
The origin of the Pycnodonteinae and relationship between gryphaeas and true pycnodontes
IGOR N. KOSENKO
Kosenko, I.N. 2018. The origin of the Pycnodonteinae and relationship between gryphaeas and true pycnodontes. Acta Palaeontologica Polonica 63 (4): 769–778.
Berriasian (Early Cretaceous) oyster Pycnogryphaea weberae from Crimea is one of the earliest known pycnodonteine oysters. Examination of shell morphology and microstructure of this species shows that it shares characters of the subfamilies Gryphaeinae and Pycnodonteinae. Shell microstructure of Pycnogryphaea weberae lacks vesicular structure and is similar to microstructure of the genus Gryphaea. At the same time the shell of this species possesses morphological characters typical of pycnodonteins: development of chomata and well-defined circumferential curb. Pycnogryphaea weberae is regarded as a transitional taxon between the subfamilies Gryphaeinae and Pycnodonteinae and referred herein to a new genus Pycnogryphaea within the Pycnodonteinae. This suggests that the subfamily Pycnodonteinae likely originated from the Late Jurassic representatives of the genus Gryphaea (Gryphaeinae) in Berriassian. First pycnodonteins preserved shell microstructure of the genus Gryphaea but developed chomata along all the perimeter of both valves.
Key words: Bivalvia, Gryphaeinae, Pycnodonteinae, Cretaceous, Berriasian, Crimea.
Igor N. Kosenko [kosenkoin@ipgg.sbras.ru], Trofimuk Institute of Petroleum Geology and Geophysics, Novosibirsk, 630090, Russia; correspondence address: Prospekt Koptyuga, 3, Novosibirsk, 630090, Russia; Novosibirsk State University, Novosibirsk, 630090, Russia.
Received 24 April 2018, accepted 17 August 2018, available online 12 September 2018.
Copyright © 2018 I.N. Kosenko. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The subfamily Pycnodonteinae was first proposed by Stenzel in 1959. Its original diagnosis includes: well-defined commissural shelf, proximally delimited by a circumferential curb; chomata short to long, branching and vermiculate; occurrence of vesicular structure; and absence of prismatic shell layer, except for the genus Neopycnodonte Stenzel, 1971 (Fig. 1). The subfamily Pycnodonteinae ranges from the Early Cretaceous to Recent (Stenzel 1971). Stenzel (1971: N1096) emphasized that the main morphological characteristic of the subfamily Pycnodonteinae is the occurrence of vesicular shell layer.
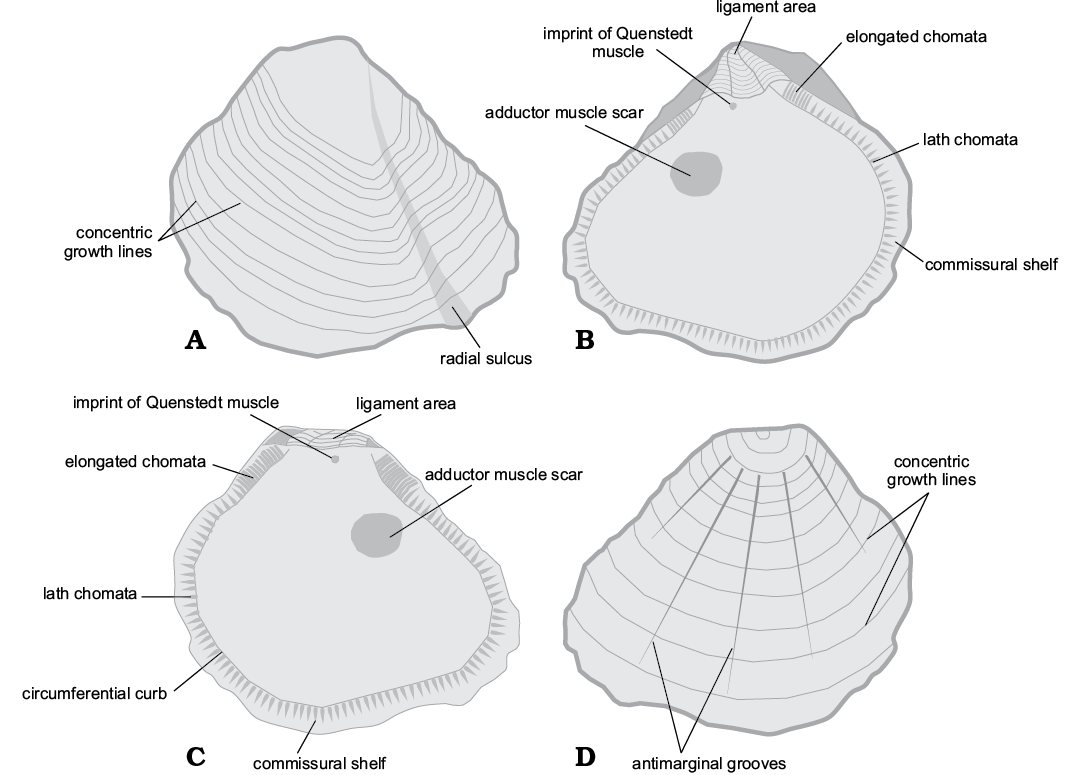
Fig. 1. Schematic drawing illustrating shell morphology of Pycnogryphaea weberae Yanin in Tschelzova, 1969. Left valve in inner (A) and outer (B) views; right valve in inner (C) and outer (D) views. The image is given approximately in natural size.
Harry (1985), Malchus (1990), Cooper (1992), and Ivanov (1995) later clarified the diagnoses of supraspecific taxa within the subfamily Pycnodonteinae. Cooper (1992) was the first to propose a hypothesized relationships within the subfamily Pycnodonteinae on the basis of the shell morphology. In this model, genus Bilobissa Stenzel, 1971 (Cooper considered Bilobissa as a genus within the Pycnodonteinae; herein Bilobissa is considered as a subgenus of Gryphaea [Gryphaeinae] as in original description of Stenzel [1971]) and genus Texigryphaea Stenzel, 1959 are closely related (Cooper 1992: fig. 1). Furthermore, the phylogenetic affinities of the subgenus Bilobissa as closely related to the Pycnodonteinae were already proposed by Stenzel (1971: N1099) in its subgeneric description. However, the stratigraphic distribution of subgenus Bilobissa and genus Texigryphaea ranges from Middle to Late Jurassic, and Albian to Cenomanian, respectively, whereas other oysters from the subfamily Pycnodonteinae were absent through Berriasian and Albian successions. This problem of a large temporal gap in the occurrence of Pycnodontein oysters was later resolved with the discovery of the species Pycnodonte miranda Bogdanova, 1980, in the Berriasian–Valanginian successions of the Mangyshlak Region (north-western Kazakhstan), as well as Gryphaea weberae Yanin in Tschelzova, 1969 in the Berriasian of Crimea. Later, Bogdanova (in Arkadiev et al. 1997) and Yanin (in Arkadiev et al. 2012) transferred G. weberae to the genus Pycnodonte Fisher de Waldheim, 1835. Thus, Pycnodonte weberae is one of the earliest pycnodontein oysters.
