
A middle Eocene seep deposit with silicified fauna from the Humptulips Formation in western Washington State, USA
FRIDA HYBERTSEN and STEFFEN KIEL
Hybertsen, F. and Kiel, S. 2018. A middle Eocene seep deposit with silicified fauna from the Humptulips Formation in western Washington State, USA. Acta Palaeontologica Polonica 63 (4): 751–768.
Carbonate blocks with silicified fossils were recovered from a newly recognized cold seep deposit, the Satsop Weatherwax site, in the basal Humptulips Formation, along the West Fork of Satsop River in Washington State, USA. The petrography and the stable carbon isotope signature of the carbonate, with values as low as -43.5‰, indicate that these carbonate blocks formed at an ancient methane seep. The fossils recovered from this block include five vesicomyid specimens, two fragments of a thyasirid, five specimens of the peltospirid Depressigyra, two specimens of the hyalogyrinid Hyalogyrina, 25 specimens of the neritimorph Thalassonerita eocenica, and three limpet specimens of two different species. Five species can be described as new: Nuculana acutilineata (Nuculanoidea), Desbruyeresia belliatus (Provannidae), Provanna fortis (Provannidae), Orbitestella dioi (Orbitestellidae), and Leptochiton terryiverseni (Polyplacophora). Other fossils recovered from this site are numerous serpulid tubes, echinoid spines, one brachiopod fragment and two neogastropods. Almost all species recovered belong to extant genera and the fauna has a modern character, but are different from species found in younger seeps in Washington State. This is the first record of an orbitestellid from an ancient cold seep deposit, the first fossil provannids from the Humptulips Formation, and the first fossil record of Desbruyeresia from North America.
Key words: Mollusca, Nuculoida, Caenogastropoda, deep sea, cold seeps, Eocene, Humptulips Formation, USA.
Frida Hybertsen [frida.hybertsen@nrm.se] and Steffen Kiel [steffen.kiel@nrm.se], Department of Palaeobiology, Swedish Museum of Natural History, Box 500 07, 104 05 Stockholm, Sweden.
Received 6 July 2018, accepted 11 October 2018, available online 28 November 2018.
Copyright © 2018 F. Hybertsen and S. Kiel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Cold-seep environments were first described in the 1980’s from the Gulf of Mexico (Paull et al. 1984) and the associated fauna was found to be supported by methane and hydrogen sulfide, the oxidation of which is the main energy source in chemosynthesis-based ecosystems. These specific environments have been since found from all over the world at wide depth range along continental margins and host a diverse community of specialized biota. Methane and hydrogen sulfide present at cold-seeps provide energy to a chemosynthetically driven community of sulfur-reducing bacteria in symbiosis with other deep-sea organisms, for example tubeworms, mollusks, and crustaceans (e.g., Dubilier et al. 2008; Freytag et al. 2001; Schweitzer and Feldmann 2008). Increased pH values at seeps enhance carbonate precipitation and preservation of carbonaceous shells which gives seep communities a high abundance of fossils although their preservational condition varies. Seep carbonates are distinguishable from non-seep carbonates by their petrographical characteristics such as wavy laminations, clotted micrite and yellow calcite (cf. Peckmann et al. 2001) and to confirm seep origin stable carbon isotopes are analyzed. When microbial oxidation of methane occurs the carbonates become depleted in 13C and thus seep carbonates yield negative δ13C-values.
The fossil record of cold-seep communities has been growing ever since the discovery of these ecosystems, and although it is far from being complete, it provides a growing body of evidence for the evolution of these faunas. Modern and Cenozoic seep communities are dominated by chemosymbiotic bivalves, such as vesicomyids and bathymodiolins, but bivalves are less common in Palaeozoic to mid-Mesozoic seep deposits, which were thought to be mostly dominated by brachiopods (Campbell and Bottjer 1995; Little et al. 1997). This view was challenged in recent years by findings of the modiomorphid bivalve Ataviaconcha Hryniewicz, Jakubowicz, Belka, Dopieralska, and Kaim, 2016 in Silurian and Devonian seep deposits in Morocco (Hryniewicz et al. 2016; Jakubowicz et al. 2017), and by kalenterid and anomalodesmatan bivalves in Triassic seep deposits (Peckmann et al. 2011; Kiel et al. 2017; Kiel 2018), providing evidence that bivalves and brachiopods co-existed in early seep-related ecosystems (Jakubowicz et al. 2017). Many of the bivalve and gastropod clades that are abundant and diverse at seeps today started appearing during the Jurassic and Cretaceous (Campbell and Bottjer 1995; Little and Vrijenhoek 2003; Kiel 2010), though most of the bivalves were infaunal. An exception to this is the large epifaunal to semi-infaunal kalenterid bivalve Caspiconcha Kelly in Kelly et al., 2000 that reached a world-wide distribution during the Early Cretaceous (Kiel and Peckmann 2008; Jenkins et al. 2013, 2018), but this genus declined in diversity and abundance towards the Late Cretaceous until its last occurrence in the Campanian (Jenkins et al. 2013, 2018). Its specific ecological niche was populated again by vesicomyid and bathymodiolin bivalves in middle Eocene (Amano and Kiel 2007; Kiel and Amano 2013) and these two clades have been dominating vent and seep sites until the present day. Cause and timing of this turnover and the rise of the modern fauna are the matter of debate. A hypothesis based on molecular age estimates suggests extinction and origination event around the Paleocene–Eocene Thermal Maximum (Vrijenhoek 2013), another hypothesis based on paleoecological pattern indicated marine sulfate concentrations as main driver (Kiel 2015). In any case, recently studied seep deposits from the Paleocene in the high Arctic revealed no vesicomyids or bathymodiolins, suggesting that the modern seep fauna did not originate before the Eocene (Hryniewicz et al. 2016).
The Cenozoic cold-seep deposits and associated faunas from deposits of western Washington State, USA have often been used to address taxonomic, geochemical and evolutionary inquiries due to their wide diversity and good preservation (e.g., Goedert and Squires 1990; Goedert et al. 2003; Kiel 2006; Kiel and Goedert 2006; Amano and Kiel 2007; Kiel and Goedert 2007; Schweitzer and Feldmann 2008). These deposits are most often found in the tectonic unit of the Coast Range terrane, which consists of early Eocene basalts at the base and middle Eocene to lower Miocene marine sediments on the top. The Humptulips Formation is the oldest seep-bearing rock formation in the Cenozoic accretionary prism of the Cascadia subduction zone, which forms the Olympic Mountains in Washington and is also exposed north and south of the Olympics, along the coast of British Columbia, Washington, and Oregon (Brandon and Vance 1992; Wells et al. 2014). Several of the oldest records of modern vent and seep-inhabiting clades have been reported from the Humptulips Formation, including the oldest record of the bathymodiolin mussel Vulcanidas? goederti Kiel and Amano, 2013 and the vesicomyid clam “Archivesica” cf. tschudi Olsson, 1931 (Amano and Kiel 2007; Kiel and Amano 2013), the thyasirid Maorithyas (Hryniewicz et al. 2017a), the neritid Thalassonerita (Squires and Goedert 1996a) and the galatheid crab Shinkaia (Schweitzer and Feldmann 2008). Many of these records have been used as calibration points in molecular phylogenetic studies of vent and seep clades (Kano et al. 2002; Lorion et al. 2013; Roterman et al. 2013; Vrijenhoek 2013; Valdés et al. 2013; Johnson et al. 2017), highlighting the importance of documenting the diversity of the seep fauna of the Humptulips Formation for understanding the origin and evolution of these unique ecosystems.
Institutional abbreviations.—LACMIP, Los Angeles County Museum, Invertebrate Paleontology, USA; NRM, Swedish Museum of Natural History, Stockholm, Sweden; UWBM, University of Washington, Burke Museum of Natural History and Culture, Seattle, USA.
Other abbreviations.—h, height; w, width.
Material and methods
The cold seep limestone boulder bearing the material reported here was found as float in the bed of the West Fork of the Satsop River (Fig. 1), approximately 240 m east and 200 m north of the southwest corner of Sec. 10, T. 21 N., R. 7 W., Grays Harbor County, Washington State, USA (map: Grisdale, Washington USGS quadrangle, 7.5 minute series, topo., 1990 provisional edition; coordinates: 47.317177°N, 123.565931°W). It is derived from the basal part of the Humptulips Formation (possibly the basal boulder conglomerate) and is most likely of middle Eocene age (Prothero 2001), approximately 40–42.5 Ma, or late Lutetian to early Bartonian (Goedert et al. 2013). This seep deposit is here called the Satsop Weatherwax seep deposit, named for the Satsop River and Weatherwax Ridge, the nearest named topographical feature, about a mile to the northwest. This is UWBM locality number B8951.
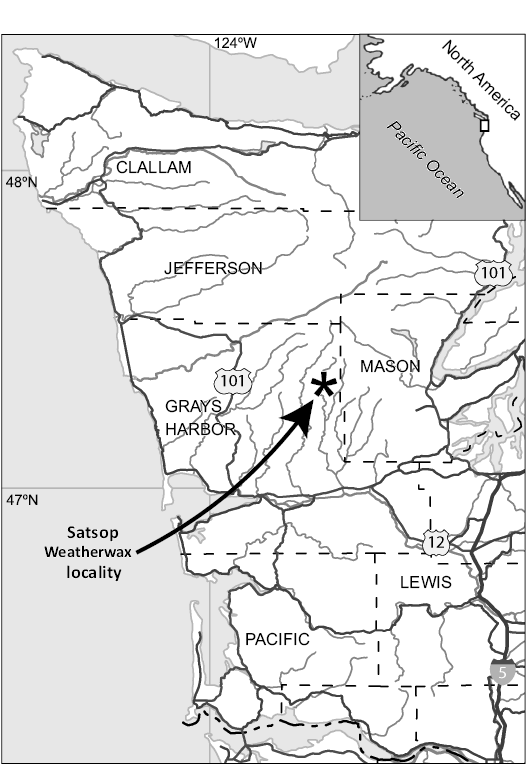
Fig. 1. Maps showing the location of the Satsop Weatherwax locality in Grays Harbor County in Washington State, USA; location of the enlarged map indicated by white rectangle.
