
A serpulid-Anodontia-dominated methane-seep deposit from the upper Miocene of northern Italy
STEFFEN KIEL, MARCO SAMI, and MARCO TAVIANI
Kiel, S., Sami, M., and Taviani, M. 2018. A serpulid-Anodontia-dominated methane-seep deposit from the Miocene of northern Italy. Acta Palaeontologica Polonica 63 (3): 569–577.
A limestone deposit with an unusual fauna is reported from the late Miocene of northern Italy (Ca’ Fornace site). The petrography of the carbonate and its distinct carbon isotope signature (with δ13C values as low as -57.6‰) clearly identify this limestone as an ancient methane-seep deposit. The dominant faunal elements are serpulid tubes belonging to Protis, and extremely inflated, medium-sized shells of the lucinid bivalve Anodontia mioinflata sp. nov. Also common is the small bathymodiolin Idas aff. tauroparva, plus some large specimens of the lucinid Lucinoma, and poorly preserved, medium-sized specimens of a possible vesicomyid bivalve, an arcid bivalve, small gastropods of the genera Laeviphitus (Elachisinidae), Anatoma (Scissurellidae), as well as desmophyliid and caryophyllid scleractinian corals. This faunal assemblage is quite distinct from the typical Miocene seep faunas in northern Italy, which are dominated by large bivalves of the Lucinidae (Meganodontia), Vesicomyidae (Archivesica), and Bathymodiolinae, possibly due to a shallower depositional depth of the Ca’ Fornace site.
Key words: Gastropoda, Scleractinia, Serpulidae, Bivalvia, Lucinidae, Bathymodiolinae, methane seep, Miocene, Italy, Apennines.
Steffen Kiel [steffen.kiel@nrm.se], Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden.
Marco Sami [marco.sami@cheapnet.it], Museo Civico di Scienze Naturali, 51, Via Medaglie D’Oro 51, 48018 Faenza, Italy.
Marco Taviani [marco.taviani@bo.ismar.cnr.it], Institute of Marine Sciences, Italian National Research Council, Via Gobetti 101, 40129 Bologna, Italy; Biology Department, Woods Hole Oceanographic Institution, 266 Woods Hole Rd, Woods Hole, MA 02543, USA; and Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy.
Received 28 February 2018, accepted 24 April 2018, available online 25 June 2018.
Copyright © 2018 S. Kiel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Miocene methane-seep deposits and their fauna are well-known from northern Italy. They are called “Calcare a Lucina” (CAL) due to the superabundance of the large lucinid bivalve Meganodontia hoernea (Des Moulins, 1868). Further common faunal elements include the lucinid Lucinoma perusina (Sacco, 1901), Bathymodiolus mussels, large vesicomyid clams of the genus Archivesica, and the most common gastropods are the neritid Thalassonerita megastoma and the vetigastropod Homalopoma domeniconii (Moroni 1966; Taviani 1994, 2011; Kiel and Taviani 2017). Similar faunal associations occur at present-day methane seeps throughout the world’s oceans (Sibuet and Olu 1998; Van Dover et al. 2002; Olu et al. 2010; Kiel 2016). The present-day Mediterranean Sea, however, represents an exception from this, its seep faunas being dominated by small bathymodiolins (Idas), small vesicomyids (Isorropodon) and the moderately sized lucinid Lucinoma kazani (Salas and Woodside 2002; Olu et al. 2004; Zitter et al. 2008; Taviani 2011; Taviani et al. 2013). Also Pliocene seep deposits in northern Italy lack the large bathymodiolins and vesicomyids and are dominated instead by smaller lucinid taxa (Monegatti et al. 2001; Cau et al. 2015; Kiel and Taviani 2018). It has been suggested that the present-day Mediterranean seep fauna originated after the Messinian salinity crisis (Taviani 2001, 2003; Olu et al. 2004). Here we report a late Miocene, pre-salinity crisis methane-seep fauna from Ca’ Fornace in northern Italy, with a quite different character compared to those presently known.
Institutional abbreviations.—BS, Museo Regionale di Scienze Naturali, Torino, Italy; MGGC, Museo Geologico Giovanni Capellini, University of Bologna, Italy; MSF, Museo Civico di Scienze Naturali, Faenza, Italy; NRM, Swedish Museum of Natural History, Stockholm, Sweden.
Other abbreviations.—CAL, Calcare a Lucina.
Geological setting
The Ca’ Fornace site providing the material discussed in this paper is located in the Sintria creek valley, next to Brisighella (Ravenna province) in the Romagna Apennines, near a bridge on the provincial road to Zattaglia, at 44°14’38.9”N, 11°44’0”E (Fig. 1). The presence of Miocene seep carbonates and related fauna in the Sintria creek area is recorded by Vai et al. (1997). For instance, Ca’ Fornace is about 1600 m downstream of the Tortonian CAL site of Pietralunga and 750 m from the Le Lame CAL outcrop (Aharon and Sen Gupta 1994; Peckmann et al. 2004). Both outcrops belong to classical CAL seep carbonates embedding the emblematic giant lucinid Meganodontia hoernea. Boulders, mostly ascribable to typical “Calcare a Lucina” and fewer atypical limestone blocks entrapping the fossils here considered, occur out of any coherent stratigraphic setting as alluvial erratics on Pliocene-age Argille Azzurre Formation. The atypical CAL limestone rocks have never been studied in any detail before (Taviani 2001: fig. 20.8) but seem deprived of the large CAL bivalves (cf. Taviani 1994; Kiel and Taviani 2017). In the current lack of stratigraphic sections hosting these atypical CAL carbonates their precise age is difficult to establish with confidence but conjecturally hypothesized to be late Miocene (Tortonian/?early Messinian) as for the other CAL in the region.
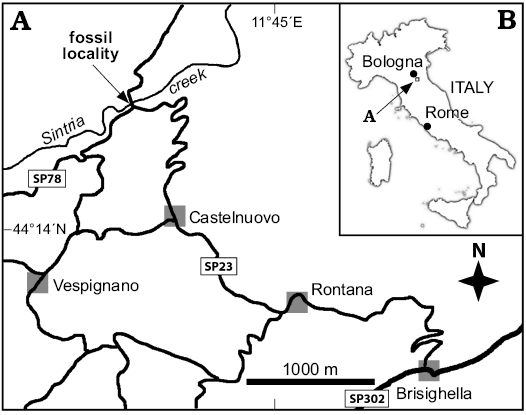
Fig. 1. Map showing location of Ca’ Fornace seep deposit in the Sintria creek valley on the map of Italy (B) and in the Sintria creek valley (A).
