
Early steps in the radiation of notoungulate mammals in southern South America: A new henricosborniid from the Eocene of Patagonia
NICOLÁS BAUZÁ, JAVIER N. GELFO, and GUILLERMO M. LÓPEZ
Bauzá, N., Gelfo, J.N., and López, G.M. 2019. Early steps in the radiation of notoungulate mammals in southern South America: A new henricosborniid from the Eocene of Patagonia. Acta Palaeontologica Polonica 64 (3): 597–607.
Here we describe a new notoungulate mammal from the early Eocene (Itaboraian SALMA) of Chubut, Argentinian Patagonia, from the localities of Las Flores and Las Violetas Farm, represented by fragments of maxilla and isolated teeth. The specimens were found in the Las Flores Formation, Río Chico Group, and assigned to the Henricosborniidae, a primitive family within the order Notoungulata. Orome deepi gen. et. sp. nov. differs from other henricosborniids in a less developed metacone column in the upper molars, a larger metaloph in the third upper molar and a larger mean size. The morphometric analysis does not show any difference between the individuals of the two studied localities. The new addition to the henricosborniid diversity in Patagonia allows to provide a better understanding of the early radiation of notoungulates in South America.
Key words: Mammalia, Notoungulata, Paleogene, Eocene, Patagonia, Argentina.
Nicolás Bauzá [bauzan@fcnym.unlp.edu.ar], División Paleontología de Vertebrados, Museo de La Plata. Paseo del Bosque s/n B1900FWA, La Plata, Buenos Aires, Argentina and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina.
Javier N. Gelfo [jgelfo@fcnym.unlp.edu.ar], División Paleontología de Vertebrados, Museo de La Plata. Paseo del Bosque s/n B1900FWA, La Plata, Buenos Aires, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina; Facultad de Ciencias Naturales y Museo de La Plata, Universidad Nacional de La Plata, Buenos Aires, Argentina.
Guillermo M. López [glopez@fcnym.unlp.edu.ar], División Paleontología de Vertebrados, Museo de La Plata. Paseo del Bosque s/n B1900FWA, La Plata, Buenos Aires, Argentina; Facultad de Ciencias Naturales y Museo de La Plata, Universidad Nacional de La Plata, Buenos Aires, Argentina.
Received 6 November 2018, accepted 25 April 2019, available online 29 July 2019.
Copyright © 2019 N. Bauzá et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Henricosborniidae Ameghino, 1901 is one of the basal families of notoungulates, the most diversified order of extinct South American native ungulates (SAnu). By now, the species recognized for this family are characterized almost exclusively by dental remains, except for some cranial specimens of Simpsonotus from the Mealla Formation in Jujuy province, Argentina (Pascual et al. 1978), and tentative assigned postcranial remains from Paso del Sapo faunal assemblage (Tejedor et al. 2009; Lorente 2015). They are recorded from several localities of early Eocene Itaboraian South American Land Mammal Age (SALMA) in Brazil, at São José de Itaboraí, and in Argentina, in several outcrops in Patagonia (e.g., Simpson, 1948; Tejedor et al. 2009), in northwest area (Pascual et al. 1978; Babot et al. 2017), and the central Andean region (López 2008).
An isolated molar from the early Paleocene of Santa Lucía Formation in Tiupampa, Bolivia (Tiupampan SALMA) was doubtfully assigned to the Henricosborniidae (Muizon et al. 1984; Muizon 1991) and its phylogenetic relationships need to be confirmed. This scarcity of notoungulates in Tiupampa, in contrast to a rich and well preserved, faunistic association of marsupials, pantodonts, and mioclaenid kollpaniinaes, could be easily explained by a paleoecological bias.
Despite this record, henricosborniids suddenly thrived in the early Eocene in different latitudes of South America. There is no certainty about the cause of notoungulates absence in the Paleocene record of Patagonia. This could be explained due to the temporal gap since the early Paleocene units or by a later migration of the group to higher latitudes. Usually recognized genera include Henricosbornia, Peripantostylops, Othnielmarshia, and Simpsonotus (McKenna and Bell 1997), with Itaboraitherium, Paginula, and Acamana being taxa commonly associated with the family (López 2008). Currently the original materials that were used to create the species Acamana ambiguus are lost (Bond and Vucetich 1983).