The rock samples were hammered into ~10×10×10 cm blocks that were soaked in a 5–10% acetic acid solution. The dissolved rock fragments were washed, dried at 60°C and then examined using a binocular microscope. Some specimens were chosen for scanning electron microscopy; they were mounted and coated with gold and photographed using a Hitachi-4300 SEM at NRM. Macrofossils were coated with ammonium chloride and photographed using a Nikon D80 for capturing different angles and Cognisys Stackshot 3X stacking image system and the Zerene Stacker software, combining 20–25 pictures for each composite image.
Samples for carbon and oxygen stable isotope analyses were extracted from the counterparts of the thin sections using a hand-held microdrill, and carbonate powders were reacted with 100% phosphoric acid at 75°C using a Kiel III online carbonate preparation line connected to a ThermoFinnigan MAT 252 mass spectrometer. All values are reported in per mil relative to PDB by assigning a δ13C value of +1.95‰ and a δ18O value of -2.20‰ to NBS19. Reproducibility was checked by replicate analysis of laboratory standards and is better than ±0.05‰.
Results
Petrography and stable isotopes.—The carbonate is dominated by dark micrite, with occasional cavities lined by rim cement and sometimes filled with lighter micrite or calcite spar (Fig. 2). Silicified mollusks are abundant in patches and are also scattered throughout the deposit. The carbon isotope signature of the carbonates ranges from -43.5 to -38.0‰, with values of the micrite being slightly more negative than those of the rim cements; the corresponding oxygen isotope values range from -7.8 to -2.5‰ (Fig. 3).

Fig. 2. Scanned polished slab of carbonate from the middle Eocene Satsop Weatherwax seep deposit showing its main petrographic features.
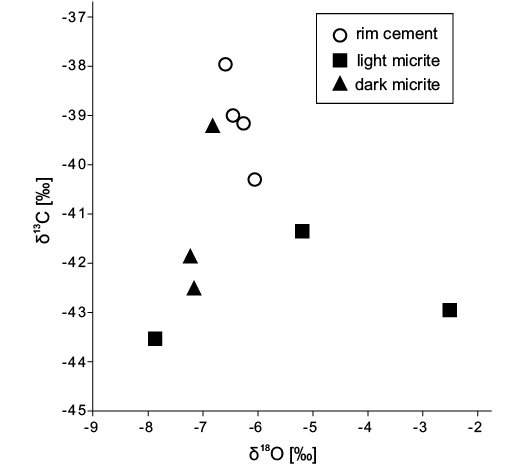
Fig. 3. Cross-plot of carbon and oxygen isotope data of the middle Eocene Satsop Weatherwax seep deposit; all values are given relative to the PDB standard.
Fauna.—The Satsop Weatherwax seep deposit contains a variety of fossils including rare echinoderms and worm tubes, and abundant molluscs of the genera typically associated with deep-sea methane seeps. Their aragonitic shells have been silicified by the dissolution of calcium carbonate and precipitation of silica occurring at deep sea methane seeps and the preservation is mostly poor, however the great abundance makes this a lucrative site for seep fossils. Most taxa are briefly discussed below; new taxa are described in detail in the Systematic Paleontology section. The full list of speciesis shown in Table. 1.
Table 1. The fossils found at the Satsop Weatherwax seep deposit.
|
Phylum/Class |
Species |
Total number |
Figure |
|
Annelida |
Serpulid tube fragments |
5 |
11B, D |
|
Bivalvia |
Vesicomyid |
5 |
5 |
|
Thyasira sp. |
8 |
4 |
|
|
Nuculana acutilineata |
>300 |
12 |
|
|
Brachiopoda |
Discinisca sp. |
1 |
11A |
|
Echinoidea |
Echinoid spines |
2 |
11E |
|
Gastropoda |
Depressigyra sp. |
5 |
7 |
|
Thalassonerita eocenica |
25 |
8 |
|
|
Gastropod limpet 1 |
2 |
6A, B |
|
|
Gastropod limpet 2 |
1 |
6C |
|
|
Desbruyeresia belliatus |
>230 |
13 |
|
|
Provanna fortis |
6 |
14 |
|
|
Colus? sp. |
2 |
9 |
|
|
Hyalogyrina sp. |
2 |
10 |
|
|
Orbitestella dioi |
2 |
15 |
|
|
Polyplacophora |
Leptochiton terryiverseni |
4 |
16 |
Bivalves: These include a thyasirid, a vesicomyid, and a new species of Nuculana described below. The thyasirid is identified as Thyasira (Thyasira) sp. because the two available shell fragments show an edentulous hinge, a deep posterior sulcus and a deep submarginal sulcus separating a distinct auricle (Fig. 4). These features distinguish it also from Maorithyas humptulipsensis Hryniewicz, Amano, Jenkins, and Kiel, 2017, which is the only other thyasirid known from the Humptulips Formation (Hryniewicz et al. 2017a). Five specimens of an unidentified vesicomyid (Fig. 5) were found, which show a typical vesicomyid hinge dentition but lack details of muscle scars and the pallial line. This makes them difficult to assign to any genus among the vesicomyids and also make their relation to the only other vesicomyid known from the Humptulips Formation—“Archivesica” cf. tschudi (Olsson, 1931) as shown in Amano and Kiel (2007) uncertain.

Fig. 4. The thyasirid bivalve Thyasira (Thyasira) sp. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185006; posterodorsal view of right valve hinge and umbo with two prominent ridges (A1); lateral view (A2); close-up on posterodorsal ridges (A3). B. NRM Mo 185007 with edentulous hinge (B1); close-up on prodissoconch (B2).

Fig. 5. Unidentified vesicomyid bivalve from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185002; left valve of young specimen showing hinge disrupted by a crack (A1); close-up on hinge and dentition (A2). B. NRM Mo 185005; right valve of young specimen showing hinge and ligament nymph (B1); close-up on hinge and dentition (B2).
Gastropods: Among the gastropods recovered were two different limpet species, the peltospirid Depressigyra sp., Thalassonerita eocenica, buccinid neogastropods and the hyalogyrinid Hyalogyrina sp. Gastropod limpet 1 (Fig. 6A, B) is low, has a rounded-rectangular to oval outline, the apex is in a subcentral position and the sculpture appears to consist of indistinct concentric growth lines only. Most similar in outline and sculpture are Cocculina rathbuni and C. ovata illustrated by McLean (1987), and C. messingi McLean and Harasewych, 1995, and this species might hence belong to the Cocculiniformia. Gastropod limpet 2 (Fig. 6C) has a tall, slender, elongate diamond-shaped shell with the apex displaced slightly to the anterior. The most similar extant species all belong to the pseudococculinid genus Tentaoculus (see McLean and Harasewych 1995 and Marshall 1986), but also Pyropelta elongata from the South China Sea (Zhang and Zhang 2017) is somewhat similar in shape. Both Tentaoculus and Pyropelta are members of the vetigastropod superfamily Lepetelloidea (see Kano et al. 2016), and therefore “Limpet 2” might belong here. Five poorly preserved specimens having a discoidal shell of two almost smooth whorls and a circular aperture are tentatively assigned to Depressigyra (Fig. 7). The Eocene to Oligocene Depressigyra? statura Goedert and Benham, 1999 from seep deposits in Washington State has been re-assigned to the genus Retiskenea Warén and Bouchet, 2001 based on protoconch morphology (Kiel 2006), and specimens reported as Depressigyra? sp. from Oligocene seep deposits in Washington State have a higher spire than the specimens reported here. The preservation of the 25 available specimens of Thalassonerita eocenica Squires and Goedert, 1996a (Neritimorpha; Fig. 8) is not ideal, but the specimens show several features characteristic for neritimorph gastropods, including the protoconch, an excavated shell interior, and also the shape of the aperture is typical for neritoids in general, and for Thalassonerita and the extant Thalassonerita naticoidea (Clarke, 1989) in particular. Unlike Thalassonerita naticoidea, Thalassonerita eocenica has spiral sculpture on the entire shell. Two specimens tentatively assigned to the buccinid neogastropod Colus (Fig. 9) are sculptured by blunt axial ribs that are strongest on the whorls’ shoulder, crossed by broad and almost flat spiral cords with narrow interspaces. With this sculpture, the specimens resemble various species of the buccinid Colus, for example C. kroeyeri and C. latericeus from the deep North Atlantic Ocean have similar axial and spiral sculpture (Bouchet and Warén 1985). Its sculpture is also very similar to that of Colus sekiuensis Kiel and Goedert, 2007 from late Eocene to Oligocene from seeps and wood falls in western Washington State, USA (Kiel and Goedert 2007). The specimens are difficult to compare, though, because the known specimens of Colus sekiuensis are moderately sized adults whereas we have only small juveniles available. Lastly, two specimens of the heterobranch Hyalogyrina sp. (Fig 10) are almost indistinguishable from shells of the extant H. globularis Warén and Bouchet, 2001 from vents of the Juan de Fuca Ridge, and from H. amphorae Warén, Carozza, and Rocchini 1996 from the Mediterranean Sea. The extant H. umbellifera Warén and Bouchet, 2001 from the Aleutian Trench is higher spired, and most other Hyalogyrina species, including the type species H. glabra Marshall, 1988 from sunken wood around New Zealand, are lower spired. A Hyalogyrina sp. previously reported from the Humptulips Formation (from LACMIP loc. 12385) has a higher whorl expansion rate than the present specimens (Kiel 2006: figs: 11-1, 11-2). In contrast to most Hyalogyrina species, the present one seems to lack the reticulate pattern on the initial part of the protoconch, but this might be a preservational issue.

Fig. 6. Gastropod limpets from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A, B. Limpet 1, possibly belonging to the Cocculiniformia. A. NRM Mo 185019 slightly oval specimen in apical (A1) and oblique (A2) views. B. NRM Mo 185020 more rectangular specimen in apical (B1) and oblique (B2) views; close-up of sculpture (B3). C. NRM Mo 185021, limpet 2 in lateral (C1) and apical (C2) views.

Fig. 7. The neomphalin gastropod Depressigyra sp. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185013; specimen with 2.5 whorls in apical (A1) and oblique (A2) views; close-up on protoconch (A3); sculpture at the aperture (A4). B. NRM Mo 185014, specimen with 2.25 whorls in apical view (B1) and apertural view, showing its circular aperture (B2), and close-up on protoconch (B3).