The aim of this study is to re-examine the morphology and microstructure of the species Pycnodonte weberae in order to understand its taxonomic affinities, and more generally the early evolution of the Pycnodonteinae. In this work, previously published data (Tschelzova 1969; Arkadiev et al. 1997, 2012) are re-examined and backed by new data on the microstructure of the species Pycnodonte weberae from the Berriasian successions of Crimea.
Institutional abbreviations.—CP MSU, Chair of Palaeontology, Moscow State University, Moscow, Russia; IPGG SB RAS, Trofimuk Institute of Petroleum Geology and Geophysics Siberian Branch of Russian Academy of Sciences, Novosibirsk, Russia; TsSGM, Central Siberian Geological Museum, Novosibirsk, Russia; VSEGEI, All Russian Research Geological Institute, St. Petersburg, Russia.
Other abbreviations.—Conv, convexity of the shell; Conv/L, convexity coefficient; H, shell height; L, shell length; L/H, elongation coefficient.
Nomenclatural acts.—The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The LSID for this publication is: urn:lsid:zoobank.org:act:DE9CA5C3-1579-460D-85CD-65A10233558F. The electronic edition of this work was published in a journal with an ISSN 0567-7920, eISSN 1732-2421, and has been archived and is available from the following digital repository: APP archive: http://www.app.pan.pl/article/item/app004942018.html.
Material and methods
The studied material was collected by Tamara N. Bogdanova (VSEGEI) in the Berriasian sections of Crimean Mountains (Pseudosubplanites grandis Subzone in the section Sarysu river; Dalmasiceras tauricum Subzone in the section Balki village; lower part of Riasanites crassicostatum Subzone in the section Chernokamenka village; beds with Symphythyris arguinensis in sections Balki village and Kuchki village; beds with Zeillerina baksanensis corresponding the uppermost Berriasian in the section Mezhgorye village) (Figs. 2, 3). The material consists of 83 left and 39 right valves of the species Pycnogryphaea weberae.
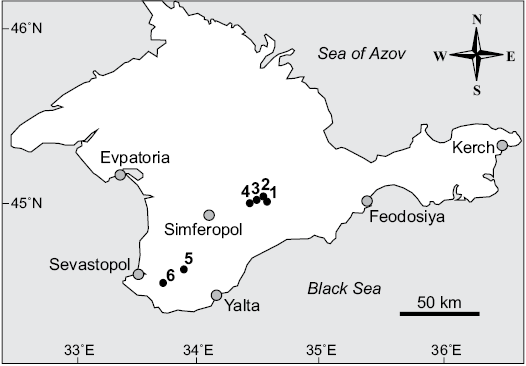
Fig. 2. Geographical position of the studied oysters locations. 1, Chernokamenka village; 2, Sarysu river; 3, Balki village; 4, Mezhgorye village; 5, Solnechnoselye village; 6, Kuchki village.
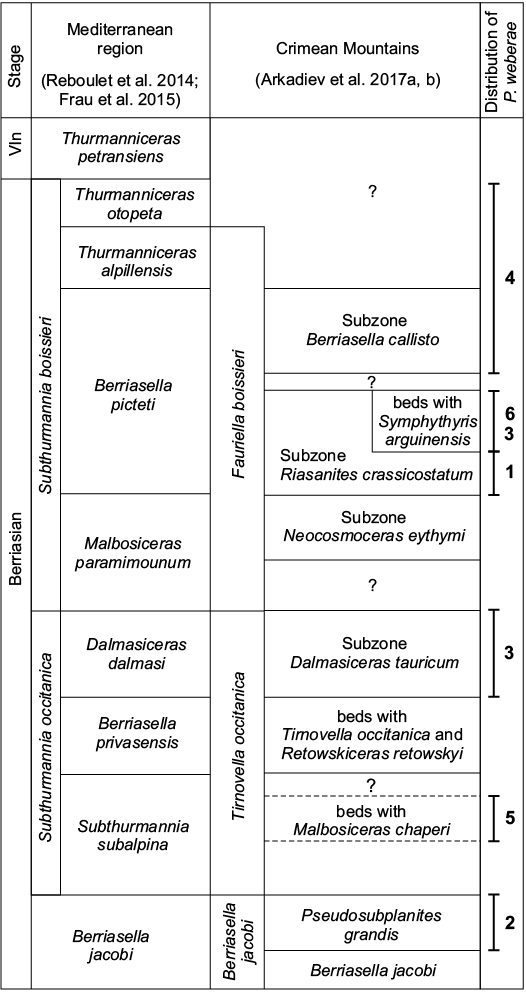
Fig. 3. Subdivision of the Berriasian stage from Crimean Mountains correlated along with Mediterranean basin, and associated distribution of the species Pycnogryphaea weberae from the different localities. 1, Chernokamenka village; 2, Sarysu river; 3, Balki village; 4, Mezhgorye village; 5, Solnechnoselye village; 6, Kuchki village; Vln, Valanginian. Stratigraphy after Arkadiev et al. 2017a, b.
Crimean upper Maastrichtian species Phygraea vesicularis (Lamarck, 1806) and Pycnodonte mirabilis (Rousseau, 1842) and Paleogene species Phygraea pseudovesicularis (Gümbel, 1861), pycnodonteine oysters were collected by INK together with Vladimir V. Arkadiev (St. Petersburg State University, Russia) and used as comparative material in the microstructural studies.
In order to understand the intraspecific variability of the species Pycnogryphaea weberae, the following morphometric parameters were measured on specimens from the Dalmasiceras tauricum Subzone (the section Balki village): shell height, shell length and convexity of the shell; calculated parameters include: elongation coefficient and convexity coefficient. Distributions graphs were prepared on the basis of the previously mentioned parameters using the freely distributed software Past 3 (Hammer et al. 2001)
The microstructure of the shells was examined in thin sections under light microscope Olympus BX60. Shell photographs were made after whitening with ammonium chloride (NH4Cl) using a camera Canon EOS 60D.
The morphological and microstructure terminology is taken from Stenzel (1971), Carter (1990), Carter et al. (2012) and Koppka (2015). The term “furcate” microstructure comes from the work of Tschelzova (1969). In longitudinal section, furcate microstructure consists on a system of tree-like branching of vertical, large, irregular calcite crystals producing irregular embranchment in the upper part of the layer (Tschelzova 1969: 25, fig. 13). Interspaces between vertical and inclined crystals are filled by thin parallel calcite lamellas (0.01–0.02 mm), which in turn are more or less parallel to the shell surface. Extinction is heterogeneous.
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Autobranchia Grobben, 1894
Cohort Ostreomorphi Férussac, 1822
Subcohort Ostreoini Férussac, 1822
Megaorder Ostreata Férussac, 1822
Superorder Ostreiformii Férussac, 1822
(= Eupteriomorphia Boss, 1982)
Order Ostreida Férussac, 1822
Suborder Ostreidina Férussac, 1822
Superfamily Ostreoidea Rafinesque, 1815
Family Gryphaeidae Vialov, 1936
Subfamily Pycnodonteinae Stenzel, 1959
Genus Pycnogryphaea nov.
