
A new euselachian shark from the early Permian of the Middle Urals, Russia
ALEXANDER O. IVANOV, CHRISTOPHER J. DUFFIN, and SERGE V. NAUGOLNYKH
Ivanov , A.O., Duffin, C.J., and Naugolnykh, S.V. 2017. A new euselachian shark from the early Permian of the Middle Urals, Russia. Acta Palaeontologica Polonica 62 (2): 289–298.
The isolated teeth of a new euselachian shark Artiodus prominens Ivanov and Duffin gen. et sp. nov. have been found in the Artinskian Stage (Early Permian) of Krasnoufimskie Klyuchiki quarry (Sverdlovsk Region, Middle Urals, Russia). The teeth of Artiodus possess a multicuspid orthodont crown with from four to nine triangular cusps; prominent labial projection terminating in a large round tubercle; distinct ornamentation from straight or recurved cristae; oval or semilunar, elongate, considerably vascularized base; dense vascular network formed of transverse horizontal, ascending, short secondary and semicircular canals. The teeth of the new taxon otherwise most closely resemble the teeth of some protacrodontid and sphenacanthid euselachians possessing a protacrodont-type crown, but differ from the teeth of all other known euselachians in the unique structure of the labial projection. The studied teeth vary in crown and base morphology, and three tooth morphotypes can be distinguished in the collection reflecting a moderate degree of linear gradient monognathic heterodonty. The range of morphologies otherwise displayed by the collection of teeth shows the greatest similarity to that described for the dentitions of relatively high-crowned hybodontids from the Mesozoic. The internal structure of the teeth, including their vascularization system is reconstructed using microtomography. The highest chondrichthyan taxonomic diversity is found in the Artinskian, especially from the localities of the Middle and South Urals.
Key words: Chondrichthyes, Elasmobranchii, teeth, Permian, Russia, Urals.
Alexander O. Ivanov [IvanovA-Paleo@yandex.ru], Institute of Earth Sciences, St. Petersburg State University, 16 Liniya 29, St. Petersburg, 199178, Russia; Kazan Federal University, 18 Kremlevskaya ul., Kazan, 420008, Russia.
Christopher J. Duffin [cduffin@blueyonder.co.uk], Palaeontology Section, Earth Science Department, The Natural History Museum, Cromwell Road, London, SW7 5BD, UK; current address: 146, Church Hill Road, Sutton, Surrey SM3 8NF, UK.
Serge V. Naugolnykh [naugolnkh@list.ru], Geological Institute of Russian Academy of Sciences, 7, Pyzhevsky per., Moscow, 119017, Russia; Kazan Federal University, 18, Kremlevskaya ul., Kazan, 420008, Russia.
Received 31 January 2017, accepted 14 March 2017, available online 31 May 2017.
Copyright © 2017 A.O. Ivanov et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
When Louis Agassiz produced his monumental pioneering work on fossil fishes (Agassiz 1833–1843), Carboniferous chondrichthyans were well represented, as were those from Late Triassic and later deposits. Chondrichthyans of what were eventually to be identified as Devonian and Permian age were, by contrast, almost totally absent. This sampling gap was later gradually to be filled by descriptions of often partial chondrichthyan skeletons and dentitions from now classic localities. These include the Cisuralian (lower Permian) of Germany, Bohemia, Oklahoma and Texas (USA); the Guadelupian (middle Permian) of East Greenland; the Lopingian (upper Permian) Marl Slate (north east England), Kupferschiefer (Germany), Productus Limestone (Pakistan) and deposits of Montpelier (Bear Lake County, Idaho, USA)(see details and references in Zangerl 1981; Ginter et al. 2010). More recently, bulk processing of sediment for microvertebrate content has vastly increased our knowledge of Mesozoic selachian faunas. Carboniferous and Devonian deposits have begun to be sampled in a similar fashion (Ginter et al. 2010), again yielding significant numbers of new taxa. This approach has only recently been applied to sequences of Permian age, but the initial results are, as expected, very encouraging (e.g., Johnson 1981, 1996; Ivanov 2005; Koot et al. 2013).
Chondrichthyan remains are both diverse and abundant in the early Permian of the Middle Urals of Russia, especially in rocks belonging to the Artinskian Stage (early Permian); the vicinity of Krasnoufimsk (Sverdlovsk Region, Middle Urals) is famous for its Artinskian fishes. Symphyseal tooth-whorls of the agassizodontid eugeneodontiform, Helicoprion bessonovi Karpinsky, 1899, were collected in Artinskian deposits exposed at Divya Gora in 1898 (Karpinsky 1899). Further abundant fish remains were later discovered in several quarries surrounding the town of Krasnoufimsk: Chigvintsevo, Krasnoufimskie Kluchiki, Ryabinovka, Selektsiya, Sobolevskiy. In addition to the symphyseal tooth-whorls of Helicoprion and numerous isolated teeth of ctenacanthids, euselachians and petalodontids (Ivanov 2014). These localities have also yielded well preserved parts of the endoskeleton of euselachian elasmobranch and cochliodont holocephalian (Ivanov 2013, 2014).
The purpose of the present paper is to describe the teeth of a new euselachian shark which have been found at Krasnoufimskie Klyuchiki quarry.
Institutional abbreviations.—PM SPU, Palaeontological Museum, Institute of Earth Sciences, St. Petersburg State University, Russia; NHMUK PV, Fossil Fish Section, Earth Science Department, Natural History Museum, London, UK.
Material and methods
Those teeth which have been separated from their enclosing matrix were studied and micrographed using Cambridge CamScan-4 and Tescan VEGA-II XMU scanning electron microscopes. One specimen with a broken crown in longitudinal section was photographed using an optical Leica MZ 16 microscope fitted with a digital camera. The internal structure of the teeth of the new taxon was reconstructed utilising a SkyScan 1172 Bruker-microCT (Center for Geo-Environmental Research and Modeling “GEOMODEL”, Research park of St. Petersburg State University). Projection images were reconstructed using NRecon software into cross section images having rotated them to an upright position in the software DataViewer. CTAnalyzer software was used to correct visualization and interpretation of the reconstructed images, and CTVox software was used to achieve 3D volume rendering.
