
The mystery of Mystriosaurus: Redescribing the poorly known Early Jurassic teleosauroid thalattosuchians Mystriosaurus laurillardi and Steneosaurus brevior
SVEN SACHS, MICHELA M. JOHNSON, MARK T. YOUNG, and PASCAL ABEL
Sachs, S., Johnson, M.M., Young, M.T., and Abel, P. 2019. The mystery of Mystriosaurus: Redescribing the poorly known Early Jurassic teleosauroid thalattosuchians Mystriosaurus laurillardi and Steneosaurus brevior. Acta Palaeontologica Polonica 64 (3): 565–579.
The genus Mystriosaurus, established by Kaup in 1834, was one of the first thalattosuchian genera to be named. The holotype, an incomplete skull from the lower Toarcian Posidonienschiefer Formation of Altdorf (Bavaria, southern Germany), is poorly known with a convoluted taxonomic history. For the past 60 years, Mystriosaurus has been considered a subjective junior synonym of Steneosaurus. However, our reassessment of the Mystriosaurus laurillardi holotype demonstrates that it is a distinct and valid taxon. Moreover, we find the holotype of “Steneosaurus” brevior, an almost complete skull from the lower Toarcian Whitby Mudstone Formation of Whitby (Yorkshire, UK), to be a subjective junior synonym of M. laurillardi. Mystriosaurus is diagnosed in having: a heavily and extensively ornamented skull; large and numerous neurovascular foramina on the premaxillae, maxillae and dentaries; anteriorly oriented external nares; and four teeth per premaxilla. Our phylogenetic analyses reveal M. laurillardi to be distantly related to Steneosaurus bollensis, supporting our contention that they are different taxa. Interestingly, our analyses hint that Mystriosaurus may be more closely related to the Chinese teleosauroid (previously known as Peipehsuchus) than any European form.
Key words: Thalattosuchia, Teleosauroidea, Mystriosaurus, Jurassic, Toarcian Posidonienschiefer Formation, Whitby Mudstone Formation, Germany, UK.
Sven Sachs [Sachs.Pal@gmail.com], Naturkunde-Museum Bielefeld, Abteilung Geowissenschaften, Adenauerplatz 2, 33602 Bielefeld, Germany; Im Hof 9, 51766 Engelskirchen, Germany (address for correspondence).
Michela M. Johnson [Michela.Johnson@ed.ac.uk] and Mark T. Young [Mark.Young@ed.ac.uk], School of GeoSciences, Grant Institute, University of Edinburgh, James Hutton Road, Edinburgh, EH9 3FE, UK.
Pascal Abel [pascal.abel@ifg.uni-tuebingen.de], Senckenberg Centre for Human Evolution and Palaeoenvironment, Eberhard-Karls-Universität Tübingen, Sigwartstraße 10, 72076 Tübingen, Germany.
Received 12 October 2019, accepted 17 April 2019, available online 12 September 2019.
Copyright © 2019 S. Sachs et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Thalattosuchians were a diverse group of aquatic crocodylomorphs, which flourished in marine and brackish ecosystems during the Jurassic and Cretaceous Periods (Buffetaut 1982; Hua and Buffetaut 1997; Young et al. 2010; Foffa et al. 2015, 2019). They are divided into two groups, teleosauroids and metriorhynchoids (sensu Young and Andrade 2009). Teleosauroids are particularly well known from numerous complete skeletons discovered in the Toarcian Posidonienschiefer Formation (Holzmaden area) of southwestern Germany (Hauff and Hauff 1981). A second, less well-known but historically important locality in southern Germany is the Altdorf region in northern Bavaria (Mäuser 2001). One of the early fossil collectors from Altdorf was Johann Friedrich Bauder, a merchant and avid naturalist. From 1770 to 1776, Bauder was mayor of Altdorf and during that time, he made one of his most important discoveries, an incomplete thalattosuchian skull. This specimen, which Bauder discovered in a quarry near Altdorf, is one of the first ever described thalattosuchian fossils and has a long and remarkable history (see below). In 1834, Johann Jakob Kaup established it as the holotype of Mystriosaurus laurillardi Kaup, 1834 (see Fig. 1). During the latter half of the 1800s, Blake (1876) described a different teleosauroid skull from the Whitby Mudstone Formation of Whitby (UK) as “Steneosaurus” brevior Blake, 1876 (see Fig. 2), a taxon that was later considered to be closely related to M. laurillardi (see below). However, the validity of Mystriosaurus has often been questioned (e.g., Westphal 1961, 1962; Steel 1973) and in recent publications this genus has been considered to be a subjective junior synonym of Steneosaurus (e.g., Westphal 1962).
During the 19th and early 20th centuries, Teleosauroidea was considered to be a generically diverse clade (e.g., Eudes-Deslongchamps 1877; Mook 1934) and numerous taxa were described (e.g., Eudes-Deslongchamps 1867–1869; Andrews 1909, 1913). However, during the latter half of the 20th century there was a trend towards taxonomic lumping, at both the genus and species levels (e.g., Westphal 1962; Steel 1973; Adams-Tresman 1987). By the end of the 20th century, only four genera were commonly recognised: Machimosaurus, Platysuchus, Steneosaurus, and Teleosaurus; there were also disagreements on whether the genera Peipehsuchus and Pelagosaurus belonged within Teleosauroidea or not (Mueller-Töwe 2005). Of these four, Platysuchus is monospecific; Teleosaurus was largely restricted to the Bathonian-aged material from England and France; Machimosaurus housed the distinctive Late Jurassic–Early Cretaceous durophagous forms, and “Steneosaurus” was a wastebasket genus for all other species (ranging from the Toarcian to the Tithonian).
However, since the beginning of the 21st century, certain teleosauroid interrelationships were tentatively being examined using a phylogenetic approach. These analyses all agree that the genus “Steneosaurus” is not monophyletic (e.g., Mueller-Töwe 2006; Jouve 2009; Young et al. 2012; Young 2014; Wilberg 2015; Fanti et al. 2016; Ősi et al. 2018: datasets 2 and 3; Foffa et al. 2019; Johnson et al. 2019). In fact, in the two widest sampled teleosauroid analyses (Ősi et al. 2018: datasets 2 and 3), the only way to make Steneosaurus monophyletic would result in Teleosauroidea being monogeneric (i.e., only Steneosaurus). This highlights the extent of the “Steneosaurus lumping problem”, which is masking the range of morphological diversity in Teleosauroidea and its alpha taxonomy (e.g., see Jouve et al. 2017). As such, the validity of the genus “Steneosaurus” is currently being worked on by MMJ, and is beyond the scope of this paper.
Both Mystriosaurus laurillardi and “Steneosaurus” brevior have been considered to be subjective junior synonyms of the ubiquitous lower Toarcian teleosauroid Steneosaurus bollensis (Jäger, 1828), at some point over the past 100 years (see below). The holotypes of both species have not been redescribed in over 100 years, nor have they been specifically compared to one another. Here we provide a detailed redescription and phylogenetic analysis of the holotype specimens of Mystriosaurus laurillardi and “Steneosaurus” brevior and demonstrate that: (i) both represent the same species (M. laurillardi) and (ii) Mystriosaurus is a distinct and valid genus.
Institutional abbreviations.—CAMSM, Sedgewick Museum, Cambridge, UK; HLMD, Hessisches Landesmuseum, Darmstadt, Germany; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; LPP (PALEVOPRIM-CVCU), Institut de Paléoprimatologie, Paléontologie humaine: Evolution et Paléoenvironnements Université de Poitiers, Poitiers, France; MMG, Staatliches Museum für Mineralogie und Geologie, Dresden, Germany; MNHN, Muséum national d’Histoire naturelle, Paris, France; MNHNL, Musée national d’histoire naturelle, Luxembourg, Luxembourg; NHMUK, Natural History Museum, London, UK; PETMG, Peterborough Museum and Art Gallery, Peterborough, UK; PMU, Evolutionsmuseet, Uppsala universitet, Uppsala, Sweden; RE, Ruhr Museum, Essen, Germany; SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany; UH, Urweltmuseum Hauff, Holzmaden, Germany.
Other abbreviations.—CI, consistency index; HI, homoplasy index; MPC, most parsimonious cladograms; OTU, operational taxonomic unit; RCI, rescaled consistency index; RI, retention index.
Historical background
Mystriosaurus laurillardi.—The first, and very detailed, description of Bauder’s find was provided in 1776 by Johann Ernst Immanuel Walch in the journal “Der Naturforscher” [The Naturalist], of which he was editor during that time. Walch also figured the fossil (Walch 1776: pl. 4: 8) and referred to it as the skull of a “Crocodill”. This identification was questioned, however, by Johann Samuel Schröter, who discussed the specimen and Walch’s (1776) conclusions in a detailed rebuttal that he published in 1780. Rather than the fossil belonging to a crocodile, Schröter (1780: 530) suggested that the Altdorf find might belong to a large ant-eating animal. Some years later, on 2 April 1783, the poet Johann Wolfgang von Goethe reported in a letter to his friend (the wealthy Darmstadt-based naturalist Johann Heinrich Merck) that Bauder was going to send him the specimen, which von Goethe apparently planned to purchase (Wagner 1835: 384). Merck, however, was able to acquire it first and paid for it the sum of 20 Louis d’or (Wagner 1835: 389). Merck (1786: 81) briefly discussed the fossil after it was in his collection and compared it with a “Gangetischen Krokodil”, which presumably refers to the Indian gavial. Merck died in 1791, and the specimen was purchased by landgrave Ludwig X in 1792. It was added to the Nature Cabinet at Darmstadt, which formed the foundation of today’s Hessisches Landesmuseum (Westphal 1962: 78). Faujas de Saint Fond (1799: pl. 54) illustrated the Darmstadt skull and followed Merck’s identification of it as a gavial. Cuvier (1808) then described it as a crocodile, a determination that he followed in his 1825 publication (Cuvier 1825: 115ff., pl. 6: 10–15). Von Sömmerring (1814) briefly described the specimen as the skull of a fossil gavial.
Krüger (1825) mentioned the name Crocodilus cylindrirostris for one species of gavial that had been found near Altdorf. This name also appeared in several later publications (e.g., Pierer 1827; Holl 1829; de la Beche 1832; Glocker 1839). Holl (1829) mentioned the names Crocodilus altorfinus and Crocodilus brevirostris when referring to material from the same locality. Both C. cylindrirostris and C. altorfinus were later considered to be subjective junior synonyms of C. brevirostris, which was subsequently referred to as Steneosaurus brevirostris (e.g., Giebel 1847). However, it is not clear if any of these species epithets specifically referred to the Darmstadt specimen, as the specimen was not mentioned in relation with any of these binomina. Furthermore, the Darmstadt specimen was not mentioned alongside other thalattosuchian material from Altdorf that was known by that time (see Von Sömmerring 1814).
