
Sauropodomorph dinosaur trackways from the Fleming Fjord Formation of East Greenland: Evidence for Late Triassic sauropods
JENS N. LALLENSACK, HENDRIK KLEIN, JESPER MILÀN, OLIVER WINGS, OCTÁVIO MATEUS, and LARS B. CLEMMENSEN
Lallensack, J.N., Klein, H., Milán, J., Wings, O., Mateus, O., and Clemmensen, L.B. 2017. Sauropodomorph dinosaur trackways from the Fleming Fjord Formation of East Greenland: Evidence for Late Triassic sauropods. Acta Palaeontologica Polonica 62 (4): 833–843.
The Late Triassic (Norian–early Rhaetian) Fleming Fjord Formation of central East Greenland preserves a diverse fossil fauna, including both body and trace fossils. Trackways of large quadrupedal archosaurs, although already reported in 1994 and mentioned in subsequent publications, are here described and figured in detail for the first time, based on photogrammetric data collected during fieldwork in 2012. Two trackways can be referred to Eosauropus, while a third, bipedal trackway may be referred to Evazoum, both of which have been considered to represent sauropodomorph dinosaur tracks. Both the Evazoum and the Eosauropus trackways are distinctly larger than other trackways referred to the respective ichnogenera. The trackmaker of the best preserved Eosauropus trackway is constrained using a synapomorphy-based approach. The quadrupedal posture, the entaxonic pes structure, and five weight-bearing digits indicate a derived sauropodiform trackmaker. Other features exhibited by the tracks, including the semi-digitigrade pes and the laterally deflected unguals, are commonly considered synapomorphies of more exclusive clades within Sauropoda. The present trackway documents an early acquisition of a eusauropod-like pes anatomy while retaining a well-developed claw on pedal digit IV, which is reduced in eusauropods. Although unequivocal evidence for sauropod dinosaurs is no older than the Early Jurassic, the present trackway provides evidence for a possible Triassic origin of the group.
Key words: Dinosauria, Sauropodomorpha, trackway, Triassic, Greenland.
Jens N. Lallensack [jens.lallensack@uni-bonn.de], Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Rheinische Friedrichs-Wilhelm Universität Bonn, Nußallee 8, 53115 Bonn, Germany.
Hendrik Klein [Hendrik.Klein@combyphone.eu], Saurierwelt Paläontologisches Museum, Alte Richt 7, D-92318 Neumarkt, Germany.
Jesper Milàn [jesperm@oesm.dk], Geomuseum Faxe/Østsjællands Museum. Østervej 2, DK-4640 Faxe, Denmark, and Natural History Museum of Denmark, Øster Voldgade 5-7, DK-1350 Copenhagen, Denmark.
Oliver Wings [dr.wings@gmail.com], Zentralmagazin Naturwissenschaftlicher Sammlungen (ZNS), Martin-Luther-Universität Halle-Wittenberg, Domplatz 4, 06108 Halle (Saale), Germany.
Octávio Mateus [omateus@fct.unl.pt], GEOBIOTEC, Faculdade de Ciências e Tecnologia, FCT, Universidade Nova de Lisboa, 2829-526 Caparica, Portuga, and Museu da Lourinhã, Rua João Luís de Moura, 2530-157 Lourinhã, Portugal.
Lars B. Clemmensen [larsc@ign.ku.dk], Department of Geosciences and Natural Resource Management, University of Copenhagen, Øster Voldgade 10, DK-1350 Copenhagen, Denmark.
Received 21 April 2017, accepted 28 June 2017, available online 4 September 2017.
Copyright © 2017 J.N. Lallensack et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Triassic body fossil record of sauropods is extremely poor. Although Isanosaurus from the Nam Phong Formation of Thailand had been described as the oldest known sauropod (Buffetaut et al. 2000), its proposed late Norian or Rhaetian age was recently questioned, possibly placing the fossil within the Jurassic (Racey and Goodall 2009; McPhee et al. 2015). The sauropod status of Late Triassic forms such as Antetonitrus is unclear, given current disagreements on the taxonomic definition of Sauropoda (McPhee et al. 2014). Thus, undisputed evidence for Sauropoda sensu stricto from the Triassic is currently lacking, with some authors arguing that sauropods diverged from other sauropodomorphs as late as close to the Triassic–Jurassic boundary (McPhee et al. 2015).
Jenkins et al. (1994) briefly mentioned the occurrence of three large, quadrupedal trackways referable to large archosaurs from the Late Triassic (Norian–Rhaetian) Fleming Fjord Formation of East Greenland. The best preserved of these trackways (herein, trackway S1) was noted to show “four clawed digits, oriented anterolaterally” and “small, crescentic impressions of the manus” (Jenkins et al. 1994: 19), and included in a sitemap showing its position amongst multiple smaller, bipedal trackways referred to the ichnogenus Grallator. These large trackways were frequently mentioned in subsequent literature, but have never been described or illustrated in detail. Lockley and Meyer (2000) suggested that these tracks are attributable to basal sauropodomorph (“prosauropod”) dinosaurs, and possibly to the ichnogenus Tetrasauropus. Lockley et al. (2001, 2006a) mention that the trackways might be those of sauropod trackmakers, with Lockley et al. (2006a) stating that the tracks are similar to those referred to the ichnogenus Eosauropus. Fieldwork carried out in 2012 was able to relocate two of the three trackways (Sulej et al. 2014; Clemmensen et al. 2016). The better-preserved trackway was identified as that of a biped with “rounded pes and weak indications of outwards-rotated claws”, and “a weak impression that could indicate the presence of a thin semilunate-shaped manus” (Clemmensen et al. 2016: 43). A second trackway, “quadrupedal, with an extreme degree of heteropody” and elongated pes imprints (Clemmensen et al. 2016: 43), was ascribed to a different type. A newly discovered third trackway showing tetradactyl pes prints was referred to cf. Evazoum, suggesting a basal sauropodomorph trackmaker (Sulej et al. 2014; Clemmensen et al. 2016). Klein et al. (2016) noted that, given the lack of a complete description and detailed figures, it cannot be excluded that these tracks might not belong to dinosaurs but to large chirotheriids. The purpose of the present paper is to describe these trackways in detail for the first time. In light of a possible sauropod affinity of the quadrupedal trackways, footprint features are compared with skeletal features of sauropodomorphs, allowing for trackmaker identification based on synapomorphies, and possibly providing additional data on the poorly known early evolutionary history of sauropods.
Other abbreviations.—LM, left manus; LP, left pes; PL, pes length; RM, right manus; RP, right pes; WAP, width of the angulation pattern.