Material and methods
Thin sections of ca. 60 micrometer thickness were prepared and viewed and photographed under an Olympus SZX10 stereomicroscope. Samples for carbon and oxygen stable isotope analyses were extracted from the counterparts of the thin sections using a hand-held microdrill, and carbonate powders were reacted with 100% phosphoric acid at 75°C using a Kiel III online carbonate preparation line connected to a ThermoFinnigan MAT 252 mass spectrometer. All values are reported in per mil relative to PDB by assigning a δ13C value of +1.95‰ and a δ18O value of -2.20‰ to NBS19. Reproducibility was checked by replicate analysis of laboratory standards and was better than ±0.05‰.
Fossil invertebrate specimens were extracted and prepared from the rock matrix using a pneumatic hammer; small gastropods were photographed using a Hitachi S-4300 scanning electron microscope, and a Cognisys Stackshot 3X stacking image system and the Zerene Stacker software, combining 20 to 25 pictures for each composite image.
Results
Petrography and stable isotopes.—The seep deposit is dominated by micrite with varying amounts of biodetritus; hand samples with large Lucinoma shells contain few serpulid tubes but cavities lined with banded and botryoidal rim cement, and with locally abundant fecal pellets (Figs. 2A, C); late diagenetic spar in the center of the cavities is rare. Hand samples dominated by serpulid tubes have virtually no cavities (Fig. 2B). Pyrite is finely dispersed throughout all samples, and occasionally occurs in clumps, especially around fecal pellets (Fig. 2C–F), and as linings on the outer surface of mollusk shells. Wood and bone fragments occur, but only sporadically. The carbon isotope values of most carbonate phases fall in a narrow band ranging from -45 to -35‰, the main exception being those of the banded botryoidal rim cement, which reach as low as -57.6‰ (Fig. 3). The corresponding oxygen isotope values range from -0.2 to +1.75‰.

Fig. 2. Petrographic character of the late Miocene Ca’ Fornace seep deposit, Italy, illustrated by scanned thin sections (A, B) and thin section micrographs (C–F). A. Cavity in micrite (m), lined with banded and botryoidal cement (bbc), and filled with fecal pellets (fp) in the upper part and micrite in the lower part. B. Serpulid tubes cemented by micrite. C. Close-up of section in A, showing banded botryoidal cement (bbc), micrite (m), fecal pellets (fp), and clumps of pyrite (p). D. Close-up of section in B, showing details of the serpulid tubes. E. Sponge fabric (sf) around bivalve shell. F. Close-up of sponge fabric, also showing scattered pyrite framboids (p).

Fig. 3. Isotope plot; all values against the PDB standard.
Fauna.—The dominant macrofauna are serpulid worms, represented by rare large tubes (Fig. 4B) and abundant small specimens (Protis sp.) indistinguishable from those of the late Miocene Case Rovereti seep deposit (Vinn et al. 2013). Protis tubes rarely seem to attach to one another (Fig. 2D) and they are also only rarely attached to other substrates such as bivalve shells. Protis was seen in direct association with all other species, except for the large Lucinoma (Fig. 5A). A large serpulid tube was attached to a solitary coral (Fig. 4B).