Since Ameghino (1901) the content and relationships of Henricosborniidae have changed. Originally, Ameghino placed them within the Prosimiae, alongside the Hyopsodontidae Selenoconus (Ameghino, 1906). The last revision of the henricosborniids made by Simpson (1948) placed them, together with Notostylopidae and Arctostylopidae, in the suborder Notioprogonia, which was considered as the most primitive group of notoungulates; Simpson also identified Pantostylopidae and Selenoconus as junior synonyms of Henricosborniidae and Henricosbornia, respectively. Nowadays, the Arctostylopidae are considered as an independent radiation of mammals (Cifelli et al. 1989; Missiaen et al. 2012, but see Kondrashov and Lucas 2004), and the group Henricosborniidae–Notostylopidae is considered as paraphyletic. Still, the basal position of the henricosborniids is recovered in recent phylogenetic analyses, but due to their fragmentary record and the lack of postcranial characteristics, only a few genera (i.e., Henricosbornia and Simpsonotus) are usually used, recovering the family as paraphyletic (Billet 2011) or polyphyletic (Vera 2015).
In this work we describe a new genus and species of henricosborniid from the Las Flores Formation, Rio Chico Group in outcrops of two Patagonian localities, Las Flores, in the eastern side of Gran Barranca at the south of Colhué Huapi Lake, and Las Violetas Farm, closer to the shore and near the margin of the San Jorge basin (Raigemborn et. al. 2010). This new taxon indicates the presence of a new morphological type for the family, increasing their diversity and our knowledge of the importance of this group in the evolutionary history of notoungulates.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; CCMGE, Chernyshev’s Central Museum of Geological Exploration, Saint Petersburg, Russia; FMNH, Field Museum of Natural History, Chicago, USA; MACN, Museo Argentino de Ciencias Naturales, Buenos Aires, Argentina; MCZ, Museum of Comparative Zoology, Harvard College, Cambridge, Massachusetts, USA; MLP, Museo de La Plata, Argentina; MNHN-SCZ, Santa-Cruz collection of the Museum National d’Histoire Naturelle, Paris, France; ZIN, Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia.
Other abbreviations.—CVA, Canonical Variate Analysis; EECO, early Eocene Climatic Optimum; PCA, Principal Component Analysis; SALMA, South American Land Mammal Ages; SAnu, South American native ungulates. We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively. The number following the tooth abbreviation indicates locus position (e.g., M1 is a first upper molar, p2 a second lower premolar).
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:5B1AA50E-D2F3-4E52-BE59-4D284744D9AA
Material and methods
We compared the new remains with the holotypes and syntypes of the other henricosborniids, and materials referred to other families within the order Notoungulata and basal Paleocene Eutheria (see Appendix 1). In the case of Peripantostylops minutus a cast and photographs of complete upper and lower dental series (AMNH 28494) were used for the morphometric analysis.
The phylogenetic analyses were made with TNT version 1.5 (Goloboff et. al. 2008). The matrix utilized was modified from Billet (2011), in which we added a couple of discrete characters and six landmark configurations (see Appendix 1), one for each upper and lower molar, which we treated as continuous characters (Goloboff and Catalano 2016). Because our work is more specific than a phylogeny of the complete order Notoungulata, we excluded a great number of taxa from the original matrix, including only henricosborniids and a few examples of other families, just to get a preliminary glimpse of how Henricosborniidae relates to other groups of notoungulates. We also chose Zhelestes temirkazyk as our outgroup, since it is closer to our group of interest. Since the new remains are fragmentary and no complete dental series was recovered the implementation of a single landmark configuration for the geometric morphometric analysis was impossible. Photographs of the analyzed materials were compiled into .tps files with tpsUtil software (Rohlf 2004) and the landmarks were digitized with tpsDig software (Rohlf 2005). For the upper molars, the two-dimensional coordinates of eight landmarks were digitized over the occlusal surface. For the lower molars, a total of six two-dimensional coordinate landmarks were digitized. The landmark configurations were subjected to a Procrustes analysis (Rohlf and Slice 1990), in which the configurations are scaled to a centroid size of one, transposed and rotated, so that the sum of squared distances between corresponding landmarks is minimal. The geometric morphometric analyses were made with MorphoJ software (Klingenberg 2011), including a Principal Component Analysis (PCA) to visualize the major trends of shape variation and possible patterns of distribution of specimens along each factor, a Canonical Variate Analysis (CVA) to help emphasize those trends, and a regression test in order to discard the effects of allometry.
