraska, USA (Korth and Bailey 2006).
Description.—No new material of this species has been recovered since its original description (Korth and Bailey 2006). The described cranium, UNSM 119707, is partially distorted (dorsoventral compaction of rostrum and anterior displacement of basicranium; Korth and Bailey 2006: fig. 1), but is nearly identical to the partial cranium of P. wilsoni in all observable features (Fig. 1B).
Discussion.—P. browni differs from P. wilsoni in being slightly smaller and in the morphology of p4 (see above discussion of P. wilsoni).
Stratigraphic and geographic range.—The late Arikareean (latest Oligocene) of western Nebraska, USA.
Priusaulax senrudi (Wood, 1945)
Figs. 4, 5, Table 1.
1945 Monosaulax senrudi; Wood 1945: 2, fig. 1.
1917 Hystricops senrudi (Wood); Stout and Stone 1971: 281.
1980 non Hystricops senrudi (Wood); Cassilliano 1980: 36.
1994 Amblycastor sp. nov.; Korth 1994: 133.
1994 ?Monosaulax senrudi Wood; Korth 1994: 148.
1998 ?Oligotheriomys senrudi (Wood); Korth 1998a: 127.
2001 Anchitheriomys senrudi (Wood); Korth 2001a: 54.
2010 “Anchitheriomys” senrudi (Wood); Mörs and Hulbert 2010: 1901.
Holotype: AMNH 39415, left dentary with incisor and p4–m3.
Type locality: “On top of Fighting Butte, at the northwest tip of Chalk Buttes, section 30, Township 1 South, Range 57 East” (Wood 1945: 2), Carter County, Montana, USA.
Type horizon: Barstovian (medial Miocene), Ba1 of Janis et al. (2008).
Material.—Holotype only.
Diagnosis.—Slightly larger than P. wilsoni; additional enamel fossette posterior to hypoflexid variably present on lower cheek teeth.
Description.—The holotype and only specimen of this species has been fully described and figured by Wood (1945). No additional description is necessary.
Discussion.—Monosaulax senrudi Wood, 1945, has been included in a number of castorid genera (see above list of synonymies). Most recently, it has been referred to Anchitheriomys Roger, 1898 by Korth (2001a) and Flynn and Jacobs (2008). However, this allocation was based on the erroneous recognition of a ridged lower incisor. The incisor of the holotype was originally described as having “…fine, slightly divergent striations on the convex enamel face of the incisor...” (Wood 1945: 2). However, no such striations are visible on the incisor (Fig. 5B) clearly not the deeper, very distinct parallel ridges and grooves of the known species of Anchitheriomys (Fig. 5C; also see Korth 2001a; Mörs and Hulbert 2010). The overall morphology of the type specimen is relatively simple, and similar to that of Priusaulax. It differs from the latter only in the presence of an additional enamel fossettid posterior to the mesofossettid on p4 and m1 (Fig. 4B), not known in any specimens of H. venustus, but similar to the accessory fossettes on the upper cheek teeth of P. wilsoni (Fig. 2A). The isolated lower cheek teeth referred to P. wilsoni do not have the accessory fossettid as in P. senrudi (Wilson 1960: fig. 68). The angle of the dentary is not preserved on the type specimen, so it cannot be determined whether there is a lateral extension of the bone or a flattening of the ventral margin at the anterior margin of the angle.
The only specimen other than the holotype referred to H. senrudi was an isolated lower molar (UW 10010) from the Hemingfordian of Wyoming that is essentially unworn and does not have an accessory fossettid (Cassilliano 1980). In size, the UW specimen is closer to that of H. wilsoni (Wilson 1960: 67; Casilliano 1980: table 1). The age of the UW specimen is Hemingfordian, as is H. wilsoni, whereas the type of H. senrudi has been referred to the early Barstovian. This specimen is referred above to H. wilsoni.
Stratigraphic and geographic range.—Barstovian (middle Miocene) of Monanta, USA.