Fig. 8. The neritimorph gastropod Thalassonerita eocenica Squires and Goedert, 1996a from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185015, specimen with nicely preserved aperture. B. NRM Mo 185016, juvenile specimen; apertural view (B1); apical view showing spiral sculpture and outline of protoconch (B2); side view showing spiral sculpture (B3). C. NRM Mo 185017, moderately sized specimen; oblique view showing dissolved interior of shell (C1); apical view showing spiral and reticulate sculpture (C2), note that the distinct, fine ribbing near the aperture is most likely the result of dissolution and not sculpture; close-up of protoconch (C3). D. NRM Mo 185018, juvenile specimen in which the protoconch is preserved as internal mold, showing that its internal walls were dissolved; note also reticulate sculpture.

Fig. 9. The buccinid neogastropod Colus? sp. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185029, small specimen with well-preserved aperture and siphonal canal. B. NRM Mo 185030, larger specimen.

Fig. 10. The heterobranch gastropod Hyalogyrina sp. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185031; oblique (A1) and apical (A2) views; close-up of protoconch (A3). B. NRM Mo 185032; apertural (B1) and apical (B2) views; close-up of protoconch (B3).
Other taxa: These include the inarticulate brachiopod Discinisca sp. (Fig. 11A) showing concentric growth rings and a very fine reticulate pattern. The geographically and stratigraphically closest report is a “Discinisca sp. with fine radial sculpture” reported from the late middle Eocene Tejon Formation in southern California (Nilsen 1987: 90). Other similar Eocene species are Discinisca insularis from the Eocene London Clay (Muir-Wood 1939) and Discinisca sp. from the late Paleocene–early Eocene of New Zealand (Lee 1987). Numerous fragments of curved or coiled worm tubes often bearing trumpet-like flanges and having a maximum diameter of 5 mm, most likely belong to the serpulids (Fig. 11D). Serpulid tubes are not uncommon at fossil seeps, where the worms were either free-living or attached to various hard substrates (Vinn et al. 2013). Two echinoid spines possibly belonging to the Atelostomata, one smooth, one with regular, low, blunt spines, were found (Fig. 11E), and a few fragmentary scaphopods (Fig. 11F).

Fig. 11. Invertebrate fossils from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. The inarticulate brachiopod Discinisca sp. (NRM Br 149701); view on upper surface (A1) and close-up of sculpture (A2). B–D. Probable serpulid tubes; specimens with flanges (B, NRM An 3073; C, NRM An 3074); a partially coiled specimen lacking flanges (D, NRM An 3075). E. Spine possibly belonging to an atelostomatan echinoid (NRM Ec 34702), complete specimen (E1) and detail of sculpture (E2). F. Scaphopod with fine concentric ribs (NRM Mo 185038).
Systematic paleontology
Class Bivalvia Linneaus, 1758
Order Nuculoida Dall, 1889
Superfamily Nuculanoidea Adams and Adams, 1858
Genus Nuculana Link, 1807
Type species: Nuculana pernula (Müller, 1779); North Sea, Recent.
Nuculana acutilineata sp. nov.
Fig. 12.
Etymology: From Latin acuti, sharp and lineae, lines; in reference to the fine increments on the outer shell.
Type material: Holotype: NRM Mo 185009 consisting of a right valve with interior features. Paratypes: NRM Mo 185008, left valve; NRM Mo 185010, young articulated specimen; NRM Mo 185011, adult articulated specimen; NRM Mo 185012, right valve.
Type locality: The Satsop Weatherwax seep deposit, Washington State, USA.
Type horizon: Basal Humptulips Formation, middle Eocene.
Material.—More than 300 specimens at NRM, including type material, mostly single valves along with numerous shell fragments. Five specimens are deposited in the Burke Museum (UWBM 108856–60).
Diagnosis.—Elongated, oval and equivalve shell with slightly oval and wrinkled prodissoconch.
Description.—Prodissoconch slightly oval; sculptured by up to seven commarginal wrinkles; transition to dissoconch marked by thick prominent rib; single valve dimensions up to length 6 mm, height 2 mm and width 1.2 mm. Elongated oval dissoconch, equivalve; more than 25 fine commarginal growth lines; pointed beaks of fair size; umbo anterior (at 25% of total length); angular hinge line with taxodont teeth, at least 12 anterior and at least 18 posterior teeth found in three specimens (see Fig. 12B); anterior muscle scar pear-shaped, pallial line parallel to dorsal margin (see Fig. 12E).
Remarks.—Nuculana acutilineata does not have the elongated posterior end seen in the type taxon Nuculana pernula (Pliocene of North America and Japan to Recent). Nuculana sp. as described by Goedert and Campbell (1995) has thicker and fewer commarginal ribs on the shell than N. acutilineata. Both Nuculana elenensis (Sowerby, 1833) (Pleistocene of North America to Recent) and Nuculana taphira Dall, 1896 (Oligocene of northwestern North America to Recent) have fine and many commarginal growth lines similar to N. acutilineata, however, N. acutilineata has a more rounded posterior end and straighter anterodorsal margin than N. elenensis, and the posterior end on N. taphira is narrower than on N. acutilineata. The growth lines on Nuculana minuta Müller, 1776 (Pliocene of North America to Recent) are fine and about the same number as on N. acutilineata, but the prominent, pointy prodissoconch is different as well as the wide, marginal ridge defining the escutcheon on N. minuta. N. acutilineata does resemble the Recent Nuculana grasslei Allen 1993, possibly also associated with cold-seep carbonates of the Lincoln Creek Formation (Peckmann et al. 2002; Kiel 2006) which is characterized by a large, ornamented prodissoconch. Nuculana grasslei was re-assigned to “Tindariopsis” grasslei by Coan and Valentich-Scott (2012). The prodissoconch can be seen in our specimens with fairly strong wrinkles; however N. acutilineata has over 25 commarginal growth lines at a narrower distance than at least 20 growth lines on T. grasslei.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 12. The nuculanid bivalve Nuculana acutilineata sp. nov. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185010, almost complete young specimen with initial part intact (A1); close-up on initial part (A2). B. NRM Mo 185008, left valve in lateral view. C. NRM Mo 185009 (holotype), right valve in lateral view showing interior features. D. NRM Mo 185012, right valve specimen showing interior with hinge, faint muscle scar and pallial line. E. NRM Mo 185011; almost complete adult specimen (E1); close-up of escutcheon and dentition (E2).
Class Gastropoda Cuvier, 1797
Subclass Caenogastropoda Cox, 1959
Superfamily Abyssochrysoidea, Tomlin 1927
Family Provannidae Warén and Ponder, 1991
Remarks.—Two new species described herein have regularly coiled shells with a high spire and rounded aperture, the shell structure is sculptured with axial ribs and spiral cords. Furthermore they have a tall protoconch with sharp axial ribs and fine spirals, which places them within the family Provannidae (Kaim et al. 2008).
Genus Desbruyeresia Warén and Bouchet, 1993
Type species: Desbruyeresia spinosa Warén and Bouchet, 1993; North Fiji Basin, Recent.
Desbruyeresia belliatus sp. nov.
Fig. 13.
Etymology: From Latin belliatus, plump; referring to the appearance of the shell.
Type material: Holotype: NRM Mo 185025 consisting of an almost fully intact shell with distinct sculpture. Paratypes: NRM Mo 185022–24; NRM Mo 185026, four specimens with distinct shell features.
Type locality: The Satsop Weatherwax seep deposit, Washington State, USA.
Type horizon: Basal Humptulips Formation, middle Eocene.
Material.—Type material and approximately 230 unnumbered specimens at NRM from the type locality. Three specimens are deposited in the Burke Museum (UWBM 108851–53).
Dimensions.—Holotype: 3.78 mm high and 1.54 mm wide.
Diagnosis.—Thick and sturdy high-spired shell with rounded whorls, strong axial sculpture forming nodes at axial-spiral intersections, apical angle approximately 45° reaching at least 5 mm in height and 3 mm in width.
Description.—Protoconch: up to 4 whorls; protoconch is decollated, silicified calcareous plug filling topmost whorl; maximum diameter 0.5 mm, minimum height 0.5 mm, sculptured by reticulate pattern, ~30 prominent, slightly ophistocline axial ribs, 14 weaker spiral ribs across whorl; aperture not thickened, teleoconch transition visible on fourth whorl. Teleoconch: up to four whorls; sculptured by reticulate pattern, ~12 slightly opisthocline axial ribs, at least four spiral ribs, prominent nodes in oblique angle toward apex at intersections; adapical row of nodes on each whorl strong, successively weaker towards base of whorl; whorls slightly convex, incised sutures; basal margin marked by thick spiral cord; base has up to 5 spiral cords, downwards decreasing in strength; aperture oval with short, rounded siphonal notch; columella smooth; thin parietal callus.