Specimens still adhering to the matrix were observed and photographed using a Leica M205C optical microscope and Leica DFC425 C digital microscope camera. This system was used to produce high-resolution pictures of objects that have a high 3-D type character. Using the multi-focus option and Leica application suite (LAS) software the clearest images are retained and combined so that the object has optimum focus. The system provides a scale but the object size was checked directly with the specimen next to a stage graticule measuring to the nearest 0.1 mm.
Geological setting
Krasnoufimskie Klyuchiki quarry is located 1 km east of the village of Klyuchiki and around 7 km to the north of the town of Krasnoufimsk, Krasnoufimsk District, Sverdlovsk Region, Russia (Fig. 1). This area belongs to the Yuryuzan-Ay Depression of the Middle Urals. The quarry extends over an area of 150 m2 and now exposes deposits of the Divya Formation (Sarginskian Regional Stage, Upper Substage of the Artinskian; for details of the regional stratigraphy see Chuvashov and Dyupina 1973; Chuvashov et al. 1990; Naugolnykh 2014). The deposits exposed in the quarry consist of platy marls of grey to yellow-grey or blue-grey color, with intercalations of clay, siltstone, and limestone. The general thickness of the exposed deposits is approximately 6 m.
The palaeontologically richest interval is in the middle part of the section and corresponds to the relatively dark blue-grey marls. The assemblage of fossils from Krasnoufimskie Klyuchiki quarry includes the following higher plants: the lycopodiopsid, Ufadedron ufaensis Naugolnykh, 2014; Paracalamites sp., an equisetophyte; the peltasperm Permocallipteris sp.; the vojnovskyan gymnosperms Rufloria sp. and Lepeophyllum sp. (or so-called “Angarian cordaits”; see Naugolnykh 2001); isolated seeds of the early coniferophytes Cardiocarpus cordatus (Eichwald) Schmalhausen, 1887, Catpolithes sp., and Cordaicarpus sp.; invertebrates: conulariids; brachiopods; bivalves; the gastropod Worthenia aff. burtasorum (Golovkinsky, 1868); nautiloids Orthoceratites siphonocentralis Krotov, 1885 and Metacoceras artiense Kruglov, 1928; the ammonoids Medlicottia orbignyana (Verneuil, 1845), Uraloceras suessii (Karpinsky, 1889), and Paragastrioceras jossae (Verneuil, 1845); the trilobite Ditomopyge artinskense (Weber, 1933); chondrichthyan and actinopterygian fishes, and various trace fossils.
Actinopterygian remains include the skull bones and scales of Rhadinichthys flexuosus Yankevich, 2001 and Acrolepis chuvashovi Yankevich, 2001 (Yankevich 2001). The chondrichthyan component of the fauna varies extensively in quality of preservation from isolated teeth, scales and fin spines to well preserved endoskeletons. Chondrichthyans are represented by the teeth of the symmoriid Stethacanthus sp., the ctenacanthid Glikmanius occidentalis (Leidy, 1859), the new euselachian described here, the eugeneodontids Campodus krasnopolskyi Kozlov, 2000 and Uralodus zangerli Kozlov, 2000, petalodontids including Tanaodus sp. and Permopetalodus frederixi Kozlov, 2000, orodontids and helodontids; the symphyseal tooth-whorls of the eugeneodontiforms Helicoprion bessonovi Karpinsky, 1899 and Parahelicoprion clerci (Karpinsky, 1916); sphenacanthid fin spine; fragment of hybodontoid skeleton including parts of the neurocranium, jaw, gill arch elements, teeth and fin spines; jaws, pectoral fin skeleton and tooth plates of a cochliodontid; “Listracanthus”-type spines (Kozlov 2000; Ivanov 2013, 2014).
The teeth of the new euselachian shark described here are scarce components in the palaeontologically richest interval in the section. The total of 26 teeth comes from two collections. The first collection was made by Olga V. Abrosimova, AOI, and SVN. The second collection was made by Vladimir A. Kozlov and passed to CJD for description in 2002.
Fig. 1. Maps of Ural (A) and Krasnoufimsk area (B; source of the map from https://yandex.ru/maps/) showing the location of Krasnoufimskie Klyuchiki quarry.
Systematic palaeontology
Class Chondrichthyes Huxley, 1880
Subclass Elasmobranchii Bonaparte, 1838
Cohort Euselachii Hay, 1902
Order and family incertae sedis
Genus Artiodus Ivanov and Duffin nov.
Etymology: From the Arti River, the stratotype area for Artinskian deposits, and Greek όδούς, tooth.
Type species: Artiodus prominens Ivanov and Duffin gen. et sp. nov.; see below.
Diagnosis.—As for species, by monotypy.
Stratigraphic and geographic range.—Late Artinskian (early Permian) of the Middle Urals (Russia).
Artiodus prominens Ivanov and Duffin sp. nov.
Figs. 2–5.
Etymology: From Latin prominens, projecting.
Holotype: PM SPU 81-1, isolated tooth (Fig. 2A).
Type locality: Krasnoufimskie Klyuchiki quarry, town of Krasnoufimsk, Sverdlovsk Region, Middle Urals, Russia.
Type horizon: Divya Formation, Sarginskian Regional Stage, late Artinskian, early Permian.
Material.—14 teeth (PM SPU 81-1–14) and 12 teeth (NHMUK PV P65426, P65427, P65450–59).
Diagnosis.—Multicuspid teeth with pyramidal, flattened labio-lingually, orthodont crown bearing from four to nine triangular cusps: large central, moderate intermediate and small lateral; well-developed lateral carina; strongly developed, prominent labial projection terminating in a large round tubercle and separated from the crown and base surfaces by a depression; distinct ornamentation from straight or recurved cristae of different length; oval or semilunar, elongate, considerably vascularized base with extended lingual part; dense vascular network formed of transverse horizontal, ascending, short secondary and semicircular canals.