Due to the trend to form lophids in the dental morphology of the henricosborniids, and, the shear and wear of the occlusal surface of the teeth, it is not always possible to define a single point in each principal cusp to establish landmarks. We chose the maximum inflection points of the tooth outline, that is the labial end for labial cusp or structures, and the lingual end for lingual ones (Fig. 1). We discard those specimens where the wear was too high for the recognition of the main structures. In order to have the same number of landmarks in every structure, in the case of the last upper molar (i.e., M3) the sixth landmark was relocated to the lingual end of the metaloph. Landmarks 7 and 8 (in upper molars) were only considered in henricosborniids for the geometric morphometric analysis, not for the phylogenetic analysis.
Measures were taken manually with a Schwyz electronic digital caliper, considering maximum mesio-distal length and maximum labo-lingual width for each tooth.
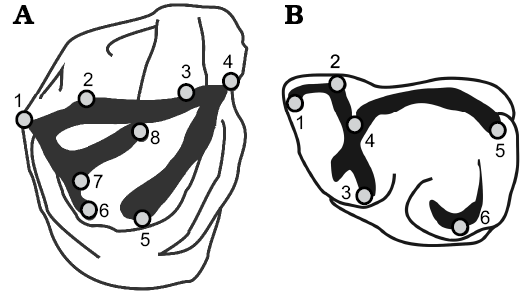
Fig. 1. Landmark configurations maped onto a hypothetical generalized henricosborniid. A. Upper molar: 1, metastyle; 2, metacone; 3, paracone; 4, parastyle; 5, protocone; 6, hypocone; 7, distolingual end of crochet; 8, mesiolabial end of crochet. B. Lower molar: 1, labial end of paralophid; 2, labial end of metalophid; 3, lingual end of metalophid; 4, mesial end of cristid obliqua; 5, hypoconulid; 6, entoconid/lingual end of entolophid.
Geographical and geological setting
The specimens of the new taxon here described came from the levels of Las Flores Formation, which is part of the Paleogene Rio Chico Group, together with the lower levels of the Las Violetas and Peñas Coloradas formations and, the upper Koluel-Kaike Formation (Raigemborn et al. 2010). The unit has its greatest thickness in the central area of the Golfo San Jorge basin in Chubut Province, where localities like Cañadón Hondo present a thickness of 57 meters, decreasing from east to west. The specimens here described came from two localities where Las Flores Formation outcrops. The first belongs to the type section of the unit placed in the east side of Gran Barranca at the south of Colhué Huapi lake, at the homonym locality, and with a thickness of 44 meters.
The other locality belongs to the outcrops of Las Flores Formation in the Las Violetas Farm, located around 150 km NE from Las Flores, near the national route 3. In Las Violetas Farm, several isolated exposures of the Las Flores Formation, no more than 15 meters thick, unconformably overlie Las Violetas Formation (Raigemborn et al. 2010).
Systematic palaeontology
Mammalia
Order Notoungulata Roth, 1903
Family Henricosborniidae Ameghino, 1901
Genus Orome nov.
ZooBank LSID: urn:lsid:zoobank.org:act:3B4338D2-F48F-411C-9370- 9F48F198BA5C
Etymology: Named after Oromë, one of the Valar in J.R.R. Tolkien’s The Silmarillion.
Type species: Orome deepi sp. nov.; monotypic, see below.
Diagnosis.—Same as for the only described species.