Remarks.—Distinguishable from Provanna based on its more slender shell (Warén and Bouchet 1993) and the shell has “axial ribs, spiral cords, knobs and occasionally short spines” (Desbruyères et al. 2006), features that most of which are present in the species from Satsop Weatherwax, making it a likely member of Desbruyeresia. Type species Desbruyeresia spinosa Warén and Bouchet, 1993 is characterized by prominent spines forming at the crossings of the axial and spiral ribs on the whorls of the teleoconch, which can also be seen in Desbruyeresia cancellata Warén and Bouchet, 1993. No spines can be seen on D. belliatus, instead the shell structure more resembles that of Desbruyeresia melanioides Warén and Bouchet, 1993 with small nodes forming at the intersections of the ribs. D. melanioides have slightly curved axial ribs, but not as strongly ophistocline as seen on D. belliatus. The whorls of Desbruyeresia marianaensis have six or more spinal cords and there are no prominent nodes at the intersections of the apical and spinal cords as on D. belliatus. Desbruyeresia antiqua Bandel and Kiel, 2000 from a Late Cretaceous slump deposit in Spain is quite different from Desbruyeresia belliatus by having sharp axial ribs with a single, central row of spines. The species is based on a single specimen and its placement in Desbruyeresia has been questioned (Kaim et al. 2008), but without further material, these doubts can neither be confirmed nor rejected. The reticulate pattern on Desbruyeresia chamorrensis Chen, Ogura, and Okutani, 2016 is made up by three spiral cords whereas in D. belliatus there are at least four and D. belliatus is generally smaller. The reticulate pattern on the protoconch and the pointed nodes on the spire of D. belliatus closely resembles that of Desbruyeresia kanajirisawensis Kaim, Jenkins, and Warén, 2008 from the Cretaceous of Japan (Kaim et al. 2008) and the calcareous plug described in D. kanajirisawensis is also present in D. belliatus, however D. kanajirisawensis displays six nodes making up a tight reticulate pattern whereas D. belliatus is of larger size and only has five nodes making up a wider pattern.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 13. The provannid gastropod Desbruyeresia belliatus sp. nov. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185022; almost complete specimen showing protoconch with 3 whorls (A1); close-up on protoconch (A2); spire with 2.5 whorls and the spire base with short siphonal notch, arrows pointing to spiral cords (A3). B. NRM Mo 185023; protoconch with 3.5 whorls showing the protoconch-teleoconch transition (B1); close-up of protoconch sculpture (B2); close-up of protoconch-teleoconch transition (arrow) (B3). C. NRM Mo 185024; decollated protoconch and calcareous plug (C1); close-up on protoconch (C2). D. NRM Mo 185025 (holotype) showing protoconch with 3 whorls and teleoconch with 3.5 whorls and an oval aperture. E. NRM Mo 185026, Close-up of specimen with protoconch-teleoconch transition (arrow).
Genus Provanna Dall, 1918
Type species: Trichotropis (Provanna) lomana Dall, 1918; Magdalena Bay, USA, Recent.
Provanna fortis sp. nov.
Fig. 14.
Etymology: From Latin fortis, reinforced; referring to the appearance of the shell.
Type material: Holotype: NRM Mo 185028 with one intact whorl of protoconch and distinct nodes on the teleoconch. Paratype: NRM 185027 with distinct spiral cords on the base and aperture with siphonal notch.
Type locality: Satsop Weatherwax seep deposit, Washington State, USA.
Type horizon: Basal Humptulips Formation, middle Eocene.
Material.—Type material and 5 other specimens. Two specimens deposited in the Burke Museum (UWBM 108854–55).
Dimensions.—Holotype is approximately 2.3 mm in height and 1.7 mm in width.
Diagnosis.—Sturdy, high-spired shell with prominent, angular whorls with spiny nodes at intersection of the axial and spiral ribs, apical angle of approximately 45° reaching at least 2.3 mm in height and 1.7 mm in width.
Description.—Protoconch: at least one whorl; maximum diameter 0.5 mm; sculptured by reticulate pattern, ~24 prominent, slightly sinuous axial ribs, ~16 fine spiral cords; transition to teleoconch unknown. Teleoconch: up to three whorls, sculptured by reticulate pattern, 14 orthocline axial ribs, two prominent and equally strong spiral ribs, blunt, short spines at intersections; spines more prominent on lower whorls; axial ribs start at the upper suture and fade below the lower spiral rib; irregularly spaced, fine spiral treads on whorl flank, fine axial growth increments; basal margin marked by distinct spiral rib, without spines; two spiral ribs of decreasing strength on base of the whorl; aperture broadly oval; short siphonal notch bordered by low ridge; columella smooth, thin callus on inner lip.
Remarks.—The strong angulation and spiny appearance of Provanna fortis can also be seen in other provannids such as for example Provanna ios Warén and Bouchet, 1986 and Provanna muricata Warén and Bouchet, 1986 from the East Pacific region (Warén and Bouchet 1986). Where the first one has one distinct spiral keel, P. muricata has developed two ribs on the spire which can also be seen in the P. fortis sp. nov. The protoconch of P. muricata was not recovered from the specimen described by Warén and Bouchet (1986) which aggravates a distinction between these two species. Provanna antiqua Squires, 1995 from Eocene and Oligocene seep deposits in western Washington shows a reticulate pattern but lacks spines and has rounded whorls instead of the prominent angulation seen on P. fortis. Provanna alexi Amano and Little, 2014 and Provanna hirokoae Amano and Little, 2014 from the middle Miocene of Japan (Amano and Little 2014) both lack the strong angulation seen on P. fortis. Provanna marshalli Saether, Little, and Campbell, 2010 from early to middle Miocene of New Zealand is similar to P. antiqua, also lacks strong angulation and has more spiral ribs on the whorls than P. fortis. Provanna urahoroensis Amano and Jenkins, 2013 reported from Oligocene deposits in Japan does not have the distinct reticulate pattern or anything resembling the spines on P. fortis. Provanna nakagawensis Kaim, Jenkins, and Hikida, 2009 from the Upper Cretaceous of Japan is sculptured by a distinct reticulate pattern, lacks the spines and does not have the shelf-structure made up by strong angulation as seen on the lowermost whorl of P. fortis. Kaim et al. (2008) also report Provanna tappuensis Kaim, Jenkins, and Warén, 2008 from the Cretaceous of Japan, which shows strong axial and spiral sculpture, although the spiral base has an increasing number of spiral ribs; up to five, beneath the spiral base where the P. fortis only has up to three spiral ribs in adult specimens and is therefore described as a new species.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 14. The provannid gastropod Provanna fortis sp. nov. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185027, teleoconch with 2.5 whorls (A1); basal view showing spiral cords and aperture with siphonal notch (A2). B. NRM Mo 185028 (holotype); part of protoconch and teleoconch with 3 strongly angular whorls with spines (B2); close-up on protoconch (B1); spiral suture and fading axial rib (B3); growth increments (B4).
Subclass Heterobranchia Burmeister 1837
Superfamily Orbitestelloidea Iredale, 1917
Family Orbitestellidae Iredale, 1917
Genus Orbitestella Iredale, 1917
Type species: Cyclostrema (Orbitestella) bastowi Gatliff, 1906; Western Port Bay, Australia, Recent.
Orbitestella dioi sp. nov.
Fig. 15.
Etymology: Named after a musician Ronnie James Dio (1942–2010).
Type material: Holotype: NRM Mo 185034 with two mostly intact whorls with defined sculpture. Paratype: NRM Mo 185033 with elevated spire and intact protoconch.
Type locality: The Satsop Weatherwax seep deposit, Washington State, USA.
Type horizon: Basal Humptulips Formation, middle Eocene.
Material.—Type material only.
Dimensions.—Holotype: 0.75 mm in height and 1.5 mm in width.
Diagnosis.—Discoidal shell with wide umbilicus, a slightly elevated spire, a rounded-pentagonal whorl profile with an apical angle of 105°, with dimensions at least 0.75 mm in height and a diameter of 1.5 mm.
Description.—Protoconch diameter approximately 0.2 mm, terminates in thin varix, probably smooth. Teleoconch has three whorls with ~25 prominent, evenly spaced axial ribs, at least 12 weaker spiral ribs in reticulate pattern on spire; sinuous growth lines, ophistocline on shoulder, prosocyrt on periphery. Umbilicus deep, width around 1/3 of shell diameter, sculptured with strong axial ribs and finer spiral ribs; rounded aperture with sinuous peristome.
Remarks.—Shares the general shell sculpture and size dimensions with the Recent Orbitestella patagonica Simone and Zeyala, 2004 but has fewer axial ribs and less prominent spiral ribs. In O. patagonica the spiral ribs are more prominent than the axial ribs, which is the opposite from O. dioi sp. nov. A similar reticulate pattern on the outer surface of the shell is also found on the Recent Orbitestella bermudezi Aguayo and Borro, 1946 along with a sinuous peristome and rounded aperture, however in O. bermudezi the intersections between the axial ribs and spiral cords form pointed nodules and the aperture is subtriangular, which differs from O. dioi sp. nov. The prominent vertical ribs on the whorls of Orbitestella granulata Lozouet, 1998, from the Oligocene of France (Lozouet 1998) are coarser than on O. dioi and the former is also missing a reticulate ornamentation. Orbitestella palaiopacifica Squires and Goedert, 1996b from the Eocene of the Crescent Formation, North America lacks the spiral ribs on the whorls as well as an elevated spire. The organization of the growth lines on O. dioi follows the same pattern as the Recent Lurifax goederti (Kiel, 2006) but lacks the strong angulation and prominent spiral cords. Lurifax was placed within the family Orbitestellidae (Warén and Bouchet, 2001). Recently Lauridsen et al. (2014) reported an orbitestellid from the Faxe Formation, Denmark that is larger than our specimens and with a distinctively larger protoconch. O. dioi shares many features with the orbitestellids compared above, however, it does not fit to any already described species and therefore we describe this new species.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 15. The orbitestellid gastropod Orbitestella dioi sp. nov. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA. A. NRM Mo 185033; specimen with 2.25 whorls in apical view (A1); apertural view showing a rounded aperture (A2); close-up on protoconch, arrow indicates protoconch-teleoconch transition (A3). B. NRM Mo 185034 (holotype) with roughly 2 whorls in apical (B1) and oblique (B2) views; close-up on protoconch (B3).
Class Polyplacophora Gray, 1821
Order Lepidopleurida Thiele, 1909
Family Leptochitonidae Dall, 1889
Genus Leptochiton Gray, 1847
Type species: Leptochiton asellus (Gmelin, 1791); Telemark, Norway, Recent.
Leptochiton terryiverseni sp. nov.
Fig. 16.
Etymology: For Terry Iversen, who helped in collecting this material.
Type material: Holotype: NRM Mo 185036 consisting of three articulated intermediate valves. Paratypes: NRM Mo 185035, NRM Mo 185037, UWBM 108862, three isolated intermediate valves.
Type locality: The Satsop Weatherwax seep deposit, Washington State, USA.
Type horizon: Basal Humptulips Formation, middle Eocene.
Material.—The type material only.
Dimensions.—The largest specimen is 6.7 mm wide and 3 mm long.
Diagnosis.—Leptochiton having moderately elevated, round-backed valves with angulation, lacking jugal area of the tegmentum; pleural area with fine, subparallel rows of granules; lateral areas with irregular but distinct commarginal ridges.
