
Lucinid bivalves from Miocene hydrocarbon seep sites of eastern North Island, New Zealand, with comments on Miocene New Zealand seep faunas
KAZUTAKA AMANO, CRISPIN T.S. LITTLE, and KATHLEEN A. CAMPBELL
Amano, A., Little, C.T.S., and Campbell, K.A. 2018. Lucinid bivalves from Miocene hydrocarbon seep sites of eastern North Island, New Zealand, with comments on Miocene New Zealand seep faunas. Acta Palaeontologica Polonica 63 (2): 371–382.
Three new fossil species of lucinids, Meganodontia haunuiensis, Elliptiolucina neozelandica, and Lucinoma saetheri, are described from lower to middle Miocene hydrocarbon seep carbonates from north and south of Hawke Bay, eastern North Island, New Zealand. Of these taxa Meganodontia haunuiensis is confined to seep sites south of Hawke Bay, while Elliptiolucina neozelandica comes only from the seep sites north of Hawke Bay. Using ecological information from modern bivalve species, we suggest the southern area seep sites formed in shallower waters than the northern sites. Among the lucinids, Meganodontia haunuiensis is one of the oldest records of the genus, and its distribution has shrunk from worldwide in the Miocene times to only around Taiwan today. Elliptiolucina neozelandica is the second oldest species in the genus. Since the Miocene the distribution of Elliptiolucina has narrowed but its habitat range has increased to both seeps and sandy environments, mainly around the Philippines.
Key words: Mollusca, Bivalvia, Lucinidae, hydrocarbon seep, Miocene, New Zealand.
Kazutaka Amano [amano@juen.ac.jp], Department of Geoscience, Joetsu University of Education, 1 Yamayashiki, Joetsu 943-8512, Japan.
Crispin T.S. Little [earctsl@leeds.ac.uk], School of Earth and Environment, University of Leeds, Leeds LS2 9JT, UK.
Kathleen A. Campbell [ka.campbell@auckland.ac.nz], School of Environment, Faculty of Science, University of Auckland, Private Bag 92019, Auckland Mail Centre, Auckland 1142, New Zealand.
Received 22 January 2018, accepted 9 March 2018, available online 28 March 2018.
Copyright © 2018 K. Amano et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The bivalve family Lucinidae is a species-rich taxon in modern and ancient hydrocarbon seep communities (e.g., Taylor and Glover 2006, 2010; Kiel 2010, 2013). Molecular work indicates that the lucinids can be grouped into eight subfamilies: Pegophyseminae Taylor and Glover, 2011, Leucosphaerinae Taylor and Glover, 2011, Myrteinae Chavan, 1969; Monitilorinae Taylor and Glover, 2011, Fimbriinae Nicol, 1950, Codakiinae Iredale, 1937, Lucininae Fleming, 1828, and ?Milthinae Chavan, 1969 (Taylor et al. 2011). Of these subfamilies, many fossil species from seep environments belong to the Myrteinae and Codakiinae (Kiel 2013).
Until now, only a few modern and fossil lucinid species have been recorded from hydrocarbon seep sites from North Island, New Zealand. Campbell et al. (2010) illustrated Lucinoma galathea (Marwick, 1953) from modern seeps off the Hikurangi Margin. Campbell et al. (2008) listed Lucinoma aff. taylori from the Miocene Rocky Knob seep site in the Gisborne region (Fig. 1). Saether et al. (2016) figured Meganodontia? sp. from several seep sites in the Hawke’s Bay region east of Dannevirke (Fig. 1). Kiel et al. (2013) illustrated unidentified species belonging to the genera Ezolucina? and Nipponothracia? from Cenomanian to Campanian (Cretaceous) North Island seep deposits. Specimens of Nipponothracia? from the Cenomanian Port Awanui seep site on the Raukumara Peninsula were subsequently described as Amanocina raukumara by Kiel (2013).
The Miocene seep deposits of North Island occur in two main areas north and south of Hawke Bay, respectively, north of Gisborne and east of Dannevirke (Fig. 1). The majority of the molluscan taxa from these seep sites have been described by Saether et al. (2010a, b, 2012, 2016) and Amano et al. (2014, 2015), except for the lucinids. In this paper, we describe and illustrate the Miocene seep lucinids from North Island, and briefly discuss the wider paleoenvironmental and regional paleobiogeographical context of the Miocene New Zealand seep communities.
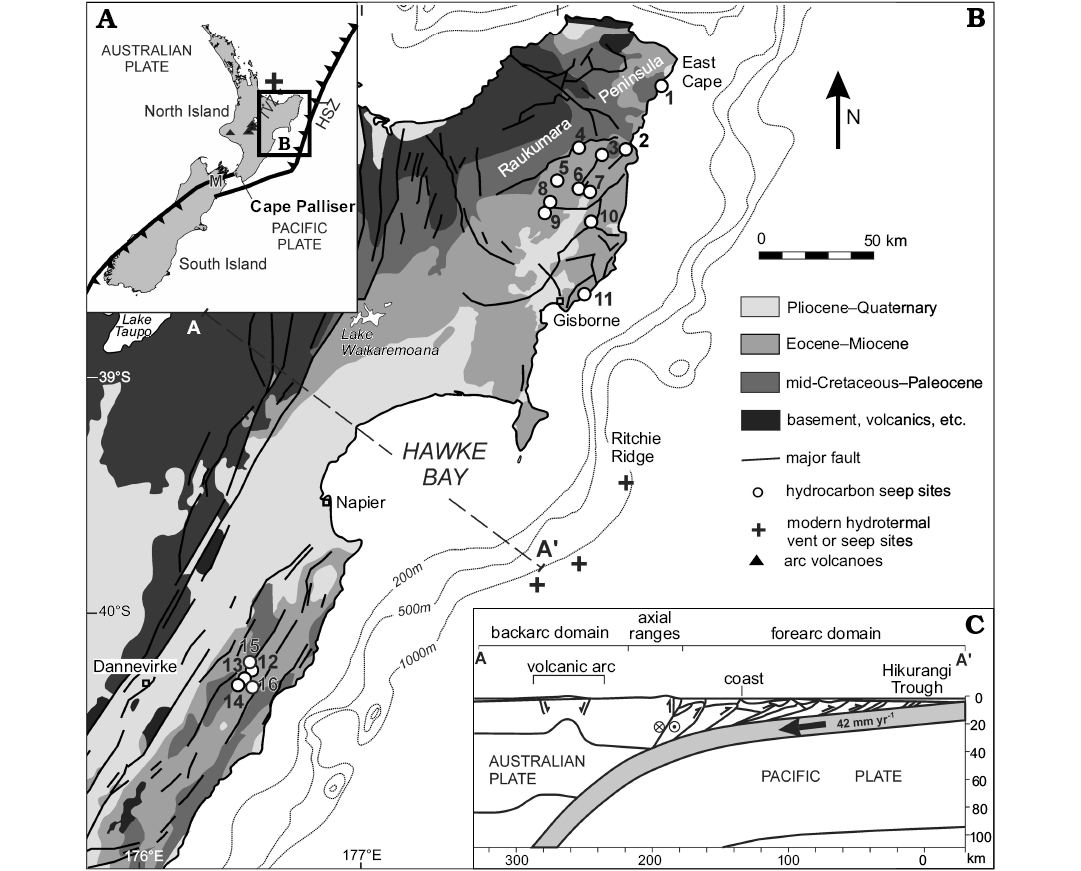
Fig. 1. Overview of the geology of the East Coast, North Island, New Zealand (A, B), showing locations of known Miocene hydrocarbon seep sites. Northern sites: 1, Waiapu; 2, Waipiro; 3, Karikarihuata; 4, Bexhaven; 5, Tauwhareparae; 6, Puketawa; 7, Totaranui; 8, Moonlight North; 9, Rocky Knob; 10, Waikairo; 11, Turihaua. Southern sites: 12, Wanstead; 13, Ugly Hill; 14, Haunui; 15, Ngawaka; 16, Wilder; HSZ, Hikurangi subduction zone; M, Marlborough; TVZ, Taupo Volcanic Zone. Also shown are some of the modern offshore hydrocarbon seep sites of the Hikurangi subduction margin, and a hydrothermal vent location (inset). C. Cross-section (modified from Barnes et al. 2010) shows transpressive subduction of the Pacific Plate beneath the Australian Plate, and relationships of tectonic elements of the northern New Zealand plate margin (see Campbell et al. 2008 and Barnes et al. 2010 for further details on geologic context). The fossil lucinids of this study are described from fossil seep locations 4, 8, 9, 12–15.