Stratigraphic and geographic range.—Las Flores Formation (Río Chico Group) from Las Flores and Las Violetas Farm localities, Chubut Province, Argentina (Fig. 2); Ypresian (early Eocene), Itaboraian SALMA (sensu Woodburne et. al. 2014).

Fig. 2. Geochronology of SALMAs and litostratigraphic units mentioned in the text modified from Gelfo et al. (2009) and Krause et al. (2017). The formations outcrop in different localities: 1, Northwest Patagonia, Argentina; 2, Tiupampa, Bolivia; 3, San Jorge Basin, Patagonia, Argentina. Abbreviations: C., Cocatherium; Fm., Formation.
Orome deepi sp. nov.
Fig. 3.
ZooBank LSID: urn:lsid:zoobank.org:act:BCE56DA5-4984-4452-B9F2- 1197BD865176
Etymology: Named after the British rock band Deep Purple, and in allusion to Las Violetas Farm, the locality in which the holotype was found (Spanish violeta, purple).
Holotype: MLP-90-II-5-1631, fragment of left maxilla with dP4–M1 (Fig. 3).
Type locality: Las Violetas Farm locality (45°02’00.3”S 66°59’25.6”W), Chubut Province, Argentina.
Type horizon: Ypresian, early Eocene, Las Flores Formation, Río Chico Group, Itaboraian SALMA (sensu Woodburne et. al. 2014).
Material.—MLP-90-II-5-1632, fragment of left maxilla with M1–M2; MLP-90-II-5-1633, fragment of right maxilla with M1–M2; and MLP-90-II-5-1634, fragment of right maxilla with M1–M2 from type locality and horizon. MLP-79-I-5-47, isolated right M3; MLP-90-II-5-1635, isolated left M1; MLP-90-II-5-1636, isolated left M1, MLP-90-II-5-1637, fragment of left maxilla with broken P4, M1 and M2; and MLP-90-II-5-1638, fragment of left maxilla with broken M1 and M2, from Las Flores locality, Chubut Province, Argentina; Ypresian (early Eocene), Las Flores Formation (Río Chico Group), Itaboraian SALMA.
Diagnosis.—Medium-sized henricosborniid (see dimensions in Table 1) comparable to Simpsonotus praecursor in size. Molars with metacone column weaker than in Henricosbornia, but stronger than in Notostylops. The middle portion of the ectoloph is slightly convex to straight in occlusal view. M3 with metaloph longer than other henricosborniids, reaching the protocone.
Table 1. Size comparison and mean dimensions (in mm) of Orome deepi gen. et sp. nov. from Las Violetas Farm and Las Flores.
|
Locality |
Specimen |
Locus |
Length |
Width |
|
Las Violetas |
MLP-90-II-5-1631 |
dP4 |
5.57 |
6.38 |
|
M1 |
6.62 |
8.78 |
||
|
MLP-90-II-5-1632 |
M2 |
7.16 |
8.86 |
|
|
MLP-90-II-5-1633 |
M1 |
4.67 |
6.01 |
|
|
M2 |
5.3 |
6.36 |
||
|
MLP-90-II-5-1634 |
M2 |
6.16 |
7.18 |
|
|
Las Flores |
MLP-90-II-5-1635 |
M1 |
5.91 |
7.66 |
|
MLP-90-II-5-1636 |
M2 |
6.12 |
7.08 |
|
|
MLP-90-II-5-1637 |
M1 |
4.68 |
6.56 |
|
|
MLP-90-II-5-1638 |
M2 |
5.69 |
7.34 |
|
|
MLP-79-I-5-47 |
M3 |
4.57 |
5.89 |
|
|
Orome deepi (mean) |
dP4 |
5.57 |
6.38 |
|
|
M1 |
5.47 |
7.2525 |
||
|
M2 |
6.086 |
7.364 |
||
|
M3 |
4.57 |
5.89 |
||
Description.—The dP4 (MLP-90-II-5-1631; Fig. 3) presents an advanced wear and, in occlusal view, is trapezoidal in shape and molarized. The protocone and the hypocone are joined at the base and originating a central fossa open lingually. As a result of wear, the crochet reaches the protoloph and the distolabial fossette is shallow and oval in outline. In labial view the ectoloph is bulbous, curved inwards in the medial region and shows parastyle paracone and metacone columns well developed. The mesial and distal cinguli are weak.