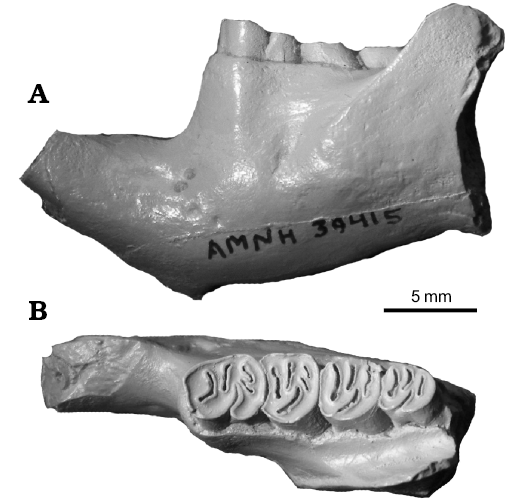
Fig. 4. Holotype of castoroidine beaver Priusaulax senrudi (Wood, 1945), AMNH 39415 (cast) from Barstovian (middle Miocene) Chalky Buttes, Montana, USA. Dentary in lateral view (A) and dorsal view with p4–m3 in occlusal view (B).
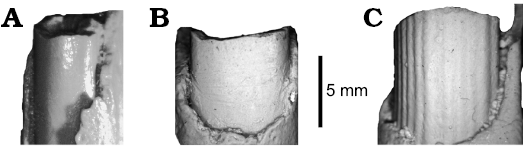
Fig. 5. Lower incisors of castoroidine beaver Priusaulax and Anchitheriomys in anterior views. A. Priusaulax wilsoni sp. nov., UNSM 119711 (holotype) from early Hemingfordian (early Miocene) Pawnee Creek Formation, Colorado, USA. B. Priusaulax senrudi (Wood, 1945), AMNH 39415 (holotype) from Barstovian (middle Miocene) Chalky Buttes, Montana, USA. C. Anchitheriomys stouti Korth, 2001a, AMNH 64017 (holotype) from Hemingfordian (early Miocene) Running Water Formation, Nebraska, USA.
Genus Hystricops Leidy, 1858
Type species: Hystricops venustus Leidy, 1858, “Niobrara River, Loup Fork beds of Hayden and Leidy’s survey. Exact locality unkown.” (Stirton 1935). Believed to be Clarendonian (late Miocene), Ash Hollow Formation, Nebraska, USA.
Diagnosis.—Hystricopine; markedly larger with much more robust dentary than Priusaulax; cheek teeth slightly higher-crowned; lower incisor with flattened anterior enamel surface, nearly as wide as high.
Discussion.—Hystricops is allied here with Priusaulax based on shared morphologies of the cheek teeth (large size; mesodont cheek teeth with simple occlusal morphology [one re-entrant and three transversely elongated fossettes], and dentary (the castoroid-like morphology of the mandibular processes [zig-zag pattern]) and derived castorid characters of the palate (posterior palatine foramen within the maxillary-palatine suture, grooved palate). Hystricops differs from Priusaulax in its much larger size and more robust dentary, higher-crowned cheek teeth, and having the lower incisor flattened anteriorly, and nearly as wide as high (narrower in Priusaulax).
Stratigraphic and geographic range.—Late Barstovian (middle Miocene; Ba2) to Clarendonian (late Miocene; Cl2) of Nebraska and Clarendonian (late Miocene; Cl1) to Hemphillian (latest Miocene; Hh2) of Oregon.
Hystricops venustus Leidy, 1858
Figs. 6–8; Tables 1, 2.
1858 Hystrix (Hystricops) venustus; Leidy 1858: 22.
1902 Erethizon venustus (Leidy); Hay 1902: 733.
1935 Hystricops venustus Leidy; Stirton 1935: 414, figs. 60, 61.
1963 ?Hystricops sp.; Shotwell and Russell 1963: 47, figs. 50, 51, table 7.
Holotype: USNM 1180, right m1 or m2 and left p4.
Type locality: Uncertain locality in “Niobrara River, Loup Fork beds, horizon F of Hayden and Leidy’s survey” (Stirton 1935: 414).
Type horizon: Late Barstovian and late Clarendonian (middle to early–late Miocene), B2–Cl3 of Janis et al. (2008).