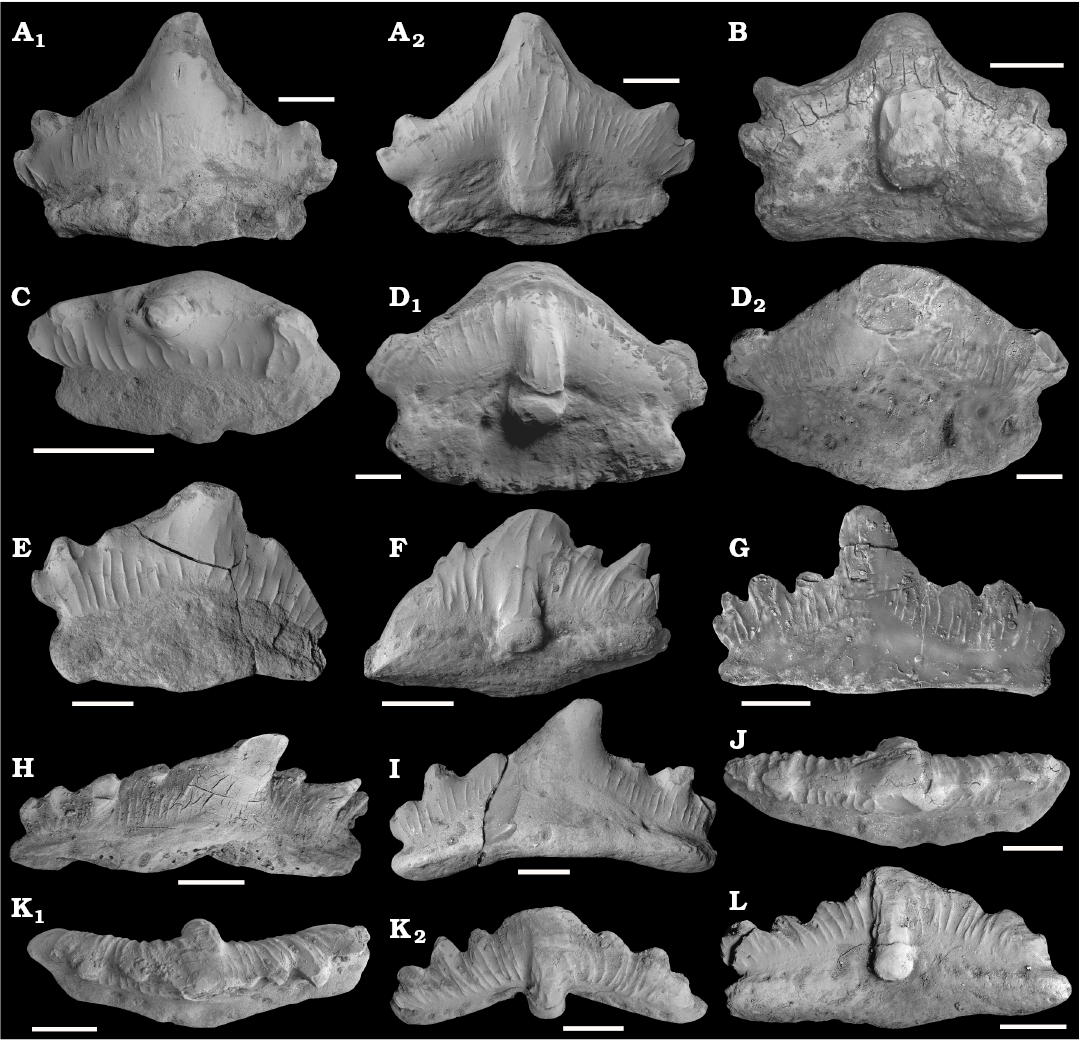
Fig. 2. Teeth of the euselachian shark Artiodus prominens Ivanov and Duffin gen. et sp. nov. from the Artinskian (Early Permian) of Krasnoufimskie Klyuchiki quarry (Middle Urals, Russia). A. PM SPU 81-1, holotype, lingual (A1) and labial (A2) views. B. PM SPU 81-2, labio-basal view. C. PM SPU 81-3, occlusal view. D. PM SPU 81-4, labio-basal (D1) and lingual (D2) views. E. PM SPU 81-5, lingual view. F. PM SPU 81-6, labio-basal view. G. PM SPU 81-7, lingual view. H. PM SPU 81-8, lingual view. I. PM SPU 81-9, lingual view. J. PM SPU 81-10, occlusal view. K. PM SPU 81-11, occlusal (K1) and labial (K2) views. L. PM SPU 81-12, labio-basal view. Scale bars 1 mm. All SEM images.
Description.—The teeth range in size from 2 to 7 mm mesio-distally. The teeth have a multicuspid, pyramidal crown and a base (or root) which is extended lingually (Figs. 2D, E, 3G, I–L). The crown is labio-lingually compressed, elongated mesio-distally, slightly inclined lingually (Fig. 2C), and bears distinct cusps, separated from each other by notches, and fused basally (Figs. 2, 3). The central cusp is considerably higher and wider than the moderate intermediate and small lateral cusplets. The height of the cusps decreases mesially and distally away from the central cusp, but some intermediate cusps are slightly higher than surrounding intermediate or lateral ones (Figs. 2F, 3A, D, F). A number of cusps in the crown varies from four to nine but is usually five or eight.
The crown is either symmetrical, with same number of intermediate and lateral cusps on both mesial and distal sides of the crown (e.g., Figs. 2A, 3G), or asymmetrical. In asymmetrical teeth, the central cusp is distally inclined and flanked by a greater number of mesial cusps than distal ones (e.g., 4 mesial cusps and 3 distal cusps in NHMUK PV P65450; Fig. 3D). The cusps are triangular in labial and lingual views, oval in cross section, and a lateral carina or occlusal crest separates the labial from the lingual face of the crown, forming a moderately well-developed cutting edge. The lateral cusps are often incurved (Fig. 3J). In symmetrical teeth, their long axes usually diverge from that of the central cusp (Fig. 2A, B). In asymmetrical teeth, the long axes of the lateral and intermediate cusps are usually sub-parallel to that of the central cusp (Figs. 2F, 3A, C, D), especially mesially.