In 1832, von Meyer (1832: 106) established the taxon Streptospondylus altdorfensis for material from France and Germany. This taxon, however, is a chimera represented by both thalattosuchian and theropod dinosaur remains (Allain 2001). Von Meyer (1832) considered Cuvier’s (1801) “Gavial de Honfleur” as the holotype of S. altdorfensis but also added the Darmstadt specimen to the hypodigm without providing an explanation (von Meyer 1832: 226–227).
In 1834, Johann Jakob Kaup, by then assistant at the Nature Cabinet in Darmstadt, assembled a catalogue of plaster casts within the Darmstadt collection. Original specimens were listed in the additions of this catalogue and here Kaup (1834) first mentioned the name Mystriosaurus laurillardi specifically for the skull from Altdorf. Based upon a letter from Kaup, a brief note on the existence of the M. laurillardi specimen was also published by Münster (1834). Kaup (1837) provided a more detailed description, listing the diagnostic characters. In this publication, additional cranial and postcranial material from the same locality at Altdorf is mentioned, which by then was housed in the collection of the Graf zu Münster (the current whereabouts of this material are unknown). Kaup (1837) later referred other specimens from Germany and England to Mystriosaurus and established several new species.
Wagner (1850: 595), however, pointed out that the diagnostic characters given by Kaup in Bronn (1837) are not sufficient as they also occur in Teleosaurus. Still Wagner (1850) retained Mystriosaurus as a valid genus, but mainly because Teleosaurus derives from younger strata. d’Alton and Burmeister (1854) discussed Wagner’s (1850) conclusion and considered Teleosaurus chapmani as being a subjective junior synonym of Mystriosaurus laurillardi. Eudes-Deslongchamps (1863–1869) also questioned the validity of Mystriosaurus and established a new combination for the type species: Teleosaurus (Steneosaurus) laurillardi; a view which only some (e.g., Lydekker 1888) followed. Eudes-Deslongchamps (1877: 4) included in his “téléosauriens” numerous valid genera: Pelagosaurus, Mystriosaurus, Teleosaurus, Steneosaurus, Spatocranius, Teleidosaurus, Aeolodon, and Metriorhynchus. The teleosauroids in Mook (1934) were similarly diverse: Pelagosaurus, Teleosaurus, Teleidosaurus, Aeolodon, Crocodileimus, Gnathosaurus, Steneosaurus, Mycterosaurus (an error in print, should have been Mycterosuchus), and Mystriosaurus. Note, Mook (1934) considered Machimosaurus to be a goniopholidid (an opinion common until the work of Krebs 1967).
The generic name Mystriosaurus was broadly accepted in various publications during the 19th and early 20th centuries (e.g., Bronn 1842; Wagner 1850; d’Alton and Burmeister 1854; Winkler 1878; Woodward 1885; Drevermann 1914; Hauff 1921; Berckhemer 1929; Mook 1934; Hölder 1955; Huene 1956). It has only been since Westphal’s (1961, 1962) revisions of the Lower Jurassic thalattosuchians of Germany and England that Mystriosaurus laurillardi has been widely accepted to be a subjective junior synonym of Steneosaurus bollensis (e.g., the review of crocodylomorphs by Steel 1973 follows that taxonomic change made by Westphal 1962). Moreover, Mystriosaurus is rarely mentioned in more recent revisions of Early Jurassic European thalattosuchians (e.g., Mueller-Töwe 2006; Young and Steel in press).
“Steneosaurus” brevior.—Compared to the historical background of Mystriosaurus laurillardi (see above), that of “Steneosaurus” brevior is relatively straightforward. The holotype specimen was discovered in Whitby (UK) (although when and by whom is unknown) and was purchased by the British Museum of London (now the Natural History Museum London) in 1840. The specimen was long referred to as the manuscript name of Teleosaurus brevior, established by Richard Owen (Blake 1876; Woodward 1885). Blake (1876) identified it as belonging to the genus Steneosaurus and was the first to describe and name the specimen, as “Steneosaurus” brevior. However, Blake apparently briefly considered the possibility of it belonging to Teleosaurus chapmani (see Vignaud 1995). Owen (1884) reprinted his earlier 1841 paper on T. chapmani, but also included information on the holotype of “S”. brevior, which he still referred to as T. brevior. Woodward (1885: 499) suggested Owen (1884) had reprinted his 1841 work “without attempting to incorporate the results of recent Continental research”. Woodward (1885: 499) went on to discuss the recent teleosauroid taxonomic changes made by Eudes-Deslongchamps (1863–1869, 1877), and stated that in following them “S.” brevior “would be relegated to Mystriosaurus”. In his overview taxonomic table, Woodward (1885) had both T. chapmani and S. brevior as species of Mystriosaurus. Westphal (1961, 1962) used the binomen “Steneosaurus” brevior, although he was unsure about the taxonomic status of the species, hypothesising that “S.” brevior could be basal to S. bollensis. However, Steel (1973) listed “S.” brevior as one of the numerous subjective junior synonyms of S. bollensis. Walkden et al. (1987) again were unsure about the taxonomic status of “S.” brevior, as they too raised the possibility it was synonymous with S. bollensis. More recently, “S.” brevior has been recognised as a distinct taxon (e.g., Vignaud 1995; Mueller-Töwe 2006; Pierce et al. 2009; Young and Steel in press). Young and Steel (in press) concluded that the only UK museum specimen that could be definitively referred to “S.” brevior is the holotype (NHMUK PV OR 14781). They highlighted that previous studies had erroneously referred all massive specimens to this species, and that they were in fact very large and robust individuals of S. bollensis.
Young and Steel (in press) stated that part of the taxonomic confusion of this species originated from Blake (1876), who referred a second large partial skull (NHMUK PV R 756) to “S.” brevior when originally describing the species. However, Young and Steel (in press) referred this specimen to S. bollensis based on their revised species diagnoses of the thalattosuchians from the Whitby Mudstone Formation. Furthermore, they stated that all other fragmentary museum-based specimens previously referred to “S.” brevior must be considered Teleosauroidea indeterminate.
Interestingly, there is a potentially older name for this species. Edward Charlesworth announced a new species, Teleosaurus ischnodon, at the 24th meeting of the British Association for the Advancement of Science held in Liverpool in 1854. Unfortunately, Charlesworth seems to have never formally published the name, or given a diagnosis (Blake 1876; Woodward 1885). Thus, it is a nomen nudum. Blake (1876: 245) mentioned when establishing “S.” brevior, that T. ischnodon could have been the same species. Unfortunately, there is no way of establishing what specimen was intended to be the type specimen, if it has survived to the present day.
Systematic palaeontology
Thalattosuchia Fraas, 1901
Teleosauroidea Geoffroy Saint-Hilaire, 1831
Genus Mystriosaurus Kaup, 1834
Type species: Mystriosaurus laurillardi Kaup, 1834, Altdorf bei Nürnberg, lower Toarcian, Lower Jurassic.
Mystriosaurus laurillardi Kaup, 1834
Figs. 1–4.
1834 Mystriosaurus laurillardi gen. et sp. nov.; Kaup 1834: p. 28 [not figured].
1876 Steneosaurus brevior sp. nov.; Blake 1876: pl. 1:1, 2.
Holotype: HLMD V946-948, an incomplete cranium and mandible.
Type locality: Altdorf bei Nürnberg, Nürnberger Land district, Bavaria, southern Germany.
Type horizon: Posidonienschiefer Formation, Schwarzjura Group, probably Harpoceras serpentinum Sub-Mediterranean Ammonite Zone, lower Toarcian, Lower Jurassic (Page 2003; German Stratigraphic Commission 2016).
Emended diagnosis.—Teleosauroid crocodylomorph with the following unique combination of characters (proposed autapomorphic characters indicated by an asterisk): robust skull with well-developed and extensive ornamentation on the premaxillae, maxillae, nasals*, frontal, prefrontal, lacrimal, jugal*,postorbital and squamosal*; frontal ornamentation composed of small sub-circular to elongate pits that are closely spaced or that fuse and become a ridge-groove pattern; mesorostrine snout (preorbital length is approximately 66% of skull length), which is broad near the orbits but narrows and becomes slightly more slender anteriorly; slight constriction of the snout in front of the orbits; large and numerous neurovascular foramina on the premaxillae, maxillae and dentaries (shared with Machimosaurini); external nares oriented anteriorly*; the anterior and anterolateral margins of the premaxilla are orientated anteroventrally and extend ventrally in lateral view (shared with Mycterosuchus nasutus Andrews, 1913, Platysuchus multiscrobiculatus Berckhemer, 1929, the Chinese teleosauroid previously referred to as Peipehuschus (see Li 1993), Bathysuchus megarhinus Hulke, 1871; Foffa et al. 2019); antorbital fenestrae almost equidistant to the orbits and alveolar margins (shared with P. multiscrobiculatus); the antorbital fenestrae are large relative to the orbits (anteroposterior length equals 25% of orbital length), being slightly anteroposteriorly elongated and subcircular in shape; supratemporal fossae form an approximate isosceles trapezoid-shape*; anterolateral margin of supratemporal fossae (i.e., lateral processes of the frontal and the dorsal processes of the postorbitals) noticeably inclined anterolaterally (shared with Mycterosuchus nasutus, Platysuchus multiscrobiculatus, Chinese teleosauroid, Bathysuchus megarhinus); the anterolateral corners of the supratemporal fossae extend significantly lateral to the lateral margin of the orbits (= the anterior region of the supratemporal fenestrae are very wide); medial margin of supratemporal arch is straight, with no significant concavity*; the orbits are subcircular in shape, and are dorsolaterally orientated; postorbital reaches the orbit posteroventral margin (shared with P. multiscrobiculatus, Chinese teleosauroid, B. megarhinus); short frontal anteromedial process, being noticeably shorter than the prefrontals when seen in dorsal view; ventral border of angular horizontal and poorly curved (shared with Steneosaurus gracilirostris Westphal, 1961); mandibular fenestra poorly elliptic*; four teeth per premaxilla; large, slightly robust teeth that have numerous conspicuous apicobasally aligned ridges, and a pointed apex, with some anteriorly-placed tooth crowns being procumbent*.
Material.—NHMUK PV OR 14781, an almost complete skull associated with a fragmentary limb bone (holotype of “Steneosaurus” brevior) from Whitby, Yorkshire, UK; Mulgrave Shale Member, Whitby Mudstone Formation, Lias Group, Harpoceras serpentinum Sub-Boreal Ammonite Zone, lower Toarcian, Lower Jurassic (Blake 1876).