Etymology: Combination of two gryphaeid generic names Pycnodonte, and Gryphaea, since the new genus shares characteristics of both.
Type species: Gryphaea weberae Yanin in Tschelzova, 1969, Berriasian of Crimea, section Sarysu river.
Species included: Type species only.
Emended diagnosis.—As for the type species (by monotypy).
Description (microstructure).—Outer shell layer of left valve consists in furcate microstructure. Inner shell layer of left valve consists mainly in irregular complex crossed foliated microstructure with sublayers of simple regular foliated microstructure. Outer shell layer of right valve consists in prismatic microstructure. Inner shell layer of right valve consists in simple regular foliated and irregular complex crossed foliated microstructures with chomata influenced crossed foliated microstructure. Chambering absent. Vesicular microstructure absent.
Remarks.—Late Triassic to Late Jurassic (or even Early Cretaceous according to Zakharov 1966 and Kosenko 2017) Gryphaea Lamarck, 1801, (Gryphaeinae) characterized by a gryphaeoid shell shape, orthogyrate beaked umbo, circular posterodorsally located posterior adductor muscle scar and radial grooves on the right valve, appears to be very similar to Pycnogryphaea. Pycnogryphaea differs from Gryphaea by chomata developed all along the circumferential curb. Due to the presence of chomata Pycnogryphaea is regarded here within the Pycnodonteinae. However, it differs from other genera of the Pycnodonteinae by the absence of vesicular structure in shell microstructure.
The Albian to Cenomanian Texigryphaea Stenzel, 1959, appears to be similar to Pycnogryphaea by the lath chomata developed along the entire circumferential curb. However, Texigryphaea differs by commonly anterodorsally compressed left valve with median radial keel. Several genera of the Pycnodonteinae, such as Crenostrea Marwick, 1931 (Paleogene of New Zealand), “Pycnodonte” gr. newberryi (Stanton, 1893) (early Late Cretaceous of North America), Labrostrea Vialov, 1945 (Late Cretaceous–Paleogene, distributed worldwide), Phygraea Vialov, 1936 (Late Cretaceous–Paleogene, worldwide), and Costeina Vialov, 1936 (Late Cretaceous of North America and Europe) had developed a gryphaeoid shell shape. However, all these taxa differ from Pycnogryphaea by the presence of a vesicular structure and chomata developed only near the ligament area. Another peculiar character of Pycnogryphaea is absence of simple herringbone regularly foliated structure typical for most Pycnodonteinae (Aqrabawi 1993; Yakushin and Ivanov 2001).
The Late Jurassic Circunula Koppka, 2015, questionably referred by Koppka (2015) to the Pycnodonteinae, recorded from the Oxfordian to Kimmeridgian of Switzerland and France, the Kimmeridgian of Germany and Crimea and the Tithonian of Czech Republic, differs from Pycnogryphaea by a large attachment area, the disappearance of chomata in adult growth stage and the non gryphaeoid shell shape.
Stratigraphic and geographic range.—Berriasian of Crimea.
Pycnogryphaea weberae (Yanin in Tschelzova, 1969)
Fig. 4.
1969 Gryphaea weberae Yanin sp. nov.; Tschelzova 1969: 56, pl. 4: 6, 7–8 (microstructure).
1997 Pycnodonte weberae (Yanin in Tschelzova, 1969); Arkadiev et al. 1997: 78, pl. 18: 8.
2012 Pycnodonte weberae (Yanin in Tschelzova, 1969); Arkadiev et al. 2012: 251, pl. 43: 16.
Type material: Holotype CP MSU 331/16 (Tschelzova 1969: pl. 4: 6).
Type locality: Central Crimea, sections Sarysu river and Balki village.
Type horizon: Middle Berriasian.
Material.—11 left valves and 4 right valves, section Sarysu river, Pseudosubplanites grandis Zone; 15 left valves, section Solnechnoselye village, beds with Malbosiceras chaperi; 53 left valves and 22 right valves, section Balki village, Dalmasiceras tauricum Zone; 1 juvenile left valve and 2 right valves, section Chernokamenka village, lower part of the Riasanites crassicostatum Subzone; 1 left valve and 5 right valves, section Balki village, beds with Symphythyris arguinensis; 1 left valve and 3 right valves, section Kuchki village, beds with Symphythyris arguinensis; 1 left valve and 3 right valves, section Mezhgorye village, beds with Zeillerina baksanensis (base of the Berriasella callipso Subzone to the Berriasian–Valanginian boundary). All material is housed in the Central Siberian Geological Museum, and referenced as collection TsSGM 2088 (TsSGM 2088/1–110).
Emended diagnosis.—Shell medium-sized, strongly inequivalve and thick. Left valve gryphaeoid, strongly convex with well-defined posterior sulcus and posterior flange, right valve flat or slightly concave. Outlines triangular or subtriangular. Attachment area very small. Left valve umbo prominent, rising well above hinge line. Both valves with concentric growth squamae, right valve with weak antimarginal grooves. Left valve lacks umbonal cavity. Ligament area triangular with triangular resilifer. Posterior adductor scar located posterodorsally, rounded (gryphaeate). Quenstedt muscle scar well distinguishable on right valve, located ventrally from resilifer. Commissural shelf narrow, well-defined, delimited by circumferential curb. Chomata developed all along the circumferential curb. Elongated chomata located near ligament area, simple lath chomata developed below elongated chomata.
Description.—Left valve gryphaeoid, moderately to highly convex (convexity coefficient Conv/L in studied samples varies from 0.36 to 0.80; see Fig. 5C), triangular to oval in outline, sometimes with opisthogyrate beak, larger than the right valve; right valve flat or slightly concave. Examined specimens up to 66 mm high and up to 72 mm long (Fig. 5A); elongation coefficient (L/H) varies in studied sample from 0.66 to 1.22 (Fig. 5B). Left valve covered by commarginal growth lines; well-defined posterior sulcus. Attachment area very small (diameter up to 10 mm in shell with 42 mm high and 47 mm long; usually not distinguishable). Right valve with well-defined commarginal growth lines and antimarginal grooves.
Ligament area: small, triangular; initially prosogyrate in some specimens; resilifer triangular, weakly concave with slightly convex bourrelets on both valves, less elevated on the right valve. Plane of ligament area depends on the degree of beak development: from almost perpendicular to the commissure plane in shells with a well-developed beak, to subparallel to the commissure plane in shells having a non-developed beak.
Internal shell characters: Umbonal cavity absent. Posterior adductor muscle rounded, relatively small, posterodorsal. Quenstedt muscle scar well distinguishable on the right valve, approximately located approximately under the resilifer (Fig. 4A–C, E, F). Commissural shelf narrow, well-defined, delimited by the circumferential curb. Chomata developed on commissural shelf all along the circumferential curb. Elongated chomata located near the ligament area, simple lath chomata developed below the elongated chomata.