The labial side of the teeth bears a strongly developed projection restricted to the lower part of the crown at the base of the central cusp, and continues to the base. This labial projection is separated from the labial face of the teeth by deep grooves on either side, and terminates in a large rounded tubercle sharply separated from the mesial and distal flanks of the crown and base (Fig. 2B, D1, F, L). The tubercle is made up mainly of coronal tissues, but the basal part is formed by a thin sliver of base tissue. The two tissues meet at a flat crest which runs circumferentially around the tubercle low down on the tubercle margin (Fig. 5A, D). The boundary between the crown and the base, or the crown/root junction, is distinct, and marked by a groove on both labial and lingual sides; as described above, the junction can also be traced around the labial projection.
The ornamentation of the crown comprises relatively coarse cristae or vertical ridges of varying length, following trajectories that may be straight, curved or wavy. The labial face of the crown is generally more heavily and densely ornamented than the lingual face, with the longest cristae generally being located toward the central sections of the cusps. In many specimens, the ridges begin just above the crown/root junction and ascend the crown toward the cusp apices or occlusal crest. In some cases the ridges attain the apices of the central and lateral cusps (Figs. 2E–G, K, L, 3), while in many others they terminate approximately two-thirds the way up the crown (Figs. 2A1, C, 3B). The coarser ridges may bifurcate basally (Fig. 2A2, K2). Short, impersistent cristae are often interspersed between the coarser examples, and may be confined to the upper parts of the crown (Fig. 2A). A very coarse ridge commonly surmounts the crest of the labial projection either in a central position (Figs. 2F, 3A, D) or rather offset laterally, towards the flanks of the projection (Fig. 2A2, K2). Short ridges also adorn the lateral margins of the labial projection (Figs. 2A2, D1, F, K2, 3A–F). The upper part of the central cusp on the lingual side is frequently smooth (Fig. 2A1, C, E, I).
The tooth base is oval or semilunar in outline, elongated mesio-distally, arched, convex on the occlusal side and concave on the basal one (Figs. 2B, D1, I, K2, 5D). It is lingually offset from the crown underside. The base bears the numerous large and small foramina marking the entrances and exits of the vascular canals which perforate the base. The foramina tend to be more numerous on the labial side, where they are commonly organized into a longitudinal row just beneath the crown/base junction (Figs. 2F, L, 3A–F). Lingually, a relatively large median foramen may be present on the linguo-basal border beneath the central cusp (Fig. 3J, K). Larger foramina are also present on the mesial and distal flanks of the lingual face of the base, often interspersed with smaller examples (Figs. 3J, K, 5B). The foramina on both labial and lingual walls of the base enter the base tissues horizontally. A few foramina open onto the basal surface of the base.

Fig. 3. Teeth of the euselachian shark Artiodus prominens Ivanov and Duffin gen. et sp. nov. from the Artinskian (Early Permian) of Krasnoufimskie Klyuchiki quarry (Middle Urals, Russia). A. NHMUK PV P65426. B. NHMUK PV P65452. C. NHMUK PV P65427. D. NHMUK PV P65450. E. NHMUK PV P65451. F. NHMUK PV P65455. G. NHMUK PV P65457. H. NHMUK PV P65456. I. NHMUK PV P65458. J. NHMUK PV P65454. K. NHMUK PV P65453. L. NHMUK PV P65459. A–F, labial views; G–L, lingual views. Scale bars 1 mm.
Tooth histology: The crown is composed of orthodentine with numerous, strongly branched dentine tubules and a shiny layer of enameloid (Fig. 4B). The base contains considerably vascularized trabecular dentine but the labial projection comprises compact dentine which lacks canals and dentine tubules. The vascularization system consists of a dense network of internal canals occupying the whole tooth base. The vascularization network comprises transverse horizontal, ascending, secondary and semicircular canals with small ramifications (Fig. 5C–H). The horizontal canals include a wide main canal and narrower moderate canals. They pass across the base from the lingual torus to the labial groove. The ascending canals are vertical or inclined; the main, central is wide and, together with narrower lateral canals occupies and fills the upper part of the base. Two main, vertical ascending canals start their trajectories from the mid basal side of the base and run to the crown/base junction under the central cusp. Some inclined ascending canals open on the lingual side of the base at the crown/base junction. The short secondary canals are connected the horizontal canals by means of ascending canals. A narrow circular canal is confined to the labial projection and opens in the grooves forming the flanks of the projection.
