
Revealing body proportions of the enigmatic choristodere reptile Khurendukhosaurus from Mongolia
RYOKO MATSUMOTO, KHISHIGJAV TSOGTBAATAR, SHINOBU ISHIGAKI, CHINZORIG TSOGTBAATAR, ZORIG ENKHTAIVAN, and SUSAN E. EVANS
Matsumoto, R., Tsogtbaatar, K., Ishigaki, S., Tsogtbaatar, C., Enkhtaivan, Z., and Evans, S.E. 2019. Revealing body proportions of the enigmatic choristodere reptile Khurendukhosaurus from Mongolia. Acta Palaeontologica Polonica 64 (2): 363–377.
Khurendukhosaurus is an enigmatic genus of choristodere, recorded from the Lower Cretaceous of East Asia, Mongolia, and Siberian Russia. Until now, it was known only from isolated skull and postcranial elements, limiting comparison with other genera. Three major morphotypes have been recognised within Choristodera: longirostrine neochoristoderes with short-necks, and brevirostrine non-neochoristoderes with either short or long necks. The morphotype of Khurendukhosaurus was uncertain, although it had been inferred to be long-necked, based on cervical and caudal vertebral morphology shared with the Chinese Hyphalosaurus and on the results of phylogenetic analysis that placed it within a clade of Sino-Japanese long-necked taxa. Newly discovered material from the Mongolian type locality, Khuren-Dukh, preserves most major postcranial elements of a single individual. This specimen confirms that Khurendukhosaurus belongs to the long-necked morphotype, in having at least 13 cervical vertebrae. Moreover, a new phylogenetic analysis supports the placement of Khurendukhosaurus as a sister group of the Hyphalosaurus + Shokawa clade. Based on the new material, Khurendukhosaurus is estimated to have been roughly 1 m in total length, placing it at the upper end of the size range for long-necked choristoderes.
Key words: Reptilia, Choristodera, Khurendukhosaurus, Cretaceous, Mongolia, Khuren-Dukh.
Ryoko Matsumoto [r-matsumoto@nh.kanagawa-museum.jp], Kanagawa Prefectural Museum Natural History, 499 Iryuda, Odawara, Kanagawa, 250-0031, Japan.
Khishigjav Tsogtbaatar [tsogtbaatar@ipg.mn], Chinzorig Tsogtbaatar [chinzorig.tsogtbaatar@gmail.com], Zorig Enkhtaivan [zorig.enkhtaivan99@gmail.com], Institute of Paleontology and Geology, Mongolian Academy of Sciences, S. Danzan Street 3/1, Ulaanbaatar-15160, Mongolia.
Shinobu Ishigaki [isgk-7591@wind.email.ne.jp], Okayama University of Science, 1-1 Ridai-cho, Kita-ku, Okayama, 700-0005, Japan.
Susan E. Evans [ucgasue@ucl.ac.uk], Department of Cell and Developmental Biology, UCL, University College London, Gower Street, London WC1E 6BT, UK.
Received 21 October 2018, accepted 19 December 2018, available online 21 March 2019.
Copyright © 2019 R. Matsumoto et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The freshwater choristoderes were distributed throughout Laurasia from the Middle Jurassic to the Miocene, with a putative extension to Gondwana based on incomplete jaw elements, dorsal and caudal vertebral centra from North Africa (Haddoumi et al. 2016). Research over the last three decades has expanded knowledge of choristodere taxonomic diversity and distribution (e.g., Matsumoto and Evans 2010). To date, 12 genera and 25 species of choristoderes have been described (Matsumoto et al. 2019), and three primary morphotypes are recognized: short-necked longirostrine taxa, short-necked brevirostrine taxa, and long-necked brevirostrine taxa. The Early Cretaceous in Asia seems to have been an important time period in the evolutionary history of choristoderes, with all three morphotypes represented among 8 genera and 13 species, including Ikechosaurus (Sigogneau-Russell 1981; Brinkman and Dong 1993; Liu 2004) and Tchoiria (Efimov 1979; Ksepka et al. 2005), the first members of the derived Neochoristodera (sensu Evans and Hecht 1993).
Khurendukhosaurus orlovi, the type species of the genus, was first described in 1984 from the Early Cretaceous (Albian) of Khuren-Dukh, Mongolia, based on disarticulated postcranial elements (Sigogneau-Russell and Efimov 1984: table 1, PIN 3386/3). A second species, Khurendukhosaurus bajkalensis, was described based on a scapulocoracoid and rib from the Early Cretaceous (Barremian?–Aptian) Mogoito locality of Russia (Efimov 1996). However this species was later designated as indeterminate, due to a lack of diagnostic characters (Efimov and Storrs 2000). Further specimens have subsequently been found from the Murtoi Formation (Skutschas 2008; Skutschas and Vitenko 2017) including isolated skull elements (Table 1), and also from the Khuren-Dukh Formation (Matsumoto et al. 2009). These new specimens link this enigmatic taxon with other non-neochoristoderes (e.g., Matsumoto et al. 2009). A preliminary phylogenetic analysis placed Khurendukosaurus in an unresolved polytomy with Neochoristodera (Skutschas 2008), but subsequent analyses have weakly supported a relationship between Khurendukhosaurus and the long-necked Sino-Japanese Hyphalosauridae (Hyphalosaurus, Shokawa) (e.g., Matsumoto et al. 2009, 2019; Skutschas and Vitenko 2017).
Skutschas (2008) considered Khurendukhosaurus to be at least semi-terrestrial, based on closed neurocentral sutures, non-pachyostotic dorsal ribs, and a co-ossified scapula and coracoid. However, with the recovery of additional elements, new characters such as unfused sacral ribs, tall caudal neural spines from a deep swimming tail, and weak development of the distal humerus (Matsumoto et al. 2009) supported a largely aquatic lifestyle. This hypothesis was tested by a histological study of the humerus, rib, and gastralia that showed a specialised pachyosteosclerotic structure indicative of an aquatic lifestyle (Skutschas and Vitenko 2017). However Khurendukhosaurus remained incompletely known from a few partial elements belonging to different individuals. The recovery of new material from a single individual provides important new data that increases our understanding of the general morphology, body proportions and size of Khurendukhosaurus.
Institutional abbreviations.—BDL, Menat Museum Bord Du Lac, France; BMNHC, Beijing Museum of Natural History, Beijing, China; DR, Dalian Natural History Museum, Dalian, China; GMV, Geological Museum of China, vertebrate fossil collections, Beijing, China; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; LPMC, Liaoning Paleontological Museum of China, Shenyang, China; LMCCE, Laboratory of Mesozoic and Cenozoic Continental Ecosystems, Tomsk State University, Tomsk, Russia; MPC, Mongolian Paleontology Collection, Institute of Paleontology and Geology, Ulaanbaatar, Mongolia; PIN, A.A. Borissyak Palaeontological Institute, Russian Academy of Sciences, Moscow, Russia; ZIN PH, Paleoherpetological collection, Zoological Institute of the Russian Academy of Sciences, Saint Petersburg, Russia.
Other abbreviations.—MPT, most parsimonius tree; pobf, postorbitofrontal; TNT, Tree analysis using New Technology.
Geological setting, material
and methods
The Khurendukhosaurus specimens described here were collected from the Khuren-Dukh Formation in 1996 by a joint project team of the Institute of Paleontology and Geology (formerly the Mongolian Paleontology Center), Ulaanbaatar, and the Hayashibara Museum of Natural Sciences, Okayama, Japan. Isolated postcranial elements from the locality were described in 2009 (Matsumoto et al. 2009), but new specimens have subsequently been prepared from a single matrix block. All postcranial elements were closely associated and of similar size to one another, with no duplication of parts. We are therefore confident that they belong to a single individual. The block also contained an isolated postorbitofrontal, probably of the neochoristoderan Tchoiria that is known from the same locality (Efimov 1975) (Fig. 1). This postorbitofrontal is stout and L-shaped, with an elongated squamosal process. It is much larger than the postcranial elements (pobf; 90 mm in length) and clearly belonged to a different, unrelated, individual.
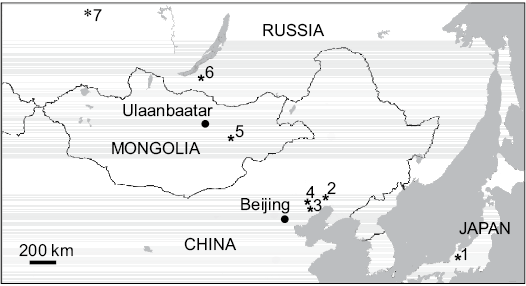
Fig. 1. Map showing Khuren-Dukh Formation, Mongolia and localities where long-necked choristoderes have been found. 1, Shokawa ikoi, Okurodani Formation, Barremian–Aptian (Evans and Manabe 1999); 2, Hyphalosaurus baitaigouensis, Baitaigou, Yixian Formation, Barremian–Aptian (Ji et al. 2004); 3, Hyphalosaurus sp., Luojiagou, Yixian Formation, Barremian–Aptian (Zhang et al. 2012; the Tianyu specimens with locality information are all from Badaoyingzi, Luojiagou); 4, Hyphalosaurus lingyuanensis, Dawangzhangzi, Yixian Formation, Barremian–Aptian (Fanzhangzi) (Gao et al. 1999; Gao and Ksepka, 2008); 5, Khurendukhosaurus orlovi and Khurendukhosaurus sp., Khuren-Dukh Formation, middle–late Albian (Sigogneau-Russell and Efimov 1984; Efimov and Storrs 2000); 6, Khurendukhosaurus sp., Mogoito, Murtoi Formation, Barremian?–Aptian (Efimov 1996; Skutschas 2008; Skutschas and Vitenko 2017); 7, cf. Khurendukhosaurus sp., Bol’shoi Kemchung 3, Ilek Formation, Early Cretaceous (Skutschas and Vitenko 2017).