Fig. 4. Fauna of the late Miocene, Ca’ Fornace seep deposit, Italy. A. The Anodontia-serpulid-association. B. Specimens of the large serpulid (ls) among numerous specimens of the small serpulid Protis sp. C. Scleractinian corals; car, caryophillid; den, dendrophyllid; all specimens housed at MSF.
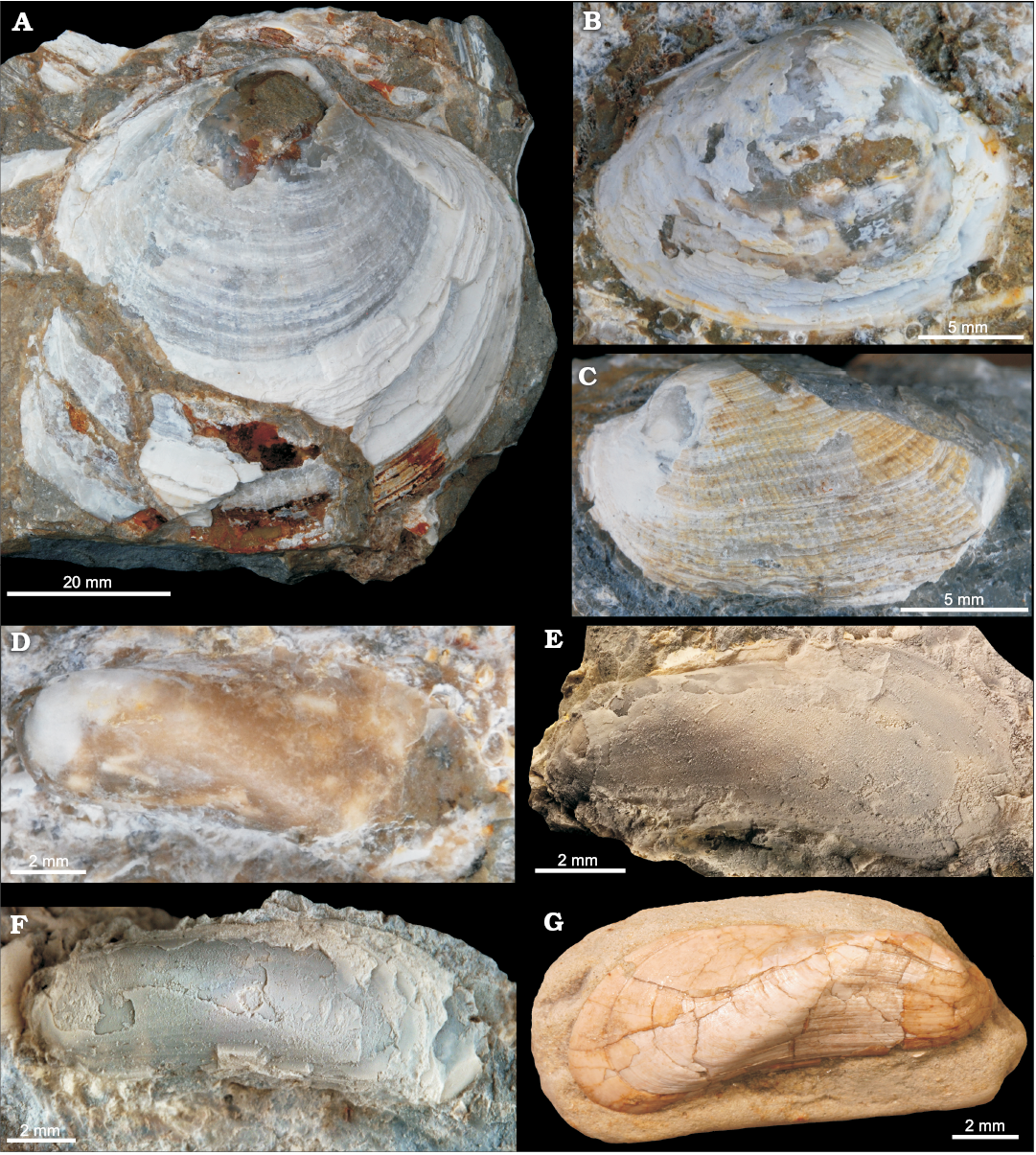
Fig. 5. Bivalves from the late Miocene Ca’ Fornace seep deposit, Italy (A–F). A. MSF 2132, large Lucinoma sp., view of right valve. B. MSF 2133, vesicomyid?, view of right valve. C. MSF 2134, arcid bivalve ?Asperarca sp., view of left valve. D–F. Idas aff. tauroparva (Sacco, 1898), medium-sized specimens consisting of left valves. D. MGGC.22005. E. MGGC.22006. F. MSF 2135. G. BS.117.03.016-1, type specimen of Modiola exbrocchii var. tauroparva Sacco, 1898, from the early Miocene of Sciolze, Italy.
Regarding chemosymbiotic bivalves, most common are the lucinid “Anodontia” mioinflata sp. nov. followed by the bathymodiolin Idas aff. tauroparva (Sacco, 1898), originally described as Modiola exbrocchii var. tauroparva (Fig. 5D–F), both of which have so far not been reported from any Miocene seep deposit in Italy. Other chemosymbiotic bivalves are far less common and include Lucinoma sp. and few specimens of a medium-sized putative vesicomyid, potentially belonging to Isorropodon (Fig. 5B). In addition, we found one specimen of a non-chemosymbiotic arcid bivalve (?Asperarca sp., Fig. 5C). Noteworthy is the occurrence of the gastropod Laeviphitus sp. (Fig. 6D, E); species of this genus have been reported from several Recent vent and seep settings world-wide, but not yet from any fossil example of these habitats. Further gastropods include small specimens belonging to Anatoma (Scissurellidae) (Fig. 6A), perhaps a Hyalogyrina (Fig. 6B), and a small “skeneid” (Fig. 6C). Specimens of solitary scleractinian corals (Fig. 4C) are not uncommon and are mostly found in hand samples that contain only few serpulid tubes. Structures resembling that of “keratose” sponge fabric (cf. Luo and Reitner 2014) were seen associated with serpulid tubes and a bivalve shell (Fig. 2E, F). The full list of species found at the Ca’ Fornace seep deposit is given in Table 1.
Table 1. List of species found at the Ca’ Fornace seep deposit.
|
Species (family) |
Figure |
|
|
Annelida |
||
| |
Protis sp. (Serpulidae) |
5A |
| |
large serpulid |
5B |
|
Bivalvia |
||
| |
arcid (?Asperarca sp.) |
6C |
| |
Idas aff. tauroparva (Bathymodiolinae) |
8 |
| |
“Anodontia” mioinflata (Lucinidae) |
9 |
| |
Lucinoma (Lucinidae) |
6A |
| |
Isorropodon? (Vesicomyidae?) |
6B |
|
Gastropoda |
||
| |
Anatoma (Scissurellidae) |
7A |
| |
skeneid |
7C |
| |
Laeviphitus (Elachisinidae) |
7D,E |
| |
Hyalogyrina? (Hyalogyrinidae) |
7B |
|
Cnidaria |
||
| |
Caryophylliidae |
5C |
| |
Dendrophyllidae |
5C |
|
Porifera |
||
| |
“keratose” fabric |
3C, D |
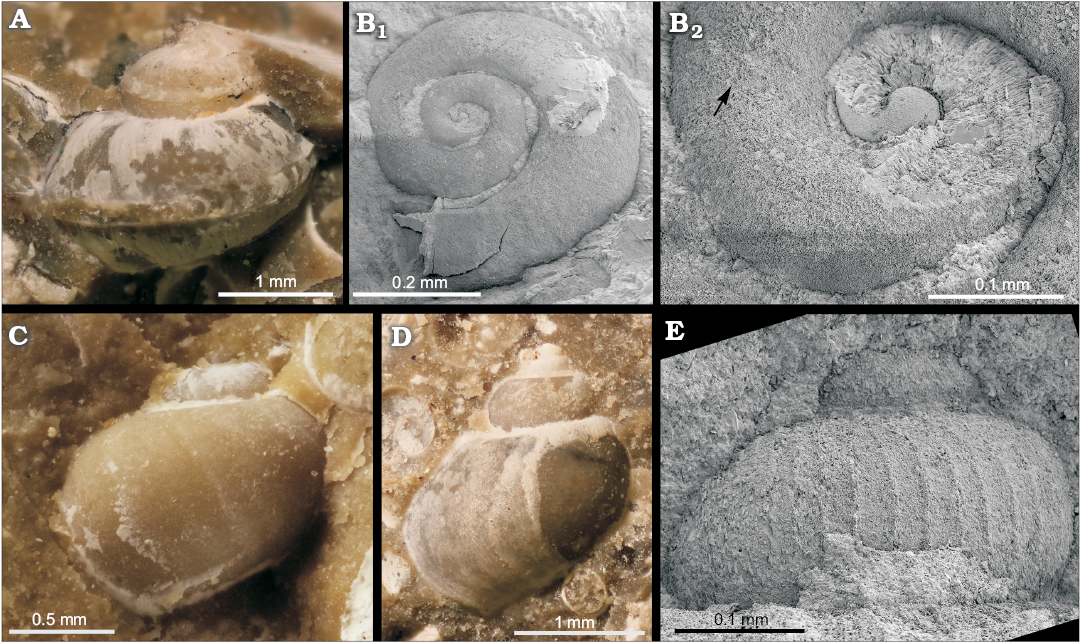
Fig. 6. Gastropods from the late Miocene Ca’ Fornace seep deposit, Italy. A. MSF 2136, Anatoma sp.; lateral view with selenizone visible at the whorl angulation. B. MSF 2137, Hyalogyrina? sp.; view of top of whole shell (B1), close-up of early whorls (B2), arrow indicates transition from protoconch to teleoconch. C. MSF 2138, skeneiform gastropod, lateral view. D, E. Laeviphitus sp. D. MSF 2139, adapertural side of adult shell. E. MSF 2140, close-up of protoconch.