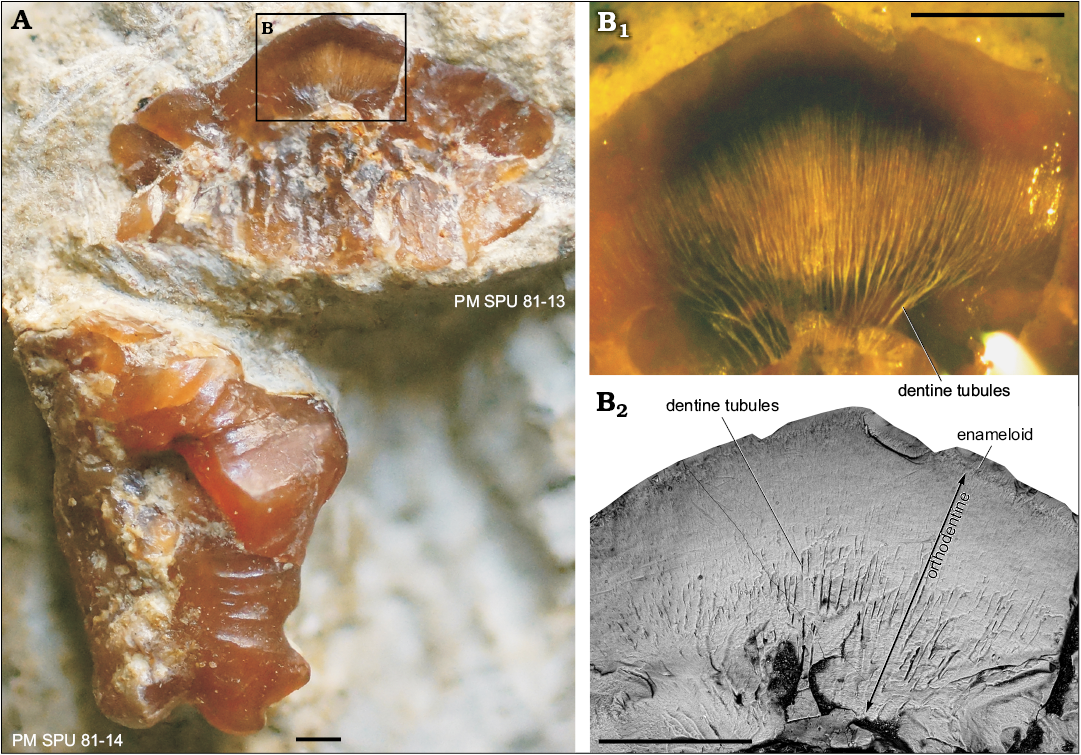
Fig. 4. Teeth of the euselachian shark Artiodus prominens Ivanov and Duffin gen. et sp. nov. from the Artinskian (Early Permian) of Krasnoufimskie Klyuchiki quarry (Middle Urals, Russia). A. PM SPU 81-13, tooth with broken crown in the longitudinal section in lingual view and PM SPU 81-14 in labial view. B. PM SPU 81-13, details of the crown; optical microscope photograph in aniseed oil (B1) and SEM microphotograph (B2). Scale bars 500 µm.
Variation and heterodonty: The teeth of the new taxon vary in the height of central cusp, the number of intermediate and lateral cusp pairs, the symmetry of the crown, the degree of cusp inclination, and the development of crown ornamentation. The teeth can be separated into three morphotypes based on the known dentitions of euselachians such as Hamiltonichthys mapesi Maisey, 1989 (Maisey 1989), Hybodus basanus Egerton, 1845 (Maisey 1983), Hybodus reticulatus Agassiz, 1837 (Maisey 1987), and Hybodus brevicostatus Patterson, 1966 (Patterson 1966).
Morphotype 1 (four teeth): symmetrical teeth with the same number of intermediate and lateral cusplets on both mesial and distal flanks of the crown. The central cusp is very high with a wide base, and is flanked by one pair of intermediate cusps and one pair of lateral ones. The labial projection is relatively wide. The mesial and distal flanks of the crown extend beyond the limits of the base, meaning the crown is wider mesio-distally than the base. The latter projects very strongly in a lingual direction from the crown underside (Figs. 2A, B, D1, D2, 3L). The height of the crown is up to four times the width of the base. These teeth probably come from a symphyseal or anterior position in the dentition, based on their symmetrical form and similar patterns of heterodonty in the dentitions of relatively high-crowned hybodontid sharks such as H. basanus, H. brevicostatus, and H. reticulatus (Maisey 1983, 1987; Patterson 1966).
Morphotype 2 (two teeth): comprises smaller teeth in our collection, possessing a slightly asymmetrical crown with a short central cusp, flanked by only one pair of intermediate cusps. The labial projection, although prominent, is narrow, and lingual extension of the base is moderately well developed (Fig. 2C, E). The height of the crown and the width of the base are sub-equal. By reference to the heterodonty shown by relatively well-known high-crowned hybodont dentitions, these teeth are probably antero-laterals or posteriors.
Morphotype 3 (20 teeth): These teeth are characterized by a more strongly asymmetrical crown with distally inclined cusps. The central cusp is of moderate size and flanked by two or three intermediate cusps and one lateral cusp pair. The number of mesial intermediate cusps is greater than the number of distal cusps. The labial projection is narrow and very prominent. The lingual extension of the base is relatively short (Figs. 2F–L, 3A–K, 4). Bearing in mind the heterodonty displayed in certain hybodont dentitions, these teeth are probably lateral and postero-lateral teeth.
Although based upon a relatively small number of isolated teeth, the pattern of moderate heterodonty suggested here for Artiodus prominens is an example of simple linear gradient monognathic heterodonty (Duffin and Ward 1983) in which the following changes take place distally through the dentition of a single jaw ramus: (i) symmetrical symphyseal teeth give way to successively more asymmetrical forms distally; (ii) the central cusp is upright in symphyseal teeth, becoming more distally inclined away from the symphysis; (iii) relative to overall tooth length, the height of the central cusp reduces distally through the dentition; (iv) the number of intermediate cusps increases from one to three pairs; and (v) the labial projection becomes narrower.
The collection of teeth available to use shows no evidence of dignathic, gynandric or ontogenetic heterodonty. A greater number and morphological range of isolated teeth, or fully articulated dentitions of Artiodus are needed for better definition of the types of heterodonty which might be displayed in this taxon.
Stratigraphic and geographic range.—Late Artinskian (early Permian) of the Middle Urals (Russia).