The Khurendukhosaurus postcranial elements from the block include: seven cervical vertebrae; eight dorsal vertebrae; six caudal vertebrae; a cervical rib; nineteen dorsal ribs; several gastralia; the right and left humeri and radii; a scapulocoracoid and interclavicle; the right and left ilia, pubes, and ischia; the right and left femora, right tibia, and left fibula (Fig. 2).
The Khuren-Dukh Formation is considered to be middle–late Albian in age (Hicks et al. 1999; Nichols et al. 2006). The Formation is sub-divided into three layers that reflect changes in depositional environment, from a meandering river (lower member), through a lake, swamp, and flood plain system (middle member), to a river delta (upper member) (Ito et al. 2006; Sha et al. 2006). However, the Khurendukhosaurus specimens were collected before this environmental study was conducted. Based on the locality data, the material probably came from the lower member, but this remains uncertain.
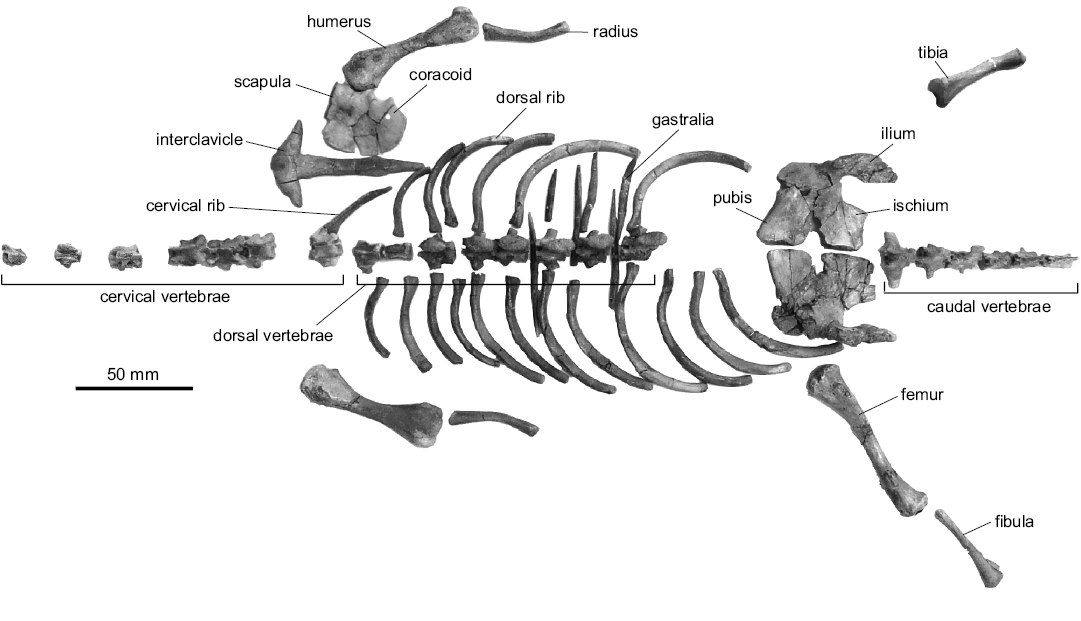
Fig. 2. All elements of the newly discovered specimen of the choristoderan reptile Khurendukhosaurus sp. (MPC-MX 1/107) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia. The bones are disarticulated but preserved in close association. This image shows the elements arranged in anatomical position.
Systematic palaeontology
Class Reptilia Linnaeus, 1758
Order Choristodera Cope, 1884
Family unknown
Genus Khurendukhosaurus Sigogneau-Russell and Efimov, 1984
Type species: Khurendukhosaurus orlovi (Sigogneau-Russell and Efimov, 1984); Albian, Khuren-Dukh Formation, Mongolia.
Emended diagnosis.—A medium-sized choristodere characterized by the following combination of characters: marginal tooth bases mediolaterally extended; basal tubera of the basioccipital moderately expanded laterally; cervical vertebral count at least 13; neural spines of cervical vertebrae short; zygapophyses nearly horizontal; neurocentral sutures closed; neural spines of dorsal vertebrae anteroposteriorly elongated with transversely expanded, rugose spine tables; tall narrow caudal vertebral neural spines; waisted ilium with an almost rectangular iliac blade patterned by coarse anteroposterior ridges.
Stratigraphic and geographic range.—Lower Cretaceous (Albian, Late Barremian–Middle Aptian), Mongolia and Russia.
Khurendukhosaurus sp.
Material.— MPC-MX 1/107, partial skull elements and postcranium from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia (see Table 1 for list of referred specimens and localities).
Table 1. Summary of described Khurendukhosaurus specimens from Mongolia (Khuren-Dukh locality) and Russia (Mogoito and Bol’shoi Kemchung 3 localities).
|
Locality |
Unit |
Stage |
Identity |
Material |
Specimen |
Reference |
|
Khuren- |
Khuren- |
Albian |
Khurendukhosaurus orlovi |
clavicle, interclavicle, coracoid, cervical and dorsal vertebrae, humerus, femur |
PIN 3386/3-1, 3386/3-2, 3386/3-3, 3386/3-4, 3386/3-5, 3386/3-6, 3386/3-7, 3386/3-8, 3386/3-13 (holotype), 3386/6-1-1, 3386/6-1-5, 3386/6-2-1, 3386/6-2-2, 3386/6-3 |
|
|
Khuren- |
Khuren- |
Albian |
Khurendukhosaurus orlovi |
cervical vertebra, dorsal vertebra, sacral vertebra, caudal vertebra, dorsal rib, 2nd sacral rib, ilium, femur, tibia |
MPC-MX1/103, 104, 105, 106 |
|
|
Khuren- |
Khuren- |
Albian |
Khurendukhosaurus sp. |
Seven cervical vertebrae, eight dorsal vertebrae, six caudal vertebrae, cervical rib, nineteen dorsal ribs, several gastralia, right and left humeri, right and left radii, scapulocoracoid, interclavicle, right and left ilia, right and left pubes, right and left ischia, right and left femora, right and left tibiae, right and left fibulae |
MPC-MX1/107 |
this paper |
|
Mogoito |
Murtoi |
Barremian? |
Khurendukhosaurus
|
scapulocoracoid, rib |
PIN 2234/201 (holotype) |
|
|
Mogoito |
Murtoi |
Barremian? |
Khurendukhosaurus sp. |
basioccipitals, exoccipital, partial maxilla, partial dentary,cervical vertebra, dorsal vertebra, sacral centrum, caudal vertebrae, dorsal rib, scapulocoracoid, clavicle, anterior part of interclavicle, humerus, radius, ulna, fibula, fragment of gastralia |
ZIN PH 1/25, 2/25, 3/25, 4/25, 8/25, 9/25, 10/25, 11/25, 12/25, 13/25, 14/25, 15/25, 16/25, 17/25, 18/25, 19/25, 20/25, 21/25, 22/25, 23/25, 24/25, 25/25; PIN 2234/201b |
|
|
Mogoito |
Murtoi |
Barremian? |
Khurendukhosaurus sp. |
postorbitofrontal, postorbital |
ZIN PH 26/25, 27/25 |
|
|
Bol’shoi Kemchung 3 |
Ilek |
Early |
Khurendukhosaurus sp. |
dorsal vertebra, sacral vertebra |
LMCCE 15/4, 21/4 |
Description.—Vertebrae: Seven cervical (excluding the atlas and axis), eight dorsal, and six caudal vertebrae are preserved in this specimen, MPC-MX 1/107 (Figs. 3–5). These vertebrae are characterized by their closed neurocentral sutures and relatively long (i.e., longer than high) amphiplatyan centra. The cervical vertebrae retain a notochordal pit on the posterior articular surface, but not on the anterior surface. Dorsal and caudal vertebrae vary in the presence of the pit, and some retain the pit at both ends of the centrum. Thus the appearance and position of the notochordal pit may vary within a single individual.
Of the seven cervical vertebrae preserved only three (Fig. 3D–F) are associated. Based on morphology, these may be from the mid-neck region. Although the total number of cervical vertebrae remains uncertain, this new specimen implies that Khurendukhosaurus had more than nine cervicals as the preserved elements do not include the atlas or axis. Three cervical vertebrae D–F (Fig. 3) may form a consecutive series, but there are gaps between the other cervical vertebrae that suggest missing elements, given the sharp rather than gradual changes in the size of the centrum articular surfaces, centrum length, and position, and size of rib para- and diapophyses. Thus the minimum estimated cervical number is thirteen: atlas+axis, anterior cervical A + (missing cervical CV4?) + anterior cervical B + (missing cervical CV6?) + anterior cervical C + (missing cervical CV8?) + middle cervical D–F + (missing cervical CV12?) + posterior cervical G). The rib diapophyses are almost horizontal in anterior cervicals, and become vertical in the posterior part of the neck. In addition, anterior cervicals have a strong midventral keel that gives the articular surface of the centrum an oval shape in anterior view. In the posterior cervicals (e.g., Fig. 3G1, G3), the centrum becomes almost circular with loss of the central keel. Several middle to posteriorly located cervical vertebrae (Fig. 3E2, F2, G2) bear small spinous processes below the postzygapophyses, without accessory facets (Fig. 3E2, F2, G2). Moreover, small accessory processes are developed between the posterior zygapophyses at the midline (Fig. 3E3, F3, G3), a feature also recognized in the dorsal vertebrae of the Russian Khurendukhosaurus (Skutschas 2008). The neural spines of the cervical vertebrae are low, and the spine table is narrower than that of the dorsal vertebrae. The angle enclosed by the anterior and/or posterior zygapophyses changes along the cervical vertebral column from 135° to 161° (Fig. 3C–G), indicating that the zygapophysial surfaces change from oblique to almost horizontal. The most posterior cervical vertebra (Fig. 3G) resembles dorsal vertebrae in having a dorsally located parapophysis close to the diapophysis (but not fused), and a centrum with a smooth ventral surface, and a circular articular surface.