Discussion
The petrography of the Ca’ Fornace seep deposit is similar to that of many other Cenozoic seep deposits in the Mediterranean region (Peckmann et al. 1999; Natalicchio et al. 2015) and world-wide (Peckmann et al. 2003; Campbell et al. 2008; Amano et al. 2010; Kiel and Hansen 2015). The very negative δ13C values of the carbonate (down to -57.6‰) clearly indicates that the oxidation of biogenic rather than thermogenic methane lead to carbonate precipitation at this site (Whiticar 1999; Peckmann and Thiel 2004). These δ13C values are among the lowest ever reported from any fossil seep carbonate. Three further sites with values around -58‰ are known: the Miocene Montepetra site in northern Italy (Taviani 1994), concretions with Conchocele from the Cretaceous of Seymour Island, Antarctica (Little et al. 2015), and a Pliocene site in the Ogikubo Formation in Nagano Prefecture, Japan, with a δ13C value of -58.8‰ (Majima et al. 2005).
With its abundant serpulid tubes and Anodontia specimens, the fauna of the Ca’ Fornace seep deposit is very different from any other Miocene seep deposit in northern Italy. Only at the late Miocene Case Rovereti site occur patches with similarly abundant tubes of the serpulid Protis (Vinn et al. 2013). Serpulids are known from modern seep sites, even in abundance, but they seem to lack particular adaptations to the seep environment (cf. Vinn et al. 2013). Quite likely they are suspension-feeders taking advantage of the abundance of bacterioplankton at seeps (cf. LaRock et al. 1994).
Among the bivalves, only the large Lucinoma is shared with other sites, but the dominant lucinid Anodontia was previously unreported from Italian Miocene seeps (Moroni 1966; Taviani 2014; Kiel and Taviani 2017). Also the moderately small, oval vesicomyid from Ca’ Fornace (Isorropodon?) differs markedly from the large and elongate Archivesica species, and also from the large, inflated “Pliocardia” italica, reported from other Miocene CAL deposits (Kiel and Taviani 2017). Vents and seeps are typically settled by large bathymodiolins whereas small bathymodiolins such as Idas typically colonize whale and wood falls (Distel et al. 2000; Lorion et al. 2013). This pattern can also be seen in the fossil record (Goedert and Squires 1990; Goedert et al. 1995; Amano and Little 2005; Kiel and Amano 2013; Jenkins et al. 2018), including the Neogene of Italy (Moroni 1966; Bertolaso and Palazzi 1994; Dominici et al. 2009; Danise et al. 2016; Kiel and Taviani 2017). The occurrence of Idas aff. tauroparva at the Ca’ Fornace seep site is thus remarkable, also because in the present-day Mediterranean Sea, Idas is widespread not only at whale falls but also at seeps (Olu et al. 2004; Ritt et al. 2010, 2012; Giusti et al. 2012). The only other fossil occurrence of Idas at a seep is Idas olympicus from an early Oligocene site in the Pysht Formation in western Washington State, USA (Kiel and Amano 2013). The small bathymodioline Adipicola apenninica Danise, Bertolaso, and Dominici, 2016, described from a Serravallian whale fall in the Emilian Apennines (Danise et al. 2016) is easily distinguished from Idas aff. tauroparva by its much longer anterior shell margin, and its more strongly curved shell.
Remarkable is the occurrence of the minute deep-water elachisinid gastropod Laeviphitus. A few species are known from extant vents and seeps world-wide, including Laeviphitus atlanticus from hydrothermal vents at the Mid-Atlantic Ridge (Warén and Bouchet 2001), L. japonicus Okutani, Fujikura, and Sasaki, 1993, from hydrothermal vents off Japan, and L. verduini van Aartsen, Bogi, and Giusti, 1989, from unspecified habitat in the Tyrrhenian Sea; plus further records of Laeviphitus sp. from seep off New Zealand and from brine pools in the Red Sea (Vestheim and Kaartvedt 2016). Regarding its paleontological history, Laeviphitus sp. was reported from the Danian (early Paleocene) cold-water coral reef facies at Faxe in Denmark (Lauridsen and Schnetler 2014) while Laeviphitus aquitanicus, has been described from the Burdigalian (early Miocene) of France (Lozouet 2015). Along with the Italian material discussed here, these records document a remarkably long geological history of this elusive genus.
Corals are not uncommon at extant seep sites (Hovland and Thomsen 1997; Hovland and Risk 2003; León et al. 2007; Lindberg et al. 2007), though their relationship to seepage is debated. An at least partial reliance on the seeping hydrocarbons has been suggested (Hovland and Thomsen 1997; Hovland et al. 1998; Hovland and Mortensen 1999), but stable isotope work in the Gulf of Mexico suggests that Lophelia pertusa branching scleractinian corals use exposed seep carbonate for attachment only after most seepage has ceased (Becker et al. 2009). Depending upon species, solitary corals may be equally attached to hard substrates or live partially embedded in mud. At any event, solitary corals have been uncommonly reported from fossil seep deposits in Washington and Oregon (Goedert and Peckmann 2005), California (Schwartz et al. 2003), and New Zealand (Campbell et al. 2008). Colonial, including dendrophyllids and caryophillid solitary corals also occur in Neogene deep-water cold seep sites in the northern Apennines (Monegatti et al. 2001; Vertino et al. in press).