Material.—F:AM 64048–64050 from Pratt Quarry, the Merritt Dam Member, Ash Hollow Formation (late Clarendonian), Brown County, Nebraska, USA. F:AM 65026, left maxillary fragment with P4–M1; from Gallop Quarry, the Merritt Dam Member, Ash Hollow Formation (late Clarendonian), Cherry County, Nebraska, USA (Skinner and Johnson 1984; Korth 1997, 1998b). F:AM 64041, left maxilla with P4–M3 from East Kat Quarry, Xmas-Kat Channels (late Clarendonian), Cherry County, Nebraska, USA (Skinner and Johnson 1984); three dentaries with p4–m2 in the F:AM collections (Korth 1997:101). UNSM 75995, left dentary with p4–m1 from Cedar Creek Quarry, Valentine Formation (late Barstovian), Webster County, Nebraska, USA (UNSM locality Wt-102; horizon equivalent to that of Myers Farm: Holman 1977). Ten isolated cheek teeth (UNSM 101660, p4; UNSM 101661, 101662, 101775, m1 or m2; UNSM 101663, 101664, 101667, m3; UNSM 101655, 101666, P4; UNSM 101668, 101663, M1 or M2) and UNSM 101747, an incisor fragment (Korth 1998b: 313), all from Pratt Quarry, the Merritt Dam Member, Ash Hollow Formation (late Clarendonian), Brown County, Nebraska, USA. UOMNH F-2038 and UOMNH F-5802, left P4s, from Black Butte local fauna, Juntura Formation (Clarendonian), Malheur County, Oregon (Bowen et al. 1963; Shotwell and Russell 1963).
Emended diagnosis.—Slightly smaller than H. browni; striae not as deep on cheek teeth as in H. browni (fossettes [-ids] form at earlier stages of wear).
Table 2. Dental measurements (in mm) of Hystricops venustus. Abbreviations: N, number of specimens; M, mean; Min, minimum measurement; Max, maximum measurement; SD, standard deviation; CV, coefficient of variation; L, maximum anteroposterior length; W, maximum transvserse width. Data compiled from Shotwell and Russell (1963: table 1) and Korth (1997: table 3; 1998a: table 6) along with additional new measurements.
| |
P4L |
P4W |
M1L |
M1W |
M2W |
M2W |
M3L |
M3W |
P4–M3 |
p4L |
p4W |
m1L |
m1W |
m2L |
m2W |
m3L |
m3W |
p4–m2 |
|
N |
6 |
6 |
4 |
4 |
1 |
1 |
1 |
1 |
1 |
6 |
6 |
8 |
8 |
3 |
3 |
2 |
3 |
4 |
|
M |
8.94 |
9.58 |
6.55 |
7.06 |
7.20 |
6.96 |
5.70 |
6.04 |
28.50 |
10.54 |
8.42 |
7.23 |
7.79 |
7.00 |
7.41 |
6.46 |
6.05 |
26.09 |
|
Min |
8.42 |
7.89 |
5.74 |
6.41 |
– |
– |
– |
– |
– |
9.00 |
7.08 |
6.37 |
6.98 |
6.15 |
6.71 |
5.56 |
5.80 |
24.42 |
|
Max |
10.46 |
10.50 |
7.45 |
7.81 |
– |
– |
– |
– |
– |
11.90 |
9.39 |
8.00 |
8.33 |
7.80 |
7.81 |
7.35 |
6.51 |
27.00 |
|
SD |
0.79 |
0.94 |
0.72 |
0.57 |
– |
– |
– |
– |
– |
0.96 |
0.91 |
0.52 |
0.48 |
– |
– |
– |
– |
1.17 |
|
CV |
8.84 |
9.83 |
10.93 |
8.14 |
– |
– |
– |
– |
– |
9.07 |
10.86 |
7.23 |
6.17 |
– |
– |
– |
– |
4.47 |