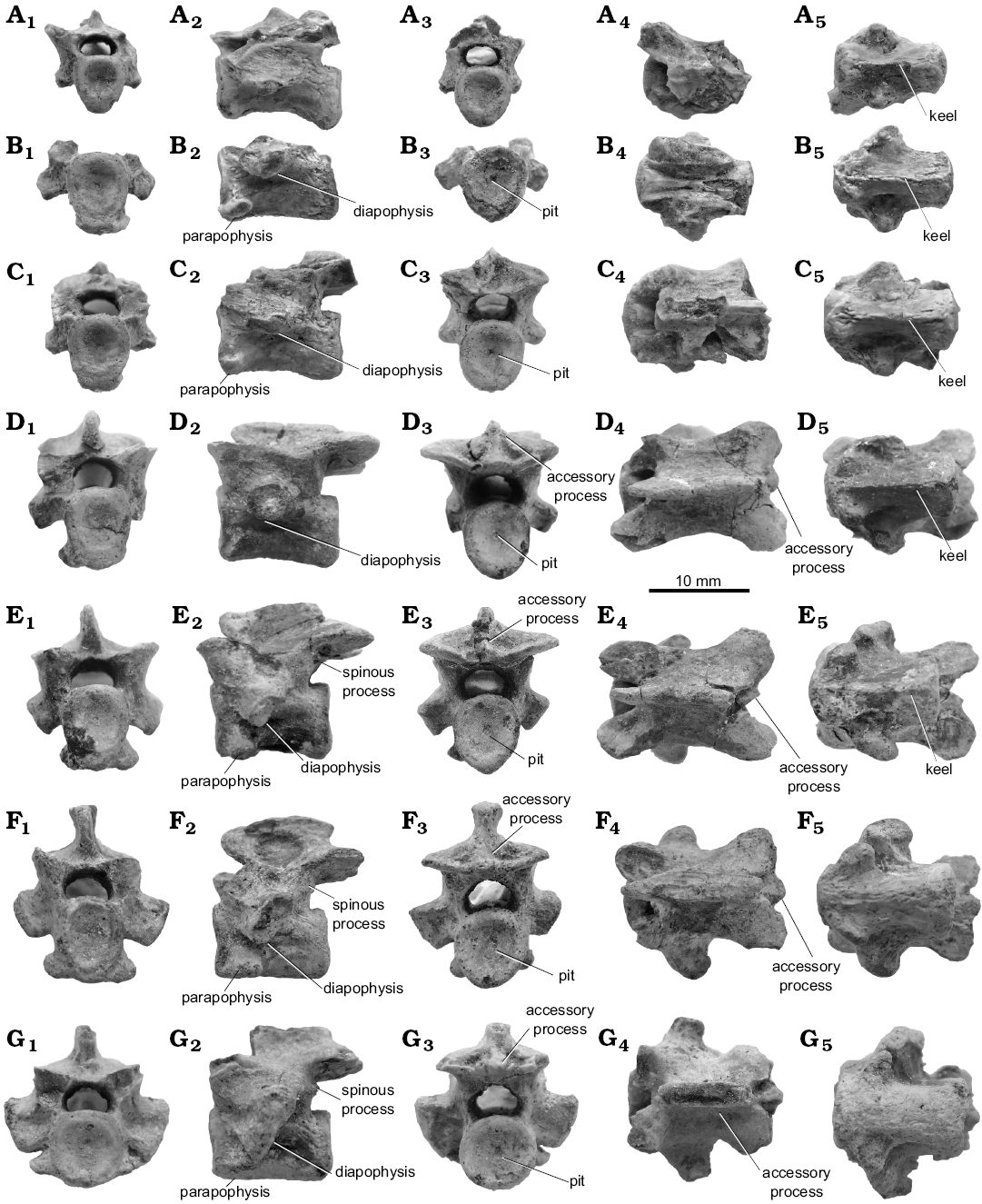
Fig. 3. Cervical vertebrae (A–G) of choristodere reptile Khurendukhosaurus sp. (MPC-MX 1/107) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia; in anterior (A1–G1), lateral (A2–G2), posterior (A3–G3), dorsal (A4–G4), and ventral (A5–G5) views. The precise anatomical order of these seven cervical vertebrae is uncertain, but vertebrae D, E, and F may be associated.
Choristoderes usually have sixteen dorsal vertebrae (e.g., Monjurosuchus, Philydrosaurus, Hyphalosaurus [some exceptions in H. baitaigouensis]; Gao et al. 2000; Gao and Fox 2005; Gao and Ksepka 2008). This new Khurendukhosaurus material includes six nearly complete dorsal vertebrae and two isolated centra (Fig. 4). Each centrum is longer than high, and has a rough ventral surface and circular articular surfaces. The notochordal canal is closed, but pits remain in some vertebrae as noted above. The neural arch and centrum are fused with no trace of a suture. The neural spines are rectangular in lateral view (longer than high), with spine tables that are swollen with a rugose dorsal surface, as previously recorded in Khurendukhosaurus (e.g., Skutschas 2008; Matsumoto et al. 2009) and in the Japanese long necked genus Shokawa (Evans and Manabe 1999). The neural spine angle varies between vertebrae, being vertical or inclined either anteriorly or posteriorly. The anterior margin of the neural spine is notched at the base (constricted spine), this space accommodating a small flange borne between the postzygapophyses of the preceding vertebra. This device effectively interlocks the dorsal vertebrae, limiting rotation. Ventral to the postzygapophyses some dorsal vertebrae bear small spinous processes without accessory facets (Fig. 4B2, E2). All dorsal vertebrae lack parapophyses, the rib articulation being limited to a synapophysis carried on a short transverse process. In lateral view, these transverse processes are seen to be inclined slightly posterodorsally. Posteroventral to the synapophysis some dorsal vertebrae bear a shallow fossa (Fig. 4). The enclosed zygapophyseal angle is roughly 90° (78–105°), more acute than in the cervical vertebrae. The trunk length of this individual is estimated as roughly ~200 mm, with a width of more than 100 mm.
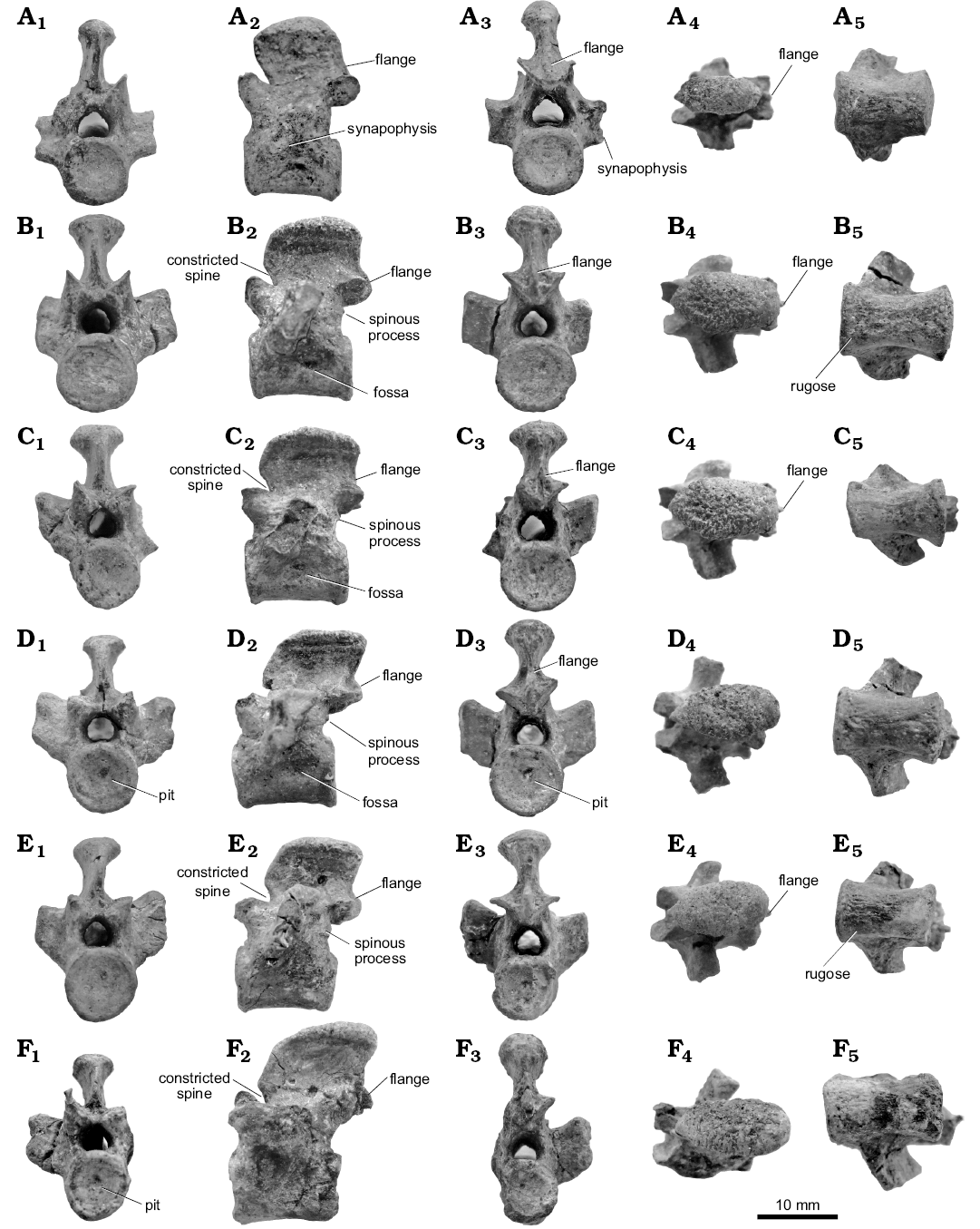
Fig. 4. Dorsal vertebrae (A–F) of choristodere reptile Khurendukhosaurus sp. (MPC-MX 1/107) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia; in anterior (A1–F1), lateral (A2–F2), posterior (A3–F3), dorsal (A4–F4), and ventral (A5–F5) views.