Geological setting
Lake deposits of the Late Triassic Fleming Fjord Formation of central East Greenland have yielded diverse vertebrate remains, including both body and trace fossils. Body fossils include selachians, bony fishes, temnospondyl amphibians, mammaliaforms, possible sphenodonts and lepidosaurs, testudinatans, phytosaurs, aetosaurs, a possible rauisuchian, pterosaurs, and dinosaurs (theropods and basal sauropodomorphs), while the ichnofauna includes a large number of theropod-like (Grallator), sauropodomorph-like, and stem-crocodylian archosaur trackways (Sulej et al. 2014; Clemmensen et al. 2016; Klein et al. 2016). The three sauropodomorph-like trackways described herein come from the Track Mountain locality at Wood Bjerg-Macknight Bjerg (Fig. 1) along Carlsberg Fjord near the eastern margin of the basin (Clemmensen et al. 2016). Trackway S1 (71°24.853’ N, 22°33.322’ W; 534 m above sea level) and trackway S2 (71°24.955’ N, 22°32.952’ W) represent quadrupedal trackways, while trackway S3 (71°24.857’ N, 22°33.334’ W) is bipedal. The tracks are preserved as concave epireliefs on a thin, laterally extensive siltstone bed that is under- and overlain by red mudstone. The siltstone bed has a thickness between 1 and 5 cm, is multistorey and contains wave-formed structures including even lamination and tiny wave ripples. The upper surface is cut by polygonal mudcracks and reveals micro-ridges of uncertain origin. Tridactyl Grallator tracks are seen locally in close association with the sauropodomorph-like tracks. Selected high-resolution photographs showing the described trackways in the field are provided as SOM 1 (Supplementary Online Material available at http://app.pan.pl/SOM/app62-Lallensack_etal_SOM.pdf). During Late Triassic times, the study area was located at the northern rim of the Pangaean supercontinent at about 40 ͦ N (Clemmensen et al. 2016). The track-bearing siltstone bed is situated in the lowermost part of the Tait Bjerg Beds of the Ørsted Dal Member within the Fleming Fjord Formation (Clemmensen et al. 1998, 2016). The Ørsted Dal Member has been dated to the late Norian–early Rhaetian, based on invertebrate fossils, land-derived palynomorphs, and paleomagnetic data; the track-bearing bed probably formed around 208 mya (Clemmensen 1980; Kent and Clemmensen 1996; Clemmensen et al. 1998, 2016). The track-bearing part of the succession was deposited in an ephemeral to semi-perennial lake with annual and longer-term fluctuations in lake water. The siltstone bed records flooding events of mudflats by lake water (Clemmensen et al. 1998).
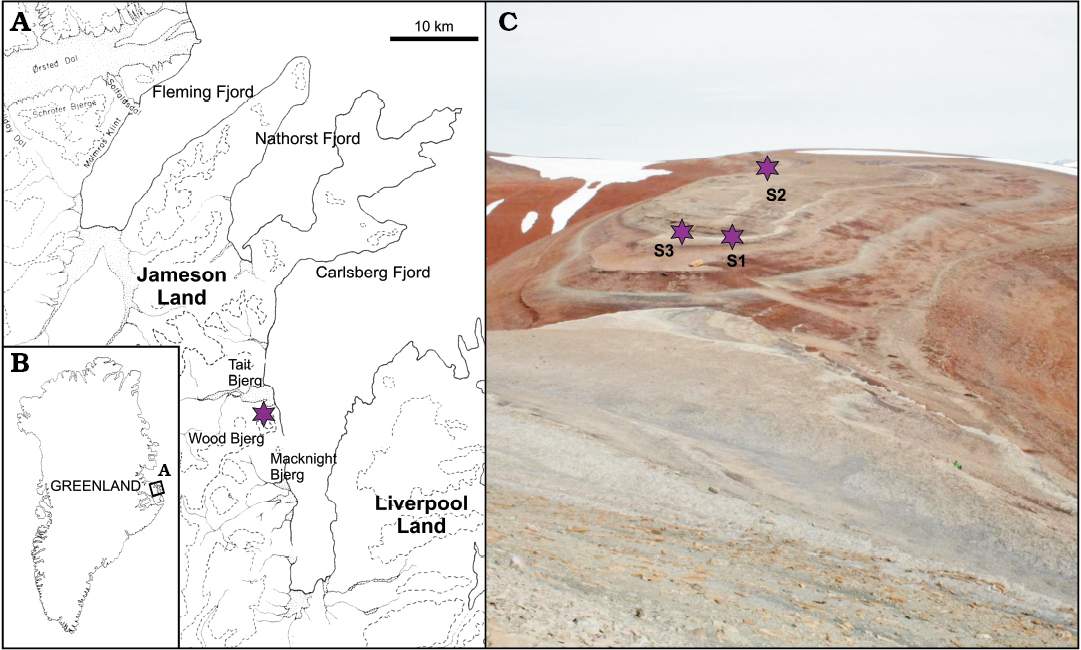
Fig. 1. A. Location of the “Track Mountain” locality (star) on a ridge on the northeastern slope of Wood Bjerg in the Late Triassic sediments at the west side of Carsberg Fjord. B. Location of Jameson Land (A) in central East Greenland. C. Photograph of the “Track Mountain” locality showing the approximate location of trackways S1, S2, and S3 (view towards the east).
Material and methods
The three trackways described herein (S1, S2, S3) were found in close proximity to each other (Fig. 1C). Descriptions and measurements are based on high-resolution photogrammetric 3D-models generated from photographs taken during fieldwork following the procedures described by Mallison and Wings (2014). Photographs of trackway S1 were taken by OW using a Canon EOS 30D digital SLR camera (8.19 Mpx), while those of trackways S2 and S3 were taken by JM using a Canon EOS 400D digital SLR camera (10 Mpx). Photogrammetric models were generated using Agisoft PhotoScan Professional 1.2.4 (www.agisoft.com). The horizontal plane of the model was automatically determined using the free software CloudCompare 2.6 (www.cloudcompare.org). Orthographic depth-color maps were generated using the free software Paraview 5.0 (www.paraview.org). Trackway parameters and footprint dimensions were calculated using standard trigonometric functions, based on xy-coordinates taken from the depth-color maps using the free software Inkscape 0.48 (www.inkscape.org). Measurements were taken according to Marty (2008). Terminology follows, until otherwise noted, that of Leonardi (1987). Photogrammetric models, including textured high-resolution .ply files and PDFs detailing the process of model generation, are provided as SOM 2–4.