Description.—The holotype of Mystriosaurus laurillardi (HLMD V946-948) is a three-dimensionally preserved skull (Fig. 1) that measures approximately 650 mm along the dorsal midline. HLMD V946-948 consists of an isolated rostrum and the posterior part of the skull, preserved as a slab and counter-slab. The majority of the skull roof, the occipital part and the rear portion of the mandible are either missing or obscured by matrix. Infillings of the internarial cavities are preserved in the counter-slab. The block with the post-symphyseal mandibular portion has two carved holes, which Kaup in Bronn and Kaup (1841) discussed as Gaumenlöcher (“palatal fenestrae”). The anterior rostrum (premaxillae, maxillae, and dentary) are cemented together.

Fig. 1. Teleosauroid thalattosuchian Mystriosaurus laurillardi Kaup, 1834 (HLMD V946-948, holotype), lower Toarcian of Altdorf (southern Germany); skull in dorsal (A1) and lateral (A2) views, midsection of the skull in ventral view (A3), and of the mandible in dorsal view (A4).
NHMUK PV OR 14781 comprises a nearly complete cranium and mandible (Fig. 2) which are adhered together tightly so that the cranium in palatal view and mandible dorsal view are inaccessible. Both the left posterior region and occipital of the cranium are not preserved; in addition, the right posterior portion of the cranium is partially covered with matrix (mainly seen in dorsal view; Fig. 2A2). In ventral view (Fig. 2A3), the posterior part of the cranium is likewise covered in matrix, and therefore cannot be described at present. In dorsal view, there is a limb bone (Fig. 2A2) preserved in the matrix; however, only the shaft is well-preserved, making identification difficult (the anterior part appears to resemble the humeral head found in other teleosauroids, making this bone a possible humerus). All information pertaining to the jugal, prefrontal, lacrimal, frontal, postorbital, squamosal, quadratojugal, and surangular was inferred from NHMUK PV OR 14781.
Cranium: The cranium of NHMUK PV OR 14781 is approximately 595 mm from the anterior tip of the rostrum to the anterior margin of the orbit. The overall appearance of the preorbital skull is mesorostrine and is slightly more robust, shorter, and rugose than those seen in the other Toarcian teleosauroids (e.g., Steneosaurus bollensis, SMNS 51753; S. gracilirostris, NHMUK PV OR 14792; Platysuchus multiscrobiculatus, SMNS 9930). The anterior premaxillae and dentaries are laterally expanded similarly to the condition in other teleosauroids (e.g., Andrews 1909, 1913; Hua 1999; Young et al. 2014;Johnso n et al. 2017). The orbits are large, circular and anteroposteriorly oriented (Fig. 2A1, A2), and in dorsal view the partially preserved right supratemporal fenestra is sub-rectangular in shape, with the anteroposterior axis being more than 10% longer than the lateromedial axis. The anterolateral margin of the supratemporal fossae are inclined anterolaterally; the anterolateral corners of the supratemporal fossae extend noticeably more anterior than the anteromedial corners of the supratemporal fossae (Fig. 2A2). The medial margin of the supratemporal arch is relatively straight in dorsal view.
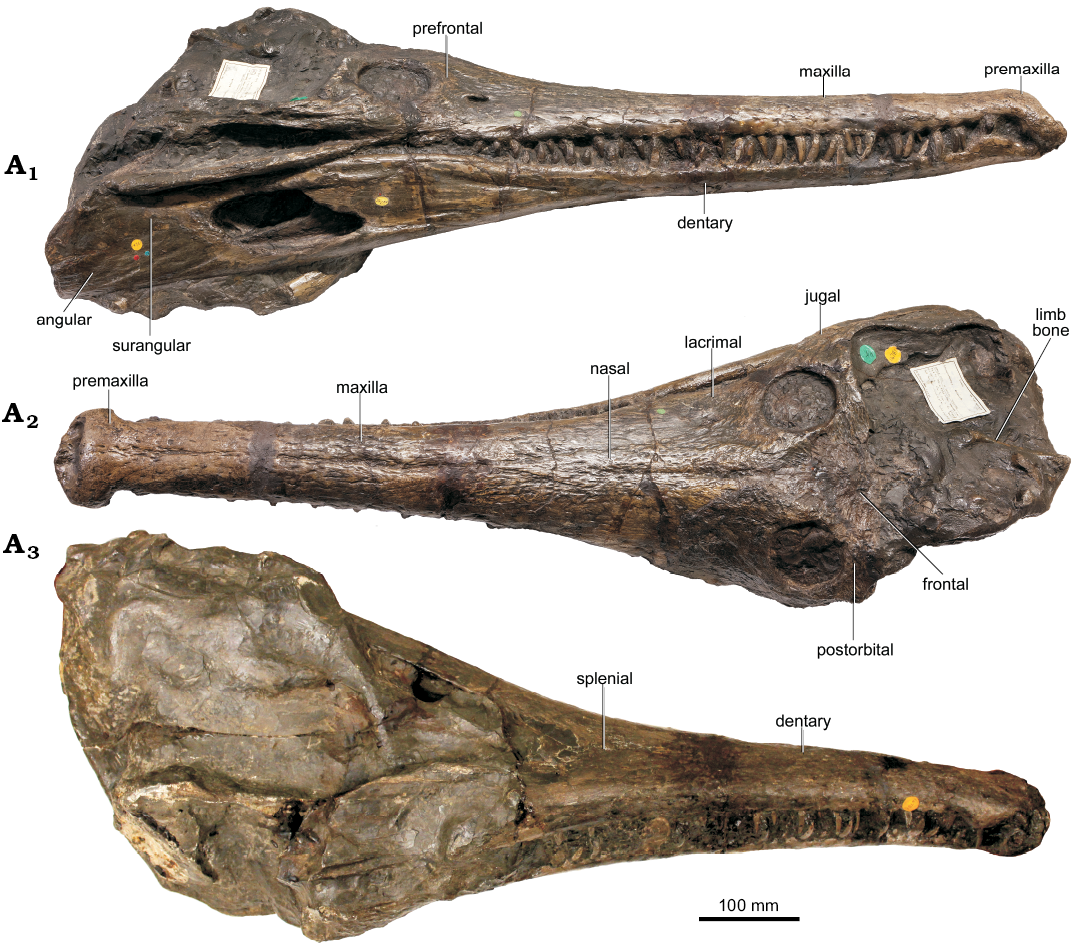
Fig. 2. Teleosauroid thalattosuchian Mystriosaurus laurillardi Kaup, 1834 (NHMUK PV OR 14781, holotype of Steneosaurus brevior Blake, 1876, lower Toarcian of Whitby (Yorkshire, UK); skull in lateral (A1), dorsal (A2), and ventral (A3) views.
Premaxilla: The premaxillae (Figs. 1–3) in both HLMD V946-948 and NHMUK PV OR 14781 are large and dorsoventrally deep. They surround the external narial opening and are oriented anteriorly. The external nares are laterally expanded and their posterior margins do not reach beyond the third premaxillary alveolar pair. In NHMUK PV OR 14781, there is a small premaxillary septum protruding into the nares, giving them a slight B-shape in anterior view (Fig. 3B2). This septum is broken off in HLMD V946-948. The anterior two-thirds of the premaxilla is laterally expanded, anteroposteriorly shortened and the anterior margin is ventrally deflected, giving the snout a scoop-like appearance. In dorsal view, the premaxilla-maxilla suture is slightly interdigitating and rounded. In lateral view, the anterior and anterolateral premaxillary margins are orientated anteroventrally and extend ventrally (which is similar to Platysuchus multiscrobiculatus, SMNS 9930 and differs from Steneosaurus bollensis, RE 551.762.130 A 0248 (see SOM: fig. S1, Supplementary Online Material available at http://app.pan.pl/SOM/app64_3-Sachs_etal_SOM.pdf). The dentigerous anterior portion of the conjoint premaxillae forms a rosette with undulated margins. A prominent diastema is present that starts after the last premaxillary tooth position and here the premaxillae are transversely constricted (Figs. 1, 2). The posterodorsal portions of the premaxillae together form a prominent tip that terminates at the level of the third maxillary tooth position and separates the maxillae anterodorsally. In both HLMD V946-948 and NHMUK PV OR 14781, the mandible obscures the majority of the ventral view of the premaxillae; however, in both ventral views (Fig. 3C, D), the lateral margins of four alveoli are visible. The first and second alveoli are placed anteriorly, and are nearly confluent. A small, slightly indented gap is present between the first premaxillary tooth and the midline suture, and between the second and third premaxillary teeth, there is an additional, larger gap that is indented in ventral view. The third premaxillary tooth is prominent, more so when compared to the other premaxillary teeth. Between the third and fourth premaxillary teeth there is a prominent concave gap, such as in other teleosauroids (e.g., Steneosaurus leedsi Andrews, 1909, NHMUK PV R 3806). The last premaxillary tooth is the smallest of the four teeth. Both premaxillae are well ornamented with conspicuous pits and grooves (Figs. 1, 2). Best seen in NHMUK PV OR 14781, there is a cluster of large circular foramina along the anterior and lateral margins of the external nares visible in dorsal, lateral and anterior views (Figs. 2, 3).
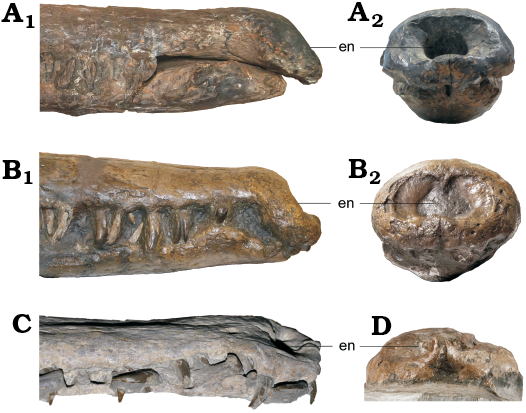
Fig. 3. Comparison between teleosauroid thalattosuchians Mystriosaurus laurillardi Kaup, 1834 (HLMD V946-948, holotype), lower Toarcian of Altdorf (southern Germany) (A), Mystriosaurus laurillardi Kaup, 1834 (NHMUK PV OR 14781, holotype of Steneosaurus brevior Blake, 1876, lower Toarcian of Whitby (Yorkshire, UK) (B), and Steneosaurus bollensis (Jäger, 1828) (PMU R161, RE 551.762.130 A 0248), lower Toarcian of Holzmaden (southwestern Germany) (C, D, respectively). Anterior portion of rostrum in lateral (A1, B1, C) and anterior (A2, B2, D) views, showing the ventrally deflected premaxilla and the anteriorly facing external naris (en). Not to scale.