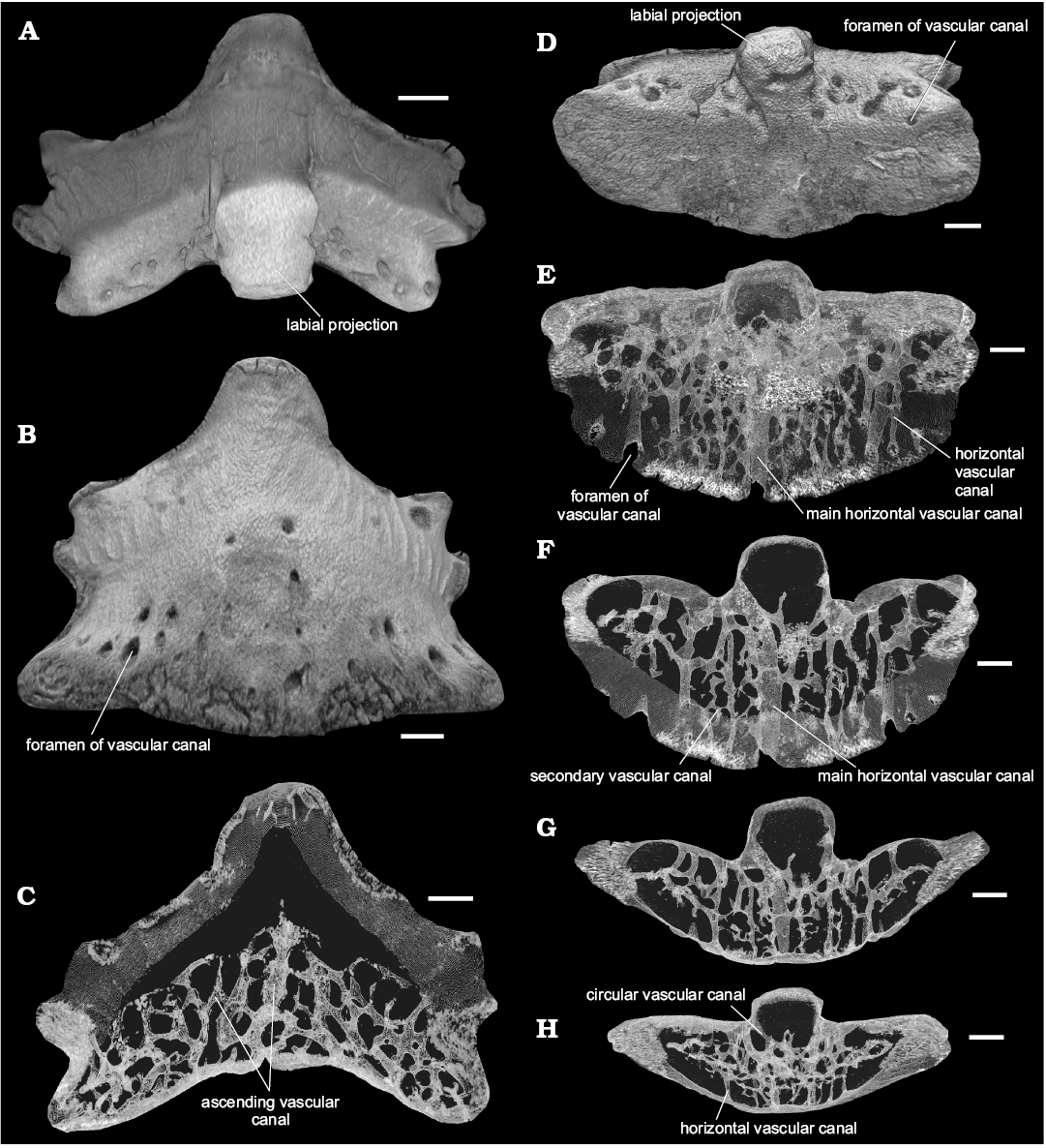
Fig. 5. Tooth (PM SPU 81-2) of the euselachian shark Artiodus prominens Ivanov and Duffin gen. et sp. nov. from the Artinskian (Early Permian) of Krasnoufimskie Klyuchiki quarry (Middle Urals, Russia). Micro-CT virtual model in labial (A), lingual (B), and basal (D) views. Virtual sections: longitudinal section of the tooth (C); section series of the base (F–H). Virtual model of the tooth in basal view with transparent dental tissues (E). Scale bars 500 µm.
Concluding remarks
Dentition of Artiodus.—The teeth of euselachians share a highly vascularized base and possess crowns displaying different types of consolidation. The three types of tooth crown based on degree of cusp fusion and crown consolidation which have been suggested for euselachian teeth are: hybodont, protacrodont, and orodont (Ginter et al. 2010). The last of these, comprising a completely consolidated crown lacking separated cusps is perhaps more correctly named “polyacrodont” for the euselachian teeth. The teeth of Artiodus gen. nov. differ from the teeth of all other known euselachians in the unique structure of the labial projection which projects from the lower part of the crown to the base and has a distinctive histology. The incorporation of a thin strip of base tissue into the underside of the labial projection is a condition known in only one other euselachian, so far as we are aware—Reesodus wirksworthensis (Duffin, 1985) from the early Carboniferous of Great Britain (Duffin 1985). The genus Reesodus was erected for teeth described from the middle Permian of Oman as the new species, R. underwoodi Koot, Cuny, Tintori, and Twitchett, 2013 (Koot et al. 2013). These authors included Reesodus wirksworthensis (Duffin, 1985) and R. pectinatus (Lebedev, 1996) into the genus from Lissodus, an association which we consider to be highly doubtful. In spite of the incorporation of base tissue onto the lower part of the labial projection in both Artiodus and R. wirksworthensis, the two taxa are clearly different on a number of counts; Artiodus is high-crowned whereas R. wirksworthensis in consistently low-crowned throughout the dentition and the tooth proportions and ornamentation are completely different in the two taxa.
The teeth of Artiodus prominens Ivanov and Duffin gen. et sp. nov. otherwise most closely resemble the teeth of some protacrodontid and sphenacanthid euselachians possessing a protacrodont-type crown; the cusps on the pyramidal crown are separated from each other by deep notches. The teeth of neither protacrodontids nor sphenacanthids develop a labial projection, however. The possible hybodontiform Roongodus phijani Hairapetian and Ginter, 2009 has teeth with a prominent lingual projection (Hairapetian and Ginter 2009). The teeth of some polyacrodontids and lonchidiids also characteristically possess a labial projection (peg) (Duffin 2001; Ginter et al. 2010; Cappetta 2012). The projection found in the teeth of Artiodus, however, is unique in being strongly accentuated basally. In addition, the crowns of polyacrodontid and lonchidiid teeth are more consolidated: the cusps are fused and not separated from each other by deep intervening notches (see for example, Reesodus; Koot et al. 2013: fig. 7). Furthermore, the height of the crown and the width of the base are virtually equal to each other throughout the dentition in polyacrodontid and lonchidiid sharks, but teeth from putative anterior positions in the dentition of Artiodus have crowns which are more extended than the base.