Water depth has a great impact on the taxonomic composition of seep faunas (Sahling et al. 2003; Cordes et al. 2010) and such pattern is also known from the fossil record (Amano et al. 2010; Kiel 2010). Indeed, several lines of evidence suggest that the unusual character of the Ca’ Fornace seep fauna might relate to a shallower depositional depth compared to other CAL sites, possibly in the order of 150–300 m water depth: (i) extant species of Anodontia live mostly in shallow water with only a few species occurring in depth down to 500 m (Taylor and Glover 2005), and Anodontia is the most common bivalve at Ca’ Fornace; (ii) also common at Ca’ Fornace is Idas, and extant Idas tends to occur in shallower water than the larger bathymodiolin genera (Distel et al. 2000; Lorion et al. 2013); (iii) likewise, also the absence of large vesicomyids and bathymodiolins from the Ca’ Fornace site is in line with this “shallower water” hypothesis.
Conclusions
The fossiliferous limestone found at Ca’ Fornace is here identified as ancient seep deposit based on its petrography and its very negative δ13C values (as low as -57.6‰), which indicate the oxidation of biogenic rather than thermogenic methane. Its fauna is quite distinct from that of other Miocene seep deposits in northern Italy, especially due to the dominance of the lucinid Anodontia, and the presence of the small bathymodiolin Idas. This distinctive character might be the result of a shallower depositional depth of the Ca’ Fornace site compared to other “Calcare a Lucina” seep faunas in Italy. The Ca’ Fornace seep fauna broadens our knowledge of the diversity of seep faunas in the Italian Miocene.
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Heterodonta Neumayr, 1884
Family Lucinidae Fleming, 1828
Genus Anodontia Link, 1807
Type species: Anodontia alba Link, 1807 (type by monotypy); Recent, Caribbean Sea.
“Anodontia” mioinflata sp. nov.
Fig. 7.
Etymology: Combination of the age of the species (Miocene) with the globosity of the shell.
Type material: Holotype: MGGC.22003, well-preserved, articulated shell. Paratypes: MGGC.22004 (two disarticulated valves embedded in rock matrix), MSF 2141 (internal mold of right valve), MSF 2142 (articulated specimen with shell remains). All from the type locality and horizon.
Type locality: The Ca’ Fornace seep deposit in the Sintria creek valley, northern Italy.
Type horizon: Presumably late Miocene.
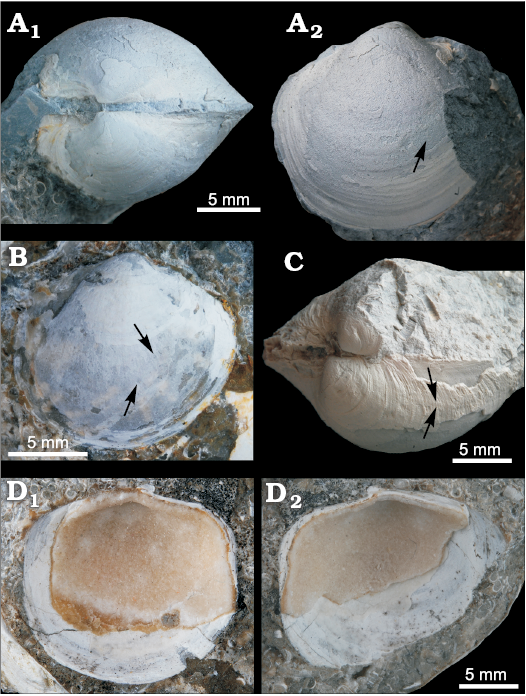
Fig. 7. The lucinid bivalve “Anodontia” mioinflata sp. nov. from the late Miocene Ca’ Fornace seep deposit, Italy. A. MGGC.22003, holotype, an articulated specimen, view on the dorsal side (A1) and on the right valve (A2), arrow indicating lower end of anterior adductor muscle scar. B. MSF 2141, paratype, right valve embedded in rock matrix, arrows indicate anterior adductor muscle scar. C. MSF 2142, paratype, view on posterodorsal side, arrows indicate the double line delimiting the posterodorsal area. D. MGGC.22004, paratype, the outer side embedded in rock matrix and carbonate mineral deposits on the inside, but showing the general shell outline; left valve (D1) and right valve (D2).
Material.—The type material, and 11 specimens housed at MSF from the type locality and horizon.
Dimensions.—The holotype is 19 mm long, 17.5 mm high, and 13.5 mm wide (articulated); the largest specimen is 23 mm long.
Diagnosis.—Subcircular, strongly inflated shell (T/L-ratio ca. 0.81–0.88); posterodorsal area large, marked by distinct ridge running from umbo to posteroventral margin; anterior adductor muscle scar elongate and long, reaching well below midline of shell, deviating from the pallial line by ca. 20°.
Description.—Solid, medium-sized, subcircular, strongly inflated shell (L/H-ratio ca. 1.1; T/L-ratio ca. 0.81–0.88); beaks prominent but only slightly elevated above hinge line; both anterior and posterior margins slightly truncate, ventral margin well-rounded; posterodorsal area large, marked on the exterior by a fine double line, and on the interior by a distinct ridge; lunule indistinct, if any; surface sculpture of fine commarginal growth lines. Anterior adductor muscle scar thin, elongate, reaching down to about 66–70% of shell height, detached from pallial line for most of its length, at an angle of ca. 20°.
Remarks.—A large number of species often differentiated through subtle morphological shell characters have been recognized in the edentulous lucinid genus Anodontia (Taylor and Glover 2005). Most living species in Anodontia inhabit shallow settings, rarely reaching down to 500 meters and none is known from bathyal seep habitats. Fossil specimens of this genus in the Mediterranean basin are currently assigned to the extant A. fragilis Philippi, 1836 and relates to shallow water situations (Chirli 2015); however, some fossil attributions of A. fragilis are questioned by Taylor and Glover (2005) and identified as Leucosphaera. Our material consists of very globose shells definitely different from any known fossil representative, and its deep-water seep habitat is almost unique although the genus has been recently discovered also in a Pliocene cold seep (Kiel and Taviani 2018). We believe that this warrants the institution of a new species. The preservation state of our material suggests a prudential collocation in “Anodontia” sensu lato.