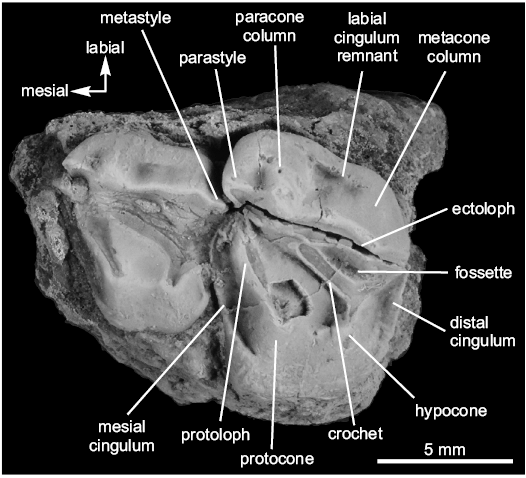
Fig. 3. Holotype of notoungulate mammal Orome deepi gen. et sp. nov. (MLP-90-II-5-1631) from the early Eocene of Las Violetas Farm, Argentina. Left dP4 and M1 in occlusal view.
The M1 (MLP-90-II-5-1631) shows a pattern similar to dP4, but, in contrast, the ectoloph is only slightly curved in the mesostyle region, with a strong parastyle that is projected mesially over the preceding tooth, and a weaker metastyle. Both the paracone and metacone column are identifiable in the labial side of the ectoloph, but the metacone is significantly weaker. There is a short basal shelf between these columns as remnant of a labial cingulum. The crochet is long, contacting with the medial portion of the ectoloph, and isolating a distolabial fossette. The mesial and distal cinguli are strong and well developed. Even though MLP-90-II-5-1631 is a juvenile specimen, the M1, being part of the definitive dentition, contains the same information as in adult specimens (or even more, given the great degree of conservation). The M1 in MLP-90-II-5-1633 (Fig. 4C) is heavily worn, making the outline very rounded, and in the crown only the crochet is distinguishable. MLP-90-II-5-1634 (Fig. 4A) is less worn out than MLP-90-II-5-1633, but the M1 is only preserved from crochet to lingual end. MLP-90-II-5-1635 (Fig. 4G) and MLP-90-II-5-1636 (Fig. 4E) are well preserved, with the features of the genus easily noticeable. The crown in MLP-90-II-5-1635 presents a unified surface between protoloph, metaloph, and crochet, similar to the dP4 in MLP-90-II-5-1631. MLP-90-II-5-1637 (Fig. 4D) and MLP-90-II-5-1638 (Fig. 4F) are as worn out as MLP-90-II-5-1633, being only recognizable by size and the morphology of the ectoloph.
The M2 (MLP-90-II-5-1632; Fig. 4B) follows the same pattern as dP4 and M1, differing in the weaker development of the metacone column in the ectoloph. MLP-90-II-5-1634 (Fig. 4C) presents a M2 with a wider crochet that is confluent with the ectoloph. The distolabial fossette is also wider.
The M3 (MLP-79-I-5-47; Fig. 4H) is a triangular shaped tooth. The distinction between the metacone and metastyle is not clear. As in the M2, the metacone column is poorly developed. The long crochet reaches the vertex where ectoloph and protoloph join together. The metaloph is also long, reaching the lingual limit of the protocone, but with a weak hypocone. The mesial and distal cinguli are strongly developed, the first being longer.
Stratigraphic and geographic range.—Ypresian (early Eocene), Las Flores Formation, Río Chico Group, Itaboraian SALMA. Patagonia outcrops in Chubut Province, Argentina.