Description.—Elongate-oval outline, valves of moderate elevation (h/w ~0.33), round-backed with an angulation, side slopes slightly convex; jugal area of tegmentum absent, pleural area sculptured by fine granules arranged in subparallel, longitudinal rows; sculpture changes at abruptly at diagonal ridges, granules on lateral areas stronger, arranged somewhat irregularly near the middle, but forming distinct commarginal ridges toward the lateral margins. The apophyses are separated by a broad jugal sinus.
Remarks.—Squires and Goedert (1995) reported the extant Leptochiton alveolus (Lovén, 1846) from Eocene–Oligocene seep deposits in western Washington. Their illustrated specimens are from the Oligocene Lincoln Creek Formation and show a quincunx arrangement of granules on the tegmentum, which is different from the longitudinal arrangement of granules in L. terryiverseni n. sp. Also mentioned by Squires and Goedert (1995) were two valves of L. alveolus from seep deposit LACMIP loc. 12385 in the Humptulips Formation, but they did not illustrate them. Hence it remains unclear whether they belong to the species from the Lincoln Creek Formation or to L. terryiverseni. Extant species similar to L. terryiverseni include L. micropustulosus Kaas, 1984 from ca. 1135 to 1236 m depth on the Barbados Prism (Kaas 1984) and Leptochiton sarsi Kaas, 1981 from Scandinavian waters (Kaas 1981; Dell’Angelo et al. 2009), both of which differ from L. terryiverseni by the granulae on the tegmentum gently curving around the diagonal ridges, in contrast to the sharp change in granulae arrangement in L. terryiverseni. Leptochiton tenuidontus Saito and Okutani, 1990 from a hydrothermal vent site in 1395 m depth in the Okinawa Trough (Saito and Okutani 1990) shows a similar arrangement of the granulae, but its valves are less elevated than those of L. terryiverseni.
Among fossil species of Leptochiton, an Eocene specimen from Seymour Island, Antarctic Peninsula (López Cabrera and Olivero 2011), has the longitudinally arranged granule distributed over a much broader area on the tegmentum than in L. terryiverseni, and laterally the granulae tend to radiate outward. In the late Eocene Leptochiton sp. reported from the Lincoln Creek Formation in western Washington (Dell’Angelo et al. 2011), the granulae on the anterior-lateral sides tend to radiate outward rather than being longitudinally arranged, as in L. terryiverseni. Recently, that specimen was hesitantly assigned to the extant L. cascadiensis (Sigwart and Chen 2017). The wood-associated Leptochiton lignatilis Bertolaso, Garilli, Parrinello, Sosso, and Dell’Angelo, 2015 from the middle to late Miocene of northern Italy has irregularly arranged granulae (Bertolaso et al. 2015) in contrast to the longitudinal arrangement in L. terryiverseni. Early Oligocene specimens from France assigned to Leptochiton cf. algesirensis (Capellini, 1859) have outwardly directed beaded riblets rather than longitudinally arranged granules (Cherns and Schwabe 2017). Finally, the Paleocene Leptochiton faxensis Sigwart, Andersen, and Schnetler 2007 has a quincunx arrangement of granules.
Stratigraphic and geographic range.—Type locality and horizon only.
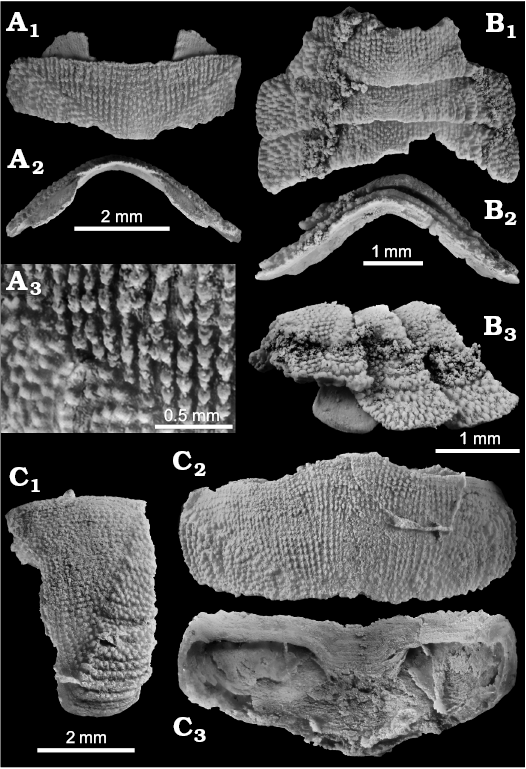
Fig. 16. The lepidopleurid polyplacophoran Leptochiton terryiverseni sp. nov. from the middle Eocene Satsop Weatherwax seep deposit, Washington State, USA; all specimens represented by intermediate valves. A. NRM Mo 185035 (paratype); isolated valve with preserved apophyses in dorsal (A1) and anterior (A2) views; close-up of sculpture (A3). B. NRM Mo 185036 (holotype), three articulated valves in dorsal (B1), posterior (B2), and lateral (B3) views. C. NRM Mo 185037 (paratype); isolated valve showing the broad jugal sinus in lateral (C1), dorsal (C2), and ventral (C3) views.
Discussion
Petrography and stable isotopes.—The petrography and stable isotope signature of the Satsop River seep deposit is similar to that of other seep deposits in western Washington (Peckmann et al. 2002; Goedert et al. 2003; Kuechler et al. 2012). The negative carbon isotope signature (with values as low as -43.5‰) clearly indicate that carbonate was related to the oxidation of biogenic methane (Peckmann and Thiel 2004). Also the silicification of the fossils is most likely related to methane oxidation: the anaerobic oxidation (AOM) of methane causes an increase in alkalinity, thereby inducing the dissolution of silica skeletons; once AOM ceases the pH drops again, leading to dissolution of aragonite minerals (including shells) on the one hand, and to silica precipitation on the other (Kuechler et al. 2012; Smrzka et al. 2015; Miyajima et al. 2016). Virtually all silicified fossils reported here are mollusks that originally had an aragonitic shell (Carter 1990; Vinn et al. 2008); fossils of taxa with calcitic shells, such as ostracods, are notably absent.
Fauna.—Most mollusk species from the Satsop Weatherwax seep deposit belong to Recent genera, highlighting the modern character of this ca. 40–42.5 million year old seep fauna. Furthermore, although not assignable to any known genus, the Satsop Weatherwax seep deposit contains vesicomyid bivalves, a clade that is unknown from seeps of older age, including the two known Paleocene examples (Schwartz et al. 2003; Hryniewicz et al. 2016). There are nine described fossil species of Provannidae and they are mostly reported from hydrocarbon seep deposits, with a few exceptions of reports from other chemosynthetic environments such as wood falls or whale falls. Provannid gastropods have not been reported from the Humptulips Formation before. They precede Provanna antiqua Squires, 1995 from late Eocene Bear River deposit and are therefore the two geochronologically oldest provannid gastropods associated with deep-sea methane seeps in North America. Only six specimens of P. fortis were recovered, but all specimens show distinctive characters such as the prominent angulation of the spire and spiny whorls. In contrast, the more than 230 specimens of D. belliatus show rounded, spineless whorls and a protoconch with a calcareous plug as distinguishing features. This gives enough support to the notion that these fossils represent two new species and further extends the fossil record of the family Provannidae. Orbitestella dioi differs from the previously described Orbitestella palaiopacifica Squires and Goedert, 1996b from the Eocene of western Washington, because it has an elevated spire and spiral ribs crossing the axial ribs in a reticulate pattern. It is the first representative of the genus Orbitestella reported from the Humptulips Formation and Recent relatives have not been reported in association with present day vents and seeps. Our new material of Thalassonerita eocenica confirms that this species is a neritoid and does not belong to the vetigastropod genus Sahlingia, as suggested earlier (Warén and Bouchet 2001). This indicates that Phenacolepidae, the family which includes the extant Thalassonerita naticoidea, colonized deep-water habitats at latest in the middle Eocene (Kano et al. 2002). Their phylogenetic relationships to geologically older neritids from Cretaceous and Jurassic methane seeps (Kiel and Peckmann 2008; Kiel et al. 2010) remains unclear due to the poor preservation of those Mesozoic examples.
Whereas several gastropod and bivalve groups, including thyasirids, colloniids, and provannids, colonize both seeps and wood falls (Warén and Bouchet 2001; Gaudron et al. 2010) and are known to have been doing so since the Late Cretaceous (Kaim et al. 2008, 2014; Kiel et al. 2009), this seems to be different among polyplacophorans. Chitons have been reported from Middle Jurassic wood-fall sites (Kaim 2011; Sirenko 2013) and even a Carboniferous origin for the colonization of wood falls has been suggested (Sirenko 2004). But although Cretaceous seeps are increasingly well sampled (Campbell and Bottjer 1993, Campbell et al. 2002; Kaim et al. 2009, 2013; Kiel et al. 2013, 2017; Agirrezabala et al. 2013; Little et al. 2015; Hryniewicz et al. 2015; Meehan and Landman 2016), there is not a single report of seep-inhabiting chitons from this period. Whether this (still) reflects a sampling bias or if the adaptation of lepidopleuran chitons to seeps is a post-Paleocene affair, as in many other invertebrate groups (Vrijenhoek 2013; Kiel 2015), remains to be tested.
Conclusions
Overall the Satsop Weatherwax seep fauna has a modern character. But compared to the well-known late Eocene to Oligocene seep faunas in Washington, most taxa of the middle Eocene Satsop Weatherwax seep deposit, and the Humptulips Formation in general, are different species. Examples include the two new provannids Desbruyeresia belliatus and Provanna fortis, the new Leptochiton terryiverseni, and Thalassonerita eocenica, all of which are not known from the younger seeps. Previously published examples include the bathymodiolin Vulcanidas goederti Kiel and Amano, 2013 and Maorithyas humptulipsensis Hryniewicz, Amano, Jenkins, and Kiel, 2017, both from seep deposits in the Humptulips Formation. In these cases, not even the genera are known from the younger seep sites in western Washington, only members of their respective families and subfamilies (Kiel and Amano 2013; Hryniewicz et al. 2017a).