Acknowledgements
We thank Agostina Vertino (Ghent University, Belgium) and Helmut Zibrowius (formerly Station Marine d’Endoume, Marseille, France) for aid with coral identifications, and Daniele Ormezzano (Museo Regionale di Scienze Naturali, Torino, Italy) for access to, and help with, the collection under his care. We are also grateful to the Museo Civico di Scienze Naturali, Faenza for making available their collection for this study. The reviewers Robert Jenkins (University of Kanazawa, Japan) and Cris Little (University of Leeds, UK) are thanked for their help to improve the manuscript. Financial support was provided by the Swedish Research Council (Vetenskapsrådet) through grant 2016-03920 to SK. This is Ismar-CNR, Bologna scientific contribution n. 1961.
References
van Aartsen, J.J., Bogi, C., and Giusti, F. 1989. Remarks on the genus Benthonella (Rissoidae) in Europe, and the description of Laeviphitus (nov. gen.) verduini (nov. spec.) (Epitonidae). La Conchiglia 246–249: 19–22.
Aharon, P. and Sen Gupta, B.K. 1994. Bathymetric reconstructions of the Miocene-age “calcari a Lucina” (Northern Apennines, Italy) from oxygen isotopes and benthic Foraminifera. Geo-Marine Letters 14: 219–230. Crossref
Amano, K. and Little, C.T.S. 2005. Miocene whale-fall community from Hokkaido, northern Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 215: 345–356. Crossref
Amano, K., Jenkins, R.G., Aikawa, M., and Nobuhara, T. 2010. A Miocene chemosynthetic community from the Ogaya Formation in Joetsu: evidence for depth-related ecologic control among fossil seep communities in the Japan Sea back-arc basin. Palaeogeography, Palaeoclimatology, Palaeoecology 286: 164–170. Crossref
Becker, E.L., Cordes, E.E., Macko, S.A., and Fisher, C.R. 2009. Importance of seep primary production to Lophelia pertusa and associated fauna in the Gulf of Mexico. Deep-Sea Research I 56: 786–800. Crossref
Bertolaso, L. and Palazzi, S. 1994. La posizione sistematica di Delphinula bellardii Michelotti, 1847. Bolletino Malacologico 29: 291–302.
Campbell, K.A., Francis, D.A., Collins, M., Gregory, M.R., Nelson, C.S., Greinert, J., and Aharon, P. 2008. Hydrocarbon seep-carbonates of a Miocene forearc (East Coast Basin), North Island, New Zealand. Sedimentary Geology 204: 83–105. Crossref
Cau, S., Franchi, F., Roveri, M., and Taviani, M. 2015. The Pliocene-age Stirone River hydrocarbon chemoherm complex (Northern Apennines, Italy). Marine and Petroleum Geology 66: 582–595. Crossref
Chirli, C. 2015. Malacofauna Pliocenica Toscana, Vol. 11. Bivalvia, Heteroconcha Hertwig, 1895. 234 pp. The Author, Firenze.
Cordes, E.E., Hourdez, S., and Roberts, H.H. 2010. Unusual habitats and organisms associated with the cold seeps of the Gulf of Mexico. In: S. Kiel (ed.), The Vent and Seep Biota, 315–332. Springer, Heidelberg.
Danise, S., Bertolaso, L., and Dominici, S. 2016. Bathymodioline mussel dominated Miocene whale fall from Italy. Bolletino della Società Paleontologica Italiana 55: 1–7.
Distel, D.L., Baco, A.R., Chuang, E., Morrill, W., Cavanaugh, C.M., and Smith, C.R. 2000. Do mussels take wooden steps to deep-sea vents? Nature 403: 725–726. Crossref
Dominici, S., Cioppi, E., Danise, S., Betocchi, U., Gallai, G., Tangocci, F., Valleri, G., and Monechi, S. 2009. Mediterranean fossil whale falls and the adaptation of mollusks to extreme habitats. Geology 37: 815–818. Crossref
Fleming, J. 1828. A History of British Animals, Exhibiting the Descriptive Characters and Systematical Arrangement of the Genera and Species of Quadrupeds, Birds, Reptiles, Fishes, Mollusca and Radiata of the United Kingdom; Including the Indigenous, Extirpated, and Extinct Kinds; Together With Periodical and Occasional Visitants. xxiii + 554 pp. Bell & Bradfute, Edinburgh.
Giusti, F., Mietto, P., and Sbrana, C. 2012. Il genere Idas (Mytilidae, Bathymodiolinae) in Mediterraneo, con la descrizione di quattro nuove specie. Bollettino Malacologico 48: 122–135.
Goedert, J.L. and Peckmann, J. 2005. Corals from deep-water methane-seep deposits in Paleogene strata of Western Oregon and Washington, U.S.A. In: A. Freiwald and J.M. Roberts (eds.), Cold-Water Corals and Ecosystems, 27–40. Springer, Berlin.
Goedert, J.L. and Squires, R.L. 1990. Eocene deep-sea communities in localized limestones formed by subduction-related methane seeps, southwestern Washington. Geology 18: 1182–1185. Crossref
Goedert, J.L., Squires, R.L., and Barnes, L.G. 1995. Paleoecology of whale-fall habitats from deep-water Oligocene rocks, Olympic Peninsula, Washington State. Palaeogeography, Palaeoclimatology, Palaeoecology 118: 151–158. Crossref
Hovland, M. and Mortensen, P.B. 1999. Norske korallrev of prosesser i havbunnen. 155 pp. John Grieg Forlag, Bergen.