Six anterior–middle caudal vertebrae are preserved, three of which retain the neural spine and have a closed neurocentral suture (Fig. 5). The notochordal canal is closed but a pit remains on the anterior and/or posterior of the centrum (Fig. 5A, B, D, F). The anterior caudal vertebra (Fig. 5A) is distinguished from other caudal vertebrae in lacking grooves on the ventral surface of the centrum and it possesses weakly developed facets for the haemal arch on the posterior margin. This combination of features implies it is an anterior post-pygal vertebra. The neural spine is nearly vertical and is relatively shorter than those of subsequent caudal vertebrae. Its lateral surface bears a rough ridge that may indicate a line/limit of muscle attachment (Fig. 5A2). In subsequent caudal vertebrae (Fig. 5A, B), a vertical flange is developed between anterior and/or posterior zygapophyses. The spine tables are convex with rugosities, but they gradually become narrower further posteriorly. The centrum bears a ventral groove flanked by paired anteroposterior ridges and the attachments for the haemal arches straddle this groove at the posterior edge of the centrum. Where present, the caudal ribs are fused to the centrum, and they taper ventrolaterally at roughly 30° to the horizontal. The zygapophysial facets are almost vertical, with the posterior facet smaller than the anterior one.

Fig. 5. Caudal vertebrae (A–F) of choristodere reptile Khurendukhosaurus sp. (MPC-MX 1/107) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia; in anterior (A1–F1), lateral (A2–F2), posterior (A3–F3), dorsal (A4–F4), and ventral (A5–F5) views. Based on morphology, the vertebrae are arranged in an approximate proximal to distal sequence.
Ribs and gastralia: Cervical rib: One right cervical rib is preserved. At 34.48 mm, it is 2.8 times the length of an average cervical centrum (12.49 mm). The rib has two proximal heads: a figure-of-eight shaped facet for the diapophysis and a small rounded facet on the tuberculum for the parapophysis (Fig. 6A). This short rib is weakly curved posteriorly and has a sharp distal end. The tuberculum edge bears a well-developed crest for intercostal muscle attachment (Fig. 6A2), and there are additional keels along the dorsal and ventral margins. In Hyphalosaurus, anterior cervical ribs are no longer than the associated vertebral centrum, but they gradually increase in length to become more than twice centrum length at the level of the pectoral girdle (IVPP V11075, RM personal observation). These observations suggest that the isolated cervical rib in Khurendhukosaurus was from the posterior part of the neck.
Dorsal ribs: There are nineteen nearly complete dorsal ribs, seven from the right and twelve from the left, as well as fifteen rib fragments. The well-preserved ribs bear a single proximal head and rounded distal ends with a pitted surface indicating the presence of costal cartilages. Anterior dorsal ribs are roughly half the length of the posterior ribs (Fig. 6B1). The mid-shaft is slightly swollen and the rib then tapers towards its distal end (Fig. 6B1). The middle ribs are elongated with curvature around the mid-shaft (Fig. 6B2). Some of these dorsal ribs are of roughly similar diameter throughout the shaft, but several ribs are expanded at the distal tip. Long ribs bear an indentation on the posterodorsal surface of the mid-shaft (Fig. 6B3). A similar structure on the rib in the neochoristodere Simoedosaurus was described as a “scar” (Erickson 1987), and interpreted as an anchoring site for the gastral complex (Erickson 1987).
Gastralia: Each row of gastralia consists of at least three pieces (Fig. 6C). The median segment is almost straight (at least 52.64 mm in length) and has tapered ends that bear facets for lateral gastral elements (Fig. 6E). The lateral segment is shorter than the median part, and the medial facet is roughly half of its length (39.21 mm in total; facet 17.35 mm). The distal end of this component is gently curved and bears a weakly developed a keel along the posterior surface, as found in Champsosaurus (Erickson 1985). The facet surface is concave and accommodates a ridged facet on the median gastral element, locking the two together. Although the left lateral gastralia element is incomplete, the length of one set of associated gastralia can be estimated by reflecting the right side, giving a body width of about 103 mm for this individual Khurendukhosaurus.
A schematic section through the Khurendukhosaurus trunk is reconstructed based on middle or posterior ribs and nearly complete gastralia (Fig. 6C), and compared with a similar profile for Champsosaurus (modified from Erickson 1985; Fig. 6C). The distal ends of the dorsal ribs of Khurendukhosaurus are posteroventrally directed, forming a solid cage for the viscera. The lateral gastralia curve dorsolaterally to meet the rib cage, but the central element is flat giving a rather D-shaped cross-section (Fig. 6C). Champsosaurus has a more spindle-shaped trunk, formed by straighter dorsal ribs and with the central element of the gastralia moderately curved (Fig. 6D). However these shapes may change slightly along the trunk.
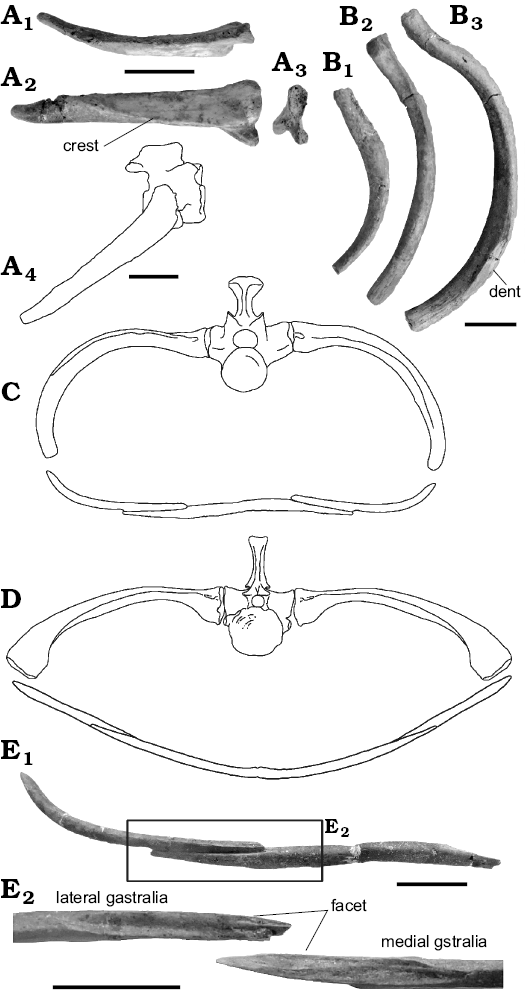
Fig. 6. Ribs and gastralia of choristodere reptile Khurendukhosaurus sp. (MPC-MX 1/107, A–C, E) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia and Champsosaurus gigas (D) from Late Paleocene, Wannagan Creek Quarry, USA. A. A cervical rib in dorsal (A1), anterior (A2), and medial (A3) views, reconstruction of cervical rib articulated with a cervical vertebra in lateral view (A4). B. Dorsal ribs (B1–B3) in anterior view. C, D. Sections through the trunk of dorsal vertebrae (number unknown) (C) and eighth dorsal vertebra (D, modified from Erickson 1985). E. Gastralia in anterior view (E1), with an enlargement of their articular surfaces (E2). Scale bars 10 mm. C, D not to scale.
Pectoral girdle and forelimb: The clavicles are not preserved, but the interclavicle is nearly complete (Fig. 7A). The proximal portion is rhomboid and the ventral surface is slightly convex at the centre (Fig. 7A1, A2). The clavicular facets are continuous across the midline, and the surface of each facet is ridged (Fig. 7A3–A5). The interclavicular stem is nearly twice the length of the proximal head. It is essentially parallel-sided, but the stem is weakly waisted around its midpoint, and it gradually tapers towards the distal end (Fig. 7A1–A3). On the ventral surface, the head is delineated from the shaft by the presence of a pair of distinct concavities that are separated in the midline by a weak keel that gradually reduces in height towards the posterior end (Fig. 7A1, A2). These concavities probably mark the attachment of paired pectoralis muscles. This keel is clear in the Russian specimen (Skutschas 2008).
Part of the right scapulocoracoid is preserved. The suture between the scapula and coracoid is visible on the dorsal surface, but it is obscured on the ventral side (Fig. 7B). The blade of the scapula is missing. The coracoid forms a rounded plate of smooth bone, but it increases in thickness laterally, close to the junction with the scapula. More than half part of the shallow glenoid cavity is formed by the coracoid (Fig. 7B4, B5). Lateral to the glenoid cavity, the scapula forms a rough, projecting supraglenoid tubercle. The infraglenoid tubercle is located medial to the glenoid cavity and comprises a dorsally facing rugosity (Fig. 7B4, B5). On the dorsal surface, a shallow concavity along the line of the scapulocoracoid suture might be for the sub-coracoideus muscle, as suggested by Sigogneau-Russell (1981) (Fig. 7B2, B3).