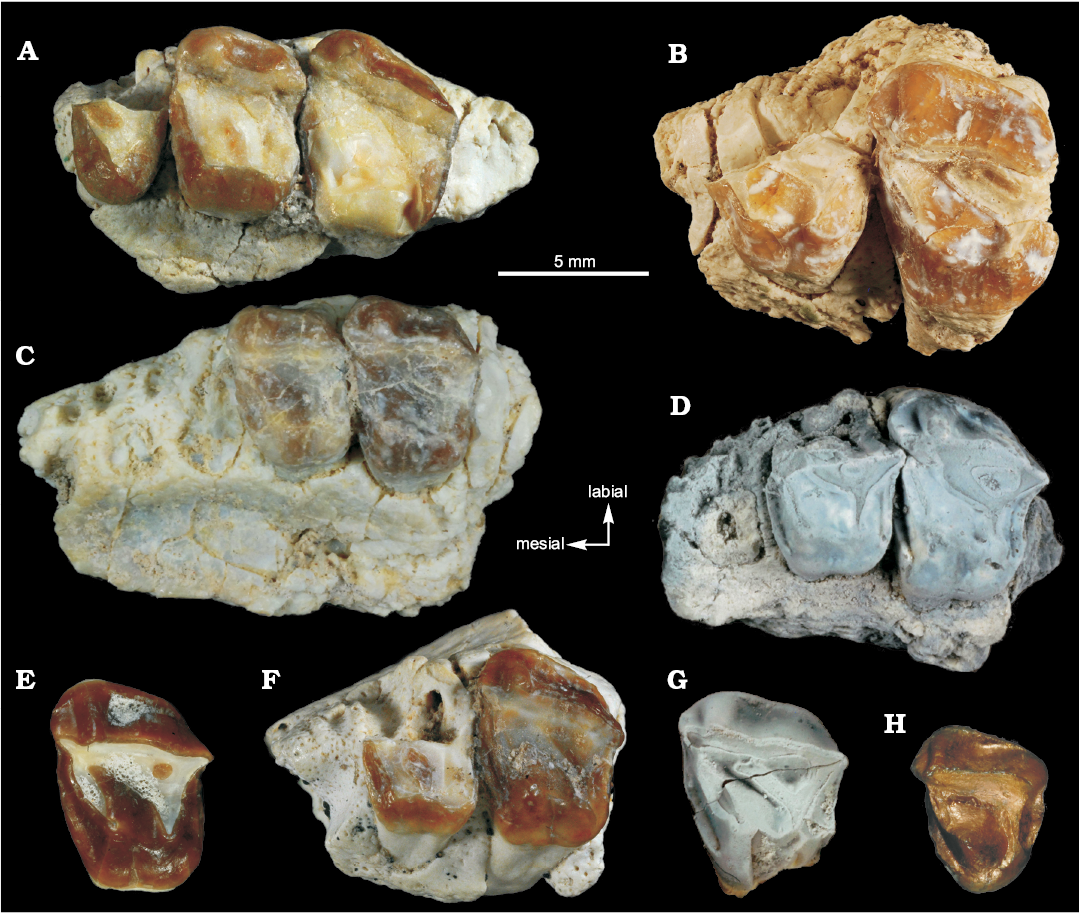
Fig. 4. Notoungulate mammal Orome deepi gen. et sp. nov. from the early Eocene of Las Violetas Farm (A–C) and Las Flores (D–H), Argentina, all tooth in occlusal view. A. MLP-90-II-5-1633, fragment of right maxilla with M1–M2. B. MLP-90-II-5-1632, fragment of left maxilla with M1–M2. C. MLP-90-II-5-1634, fragment of right maxilla with M1–M2. D. MLP-90-II-5-1637, fragment of left maxilla with broken P4, M1, and M2. E. MLP-90-II-5-1636, isolated left M1. F. MLP-90-II-5-1638, fragment of left maxilla with broken M1 and M2. G. MLP-90-II-5-1635, isolated left M1. H. MLP-79-I-5-47, isolated right M3. A, B, C, and H were horizontaly mirrored.