Description.—The upper dentition of H. venustus has been known previously from isolated cheek teeth (Korth 1998b: fig. 6F–H). The two specimens of partial maxillae: F:AM 65026 and F:AM 64041, retain P4–M1 and P4–M3, respectively (Fig. 6). Based on the stage of wear, F:AM 64041 is a much younger individual than F:AM 65024 because in the former, the mesoflexus, metaflexus, and paraflexus remain open on P4 and M3. On M1 and M2 of this specimen (Fig. 6A) the only buccal flexus that remains open is the mesoflexus. On M1 and M2, the parafossette and metafossette have been formed. All of the upper cheek teeth of F:AM 64041 have very a long mesoflexus that nearly reaches the lingual edge of the tooth and curves slightly posteriorly at its lingual end. The remaining parafossette and metafossette are transversely elongated and less than half the width of the tooth in length. M3 has a greatly reduced posterolingual corner of the tooth, resulting in a nearly circular occlusal outline of the tooth. In the heavily worn specimen, F:AM 65026, the P4 appears more elongated than in the little-worn specimen (Fig. 6B). The only flexus that remains open is the hypoflexus. All others have been closed buccally and formed fossettes, the mesofossette being the longest. The dimensions of the tooth are greater than those of the little-worn specimen. On the M1 of F:AM 65026, no flexi remain open. Only two fossettes remain; the mesofossette and the hypofossette. The tooth is much shorter (anteroposteriorly) than the unworn M1. The difference in size and proportions of the cheek teeth of these two maxillary specimens is clearly due to the stage of wear, which demonstrates the change in proportions of the teeth as they wear (molars become shorter, P4 increases in all dimensions).
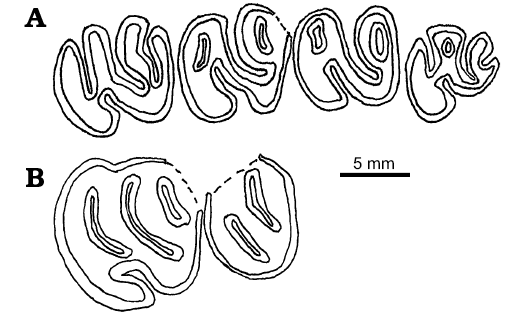
Fig. 6. Occlusal views of upper cheek teeth of castoroidine beaver Hystricops venustus Leidy, 1858, from Clarendonian (late Miocene) Ash Hollow Formation, Nebraska, USA. A. F:AM 64041, left P4–M3. B. F:AM 65026, left P4–M1.
No cranial material has previously been described for Hystricops. F:AM 64041 preserves the base of the zygomatic arch and much of the palate (Fig. 7A). The zygomatic arch is sciuromorphous and originates lateral to the P4 and M1, typical of castorids. The palate has two gentle grooves that run anteroposteriorly on either side of the mid-line of the palate. The maxillary-palatine suture extends anterior to the level of the anterior edge of M1. The posterior palatine foramina are along the suture. Little else can be determined from the specimens available.
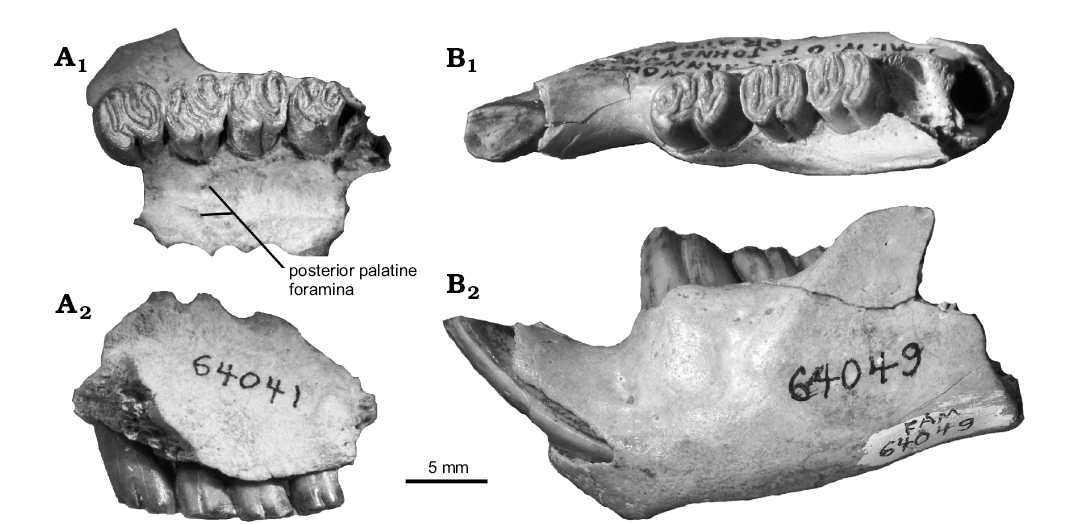
Fig. 7. Maxilla and dentary of castoroidine beaver Hystricops venustus Leidy, 1858, from Clarendonian (late Miocene) Ash Hollow Formation, Nebraska, USA. A. F:AM 64041, left maxilla with P4–M3 in occlusal (A1) and lateral (A2) views. B. F:AM 64049, left dentary with i1, p4–m3 in occlusal (B1) and lateral (B2) views.