Both humeri are preserved (Fig. 7C, D). The proximal and distal heads are of roughly equal width. The mid-shaft is constricted, rhomboid in cross section, and the distal end is twisted roughly 45° in relation to the proximal head. The proximal condyle is dorsally expanded (Fig. 7C1) flanked by roughly symmetrical ecto- and entotuberosities that are separated from the central condyle by distinct notches (Fig. 7C2, D1). This morphology is also recognized in the problematic taxon Irenosaurus egloni (Efimov 1983), but the proximal head is relatively flat in the holotype of Khurendukhosaurus orlovi (Sigogneau-Russell and Efimov 1984) and in the more recently found Khurendukhosaurus sp. from Russia (Skutschas 2008). In ventral view, the shallow concavity between the humeral crest and the deltopectoral crest has a roughened surface for the attachment of the deltoid muscle (Fig. 7C2, D1). Below the crest, a weak ridge extends toward the ectocondyle, developing a roughened surface around the mid-shaft (Fig. 7C2, D1). The distal head is well ossified and bears identical ecto-, ento-, and entepicondyles (Fig. 7C3). Both an entepicondylar foramen and groove are absent. The shallow ectepicondylar groove for the radial nerve separates the ectocondyle and the ectepicondyle. Moreover the ectocondyle is expanded posterolaterally. The radial supracondylar ridge is developed along the distal one third of the humerus in posteroventral view, and presumably marks the origin of the brachio-radialis or extensor carpitalia.
Both radii are preserved, but the ulnae are missing (Fig. 7E, F). The radius is about 60% of the length of the humerus, and is strongly curved from the middle to the distal end of the shaft. The proximal head is wider than the distal head and bears an oval facet with a shallow concavity (Fig 7E1, E4). Two distinct crests, upper and lower, are recognized as described in the Russian species (Skutschas 2008). The upper crest lies on the posterolateral surface of the proximal shaft (Fig. 7E2, F1). The lower crest begins on the anterior two thirds of the shaft, and runs along the lateral margin toward the distal end (Fig. 7E). In Simoedosaurus, there is a single keel that may have separated the attachments of the brachio-radialis from more medially attaching muscles (Sigogneau-Russell 1981).
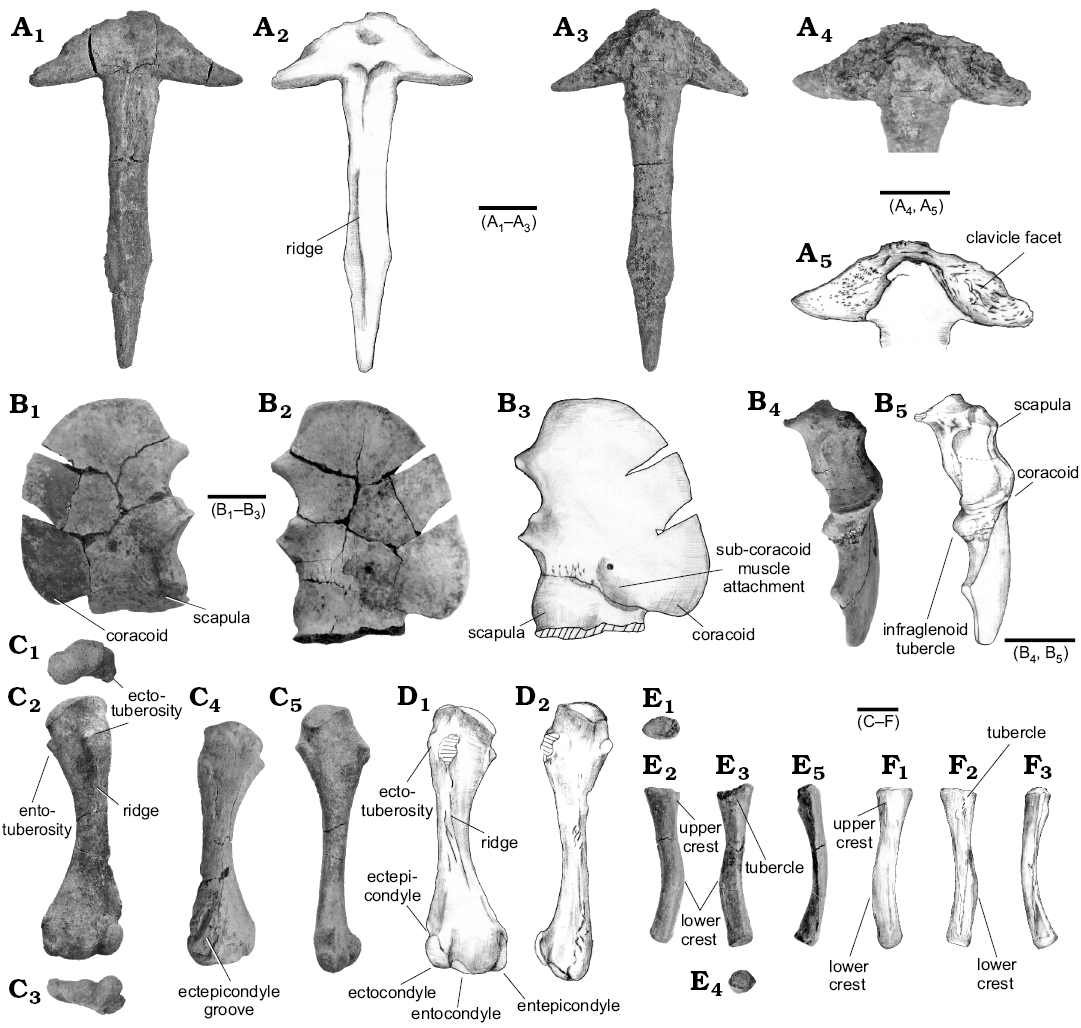
Fig. 7. Pectoral girdle and forelimb of choristodere reptile Khurendukhosaurus sp. (MPC-MX 1/107) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia. A. Interclavicle in ventral (A1, A2) and dorsal (A3) views, clavicle facets (A4, A5). B. Right scapulocoracoid in ventral (B1), dorsal (B2, B3), and lateral (B4–B5) views. C. Right humerus, proximal end in proximal view (C1) and dorsal (C2) views, distal end in distal view (C3) and ventral (C4) views. D. Left humerus in dorsal (D1) and ventral (D2) views. E. Right radius in proximal (E1), anterior (E2), posterior (E3), distal (E4), and lateral (E5) views. F. Left radius in anterior (F1), posterior (F2), and lateral (F3) views. Photographs (A1, A3, A4, B1, B2, B4, C, E); explanatory drawings (A2, A5, B3, B5, D, F). Scale bars 10 mm.
The pelvic girdle and hind limb: The three elements of the pelvic girdle (ilium, pubis, ischium) are articulated on both sides (Fig. 8A). The pubis and ischium of Khurendukhosaurus are described here for the first time. Sutures between these three elements are visible on the medial surface (Fig. 8A2). The dorsal edge of the iliac blade is probably damaged, and the original blade may have been rectangle in outline. The only comparable ilium of Khurendukhosaurus was found at the same locality as this new specimen (MPC-MX1/103; Matsumoto et al. 2009). These two ilia share similarities in the positions of muscle attachments, such as the small anterior tubercle for the iliotibialis muscle and a weak ridge on the dorsal acetabular margin for the iliofemoralis. Moreover the medial surface of the blade is ornamented with anteroposteriorly developed ridges, and bears two concavities for sacral rib articulations. The first facet lies along the anterodorsal edge of the blade. The oval concavity for sacral rib two is clear and is expanded anteroposteriorly along the ventral margin of the blade, but there is no obvious facet for a third sacral (Fig. 8A5, A6).
The pubis is a rectangular plate with rounded lateral and medial rims. A small obturator foramen perforates the plate (Fig. 8A2). The medial margin of the pubic plate is flat and is much thinner than the rest of the plate (Fig. 8A1, A2). The posterior margin is straight and is shorter than the anterior margin. It has a long articulation with the ischium. The anterior margin of the ischium is incomplete on both sides and the presence or absence of a thyroid fenestra cannot be confirmed on this specimen. Khurendukuhosaurus has a typical choristoderan ischium, forming a square plate with a sharp apex on the posterior margin. The ischium is slightly narrower than the pubis, but the ischial plate (5.5 mm) is thicker (2.7 mm) at the midline. The posterior margin of the ischium bears a weakly developed tubercle like that found in Lazarussuchus and Simoeodosaurus (RM personal observation).
Both femora are complete. They are slender bones in which the distal head is twisted on the proximal head by about 70°, as previously described in MPC-MX1/103 (Matsumoto et al. 2009). However the femur of the new specimen is shorter (76.7 mm) than that of MPC-MX1/103 (96.6 mm), and the shaft is somewhat straighter. In Monjurosuchus, the degree of sigmoid curvature of the femur increases through ontogeny, with the bone straight in juveniles and sigmoid in mature adults (Gao et al. 2000). Thus this difference between specimens of Khurendukhosaurus may be size/age related. The well-ossified proximal head is separated from the internal trochanter, which is at roughly 90° to the proximal head (Fig. 8B). The proximoventral and proximodorsal surfaces are rugose (Fig. 8B). The former is possibly the site of insertion of m. puboischiofemoralis externus and the latter for the puboischiofemoralis internus, as suggested for Simoedosaurus (Sigogneau-Russell 1981).
The femur is roughly 120% of the humeral length, and thus of similar proportions to that of other non-neochoristoderes: Hyphalosaurus, ~120–138% (holotype of H. lingyuaensis and H. sp. IVPP V 12696); Monjurosuchus, 130–140% (Gao et al. 2000); Philydrosaurus, 140% (Gao and Fox 2005); Lazarussuchus, 121% (Hecht 1992).