Comparison and phylogenetic analysis
Even though the specimens present a set of features that link them within the henricosborniids (general appearance of the occlusal face of the premolars and molars, presence of a metacone column in the ectoloph, crochet and M3 triangular shaped), they also show unique characteristics that separates them from the rest of the family. The only non-deciduous premolar is a lingual fragment of P4 (MLP-90-II-5-1637) without hypocone, showing a contour compatible with the family. On the other hand, the dP4 shows differences with the other members of the family, due to its trapezoidal outline, resulting from the development of an hypocone, and the presence of a metacone column in the ectoloph. These differences can be explained by the deciduous nature of this tooth, considering its thinner enamel and the higher height of the crown in labial view, in contrast to the M1, which denotes an early eruption. Aside from this specimen, there is only one record of a deciduous premolar assigned to the family, AMNH 28971, which was described as having “well-developed crochet, protocone and hypocone not joined, crown unusually narrow (less transverse), perhaps a deciduous molar” by Simpson (1948: 153). In the molars, the metacone column, while present, is weaker than in most of the henricosborniids genera, but not quite like Notostylops species where, seemingly, it does not develop. The general size is greater than the mean for the group, with the exception of Simpsonotus (S. praecursor is similar and S. major is larger) (Fig. 5). The only M3 (MLP 79-I-5-47) assigned to O. deepi is similar in size to the M3 of Henricosbornia lophodonta (MACN 10808), but in contrast, the ectoloph arrangement is similar to the M2 of other specimens of O. deepi, where the metacone column is poorly developed. Like the other Henricosborniidae, the M3 is triangular in outline, with a straight ectoloph but without a clear sulcus that separates the metacone region from the metastyle This sulcus is not always evident in the M3 of members of the family, but in O. deepi the distal end of the ectoloph is almost flat, due to its weaker metacone column. The metaloph surpasses the crochet, reaching the lingual extreme of the tooth. This makes it longer than in other forms of henricosborniids, where the metaloph only reaches the crochet.
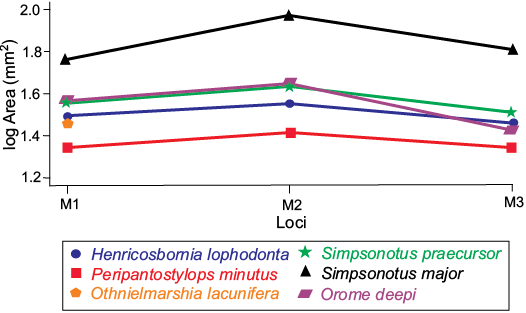
Fig. 5. Size comparison between members of Henricosborniidae and Orome deepi gen. et sp. nov.
The phylogenetic analysis (Fig. 6) resulted in a single most parsimonious tree of 153.71853 steps, with Henricosborniidae being recovered as a monophyletic group and as the sister group of Notostylops, confirming their basal position within the order. Orome is showed as the sister taxon of Peripantostylops, and this clade is located crownward within the family. Resampling values are remarkably low for the relationships mentioned above and even negative in some inner nodes of the clade.
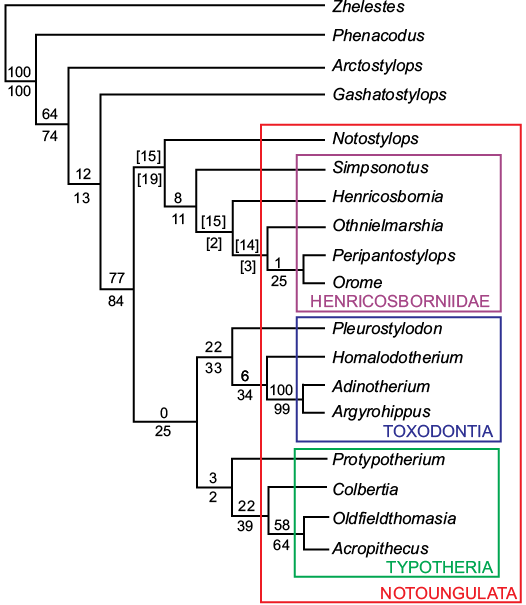
Fig. 6. Phylogenetic analysis. Resampling values of Bootstrap above the branches and Jacknife below the branches. Values between brackets are negative resampling values.