Description
Trackway S1.—It consists of five consecutive pes and four manus impressions (Fig. 2, SOM 1, 2) leading towards the south-west. The layer preserving the first pes-manus set is destroyed by erosion in the remaining section of the trackway, revealing a second, slightly lower layer. It is unclear on which of these layers the animal walked on. Well preserved claw impressions might indicate that the footprints of the lower layer represent true tracks, with the more indistinct pes-manus set of the upper layer preserved as an overtrack. Alternatively, the lower layer could represent a subsurface layer preserving undertracks or deep tracks that recorded the foot anatomy more precisely than the, in this case incompetent, surface layer (Gatesy 2003; Milàn and Bromley 2006). The pes-manus sets RP1/RM1 and RP2/RM2 are the best preserved, while LP3 appears markedly more elongated than the remaining pes footprints, probably due to sediment drawn into the footprint during track formation. The trackway shows a narrow gauge (sensu Marty 2008), with a WAP/PL-ratio of 0.85. The pes pace angulation (128°) is slightly larger than the manus pace angulation (115°). The manus impressions RM1 and RM2 are broader than long and gently arched with a concave posterior margin, although this could be the result of overstepping by the pes. Clear manus digit impressions are not identifiable, possibly due to imperfect preservation. The right manus impressions are directed anteriorly, while the left ones are slightly rotated outwards. Heteropody, i.e., the ratio between the pes and manus footprint area, is about 1:3. The pes footprints, measuring 42 cm in length on average, are oval in shape and consistently rotated outwards by about 30°. Five digit impressions can be observed in the pes. The trace of digit I can be clearly identified only in RP2, where it appears broader and more protruding than the remaining digit impressions, leaving a pronounced displacement rim. In RP1, the trace of digit I is indistinct and shorter than the trace of digit II, either because of incomplete preservation, or because the ungual of digit I was deflected laterally or postero-laterally due to strong flexion, and therefore left no trace. Claw marks of digits II–IV are well preserved in RP1 and RP2, extending from the anterior half of the lateral margin of the footprint, directed laterally with respect to the long axis of the footprint and posterolaterally with respect to the direction of travel. The claw marks probably represent the impressions of the medial sides of the claws, as indicated by their broad bases and curved tips, and not the ventral sides as would be expected in anteriorly directed claws. A small semi-circular bulge, which is well separated from claw impression IV by a sediment bar, can be consistently found slightly below the midlength of the footprint, and is here interpreted as the clawless digit impression V.
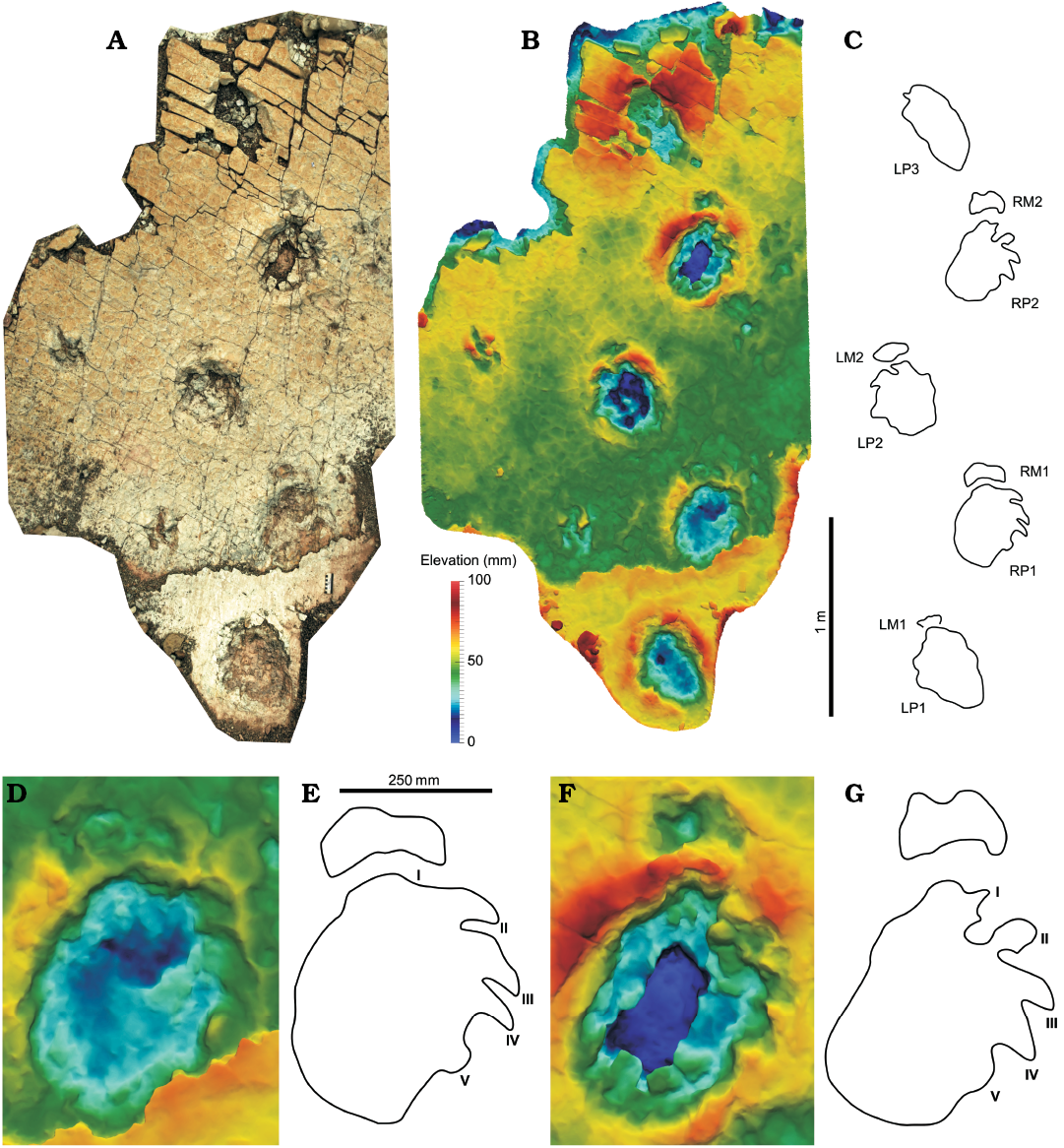
Fig. 2. A–C. Trackway S1 (Eosauropus sp.), here attributed to a sauropod trackmaker based on pedal synapomorphies; trackmaker is moving towards the south-west. Two consequtive pes impressions of a tridactyl Grallator trackway can be seen left to the S1 trackway. D, E. Detail of representative pes-manus set RP1/RM1. F, G. Detail of representative pes-manus set RP2/RM2. Photogrammetric orthophoto (A), depth-color images (B, D, F), interpretative drawings (C, E, G). Abbreviations: LM, left manus; LP, left pes; RM, right manus; RP, right pes.
Trackway S2.—It consists of four consecutive pes-manus pairs (Fig. 3, SOM 1, 3) leading towards the south-west, located ca. 290 m away from trackway S1. With a pes length of 41 cm, it is very similar in size to trackway S1. When compared with the latter, the pes impressions are much more elongated, and surrounded by broad displacement rims. The pes impressions are also more indistinct. While the medial margin of the pes prints is only slightly irregular, the lateral side shows irregular extensions which suggest the presence of digits, although the identification of specific digit impressions is not possible. All manus impressions are clearly visible, surrounded by displacement rims, and are either oval in shape or show a concave posterior margin. The trackway is somewhat narrower than trackway S1, with a WAP/PL ratio of 0.7. As in trackway S1, the pes pace angulation (137°) is higher than the manus pace angulation (128°). Pes prints are rotated outwards relative to the trackway midline by about 24° on average, only slightly less than in trackway S1, while the rotation angles of the manus prints are variable, being strongly rotated outwards in LM1 and slightly inwards in LM2.