Hovland, M. and Risk, M. 2003. Do Norwegian deep-water coral reefs rely on seeping fluids? Marine Geology 198: 83–96. Crossref
Hovland, M. and Thomsen, E. 1997. Cold-water corals—are they hydrocarbon seep related? Marine Geology 137 159–164. Crossref
Hovland, M., Mortensen, P.B., Brattegard, T., Strass, P., and Rokoengen, K. 1998. Ahermatypic coral banks off Mid-Norway: evidence for a link with seepage of light hydrocarbons. Palaios 13: 189–200. Crossref
Jenkins, R.G., Kaim, A., Amano, K., Sakurai, K., and Matsubara, K. 2018. A new Miocene whale-fall community dominated by bathymodiolin mussel Adipicola from Hobetsu area, Hokkaido, Japan. Paleontological Research 22: 105–111. Crossref
Kiel, S. 2010. On the potential generality of depth-related ecologic structure in cold-seep communities: Cenozoic and Mesozoic examples. Palaeogeography, Palaeoclimatology, Palaeoecology 295: 245–257. Crossref
Kiel, S. 2016. A biogeographic network reveals evolutionary links between deep sea hydrothermal vent and methane seep faunas. Proceedings of the Royal Society B 283: 20162337. Crossref
Kiel, S. and Amano, K. 2013. The earliest bathymodiolin mussels: Evaluation of Eocene and Oligocene taxa from deep-sea methane seep deposits in western Washington State, USA. Journal of Paleontology 87: 589–602. Crossref
Kiel, S. and Hansen, B.T. 2015. Cenozoic methane-seep faunas of the Caribbean region. PLoS ONE 10: e0140788. Crossref
Kiel, S. and Taviani, M. 2017. Chemosymbiotic bivalves from Miocene methane-seep carbonates in Italy. Journal of Paleontology 91: 444–466. Crossref
Kiel, S. and Taviani, M. 2018. Chemosymbiotic bivalves from the late Pliocene Stirone River hydrocarbon seep complex in northern Italy. Acta Palaeontologica Polonica 63: 557–568. Crossref
LaRock, P.A., Hyun, J.-H., and Bennison, B.W. 1994. Bacterioplankton growth and production at the Louisiana hydrocarbon seeps. Geo-Marine Letters 14: 104–109. Crossref
Lauridsen, B.W. and Schnetler, K.I. 2014. A catalogue of Danian gastropods from the Baunekule facies, Faxe Formation, Denmark. Geological Survey of Denmark and Greenland Bulletin 32: 1–117.
León, R., Somoza, L., Medialdea, T., González, F.J., Diaz-del-Río, V., Fernández-Puga, M.d.C., Maestro, A., and Mata, M.P. 2007. Sea-floor features related to hydrocarbon seeps in deepwater carbonate-mud mounds of the Gulf of Cádiz: from mud flows to carbonate precipitates. Geo-Marine Letters 27: 237–247. Crossref
Lindberg, B., Berndt, C., and Mienert, J. 2007. The Fugløy Reef at 70°N; acoustic signature, geologic, geomorphologic and oceanographic setting. International Journal of Earth Sciences 96: 201–213. Crossref
Link, H.F. 1806–1808. Beschreibung der Naturalien. 1: 160 pp., 2: 30 pp., 3: 38 pp. Sammlung der Universität zu Rostock, Rostock.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis,locis. Vol. 1. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Little, C.T.S., Birgel, D., Boyce, A.J., Crame, J.A., Francis, J.E., Kiel, S., Peckmann, J., Pirrie, D., Rollinson, G.K., and Witts, J.D. 2015. Late Cretaceous (Maastrichtian) shallow water hydrocarbon seeps from Snow Hill and Seymour Islands, James Ross Basin, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 418: 213–228. Crossref
Lorion, J., Kiel, S., Faure, B.M., Masaru, K., Ho, S.Y.W., Marshall, B.A., Tsuchida, S., Miyazaki, J.-I., and Fujiwara, Y. 2013. Adaptive radiation of chemosymbiotic deep-sea mussels. Proceedings of the Royal Society B 280: 20131243. Crossref
Lozouet, P. 2015. Nouvelles espèces de gastéropodes (Mollusca: Gastropoda) de l’Oligocène et du Miocène inférieur d’Aquitaine (Sud-Ouest de la France). Partie 5. Cossmanniana 17: 15–84.
Luo, C. and Reitner, J. 2014. First report of fossil “keratose” demosponges in Phanerozoic carbonates: preservation and 3D reconstruction. Naturwissenschaften 101: 467–477. Crossref
Majima, R., Nobuhara, T., and Kitazaki, T. 2005. Review of fossil chemosynthetic assemblages in Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 227: 86–123. Crossref
Monegatti, P., Raffi, S., Roveri, M., and Taviani, M. 2001. One day trip in the outcrops of Castell’Arquato Plio-Pleistocene Basin: from the Badland of Monte Giogo to the Stirone River. In: P. Monegatti, F. Cecca, and S. Raffi (eds.), International Conference Paleobiogeography and Paleoecology (2001: Piacenza, Italy, and Castell’Arquato, Italy), May 31–June 2 2001, Excursion Guidebook, 1–22. Elsevier, Paris.
Moroni, M.A. 1966. Malacofauna del “Calcare a Lucine” di S. Sofia-Forlì. Palaeontographica Italica 60: 69–87.
Natalicchio, M., Peckmann, J., Birgel, D., and Kiel, S. 2015. Seep deposits from northern Istria, Croatia: a first glimpse into the Eocene seep fauna of the Tethys region. Geological Magazine 152: 444–459. Crossref
Neumayr, M. 1884. Zur Morphologie des Bivalvenschlosses. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften 88: 385–419.
Okutani, T., Fujikura, K., and Sasaki, T. 1993. New taxa and new distribution records of deepsea gastropods collected from or near the chemosynthetic communities in the Japanese waters. Bulletin of the National Science Museum Series A (Zoology) 19: 123–143
Olu, K., Cordes, E.E., Fisher, C.R., Brooks, J.M., Sibuet, M., and Desbruyères, D. 2010. Biogeography and potential exchanges among the Atlantic equatorial belt cold-seep faunas. PLoS ONE 5: e11967. Crossref
Olu, K., Sibuet, M., Fiala-Médoni, A., Gofas, S., Salas, C., Mariotti, A., Foucher, J.-P., and Woodside, J. 2004. Cold seep communities in the deep eastern Mediterranean Sea: composition, symbiosis and spatial distribution on mud volcanoes. Deep-Sea Research I 51: 1915–1936. Crossref
Peckmann, J. and Thiel, V. 2004. Carbon cycling at ancient methane-seeps. Chemical Geology 205: 443–467. Crossref
Peckmann, J., Goedert, J.L., Heinrichs, T., Hoefs, J., and Reitner, J. 2003. The late Eocene ‘Whiskey Creek’ methane-seep deposit (western Washington State) Part II: petrology, stable isotopes, and biogeochemistry. Facies 48: 241–254. Crossref
Peckmann, J., Thiel, V., Michaelis, W., Clari, P., Gaillard, C., Martire, L., and Reitner, J. 1999. Cold seep deposits of Beauvoisin (Oxfordian; southeastern France) and Marmorito (Miocene; northern Italy): microbially induced authigenic carbonates. International Journal of Earth Sciences 88: 60–75. Crossref
Peckmann, J., Thiel, V., Reitner, J., Taviani, M., Aharon, P., and Michaelis, W. 2004. A microbial mat of a large sulfur bacterium preserved in a Miocene methane-seep limestone. Geomicrobiology Journal 21: 247–255. Crossref
Philippi, R.A. 1836. Enumeratio Molluscorum Siciliae cum viventium tum in tellure Tertiaria fossilium quae in itinere suo observavit. 303 pp. + 28 pls. Simonis Schroppii et Sociorum, Berolini.