The tibia is slightly curved toward the fibula, and its proximal head is wider than the distal one (Fig. 8C). The anterior surface bears a ridge, as recognized in MPC-MX1/103, and in the lateral aspect a rugosity is apparent at the mid-shaft. The femur/ tibia proportion is 169% in the new specimen, which is the same as in MPC-MX1/103 (Matsumoto et al. 2009).
The fibula is of roughly similar length to the tibia and the shaft is straight (Fig. 8D). The proximal shaft is narrow with a rounded head (Fig. 8D1), whereas the distal end is expanded with a D-shaped profile in end view (Fig. 8D3). The shaft is flat on the lateral surface with an acute posterior edge. There are two distinctive ridges marking muscle attachments. One is on the medial surface of the proximal shaft (Fig. 8D2, D4), and the other runs along the medial edge of the mid-shaft (Fig. 8D5, D6).
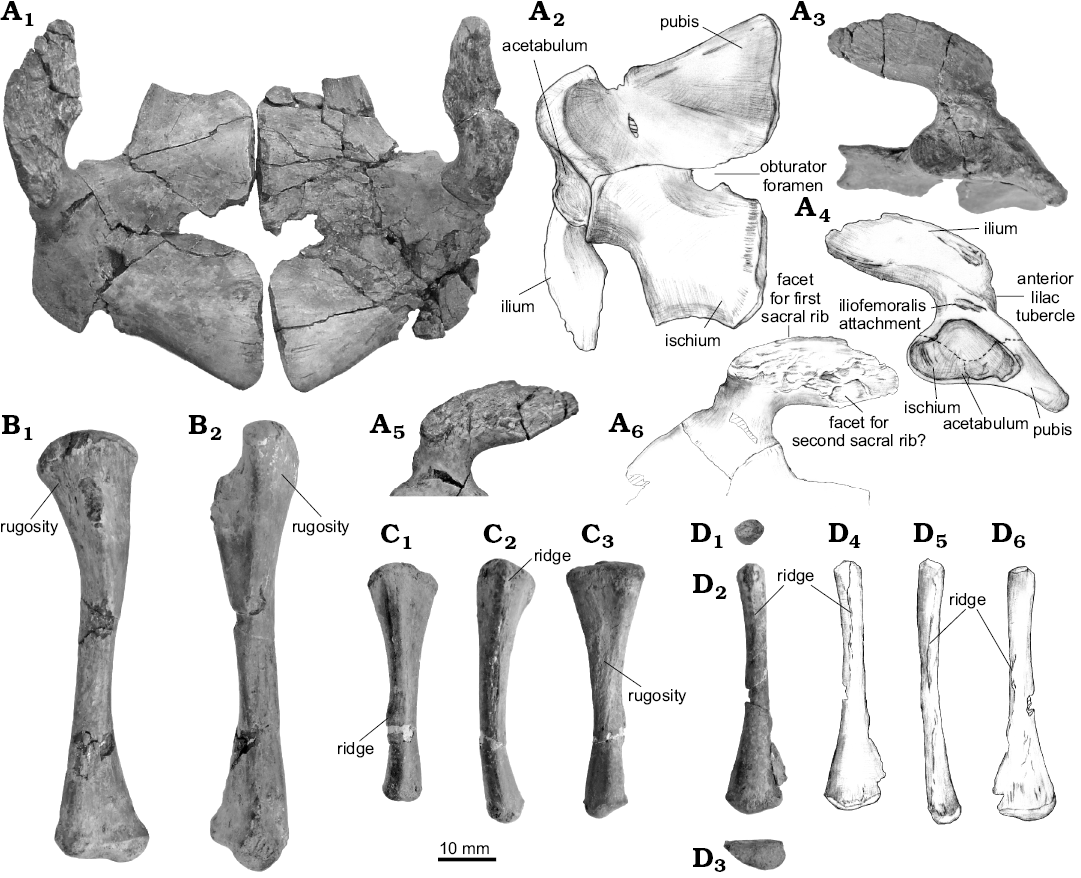
Fig. 8. Pelvic girdle and hind limbs of choristodere reptile Khurendukhosaurus sp. (MPC-MX 1/107) from middle–late Albian (Early Cretaceous), Khuren-Dukh, Mongolia. A. Associated ilium, pubis, and ischium in dorsal (A1), ventral (A2), lateral (A3, A4), and medial (A5, A6) views. B. Left femur in ventral (B1) and anterior (B2) views. C. Right tibia in lateral (C1), anterior (C2), and medial (C3) views. D. Left fibula in proximal (D1), medial (D2, D4), distal (D3), anterior (D4), and lateral (D5) views. Photographs (A1, A3, A5, B, C, D1–D3), explanatory drawings (A2, A4, A6, D4–D6).
Remarks.—The new specimen (MPC-MX 1/107) is comparable to the holotype of Khurendukhosaurus orlovi (PIN 3386/3) in the morphology of the scapulocoracoid, interclavicle, vertebrae and humerus. The fused scapulocoracoid closely matches that of the original specimens of K. orlovi. However, there are some minor differences in the vertebrae, humerus and interclavicle. In the original description (Sigogneau-Russell and Efimov 1984), the cervical and dorsal vertebrae were possibly, but not certainly, referred to Khurendukhosaurus. Later, Efimov and Storrs (2000) assigned these elements to the type series of the species. As described above, some cervical and dorsal vertebrae from the new Khuren-Dukh specimen (MPC-MX 1/107) bear small spinous processes below the posterior zygapophyses. These processes were absent in previously reported presacral vertebrae (Matsumoto et al. 2009), and there is no trace of the processes on the type vertebrae illustrated by Sigogneau-Russell and Efimov (1984). It is possible that the spinous processes occur only in limited positions along the vertebral column.
The holotype humerus (PIN 3386/3; Sigogneau-Russell and Efimov 1984) is larger than that of the new specimen (holotype 82 mm; MPC-MX 1/107, 64 mm in length), and the proximal and distal condyles are smoothly rounded. However, the humerus of MPC-MX 1/107 possesses paired tuberosities on the proximal head, and a well-ossified distal end with distinct ecto-, ento-, and entepicondyles. Comparable differences in the shape of the proximal head of the humerus have also been noted between similar sized specimens of Champsosaurus laramiensis (AMNH 982) and C. ambulator (Brown, 1905) (AMNH 983). Katsura (2004, 2007) suggested that this morphological variation could represent sexual dimorphism, but the significance of the variation in Khurendukhosaurus remains uncertain due to a lack of other comparable elements such as the femur and pectoral girdle in the Mongolian holotype and the Russian specimens (Skutschas 2008; Skutschas and Vitenko 2017).
The interclavicle of the holotype has a slender straight stem, whereas the stem of the similar-sized interclavicle of the new specimen is slightly waisted at the midshaft, and bears concavities along the midline.
Further comparison is possible with the referred Mongolian material of K. orlovi (MPC-MX1/103; Matsumoto et al. 2009) in the femur, tibia, and ilium. In MPC-MX 1/107 the femur and tibia are slightly smaller than those of MPC-MX1/103, but they are similar in their morphology. In addition the hind limb proportions (femur : tibia ratio 169%) are the same. The iliac blade is also slightly smaller in MPC-MX 1/107, but it differs in having a blade angle that is more horizontal than that of MPC-MX1/103. However these two ilia are similar in the positions of muscle attachment scars, such as the small anterior tubercle for the iliotibialis muscle and the weak ridge on the dorsal margin of the acetabulum for iliofemoralis. The iliac blade changes its morphology through ontogeny, the immature ilium being narrower and more angled than that of the adult in Champsosaurus (Erickson 1972). Moreover, in Hyphalosaurus, the blade shape varies from triangular (the large holotype of H. lingyuaensis) to square (some smaller specimens) (Matsumoto et al. 2009). Thus in Khurendukhosaurus iliac blades (MPC-MX 1/107 vs. 1/103) appear to change their angle through development, being horizontal in small individuals and vertical in larger ones, which is the opposite of the condition in Champsosaurus (Erickson 1972)
Thus the new Khurendukhosaurus specimens differ from previously described specimens from Khuren-Dukh (including the holotype specimen) in some features of the humerus (tuberosities on the proximal head and distal head) and ilium (blade angle). These differences are unlikely to result from ontogenetic variation, but further specimens are required to fully understand the variation with the genus.