Institutional abbreviations.—AU, Paleontology Collection, School of Environment, UOA; NSMT, National Science Museum of Tokyo; UOA L, bivalve specimen number, Paleontology Collection, School of Environment, UOA; UOA, University of Auckland, Auckland, New Zealand.
Other abbreviations.—AL, anterior length; H, height; L, length; W, width; Y and U, map codes in New Zealand.
Material
In the area east of Dannevirke, lucinids were collected from the Ugly Hill and Haunui seep sites of the lower Miocene Ihungia Limestone. In the area north of Gisborne, lucinid specimens were collected from the Bexhaven, Moonlight North and Rocky Knob seep sites of the middle Miocene Bexhaven Limestone.
All but four of the fossil specimens described and figured herein were collected in February 2012 by the authors, and these are deposited in the paleontology collections of the School of Environment, the University of Auckland (UOA). Additional fossils collected by UOA paleontologists between 1997–2010, also housed in the UOA collections, were examined. All the localities of the illustrated specimens are registered in the New Zealand Fossil Record File database (http://www.fred.org.nz), jointly administered by GNS Science and the New Zealand Geoscience Society. For comparative purposes, modern seep lucinids were studied, comprising specimens dredged during the 2007 joint German-New Zealand NEW VENTS Hikurangi margin cruise with the R/V SONNE (stored in the UOA collections), and some Lucinoma specimens at the Museum of New Zealand, Te Papa Tongarewa (Wellington). Fig. 2 shows terminology for description.
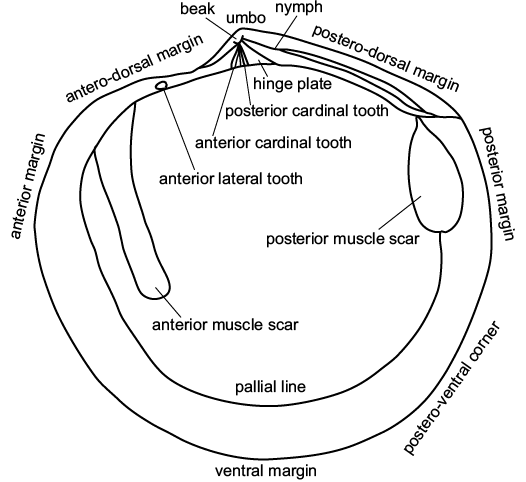
Fig. 2. Schematic drawing of a model lucinid bivalve with explanation of the main morphological terms used in this paper.
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Heterodonta Neumayr, 1884
Family Lucinidae Fleming, 1828
Subfamily Pegophyseminae Taylor and Glover
in Taylor et al., 2011
Remarks.—Huber (2015) cited the authors of this subfamily name as Taylor, Glover, Smith, Dyal, and Williams, 2011. However, in table 3 of Taylor et al. (2011), the authors were given as Taylor and Glover.
Genus Meganodontia Bouchet and Cosel, 2004
Type species: Meganodontia acetabulum Bouchet and Cosel, 2004; Recent, Tashi fishing ground, off Tashi, NE coast of Taiwan.
Remarks.—Meganodontia acetabulum Bouchet and Cosel, 2004 has a very large shell (up to 150 mm in length). Bouchet and Cosel (2004) listed the generic characters: edentulous hinge, thick-shelled, large anterior adductor muscle scar broadening ventrally, less prominent umbo and maximum height located behind the umbo. However, based on our observations of published photographs of the Italian fossil species Meganodontia hoernea (Des Moulins, 1868), this latter character seems to be absent, so it may be a species rather than generic-level character.
Within the subfamily Pegophyseminae, Pegophysema Stewart, 1930 differs from Meganodontia by having a smaller and thinner anterior adductor muscle scar, a more swollen umbo, a straighter hinge and some secondary pallial attachment scars (Bouchet and Cosel 2004; Taylor and Glover 2005). In addition, most Recent species of Pegophysema come from very shallow water sites, whereas fossil and Recent Meganodontia species come from deep-sea vent and seep sites (Bouchet and Cosel 2004; Taylor and Glover 2005, 2016; Kiel and Hansen 2015; Kiel and Taviani 2017).
Meganodontia haunuiensis sp. nov.
Fig. 3.
2016 ?Meganodontia sp.; Saether et al. 2016: 16, table 1.
Etymology: Named for the type locality (Haunui) in North Island, New Zealand.
Type material: Holotype, UOA L4781 (Fig. 3B) from locality U23/f270 (Haunui), collection AU19880. Paratypes: UOA L4777 (Fig. 3A) from locality U23/f280 (Ugly Hill), collection AU 21299; UOA L4782 (Fig. 3E), UOA L4791 (Fig. 3C), UOA L4792 (Fig. 3F) from locality U23/f270, collection AU 21298.
Type locality: Haunui seep site, North Island, New Zealand.
Type horizon: Ihungia Limestone, lower Miocene.
Dimensions.—See Table 1.
Table 1. Measurements (in mm) of Meganodontia
haunuiensis sp. nov.
Abbreviations: AL, anterior length; H, height; L, length; W, width.
|
Specimen |
Type |
L |
H |
AL |
W |
H/L |
AL/L |
W/L |
Valve |
|
UOA L4781 |
holotype |
84.6 |
73.8 |
22.1 |
– |
0.87 |
0.26 |
– |
right |
|
UOA L4777 |
paratype |
62.9+ |
63.3 |
– |
31.4 |
– |
– |
– |
left |
|
UOA L4782 |
paratype |
78.6 |
73.9 |
29.5 |
42.6 |
0.94 |
0.38 |
0.54 |
both |
|
UOA L4791 |
paratype |
78.2+ |
– |
– |
– |
– |
– |
– |
right |
|
UOA L4792 |
paratype |
74.8 |
67.9 |
22.4 |
51.1 |
0.91 |
0.30 |
0.68 |
both |
|
UOA L4778 |
|
47.5 |
47.7 |
18.1 |
20.4 |
1.00 |
0.38 |
0.43 |
both |
|
UOA L4787 |
|
67.4 |
62.0 |
29.7 |
30.0 |
0.92 |
0.44 |
0.44 |
both |
|
UOA L4788 |
|
55.0 |
49.6 |
24.0 |
– |
0.90 |
0.44 |
– |
left |
|
UOA L4789 |
|
40.2+ |
– |
– |
– |
– |
– |
– |
right |
|
UOA L4790 |
|
59.4 |
52.2 |
22.5 |
32.5 |
0.88 |
0.38 |
0.55 |
both |
Diagnosis.—Moderate-size, subcircular and strongly inflated Meganodontia with narrow lunule demarcated by sharp ridge in both valves and deeply sunken posterior lanceolate dorsal gape. Hinge rather broad and edentulous.
Description.—Shell moderately thick, moderate size up to 84.6 mm in length, subcircular but slightly longer than high (H/L = 0.87–1.00), strongly inflated (W/L = 0.43–0.68), equivalve, inequilateral. Postero-dorsal margin straight making obtuse angle with subtruncated posterior margin; ventral margin well rounded, graduating subcircular posterior margin into nearly straight dorsal margin. Umbo rather prominent but not swollen, situated at anterior one-fourth to two-fifths of shell length (AL/L = 0.26–0.44). Maximum height slightly behind umbo in holotype but located at umbo in paratype specimens. Surface sculptured by fine commarginal growth lines and some irregular commarginal undulations. Very weak groove running from umbo to postero-ventral corner. Lunule narrow, demarcated by sharp ridge in both valves and occupying postero-dorsal margin observed in right valve; posterior lanceolate dorsal gape deeply sunken and probably once occupied by ligament. Scar trace of pallial blood vessel observed in one specimen. Hinge rather broad for genus and edentulous, except for nymph separating ligament and subumbonal flat area. Anterior adductor scar rather large, elongate and stout; posterior adductor scar indistinct and subquadrate. Pallial line entire with no secondary pallial attachment scars. Inner surface of shell covered by fine radial grooves.