Maxilla: The maxillae (Figs. 1, 2) form a substantial part of the rostrum. In HLMD V946-948, the anterior portions of the maxillae are well preserved, but the dorsal midline suture is barely visible. The maxillae are elongate, transversely narrow elements, and anterodorsally separated by the premaxillae (Figs. 1, 2, 4). Their lateral margins are sub-parallel in dorsal view. The maxillae seperate the nasals from the premaxillae. The lateral maxillary margins are undulated and slightly bulged at the tooth positions, especially in the anterior part (indicating deep reception pits). In HLMD V946-948, the posterior maxillary portion is only partly preserved on the left side, adjacent to the nasal. The latter contacts the maxilla via a smooth, straight suture that extends anteromedially. All pointed tooth crowns (Figs. 1–3) are intersected and expose the pulp cavity in lateral view. In HLMD V946-948, the exact number of maxillary teeth cannot be provided, as both maxillae are incomplete. In NHMUK PV OR 14781, the precise maxillary tooth count is also difficult to determine, but there are at least 29 alveolar pairs. The maxilla is heavily and extensively ornamented with conspicuous pits and grooves. In addition, there are deep, well-spaced foramina (arranged in a single line parallel to the maxillary ventral margin) that are visible in dorsal, lateral and anterior views (best seen in NHMUK PV OR 14781). In both specimens, the mandible obscures the ventral sides of the maxillae.
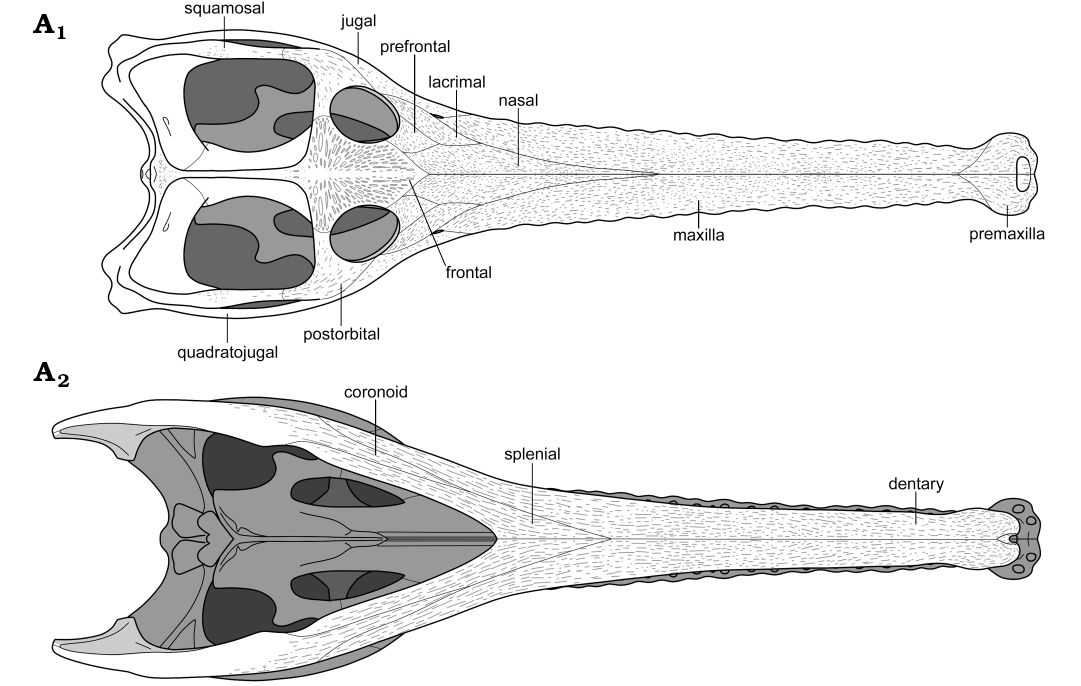
Fig. 4. Reconstruction of teleosauroid thalattosuchian Mystriosaurus laurillardi Kaup, 1834 cranium and mandible in dorsal (A1) and ventral (A2) view. The reconstructed portions are illustrated in SOM: fig. S6. Not to scale.
Jugal: The right jugal in NHMUK PV OR 14781 is complete (Fig. 2) whereas the left jugal is partially preserved (the posterior area is missing). It is triradiate, forms the lateral border of the orbit as seen in other teleosauroids (Andrews 1909, 1913), and participates in the ventral margin of the orbit. The anterior jugal is short and does not extend anteriorly past the orbits (Figs. 2, 4). The postorbital-jugal contact (best seen in right lateral view) appears to be anteroposteriorly straight (with the postorbital overlapping the jugal). The quadratojugal-jugal contact cannot be clearly seen. The jugal is also ornamented with elongate, deep grooves and small sub-oval pits.
Nasals: In both specimens, the dorsal surfaces of the nasals are exposed (Figs. 1, 2). The nasals are large triangular bones exposed on the dorsal surface of the posterior rostrum and orbital area. The straight midline suture is clearly visible and so is the anteromedially extending lateral suture to the maxilla. The overall anteroposterior elongation of the nasals is slightly shorter and mediolaterally broader than in other Toarcian teleosauroids (e.g., S. gracilirostris NHMUK PV OR 14792). The nasal anterior processes are approximately one-third of the posterior rostrum and have a well-developed near-parallel suture contact with the maxillae. There is no dorsoventral abrupt “doming” of the nasals (see Lemmysuchus obtusidens (Andrews, 1913), Johnson et al. 2017, NHMUK PV R 3168, LPP.M.21, PETMG R39). A midline concavity (“midline trench”) is present in NHMUK PV OR 14781. There is a faint internarial suture in HLMD V946-948 and NHMUK PV OR 14781, suggesting that the nasals are unfused. The nasals, similarly to the premaxillae and maxillae, are noticeably ornamented in (best seen in NHMUK PV OR 14781, Fig. 2A2), with multiple, deep grooves, which differs from other Toarcian teleosauroids (e.g., S. gracilirostris, NHMUK PV OR 14792; S. bollensis, SMNS 51563, MMG BwJ 565).
Prefrontal: The prefrontals in NHMUK PV OR 14781 are slightly sub-circular in shape, longer than wide in dorsal view (Figs. 2, 4), and ornamented with deep grooves. The prefrontal forms the anteromedial border of the orbit, and contacts the nasal and frontal medially and the lacrimal laterally (Figs. 2, 4). The prefrontal-frontal suture is irregular and the prefrontal-lacrimal contact is regular.
Lacrimal: The lacrimal in NHMUK PV OR 14781 is a large, triangular bone that constitutes the majority of the anterolateral margin of the orbits (Figs. 2, 4). The lacrimal is ornamented as in the other aforementioned bones. The paired antorbital fenestrae are large relative to the orbits, subcircular in shape and slightly anteroposteriorly elongated. The right antorbital fenestra is better preserved than the left, and it is almost equidistant between the orbit and alveolar margin, although it is still slightly closer to the orbit, which is similar in P. multiscrobiculatus (SMNS 9930), whereas in other teleosauroids (e.g., Steneosaurus leedsi, NHMUK PV R 3806) the antorbital fenestrae are much closer to the orbits (e.g., Andrews 1913). The anteroposterior length of the antorbital fenestrae is approximately 25% of the orbital anteroposterior length.
Frontal: The frontal in NHMUK PV OR 14781 is a large, dorsoventrally deep bone with no evidence of a midline suture (Figs. 2, 4). The frontal contributes to the posteromedial border of the orbit and forms the anterior medial borders of the supratemporal fenestra, as well as an almost straight vertical contact with the postorbital in dorsal and lateral views (Fig. 2A1, A2). The anterior process is short as in other teleosauroids (e.g., S. bollensis, SMNS 51753). The frontal is heavily ornamented with large pits and grooves that irradiate from the centre of the bone and extend to the anterior-most and lateral-most regions, similar to other Toarcian teleosauroids (e.g., P. multiscrobiculatus, SMNS 9930; S. bollensis, RE 551.762.130 A 0248; see SOM: fig. S2). The posterior-most portion of the frontal is not preserved.
Postorbital: The large, robust postorbital in NHMUK PV OR 14781 (Fig. 2A1, A2) reaches the posteroventral margin of the orbit (with the postorbital overlapping the jugal), and extensively forms part of the orbital ventral margin (similar to P. multiscrobiculatus SMNS 9930). The postorbital also forms the lateral and posteroventral margins of the supratemporal fenestra (Figs. 2, 4). The anterodorsal suture interdigitates tightly with the frontal and forms the posterior margin of the orbit. The postorbital is noticeably larger and more elongate than the squamosal, and the postorbital-squamosal contact is straight (best seen in lateral view). The anterodorsal area of the postorbital is slightly anteroposteriorly constricted (Fig. 2A1), whereas the rest of it is anteroposteriorly broad. The postorbital-jugal contact is difficult to discern due to the rugosities of the skull. The postorbital bar (formed by the frontal-postorbital contact) is similar to other teleosauroids (e.g., S. bollensis, SMNS 51753; Andrews 1913). It is slightly anteroposteriorly thickened, is ornamented by pits and grooves and forms the posterolateral margin of the orbit.
Squamosal: The right squamosal is preserved in NHMUK PV OR 14781 (Fig. 2A1), and is a large, elongate L-shaped bone (Fig. 4). The anterior process is anteroposteriorly elongated (in dorsal view) and forms the posterolateral border of the supratemporal fenestra. The posterolateral surface of the squamosal is concave in lateral view and convex in dorsal view (Fig. 2A2). The squamosal bar is robust and anteriorly contacts the postorbital bar (together forming the supratemporal arch). The anterior squamosal bar is ornamented with pits and grooves, as in the postorbital.
Quadratojugal: The anterior part of the right quadratojugal is preserved in lateral view in NHMUK PV OR 14781; it is a thin, elongate bone (Fig. 4).
Other cranial elements: In HLMD V946-948, remnants of the right palatine and possibly also the pterygoid are exposed in ventral view (Fig. 1A3), but the poor preservation precludes a detailed description. The palatal elements are unavailable in NHMUK PV R 14781. The parietals, prootics, laterosphenoids and quadrates are not preserved in either specimen and therefore cannot be described. In addition, the majority of bones representing the occipital are not preserved, and the basicranium is unavailable (as mentioned above).