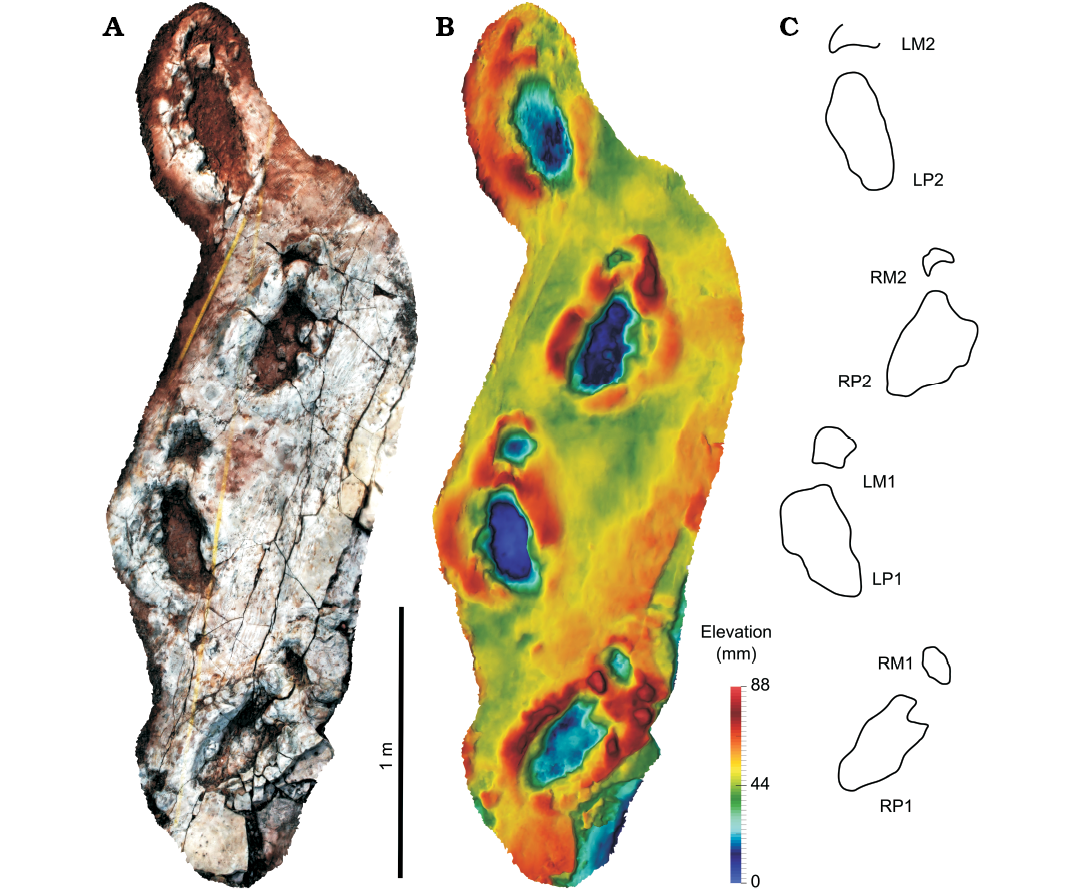
Fig. 3. Trackway S2 (Eosauropus sp.), which probably represents the same trackmaker species, or even the same individual, as trackway S1; trackmaker is moving towards the southwest. A. Photogrammetric orthophoto. B. Depth-color image. C. Interpretative drawing. Abbreviations: LM, left manus; LP, left pes; RM, right manus; RP, right pes.
Trackway S3.—It is the trackway of a biped consisting of four consecutive pes imprints (Fig. 4, SOM 1, 4), located ca. 10 m away from trackway S1. Only RP1 shows sufficient detail for inferences on the foot anatomy (Fig. 4B, C). This footprint is slightly longer (36 cm) than wide (33 cm) and shows impressions of digits I–IV, with digit impression III being the longest, followed by II and IV. Digit IV is incompletely preserved distally. Digit I is thin and strongly deformed, possibly due to mud collapse. Digit impressions I–III preserve claw traces that are curved medially. Digit impression I is somewhat separated from, and much more posterior to, digit impressions II–IV. The elongated heel impression is deep and narrow, and well defined laterally, being separated from digit IV by an embayment of the lateral track wall. The medial border of the heel impression is less well defined and separated from digit impression I by an extensive embayment. The posterior margin of the heel is distinctly rounded. The trackway shows a relatively wide gauge, with a pace angulation of 144°. Footprints are slightly rotated inwards by about 6° with respect to the direction of travel.

Fig. 4. A, B. Trackway S3 (Evazoum sp.). This bipedal trackway can possibly be attributed to a non-sauropod sauropodomorph trackmaker. Two smaller tridactyl trackways can be identified in close association with trackway S3. C, D. Detail of the best preserved pes impression RP1 (total depth represented by the color scale is 58 mm). Photogrammetric orthophoto (A), depth-color images (B, C), interpretative drawing (D).
Discussion
Number of trackmaker taxa and ichnotaxonomy.—Clemmensen et al. (2016) assigned trackways S1 and S2 to separate types, suggesting different sauropodomorph trackmakers, due to the much more elongated pes shape seen in trackway S2. However, trackway S2 also differs from S1 in the more indistinct shape and the broad displacement rims, indicating different substrate conditions at the time of track formation. Indeed, LP3 of trackway S1 appears to be elongated to a similar degree seen in trackway S2, indicating that the elongated morphotype results from suction effects narrowing the footprint during track formation. Furthermore, as both trackways lead to the south-west, closely match in pes length, and are preserved on the same bedding plane ca. 290 m apart from each other, it is well possible that trackway S1 represents the continuation of trackway S2. Consequently, trackways S1 and S2 are here regarded a single track type, possibly left by the same species or even the same individual.
Trackway S3 is clearly distinct, differing from trackways S1 and S2 in a number of features, including the lack of manus prints, the anteriorly to slightly inward rotated pes prints, the constricted heel-region, the relative lengths of the digit impressions, where digit impression III is the most protruding, the claw impressions, which are directed medially rather than laterally, and the lack of an impression of digit V.
Many tetrapod trackways from the Upper Triassic have been assigned to sauropodomorph, mainly “prosauropod”, trackmakers. Widely discussed examples include the ichnotaxa Tetrasauropus and Pseudotetrasauropus from the Lower Elliot Formation of southern Africa (Ellenberger 1972; Porchetti and Nicosia 2007). Subsequently identified trackways from Europe and North America have been attributed to the ichnotaxa Evazoum, Otozoum, and Eosauropus (Gand et al. 2000; Lockley and Meyer 2000; Nicosia and Loi 2003; Lockley et al. 2006a, b; Lucas et al. 2010; Lockley and Lucas 2013; Meyer et al. 2013). Of these track types, Tetrasauropus and Eosauropus reflect a quadrupedal locomotion. In Tetrasauropus, the pes is oriented parallel to the trackway midline, and claw traces are strongly curved inwards, whereas in trackways S1 and S2 from Greenland both the pes and the claw traces are directed outwards.