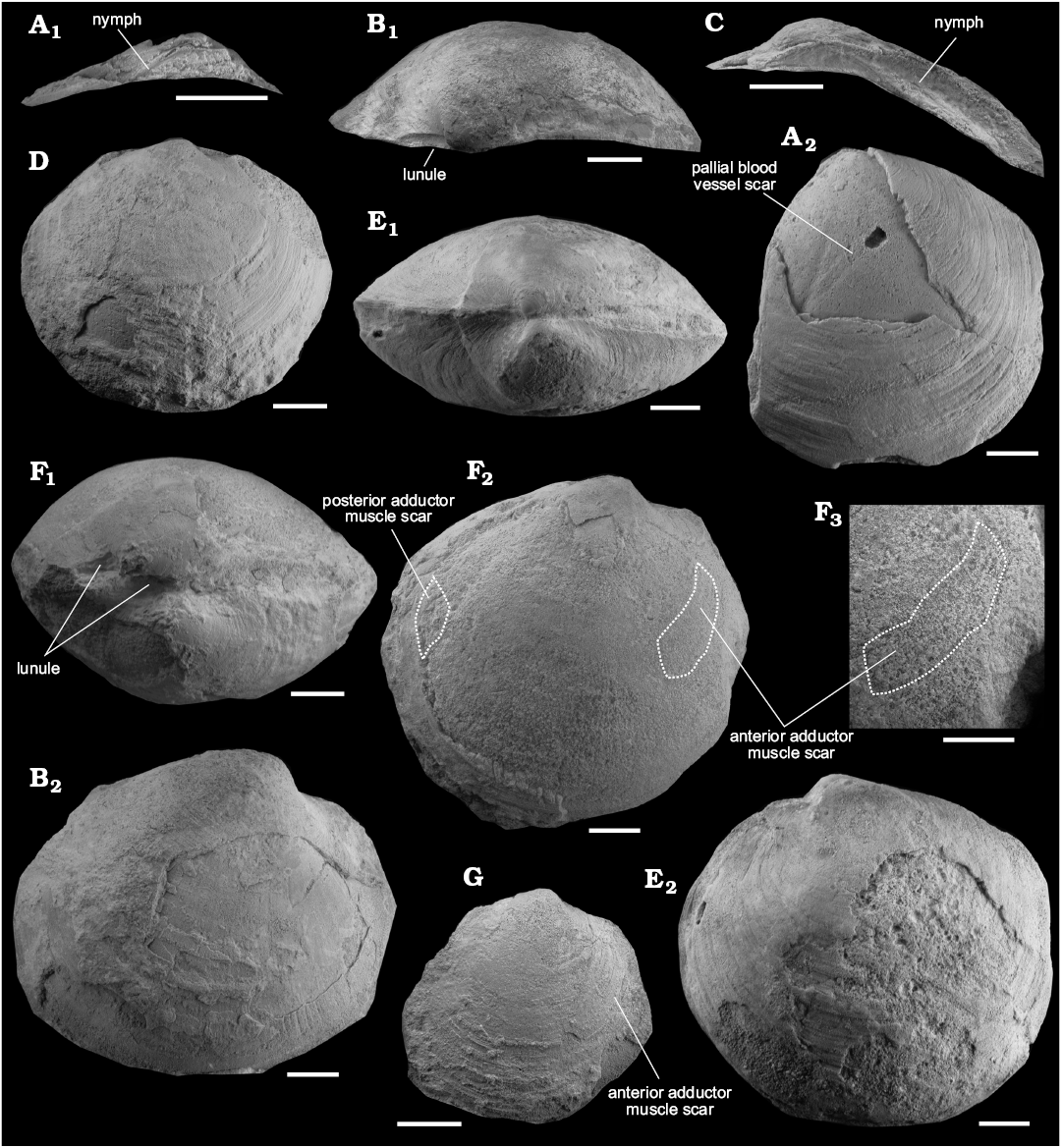
Fig. 3. Lucinid bivalve Meganodontia haunuiensis sp. nov. from the lower Miocene Ihungia Limestone at Haunui (B–G, U23/270) and Ugly Hill (A, U23/280), North Island, New Zealand. A. UOA L4777 (paratype), edentulous hinge plate of left valve (A1,), left valve (A2). B. UOA L4581 (holotype), dorsal view (B1) and frontal view (B2) of right valve. C. UOA L4791 (paratype), edentulous hinge plate of right valve. D. UOA L4790, right valve of small specimen. E. UOA L4782 (paratype), dorsal view of slightly compressed specimen (E1), left valve (E2). F. UOA L4792 (paratype), dorsal view showing symmetric lunule in both valves (F1), right valve showing internal mould (F2), enlargement of the anterior adductor muscle scar of F2 (F3). G. UOA L 4789, right valve, showing internal mould of small specimen. Scale bars 10 mm.
Remarks.—Despite having comparatively smaller shell size compared to other species in the genus, we placed these specimens into Meganodontia Bouchet and Cosel, 2004 because they have a large anterior adductor scar, a less swollen umbo than Pegophysema, a curved hinge area and no secondary pallial attachment scar. However, the broad hinge plate with a distinct posterior ridge has not been observed in other species of Maganodontia. Meganodontia hoernea (Des Moulins, 1868) from the middle to upper Miocene seep deposits of Italy is the most similar species to M. haunuiensis sp. nov. in that it also has a large shell and similar inflation (W/L = 0.31–0.71), but differs from the New Zealand species by being even larger (up to 180 mm in length) and possessing a narrower hinge plate.
Meganodontia sp. was illustrated from the Cuban early Miocene Cantera Portugalete seep site in Kiel and Hansen (2015). These specimens have similar outlines (especially their fig. 7C) to M. haunuiensis sp. nov. However, the Cuban specimens are larger (up to 120 mm in length) than M. haunuiensis sp. nov. A lack of information on the hinge area of the Cuban specimens prevents us from comparing these features.
Meganodontia haunuiensis sp. nov. can be easily distinguished from the type species of the genus, M. acetabulum, by its smaller shell (up to 84.6 mm in M. haunuiensis; up to 150 mm in M. acetabulum), more prominent umbo and much broader hinge plate.
“Anodontia (Anodontia)” waharoaensis Eagle, 1992 from Motutara, North Island, New Zealand, does not belong to Anodontia owing to its large size (L = 123 mm), but rather to the Pegophyseminae (genus uncertain). This species differs from M. haunuiensis sp. nov. by having a larger (L = 123 mm), thinner shell that is more elongated (H/L = 0.82; H/L = 0.87–1.00 in M. haunuiensis), with a narrowly rounded anterior margin.
Meganodontia haunuiensis sp. nov. resembles the Recent Indo-Pacific species, Pegophysema philippiana (Reeve, 1850) in its shell outline (H/L = 0.85–0.94 in P. phillipiana from the Kawamura Collection and NSMT-Mo 84906 stored at the National Science Museum, Tsukuba, Japan), size (max L = 71.0; Taylor and Glover 2005) and inflation (W/L = 0.52–0.74 from the Kawamura Collection and NSMT-Mo 84906 stored at the National Science Museum, Tsukuba). However, the broader hinge and larger, but narrower lunule of M. haunuiensis sp. nov. are very different from the same shell characters in P. philippiana.
Meretrix sphericula Basedow, 1902 (= Anodontia sphericula by Darragh, 1970) was described from the Miocene crystalline limestone around Edithburg, Australia. Ludbrook (1978) also described a well-preserved specimen from the Pliocene Roe Formation, Australia. This species is large in size (up to 105 mm) and has a strongly inflated shell (W/L = 0.75). However, the Australian species differs from M. haunuiensis sp. nov. in possessing a thinner shell with narrow hinge plate, a more horizontal postero-dorsal margin and a more protruding umbo.
Loripes goliath Yokoyama, 1928, from the Miocene of Taiwan has a very large shell (up to 150 mm in length) and was questionably allocated to Anodontia by Taylor and Glover (2009). This species differs from M. haunuiensis sp. nov. by having a more inflated shell and steeply sloping and straight postero-dorsal margin. Nonetheless, poor preservation and a lack of information about the internal structure of L. goliath prevent us from comparing the species further.
Stratigraphic and geographic range.—Haunui, Ugly Hill, Ngakawa, and Wanstead; Lower Miocene Ihungia Limestone, North Island, New Zealand.
Subfamily Myrteinae Chavan, 1969
Genus Elliptiolucina Cosel and Bouchet, 2008
Type species: Elliptiolucina magnifica Cosel and Bouchet, 2008; Recent, central Philippines.
Remarks.—This genus is characterized by having a subquadrate-ovate and weakly inflated shell, an edentulous hinge except for small cardinal and anterior lateral teeth in E. williamsae Glover and Taylor, 2016, and a short anterior adductor scar. Other edentulous lucinids with a weakly inflated, elongate shell include Nipponothracia Kanie and Sakai, 1997, Elongatolucina Gill and Little, 2013, and Amanocina Kiel, 2013. According to Okutani (2011), Elliptiolucina can be separated from Nipponothracia by its smaller and thicker shell and possession of a characteristic dorsal concavity in the posterior adductor muscle scar. Elongatolucina has an elongate shell with straight ventral margin, which is not seen in Elliptiolucina (see Kiel 2013). Amanocina can be easily distinguished from the other genera as it lacks a subumbonal triangular excavation and has a longer adductor muscle scar detached at a narrower angle from the pallial line (Kiel 2013).