Acknowledgments
First of all we sincerely thank James L. Goedert (Wauna, USA) and Terry Iversen (Bremerton, USA) for collecting this material and making it available to us. We greatly appreciate the continued cooperation of Green Diamond Resources Company and staff (Shelton, USA) for allowing access to the locality on their timberlands. We thank Boris Sirenko (Russian Academy of Sciences, St. Petersburg, Russia) and Bruno Dell’Angelo (Genova, Italy) for advice on Leptochiton, Frank Wiese (University of Göttingen, Germany) for help with echinoid spine identification, and Anders Warén (Swedish Museum of Natural History, Stockholm, Sweden) for discussions on gastropods. We also thank the reviewers Kazutaka Amano (Joetsu University of Education, Japan) and Krzysztof Hryniewicz (Institute of Paleobiology PAS, Warsaw, Poland) for their constructive input to improve the manuscript. Financial support was provided by Vetenskapsrådet through grant 2016-03920 to SK.
References
Adams, H. and Adams, A. 1853–58. The Genera of Recent Mollusca Arranged According to Their Organization. 1: vi–xl + 484 pp.; 2: 661 pp. John Van Voorst, London.
Agirrezabala, L.M., Kiel, S., Blumenberg, M., Schäfer, N., and Reitner, J. 2013. Outcrop analogues of pockmarks and associated methane-seep carbonates: a case study from Lower Cretaceous (Albian) of the Basque-Cantabrian Basin, western Pyrenees. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 94–115. Crossref
Aguayo, C.G. and Borro, P. 1946. Nuevos moluscos del terciario superior de Cuba. Revista de la Sociedad Malacologica “Carlos de la Torre” 4: 9–12.
Allen, J.A. 1993. A new deep-water hydrothermal species of Nuculana (Bivalvia: Protobranchia) from the Guaymas Basin. Malacologica 35: 141–151.
Amano, K. and Jenkins, R.G. 2013. A new species of Provanna (Gastropoda: Provannidae) from an Oligocene seep deposit in eastern Hokkaido, Japan. Paleontological Research 17: 325–329. Crossref
Amano, K. and Kiel, S. 2007. Fossil vesicomyid bivalves from the North Pacific region. The Veliger 49: 270–293.
Amano, K. and Little, C.T.S. 2014. Miocene abyssochrysoid gastropod Provanna from Japanese seep and whale–fall sites. Acta Palaeontologica Polonica 59: 163–172.
Bandel, K. and Kiel, S. 2000. Earliest known (Campanian) members of the Vermetidae, Provannidae and Litiopidae (Cerithioidea, Gastropoda), and a discussion of their possible relationships. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 84: 209–218.
Bertolaso, L., Garilli, V., Parrinello, D., Sosso, M., and dell’Angelo, B. 2015. A new Miocene deep-sea chiton and early evidence for Teredinidae-sustained wood-fall communities. Palaeontologica Electronica 18.2.41A: 1–15.
Bouchet, P. and Warén, A. 1985. Revision of the northeast Atlantic bathyal and abyssal Neogastropoda excluding Turridae (Mollusca, Gastropoda). Bolletino Malacologico Supplemento 1: 123–296. Crossref
Brandon, M.T. and Vance, J.A. 1992. Tectonic evolution of the Cenozoic Olympic subduction complex, Washington State, as deduced from fission track ages for detrital zircons. American Journal of Science 292: 565–636. Crossref
Burmeister, H. 1837. Handbuch der Naturgeschichte, Vol. 2, Zoologie. i–xii + 369–858 pp. Enslin, Berlin.
Campbell, K.A. and Bottjer, D.J. 1993. Fossil cold seeps. National Geographic Research & Exploration 9: 326–343.
Campbell, K.A. and Bottjer, D.J. 1995. Brachiopods and chemosymbiotic bivalves in Phanerozoic hydrothermal vent and cold seep environments. Geology 23, 321–324. Crossref
Campbell, K.A., Farmer, J.D., and Des Marais, D. 2002. Ancient hydrocarbon seeps from the Mesozoic convergent margin of California: carbonate geochemistry, fluids and palaeoenvironments. Geofluids 2: 63–94. Crossref
Capellini, J. 1859. Catalogue des oscabrions se la Méditerranée. Journal de Conchyliologie 7: 320–328.
Carter, J.G. 1990. Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends Volume I. 832 pp. Van Nostrand Reinhold, New York.
Chen, C., Ogura, T., Hirayama, H., Watanabe, H. K., Miyazaki, J., and Okutani, T. 2016. First seep-dwelling Desbruyeresia (Gastropoda: Abyssochrysoidea) species discovered from a serpentinite-hosted seep in the Southeastern Mariana Forearc. Molluscan Research 36: 277–284. Crossref
Cherns, L. and Schwabe, E. 2017. Eocene and Oligocene chitons (Polyplacophora) from the Paris and Hampshire Basins. Historical Biology [published online 10.1080/08912963.2017.1387545]. Crossref
Clarke, A.H. 1989. New mollusks from undersea oil seep sites off Louisiana. Malacology Data Net 2: 122–134.
Coan, E.V. and Valentich-Scott, P. 2012. Bivalve Seashells of Tropical West America. Marine Bivalve Mollusks from Baja California to Northern Peru. 764 pp. Santa Barbara Museum of Natural History, Santa Barbara.
Cox, L.R. 1959. Thoughts on the classification of the Gastropoda. Proceedings of the Malacological Society of London 33: 239–261.
Cuvier, G. 1797. Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la sociéte d’Histoire Naturelle de Paris, le 11 prairial an troisième. Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts, 1795 2: 433–449.
Dall, W.H. 1889. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877–78) and in the Caribbean Sea (1879–80), by the U.S. Coast Survey steamer “Blake” XXIX. Report on the Mollusca. Part 2, Gastropoda and Scaphopoda. Bulletin of the Museum of Comparative Zoology, Harvard University 18: 1–492.
Dall, W.H. 1896. Note on Leda caleata Hinds. The Nautilus 10: 70.
Dall, W.H. 1918. Description of new species of shells chiefly from Magdalena Bay, Lower California. Proceedings of the Biological Society of Washington, 31, 5–8.
Dell’Angelo, B., Bonfitto, A., and Taviani, M. 2011. Chitons (Polyplacophora) from Paleogene strata in western Washington State, U.S.A. Journal of Paleontology 85: 936–954. Crossref
Dell’Angelo, B., Giusti, F., Paolini, P., Sosso, M., and Bonfitto, A. 2009. New data on Polyplacophora from Tuscan Archipelago. I. Leptochiton sarsi Kaas, 1981 and Leptochiton pepezamorai Carmona Zalvide, Urgorri & Garcia, 2004, two species new to the Mediterranean Sea. Italian Journal of Zoology 76: 76–82. Crossref
Desbruyères, D., Segonzac, M., and Bright, M. 2006. Handbook of deep-sea hydrothermal vent fauna. Second completely revised version. Denisia 18: 1–544.
Dubilier, N., Bergin, C., and Lott, C. 2008. Symbiotic diversity in marine animals: the art of harnessing chemosynthesis. Nature Reviews Microbiology 6: 725–740. Crossref
Freytag, J.K., Girguis, P.R., Bergquist, D.C., Andras, J.P., Childress, J.J., and Fisher, C.R. 2001. A paradox resolved: sulfide acquisition by roots of seep tubeworms sustains net chemoautotrophy. Proceedings of the National Academy of Sciences 98: 13408–13413. Crossref
Gatliff, J.H. 1906. On some Victorian marine Mollusca, new species and others little-known. Proceedings of the Royal Society of Victoria, New Series 19: 1–4.
Gaudron, S.M., Pradillon, F., Pailleret, M., Duperron, S., Le Bris, N., and Gaill, F. 2010. Colonization of organic substrates deployed in deep-sea reducing habitats by symbiotic species and associated fauna. Marine Environmental Research 70: 1–12. Crossref
Gmelin, J.F. 1791. Vermes. In: J.F. Gmelin (ed.), Caroli a Linnaei Systema Naturae per Regna Tria Naturae, Ed. 13. Tome 1, Vol. 6, 3021–3910. G.E. Beer, Lipsiae.
Goedert, J.L. and Benham, S.R. 1999. A new species of Depressigyra? (Gastropoda: Peltospiridae) from cold-seep carbonates in Eocene and Oligocene rocks of western Washington. The Veliger 42: 112–116.
Goedert, J.L. and Campbell, K.A. 1995. An Early Oligocene chemosynthetic community from the Makah Formation, northwestern Olympic Peninsula, Washington. The Veliger 38: 22–29.
Goedert, J.L. and Squires, R.L. 1990. Eocene deep-sea communities in localized limestones formed by subduction-related methane seeps, southwestern Washington. Geology 18: 1182–1185. Crossref
Goedert, J.L., Peckmann, J., Benham, S.R., and Janssen, A.W. 2013. First record of the Eocene pteropod Heliconoides nitens (Gastropoda: Thecosomata: Limacinidae) from the Pacific Basin. Proceedings of the Biological Society of Washington 126: 72–82. Crossref
Goedert, J.L., Thiel, V., Schmale, O., Rau, W.W., Michaelis, W., and Peckmann, J. 2003. The late Eocene “Whiskey Creek” methane-seep deposit (western Washington State) Part I: Geology, palaeontology, and molecular geobiology. Facies 48: 223–240.
Gray, J.E. 1821. A natural arrangement of Mollusca, according to their internal structure. The London Medical Repository 15: 229–239.
Gray, J.E. 1847. A list of the genera of Recent Mollusca, their synonyma and types. Proceedings of the Zoological Record of London 15: 129–219.
Hryniewicz, K., Amano, K., Jenkins, R.G., and Kiel, S. 2017a. Thyasyrid bivalves from Cretaceous and Paleogene cold seeps. Acta Paleontologica Polonica 62: 705–728. Crossref
Hryniewicz, K., Bitner, M.A., Durska, E., Hagström, J., Hjálmarsdóttir, H.R., Jenkins, R.G, Little, C.T.S., Miyajima, Y., Nakrem, H.A., and Kaim, A. 2016. Paleocene methane seep and wood-fall marine environments from Spitsbergen, Svalbard. Palaeogeography, Palaeoclimatology, Palaeoecology 462: 41–56. Crossref
Hryniewicz, K., Hagström, J., Hammer, Ø., Kaim, A., Little, C.T.S., and Nakrem, H.A. 2015. Late Jurassic–Early Cretaceous hydrocarbon seep boulders from Novaya Zemlya and their faunas. Palaeogeography, Palaeoclimatology, Palaeoecology 436: 231–244. Crossref
Hryniewicz, K., Jakubowicz, M., Belka, Z., Dopieralska, J., and Kaim, A. 2017b. New bivalves from a Middle Devonian methane seep in Morocco: the oldest record of repetitive shell morphologies among some seep bivalve molluscs. Journal of Systematic Palaeontology 15: 19–41. Crossref
Iredale, T. 1917. More molluscan name-changes, generic and specific. Proceedings of the Malacological Society of London 12: 322–330.