Body size estimation: The new material confirms Khurendukhosaurus as a member of the long-necked choristodere morphotype, but it is problematic to estimate body size based on disarticulated material. Femora are often used for body size estimation in crocodiles (e.g., Farlow et al. 2005), but choristoderes vary in body proportions, especially in the skull and neck lengths. In contrast, the number of dorsal vertebrae is generally consistent at 16 in most choristoderes (e.g., Gao et al. 2000; Matsumoto et al. 2013), although there is some variation within Hyphalosaurus: 16 dorsal vertebrae in H. lingyuanensis, and 19 in H. bitaigouensis (Gao and Ksepka 2008). Articulated specimens of each morphotype were examined to estimate their body proportions: Ikechosaurus pijiagouensis (IVPP V 13283), Monjurosuchus splendens (IVPP V 14261, 13866; GMV 2135; DR003C; BMNHC V073), and Lazarussuchus inexpectatus (Hecht 1992) and Lazarussuchus sp. (Matsumoto et al. 2013) for the short-necked type; and Hyphalosaurus lingyuanensis and H. baitaigouensis (IVPP V17705; V12696; V12697; V14560; LPMC R-00065, 00066) for the long-necked type, including different ontogenetic stages. Trunk length was estimated from the average dorsal vertebral length and these values were compared to actual measurements of their trunk length (16 dorsal + 3 sacral vertebrae; SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app64-Matsumoto_etal_SOM.pdf). The error value was less than 10% between these two results, and so trunk length estimation based on dorsal vertebrae is considered to be valid. In long-necked Hyphalosaurus, the neck length is 80–130% of trunk length, whereas in short necked taxa (Lazarussuchus, Monjurosuchus, Ikechosaurus), the neck is only 20–50% of trunk length. The tail is generally more than twice the length of the trunk in both long-necked and short-necked morphotypes. Although an accurate cervical count is not possible in Khurendukhosaurus, its total body length (except skull) can be estimated from the average length of the eight dorsal vertebrae (i.e., 12.2 mm), and the trunk-neck and trunk-tail ratios of Hyphalosaurus. As mentioned before, cervical and dorsal vertebral numbers vary between H. lingyuanensis (19 cervical, 16 dorsal, 3 sacral vertebrae) and H. baitaigouensis (24 cervical, 19 dorsal, 3 sacral vertebrae). Each pattern was applied to Khurendukhosaurus, and the resulting estimates for total body length (without skull) were 1.02 m (based on H.linyuanensis) and 1.32 m (based on H. baitaigouensis). Thus if the new specimen of Khurendukhosaurus had similar proportions to Hyphalosaurus it may have been roughly 1.0 m in length. Moreover, previously described dorsal vertebrae of Khurendukhosaurus from the same locality (Matsumoto et al. 2009) are larger (15.5 mm in length), giving an estimated body length (excluding skull) of ~1.3 m (based on H. lingyuaensis) or 1.7 m (based on H. baitaigouensis). Thus, this preliminary study suggests that Khurendukhosaurus was a relatively large animal.
Discussion
The new Khurendukhosaurus specimen fills some missing character states in the data matrix of Matsumoto et al. (2019): cervical vertebrae count, 79 (cervical vertebral count more than 10 [1]); caudal rib shape, 102 (anterior caudal ribs straight but posterior ribs slightly curved [1]); posterior process of ischium, 115 (posterior margin develops tubercle [1]). The position of Choristodera within diapsids remains uncertain so that the selection of outgroup taxa is problematic. Moreover, the designation of outgroups has been shown to affect ingroup relationships within Choristodera (Matsumoto et al. 2009). The Late Carboniferous diapsid Petrolacosaurus (Reisz 1981) was designated as outgroup in this study (as TNT only allows one outgroup taxon), but a series of other taxa were included in the analysis to reflect varying hypotheses of relationship for choristoderes: Youngina and Araeoscelis (diapsid stem), Mesosaurus and Prolacerta (archosauromorphs), Nothosaurus and Keichousaurus (sauropterygians), and Gephyrosaurus (Lepidosauria). A second analysis used the Permian Araeoscelis (Reisz et al. 1984) as outgroup. These two analyses were run using TNT (version 1; Goloboff et al. 2008) in New Technology search mode with the Ratchet option (1000 random addition sequences: 50 iterations) followed by a Traditional search of trees in RAM for additional topologies. A Bootstrap analysis was run with 10 000 replicate resampling. The validity of Irenosaurus (Efimov 1988) and the choristoderan status of Pachystropheus (Storrs and Gower 1993) are problematic and these taxa were excluded in this analysis.
The two sets of TNT analysis (with Petrolacosaurus or Araeoscelis as outgroups) both yielded five MPTs (length = 329) with a similar overall topology. Thus the choice of Petrolacosaurus or Araeoscelis as outgroup had no effect. The basic topology of the strict consensus tree is similar to that of previous analyses (Matsumoto et al. 2019) in the monophyly of Choristodera and of Neochoristodera (Champsosaurus, Simoedosaurus, Ikechosaurus, Tchoiria), and the placement of the Middle Jurassic Cteniogenys as the sister taxon of all other Choristodera (Fig. 9). In addition, a new Late Jurassic Chinese choristodere (Matsumoto et al. 2019) is placed as the sister taxon of non-neochoristoderes (other than Cteniogenys). Thus this study confirms the positions of the new Late Jurassic Chinese choristodere and Cteniogenys. The main difference is a slightly better resolved position for Khurendukhosaurus in a monophyletic clade of long-necked species, the Hyphalosauridae (Hyphalosaurus + Shokawa). A previous study (e.g., Skutschas and Vitenko 2017) found the same topology in the 50% majority-rule consensus tree.
Khurendukhosaurus has been known from a very limited number of disarticulated postcranial elements (Table 1). Most choristoderan characters (64%) relate to the cranium, but the skull is poorly known in both Shokawa (lower jaw) and Khurendukhosaurus (partial maxilla, jaw, basioccipital, exoccipital, and postorbitofrontal). This is a major cause of the poor resolution of phylogenetic relationships between the long-necked choristoderan species. Khurendukhosaurus shares postcranial characters with the Asian Hyphalosauridae (Shokawa, Hyphalosaurus): horizontally expanded cervical zygapophyses; dorsoventrally expanded oval cervical centrum; deep groove developed on the ventral surface of caudal vertebrae. A wide rugose spine table is well developed in Shokawa, Khurendukhosaurus, and Hyphalosaurus (partially exposed in IVPP 14560). This structure is also present in Simoedosaurus and Champsosaurus but is less developed compared to the long-necked genera.

Fig. 9. Phylogenetic relationships of Khurendukhosaurus within Choristodera based on data from all available specimens. Strict consensus of five most parsimonious trees (MPTs) obtained by TNT (length = 329). Numbers in brackets indicate Bremer branch supports, followed by Bootstrap values. Abbreviations: CBU, Claude Bernard University no. Re 437, coll. Gennevaux 92813; IGM, Geological Institute of the Mongolian Academy of Sciences, Ulan Bataar, Mongolia; MNHN, Muséum National d’Histoire Naturelle, Paris; PKUP, Peking University Paleontological Collections, Beijing, China; SMM, Science Museum Minnesota, USA; TMP, Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, Canada.
Distribution of long-necked choristoderes.—Hyphalosaurus is one of the representative taxa of the Jehol biota, but is recorded only from the Yixian Formation (Barremian–Aptian; Zang and Gao 2013). The Japanese Shokawa is known from the Okurodani Formation (Barremian–Aptian; Kusuhashi et al. 2006), which is roughly equivalent in age to the Yixian Formation. Finally Khurendukhosaurus is known from Barremian?–Aptian/Albian localities in Mongolia and Russia (e.g., Skutschas 2008; Skutschas and Vitenko 2017). Thus these taxa were apparently limited to the Barremian–Albian interval. If the topology shown in the strict consensus tree is correct, then these long-necked taxa could have been derived from small lizard-like non-neochoristoderes in Asia during the Jurassic, before diversifying in the Early Cretaceous. However, all trace of these long-necked taxa had disappeared in Asia by the Late Cretaceous. Thus the geological range (Barremian?–Aptian/Albian) and geographical distribution (East Asia) of these long-necked choristoderes was apparently very limited. By contrast, although large neochoristoderes may have originated in Asia (Tchoiria, Ikechosaurus), they subsequently diversified in the Late Cretaceous–Paleocene of North America and Europe (Champsosaurus, Simoedosaurus), and small lizard-like taxa (Lazarussuchus) persisted as the very last lineage of choristoderes in the Paleocene–Miocene of Europe. Recently, a few isolated choristoderan vertebrae were found from shallow marine deposits of the Late Cretaceous Kuji Group, Northeast Japan (RM personal observation). The hiatus in the choristoderan record from the Late Cretaceous of Asia may be due to lack of deposits from temperate areas, as suggested by Matsumoto and Evans (2010). Further discoveries of choristoderan localities from both the Jurassic and Cretaceous of Asia will contribute to an increased understanding of their evolutionary history and distribution.
Conclusions
Our understanding of the enigmatic choristodere, Khurendukhosaurus, has gradually improved over the last decade. However, until now, the body proportions of this taxon remained uncertain due to a lack of specimens. New material of Khurendukhosaurus from the Mongolian type locality, Khuren-Dukh, represents an association of most major postcranial elements belonging to a single individual. This specimen confirms that Khurendukhosaurus had the long-necked choristoderan morphotype, with at least 13 cervical vertebrae. Phylogenetic analysis placed Khurendukhosaurus in a clade with other long-necked taxa (Hyphalosauridae). These long-necked genera probably arose from small lizard-like choristoderes in the Jurassic, but their record is currently limited to the Barremian?–Aptian/Albian of Asia.
Acknowledgements
RM and SEE thank Shigeru Suzuki (former Hayashibara Museum of Natural Sciences, Okayama, Japan), Makoto Manabe and Chisako Sakata (both National Museum of Nature and Sciences, Tokyo, Japan) for access to its collections in 2014, during and after special exhibition in 2013; and Pavel Skutschas (St Petersburg State University, Russia) for discussion and helpful information. We also thank Pavel Skutschas and Daniel Ksepka (Bruce Museum, Greenwich, CT, USA) for their comments on an earlier version of the manuscript. JSPS KAKENHI Grant number 26800269 allowed RM to visit Hayashibara Museum of Natural Sciences in 2014.
References
Brinkman, D.B. and Dong, Z. M. 1993. A new material of Ikechosaurus sunailinae (Reptilia: Choristodera) from the Early Cretaceous Langhongdong Formation, Ordos Basin, Inner Mongolia, and the interrelationships of the genus. Canadian Journal of Earth Sciences 30: 2153–2162. Crossref
Brown, B. 1905. The osteology of Champsosaurus Cope. American Museum of Natural History Memoirs 9: 1–26.