Elliptiolucina neozelandica sp. nov.
Fig. 4.
Etymology: Named for New Zealand, the country of the type locality.
Type material: Holotype, UOA L4771(Fig. 4A) from locality Y16/f1174 (Moonlight North), collection AU19923. Paratypes: UOA L4783 (Fig. 4B); UOA L4775 (Fig. 4C); UOA L4774 (Fig. 4D); from locality Y16/f1059 (Moonlight North), collection AU 19982. One silicone rubber cast of the internal shell features made from UOA L4775.
Type locality: Moonlight North seep site, North Island, New Zealand.
Type horizon: Bexhaven Limestone, middle Miocene.
Dimensions.—See Table 2.
Table 2. Measurements (in mm) of Elliptiolucina
sp.
Abbreviations: AL, anterior length; H, height; L, length; W, width.
|
Specimen |
Type |
L |
H |
AL |
W |
H/L |
AL/L |
Valve |
|
UOA L4771 |
holotype |
31.1+ |
24.6 |
14.4 |
12.1 |
– |
– |
both |
|
UOA L4774 |
paratype |
28.6+ |
24.1 |
12.0 |
11.0 |
– |
– |
both |
|
UOA L4775 |
paratype |
34.9+ |
27.1 |
– |
11.1+ |
– |
– |
both |
|
UOA L4783 |
paratype |
17.1 |
12.8 |
5.0 |
6.3+ |
0.75 |
0.29 |
both |
Diagnosis.—Small and compressed Elliptiolucina having a thin shell without a sulcus. Antero-dorsal margin gently sloping to subtruncated anterior margin. Lunule narrow and escutcheon deeply sunken, demarcated by sharp ridge. Hinge of right valve shallowly sunken and edentulous with nymph.
Description.—Shell size rather small for genus, up 34.9 mm in length; rather thin-shelled, elongate ovate (H/L = 0.75), compressed, equivalve and inequilateral. Antero-dorsal margin gently sloping, making right angle with subtruncated anterior margin; ventral margin broadly arched and subparallel with dorsal margin; postero-dorsal margin nearly straight, leading into subtruncated posterior margin. Distinct but shallow sulcus present from beak to antero-ventral corner; shallow depression from beak running in postero-ventral direction. Surface largely smooth, sculptured by growth lines and about four very low and dense commarginal threads between growth lines. These growth lines turning into lamellated ribs near anterior and posterior dorsal margins. Umbo not prominent, prosogyrate and located at anterior one-third of shell length in one small entire specimen. Lunule narrow, lanceolate, shallowly sunken. Escutcheon lanceolate, deeply sunken and demarcated by sharp ridge. Hinge of right valve shallowly excavated and edentulous with nymph. Pallial line entire. Anterior adductor muscle scar elongate, long and slightly detached from pallial line; posterior adductor muscle scar large, subquadrate and rather deeply depressed on dorsal side.

Fig. 4. Lucinid bivalve Elliptiolucina neozelandica sp. nov. from the middle Miocene Bexhaven Limestone at Moonlight North (A, Y16/f1174; B–D, Y16/1059), North Island, New Zealand. A. UOA L4771 (holotype), dorsal view (A1), right (A2) and left (A3) valves. B. UOA L4783 (paratype), dorsal view (B1), right valve (B2), internal mould of left valve (B3), enlargement of anterior adductor muscle scar in B2 (B4). C. UOA L4775 (paratype), edentulous hinge plate of right valve (C1), left valve (C2). D. UOA L4774 (paratype), dorsal view (D1), right (D2) and left (D3) valves. Scale bars 10 mm.
Remarks.—The outlines, hinge characters, and shape of the posterior adductor scar show that the specimens belong to the genus Elliptiolucina, except for having rather thin shells. This is the first record of fossil Elliptiolucina in New Zealand.
Elliptiolucina neozelandica sp. nov. is most similar to the Recent Philippines Sea species, E. labeyriei Cosel and Bouchet, 2008 in having a relatively small, rather thin shell, a narrow lunule and a similar outline. However, E. labeyriei has a distinct and wide sulcus from the beak to the postero-ventral corner, which is not seen in E. neozelandica sp. nov.
Elliptiolucina williamsae Glover and Taylor, 2016 from the central Philippines lives in rather shallow water (280–679 m depth) compared to other species in the genus. This species also has a small, thin shell (up to 44.1 mm), as in E. neozelandica sp. nov. However, the Recent species has a well-rounded ventral margin with a cardinal tooth and an anterior lateral tooth.
Elliptiolucina washingtonia Kiel, 2013 from the upper Oligocene Lincoln Creek Formation in Washington State (Kiel 2013) is up to 140 mm long, larger than any of the extant species, and larger than E. neozelandica sp. nov.
Elliptiolucina hetzeli (Martin, 1933) from the upper Miocene formation of Buton Island, Indonesia, redescribed by Kiel (2013), can be separated from E. neozelandica sp. nov. by its larger shell (more than 48 mm) and acutely rounded posterior margin.
Another Indonesian species, the living E. virginiae Cosel and Bouchet, 2008, can be easily separated from E. neozelandica sp. nov. by its larger size (up to 78 mm long), truncated posterior margin and narrow hinge plate.
The Recent species, Elliptiolucina ingens (Okutani, 2011) from off Amami-Oshima, southwestern Japan, differs from E. neozelandica sp. nov. in having a larger (up to 98.4 mm in length; Kuhara et al. 2014) and thick shell with a rounded posterior margin.
Stratigraphic and geographic range.—Moonlight North seep site; middle Miocene Bexhaven Limestone, North Island, New Zealand.
Subfamily Codakiinae Iredale, 1937
Genus Lucinoma Dall, 1901
Type species: Lucina filosa Stimpson, 1851; Recent, east coast of North America.
Lucinoma saetheri sp. nov.
Fig. 5.
2008 Lucinoma aff. taylori; Campbell et al. 2008: 90.
Etymology: Named for Kristian P. Saether who studied the hydrocarbon seep fauna of New Zealand for his PhD at the University of Auckland.
Type material: Holotype, UOA L4780 (Fig. 5D) from locality U23/f266 (Ugly Hill), collection AU19663. Paratypes: UOA L4772 (Fig. 5B) from locality Y16/f1174 (Moonlight North), collection AU 19923; UOA L4773 (Fig. 5A) from locality Y16/f1059 (Moonlight North), collection AU 19982; UOA L4779 (Fig. 5F) from locality U23/f266, collection 19663.
Type locality: Ugly Hill, North Island, New Zealand.
Type horizon: Ihungia Limestone, lower Miocene.
Dimensions.—See Table 3.
Table 3. Measurements (in mm) of Lucinoma
saetheri sp. nov.
Abbreviations: AL, anterior length; H, height; L, length; W, width.
|
Specimen |
Type |
L |
H |
AL |
W |
H/L |
AL/L |
W/L |
Valve |
|
UOA L4780 |
holotype |
67.4 |
63.3 |
26.6 |
37.8 |
0.91 |
0.38 |
0.56 |
both |
|
UOA L4779 |
paratype |
50.1 |
49.7 |
18.1 |
30.5 |
0.99 |
0.36 |
0.61 |
both |
|
UOA L4772 |
paratype |
35.2 |
29.1+ |
14.3 |
– |
– |
0.41 |
– |
right |
|
UOA L4773 |
paratype |
37.5 |
35.7 |
11.7 |
19.4 |
0.95 |
0.31 |
0.52 |
both |
|
UOA L4787 |
|
43.9 |
40.0 |
21.1 |
25.3 |
0.91 |
0.48 |
0.58 |
both |
|
UOA L4785 |
|
39.6 |
36.8 |
17.3 |
21.6 |
0.93 |
0.44 |
0.55 |
both |
|
UOA L4786 |
|
41.6 |
38.0 |
18.0 |
22.8 |
0.91 |
0.43 |
0.55 |
both |
Diagnosis.—Large, subcircular and moderately inflated Lucinoma with weak commarginal lamellate ribs, rather straight and gently oblique antero-dorsal margin, rounded anterior margin and weak posterior sulcus. Right valve hinge with very weakly bifid posterior cardinal tooth (3b) and relatively thick anterior cardinal tooth (3a). Ligament strong.