Discussion.—The cheek teeth of Hystricops venustus have been described and figured in detail elsewhere (Leidy 1856; Stirton 1935: figs. 60, 61; Korth 1997: fig. 3; Korth 1998b: fig. 6). However, an additional specimen of a dentary, UNSM 75995 (Fig. 8) preserves enough of the posterior end to show that there is a flattened ventral surface of the angle indicating the zig-zag pattern of mandibular processes characteristic of the Castoroidinae (see Korth 2001a: fig. 3B). The species is distinct from the earlier occurring Priusaulax in its much larger size (Table 1; Korth 1997: table 3; Korth 1998b: table 6) and flattened anterior incisor. H. venustus differs from the later occurring H. browni (Hemphillian) in having buccal striae on the upper cheek teeth that do not extend as far toward the base of the crown as the latter, making the flexi less persistent (Fig. 7A; Shotwell 1963: figs. 104, 105).
Shotwell and Russell (1963) described two P4s from the Clarendonian of Oregon referred to “?Hystricops sp.” Both specimens do not differ from the general occlusal morphology of P4s from Nebraska (Shotwell and Russell 1963: figs. 50, 51; Korth 1998b: figs. 6F, G). In size, the Oregon specimens are slightly larger but not appreciably so (Table 2; Shotwell and Russell 1963: table 7; Korth 1998b: table 6). The Oregon specimens are best referred to Hystricops sp., cf. H. venustus. The remainder of the specimens referred to H. venustus are from the late Clarendonian (Cl2) of northcentral Nebraska with one exception, UNSM 75995 (Fig. 8). This specimen is from the Valentine Formation of southeastern Nebraska, approximately 300 km south of the more northern occurrences. The fauna associated with this specimen is typical of that of the late Barstovian (Ba2) Valentine Formation in the northern part of the state (equivalent to Myers Farm Quarry of Voorhies 1990: table 2). This is the earliest occurrence of Hystricops. This specimen is well within the size range of the specimens of H. venustus from the later horizons, and there are no morphologic differences in the dentary, lower cheek teeth or incisor between UNSM 75995 and the type and referred specimens from the later horizons (Fig. 8).
Stratigraphic and geographic range.—Clarendonian (late Miocene) of Nebraska and Oregon, USA.