Mandible: In HLMD V946-948, the mandible is partially preserved (Fig. 1A2, A4), with sections of the posterior dentaries and articulars missing. The anterior portion of the dentary is cemented to the cranium. In NHMUK PV OR 14781, the mandible is nearly complete on the right side and partially preserved on the left (Fig. 2A1, A3). It is likewise cemented to the cranium, so the dorsal surface is not visible, as mentioned above. In NHMUK PV OR 14781, the mandible measures approximately 870 mm in length (with the articular missing). The contact including the dentary, angular and splenial (best seen in right ventral view) is formed posterior to the mandibular symphysis on the ventral side. The posterior-most area of the mandible is partially covered with ironstone matrix.
Dentary: In HLMD V946-948, the anterior parts of both dentaries and the midsection of the right mandibular ramus are preserved (Fig. 1), but the midline suture is barely visible. Due to the occlusion with the skull, only the lateral and ventral sides are exposed. The dentary (Fig. 1A2) is a slender, elongate bone making up the majority of the lateral and ventral surfaces of the mandible, as seen in many crocodylomorphs (Andrews 1909, 1913; Romer 1956; Nesbitt 2011; Johnson et al. 2017). The dentaries are overhung and demarcated anteriorly by the premaxillae (Figs. 2A3, 3A), unlike the condition seen in Steneosaurus bollensis (Fig. 3C, D). A prominent notch is present that intersects the anterior margin of the conjoint dentaries. It is unclear whether this notch is a preservational artefact or a genuine morphology, but it is found in numerous teleosauroid species (e.g., Platysuchus multiscrobiculatus, UH 1; Steneosaurus bollensis, RE 551.762.130 A 0248, SMNS 18672, SMNS 53422; Steneosaurus leedsi, NHMUK PV R 3320; Steneosaurus edwardsi Eudes-Deslongchamps, 1868, NHMUK PV R 2076, NHMUK PV R 3701). The dentaries are transversely widened at the level of the premaxillary diastema. In HLMD V946-948, at the posterior part of the symphysis, the dentaries are dorsomedially demarcated by the splenials. Posterior to the symphysis, the coronoid extends medially to the dentary along its entire length. Ventrally, a short contact with the splenial is indicated on the left side. A long suture is present towards the angular, which separates the dentary from the splenial for most of its preserved length. In HLMD V946-948, the teeth are broken off or intersected, exposing the pulp cavity. In both specimens, the alveoli are relatively small and subcircular, with intermediate spacing throughout (the interalveolar distance being slightly larger than the alveolar labiolingual width, similar to S. gracilirostris MNHNL TU515). In HLMD V946-948, the right ramus bears remnants of 32 teeth. In NHMUK PV OR 14781, the exact number of alveoli is difficult to discern, but there are at least 30 alveolar pairs. The lateral margins of the dentaries are undulated.
Splenial: The splenials (Fig. 1A4) are exposed on the dorsal and ventral side of the mandible in HLMD V946-948. Dorsally the posterior splenial sections are visible; they are slightly indented and form the middle portion of the symphyseal rostrum. A long contact with the dentary is present laterally, with the suture being barely visible. A short, pointed process extends posterior to the symphysis and contacts the anteromedial side of the coronoid. Ventrally, the splenial forms the medial side of the preserved post-symphyseal mandibular ramus and has an elongated contact to the angular. In NHMUK PV OR 14781, only the right splenial is best seen in ventral view (Fig. 2A3).
Surangular and angular: In NHMUK PV OR 14781, the surangular (Fig. 2A1) is a thin and anteroposteriorly elongate bone in lateral view. It appears to form a distinctive V-shape (in conjunction with the angular and presumably the articular) and terminates near the final alveolus of the dentary. The posterior margin of the surangular is slightly rounded and encloses an irregularly shaped mandibular fenestra. In HLMD V946-948, part of the angular is preserved ventrally in the left mandibular ramus as an elongate, transversely thin process that interferes between the splenial and dentary (Fig. 1). It terminates slightly posterior to the symphysis. In NHMUK PV OR 14781, the angular (Fig. 2) occupies a larger area than the dorsally placed surangular. The angular is dorsoventrally deeper and more robust than the surangular and has a poor dorsal curvature in lateral view. Both elements extend slightly more posteriorly than the cranium.
Dentition: Both specimens (HLMD V946-948 and NHMUK PV OR 14781) have teeth preserved in situ. In HLMD V946-948, the pulp cavity is exposed in lateral view, as mentioned above. The maxillary and dentary teeth are large and elongate (Figs. 3A, 4). In NHMUK PV OR 14781, the anterior teeth have a more pointed apex than the posterior ones, which are gently rounded. The teeth are slightly more robust than those seen in other Toarcian teleosauroids (e.g., S. gracilirostris, MNHNL TU515, S. bollensis, SMNS 15172a, SMNS 54046; P. multiscrobiculatus, SMNS 9930). They are more distally curved in the anterior section of the maxillae and dentaries. The enamel ridges are small yet well developed, parallel to one another and reach the top of the apex. There are no serrations present.
Phylogenetic analysis
Methods.—We conducted a series of phylogenetic analyses to test the evolutionary relationships of Mystriosaurus laurillardi, specifically in relation to Steneosaurus bollensis, within Thalattosuchia using a modified version of the dataset provided by Ősi et al. (2018) and Foffa et al. (2019). The dataset consists of a merged matrix combining two datasets (now known as the Hastings + Young, or H+Y, matrix). The total dataset currently consists of 142 taxa (18 of which are teleosauroids) scored for 462 characters. The cladistic analysis of the dataset was conducted using TNT 1.5 Willi Hennig Society Edition (Goloboff et al. 2008; Goloboff and Catalano 2016; this programme has been made available with the sponsorship of the Willi Hennig Society), following the methodology used in Young et al. (2016). In the first analysis, NHMUK PV OR 14781 and HLMD V946-948 were used to score a single Mystriosaurus OTU. In the second analysis, NHMUK PV OR 14781 and HLMD V946-948 were treated as separate OTUs; lastly, in the third analysis we removed the Chinese teleosauroid, and kept NHMUK PV OR 14781 and HLMD V946-948 as separate OTUs.
Results.—The first analysis produced 197 most MPCs with 1513 steps (CI = 0.417, RI = 0.846, RCI = 0.353, HI = 0.584). The strict consensus topology (Fig. 5A) recovered Mystriosaurus laurillardi in a polytomy that includes the Chinese teleosauroid (IVPP V 10098), Platysuchus multiscrobiculatus, Bathysuchus megarhinus, Mycterosuchus nasutus and Teleosaurus cadomensis Lamaroux, 1820.
The second analysis produced 213 MPCs with 1514 steps (CI = 0.417, RI = 0.846, RCI = 0.353, HI = 0.584; Fig. 5B). Both specimens formed a polytomy with the Chinese teleosauroid (IVPP V 10098). These taxa formed a subclade within a polytomy including P. multiscrobiculatus, B. megarhinus, M. Nasutus, and T. cadomensis.
The third analysis (192 MPCs with 1508 steps, CI = 0.418, RI = 0.847, RCI = 0.354, HI = 0.582) recovered NHMUK PV OR 14781 and HLMD V946-948 as sister taxa (Fig. 5C). The overall picture of crocodylomorph interrelationships in all three analyses are similar to those found in previous iterations of this dataset (see Ristevski et al. 2018; Ősi et al. 2018; Foffa et al. 2019). Furthermore, in all three analyses, neither of the Mystriosaurus specimens formed a close relationship with S. bollensis.
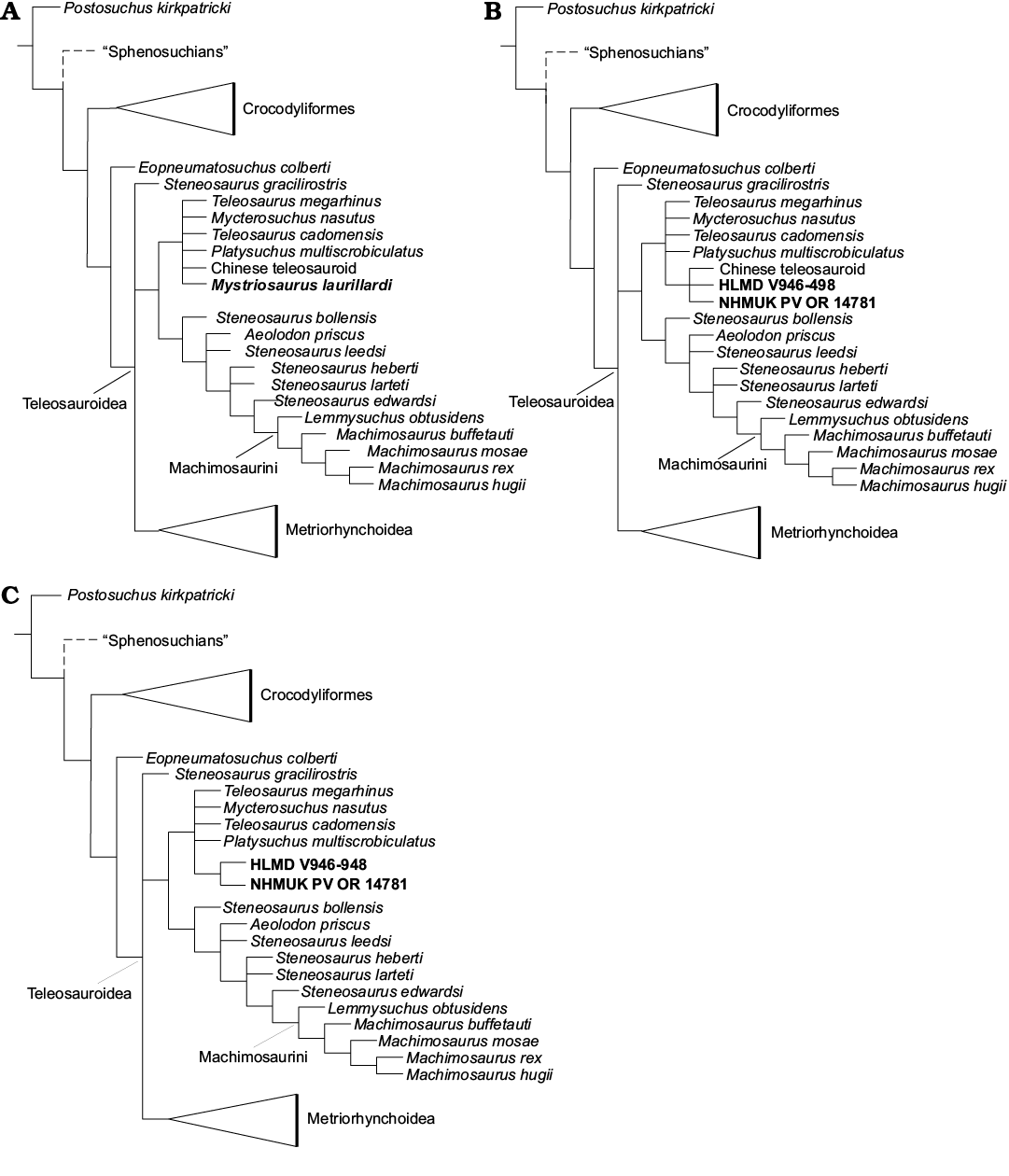
Fig. 5. Phylogenetic analyses using strict consensus trees of: A, the position of Mystriosaurus (197 MPCs, 1513 steps); B, HLMD V946-948 and NHMUK PV OR 14781 treated as two separate taxa (213 MPCs, 1514 steps); and C, HLMD V946-948 and NHMUK PV OR 14781 treated as two separate taxa and removal of the IVPP Chinese teleosauroid (192 MPCs, 1508 steps). For comparative skulls see SOM: fig. S3.