Ritt, B., Duperron, S., Lorion, J., Lazar, C.S., and Sarrazin, J. 2012. Integrative study of a new cold-seep mussel (Mollusca: Bivalvia) associated with chemosynthetic symbionts in the Marmara Sea. Deep-Sea Research I 67: 121–132. Crossref
Ritt, B., Sarrazin, J., Caprais, J.-C., Noël, P., Gauthier, O., Pierre, C., Henry, P., and Desbruyères, D. 2010. First insights into the structure and environmental setting of cold-seep communities in the Marmara Sea. Deep-Sea Research I 57: 1120–1136. Crossref
Sahling, H., Galkin, S.V., Salyuk, A., Greinert, J., Foerstel, H., Piepenburg, D., and Suess, E. 2003. Depth-related structure and ecological significance of cold-seep communities—a case study from the Sea of Okhotsk. Deep-Sea Research I 50: 1391–1409. Crossref
Salas, C. and Woodside, J. 2002. Lucinoma kazani n. sp (Mollusca: Bivalvia): evidence of a living benthic community associated with a cold seep in the Eastern Mediterranean Sea. Deep-Sea Research I 49: 991–1005. Crossref
Schwartz, H., Sample, J.C., Weberling, K.D., Minisini, D., and Moore, J.C. 2003. An ancient linked fluid migration system: cold-seep deposits and sandstone intrusions in the Panoche Hills, California, USA. Geo-Marine Letters 23: 340–350. Crossref
Sibuet, M. and Olu, K. 1998. Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins. Deep-Sea Research II 45: 517–567. Crossref
Taviani, M. 1994. The “calcari a Lucina” macrofauna reconsidered: Deep-sea faunal oases from Miocene-age cold vents in the Romagna Apennine, Italy. Geo-Marine Letters 14: 185–191. Crossref
Taviani, M. 2001. Fluid venting and associated processes. In: G.B. Vai and I.P. Martini (eds.), Anatomy of an Orogen: the Apennines and Adjacent Mediterranean Basins, 351–366. Kluwer Academic Publishers, Dordrecht.
Taviani, M. 2003. Shaping the biogeography of the Mediterranean basin: one geologist’s perspective. Biogeographia 24: 15–22.
Taviani, M. 2011. The deep-sea chemoautotroph microbial world as experienced by the Mediterranean metazoans through time. In: J. Reitner, N.-V. Quéric, and G. Arp (eds.), Advances in Stromatolite Geobiology. Lecture Notes in Earth Sciences 131, 277–295. Springer, Berlin.
Taviani, M. 2014. Marine chemosynthesis in the Mediterranean Sea. In: S. Goffredo and Z. Dubinsky (eds.), The Mediterranean Sea: Its History and Present Challenges, 69–83. Springer, Dordrecht.
Taviani, M., Angeletti, L., Ceregato, A., Foglini, F., Froglia, C., and Trincardi, F. 2013. The Gela Basin pockmark field in the strait of Sicily (Mediterranean Sea): chemosymbiotic faunal and carbonate signatures of postglacial to modern cold seepage. Biogeosciences 10: 4653–4671. Crossref
Taylor, J.D. and Glover, E.A. 2005. Cryptic diversity of chemosymbiotic bivalves: a systematic revision of worldwide Anodontia (Mollusca: Bivalvia: Lucinidae). Systematics and Biodiversity 3: 281–338. Crossref
Vai, G.B., Taviani, M., Conti, S., and Aharon, P. (eds.) 1997. Cold-E-Vent. Hydrocarbon seepage and chemosynthesis in Tethyan relic basins: products, processes and causes. In: An international field workshop to be held in Bologna and nearby Apennines. June 23–26/1997. Abstract with Program, 58 pp. Bologna.
Van Dover, C.L., German, C.R., Speer, K.G., Parson, L.M., and Vrijenhoek, R.C. 2002. Evolution and biogeography of deep-sea vent and seep invertebrates. Science 295: 1253–1257. Crossref
Warén, A. and Bouchet, P. 2001. Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44: 116–231.
Vertino, A., Taviani, M., and Corselli, C. (in press). Spatio-temporal distribution of Mediterranean cold-water corals. In: C. Orejas and C. Jiménez (eds.), Mediterranean Cold-Water Corals: Past, Present and Future. Springer, Heidelberg.
Vestheim, H. and Kaartvedt, S. 2016. A deep sea community at the Kebrit brine pool in the Red Sea. Marine Biodiversity 46: 59–65. Crossref
Whiticar, M.J. 1999. Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane. Chemical Geology 161: 291–314. Crossref
Vinn, O., Kupriyanova, E.K., and Kiel, S. 2013. Serpulids (Annelida, Polychaeta) at Cretaceous to modern hydrocarbon seeps: ecologic and evolutionary patterns. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 35–41. Crossref
Zitter, T.A.C., Henry, P., Aloisi, G., Delaygue, G., Cagatay, M.N., Mercier de Lepinay, B., Al-Samir, M., Fornacciari, F., Tesmer, M., Pekdeger, A., Wallmann, K., and Lericolais, G. 2008. Cold seeps along the main Marmara Fault in the Sea of Marmara (Turkey). Deep-Sea Research I 55: 552–570. Crossref
Acta Palaeontol. Pol. 63 (3): 569–577, 2018
https://doi.org/10.4202/app.00472.2018