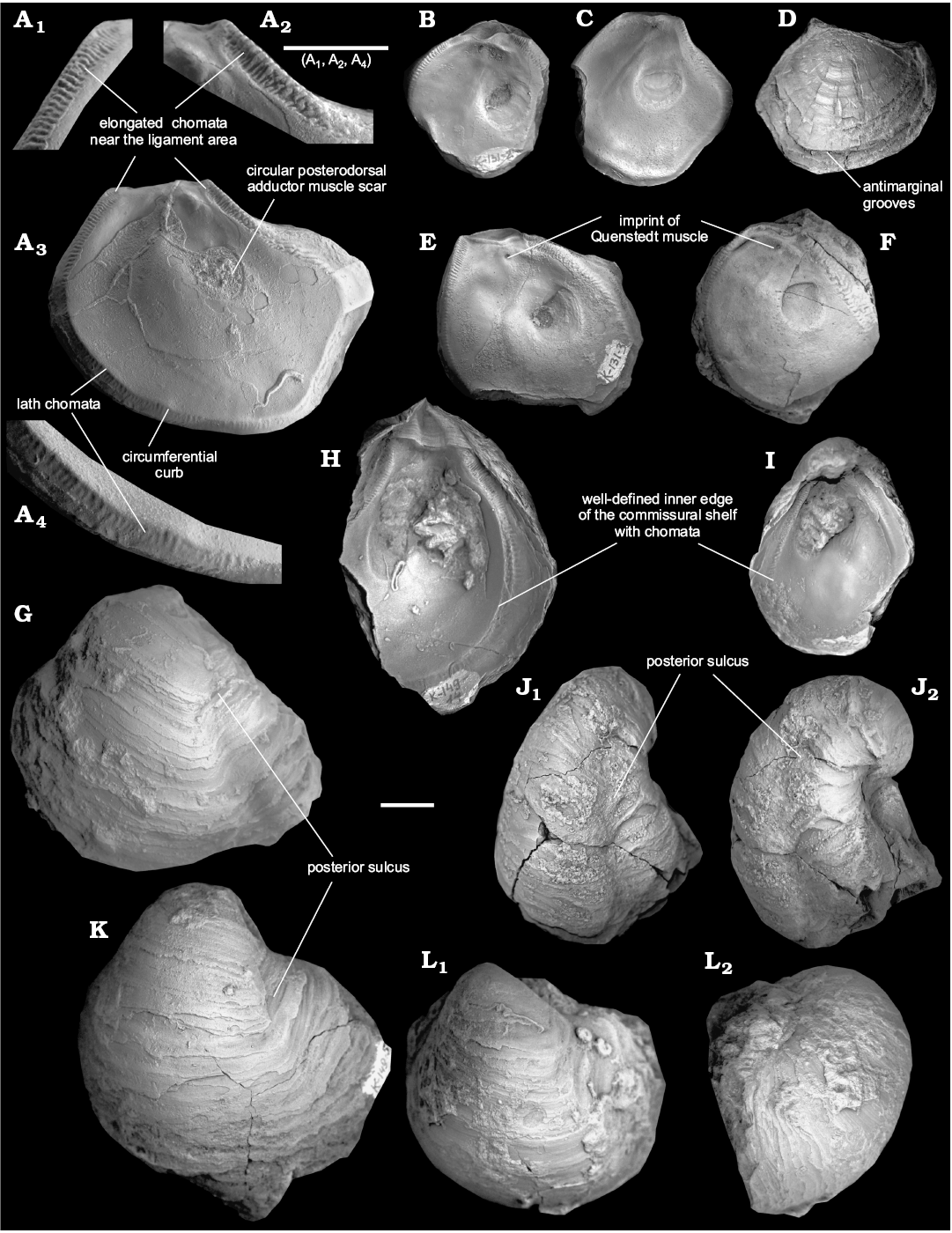
Fig. 4. Pycnodontein oyster Pycnogryphaea weberae (Yanin in Tschelzova, 1969) from Berriasian of Crimea, section Balki village, Dalmasiceras tauricum Subzone (A, C, D, F–L) and Mezhgorye village, beds with Zeillerina baksanensis (B, E). A. TsSGM 2088/28, interior of right valve (A3); elongated chomata near the hinge (A1, A2), simple lath chomata near the ventral margin of right valve (A4). B. TsSGM 2088/29, interior of right valve. C. TsSGM 2088/30, interior of right valve. D. TsSGM 2088/31, exterior of right valve. E. TsSGM 2088/34, interior of right valve. F. TsSGM 2088/32, interior of right valve. G. TsSGM 2088/3, exterior of left valve. H. TsSGM 2088/33, interior of left valve. I. TsSGM 2088/14, interior of left valve. J. TsSGM 2088/15, exterior of left valve (J1) and view from the posterior side (J2). K. TsSGM 2088/5, exterior of left valve. L. TsSGM 2088/2, exterior of left valve (L1) and view from the anterior side (L2). Scale bars 10 mm.
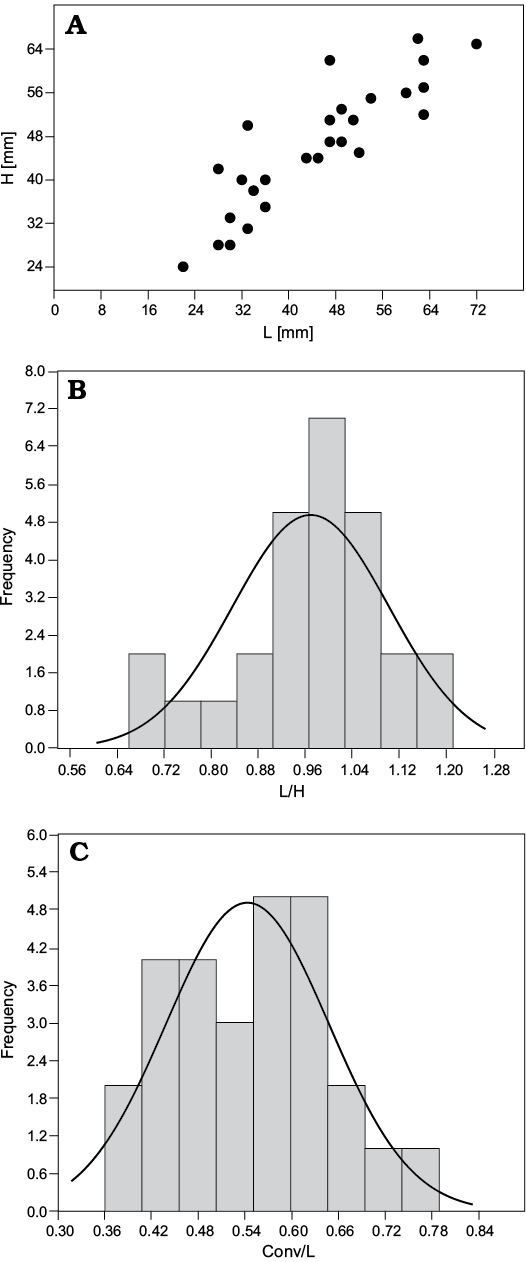
Fig. 5. Variability of pycnodontein oyster Pycnogryphaea weberae (Yanin in Tschelzova, 1969) from Berriasian of Crimea, section Balki village, Dalmasiceras tauricum Subzone. A. Length vs. height, N = 27. B. Length/height frequency, N = 27. C. Convexity/length frequency, N = 27. Conv, convexity of shell; Conv/L, convexity coefficient; H, shell height; L, shell length; L/H, elongation coefficient.