Discussion
Kaup (1837) distinguished Mystriosaurus from other fossil crocodylomorphs based on the following combination of characters: (i) long and cylindrical rostrum that curves upwards anteriorly; (ii) spoon-like skull and lower jaw expanding anteriorly; (iii) eight teeth in the conjoint premaxillae (four each); and (iv) elongated anterior portion of dentary, also with four pairs of teeth, followed by 28 additional dentary teeth per side.
Kaup in Bronn and Kaup (1841) distinguished Mystriosaurus laurillardi from other species of Mystriosaurus by these characters: (i) moderate rostrum; (ii) in the middle of the palate there is a penetrated, rim-shaped strip that is transversely intersected anterior to the palatal fenestrae; (iii) palatal fenestrae almost circular and much smaller; and (iv) an upper jaw dental formula of: 4 + 29 = 33 | 4 + 28 = 32, of which the last 6–7 are placed on top of the jaw.
In addition, Wagner (1850) provided these characters for M. laurillardi: (i) a cylindrical (“walzig”) rostrum; and (ii) alveolar edges at the same level with the palate. These characters, and in particular those listed by Kaup in Bronn and Kaup (1841), have been discussed in depth by Westphal (1962). The palatal fenestrae which Kaup in Bronn and Kaup (1841) named are in fact artefacts which probably result from an improper preparation. In addition, the rim-shaped strip in the middle of the palate likewise is an artefact. Westphal (1962) also noticed some peculiar features, such as the strongly ventrally deflected anterior ends of the premaxillae, that he considered as the most extreme case in S. bollensis (note that Westphal [1962] referred to Mystriosaurus as S. bollensis). Moreover, Westphal (1962) also considered the largest mediolateral width compared to the smallest mediolateral width of the premaxillae to be unique. Nonetheless, Westphal (1962) concluded that M. laurillardi is indistinguishable from a large individual of S. bollensis, and thus needed to be referred to that particular taxon.
Mystriosaurus laurillardi (HLMD V946-948 and NHMUK PV OR 14781) shares the following combination of characteristics with various other taxa: (i) mesorostrine skull (shared with the Chinese teleosauroid, IVPP V 10098; S. edwardsi, NHMUK PV R 2865, PETMG R178; and Machimosaurini); (ii) large and numerous foramina across the premaxillae, maxillae and dentaries (shared with Machimosaurini; e.g., Johnson et al. 2019); (iii) external nares oriented anteriorly (P. multiscrobiculatus, SMNS 9930; B. megarhinus, NHMUK PV OR 43086, unnumbered LPP specimen; T. cadomensis, Eudes-Deslongchamps 1867–1869; M. nasutus, CAMSM J.1420; and the Chinese teleosauroid, IVPP V 10098; all have the external nares oriented anterodorsally); (iv) anterior and anterolateral premaxillary margins are orientated anteroventrally and extend ventrally (shared with P. multiscrobiculatus, SMNS 9930; B. megarhinus, NHMUK PV OR 43086, unnumbered LPP specimen; T. cadomensis, Eudes-Deslongchamps 1867–1869; M. nasutus, CAMSM J.1420; and the Chinese teleosauroid, IVPP V 10098); (v) circular orbits (shared with M. nasutus, NHMUK PV R 2167; T. cadomensis, MNHN.F AC 8746; and Machimosaurini); (vi) four premaxillary alveoli (thalattosuchian symplesiomorphy); and (vii) 29–30 dentary alveoli (shared with S. bollensis, GPIT-RE-9425; S. edwardsi, NHMUK PV R 3701; and L. obtusidens, NHMUK PV R 3168).
However, it is interesting to note that, in our phylogenetic analyses, when HLMD V946-948 and NHMUK PV OR 14781 are treated as two separate OTUs, Mystriosaurus is found to be more closely related to the Chinese teleosauroid (IVPP V 10098; Fig. 5B) than any taxa from Europe.
As discussed above, M. laurillardi has previously been considered to be a subjective junior synonym of S. bollensis. However, multiple characters differentiate the two taxa: (i) M. laurillardi (NHMUK PV OR 14781) has a heavily ornamented and rugose skull, whereas the skulls of S. bollensis (e.g., MMG BwJ 565, RE 551.762.130 A 0248; SOM: fig. S2; SMNS 18672, SMNS 51563, SMNS 51753; Westphal 1962) are much less ornamented/rugose with shallower ornamentation (especially on the premaxillae, prefrontals and lacrimals, with little-to-none in the nasals and jugals; SOM: fig. S3); (ii) M. laurillardi (HLMD V946-948, NHMUK PV OR 14781) has two parallel lines of large, circular foramina across the maxillae, as well as a clustering of foramina around the anterior and lateral margins of the external nares. In S. bollensis (e.g., MMG BwJ 565, PMU R161, SMNS 51563), there is no clustering of the foramina (which are small), and only one line of foramina across the maxillae (SOM: fig. S4); (iii) M. laurillardi (NHMUK PV OR 14781) is mesorostrine with a mediolaterally broader rostrum, whereas S. bollensis (e.g., GPIT-RE-9427, MMG BwJ 565, SMNS 51753) is generally longirostrine with a more slender rostrum. The rostral length of M. laurillardi is approximately 66%, whereas S. bollensis specimens of equal size (e.g., GPIT-RE-9427) are 74–75%. However, it is important to note that M. laurillardi, NHMUK PV OR 14781, has a rostral length of approximately 580 mm (from the anterior premaxillae to the anterior rim of the orbits), whereas S. bollensis ranges between 391–902 mm; (iv) the external nares in M. laurillardi (HLMD V946-948, NHMUK PV OR 14781) are oriented anteriorly, whereas the external nares in S. bollensis (e.g., GPT-RE-9427, MMG BwJ 565, RE 551.762.130 A 0248) are oriented dorsally (Fig. 3; SOM: figs. S1, S2); (v) in M. laurillardi (HLMD V946-948, NHMUK PV OR 14781), the anterior and anterolateral premaxillary margins are orientated anteroventrally and extend ventrally (SOM: fig. S4A, B). In S. bollensis (e.g., SMNS 18672, RE 551.762.130 A 0248, SMNS 80235), the anterior and anterolateral premaxillary margins are not sub-vertical nor extend ventrally (SOM: fig. S4E); (vi) the supratemporal fenestrae in M. laurillardi (NHMUK PV OR 14781) are sub-rectangular in shape. In S. bollensis (e.g., MMG BwJ 565, PMU R161, SMNS 18672) the supratemporal fenestrae are more anteroposteriorly elongated than in Mystriosaurus (SOM: fig. S5); (vii) the anterolateral margins of the supratemporal fossae are inclined anterolaterally in M. laurillardi (NHMUK PV OR 14781), whereas S. bollensis (e.g., MMG BwJ 565, RE 551.762.130 A 0248, SMNS 51753) lacks this inclination (SOM: fig. S5); (viii) in dorsal view, the supratemporal arch seen in M. laurillardi (NHMUK PV OR 14781) is nearly straight (vertical), whereas in S. bollensis (e.g., RE 551.762.130 A 0248, SMNS 51563) it is noticeably more convex (SOM: figs. S2, S5); (ix) the orbits of M. laurillardi (NHMUK PV OR 14781) are circular in shape, whereas in S. bollensis (e.g., RE 551.762.130 A 0248, SMNS 51753, SMNS 18672) they are more elongate and anteroposteriorly ellipsoid (SOM: figs. S3, S5A, B); (x) in M. laurillardi (NHMUK PV OR 14781), the postorbital reaches the orbital posteroventral margin and extensively forms part of the orbital ventral margin. In S. bollensis (e.g., RE 551.762.130 A 0248, SMNS 51753), the postorbital is excluded from the posteroventral margin of the orbit; (xi) in M. laurillardi (NHMUK PV OR 14781), the angular is very slightly curved (nearly horizontal), whereas in S. bollensis (e.g., SMNS 18672) is it subtly and gently posterodorsally curved (SOM: figs. S1, S5); (xii) in M. laurillardi (NHMUK PV OR 14781), the mandibular symphysis measures less than half, but >0.33 of the total mandible length (between 0.3 and 0.45). In S. bollensis (SMNS 53422), the mandibular symphysis is just under half of the mandible length (between 0.45 and 0.5); (xiii) the external mandibular fenestrae are poorly elliptical in M. laurillardi (NHMUK PV OR 14781), whereas in S. bollensis (e.g., SMNS 51563, SMNS 51957) they are highly elliptical (SOM: figs. S1B, C, S5C, D); (xiv) in M. laurillardi (HLMD V946-948, NHMUK PV OR 14781), the maxillary teeth are subtly procumbent, whereas in S. bollensis (SMNS 53422, SMNS 51563) they are noticeably procumbent. However, the utility of this character is only present in specimens that have experienced little-to-no post-mortem crushing or shearing.
In addition, we note slight differences between NHMUK PV OR 14781 and HLMD V946-948: (i) the maxillary anterior alveoli shape is noticeably sub-oval in NHMUK PV R 14781, as opposed to slightly more oval in HLMD V 946-948; (ii) the interalveolar spacing between the D2–D3 is longer than the interalveolar spacing between the D1 and D2 in HLMD V946-948; (iii) the anterior maxillary tooth crown tip is slightly less pointed than those seen in NHMUK PV OR 14781; and (iv) the nasals are more heavily ornamented in NHMUK PV OR 14781. However, we currently do not consider these differences to be sufficient to establish two Mystriosaurus species, especially as the range of intraspecific variability, ontogenetic variation and sexual dimorphism is not known for this genus. Moreover, the skull roof is not well preserved in HLMD V946-948, thus the flatter and less ornamented nasal (compared to NHMUK PV OR 14781) could be a preservational artefact.