The dentition of Artiodus prominens Ivanov and Duffin gen. et sp. nov. displays moderate heterodonty. Complete dentitions are relatively rare among known Palaeozoic euselachians but include the Devonian Protacrodus; the Carboniferous Dabasacanthus, Hamiltonichthys, Onychoselache, and Tristychius, and the Permian Gansuselache (Ginter et al. 2010). Whilst some heterodonty is observed in the dentition of Hamiltonichthys (Maisey 1989) and Onychoselache (Dick and Maisey 1980), the tooth morphotypes in the jaws of these taxa are not strongly differentiated from each other as in Artiodus. In the Palaeozoic, only representatives of lonchidiids and polyacrodontids known from isolated teeth (Johnson 1981; Duffin 2001) possess heterodonty resembling the condition described above for Artiodus. The range of morphologies otherwise displayed by the collection of teeth from the Permian of the Urals shows the greatest similarity to that described for the dentitions of relatively high-crowned hybodontids from the Mesozoic.
The assemblage of Artinskian chondrichthyans.—Early Permian chondrichthyans are quite diverse in Palaeozoic fish faunas. The highest chondrichthyan taxonomic diversity is found in the Artinskian, especially from the localities of the Middle and South Urals (Ivanov 2005). The chondrichthyan assemblages from the Artinskian of the Middle and South Urals include numerous elasmobranch and euchondrocephalian taxa, such as symmoriiforms Cobelodus sp., Denaea cf. D. wangi Wang, Jin, and Wang, 2004, Kungurodus obliquus (Ivanov, 2005), Stethacanthus altonensis (St. John and Worthen, 1875), Stethacanthulus decorus (Ivanov, 1999); ctenacanthiforms Glikmanius occidentalis (Leidy, 1859), Heslerodus sp.; the jalodontid Adamantina foliacea Ivanov, 1999; euselachians: Artiodus prominens Ivanov and Duffin gen. et sp. nov., Sphenacanthus sp., the lonchidiid “Lissodus” sp., the polyacrodontid “Polyacrodus” sp.; neoselachians—the synechodontiform Synechodus antiquus Ivanov, 2005, anachronistids Cooleyella amazonensis Duffin, Richter, and Neis, 1996, Cooleyella cf. C. fordi (Duffin and Ward, 1985), Cooleyella sp.; the petalodontiforms Permopetalodus frederixi Kozlov, 2000, Tanaodus sp., Chomatodus sp.; eugeneodontiforms Helicoprion bessonovi Karpinsky, 1899, Parahelicoprion clerci (Karpinsky, 1916), Campodus krasnopolskyi Kozlov, 2000, Campodus sp., Uralodus zangerli Kozlov, 2000; Orodus sp.; a helodontid; the holocephalians Psephodus sp. and a cochliodontid.
The Artinskian chondrichthyan assemblage contains many taxa with a wide geographical distribution, being found in many regions of the world, as well as several endemic forms (Artiodus prominens Ivanov and Duffin gen. et sp. nov., Permopetalodus frederixi, Parahelicoprion clerci, Campodus krasnopolskyi, Uralodus zangerli). Some taxa from the assemblage such as Cobelodus, Denaea, Stethacanthus, Stethacanthulus, Glikmanius, Heslerodus, Adamantina, Sphenacanthus, Cooleyella, Tanaodus, Chomatodus, Campodus, Orodus, Psephodus appeared in the Carboniferous and persisted into the early Permian. Other taxa (oldest synechodontiform Synechodus antiquus, euselachian Artiodus prominens, petalodontiform Permopetalodus frederixi, and eugeneodontiforms Helicoprion bessonovi, Parahelicoprion clerci, Campodus krasnopolskyi, and Uralodus zangerli) occur only in the Artinskian Stage. The taxonomic diversity of chondrichthyans increased during the middle Permian of Urals and the East European Platform, although some groups (symmoriiforms, ctenacanthiforms, anachronistids, petalodontiforms, helodontiforms, orodontiforms, and others) are quite rare or even disappeared altogether. A few Artinskian chondrichthyan taxa (Sphenacanthus, “Lissodus”, “Polyacrodus”, Synechodus) persisted to the end of the Permian. The early Permian fish faunas of the Urals do not contain xenacanthimorphs, which were abundant in assemblages of the same age from Western Europe and North America, but do include diverse symmoriiforms, euselachians and edestids.
Acknowledgements
AOI is most grateful to Olga Abrosimova (Krasnoufimsk, Russia) for assistance in the collecting the teeth in the field, Roman Rakitov (Borissiak Paleontological Institute of the Russian Academy of Sciences, Moscow) for assistance during SEM imaging and Sergey Nilov (Institute of the Earth Sciences, St. Petersburg State University, Russia) for the microtomographic sections and reconstruction. The scientific research completed by AOI was performed at the Center for Geo-Environmental Research and Modeling (GEOMODEL) of the Research Park of St. Petersburg State University. CJD would like to thank Tom Davies and Mike Benton (both of Bristol University, UK) for the use of the Leica photomicroscope. We thank Michał Ginter (Warsaw University, Poland) and anonymous reviewer for their helpful reviews of the manuscript. The study was partly supported by the Russian Foundation for Basic Research, project No. 14-04-01507a; by a grant from the Saint-Petersburg State University No. 0.38.292.2015. The work has been performed according to the Russian Government Program of Competitive Growth of Kazan Federal University.
References
Agassiz, J.L.R. 1833–1843. Recherches sur les Poissons Fossiles. 5 volumes and atlas. 422 pp. Imprimerie de Petitpierre, Neuchâtel.