Microstructure: Outer shell layer of left valve characterized by furcate microstructure (around 300 μm) (Fig. 6D). Inner shell layer of left valve characterized by different types of irregular complex crossed foliated microstructure: homogeneous to “mosaic” irregular complex crossed foliated (Fig. 6D), low angle irregular complex crossed foliated (Fig. 6B, E, F) and dominated by high angle irregular complex crossed foliated (Figs. 6C, C, E, 7A2). Myostracum of left valve with irregular simple prismatic structure (Fig. 7A2). Outer layer of right valve consists of coarse simple prismatic microstructure (around 100 μm) (Fig. 7B3). Inner layer consists of simple regular foliated microstructure (Fig. 7A3, A4, B2, B3) with areas of chomata-influenced crossed foliated microstructure (Fig. 7B2) lapsing to low angle irregular complex crossed foliated (Fig. 7A3, A4); inflated parts of right valve consist of high angle irregular complex crossed foliated (Fig. 7A4).
Remarks.—The species Pycnogryphaea weberae was for the first time described by Yanin in the book by Tschelzova (1969), and was attributed to the genus Gryphaea. Tschelzova (1969) studied the microstructure of Pycnogryphaea weberae and showed that inner shell layer was defined by a regular foliated microstructure with rare empty chambers and subrhombic microstructure, which includes various types of irregular complex crossed foliated microstructure. Furcate microstructure was observed on the outer shell layer. Such microstructures are typical for the genus Gryphaea, and the species was included into Gryphaea by Yanin (in Tschelzova 1969). The current study of microstructure of Pycnogryphaea weberae agrees with the original description of Tschelzova (1969) (Figs. 6, 7). The presence of chomata mentioned on its original description (Yanin in Tschelzova, 1969), later encouraged Tamara N. Bogdanova (Arkadiev et al. 1997) and Boris T. Yanin (Arkadiev et al. 2012) to assign this species to the genus Pycnodonte.
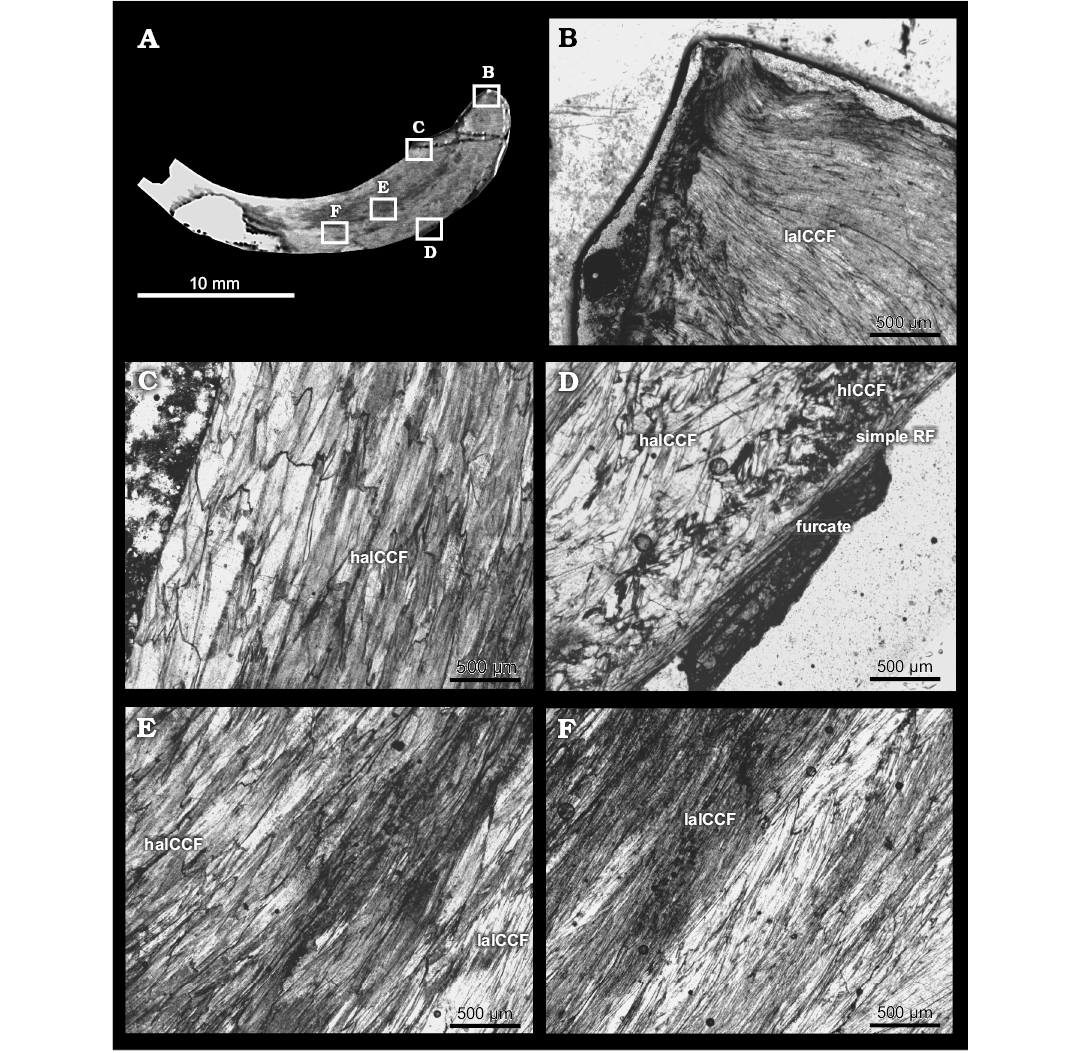
Fig. 6. Shell microstructure of pycnodontein oyster Pycnogryphaea weberae (Yanin in Tschelzova, 1969) from Berriasian of Crimea, section Balki village, Dalmasiceras tauricum Subzone. Thin section K-151-29 of left valve TsSGM 2088/35 with locations of photomicrographs (A); umbonal region represented by laCCF (B); internal part represented by haICCF (C); external part: endostracum consists of “furcate” microstructure, simple regular foliated microstructure (simple RF), hICCF and haICCF (D); central part represented by laICCF and haICCF (E, F). haICCF, high angle irregular complex crossed foliated microstructure; laICCF, low angle complex crossed foliated microstructure; simple RF, simple regular foliated microstructure.