Cope, E.D. 1884. The Choristodera. American Naturalist 17: 815–817.
Efimov, M.B. 1975. Champsosaurs from the Lower Cretaceous of Mongolia [in Russian]. Trudy Sovmestnoj Sovetsko-Mongol’skoj Paleontologičeskoj Ekspedicji 2: 84–93.
Efimov, M.B. 1979. Tchoiria (Champsosauridae) from the Early Cretaceous of Khamaryn-Khural, MNR [in Russian]. Trudy Sovmestnoj Sovetsko-Mongol’skoj Paleontologičeskoj Ekspedicji 8: 56–57.
Efimov, M.B. 1983. Champsosaurs of Central Asia [in Russia]. Trudy Sovmestnoj Sovetsko-Mongol’skoj Paleontologičeskoj Ekspedicji 24: 67–75.
Efimov, M.B. 1988. The fossil crocodiles and champsosaurs of Mongolia and USSR [in Russian]. Trudy Sovmestnoj Sovetsko-Mongol’skoj Paleontologičeskoj Ekspedicji 36: 1–105.
Efimov, M.B. 1996. Champsosaurid from the Lower Cretaceous of Burytia [in Russian]. Paleontologičeskij žurnal 1996 (1): 122–123.
Efimov, M.B. and Storrs, G.W. 2000. Choristodera from the Lower Cretaceous of northern Asia. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 390–401. Cambridge University Press, Cambridge.
Erickson, B.R. 1972. The lepidosaurian reptile Champsosaurus in North America. Monograph, Science Museum of Minnesota (Paleontology) 1: 1–91.
Erickson, B.R. 1985. Aspects of some anatomical structures of Champsosaurus Cope (Reptilia: Eosuchia). Journal of Vertebrate Paleontology 5: 111–127. Crossref
Erickson, B.R. 1987. Simoedosaurus dakotensis, new species, a diapsid reptile (Archosauromorpha; Choristodera) from the Paleocene of North America. Journal of Vertebrate Paleontology 7: 237–251. Crossref
Evans, S.E. and Hecht, M.K. 1993. A history of an extinct reptilian clade, the Choristodera: longevity, Lazarus-taxa, and the fossil record. Evolutionary Biology 27: 323–338. Crossref
Evans, S.E. and Manabe, M. 1999. A choristoderan reptile from the Lower Cretaceous of Japan. Special Papers in Palaeontology 60: 101–119.
Farlow, J.O., Hurlburt, G.R., Elsey, R.M., Britton, A.R.C., and Langston, W. Jr. 2005. Femoral dimensions and body size of Alligator mississsippiensis: estimating the size of mesoeucrocodylians. Journal of Vertebrate Paleontology 25: 354–369. Crossref
Gao, K., and Fox, R.C. 2005. A new choristodere (Reptilia: Diapsida) from the Lower Cretaceous of western Liaoning Province, China, and phylogenetic relationships of Monjurosuchidae. Zoological Journal of the Linnean Society 145: 427–444. Crossref
Gao, K. and Ksepka, D.T. 2008. Osteology and taxonomic revision of Hyphalosaurus (Diapsida: Choristodera) from the Lower Cretaceous of Liaoning, China. Journal of Anatomy 212: 747–768. Crossref
Gao, K., Evans, S.E., Ji, Q., Norell, M., and Ji, S. 2000. Exceptional fossil material of a semi-aquatic reptile from China: the resolution of an enigma. Journal of Vertebrate Paleontology 20: 417–421. Crossref
Gao, K., Tang, Z., and Wang, X. 1999. A long-necked diapsid reptile from the Upper Jurassic/Lower Cretaceous of Liaoning Province, northeastern China. Vertebrata PalAsiatica 37: 1–8.
Goloboff, P., Farris, J., and Nixon, K. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Haddoumi, H., Allain, R., Meslouh, S., Metais, G., Monbaron, M., Pons, D., Rage, J.-C., Vullo, R., Zouhri, S., and Gheerbrant, E. 2016. Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): first continental flora and fauna including mammals from the Middle Jurassic of Africa. Gondwana Research 29: 290–319. Crossref
Hecht, M.K. 1992. A new choristodere (Reptilia, Diapsida) from the Oligocene of France: an example of the Lazarus effect. Geobios 25: 115–131. Crossref
Hicks, C.L., Webb, D.L., Nichols, D.J., and Watabe, M. 1999. Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia. Cretaceous Research 20: 829–850. Crossref
Ito, M., Matsukawa, M., Saito, S., and Nicholas, D.J. 2006. Facies architecture and paleohydrology of a synrift succession in the Lower Cretaceous Choyr Basin, southeastern Mongolia. Cretaceous Research 27: 226–240. Crossref
Ji, Q., Chen, W., Wang, W.L., Jin, X.C., Zhang, J.P., Liu, Y.Q., Zhang, H., Yao, P.Y., Ji, S.A., Yuan, C.X., Zhang, Y., and You, H.L. 2004. Mesozoic Jehol Biota of Western Liaoning, China [in Chinese]. 375 pp. Geological Publishing House, Beijing.
Katsura, Y. 2004. Sexual dimorphism in Champsosaurus (Diapsida, Choristodera). Lethaia 37: 245–253. Crossref
Katsura, Y. 2007. Fusion of sacrals and anatomy in Champsosaurus (Diapsida, Choristodera). Historical Biology 19: 263–271. Crossref
Kusuhashi, N., Matsumoto, A., Murakami, M., Tagami, T., Hirata, T., Iizuka, T., Handa, T., and Matsuoka, H. 2006. Zircon U-Pb ages from tuff beds of the upper Mesozoic Tetori Group in the Shokawa district, Gifu Prefecture, central Japan. The Island Arc 15: 378–390. Crossref
Ksepka, T.D., Gao, K., and Norell, A.M. 2005. A new Choristodera from the Cretaceous of Mongolia. American Museum Novitates 3648: 1–22. Crossref
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Salvius, Stockholm. Crossref
Liu, J. 2004. A nearly complete skeleton of Ikechosaurus pijiagouensis sp. nov. (Reptilia: Choristodera) form the Jiufotang Formation (Lower Cretaceous) of Liaoning, China. Vertebrata PalAsiatica 42: 120–129.
Matsumoto, R. and Evans, S.E. 2010. Choristoderes and the freshwater assemblages of Laurasia. Iberian Journal of Geology 36: 253–274. Crossref
Matsumoto, R., Dong, L., Wang, Y., and Evans. S.E. 2019. The first record of a nearly complete choristodere (Reptilia: Diapsida) from the Upper Jurassic of Hebei Province, China. Journal of Systematic Palaeontology [published online, https://doi.org/10.1080/14772019.2018.1494220]. Crossref
Matsumoto, R., Suzuki, S., Tsogtbaatar, K., and Evans, S.E. 2009. New material of the enigmatic reptile Khurendukhosaurus (Diapsida: Choristodera) from Mongolia. Naturwissenschaften 96: 233–242. Crossref
Matsumoto, R., Buffetaut, E., Escuillie, F., Hervet, S., and Evans S.E. 2013. New Material of the choristodere Lazarussuchus (Diapsida, Choristodera) from the Paleocene of France. Journal of Vertebrate Paleontology 33: 319–339. Crossref
Nichols, D.J., Matsukawa, M., and Ito, M. 2006. Palynology and age of some Cretaceous nonmarine deposits in Mongolia and China. Cretaceous Research 27: 241–251. Crossref
Reisz, R.R. 1981. A diapsid reptile from the Pennsylvanian of Kansas. University of Kansas Paleontological Contributions, Special Publication 7: 1–74. Crossref
Reisz, R.R., Berman, D.S., and Scott, D. 1984. The anatomy and relationships of the Lower Permian reptile Araeoscelis. Journal of Vertebrate Paleontology 4: 57–67. Crossref
Sha, J., Lin, L., Chen, S., and Matsukawa, M. 2006. Some Lower Cretaceous nonmarine bivalves from fluvio-lacustrine deposits bearing dinosaur fossils in Mongolia and northeast China. Cretaceous Research 27: 262–278. Crossref
Sigogneau-Russell, D. 1981. Présence d’un nouveau Champsosauridé dans le Cretace supérieur de Chine. Comptes rendus de l’Académie des Sciences 292: 1–3.
Sigogneau-Russell, D. and Efimov, M. 1984. Un Choristodera (Eosuchia?) insolite du Crétacé Inférieur de Mongolie. Palaeontologische Zeitschrift 58: 279–294. Crossref
Skutschas, P.P. 2008. A choristoderan reptile from the Lower Cretaceous of Transbaikalia. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 247: 63–78. Crossref
Skutschas, P.P. and Vitenko, D.D. 2017. Early Cretaceous choristoderes (Diapsida, Choristodera) from Siberia, Russia. Cretaceous Research 77: 79–92. Crossref
Storrs, G.W. and Gower, D.J. 1993. The earliest possible choristodere (Diapsida) and gaps in the fossil record of semi-aquatic reptiles. Journal of the Geological Society of London 150: 1103–1107. Crossref
Zang, W. and Gao, K. 2013. Early Cretaceous evolution of choristoderes in western Liaoning based on geographic and stratigraphic evidence. Journal of Palaeogeography 16: 205–216.
Zhang, L., Zhang, L., Yang, J., Guo, S., Wang, W., Zheng, Y., Ding, Q., and Cheng, L. 2012. Division of the Lower Cretaceous Yixian Formation and its ostracod fossil in Jianchang Basin, western Liaoning. Geology and Resources 21: 81–92.
Acta Palaeontol. Pol. 64 (2): 363–377, 2019
https://doi.org/10.4202/app.00561.2018