In addition, Mueller-Töwe (2006) assigned another complete yet slightly distorted skull, along with several associate vertebrae, from the Toarcian Posidonienschiefer Formation of Holzmaden (Germany) to “Steneosaurus” brevior (Fig. 6). This specimen (UH 7), which was described by Mueller-Töwe (2006), shares several characters with Mystriosaurus laurillardi. These include a robust skull with a mesorostrine snout, frontals that are ornamented by pits that fuse and expand into elongate grooves, subcircular shaped and dorsolaterally placed orbits, and large antorbital fenestrae. In addition, the anterior margin of the right supratemporal fenestra is placed more lateral than the lateral edge of the corresponding orbit. However, due to the distortion several key characters of Mystriosaurus remain unclear. This includes in particular the ventrally protruding anterior premaxillary portions and the anteriorly oriented external nares. Therefore, we tentatively identify this specimen as ?Mystriosaurus sp.

Fig. 6. Photograph of teleosauroid thalattosuchian specimen (UH 7), lower Toracian of Holzmaden (southwestern Germany), which was described by Mueller-Töwe (2006) as “Steneosaurus” brevior Blake, 1876, and which we herein refer to tentatively as ?Mystriosaurus sp.
Conclusions
The presented combination of characters clearly demonstrates that Mystriosaurus is a distinct and valid genus. Recognition of this taxon, and its presence in England and Germany, furthers our understanding of European teleosauroid biodiversity during the Toarcian. From lower Toarcian deposits of England, Luxembourg, and Germany, a diverse array of teleosauroids have been recovered: the lateral-orbited and small-bodied longirostrine species Steneosaurus gracilirostris; the large-bodied longirostrine form Steneosaurus bollensis; the mesorostrine and robust toothed taxon Mystriosaurus laurillardi, and the heavily armoured and longirostrine Platysuchus multiscrobiculatus (Westphal 1962; Johnson et al. 2018; Young and Steel in press). Although these taxa are found in many of the same formations, their geographical distributions need to be re-examined based on our developing understanding of their comparative anatomies. Nevertheless, our re-description of Mystriosaurus further highlights that teleosauroids had already achieved ecological diversity through niche partitioning, and large body-size by the early Toarcian.
Acknowledgements
We thank Matt Riley (CAMSM), Oliver Sandrock, Marisa Blume, and Mario Drobek (all HLMD), Xing Xu and Lizhao Zhang (both IVPP), Géraldine Garcia and Frank Guy (both LPP), Markus Wilmsen (MMG), Ronan Allain (MNHN), Ben Thuy and Robert Weis (both MNHNL), Susie Maidment (NHMUK), Glenys Wass (PETMG), and Rainer Schoch and Erin Maxwell (both SMNS) for access to specimens under their care. Thanks to Benjamin Kear (PMU) for providing photos of PMU R161, and to Achim Reisdorf, Rainer Rothenberg, and Ole Berndt (all RE) for photos of RE 551.762.130 A 0248. Julia Beier (Hamburg, Germany) is thanked for producing the reconstruction of the Mystriosaurus skull in Fig. 4. Stéphane Hua (Vauréal, France) provided some translations of old French texts. MMJ is supported by a Natural Sciences and Engineering Research Council of Canada grant (PGS D3-487581-2016) and an additional museum visit was funded by the Richard Owen Research Fund by the Palaeontographical Society. MTY is supported by a Leverhulme Trust Research Project grant (RPG-2017-167). Finally we thank the reviewers Attila Ösi and Stéphane Jouve for their useful comments which helped to improve the manuscript.
References
Adams-Tresman, S.M. 1987. The Callovian (Middle Jurassic) teleosaurid marine crocodiles from central England. Palaeontology 30: 195–206.
Allain, R. 2001. Redescription de Streptospondylus altdorfensis, le dinosaure théropode de Cuvier, du Jurassique de Normandie. Geodiversitas 23: 349–367.
Andrews, C.W. 1909. On some new steneosaurs from the Oxford Clay of Peterborough. Annual Magazine of Natural History 3: 299–308.Crossref
Andrews, C.W. 1913. A descriptive catalogue of the marine reptiles of the Oxford Clay, Part 2. 206 pp. British Museum (Natural History), London.
Berckhemer, F. 1929. Beiträge zur Kenntnis der Krokodilier des schwäbischen oberen Lias. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie. Beilagen Band 64B: 1–59.
Blake, J.F. 1876. Reptilia. In: R. Tate and J.F. Blake (eds.), The Yorkshire Lias, 243–254. John van Voorst, London.
Bronn, H.G. 1842. Über die fossilen Gaviale der Lias-Formation und der Oolithe. Archiv für Naturgeschichte 8: 77–82.
Bronn, H.G. and Kaup, J.J 1841. Abhandlungen über die gavialartigen Reptilien der Lias-Formation. 47 pp. E. Schweizerbart‘sche Verlagshandlung, Stuttgart.
Buffetaut, E. 1982. Radiation évolutive, paléoécologie et biogéographie des crocodiliens mésosuchiens. Mémoires de la Société Géologique de France (Nouvelle Série) 142 (60): 1–88.
Cuvier, G. 1801. Sur une nouvelle espèce de Crocodile fossile. Bulletin des Sciences, Société Philomathique de Paris 2: 159.
Cuvier, G. 1808. Sur les ossemens fossiles de Crocodiles. Annales du Muséum d’Histoire naturelle 12: 73–110.
Cuvier, G. 1825. Recherches sur les ossemens fossiles. 3ème édition. 547 pp. Dufour et d’Ocagne, Paris.
d´Alton, E. and Burmeister, H. 1854. Der fossile Gavial von Boll in Württemberg, mit Bezugnahme auf die lebenden Krokodilinen nach seiner gesammten Organisation zoologisch geschildert. 82 pp. C. Graeger, Halle.
de la Beche, H.T. 1832. Handbuch der Geognosie. 612 pp. Duncker und Humblot, Berlin.
Drevermann, F. 1914. Die Meersaurier im Senckenbergischen Museum. Bericht der senckenbergischen naturforschenden Gesellschaft 45 (Sonderheft): 35–48.
Eudes-Deslongchamps, E. 1863–1869. Prodrome des Téléosaurines du Calvados. Notes Paléontologiques 1: 95–354.
Eudes-Deslongchamps, E. 1867–1869. Notes Paléontologiques. 392 pp. F. le Blanc-Hardel and Savy, Caen.
Eudes-Deslongchamps, E. 1868. Note sur le squelette et la restauration du Teleosaurus cadomensis. Bulletin de la Société Linnéenne de Normandie 2e série 2 1868: 381–473.
Eudes-Deslongchamps, E. 1877. Le Jura Norman. 60 pp. F. Savy, Paris.
Fanti, F., Miyashita, T., Cantelli, L., Mnasri, F., Dridi, J., Contessi, M., and Cau, A. 2016. The largest thalattosuchian (Crocodylomorpha) supports teleosaurid survival across the Jurassic–Cretaceous boundary. Cretaceous Research 61: 263–274. Crossref
Faujas de Saint Fond, B. 1799. Histoire naturelle de la montagne St. Pierre de Maestricht. 184 pp. J. Jansen, Paris.
Foffa, D., Johnson, M.M., Young, M.T., Steel, L., and Brusatte, S.L. 2019. Revision of the Late Jurassic deep-water teleosauroid crocodylomorph Teleosaurus megarhinus Hulke, 1871 and evidence of pelagic adaptations in Teleosauroidea. PeerJ 7: e6646. Crossref
Foffa, D., Young, M.T., and Brusatte, S.L. 2015. Evidence of macrophagous teleosaurid crocodylomorphs in the Corallian Group (Oxfordian, Late Jurassic) of the UK. PeerJ 3: e1497.Crossref
Fraas, E. 1901. Die Meerkrokodile (Thalattosuchia n. g.) eine neue Sauriergruppe der Juraformation. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 57: Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 57: 409–418.
German Stratigraphic Commission 2016. Stratigraphic Table of Germany. Deutsches GeoForschungsZentrum, Potsdam.
Geoffroy Saint-Hilaire, E. 1831. Recherches sur de grands sauriens trouvés à l’état fossile aux confins maritimes de la Basse-Normandie, attribués d’abord au crocodile, puis déterminés sous les noms de Teleosaurus et Steneosaurus. Mémoires de l’Académie des Sciences 12: 1–138.
Giebel, C. 1847. Fauna der Vorwelt mit steter Berücksichtigung der lebenden Thiere. 511 pp. F.A. Brockhaus, Leipzig. Crossref
Glocker, E.F. 1839. Grundriss der Mineralogie, mit Einschluss der Geognosie und Petrefactenkunde. 993 pp. Schrag, Nürnberg.
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Hauff, B. 1921. Untersuchung der Fossilfundstätten von Holzmaden im Posidonienschiefer des Oberen Lias Württembergs. Palaeontographica 64: 1–42.
Hauff, B. and Hauff, R.B. 1981. Das Holzmadenbuch. 136 pp. Hauff-Stiftung, Holzmaden.
Hölder, H. 1955. Ein Jura-Krokodil mit Magensteinen. Aus der Heimat 63: 84–88.
Holl, F. 1829. Handbuch der Petrefactenkunde Part 1. 416 pp. Hilscher, Dresden.
Hua, S. 1999. Le crocodilien Machimosaurus mosae (Thalattosuchia, Teleosauridae) du Kimmeridgien du Boulonnais (Pas de Calais, France). Palaeontographica A252: 141–170.