Description.—Shell large, up to 67.4 mm in length, rather thick, subcircular (H/L = 0.91–0.99), moderately inflated (W/L = 0.52–0.61), equivalve and slightly inequilateral. Antero-dorsal margin rather straight and gently oblique, making a near right angle turn at anterior margin end, and well-rounded ventral margin; postero-dorsal margin broadly curved, moving into posterior margin at obtuse angle; ventral margin semi-circular. Very weak sulcus present from umbo to postero-ventral corner in large specimens, including holotype and one paratype (UOA L4779). Surface sculptured with regular and widely spaced, thin commarginal lamellae; interspaces with two or three dense commarginal threads. In large specimens, these lamellae becoming low and nearly equal in strength with interstitial threads. Umbo prominent above dorsal line, prosogyrate and located at anterior one-third to two-fifths of shell length (AL/L = 0.31–0.48). Lunule lanceolate, sunken; demarcated by sharp ridge. Ligament strong, occupying about 70–80% of postero-dorsal length. Hinge of right valve with two cardinal teeth and one anterior lateral tooth; anterior tooth (3a) small, but relatively thick, oblique anteriorly; posterior tooth (3b) stout, very weakly bifid, oblique posteriorly; anterior lateral tooth (AI) very weak. Pallial line entire. Anterior adductor muscle scar elongate and long; posterior adductor muscle scar rather large and pyriform.
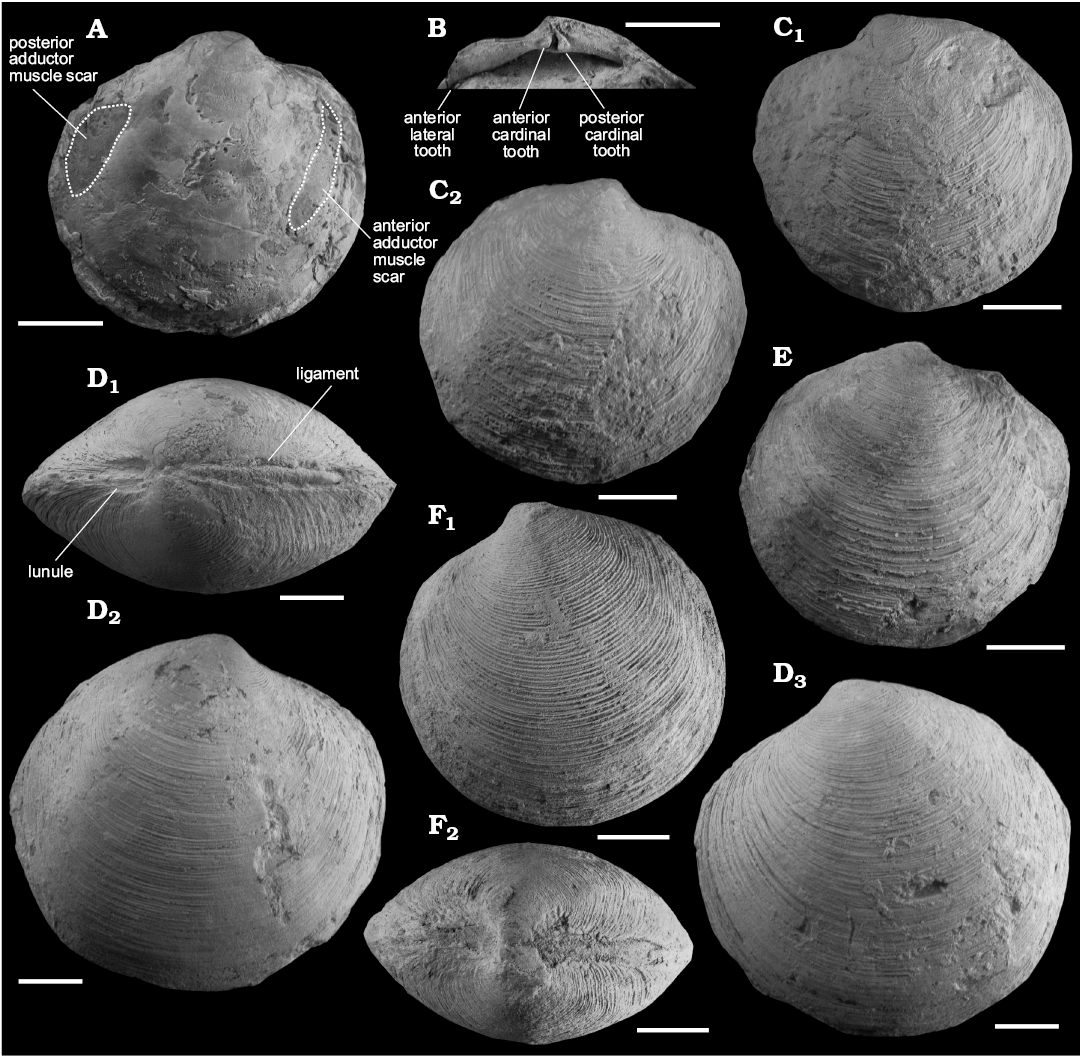
Fig. 5. Lucinid bivalve Lucinoma saetheri sp. nov. from the lower Miocene Ihungia Limestone at Ugly Hill (D, F, U23/266) and the middle Miocene Bexhaven Limestone at Moonlight North (A, C, E, Y16/1059; B, Y16/f1174), North Island, New Zealand. A. UOA L4773 (paratype), internal mould of right valve. B. UOA L 4772 (paratype), hinge plate of right valve. C. UOA L4785, left (C1) and right (C2) valves. D. UOA L4780 (holotype), dorsal view (D1), right (D2) and left (D3) valves. E. UOA L4786, right valve. F. UOA L 4779 (paratype), left valve showing lamellated commarginal riblets (F1), dorsal view (F2). Scale bars 10 mm.
Remarks.—The shell size of specimens in the seep deposits south of Hawke Bay (such as at the type locality) is often larger than those in the seeps north of Hawke Bay, up to 67.4 mm in length in the former and up to 45.0 mm in length in the latter.
Campbell et al. (2008) listed Lucinoma aff. taylori from the Rocky Knob seep deposit. However, lucinid specimens from this site are not Lucinoma taylori (Powell, 1935), instead belonging to L. saetheri sp. nov.
As noted above, Lucinoma saetheri sp. nov. does somewhat resemble L. taylori (Powell, 1935) from the lower Miocene Trikohua Formation of the Motutara area, west coast of Auckland, New Zealand. However, L. taylori is not as large as Lucinoma saetheri sp. nov. and has a shorter antero-dorsal margin, a broadly arched ventral margin, a more distinct posterior sulcus, and a less inflated shell (W/L = 0.46 in holotype; W/L = 0.52–0.61 in L. saetheri sp. nov.).
Lucinoma galathea (Marwick, 1953) is a Recent species living in and around New Zealand, and differs from L. saetheri sp. nov. by having a distinctly bifid and more oblique posterior tooth (3b), a strong anterior lateral tooth, a subtruncated anterior margin, and being smaller (maximum size up to 51.5 mm; up to 67.4 mm in L. saetheri sp. nov.).
Another large, living Lucinoma species, L. taiwanensis Cosel and Bouchet, 2008, can be easily separated from L. saetheri sp. nov. by having a distinct ridge from the beak to the anterior ventral corner.
Lucinoma perusinum (Sacco, 1901) from the middle to upper Miocene seep carbonates of northern Italy was described as L. perucina by Kiel and Taviani (2017) who allocated this species to the genus Lucinoma rather than the original designation as Dentilucina perusina by Sacco (1901). We agree with their allocation, but as Lucinoma is a neutral noun, the species name should be changed to perusinum. This Italian species is similar to L. saetheri sp. nov. in having a similar condition of the umbo and outline. However, the Italian species has a straight postero-dorsal margin and a more elongate lunule than L. saetheri sp. nov.
Lucinoma acutilineatum (Conrad, 1849) is a common species of Lucinoma from the lower Miocene seep sites of Japan. Specimens from the Kokozura Formation have a large shell (up to 58.5 mm in length; Kamada 1962). However, L. acutilineatum (Conrad, 1849) has some regularly lamellate commarginal ribs, even in large specimens, and a distinctly truncated posterior margin, so differs from L. saetheri sp. nov.
Stratigraphic and geographic range.—Ugly Hill and Haunui; lower Miocene Ihungia Limestone. Bexhaven, Moonlight North and Rocky Knob; middle Miocene Bexhaven Limestone. North Island, New Zealand.