Cappetta, H. 2012. Chondrichthyes. Mesozoic and Cenozoic Elasmobranchii: Teeth. In: H.-P. Schultze (ed.), Handbook of Palaeoichthyology, Volume 3E, 1–512. Verlag Dr. Friedrich Pfeil, München.
Chuvashov, B.I. [Čuvašov, B.I.] and Dyupina, G.V. [Dûpina, G.V.] 1973. Verhnepaleozojskie terrigennye otloženiâ zapadnogo sklona Srednego Urala. 218 pp. Nauka, Mokva.
Chuvashov, B.I. [Čuvašov, B.I.], Dyupina, G.V. [Dûpina, G.V.], Mizens, G.A., and Chernykh, V.V. [Černyh, V.V.] 1990. Opornye razrezy verhnego karbona i rannej permi zapadnogo sklona Urala i Preduralâ. 369 pp. Ural Branch of Academy of Sciences Publishing House, Sverdlovsk.
Dick, J.R.F. and Maisey, J.G. 1980. The Scottish Lower Carboniferous shark Onychoselache traquairi. Palaeontology 23: 363–374.
Duffin, C.J. 1985. Revision of the hybodont selachian genus Lissodus Brough (1935). Palaeontographica Abteilung A 188: 105–152.
Duffin, C.J. 2001. Synopsis of the selachian genus Lissodus Brough, 1935. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 221: 145–218.
Duffin, C.J. and Ward, D.J. 1983. Neoselachian sharks teeth from the Lower Carboniferous of Britain and the Lower Permian of the U.S.A. Palaeontology 26: 93–110.
Ginter, M., Hampe, O., and Duffin, C.J. 2010. Chondrichthyes Paleozoic Elasmobranchii: Teeth. In: H.-P. Schultze (ed.), Handbook of Palaeoichthyology, Volume 3D, 1–168. Verlag Dr. Friedrich Pfeil, München.
Hairapetian, V. and Ginter, M. 2009. Famennian chondrichthyan remains from the Chahriseh section, central Iran. Acta Geologica Polonica 59: 173–200.
Ivanov, A. 2005. Early Permian chondrichthyans of the Middle and South Urals. Revista Brasileira de Paleontologia 8: 127–138. Crossref
Ivanov, A.O. 2013. A new findings of the Early Permian shark endoskeleton [in Russian]. In: S.V. Naugolnyh (ed.), Obekty Paleontologičeskogo i Geologičeskogo Naslediâ i Rol' Muzeev v Ih Izučenii i Ohrane, 35–41. Kungur Historical-Architecture and Art Museum, Kungur.
Ivanov, A.O. 2014. Artinskian radiation of chondrichthyan fishes. In: D.K. Nurgaliev (ed.), International Kazan Golovkinsky Stratigraphic Meeting, Kazan, Russia, Abstract, 41–42. Kazan Federal University Publishing House, Kazan.
Johnson, G.D. 1981. Hybodontoidei (Chondrichthyes) from the Wichita-Albany Group (Early Permian) of Texas. Journal of Vertebrate Paleontology 1: 1–41. Crossref
Johnson, G.D. 1996. Vertebrate microfossils from the Lueders Formation, Albany Group, and the faunal transition from the Wichita Group into the Clear Fork Group, Lower Permian of Texas. Modern Geology 20: 371–382.
Karpinsky, A.P. 1899. Uber die Reste von Edestiden und die neue Gattung Helicoprion. Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft, Series 2 36: 361–475.
Koot, M.B., Cuny, G., Tintori, A., and Twitchett, J.J. 2013. A new diverse shark fauna from the Wordian (Middle Permian) Khuff Formation in the interior Haushi-Huqf area, Sultanate of Oman. Palaeontology 56: 303–343. Crossref
Kozlov, V.A. 2000. About new findings of sharks’ teeth in the Artinskian deposits of the Cisurals [in Russian]. In: B.I. Čuvašov (ed.), Materialy po stratigrafii i paleontologii Urala, 4, 148–153. Ural Branch of Academy of Sciences Publishing House, Ekaterinburg.
Maisey, J.G. 1983. Cranial anatomy of Hybodus basanus Egerton from the Lower Cretaceous of England. American Museum Novitates 2758: 1–64.
Maisey, J.G. 1987. Cranial anatomy of the Lower Jurassic shark Hybodus reticulatus (Chondrichthyes: Elasmobranchii), with comments on hybodontid systematics. American Museum Novitates 27 878: 1–39.
Maisey, J.G. 1989. Hamiltonichthys mapesi, g. & sp. nov. (Chondrichthyes; Elasmobranchii), from the Upper Pennsylvanian of Kansas. American Museum Novitates 2931: 1–42.
Naugolnykh, S.V. 2001. Morphology and systematics of representatives of Vojnovskyales. Paleontological Journal 35: 545–556.
Naugolnykh, S.V. 2014. A new genus of male cones of voltzialean affinity, Uralostrobus voltzioides nov. gen., nov. sp., from the Lower Permian of the Urals (Russia). Geobios 47: 315–324. Crossref
Patterson, C. 1966. British Wealden sharks. Bulletin of the British Museum (Natural History), Geology 2 (7): 283–359.
Yankevich, D.I. [Ânkevič, D.I.] 2001. New actinopterygian fishes from the Permian Artinskian and Ufimian deposits of the western Cisurals and their stratigraphical significance [in Russian]. In: B.I. Čuvašov (ed.), Materialy po stratigrafii i paleontologii Urala, 6, 100–105. Ural Branch of Academy of Sciences Publishing House, Ekaterinburg.
Zangerl, R. 1981. Chondrichthyes I. Paleozoic Elasmobranchii. In: H.-P. Schultze (ed.), Handbook of Palaeoichthyology, Volume 3A, 1–115. Gustav Fischer Verlag, Stuttgart.
Acta Palaeontol. Pol. 62 (2): 289–298, 2017
https://doi.org/10.4202/app.00347.2017