Hua, S. and Buffetaut, E. 1997. Crocodylia. In: J.M. Callaway and E.L. Nicholls (eds.), Ancient Marine Reptiles, 357–374. Academic Press, San Diego. Crossref
Hulke, J.W. 1871. Note on a fragment of a teleosaurian snout from Kimmeridge Bay, Dorset. Quarterly Journal of the Geological Society, London 27: 442–443. Crossref
Jäger, G.F. 1828. Über die fossile Reptilien, welche in Württemberg aufgefunden worden sind. 48 pp. J.B. Metzler, Stuttgart. Crossref
Johnson, M.M., Young, M.T., and Brusatte, S.L. 2019. Re-description of two contemporaneous mesorostrine teleosauroids (Crocodylomorpha, Thalattosuchia) from the Bathonian of England, and insights into the early evolution of Machimosaurini. Zoological Journal of the Linnean Society zlz037 [published online, https://doi.org/10.1093/zoolinnean/zlz037] Crossref
Johnson, M.M., Young, M.T., Steel, L., Foffa, D., Smith, A.S., Hua, S., Havlik, P., Howlett, E.A., and Dyke, G. 2017. Re-description of “Steneosaurus” obtusidens Andrews, 1909, an unusual macrophagous teleosaurid crocodylomorph from the Middle Jurassic of England. Zoological Journal of the Linnean Society 1: 1–34. Crossref
Jouve, S. 2009. The skull of Teleosaurus cadomensis (Crocodylomorpha; Thalattosuchia), and phylogenetic analysis of Thalattosuchia. Journal of Vertebrate Paleontology 29: 88–102. Crossref
Jouve, S., Mennecart, B., Douteau, J., and Jalil, N.-E. 2017. Biases in the study of relationships between biodiversity dynamics and fluctuation of environmental conditions. Palaeontologica Electronica 20: 1–21. Crossref
Kaup, J.J. 1834. Verzeichniss der Gypsabgüsse von den ausgezeichnetsten urweltlichen Thierresten des Grossherzoglichen Museums zu Darmstadt. 28 pp. Diehl, Darmstadt.
Kaup, J.J. 1837. Mystriosaurus Kaup, Löffel-Saurier. In: G. Bronn (ed.), Leathaea Geognostica oder Abbildungen und Beschreibungen der für die Gebirgs-Formation bezeichnendsten Versteinerungen. Band 1. 544 pp. Schweizerbart, Stuttgart.
Kaup, J.J. 1841. Über Mystriosaurus-Reste aus dem Lias-Kalk von Altdorf. In: G. Bronn and J.J. Kaup (eds.), Abhandlungen über die gavial-artigen Reptilien der Lias-Formation. 47 pp. E. Schweizerbart‘sche Verlagshandlung, Stuttgart.
Krebs, B. 1967. Der Jura-Krokodilier Machimosaurus H. v. Meyer. Paläontologische Zeitschrift 41: 46–59. Crossref
Krüger, J.F. 1825. Urweltliche Naturgeschichte der organischen Teiche: In alphabetischer Ordnung. Vol. 1. 406 pp. Gottfried Basse, Quedlinburg.
Lamouroux, M. 1820. Sur le crocodile fossile trouvédans les carrières du bourg d’Allemagne, à un quart de lieue de Cean. Annales Générales des Sciences Physiques 3:160–164.
Li, J. 1993. A new specimen of Peipehuschus teleorhinus from Ziliujing Formation of Daxian, Sichuan. Vertebrata PalAsiatica 31: 85–94.
Lydekker, R. 1888. Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History). Vol. 1. 309 pp. British Museum (Natural History), London.
Mäuser, M. 2001. Altdorf bei Nürnberg. In: W.K. Weidert (ed.), Klassische Fundstellen der Paläontologie, 4, 97–110. Goldschneck, Korb.
Merck, J. 1786. Von dem Krokodil mit dem langen Schnabel, Crocodilus maxillis elongatis teretibus fubcylindricis Gronov. Hessische Beiträge zur Gelehrsamkeit und Kunst 5–7: 73–87.
Mook, C.C. 1934. The evolution and classification of the Crocodilia. The Journal of Geology 42: 295–304. Crossref
Mueller-Töwe, I.J. 2005. Phylogenetic relationships of the Thalattosuchia. Zitteliana A 45: 211–213.
Mueller-Töwe, I.J. 2006. Anatomy, Phylogeny, and Palaeoecology of the Basal Thalattosuchians (Mesoeucrocodylia) from the Liassic of Central Europe. 369 pp. Unpublished Ph.D. Thesis, Universität Mainz, Mainz.
Münster, G. 1834. Mitteilungen. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1834: 539.
Nesbitt, S.J. 2011. The early evolution of Archosauria: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Ősi, A., Young, M.T., Galácz, A., and Rabi, M. 2018. A new large-bodied thalattosuchian crocodyliform from the lower Jurassic (Toarcian) of Hungary, with further evidence of the mosaic acquisition of marine adaptations in Metriorhynchoidea. PeerJ 6: e4668. Crossref
Owen, R. 1884. A History of British Fossil Reptiles. Vol. 3. 199 pp. Cassel & Company, London.
Page, K. 2003. The Lower Jurassic of Europe: its subdivision and correlation. Geological Survey of Denmark and Greenland Bulletin 1: 23–59.
Pierer, H. 1827. Encyclopädisches Wörterbuch der Wissenschaften, Künste und Gewerbe, Band 8. 734 pp. Literatur Comptoir, Altenburg.
Pierce, S.E., Angielczyk, K.D., and Rayfield, E.J. 2009. Morphospace occupation in thalattosuchian crocodylomorphs: skull shape variation, species delineation and temporal patterns. Palaeontology 52: 1057–1097. Crossref
Ristevski, J., Young, M.T., Andrade, M.B., and Hastings, A.K. 2018. A new species of Anteophthalmosuchus (Crocodylomorpha, Goniopholididae) from the Lower Cretaceous of the Isle of Wight, United Kingdom, and a review of the genus. Cretaceous Research 84: 340–383. Crossref
Romer, A.S. 1956. Osteology of the Reptiles. 772 pp. University of Chicago Press, Chicago.
Schröter, J.S. 1780. Von dem vermeintlichen Kopfscelet eines Crocodills. Journal für die Liebhaber des Steinreichs und der Konchnliologie 6: 522–530.
Steel, R. 1973. Crocodylia. In: O. Kuhn (ed.), Encyclopedia of Palaeoherpetology. Teil 16. 116 pp. Gustav Fischer Verlag, Stuttgart.
Vignaud, P. 1995. Les Thalattosuchia, crocodiles marins du Mésozoique: Systématique phylogénétique, paléoécologie, biochronologie et implications paléogéographiques. 271 pp. Unpublished Ph.D. Thesis, Université de Poitiers, Poitiers.
Von Huene, F. 1956. Paläontologie und Phylogenie der Niederen Tetrapoden. 716 pp. Gustav Fischer, Jena.
Von Meyer, H. 1832. Palaeologica zur Geschichte der Erde und ihrer Geschöpfe. 506 pp. Schmerber, Frankfurt am Main.
Von Sömmerring, S.T. 1814. Über den Crocodilus priscus oder über ein in Baiern versteinert gefundenes Krokodil, Gavial der Vorwelt. Denkschriften der Königlichen Akademie der Wissenschaften zu München Classe 4: 1–74.
Wagner, K. 1835. Briefe an Johann Heinrich Merck von Göthe, Herder, Wieland und anderen bedeutenden Zeitgenossen. 528 pp. Diehl, Darmstadt.
Wagner, A. 1850. Die fossilen Überreste gavialartiger Saurier aus der Liasformation in der königlichen paläontologischen Sammlung zu München. Abhandlungen der Mathematisch-Physikalischen Klasse der Königlich Akademie der Wissenschaften 5: 511–606.
Walch, J.E.I. 1776. Von dem versteinerten Kopfscelet eines Crocodills. Der Naturforscher 10: 279–284.
Walkden, G.M., Fraser, N.C., and Muir, J. 1987. A new specimen of Steneosaurus (Mesosuchia, Crocodilia) from the Toarcian of the Yorkshire Coast. Proceeding of the Yorkshire Geological Society 46: 279–287. Crossref
Westphal, F. 1961. Zur Systematik der deutschen und englischen Lias- Krokodilier. Neues Jahrbuch für Geologie und Paläontologie 113: 207–218.
Westphal, F. 1962. Die krokodilier des Deutschen und Englischen oberen Lias. Palaeontographica A116: 23–118.
Wilberg, E.W. 2015. A new metriorhynchoid (Crocodylomorpha, Thalattosuchia) from the Middle Jurassic of Oregon and the evolutionary timing of marine adaptations in thalattosuchian crocodylomorphs. Journal of Vertebrate Paleontology 35: e902846. Crossref
Winkler, T.C. 1878. Etude sur le Genre Mystriosaurus et description de deux exemplaires nouveaux de ce genre. Archives du Museum Teyler 4: 49–132.
Woodward, A.S. 1885. On the literature and nomenclature of British fossil Crocodilia. Geological Magazine 3: 496–510. Crossref
Young, M.T. 2014. Filling the “Corallian Gap”: re-description of a metriorhynchid crocodylomorph from the Oxfordian (Late Jurassic) of Headington, England. Historical Biology 26: 80–90. Crossref
Young, M.T. and Andrade, M.B. 2009. What is Geosaurus? Redescription of Geosaurus giganteus (Thalattosuchia: Metriorhynchidae) from the Upper Jurassic of Bayern, Germany. Zoological Journal of the Linnean Society 157: 551–585. Crossref
Young, M.T. and Steel, L. (in press). Reptiles—crocodylomorphs. In: A. Lord and M. Munt (eds.), Fossils from the Lias of the Yorkshire Coast. Palaeontological Association Field Guide, London.
Young, M.T., Brusatte, S.L., Andrade, M.B., Desojo, J.B., Beatty, B.L., Steel, L., Fernández, M.S., Sakamoto, M., Ruiz-Omeñaca, J.I., and Schoch, R.R. 2012. The cranial osteology and feeding ecology of the metriorhynchid crocodylomorph genera Dakosaurus and Plesiosuchus from the Late Jurassic of Europe. PloS ONE 7: e44985. Crossref
Young, M.T., Brusatte, S.L., Ruta, M., and Andrade, M.B. 2010. The evolution of Metriorhynchoidea (Mesoeucrocodylia, Thalattosuchia): an integrated approach using geometricsgeometric morphometrics, analysis of disparity, and biomechanics. Zoological Journal of the Linnean Society 158: 801–859.
Young, M.T., Hastings, A.K., Allain, R., and Smith, T.J. 2016. Revision of the enigmatic crocodyliform Elosuchus felixi de Lapparent de Broin, 2002 from the Lower–Upper Cretaceous boundary of Niger: potential evidence for an early origin of the clade Dyrosauridae. Zoological Journal of the Linnean Society 179: 377–403. Crossref
Young, M.T., Hua, S., Steel, L., Foffa, D., Brusatte, S.L., Thüring, S., Mateus, O., Ruiz-Omeñaca, J.I., Havlik, P., Lepage, Y., and Andrade, M.B. 2014. Revision of the Late Jurassic teleosaurid genus Machimosaurus (Crocodylomorpha, Thalattosuchia). Royal Society Open Science 1: 140222. Crossref
Acta Palaeontol. Pol. 64 (3): 565–579, 2019
https://doi.org/10.4202/app.00557.2018