Jakubowicz, M., Hryniewicz, K., and Belka, Z. 2017. Mass occurrence of seep-specific bivalves in the oldest-known cold seep metazoan community. Scientific Reports 7: 14292. Crossref
Jenkins R.G., Kaim A., Hikida Y., and Kiel S. 2018. Four new species of the Jurassic to Cretaceous seep-restricted bivalve Caspiconcha and implications for the history of chemosynthetic communities. Journal of Paleontology 92: 596–610. Crossref
Jenkins, R.G., Kaim, A., Little, C. T. S., Iba, Y., Tanabe, K., and Campbell, K.A. 2013. Worldwide distribution of modiomorphid bivalve genus Caspiconcha in late Mesozoic hydrocarbon seeps. Acta Palaeontologica Polonica 58: 357–382.
Johnson, S.B., Krylova, E.M., Audzijonyte, A., Sahling, H., and Vrijenhoek, R.C. 2017. Phylogeny and origins of chemosynthetic vesicomyid clams. Systematics and Biodiversity 15: 346–360 Crossref
Kaas, P. 1981. Scandinavian species of Leptochiton Gray, 1847 (Mollusca, Polyplacophora). Sarsia 66: 217–229. Crossref
Kaas, P. 1984. A new species of Leptochiton Gray, 1847 (Mollusca: Polyplacophora) from the South Barbados accretionary prism. Zoologische Mededelingen Leiden 68: 45–47.
Kaim, A. 2011. Non-actualistic wood-fall associations from Middle Jurassic of Poland. Lethaia 44: 109–124. Crossref
Kaim A., Hryniewicz K., Little C.T.S., and Nakrem H.A. 2017. Gastropods from the Late Jurassic–Early Cretaceous seep deposits in Spitsbergen, Svalbard. Zootaxa 4329: 351–374. Crossref
Kaim, A., Jenkins, R.G., and Hikida, Y. 2009. Gastropods from Late Cretaceous hydrocarbon seep deposits in Omagari and Yasukawa, Nakagawa area, Hokkaido, Japan. Acta Palaeontologica Polonica 54: 463–690. Crossref
Kaim, A., Jenkins, R.G., and Warén, A. 2008. Provannid and provannid-like gastropods from Late Cretaceous cold seeps of Hokkaido (Japan) and the fossil record of the Provannidae (Gastropoda: Abyssochrysoidea). Zoological Journal of the Linnean Society 154: 421–436. Crossref
Kaim, A., Jenkins, R.G., Tanabe, K., and Kiel, S. 2014. Mollusks from late Mesozoic seep deposits, chiefly in California. Zootaxa 3861: 401–440. Crossref
Kaim, A., Skupien, P., and Jenkins, R.G. 2013. A new Lower Cretaceous hydrocarbon seep locality from Czech Carpathians and its fauna. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 42–51. Crossref
Kano, Y., Chiba, S., and Kase, T. 2002. Major adaptive radiation in neritopsine gastropods estimated from 28S rRNA sequences and fossil records. Proceedings of the Royal Society B 269: 2457–2465. Crossref
Kano, Y., Takano, T., Schwabe, E., and Warén, A. 2016. Phylogenetic position and systematics of the wood-associate limpet genus Caymanabyssia and implications for ecological radiation into deep-sea organic substrates by lepetelloid gastropods. Marine Ecology 37: 1116–1130. Crossref
Kelly, S.R.A., Blanc, E., Price, S.P., and Whitham, A.G. 2000. Early Cretaceous giant bivalves from seep-related limestone mounds, Wollaston Forland, Northeast Greenland. In: E.M. Harper, J.D. Taylor, and J.A. Crame (eds.), The Evolutionary Biology of the Bivalvia, 227–246. Geological Society, London.
Kiel, S. 2006. New records and species of molluscs from Tertiary cold-Seep carbonates in Washington State, USA. Journal of Paleontology 80: 121–137. Crossref
Kiel, S. 2010. The fossil record of vent and seep mollusks. In: S. Kiel (ed.), The Vent and Seep Biota. Aspects from Microbes to Ecosystems, 255–277. Springer, Dordrecht. Crossref
Kiel, S. 2015. Did shifting seawater sulfate concentrations drive the evolution of deep-sea vent and seep ecosystems? Proceedings of the Royal Society B 282: 20142908. Crossref
Kiel, S. 2018. Three new bivalve genera from Triassic hydrocarbon seep deposits in southern Turkey. Acta Palaeontologica Polonica 63. 221–234. Crossref
Kiel, S., Amano, K., Hikida, Y., and Jenkins, R.G. 2009. Wood-fall associations from Late Cretaceous deep-water sediments of Hokkaido, Japan. Lethaia 42: 74–82. Crossref
Kiel, S. and Amano, K. 2013. The earliest bathymodiolin mussels: Evaluation of Eocene and Oligocene taxa from deep-sea methane seep deposits in western Washington State, USA. Journal of Paleontology 87: 589–602. Crossref
Kiel, S. and Goedert, J.L. 2006. Deep-sea food bonanzas: Early Cenozoic whale-fall communities resemble wood-fall rather than seep communities. Proceedings of the Royal Society B 273: 2625–2631. Crossref
Kiel, S. and Goedert, J.L. 2007. Six new mollusk species associated with biogenic substrates in Cenozoic deep-water sediments in Washington State, USA. Acta Palaeontologica Polonica 52: 41–52.
Kiel, S. and Peckmann, J. 2008. Paleoecology and evolutionary significance of an Early Cretaceous Peregrinella-dominated hydrocarbon-seep deposit on the Crimean Peninsula. Palaios 23: 751–759. Crossref
Kiel, S., Birgel, D., Campbell, K.A., Crampton, J.S., Schiøler, P., and Peckmann, J. 2013. Cretaceous methane-seep deposits from New Zealand and their fauna. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 17–34. Crossref
Kiel, S., Campbell, K.A., and Gaillard, C. 2010. New and little known mollusks from ancient chemosynthetic environments. Zootaxa 2390: 26–48. Crossref
Kiel, S., Krystyn, L., Demirtaş, F., Koşun, E., and Peckmann, J. 2017. Late Triassic mollusk-dominated hydrocarbon-seep deposits from Turkey. Geology 45: 751–754. Crossref
Kuechler, R.R., Birgel, D., Kiel, S., Freiwald, A., Goedert, J.L., Thiel, V., and Peckmann, J. 2012. Miocene methane-derived carbonates from southwestern Washington, USA, and a model for silicification at seeps. Lethaia 45: 259–273. Crossref
Lauridsen, B.W. and Schnetler, K.I. 2014. A catalogue of Danian gastropods from the Baunekule facies, Faxe Formation, Denmark. Geological Survey of Denmark and Greenland Bulletin 32: 1–117.
Lee, D.E. 1987. Cenozoic and recent inarticulate brachiopods of New Zealand: Discinisca, Pelagodiscus and Neocrania. Journal of the Royal Society of New Zealand 17: 49–72. Crossref
Link, D.H.F. 1807–1808. Beschreibung der Naturalien-Sammlung der Universität zu Rostock. Part 1: 160 pp., Part 2: 30 pp., Part 3: 38 pp. Adlers Erben, Rostock.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Salvius, Stockholm.
Little, C.T.S. and Vrijenhoek, R.C. 2003. Are hydrothermal vent animals living fossils? Trends in Ecology and Evolution 18: 582–588. Crossref
Little, C.T.S., Herrington, R.J., Maslennikov, V.V., Morris, N.J., and Zaykov, V.V. 1997. Silurian hydrothermal vent community from the southern Urals, Russia. Nature 385: 146–148. Crossref
Little, C.T.S., Birgel, D., Boyce, A.J., Crame, J.A., Francis, J.E., Kiel, S., Peckmann, J., Pirrie, D., Rollinson, G.K., and Witts, J.D. 2015. Late Cretaceous (Maastrichtian) shallow water hydrocarbon seeps from Snow Hill and Seymour Islands, James Ross Basin, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 418: 213–228. Crossref
López Cabrera, M.I. and Olivero, E.B. 2011. An Eocene articulated Polyplacophora (Mollusca) from the La Meseta Formation, Antarctica and the stratigraphy of the fossil-bearing strata. Journal of Paleontology 85: 970–976. Crossref
Lorion, J., Kiel, S., Faure, B.M., Masaru, K., Ho, S.Y.W., Marshall, B.A., Tsuchida, S., Miyazaki, J.-I., and Fujiwara, Y. 2013. Adaptive radiation of chemosymbiotic deep-sea mussels. Proceedings of the Royal Society B 280: 20131243. Crossref
Lovén, S.L. 1846. Molluscorum litora Scandinaviae occidentalia habitantium. Öfversigt af Kongliga Vetenskaps Akademiens Förhandlingar 3: 134–160, 182–204.
Lozouet, P. 1998. Nouvelles espèces de Gastéropodes (Mollusca: Gastropoda) de l’Oligocène et du Miocène inférieur de l’Aquitaine (sud-ouest de la France). Cossmanniana 5: 61–102.
Marshall, B.A. 1988. Skeneidae, Vitrinellidae and Orbitestellidae (Mollusca: Gastropoda) associated with biogenic substrata from bathyal depths off New Zealand and New South Wales. Journal of Natural History 22: 949–1004. Crossref
Marshall, B.A. 1986. Recent and Tertiary Cocculinidae and Pseudococculinidae (Mollusca: Gastropoda) from New Zealand and New South Wales. New Zealand Journal of Zoology 12: 505–546.
McLean, J.H. 1987. Taxonomic descriptions of cocculinid limpets (Mollusca, Archaeogastropoda): two new species and three rediscovered species. Zoologica Scripta 16: 325–333. Crossref
McLean, J.H. and Harasewych, M.G. 1995. Review of western Atlantic species of cocculinid and pseudococculinid limpets, with descriptions of new species (Gastropoda: Cocculiniformia). Contributions in Science, Natural History Museum of Los Angeles County 453: 1–33.