Discussion
Characteristics of the Miocene New Zealand seep faunas.—Table 4 shows the molluscan taxa described so far from the Miocene seep sites in eastern North Island of New Zealand, based on this study and data from Saether et al. (2010a, b, 2012, 2016) and Amano et al. (2014, 2015). The species composition differs to the north and south of Hawke Bay. Bathymodiolus heretaunga, Gigantidas coseli, and Provanna marshalli are shared by both areas, but are predominant in the northern area. Some of the taxa intimately linked to seepage are confined to the northern seep sites: solemyids, Elliptiolucina neozelandica, Thyasira beui, Pliocardia? sp., and “Serradonta” kimberleyae, together with vagrant species: the taxodont bivalves Lamellinucula and Nuculana, the limid Acesta, the mud pecten Parvamussium and the cardiid Pratulum. In contrast, lucinids are predominant members of the southern seep sites. Of these, Maganodontia haunuiensis has been found only from the southern sites; whereas Lucinoma saetheri is common to both areas.
Table 4. Occurence of molluscan fossils from the seep sites in North Island, New Zealand (Amano et al. 2014, 2015; Saether et al. 2016; this study). Localities: BX, Bexhaven; KK, Karikarihuata; MN, Moonlight North; PK, Puketawa; RK, Rocky Knob; TH, Turihaua; TT, Totaranui; TW, Tauwhareparae; HA, Haunui; NG, Ngawaka; UH, Ugly Hill; WA, Wanstead; WP, Waipiro.
|
Species Localities |
North of Hawke Bay |
South of Hawke Bay |
|||||||||||
|
BX |
KK |
MN |
PK |
RK |
TH |
TT |
TW |
WP |
HA |
NG |
UH |
WA |
|
|
Lamellinucula sp. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Nuculana sp. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Solemyidae gen. et sp. indet. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Bathymodiolus (sensu lato) heretaunga |
× |
× |
× |
× |
× |
× |
× |
× |
× |
|
? |
× |
× |
|
Gigantidas coseli |
|
|
× |
|
× |
× |
|
× |
|
|
? |
× |
|
|
Acesta cf. saginata |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Parvamussium cf. maorium |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Leptomya sp. |
|
|
|
|
|
|
|
|
|
× |
|
|
|
|
Meganodontia haunuiensis sp. nov. |
|
|
|
|
|
|
|
|
|
× |
× |
× |
× |
|
Elliptiolucina neozelandica sp. nov. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Lucinoma saetheri sp. nov. |
× |
|
× |
|
× |
|
|
|
|
× |
|
× |
|
|
Thyasira beui |
× |
|
× |
|
|
× |
|
|
|
|
|
|
|
|
Pratulum quinarium |
× |
|
× |
|
|
|
|
× |
|
|
|
|
|
|
Pliocardia? sp. |
|
|
× |
× |
|
|
|
|
|
|
|
|
|
|
Notocalyptogena neozelandica |
× |
|
× |
|
× |
|
|
|
|
× |
|
× |
× |
|
Semeloidea (sensu lato) bexhavenensis |
× |
|
× |
|
|
|
|
|
|
|
|
|
|
|
“Serradonta” kimberleyae |
|
|
|
× |
× |
|
× |
|
|
|
|
|
|
|
Homalopoma? sp. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Provanna marshalli |
|
|
|
× |
× |
|
|
|
|
|
|
× |
|
|
Naticidae gen. et sp. indet. |
|
|
× |
|
|
|
|
|
|
|
|
|
|
|
Mitrella sp. |
|
|
× |
|
× |
|
|
|
|
|
|
|
|
The distribution pattern of the lucinids suggest that there was likely a palaeobathymetric difference between the northern and southern areas of Miocene seepage in eastern North Island. Elliptiolucina and some vagrant specimens occur only at the northern Moonlight North locality. Most modern Ellliptiolucina species live in the deep sea (520–2570 m) around the Philippines, Indonesia, and southern Japan (Cosel and Bouchet 2008; Okutani 2011; Kuhara et al. 2014). Of the vagrant species, Lamellinucula and Pratulum now live in shallow water around New Zealand, 5–46 m and 15–180 m, respectively (Powell 1979). However, the former lives in deeper seas of Japan (50–3000 m; Higo et al. 1999). The living depth of Nuculana ranges from 0–980 m around New Zealand (Powell 1979). Acesta saginata and Parvamussium maorinum are confined to the deep sea 1000–1400 m and 168–1300 m, respectively (Powell 1979).
The large lucinid Meganodontia occurs only in the southern seep sites. The sole Recent species, Meganodontia acetabulum, has been recorded from 246 m to 472 m water depth as dead shells (Bouchet and Cosel 2004). These shells were plausibly derived from shallower water. In addition, as described above, Lucinoma saetheri sp. nov. is larger in the southern sites than the northern ones and, according to Tunberg (1984), off Norway the Recent species Lucinoma borealis grows larger in shallower water than deeper sites. Moreover, one vagrant species from the Haunui seep site, Leptomya, now lives in shallow water (4–216 m) around New Zealand (Powell 1979).
Using the above depth ranges of living bivalves as a comparative tool, we suggest the seep sites north of Hawke Bay formed in deeper water than the seep deposits to the south of Hawke Bay, the latter inferred to have developed over a depth range of approximately 250–472 m (based on Meganodontia). This palaeobathymetric estimation is consistent with the lithological differences between seep deposits in the northern and southern areas previously noted by Campbell et al. (2008). Specifically, the southern seep deposits are sandier than those in northern areas. This lithological difference also reflects the occurrence of the two deposit feeding bivalves taxa Lamellinucula and Nuculana, which are confined to the finer-grained, northern seep deposits.
Ancient and Recent distributions of Lucinoma, Meganodontia, and Elliptiolucina.—Lucinoma first appeared in seep sites of early Oligocene age (33.9–27.8 Ma) in western Washington, USA (Goedert and Campbell 1995; Peckmann et al. 2002; Kiel 2010), and today is widely distributed among seep sites at upper bathyal depths on the Louisiana slope, off Chile, West Africa, Taiwan, Japan, and New Zealand, as well as in Mediterranean mud volcanoes and the oxygen minimum zones off California and Oman (e.g., Taylor and Glover 2010; Campbell et al. 2010). Thus, the worldwide distribution of Lucinoma indicates its adaptability to different environments, and may be one of the reasons why L. saetheri is found regionally, and in different lithologies, in Miocene seeps of New Zealand, unlike the other two lucinid genera in North Island seep deposits.
Generic lucinid compositions are similar between the southern seeps of New Zealand and the middle to late Miocene seep faunas in Italy (Kiel and Taviani 2017), containing both Meganodontia and Lucinoma. The paleobathymetry was estimated as 200–800 m for the Italian “Calcari a Lucina”, based on benthic foraminifers and oxygen isotopes (Aharon and Sen Gupta 1994). Rather shallow settings allowed Meganodontia to live in the seeps of southern New Zealand and Italy. M. haunuiensis is one of the oldest records of the genus, contemporaneous with Meganodontia sp. from the early Miocene (20.44–15.97 Ma) of Cuba. All fossil species of Meganodontia are found in hydrocarbon seep sites, while the sole modern species, M. accetablum, inhabits waters off Taiwan. The distribution of the genus has shrunk from worldwide in the Miocene to only around Taiwan today (Fig 6).
Until now, all fossil Elliptiolucina species have only been found from hydrocarbon seep sites in the Pacific. The oldest species, E. washingtonia, comes from an upper Oligocene (37.8–33.9 Ma) seep site in Washington State (Kiel 2013). The second oldest species is E. neozelandica from the middle Miocene (15.97–11.63 Ma) in New Zealand. E. hetzeli is from upper Miocene (11.63–5.333 Ma) deposits of Buton Island, Indonesia (Kiel 2013), and probably also from the lower Pliocene cold-seep deposits in Leyte, Philippines (Kase et al. 2007). In comparison, modern species of Elliptiolucina are found mainly in the Philippines, and also occur in Indonesia and Amami-Oshima, southwestern Japan (Cosel and Bouchet 2008; Okutani 2011; Kuhara et al. 2014; Glover and Taylor 2016). Of these, E. magnifica, E. labeyriei, and E. ingens were collected with vesicomyid bivalves, probably from seep sites (Cosel and Bouchet 2008; Kuhara et al. 2014). The species living in shallowest water is E. williamsae, collected from sandy substrates (Glover and Taylor 2016). The habitat of E. virginiae is unknown. The species of this genus once lived in hydrocarbon seep sites around the Pacific; since then their distribution has narrowed and their habitat has expanded to include both seep sites and also sandy environments, mainly around the Philippines (Fig. 6).