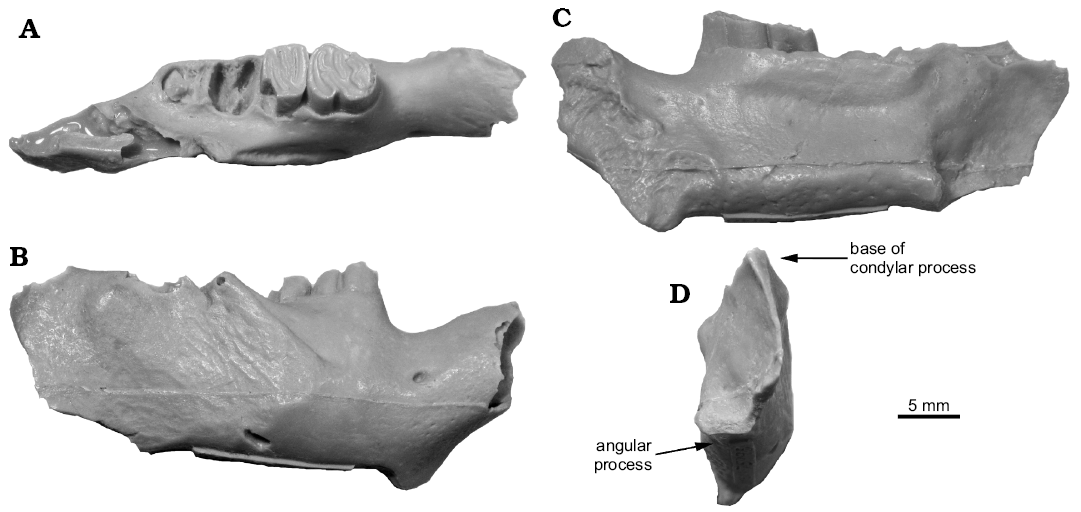
Fig. 8. Dentary of castoroidine beaver Hystricops venustus Leidy, 1858, UNSM 75995 (cast) from Barstovian (middle Miocene) Valentine Formation, southeastern Nebraska, USA, in dorsal (A), lateral (B), medial (C), and posterior (D) views.
Hystricops browni Shotwell, 1963
Table 1 (figured in Shotwell 1963: figs. 104, 105).
Holotype: UOMNH F-15696, associated left and right P4–M1.
Type locality: UOMNH locality 2239, Drewsey Formation, Harney County, Oregon, USA (Shotwell 1963).
Type horizon: Hemphillian (late Miocene), Hh2 of Janis et al. (2008).
Description.—The type and only specimen of H. browni was descriped by Shotwell (1963: 73–75). No additional specimens have been found, so no additional description is necessary.
Diagnosis.—Largest species; striae on upper cheek teeth deeper than in other species, more nearly reaching the base of the crown.
Discussion.—Shotwell (1963) fully described the type specimen of H. browni, and no additional specimens have been subsequently reported. It is similar in size to specimens of H. venustus, but the striae on the teeth more nearly reach the base of the crown (Shotwell 1963: figs. 104, 105), making the flexi more persistent than in H. venustus. It is the latest occurring species of the genus.
Stratigraphic and geographic range.—Hemphillian (latest Miocene) of Oregon, USA.
Conclusions
With the inclusion of Priusaulax along with Hystricops in the same tribe, the systematics of this group can be more closely examined. The group maintains several primitive morphologies for castorids found also in agnotocastorines: (i) long rostrum; (ii) jugal bone contacts lacrimal; (iii) infraorbital foramen low on the rostrum and posteriorly placed; (iv) crown-height of cheek teeth mesodont; (v) enamel surface of incisors smooth; (vi) anterior margin of the ascending ramus more posterior (lateral to m2), and (vii) ratio of diastema length to length of tooth row = 1.6. However, it also has several derived morphologies present in palaeocastorines, castorines, and castoroidines: (i) arched ventral surface of the upper diastema; (ii) posterior palatine foramen within the maxillary-palatine suture; (iii) ratio of incisive foramen length to diastema length less than 0.30; (iv) grooved palate; (v) P4 distinctly larger than M1; (vi) tooth rows diverge posteriorly (more than in agnotocastorines, less than in other subfamilies); (vii) occlusal surface of cheek teeth attains a single re-entrant and three enamel fossettes (-ids) (as in palaeocastorines and primitive members of castoroidines and castorines); and (viii) loss of P3.
The relatively large size of the hystricopines, (compared to contemporaneous genera) and presence of the zig-zag type of angle on the dentary ally the tribe with the subfamily Castoroidinae rather than the Castorinae that have the primitive conditions of vertically aligned angle, condyle and ascending ramus on the dentary. The diagnostic (primitive) feature of the cranium of Priusaulax that bars it from the tribe Castoroidini is the contact of the lacrimal with the jugal on the cranium. The jugal does not extend as far anteriorly in castoroidines, and does not contact the lacrimal. The dental morphology of the cheek teeth of both Priusaulax and Hystricops is primitive and similar to that of early (= primitive) castoroidines, castorines, and most palaeocastorines in crown-height and occlusal pattern.