Trackways S1 and S2 comply best with the description of Eosauropus (Lockley et al. 2006a), and can be referred to this ichnogenus. However, the pes footprints of the present trackways are about twice the size of those of the Eosauropus type trackway and distinctly larger than any other trackways referred to this ichnogenus. The outwardly rotated digit impressions of the manus seen in Eosauropus (Lockley et al. 2006a) are not discernible in the present trackways, which could be due to preservation. Trackway S3 may be referred to the ichnogenus Evazoum (Nicosia and Loi 2003; Lockley et al. 2006b), and differs from the similar ichnogenera Kalosauropus and Otozoum in the more splayed pes and the greater trackway width (cf. Nicosia and Loi 2003). Strikingly, with a pes length of 36 cm, the present trackway is distinctly larger than other tracks referred to this ichnogenus, including the type trackway (10.5 cm) as well as the larger ichnospecies Evazoum gatewayensis, which was described to be up to 23 cm in pes length, “much larger than any other Evazoum morphotypes” (Lockley and Lucas 2013: 347). Porchetti et al. (2008) suggested that Evazoum is restricted to paleolatitudes between 0° and 30° N. The present trackway suggests the occurrence of this ichnogenus at 40° N, expanding the known range of this footprint type. Evazoum tracks are commonly referred to basal sauropodomorph trackmakers (Nicosia and Loi 2003; Lockley et al. 2006b), although a bipedal crurotarsan cannot be excluded (Porchetti et al. 2008). Bipedal poposauroids similar to Effigia (cf. Nesbitt 2007) and Poposaurus (Farlow et al. 2014) show developments in the locomotor apparatus that are partly convergent to those in Dinosauromorpha. In particular, Poposaurus gracilis had a mesaxonic pes that could have left tri-tetradactyl dinosauroid footprints (Farlow et al. 2014). However, these were probably of grallatorid morphology with a relatively long impression of middle digit III, whereas in Evazoum digit impression III is only slightly longer than digit impressions II and IV. The Fleming Fjord Formation has also produced body fossils of a basal sauropodomorph, which was previously referred to Plateosaurus by Jenkins et al. (1994) and might qualify as a possible trackmaker of the described trackway. Preliminary analysis suggests this form to be distinct from Plateosaurus, recommending a suprageneric classification within Plateosauria (Marco Marzola, personal communication 2017). Given the incomplete preservation of trackway S3, a synapomorphy-based identification of the trackmaker is not attempted here.
Evolution of the manus and pes skeleton in early sauropods, and the Triassic sauropod fossil record.—The anatomy of the sauropod manus and pes is highly distinctive. The manus consists of vertically oriented metacarpals arranged in a tight semicircle, while digits are strongly shortened or, as is the case in some titanosaurs, completely absent, resulting in horseshoe-shaped manus footprints. The much larger pes is rotated outwards. The metatarsals are spreading with a semi-plantigrade posture and a fleshy heel pad as support. Five proportionally short digits contact the ground, the first three of which showing long, deep and narrow unguals decreasing in length from digit I to III, resulting in an asymmetric, entaxonic pes footprint. Unguals are reduced or absent on digit IV and absent on digit V. When flexed, the claws are laterally or posterolaterally oriented (e.g., Wilson and Sereno 1998; Bonnan 2005).
The evolution of this peculiar autopodial anatomy in early sauropods remains poorly understood given the extremely sparse body fossil record of preserved pes and manus skeletons. Although the seminal works of Wilson and Sereno (1998) and Wilson (2002, 2005) established a number of autopodial synapomorphies of Sauropoda, Eusauropoda, and Neosauropoda, newly described taxa show that some of these features are not as straightforward as previously thought.
To date, no unambiguous sauropod tracks are known from the Triassic, although a possible sauropod origin was repeatedly proposed for trackways from the Chinle Group of western North America, which are Norian–Rhaetian in age (ca. 210 Mya; Lockley et al. 2001) and ascribed to the ichnogenus Eosauropus (previously Tetrasauropus) (Lockley et al. 2001, 2006a; Wilson 2005). Eosauropus tracks feature proportionally large pes and small manus footprints, suggesting a quadrupedal locomotion. Manus footprints typically show four to five short, clawless digits arranged in a gentle crescent. Pes footprints show four claw-bearing and a fifth non-claw bearing digit, and an extensive heel-region. Claw impressions are consistently anterolaterally directed (Wright 2005; Lockley et al. 2006a). In an attempt to identify the trackmaker based on synapomorphies, Wilson (2005) proposed a sauropod origin, based on the quadrupedal gait and digitigrade manus, and even linked several track features with synapomorphies of Eusauropoda. Thus, the short digits and the extensive heel region suggest a semi-digitigrade pes (sensu Wilson 2005), which is absent in non-eusauropod sauropods such as Vulcanodon (Cooper 1984). Furthermore, Wilson (2005) proposed that the tracks show laterally directed claw impressions, which he considered a synapomorphy of eusauropods more derived than Shunosaurus. The early appearance of these features might indicate an eusauropod origin in the Triassic, implying ghost linages. However, based on a stratocladistic approach, Wilson (2005) favored the idea that these eusauropod-like characters might had evolved independently in the Eosauropus trackmakers.
More recent studies questioned a sauropod origin of the Eosauropus tracks. Bonnan and Yates (2007) suggested that quadrupedal non-sauropod sauropodomorphs closely related to sauropods may also qualify as possible trackmakers, based on the morphological similarity between Eosauropus manus footprints and the manus skeleton of Melanorosaurus. Lockley et al. (2011) noted that in Eosauropus digit impression I is relatively short in comparison with a typical sauropod pes, and suggested that this feature might indicate a “prosauropod” trackmaker. Nesbitt et al. (2007) considered the Eosauropus tracks discussed by Wilson (2005) to be indeterminate, and claimed that the synapomorphies cited by the latter author are also found in the non-dinosaurian ichnogenus Brachychirotherium (see also Lucas et al. 2010). However, the footprint morphology in the present trackway is clearly distinct from Brachychirotherium, given the asymmetric, entaxonic pes, and the extremely laterally directed claw impressions, leaving sauropodomorph dinosaurs as the most probable trackmakers.
Synapomorphy-based trackmaker identification.—In the following, we discuss and evaluate previously proposed synapomorphies which might be identifiable in the present footprints.
(i) Obligate quadrupedal posture, as indicated by the consistent appearance of manus footprints. The obligatory quadrupedal posture was considered a sauropod synapomorphy (Wilson and Sereno 1998; Wilson 2005), but it probably evolved independently in taxa close to, but not commonly regarded as sauropods, such as Melanorosaurus. Facultative quadrupedality probably appeared early in sauropodomorph evolution, as evidenced by the track record (McPhee et al. 2015).
(ii) Semi-digitigrade pes (sensu Wilson 2005) with a spreading metatarsus, in which the shortened metatarsals are held nearly horizontally, as indicated by the extensive and broad heel pad impression (Wilson 2005). This feature is considered an eusauropod synapomorphy (Upchurch 1995; Wilson 2002, 2005; Allain and Aquesbi 2008), and is absent in basal sauropods such as Vulcanodon and Tazoudasaurus, which show the plesiomorphic elongated metatarsus. As suggested by Wilson (2005), this synapomorphy can be identified in trackways referred to Tetrasauropus (= Eosauropus) from the Late Triassic of North America, suggesting either an early appearance of eusauropods or convergent evolution in sauropods and the Eosauropus trackmaker.