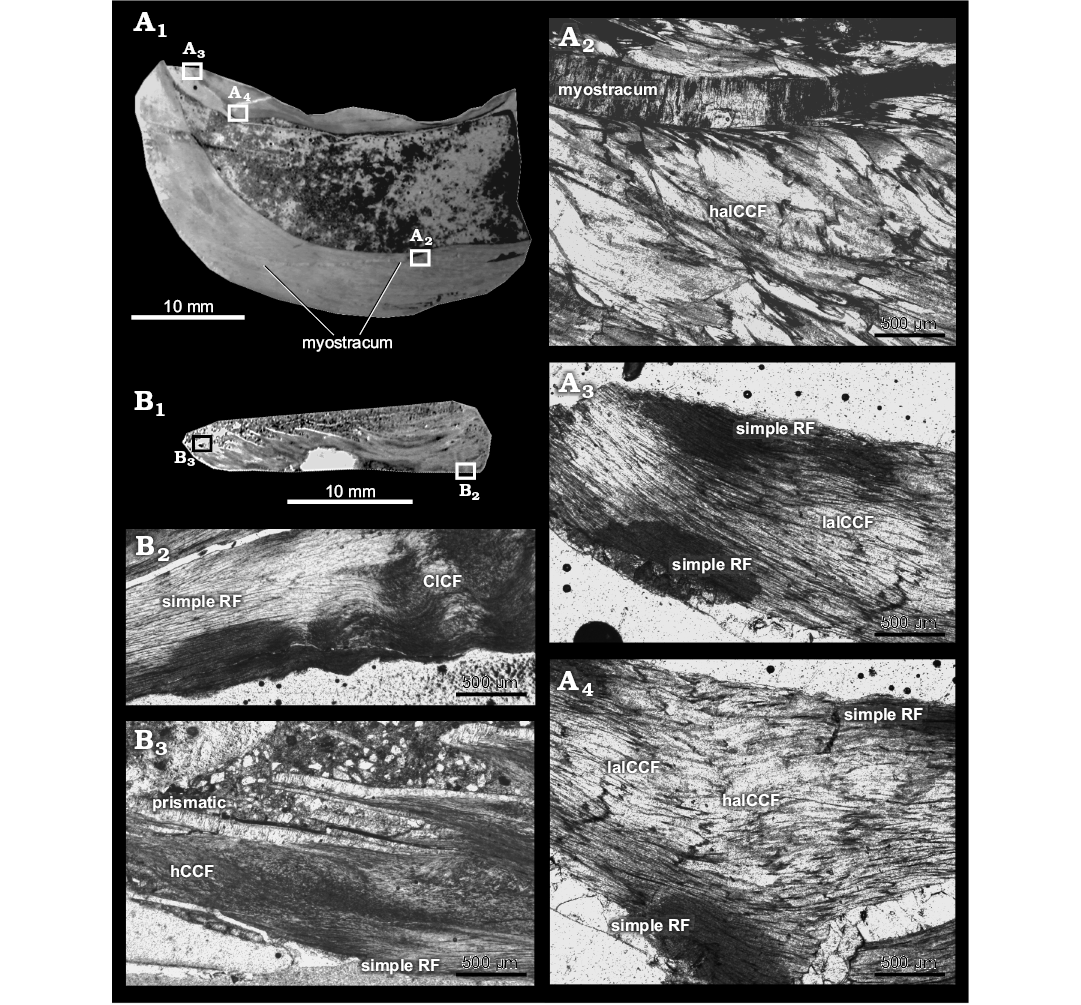
Fig. 7. Shell microstructure of pycnodontein oyster Pycnogryphaea weberae (Yanin in Tschelzova, 1969) from Berriasian of Crimea, section Balki village, Dalmasiceras tauricum Subzone. A. Thin section K-147-1 of left and right valves TsSGM 2088/36 with locations of photomicrographs (A1); inner part of left valve with myostracum consisting of irregular simple prismatic microstructure, microstructure of valve represented by haICCF (A2); umbonal part of right valve represented by simple RF and laICCF (A3); inner part of right valve represented by simple RF, laICCF and haICCF. B. Thin section K-151-10 of right valve with locations of photomicrographs (B1); umbonal part represented by simple RF and CICF (B2); outer part represented by hCCF and simple RF; exostracum consists of prismatic microstructure (B3). CICF, chomata-influenced crossed foliated microstructure; hCCF, homogeneous complex crossed foliated microstructure; haICCF, high angle irregular complex crossed foliated microstructure; laICCF, low angle complex crossed foliated microstructure; RF, regular foliated microstructure.
Discussion
Stenzel (1971: N993–N994) noted: “All Pycnodonteinae have chomata of a special character. In early, primitive genera of this subfamily (e.g., Texigryphaea), most species have low, narrow, short (1.5–2.0 mm), crowded ridgelets and corresponding grooves. On first inspection they are quite similar to those found in many Ostreinae. However, they differ in that they are crowded (about 20 per cm) and set in series and in that the difference between the right and the left valves is obliterated, that is, the ridgelets cannot be distinguished from the pits because they are so crowded”. In the present work, the chomata observed on specimens of the species Pycnogryphaea weberae are characterized by a similar structure. It agrees with the idea of Stenzel (1971) that early primitive Pycnodonteinae are defined by simple chomata, whereas more advanced Pycnodonteinae are characterized by the development of large vermiculate chomata near the hinge. No transitional Early Cretaceous oysters were found filling the gap between the true gryphaeas and pycnodontes, and Stenzel noticed (Stenzel 1971: N1077) that: “…certain traits seemingly foreshadow transitions to Texigryphaea and Pycnodonte. These traits are a broad and deep posterior sulcus dividing the left valve, an ill-defined smooth rounded radial keel running down the main body of the left valve, a slight amount of compression of the left valve in anteroposterior direction, a three-dimensional spiral growth pattern, a more opisthogyral left beak, and sharp radial grooves or gashes on the right valve”. Pycnogryphaea weberae shares all of the mentioned morphological characters typical for Late Jurassic representatives of Gryphaea (Bilobissa). It differs from the Late Jurassic Gryphaea (Bilobissa) by the development of chomata (character of the Pycnodonteinae). However, the species Pycnodonte weberae did not developed vesicular structure typical for most Pycnodonteinae (with a single exception “Pycnodonte” similis (Pusch, 1837) characterized by reduced (but not absent) vesicular structure (Machalski 1988). Thus, Pycnogryphaea weberae may be regarded as a transitional taxon between Late Jurassic Gryphaeinae and Cretaceous Pycnodonteinae. Comparisons made on shell morphology of Gryphaea (Bilobissa), Pycnogryphaea gen. nov., and Phygraea provide a basis for the reconstruction of phylogenetic line Gryphaea (Bilobissa) (Gryphaeinae; Late Jurassic) → Pycnogryphaea gen. nov. (Pycnodonteinae; Berriasian) → Phygraea (Pycnodonteinae; late Early Cretaceous).