Acknowledegements
Photos of the cranium of Hystricops wilsoni were provided by Christopher Beard (KU). The cast of the type of P. senrudi was prepared by Vernon Lee (AMNH). F:AM specimens of H. venustus were loaned by Alana Gishlick (AMNH). An earlier version of this paper was critically reviewed by Thomas Mörs (Swedish Museum of Natural History, Stockholm, Sweden) and Lawrence Flynn (Harvard University, Cambridge, Massachusetts, USA).
References
Allen, J.A. 1877. Castoroididae. In: E. Coues and J.A. Allen (eds.), Monographs of North American Roentia. United States Geological Survey of the Territories, 415–442, Government Printing Office, Washington, DC.
Bowen, R.G., Gray, W.L., and Gregory, D.C. 1963. General geology of the northern Juntura Basin. Transactions of the American Philosophical Society 53: 22–34.
Cassilliano, M. 1980. Stratigraphy and vertebrate paleontology of the Horse Creek–Trail Creek area, Laramie County, Wyoming. University of Wyoming Contributions to Geology 19: 25–68.
Flynn, L.J. and Jacobs, L.L. 2008. Castoroidea. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America. Volume 2: Small Mammals, Xenarthrans, and Marine Mammals, 391–405. Cambridge University Press, New York.
Hay, O.P. 1902. Bibliography and catalogue of the fossil Vertebrata of North America. Bulletin of the U.S. Geological Survey 179: 1–868.
Hemprich, W. 1820. Grundriss der Naturgeschichte für höhere Lehranstalten, Entworfen von Dr. W. Hemphrich. 432 pp. August Rucker, Berlin.
Holman, J.A. 1977. Upper Miocene snakes (Reptilia, Serpentes) from southeastern Nebraska. Journal of Herpetology 11: 323–335. Crossref
Janis, C.M., Gunnell, G.F., and Uhen, M.D. 2008. Introduction. In: C.M. Janis, G.F. Gunnell, and M.D. Uhen (eds.), Evolution of Tertiary Mammals of North America. Volume 2: Small Mammal, Xenarthrans, and Marine Mammals, 1–6. Cambridge University Press, New York.
Korth, W.W. 1994. The Tertiary Record of Rodents in North America. 319 pp. Plenum Press, New York. Crossref
Korth, W.W. 1997. Additional rodents (Mammalia) from the Clarendonian (Miocene) of northcentral Nebraska and a review of Clarendonian rodent biostratigraphy of that area. Paludicola 1: 97–111.
Korth, W.W. 1998a. A new beaver (Rodentia, Castoridae) from the Orellan (Oligocene) of North Dakota. Paludicola 1: 127–131.
Korth, W.W. 1998b. Rodents and lagomorphs (Mammalia) from the late Clarendonian (Miocene), Ash Hollow Formation, Brown County, Nebraska. Annals of Carnegie Museum 67: 299–348.
Korth, W.W. 2001a. A new species of Anchitheriomys (Rodentia, Castoridae) and a review of the anchitheriomyine beavers from North America. Paludicola 3: 51–55. Crossref
Korth, W.W. 2001b. Comments on the systematics and classification of the beavers (Rodentia, Castoridae). Journal of Mammalian Evolution 8: 279–296. Crossref
Korth, W.W. 2002. Review of the castoroidine beavers (Rodentia, Castoridae) from the Clarendonian (Miocene) of northcentral Nebraska. Paludicola 4: 15–24.
Korth, W.W. 2007. The skull of Nothodipoides (Castoridae, Rodentia) and the occurrence of fossorial adaptations in beavers. Journal of Paleontology 81: 1542–1546.