(iii) Laterally deflected unguals, as a result of the asymmetric position of the flexor tubercle of the unguals (Bonnan 2005). This feature was considered a synapomorphy of a clade containing Barapasaurus and more derived taxa (Wilson and Sereno 1998; Wilson 2002, 2005), or Omeisaurus and more derived taxa (Upchurch et al. 2007; Yates et al. 2010). An incipient lateral deflection of pedal ungual I, where the convex dorsal surface of the latter faces slightly medially, can be already observed in Antetonitrus (McPhee et al. 2014). Wilson (2005) identified this feature in Tetrasauropus (=Eosauropus) trackways, which, however, typically show anterolaterally oriented claw impressions not reaching the degree of deflection seen in many sauropod trackways. It has to be cautioned that the orientation of the claw marks might be to some extent influenced by the foot kinematics during footprint formation, and therefore do not necessarily mirror the exact anatomical ungual orientations of the foot. Furthermore, claw orientation is controlled by the degree of plantar flexion of the pes, and thus trackmaker behavior—while laterally deflected unguals in a flexed pes are indicative of the peculiar eusauropod pes anatomy, more anteriorly directed claw marks only indicate digit extension and do not preclude an eusauropod trackmaker (cf. Bonnan 2005; Hall et al. 2016). Nevertheless, the present trackway provides evidence of claw impressions arranged in a right angle to the pes long axis, indicating a larger degree of ungual deflection than observed in other Eosauropus trackways, closely corresponding to the condition seen in a flexed sauropod pes. Claw impressions appear to show the medial side of the claws, not the ventral side as would be expected in anteriorly directed claws.
(iv) Pes entaxony, listed as a sauropod synapomorphy by Wright (2005), and outward rotation of pes. Wilson and Sereno (1998) considered the increased robustness of metatarsal I a sauropod synapomorphy, indicating a medial shift of the weight-bearing axis. In the present trackway, the relative length of digit impression I cannot be determined with certainty, but the digit appears robust at least in RP3. Digit II is more protruding than digit III, although it cannot be determined whether this reflects the relative lengths of the individual digits. Yates et al. (2010) noted the sauropod-like metatarsal in the bipedal non-sauropod sauropodomorph Aardonyx, and suggested that a robust digit I predated the acquisition of an obligate quadrupedal gait.
Another conspicuous feature of sauropod trackways, the outward rotation of the pes, was suggested to be associated with the increased entaxony, where the more robust inner digits carry the majority of the weight. Due to this outward rotation, digit I is the leading digit despite being shorter than digits II and III. Bonnan (2005: 360) suggested that “perhaps the lateral, outward turn of the pes occurred in the neosauropod forms, with more primitive eusauropods retaining a pes orientation closer to those of other saurischian dinosaurs”. The marked pes outward rotation in the present trackway (30°), which is arguably not produced by a neosauropod trackmaker given the broadly arched manus imprints, argues against this interpretation. Outwards rotated pes impressions are also commonly observed in other trackways that possibly pertain to basal sauropods (e.g., cf. Lockley et al. 2001, 2006a; Xing et al. 2016). This feature, therefore, might be characteristic for sauropods in general or for an even more inclusive grouping.
(v) Increased length of metatarsal V, with all five digits contacting the ground and playing an active role in bearing the animal’s weight; suggested as a sauropod synapomorphy (Wilson and Sereno 1998), or as the synapomorphy of a slightly more inclusive grouping (Gongxianosaurus plus more derived taxa; Yates 2007; Yates et al. 2010). The proportionally much longer metatarsal V suggests that in footprints digit impression V would be much more protruding than in basal sauropodomorphs, where it usually would not have made contact with the ground. The position of digit impression V in the present trackway matches the sauropod pes configuration.
Other synapomorphies which can be potentially identified in tracks cannot be evaluated for the present trackway. The reduction of manual phalanges is a eusauropod synapomorphy according to Wilson and Sereno (1998), although Wilson (2005) points out that digit reduction must have begun prior to the Lower Jurassic, as indicated by trackway evidence. Both trackways S1 and S2 lack indications for free manual digits, possibly due to poor preservation. Synapomorphies related to pedal ungual I cannot be evaluated in the present trackway since claw impression I is not readily preserved. The deep and narrow morphology of pedal ungual I is considered a sauropod synapomorphy, while the increased size of this ungual is considered a synapomorphy of both Sauropoda (pedal ungual I being 25% longer than pedal ungual II) and Eusauropoda (pedal ungual I longer than metatarsal I) (Wilson and Sereno 1998). A pedal ungual II which is 90% or less the size of the ungual of pedal digit I was resolved as a synapomorphy of a clade consisting of Gongxianosaurus plus more derived taxa (Yates 2007; Yates et al. 2010). A proposed eusauropod synapomorphy, the pedal digits II to III show sickle-shaped, laterally compressed unguals similar to that of digit I (Wilson and Sereno 1998; Wilson 2002; Upchurch et al. 2007). Although appearing sickle-shaped, a comparison of claw impressions II and III with that of digit I is not possible as the latter is not preserved in the present trackway; the trackmakers however for sure lacked the distinctly dorsoventrally flattened unguals seen in Vulcanodon.
Other features exhibited by the present trackway represent plesiomorphies which are lost in derived sauropods. Although poorly preserved, the elongated manus impressions indicate metacarpals arranged in a broad arc, as exemplified by the complete articulated manus of Tazoudasaurus (Allain and Aquesbi 2008). The tightly bound, horseshoe-like manus configuration seen in later sauropods probably occurred relatively late in sauropod evolution, and was suggested to be a synapomorphy of Neosauropoda (Wilson and Sereno 1998; Wilson 2005; Allain and Aquesbi 2008). Furthermore, claw impression IV is well developed in the present trackway, but the reduction or loss of pedal ungual IV is considered a synapomorphy of Eusauropoda (Wilson and Sereno 1998; Wilson 2002; Allain and Aquesbi 2008).
The present trackway therefore exhibits a mixture of primitive and derived features. The similarities with trackways attributed to derived sauropods are intriguing, in fact, trackway S1 can be differentiated from the latter only based on the absence of the horseshoe-like manus configuration characteristic for neosauropods and the presence of a fully developed ungual on pedal digit IV, which is reduced or absent in eusauropods. The quadrupedal posture, the pentadactyly of the pes and the extent of digit V relative to digit IV, as well as the entaxony of the pes suggest a derived sauropodiform trackmaker. Other synapomorphies, including the semi-digitigrady of the pes and the laterally deflected unguals, are even suggestive to a more exclusive grouping containing sauropods more derived than Tazoudasaurus and Vulcanodon.
Conclusions
The Norian–early Rhaetian Fleming Fjord Formation of central East Greenland adds significant information to the diversity and distribution of Late Triassic vertebrate tracks. The described bipedal trackway is referred to the ichnogenus Evazoum, and might represent a non-sauropod sauropodomorph trackmaker. The two quadrupedal trackways, possibly of sauropod origin, are here referred to Eosauropus. The described tracks represent the first evidence for the presence of these ichnotaxa in Greenland, expanding their known paleogeographical range. Both track types are distinctly larger than other known tracks ascribed to these ichnogenera.