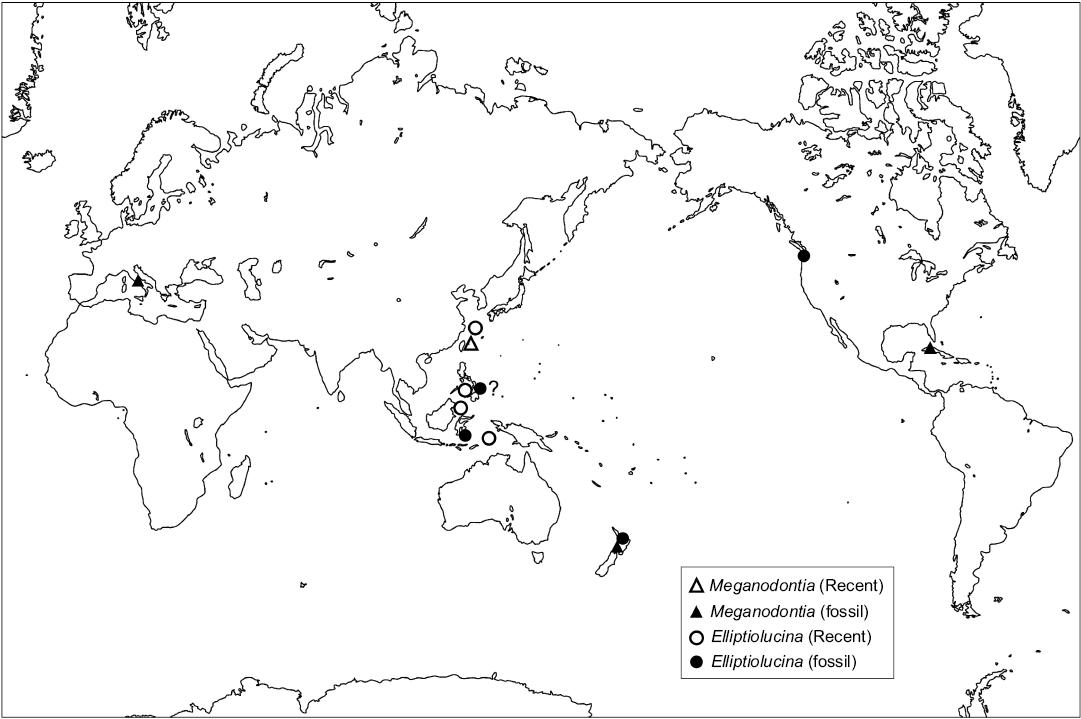
Fig. 6. Distribution of the ancient and Recent Meganodontia and Elliptiolucina. The Recent data are from Bouchet and Cosel (2004), Cosel and Bouchet (2008), and Glover and Taylor (2016). The fossil data are based on Kase et al. (2007), Kiel (2013), Kiel and Hansen (2015), Kiel and Taviani (2017), and this study.
Conclusions
This paper presents a study of lucinid bivalves from the lower to middle Miocene hydrocarbon seep carbonates in the north and south of Hawke Bay in North Island of New Zealand. Three new fossil species belonging to each of the genera Meganodontia, Elliptiolucina, and Lucinoma are described. Using modern bivalve depth ranges as a comparison, we suggest the southern area seep sites formed in shallower waters than the northern sites. Among the lucinids, the genera Meganodontia and Elliptiolucina were distributed worldwide or Circum Pacific area in the Miocene but their distribution has since narrowed to around the Philippines, Indonesia and southern Japan.
Acknowledgements
We acknowledge Kristian P. Saether (Nanjing Institute of Geology and Palaeontology, China) for his PhD study of some lucinids of New Zealand, John Taylor (The National History Museum, London, UK) for information about Meganodontia and Pegophysema, Steffen Kiel (Swedish Museum of Natural History, Stockholm, Sweden) for information on Caribbean fossil lucinids, Bruce Marshall (Te Papa Museum, Wellington, New Zealand) for showing us modern lucinid specimens from seeps of the Hikurangi margin, Neville Hudson (University of Auckland, New Zealand) for his help with fossil curation and access to material stored in the University of Auckland paleontological collections, Wilma Blom (Auckland War Memorial Museum, New Zealand) for helping us examine Powell’s and Eagles’ collections, Hiroshi Saito (National Science Museum, Tsukuba, Japan) for showing us Recent species of “Anodontia” and Elliptiolucina, and Robert G. Jenkins (Kanazawa University, Japan) for helping KA to collect some Japanese fossils. We also thank Steffen Kiel, Krzysztof Hryniewicz (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland), and an anonymous reviewer for their review and useful comments. Iain McInnes (Ernslaw One Ltd., Gisborne, New Zealand) kindly allowed access to the Moonlight North site. James and Sue Hewitt (Wanstead, New Zealand) granted access to the Haunui site, and Tim and Maggie Simcox, and Paul and Kate Dearden (Wanstead, New Zealand) gave us permission to work at Ugly Hill. This research was partly supported by a Grant-in-aid for Scientific Research from the Japan Society for the Promotion of Science (C, 23540546, 2011–2013; C, 17K05691, 2017–2019) (to KA). Field work also was supported by the Royal Society of New Zealand’s Marsden Fund (06-UOA-082) (to KAC) and a travel fund from the Royal Society (to CTSL).
References
Aharon, P. and Sen Gupta, B.K. 1994. Bathymetric reconstruction of the Miocene age “calcari a Lucina” (northern Apenines, Italy) from oxygen isotopes and benthic Foraminifera. Geo-Marine Letters 14: 219–230. Crossref
Amano, K., Little, C.T.S., Campbell, K.A., Jenkins, R.G., and Saether K.P. 2015. Paleocene and Miocene Thyasira sensu stricto (Bivalvia: Thyasiridae) from chemosynthetic communities from Japan and New Zealand. The Nautilus 129: 43–53.
Amano, K., Saether, K.P., Little, C.T.S., and Campbell, K.A. 2014. Fossil vesicomyid bivalves from Miocene hydrocarbon seep sites, North Island, New Zealand. Acta Palaeontologica Polonica 59: 421–428.
Barnes, P.M., Lamarche, G., Bialas, J., Henrys, S., Pecher, I., Netzeband, G.L., Greinert, J., Mountjoy, J.J., Pedley, K., and Crutchley, G. 2010. Tectonic and geological framework for gas hydrates and cold seeps on the Hikurangi subduction margin, New Zealand. Marine Geology 272: 26–48. Crossref
Basedow, H. 1902. Descriptions of new species of fossil Mollusca from the limestone near Edithburg (including notes by the late Professor Ralph Tate). Transactions of the Royal Society of South Australia 26: 324–325.
Bouchet, P. and Cosel, R. von 2004. The world’s largest lucinid is an undescribed species from Taiwan (Mollusca: Bivalvia). Zoological Studies 43: 704–711.
Campbell, K.A., Francis, D.A., Collins, M., Gregory, M.R., Nelson, C.S., Greinert, J., and Aharon, P. 2008. Hydrocarbon seep-carbonates of a Miocene forearc (East Coast Basin), North Island, New Zealand. Sedimentary Geology 204: 83–105. Crossref
Campbell, K.A., Nelson, C.S., Alfaro, A.C., Boyd, S., Greinert, J., Nyman, S., Grosjean, E., Logan, G.A., Gregory, M. R., Cooke S., Linke, P., Milloy, S., and Wallis, I. 2010. Geological imprint of methane seepage on the seabed and biota of the convergent Hikurangi Margin, New Zealand: Box core and grab carbonate results. Marine Geology 272: 285–306. Crossref
Chavan, A. 1969. Superfamily Lucinacea Fleming, 1828. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology Part N, Mollusca 6, Bivalvia 2, N491–N518. Geological Society of America, Boulder and University of Kansas, Lawrence.
Conrad, T.A. 1849. Description of new fresh water and marine shells. Proceedings of the Academy of Natural Sciences of Philadelphia 4: 152–156.
Cosel, R. von and Bouchet, P. 2008. Tropical deep-water lucinids (Mollusca: Bivalvia) from the Indo-Pacific: essentially unknown, but diverse and occasionally gigantic. In: V. Héros, R. Cowie, and P. Bouchet (eds.), Tropical Deep Sea Benthos, Vol. 25. Mémoires du Muséum national d’Histoire naturelle 196: 115–213.
Dall, W.H. 1901. Synopsis of the Lucinacea and of the American species. Proceedings of the U.S. National Museum 23: 779–833. Crossref
Darragh, T.A. 1970. Catalogue of Australian Tertiary Mollusca (except chitons). Memoirs of the National Museum of Victoria 31: 125–212. Crossref
Des Moulins, C. 1868, Descriptions et figures de quelques coquilles fossiles du terrain tertiaire et de la craie (Gironde, Dordogne, Royan). Actes de la Société linnéenne de Bordeaux 26: 357–379.