Korth, W.W. 2008. Cranial morphology, systematic, and succession of beavers (Rodentia, Castoridae) from the middle Miocene (late Barstovian–early Clarendonian) Valentine Formation of Nebraska, USA. Acta Palaeonotologica Polonica 53: 169–182. Crossref
Korth, W.W. and Bailey, B.E. 2006. Earliest castoroidine beaver (Rodentia, Castoridae) from the late Arikareean (early Miocene) of Nebraska. Annals of Carnegie Museum 75: 237–245. Crossref
Korth W.W. and Emry, R.J. 1997. The skull of Anchitheriomys and a new subfamily of beavers (Castoridae, Rodentia). Journal of Paleontology 71: 343–347. Crossref
Leidy, J. 1858. Notice of remains of extinct Vertebrata from the valley of the Niobrara River, collected during the exploring expedition of 1857, in Nebraska, uner the command of Lieut. G.K. Warren, U.S. Topographic Engineer, by Dr. V.F. Hayden. Proceedings of the Academy of Natural Sciences, Philadelphia 1858: 20–29.
Linneaus, C. 1758. Systema naturae per regna tria naturae, secundum class, ordines, genera, species cum characteribus, differentiis, synonymis, locis. 823 pp. Laurentii Salvii, Stockholm. Crossref
Matthew, W.D. 1902. A horned rodent from the Colorado Miocene, with a revision of the Mylagauli, beavers, and hares of the American Tertiary. Bulletin of the American Museum of Natural History 16: 291–310.
McKenna, M.C. and Bell, S.K. 1998. Classification of Mammals above the Species Level. 631 pp. Columbia University Press, New York.
Mörs, T. and Hulbert, R.C., Jr. 2010. Anchitheriomys Roger, 1898 or Amblycastor Matthew, 1918 (Rodentia, Castoridae)? Taxonomic implications of a mandible from the Miocene of Florida. Journal of Vertebrate Paleontology 30: 1899–1902. Crossref
Roger, O. 1898. Wirbeltierreste aus dem Dinotheriensande der bayerisch-schwäbischen Hochebene. Bericht des Naturwissenschaftlichen Vereins für Schwaben und Neuburg 33: 1–46.
Rybczynski, N. 2007. Castorid phylogenetics: implications for the evolution of swimming and tree-exploitation in beavers. Journal of Mammalian Evolution 14: 1–35. Crossref
Shotwell, J.A. 1963. Pliocene mammalian communities of the Juntura Basin. Transactions of the American Philosophical Society 53: 7–21.
Shotwell, J.A. and Russell, D.E. 1963. Mammalian fauna of the Upper Juntura Formation, the Black Butte local fauna. Transactions of the American Philosophical Society 53: 42–77.
Skinner, M.F. and Johnson, F.W. 1984. Tertiary stratigraphy and the Frick Collection of fossil vertebrates from north-central Nebraska. Bulletin of the American Museum of Natural History 178: 215–368.
Stout, T.M. and Stone, W.J. 1971. Fossil beavers in Tertiary caprocks in North Dakota and Montana. Geological Society of America, Abstracts 281–282. Third Annual Meeting of the Northcentral Section of the Geological Society of America.
Stirton, R. A. 1935. A review of the Tertiary beavers. University of California Publications, Geological Sciences 23: 391–458.
Voorhies, M.R. 1990. Vertebrate paleontology of the proposed Norden Reservoir area, Brown, Cherry, and Keya Paha counties, Nebraska. Technical Report 82-09. 731 pp. Division of Archeological Research, University of Nebraska, Lincoln.
Wahlert, J.H. 1977. Cranial foramina and relationships of Eutypomys (Rodentia, Eutypomyidae). American Museum Novitates 2626: 1–8.
Wilson, R.W. 1960. Early Miocene rodents and insectivores from northeastern Colorado. University of Kansas Paleontological Contributions, Vertebrata 7: 1–92.
Wood, H.E. 1945. Late Miocene beaver from sourtheastern Montana. American Museum Novitates 1299: 1–6.
Acta Palaeontol. Pol. 62 (2):
249–258, 2017
https://doi.org/10.4202/app.00339.2017