Pycnogryphaea differs from Gryphaea by the appearance of chomata. Other morphological characters of Pycnogryphaea are typical for Gryphaeinae (circular posterodorsal adductor muscle scar, gryphaeoid shell shape, beaked umbo, posterior radial sulcus). The presence of chomata is a key morphological character that distinguished the Pycnodonteinae from the Gryphaeinae. On the other hand, chomata (as also shell chambers) independently appeared several times during the evolution of Ostreoidea in the subfamilies Exogyrinae Vialov, 1936, Pycnodonteinae Stenzel, 1959, and Liostreinae Vialov, 1983 (Malchus 1998). However, other morphological characters of Pycnogryphaea are in common with Gryphaeinae and Pycnodonteinae and there are no doubts that the genus belongs to Gryphaeinae–Pycnodonteinae phylogenetic line.
The evolutionary trend from Pycnogryphaea gen. nov. to Phygraea includes the development of chambers filled by vesicular microstructure, increase of commissural shelf and the increase complexity from short simple chomata to long vermiculate chomata located only near the ligament area.
Finally, the Albian–Cenomanian genus Texigryphaea cannot be regarded as the earliest known Pycnodonteinae and may be considered as a specialized member arising from the pycnodontes branch and restricted to North and Central America (Stenzel 1971).
The Late Jurassic genus Circunula Koppka, 2015, is defined by chomata and was consequently previously questionably included in the subfamily Pycnodonteinae by Koppka (2015), but does not share morphological characters with the genera Gryphaea and Phygraea. Based on the present study, it cannot be regarded as an ancestor of the Pycnodonteinae.
Conclusions
Pycnogryphaea gen. nov. is proposed as a new genus within the subfamily Pycnodonteinae. It includes the species Pycnodonte weberae (Yanin in Tschelzova, 1969) previously attributed to the genus Pycnodonte and is defined by well-developed short simple chomata and circumferential curb (characters of the Pycnodonteinae), as well as a shell microstructure defined by irregularly complex crossed foliated and simple regularly foliated microstructures without chambering and vesicular structure (characters of the Gryphaeinae). The stratigraphic distribution of the subfamily Pycnodonteinae is extended to the early Berriasian (Berriasella jacobi Zone). The combination of typical gryphaeine shell microstructure and pycnodonteine morphology in the described specimens of the genus Pycnogryphaea suggest it may be considered as a transitional genus between both subfamilies, Gryphaeinae and Pycnodonteinae.
Acknowledgements
I am grateful to Krzysztof Hryniewicz (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland) and Susana Damborenea (National University of La Plata, Argentina) for their review of the article, useful comments and advices during the review process; Clémentine Colpaert (Novosibirsk State University, Russia) for the assistance in correction of English language. Thanks also to Vladimir Arkadiev (St. Petersburg State University, Russia) for the organization and help in fieldwork in Crimea and to Tamara Bogdanova (VSEGEI, All Russia Research Geological Institute, St. Petersburg, Russia) for the collection of Crimean oysters. Furthermore, I am indepted to Nikolaus Malchus (ICP Catalan Institute of Palaeontology Miquel Crusafont, Barcelona, Spain) for useful advices and discussions and to Petr Yan (Trofimuk Institute of Petroleum Geology and Geophysics SB RAS, Novosibirsk, Russia) for assistance with microphotography. This paper is a contribution to the UNESCO IUGS IGCP Projects 632 and 608 and it is supported by the Russian Science Foundation under grant № 18-17-00038.
References
Arkadiev, V.V [Аrkad’ev, V.V.], Atabekyan, A.A. [Аtabekân, A.A.], Baraboshkin, E.Y. [Baraboškin, E.Û.], Bogdanova, T.N., Bugrova, I.Y. [Bugrova, I.Û.], Vladimirskaya, E.V. [Vladimirskaâ, E.V.], Golovinova, M.A., Ermolaev, V.V., Klikushin, V.G., Korobkov,A.I., Krymgoltz, G.Y. [Krymgol’c, G.Â.], Lobacheva, S.V. [Lobačёva, S.V.], Titova, M.V., Tur, N.A., and Yakushina, A.A. [Âkušina, А.А.] 1997. Аtlas melovoj fauny Ûgo-Zapadnogo Kryma. 357 pp. Sankt-Peterburgskij gosudarstvennyj gornyj institut, St. Petersburg. Crossref
Arkadiev, V.V. [Аrkad’ev, V.V.], Bogdanova, T.N., Guzhikov, A.Y. [Gužikov, А.Û.], Lobacheva, S.V. [Lobačёva, S.V.], Myshkina, N.V. [Myškina, N.V.], Platonov, E.S., Savelyeva, Yu.N. [Savel’eva, Û.N.], Shurekova, O.V. [Šurekova, O.V.], and Yanin, B.T. [Ânin, B.T.] 2012. Berrias Gornogo Kryma. 473 pp. Lema, St. Petersburg. Crossref
Arkadiev, V.V., Grishchenko, V.A., Guzhikov, A.Y., Manikin, A.G., Savelieva, Y.N., Feodorova, A.A., and Shurekova, O.V. 2017a. Ammonites and magnetostratigraphy of the Berriasian–Valanginian boundary deposits from eastern Crimea. Geologica Carpathica 68: 505–516.
Arkadiev, V., Guzhikov, A., Baraboshkin, E., Savelieva, J., Feodorova, A., Shurekova, O., Platonov, E., Manikin, A. 2017b. Biostratigraphy and magnetostratigraphy of the upper Tithonian–Berriasian of the Crimean Mountains. Cretaceous Research [published online, doi: 10.1016/j.cretres.2017.07.011].
Aqrabawi, M. 1993. Oysters (Bivalvia, Pteriomorphia) of the Upper Cretaceous rocks in Jordan. Palaeontology, stratigraphy and comparison with the Upper Cretaceous oysters of Nortwest Europe. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 75: 1–135.
Bogdanova, T.N. 1980. New Early Cretaceous oysters of Mangyshlak [in Russian]. In: G.A. Stukalina (ed.), Novye vidy drevnih rastenij i bespozvonočnyh SSSR. Vol. 5, 26–28. Nauka, Moskva.
Boss, K.J. 1982. Mollusca. In: S.P. Parker (ed.), Synopsis and Classification of Living Organisms. Vol. 1, 945–1166. McGraw-Hill, New York.
Carter, J.G. 1990. Shell microstructural data for the Bivalvia. Part IV. Order Ostreoida. In: J.G. Carter (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. 1, 347–362. Van Nostrand Reinhold, New York.
Carter, J.G., Harries, P.J., Malchus, N., Sartori, A.F., Anderson, L.C., Bieler, R., Bogan, A.E., Coan, E.V., Cope, J.C.W., Cragg, S.M., Garcia-March, J.R., Hylleberg, J., Kelley, P., Kleemann, K., K K., J., McRoberts, C., Mikkelsen, P.M., Pojeta, J. Jr., Temkin, I., Yancey, T., and Zieritz, A. 2012. Illustrated Glossary of the Bivalvia. Part N, Revised, Volume 1. Treatise Online 48: 1–209.