Eagle, M.K. 1992. A new lower Miocene species of Anodontia (Mollusca: Bivalvia). Records of the Auckland Institute and Museum 29: 103–111.
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca and Radiata of the United Kingdom; Including the Indigenous, Extirpated and Extinct Kinds; Together with Periodical and Occasional Visitants. 565 pp. Bell and Bradfute, Edinburgh.
Gill, F.L. and Little, C.T.S. 2013. A new genus of lucinid bivalve from hydrocarbon seeps. Acta Palaeontologica Polonica 58: 573–578.
Goedert, J.L. and Campbell, K.A. 1995. An early Oligocene chemosynthetic community from the Makah Formation, northestern Olympic Peninsula, Washington. The Veliger 38: 22–29.
Glover, E.A. and Taylor, J.D. 2016. Lucinidae of the Philippines: highest known diversity and ubiquity of chemosymbiotic bivalves from intertidal to bathyal depths (Mollusca: Bivalvia). In: V. Héros, E. Strong, and P. Bouchet (eds.), Tropical Deep-Sea Benthos, Vol. 29. Mémoires du Muséum national d’Histoire naturelle 208: 65–234.
Huber, M. 2015. Compendium of Bivalves 2. 907 pp. Conchbooks, Harxheim.
Higo, S., Callomon, P., and Goto, Y. 1999. Catalogue and Bibliography of the Marine Shell-Bearing Mollusca of Japan. 749 pp. Elle Scientific Publications, Yao.
Iredale, T. 1937. The Middleton and Elizabeth Reefs, South Pacific Ocean. Mollusca. Australian Zoologist 8: 232–261.
Kamada, Y. 1962. Tertiary marine Mollusca from the Joban coal-field, Japan. Palaeontological Society of Japan, Special Papers 8: 1–187.
Kanie, Y. and Sakai, T. 1997. Chemosynthetic bivalve Nipponothracia, gen. nov. from the Lower Cretaceous and Middle Miocene mudstones in Japan. Venus 56: 205–220.
Kase, T., Kurihara, Y., and Hagino, K. 2007. Middle Miocene chemosynthetic thraciid Nipponothracia gigantea (Shikama, 1968) from central Japan is a large lucinid bivalve (Lucinoidea: Mollusca). The Veliger 49: 294–302.
Kiel, S. 2010. The fossil record of vent and seep mollusks. In: S. Kiel (ed.), The Vent and Seep Biota. Topics in Geobiology 33: 255–277. Crossref
Kiel, S. 2013. Lucinid bivalves from ancient methane seeps. Journal of Molluscan Studies 79: 346–363. Crossref
Kiel, S. and Hansen, B.T. 2015. Cenozoic methane-seep faunas of the Caribbean Region. PLoS ONE 10 (10): e0140788. Crossref
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Kiel, S., Birgel, D., Campbell, K.A., Crampton, J.S., Crampton, P., Schiøler, P., and Peckmann, J. 2013. Cretaceous methane-seep deposits from New Zealand and their fauna. Palaeogeograpy, Palaeoclimatology, Palaeoecology 390: 17–34. Crossref
Kuhara, T., Kano, Y., Yoshikoshi, K., and Hashimoto, J. 2014. Shell morphology, anatomy and gill histology of the deep-sea bivalve Elliptiolucina ingens and molecular phylogenetic reconstruction of the chemosynthetic Family Lucinidae. Venus 72: 13–24.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis,locis. Vol. 1. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Ludbrook, N.H. 1978. Quaternary molluscs of the western part of the Eucla Basin. Bulletin Geological Survey of Western Australia 125: 5–286.
Martin, K. 1933. Eine neue tertiäre Molluskenfauna aus dem Indischen Archipel. Leidsche Geologische Mededeelingen 6: 7–32.
Marwick, J. 1953. A Pliocene fossil found living by the Galathea expedition. New Zealand Journal of Science and Technology (B) 35: 109–112.
Nicol, D. 1950. Recent species of the lucinoid pelecypod Fimbria. Journal of the Washington Academy of Sciences 40: 82–87.
Neumayr, M. 1884. Zur Morphologie des Bivalvenschlosses. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften. Mathematisch-Naturwissenschafliche Classe 88: 385–419.
Okutani, T. 2011. Bizarre lucinid bivalves from southwestern Japan, including a new species, and relatives in adjacent waters. Venus 69: 115–122.
Peckmann, J., Goedert, J.L., Thiel, V., Michaelis, W., and Reitner, J. 2002. A comprehensive approach to the study of methane-seep deposits from the Lincoln Creek Formation, western Washington State, USA. Sedimentology 49: 855–873. Crossref
Powell, A.W.B. 1935. Tertiary Mollusca from Motutara, west coast, New Zealand. Records of the Auckland Museum 1: 327–346.
Powell, A.W.B. 1979. New Zealand Mollusca: Marine, Land and Freshwater Shells. 355 pp. Collins, Auckland.
Reeve, L.A. 1850. Monograph of the genus Lucina. In: L.A. Reeve (ed.), Conchologica Iconica, Vol. 6., pls. v–xi. Benham and Reeve, London.
Sacco, F. 1901. I molluschi dei terreni terziarii del Piemonte e della Liguria. Parte 29. 217 pp. Carlo Clausen, Torino.
Saether, K.P., Little, C.T.S., and Campbell, K.A. 2010a. A new fossil provannid gastropod from Miocene hydrocarbon seep deposits, East Coast Basin, North Island, New Zealand. Acta Palaeontologia Polonica 55: 507–517. Crossref
Saether, K.P., Little, C.T.S., Campbell, K.A., Marshall, B.A., Collins, M., and Alfaro, A.C. 2010b. New fossil mussels (Mollusca: Bivalvia: Mytilidae) from Miocene hydrocarbon seep deposits, North Island, New Zealand, with general remarks on vent and seep mussels. Zootaxa 2577: 1–45.
Saether, K.P., Little, C.T.S., Marshall, B.A., and Campbell, K.A. 2012. Systematics and palaeoecology of a new fossil limpet (Patellogastropoda: Pectinodontidae) from Miocene hydrocarbon seep deposits, East Coast Basin, North Island, New Zealand with an overview of known fossil seep pectinodontids. Molluscan Research 32: 1–15.
Saether, K.P., Sha, J., Little, C.T.S., and Campbell, K.A. 2016. New records and a new species of bivalve (Mollusca: Bivalvia) from Miocene hydrocarbon seep deposits, North Island, New Zealand. Zootaxa 4154: 1–26. Crossref
Stewart, R.B. 1930. Gabb’s California Cretaceous and Tertiary type Lamellibranchs. Academy of Natural Sciences of Philadelphia, Special Publication 3: 1–314.
Stimpson, W. 1851. Shells of New England; A Revision of the Synonymy of the Testaceous Mollusks of New England. 58 pp. Phillips, Sampson & Co., Boston.
Taylor, J.D. and Glover, E.A. 2005. Cryptic diversity of chemosymbiotic bivalves: a systematic revision of worldwide Anodontia (Mollusca: Bivalvia: Lucinidae). Systematics and Biodiversity 3: 281–338. Crossref
Taylor, J.D. and Glover, E.A. 2006. Lucinidae (Bivalvia)—the most diverse group of chemosymbiotic molluscs. Zoological Journal of the Linnean Society 148: 421–438. Crossref
Taylor, J.D. and Glover, E.A. 2009. A giant lucinid bivalve from the Eocene of Jamaica—systematics, life habitats and chemosymbiosis (Mollusca: Bivalvia: Lucinidae). Palaeontology 52: 95–109. Crossref
Taylor, J.D. and Glover, E.A. 2010. Chemosymbiotic bivalves. In: S. Kiel (ed.), The Vent and Seep Biota. Topics in Geobiology 33: 107–136. Crossref
Taylor, J.D. and Glover, E.A. 2016. Lucinid bivalves of Guadeloupe: diversity and systematics in the context of the tropical Western Atlantic (Mollusca: Bivalvia: Lucinidae). Zootaxa 4196: 301–380. Crossref
Tunberg, B. 1984. Aspects of the population ecology of Lucinoma borealis (L.) (Bivalvia) in Raunefjorden, Western Norway. Journal of Experimental Marine Biology and Ecology 81: 87–106. Crossref
Yokoyama, M. 1928. Mollusca from the oil-field of the island of Taiwan. Report Imperial Geological Survey of Japan 101: 1–112.
Acta Palaeontol. Pol. 63 (2): 371–382, 2018
https://doi.org/10.4202/app.00461.2018