Meehan, K.C. and Landman, N. 2016. Faunal associations in cold-methane seep deposits from the Upper Cretaceous Pierre Shale, South Dakota. Palaios 31: 291–301. Crossref
Miyajima, Y., Watanabe, Y., Yanagisawa, Y., Amano, K., Hasegawa, T., and Shimobayashi, N. 2016. A late Miocene methane seep deposit bearing methane-trapping silica minerals at Joetsu, central Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 455: 1–15. Crossref
Muir-Wood, H.M. 1939. Four species of Discinisca (Brachiopoda) from the Eocene of the Hampshire Basin. Proceedings of the Geologists’ Association 50: 149–158.
Müller, O.F. 1776. Zoologiae Danicae prodromus, seu animalium Daniae et Norvegiae indigenarum characters, nomina, et synonyma imprimis popularium. xxxii + 274 pp. Hallageriis, Havniae.
Müller, O.F. 1779. Zwoen wenig bekannten Muscheln, der Schinkenarche und der gerunzelten Mahlermuschel. Gesellschaft Naturforschender Freunde zu Berlin, Beschäftigengen 4: 55–59.
Nilsen, T. 1987. Stratigraphy and sedimentology of the Eocene Tejon Formation western Tehachapi and San Emigdio Mountains, California. United States Geological Survey Professional Paper 1268: 1–110. Crossref
Olsson, A.A. 1931. Contributions to the Tertiary Paleontology of Northern Peru: Part 4, The Peruvian Oligocene. Bulletins of American Paleontology 17: 97–264.
Paull, C.K., Hecker, B., Commeau, R., Freeman-Lynde, R.P., Neumann, C., Corso, W.P., Golubic, S., Hook, J.E., Sikes, E., and Curray, J. 1984. Biological communities at the Florida Escarpment resemble hydrothermal vent taxa. Science 226: 965–967. Crossref
Peckmann, J. and Thiel, V. 2004. Carbon cycling at ancient methane-seeps. Chemical Geology 205: 443–467. Crossref
Peckmann, J., Goedert, J.L., Thiel, V., Michaelis, W., and Reitner, J. 2002. A comprehensive approach to the study of methane-seep deposits from the Lincoln Creek Formation, western Washington State, USA. Sedimentology 49: 855–873. Crossref
Peckmann, J., Kiel, S., Sandy, M. R., Taylor, D. G., and Goedert, J. L. 2011. Mass occurrences of the brachiopod Halorella in Late Triassic methane-seep deposits, Eastern Oregon. Journal of Geology 119: 207–220. Crossref
Peckmann, J., Reimer, A., Luth, U., Luth, C., Hansen, B.T., Heinicke, C., Hoefs, J., and Reitner, J. 2001. Methane-derived carbonates and authigenic pyrite from the northwestern Black Sea. Marine Geology 177: 129–150. Crossref
Prothero, D.R. 2001. Chronostratigraphic calibrations of the Pacific Coast Cenozoic: a summary. The Pacific Section SEPM Special Publication 91: 377–394.
Roterman, C.N., Copley, J.T., Linse, K., Tyler, P.A., and Rogers, A.D. 2013. The biogeography of the yeti crabs (Kiwaidae) with notes on the phylogeny of the Chirostyloidea (Decapoda: Anomura). Proceedings of the Royal Society B 280: 20130718. Crossref
Saether, K.P., Little, C.T.S., and Campbell, K.A. 2010. A new fossil provannid gastropod from Miocene hydrocarbon seep deposits, East Coast Basin, North Island, New Zealand. Acta Palaeontologica Polonica 55: 507–517. Crossref
Saito, H. and Okutani, T. 1990. Two new chitons (Mollusca: Polyplacophora) from a hydrothermal vent site of the Iheya Small Ridge, Okinawa Trough, east China Sea. Venus 49: 165–179.
Schwartz, H., Sample, J.C., Weberling, K.D., Minisini, D., and Moore, J.C. 2003. An ancient linked fluid migration system: cold-seep deposits and sandstone intrusions in the Panoche Hills, California, USA. Geo-Marine Letters 23: 340–350. Crossref
Schweitzer, C.E. and Feldmann, R.M. 2008. New Eocene hydrocarbon seep decapod crustacean (Anomura: Galatheidae: Shinkaiinae) and its paleobiology. Journal of Paleontology 82: 1021–1029. Crossref
Sigwart, J.D. and Chen, C. 2017. Life history, patchy distribution, and patchy taxonomy in a shallow-water invertebrate (Mollusca: Polyplacophora: Lepidopleurida). Marine Biodiversity [published online, 10.1007/s12526-017-0688-1].
Sigwart, J.D., Andersen, S.B., and Schnetler, K.I. 2007. First record of a chiton from the Palaeocene of Denmark (Polyplacophora: Leptochitonidae) and its phylogenetic affinities. Journal of Systematic Palaeontology 5: 123–132. Crossref
Sigwart, J.D., Schwabe, E., Saito, H., Samadi, S., and Giribet, G. 2010. Evolution in the deep sea: a combined analysis of the earliest diverging living chitons (Mollusca: Polyplacophora: Lepidopleurida). Invertebrate Systematics 24: 560–572. Crossref
Simone, L.R.L. and Zelaya, D.G. 2004. A new Orbitestella (Gastropoda: Heterobranchia: Orbitestellidae) from Tierra del Fuego, Argentina. The Nautilus 118: 160–166.
Sirenko, B.I. 2004. The ancient origin and persistence of chitons (Mollusca, Polyplacophora) that live and feed on deep submerged land plant matter (xylophages). Bolletino Malacologico Supplemento 5: 105–110.
Sirenko, B.I. 2013. Four new species and one new genus of Jurassic chitons (Mollusca: Polyplacophora: Lepidopleurida) from the Middle Russian Sea. Proceedings of the Zoological Institute RAS 317: 30–44.
Smrzka, D., Kraemer, S.M., Zwicker, J., Birgel, D., Fischer, D., Kasten, S., Goedert, J.L., and Peckmann, J. 2015. Constraining silica diagenesis in methane-seep deposits. Palaeogeography, Palaeoclimatology, Palaeoecology 420: 13–26. Crossref
Sowerby, G.B., I 1833. [… the collection of shells formed by Mr. Cuming on the western coast of South America][Archidae]. Proceedings of the Zoological Society of London 1 (for 1833): 16–22.
Squires, R.L. 1995. First fossil species of the chemosynthetic-community gastropod Provanna: Localized cold-seep limestones in Upper Eocene and Oligocene rocks, Washington. The Veliger 38: 30–36.
Squires, R.L. and Goedert, J.L. 1995. An extant species of Leptochiton (Mollusca: Polyplacophora) in Eocene and Oligocene cold-seep limestones, Olympic Peninsula, Washington. The Veliger 38: 47–53.
Squires, R.L. and Goedert, J.L. 1996a. A new species of Thalassonerita? (Gastropoda: Neritidae?) from a Middle Eocene cold-seep carbonate in the Humptulips Formation, Western Washington. The Veliger 39: 270–272.
Squires, R.L. and Goedert, J. L. 1996b. New species of small to minute gastropods of Early Eocene Age from the Crescent Formation, Black Hills, Southwest Washington. The Veliger 39, 226–240.
Thiele, J. 1909. Revision des Systems der Chitonen. Zoologica Stuttgart 22: 1–132.
Tomlin, J.R.l.B. 1927. Reports on the marine Mollusca in the collections of the South African Museum II. Families Abyssochrysidae, Oocorythidae, Haliotidae, Tonnidae. Annals of the South African Museum 25: 77–83.
Valdés, F., Sellanes, J., and D’Elía, G. 2013. Phylogenetic position of vesicomyid clams from a methane seep off central Chile (~36°S) with a molecular timescale for the diversification of the Vesicomyidae. Zoological Studies 51: 1154–1164.
Vinn, O., Kupriyanova, E.K., and Kiel, S. 2013. Serpulids (Annelida, Polychaeta) at Cretaceous to modern hydrocarbon seeps: ecologic and evolutionary patterns. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 35–41. Crossref
Vinn, O., Mutvei, H., Ten Hove, H.A., and Kirsimäe, K. 2008. Ultrastructure and mineral composition of serpulid tubes (Polychaeta, Annelida). Zoological Journal of the Linnean Society 154: 633–650. Crossref
Vrijenhoek, R.C. 2013. On the instability and evolutionary age of deep-sea chemosynthetic communities. Deep-Sea Research II 92: 189–200. Crossref
Warén, A., and Bouchet, P. 1986. Four new species of Provanna Dall (Prosobranchia, Cerithiacea?) form East Pacific hydrothermal sites. Zoologica Scripta 15: 157–164. Crossref
Warén, A., and Bouchet, P. 1993. New records, species, genera, and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta 22: 1–90. Crossref
Warén, A. and Bouchet, P. 2001. Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44: 116–231.
Warén, A. and Ponder, W.F. 1991. New species, anatomy, and systematic position of the hydrothermal vent and hydrocarbon seep gastropod family Provannidae fam. n. (Caenogastropoda). Zoologica Scripta 20: 56–102. Crossref
Warén, A., Carozza, F., and Rocchini, R. 1996. Description of two new species of Hyalogyrinidae (Gastropoda, Heterobranchia) from the Mediterranean. Bolletino Malacologico 32: 57–66.
Wells, R.E., Bukry, D., Friedman, R.M., Pyle, D., Duncan, R., Haeussler, P., and Wooden, J.L. 2014. Geologic history of Siletzia, a large igneous province in the Oregon and Washington Coast Range: Correlation to the geomagnetic polarity time scale and implications for a long-lived Yellowstone hotspot. Geosphere 10: 692–719. Crossref
Zhang, S. and Zhang, S. 2017. Description of Pyropelta elongata sp. nov. (Gastropoda, Pyropeltidae) from a methane seep area in the South China Sea. American Malacological Bulletin 35: 51–54. Crossref
Acta Palaeontol. Pol. 63 (4): 751–768, 2018
https://doi.org/10.4202/app.00525.2018