Cooper, M.R. 1992. Pycnodonteine oysters from the Upper Cretaceous of Zululand. Durban Museum Novitates 17: 23–57.
Férussac, A.E.J.A. de 1821–1822. Tableaux Systèmatiques des Animaux Mollusques suivis d’un Prodrome Général pour tous les Mollusques Terrestres ou Fluviatiles Vivants ou Fossiles. Première Partie, Tableaux Systématiques Généraux. 114 pp. Arthus-Bertrand, Paris.
Fisher de Waldheim, G. 1835. Lettre à M. le Baron de Férussac sur quelques genres de coquilles du Muséum Demidoff et en particulier sur quelques fossils de la Crimée. Bulletin de la Société Impériale des Naturalistes de Moscou 8: 99–123.
Frau, C., Bulot, L.G., and Wimbledon, W.A.P. 2015. Upper Tithonian Himalayitidae Spath, 1925 (Perisphinctoidea, Ammonitina) from le Chouet (Drôme, France): implications for the systematics. Geologica Carpathica 66: 117–132. Crossref
Grobben, C. 1894. Zur Kenntniss der Morphologie, der Verwandtschaftsverhältnisse und des Systems der Mollusken. Kaiserliche Akademie der Wissenschaften (Mathematisch-Naturwissenschaftlichen Classe), Sitzungsberichte 103 (1): 61–86.
Hammer, O., Harper, D.A.T., and Ryan, P.D. 2001. PAST: Paleontological Statistics software package for education and data analysis. Palaeontologia Electronica 4 (1): 1–9.
Harry, H.W. 1985. Synopsis of the supraspecific classification of living oysters (Bivalvia: Gryphaeidae and Ostreidae). The Veliger 28: 121–158.
Ivanov, A.V. 1995. New taxa of the suborder Exogyrina (Ostreoida, Bivalvia) [in Russian]. Paleontologičeskij žurnal 3: 26–36.
Koppka, J. 2015. Revision of the Bivalvia from the Upper Jurassic Reuchenette Formation, Northwest Switzerland—Ostreoidea. Zootaxa 3927: 1–117.
Kosenko, I.N. 2017. Late Jurassic–Early Cretaceous oysters from Siberia: A systematic review. Acta Palaeontologica Polonica 62: 759–778. Crossref
Lamarck, J.B. de 1801. Systéme des animaux sans vertébres, ou table général des classes, des orders et des genres de ces animaux. 432 pp. Deterville, Paris.
Lamarck, J.-B. de 1806. Mémoires sur les fossiles des environs de Paris. Explication des planches relatives aux coquilles fossiles des environs de Paris. Annales du Muséum d’Histoire Naturelle 8: 156–166.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Machalski, M. 1988. Redescription of a Danian oyster Pycnodonte simile (Pusch, 1837) from Poland. Acta Palaeontologica Polonica 33: 73–83.
Malchus, N. 1990. Revision der Kreide-Austern (Bivalvia, Pteriomorphia) Ägyptens (Biostratigraphie, Systematik). Berliner Geowissenschaftliche Abhandlungen, Reiche A 125: 1–231.
Malchus, N. 1998. Multiple parallel evolution and phylogenetic significance of shell chambers and chomata in the Ostreoidea (Bivalvia). In: P.A. Johnston and J.W. Haggart (eds.), Bivalves: an Eon of Evolution, 393–407. University of Calgary Press, Calgary.
Marwick, J. 1931. The Tertiary Mollusca of the Gisborne District. New Zealand Geological Survey, Paleontological Bulletin 13: 1–177.
Pusch, G.G. 1837. Polens Paläontologie: oder, Abbildung und Beschreibung der vorzüglichsten und der noch unbeschriebenen Petrefakten aus den Gebirgsformationen in Polen, Volhynien und den Karpathen nebst einigen allgemeinen Beiträgen zur Petrefaktenkunde und einem Versuch zur Vervollständigung der Geschechte des europäischen Auer-Ochsen. 214 pp. Schweizerbart’s Verlagshandlung, Stuttgart.
Rafinesque, C.S. 1815. Analyse de la nature ou tableau de l’Université et des corps organisés, etc. 223 pp. Jean Barravecchia, Palermo.
Reboulet, S., Szives, O., Aguirre-Urreta, B., Barragán, R., Company, M., Idakieva, V., Ivanov, M., Kakabadze, M.V., Moreno-Bedmar, J.A., Sandoval., J., Baraboshkin E.Y., Çağlar, M.K., Fözy, I., González-Arreola, C., Kenjo, S., Lukeneder, A., Raisossadat, S.N., Rawson, P.F., and Tavera, J.M. 2014. Report on the 5th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Kilian Group (Ankara, Turkey, 31st August 2013). Cretaceous Research 50: 126–137.
Rousseau, L. 1842. Description des principaux fossiles de la Crimée. In: A. de Démidoff (ed.), Voyage dans la Russie méridionale et la Crimée. Tome 2, 781–823. Ernest Bourdin, Paris.
Stanton, T.W. 1893. The Colorado formation and its invertebrate fauna. Bulletin of the United States Geological Survey 106: 1–288. Crossref
Stenzel, H.B. 1959. Cretaceous oysters of southwestern North America. In: International Geological Congress, XX Session, Mexico City, 1956, El Sistema Cretácico 1: 15–37.
Stenzel, H.B. 1971. Oysters. In: R.C. Moore (ed.), Treatise on Invertabrate Paleontology (N) Mollusca 6, Bivalvia 3, N953–N1224. University of Kansas, Lawrence and Geological Society of America, Boulder.
Tschelzova, N.A. [Čeltsova, N.A.] 1969. Značenie mikrostruktury rakoviny melovyh ustric dlâ ih sistematiki. 83 pp. Nauka, Moskva.
Vialov, O.S. 1936. On the classification of the oysters [in Russian]. Doklady Akademii Nauk SSSR 4 (1): 2–5.
Vialov, O.S. 1945. New oysters from the Paleogene of the Trans-Caspian region [in Russian]. Doklady Akademii Nauk SSSR 48 (3): 200–203.
Vialov, O.S. 1983. General classification of oysters [in Ukrainian]. Doklady Аkademii Nauk Ukrainskoj SSR, Seriâ B 11: 6–8.
Yakushin, L.N. [Âkušin, L.N.] and Ivanov, A.V. 2001. Kratkij atlas pozdnemelovyh dvustvorčatyh mollûskov (Pectinoida, Ostreoida) ûgo-vostoka Vostočno-Evropejskoj platformy.116 pp. Naučnaâ kniga, Saratov.
Zakharov, V.A. [Zaharov, V.A.] 1966. Pozdneûrskie i rannemelovye dvustvorčatye mollûski severa Sibiri (otrâd Anisomyaria) i usloviâ ih suŝestvovaniâ. 189 pp. Nauka, Moskva.
Acta Palaeontol. Pol. 63 (4): 769–778, 2018
https://doi.org/10.4202/app.00494.2018