The early evolution of sauropods is poorly known, and undisputed evidence for an appearance of this clade prior to the Jurassic is currently lacking. The quadrupedal trackways described herein suggest the adoption of a sauropod-like foot posture, where the pes is semi-digitigrade, oriented antero-laterally, and shows laterally deflected unguals, as early as the late Norian–early Rhaetian. Pedal synapomorphies (quadrupedal posture, entaxonic pes, five weight-bearing digits) suggest a derived sauropodiform trackmaker, with two synapomorphies (semi-digitigrade pes, laterally deflected unguals) suggesting a sauropod trackmaker more derived than Tazoudasaurus and Vulcanodon. These trackways, therefore, might provide further evidence for the appearance of sauropods as early as the Late Triassic, and represent the first evidence of this group from Greenland. It cannot be excluded, however, that the trackmaker taxon was a non-sauropod sauropodiform which convergently acquired a eusauropod-like pes that is both semi-digitigrade and shows laterally deflected unguals (cf. Wilson 2005).
Two of the aforementioned features of the present trackways (semi-digitigrade pes and laterally directed unguals) have been considered eusauropod synapomorphies, while the retention of a large ungual on digit IV represents a plesiomorphic feature thought to be absent in eusauropods. This previously unknown combination of characters might therefore indicate mosaic evolution in the pes skeleton of early sauropods.
Acknowledgements
We thank P. Martin Sander (Steinmann Institute, University of Bonn, Bonn, Germany) for constructive comments on an early version of the manuscript. Nadia Rosendal Nielsen (Geocenter Møns Klint, Borre, Denmark) is thanked for logistic support and contact to the media; Jan Schulz Adolfssen (Ministry of Mineral Resources, Nuuk, Greenland) and Nicole Klein (Steinmann Institute, University of Bonn, Bonn, Germany) are thanked for help in the field. Jakob Søndergaard, POLOG, was in charge of safety during the expedition, and a great help during the excavations. Finally, we thank Nils Natorp, Director of the Geocenter Møns Klint, for organizing the expedition. Blair McPhee (University of the Witwatersrand, Johannesburg, South Africa) and an anonymous reviewer provided constructive comments that helped to improve the manuscript substantially. Furthermore, we are grateful for the generous support to the 2012 expedition from the following foundations: Dronning Margrethes og Prins Henriks Fond, Arbejdsmarkedets Feriefond, Oticon Fonden, Knud Højgaards Fond, Louis Petersens Legat, Det Obelske Familiefond, and Ernst og Vibeke Husmans Fond. POLOG provided professional expedition logistics and camp solutions.
References
Allain, R. and Aquesbi, N. 2008. Anatomy and phylogenetic relationships of Tazoudasaurus naimi (Dinosauria, Sauropoda) from the late Early Jurassic of Morocco. Geodiversitas 30: 345–424.
Bonnan, M.F. 2005. Pes Aanatomy in sauropod dinosaurs: implications for functional morphology, evolution, and phylogeny. In: V. Tidwell and K. Carpenter (eds.), Thunder-Lizards: The Sauropodomorph Dinosaurs, 346–380. Indiana University Press, Bloomington.
Bonnan, M.F. and Yates, A.M. 2007. A new description of the forelimb of the basal sauropdomorph Melanoosaurus: implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs. Special Papers in Palaeontology 77: 157–168.
Buffetaut, E., Suteethorn, V., Cuny, G., Tong, H., Le Loeuff, J., Khansubha, S., and Jongautchariyakul, S. 2000. The earliest known sauropod dinosaur. Nature 407: 72–74. Crossref
Clemmensen, L.B. 1980. Triassic lithostratigraphy of East Greenland between Scoresby Sund and Kejser Franz Josephs Fjord. Bulletin Grønlands Geologiske Undersøgelse 139: 1–56.
Clemmensen, L.B., Kent, D.V., and Jenkins Jr., F.A. 1998. A Late Triassic lake system in East Greenland: facies, depositional cycles and palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 135–159. Crossref
Clemmensen, L.B., Milàn, J., Adolfssen, J.S., Estrup, E.J., Frobøse, N., Klein, N., Mateus, O., and Wings, O. 2016. The vertebrate-bearing Late Triassic Fleming Fjord Formation of central East Greenland revisited: stratigraphy, palaeoclimate and new palaeontological data. In: B.P. Kear, J. Lindgren, J.H. Hurum, J. Milàn, and V. Vajda (eds.), Mesozoic Biotas of Scandinavia and its Arctic Territories. Geological Society, London, Special Publications 434: 31–47. Crossref
Cooper, M.R. 1984. A reassessment of Vulcanodon karibaensis Raath (Dinosauria: Saurischia) and the origin of the Sauropoda. Palaeontologia Africana 25: 203–231.
Ellenberger, P. 1972. Contribution à la classification des Pistes de Vertébrés du Trias: les types du Stormberg d’Afrique du Sud (I). Palaeovertebrata, Memoire Extraordinaire: 1–104.
Farlow, J.O., Schachner, E.R., Sarrazin, J.C., Klein, H., and Currie, P.J. 2014. Pedal proportions of Poposaurus gracilis: convergence and divergence in the feet of archosaurs. Anatomical Record 297: 1022–1046. Crossref
Gand, G., Vianey-Liaud, M., Demathieu, G., and Garric, J. 2000. Deux nouvelles traces de pas de dinosaures du Trias supérieur de la bordure Cévenole (La Grand-Combe, Sud-Est de la France). Geobios 33: 599–624. Crossref
Gatesy, S.M. 2003. Direct and indirect track features: what sediment did a dinosaur touch? Ichnos 10: 91–98. Crossref
Hall, L.E., Fragomeni, A.E., and Fowler, D.W. 2016. The flexion of sauropod pedal unguals and testing the substrate grip hypothesis using the trackway fossil record. In: P.L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks: The Next Steps, 138–151. Indiana University Press, Bloomington.
Jenkins, F.J., Shubin, N.H., Amarel, W.W., Gatesy S.M., Schaff, C.R., Clemmensen, L.B., Downs, W.R., Davidson, A.R., Bonde, N.C., and Osbaeck, F. 1994. Late Triassic continental vertebrates and depositional environments of the Fleming Fjord Formation, Jameson Land, east Greenland. Meddelelser Om Grønland, Geoscience 32: 1–25.
Kent, D.V. and Clemmensen, L.B. 1996. Paleomagnetism and cycle stratigraphy of the Triassic Fleming Fjord and Gipsdalen formations of East Greenland. Bulletin of the Geological Society of Denmark 42: 121–136.
Klein, H., Milàn, J., Clemmensen, L.B., Frobøse, N., Mateus, O., Klein, N., Adolfssen, J.S., Estrup, E.J., and Wings, O. 2016. Archosaur footprints (cf. Brachychirotherium) with unusual morphology from the Upper Triassic Fleming Fjord Formation (Norian–Rhaetian) of East Greenland. In: B.P. Kear, J. Lindgren, J.H. Hurum, J. Milàn, and V. Vajda (eds.), Mesozoic Biotas of Scandinavia and its Arctic Territories. Geological Society, London, Special Publications 434: 71–85.
Leonardi, G. 1987. Discussion of the terms and methods. In: G. Leonardi (ed.), Glossary and Manual of Tetrapod Footprint Palaeoichnology, 43–51. Publicação do Departemento Nacional da Produção Mineral Brasil, Brasília.
Lockley, M.G. and Lucas, S.G. 2013. Evazoum gatewayensis, a new Late Triassic archosaurian ichnospecies from Colorado: implications for footprints in the ichnofamily Otozoidae. In: L.H Tanner, J.A. Spielmann, and S.G. Lucas (eds.), The Triassic System. New Mexico Museum of Natural History and Science Bulletin 61: 345–353.
Lockley, M.G. and Meyer, C. 2000. Dinosaur Tracks and Other Fossil Footprints of Europe. 323 pp. Columbia University Press, New York.
Lockley, M.G., Lucas, S.G., and Hunt, A. P. 2006a. Eosauropus, a new name for a Late Triassic track: Further observations on the Late Triassic ichnogenus Tetrasauropus and related forms, with notes on the limits of interpretation. In: J.D. Harris, S.G. Lucas, J.A. Spielmann, M.G. Lockley, A.R.C. Milner, and J.I. Kirkland (eds.), The Triassic–Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37: 192–198.
Lockley, M.G., Lucas, S.G. and Hunt, A.P. 2006b. Evazoum and the renaming of northern hemisphere “Pseudotetrasauropus”: implications for tetrapod ichnotaxonomy at the Triassic–Jurassic boundary. In: J.D. Harris, S.G. Lucas, J.A. Spielmann, M.G. Lockley, A.R.C. Milner, and J.I. Kirkland (eds.), The Triassic–Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37: 199–206.
Lockley, M.G., Wright, J.L., Hunt, A.P., and Lucas, S.G. 2001. The Late Triassic sauropod track record comes into focus: old legacies and new paradigms. New Mexico Geological Society Guidebook 52: 181–190.
Lockley, M.G., Hups, K., Cart, K., and Gerwe, S. 2011. A zone of sauropodomorph footprints in the basal Wingate Sandstone (latest Triassic) of western Colorado and eastern Utah: Is Eosauropus a common ichnogenus in this region? New Mexico Museum of Natural History and Science Bulletin 53: 337–343.
Lucas, S.G., Spielmann, J.A., Klein, H., and Lerner, A.J. 2010. Ichnology of the Upper Triassic Redonda Formation (Apachean) in east-central New Mexico. New Mexico Museum of Natural History and Science Bulletin 47: 1–75.
Mallison, H. and Wings, O. 2014. Photogrammetry in paleontology—a practical guide. Journal of Paleontological Techniques 12: 1–31.
Marty, D. 2008. Sedimentology, taphonomy, and ichnology of Late Jurassic dinosaur tracks from the Jura carbonate platform (Chevenez-Combe Ronde tracksite, NW Switzerland): Insights into the tidal-flat palaeoenvironment and dinosaur diversity, locomotion, and palaeoecology. GeoFocus 21: 1–278.
McPhee, B.W., Bonnan, M.F., Yates, A.M., Neveling, J., and Choiniere, J.N. 2015. A new basal sauropod from the pre-Toarcian Jurassic of South Africa: Evidence of niche-partitioning at the sauropodomorph–sauropod boundary? Scientific Reports 5: 13224. Crossref
McPhee, B.W., Yates, A.M., Choiniere, J.N., and Abdala, F. 2014. The complete anatomy and phylogenetic relationships of Antetonitrus ingenipes (Sauropodiformes, Dinosauria): implications for the origins of Sauropoda. Zoological Journal of the Linnean Society 171: 151–205. Crossref
Meyer, C.A., Marty, D., Thüring, B., Stecher, R., and Thüring, S. 2013. Dinosaurierspuren aus der Trias der Bergüner Stöcke (Parc Ela, Kanton Graubünden, SE Schweiz). Mitteilungen der Naturforschenden Gesellschaften beider Basel 14: 135–144.
Milàn, J. and Bromley, R.G. 2006. True tracks, undertracks and eroded tracks, experimental work with tetrapod tracks in laboratory and field. Palaeogeography, Palaeoclimatology, Palaeoecology 231: 253–264. Crossref
Nesbitt, S.J. 2007. The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History 302: 1–84. Crossref
Nesbitt, S.J., Irmis, R.B., and Parker, W.G. 2007. A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology 5: 209–243. Crossref
Nicosia, U. and Loi, M. 2003. Triassic footprints from Lerici (La Spezia, northern Italy). Ichnos 10: 127–140. Crossref
Porchetti, S.D. and Nicosia, U. 2007. Re-examination of some large early Mesozoic tetrapod footprints from the African collection of Paul Ellenberger. Ichnos 14: 219–245. Crossref
Porchetti, S.D., Nicosia, U., Mietto, P., Petti, F.M., and Avanzini, M. 2008. Atreipus-like footprints and their co-occurrence with Evazoum from the upper Carnian (Tuvalian) of Trentino-Alto Adige. Studi Trentini Di Scienze Naturali: Acta Geologica 83: 277–287.
Racey, A. and Goodall, J.G. 2009. Palynology and stratigraphy of the Mesozoic Khorat Group red bed sequences from Thailand. Geological Society, London, Special Publications 315: 69–83. Crossref
Sulej, T., Wolniewicz, A., Bonde, N., Błażejowski, B., Niedźwiedzki, G., and Tałanda, M. 2014. New perspectives on the Late Triassic vertebrates of East Greenland: preliminary results of a Polish-Danish palaeontological expedition. Polish Polar Research 35: 541–552. Crossref
Upchurch, P. 1995. The evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London B: Biological Sciences 349: 365–390. Crossref
Upchurch, P., Barrett, P.M., and Galton, P.M. 2007. A phylogenetic analysis of basal sauropodomorph relationships: implications for the origin of sauropod dinosaurs. Special Papers in Palaeontology 77: 57.
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 215–275. Crossref
Wilson, J.A. 2005. Integrating ichnofossil and body fossil records to estimate locomotor posture and spatiotemporal distribution of early sauropod dinosaurs: a stratocladistic approach. Paleobiology 31: 400–423. Crossref
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology 18 (Supplement to Number 2): 1–79.
Wright, J.L. 2005. Steps in understanding sauropod biology. In: K.C. Rogers and J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, 252–285. University of California Press, Berkeley.
Xing, L.D., Lockley, M.G., Zhang, J., Klein, H., Li, D., Miyashita, T., Li, Z., and Kümmell, S.B. 2016. A new sauropodomorph ichnogenus from the Lower Jurassic of Sichuan, China fills a gap in the track record. Historical Biology 28: 881–895. Crossref
Yates, A.M. 2007. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria). Special Papers in Palaeontology 77: 9–55.
Yates, A.M., Bonnan, M.F., Neveling, J., Chinsamy, A., and Blackbeard. M.G. 2010. A new transitional sauropodomorph dinosaur from the Early Jurassic of South Africa and the evolution of sauropod feeding and quadrupedalism. Proceedings of the Royal Society of London B: Biological Sciences 277: 787–794. Crossref
Acta Palaeontol. Pol. 62 (4): 833–843, 2017
https://doi.org/10.4202/app.00374.2017