The only autapomorphy of O. deepi, besides geometric morphometric data, is the morphology of the middle portion of the ectoloph, which was one of the discrete characters added. Because of that, we realized a second analysis excluding this character from the matrix, which resulted in four most parsimonious trees of 147.04364 steps, all showing Orome closely grouped with the other henricosborniids and, in three of them recovered as the sister taxon of Peripantostylops, differing in the inner relations of the family (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app64-Bauza_etal_SOM.pdf). Although Henricosborniidae is not recovered as a monophyletic group, we decided to be conservative and place the new taxon as part of this group, since a complete revision of the family is in progress. Still, the fact that the addition of a single character could potentially resolve the relations between the henricosborniids, defining their monophyly, is worth noticing, and evidences the need of a better understanding for characters coded in future matrices.
In general terms, the specimens from Las Flores are smaller (Table 1) than the ones from Las Violetas Farm. We made a couple of geometric morphometric analyses comparing the previously described members of the family with the new specimens in order to see if the two subsamples belong to the same morphotype. The regression test shows only 4.1270% of the shape variation predicted by allometry, which can be considered as non-significant. The PCA (Fig. 7A) shows 88% of the variance contained in the first six Principal Components (see SOM), but no particular grouping is evident. In order to maximize the variance we realized a CVA (Fig. 7B), which shows three well-defined groups: (i) Simpsonotus, (ii) Orome, (iii) Henricosbornia–Peripantostylops–Othnielmarshia. Both analyses are consistent in showing that, even though the difference in size between the specimens of the two localities is evident, it is not significant enough to affect the tooth shape (and by that extent, the relationship between the morphological features).
The geometric morphometric analyses fail to highlight the differences between Orome and the other henricosborniids, so no specific landmark configuration can be defined as diagnostic of the new genus, but both (PCA and CVA) allow us to assign without any doubt the isolated teeth from Las Flores locality mentioned above to Orome.

Fig. 7. Geometric morphometric analyses. A. Principal Component Analysis. B. Canonical Variate Analysis.
Discussion
The presence of the new genus Orome, particularly for the outcrops of Las Violetas Farm locality, could be useful to understand the temporal position of this faunistic assemblage. The stratigraphy in Las Violetas Farm was difficult to define and so, the correlations and comparisons with other mammals from the Rio Chico Group deserve a brief discussion. The outcrops at Las Violetas Farm were first mentioned by Simpson (1935: 14) as “... sands of Río Chico character.” where “... in hard sandstone, rare mammal bones occur … found nothing more exactly determinable than a dasypod scute [...] but is mentioned as another possible site for Rio Chico mammals, worthy of further investigations.” Although three different zones and faunas were identified by Simpson (1935) for his Rio Chico Formation (now Rio Chico Group), which were, from base to top, Carodnia, Kibenikhoria, and Ernestokokenia chaishoer, no particular correlation was done for the level where the scute (AMNH 28488) was found in Las Violetas Farm.
Andreis et al. (1975) and Andreis (1977) seem to have described this particular level in Las Violetas Farm, as the Visser Member of the Río Chico Formation, but without any mention of their fossil content. Legarreta and Uliana (1994) reorganized the units of the San Jorge basin and upgraded Rio Chico to a Group category, dividing it from base to top in: Peñas Coloradas, Las Flores and Koluel-Kaike formations, correlating them with the biostratigraphic subdivision of Simpson (1935), but they did not argue about the fossil levels outcroping in this farm. Following this last stratigraphic organization, Raigemborn (2007) added Las Violetas Formation at the base of the group, which was equivalent to Las Violetas Member of Andreis et al. (1975). This formation was identified in Cañadón Hondo, El Sol, and Las Violetas farms, but not in the southeast part of the basin, like Punta Peligro, where the unit was considered eroded, prior to the deposition of Peñas Coloradas Formation. At Las Violetas Farm, Raigemborn (2007) identified the fossil level, as part of Peñas Coloradas Formation, a unit correlated with Carodnia Zone (Legarreta and Uliana 1994; Bond et al. 1995). Nevertheless, the preliminary analysis of the mammals found in the upper Rio Chico outcrops at Las Violetas Farm locality indicated the presence of a different and younger fauna than those expected for the Carodnia zone, originally characterized by the presence of the Xenungulata Carodnia feruglioi in the locality of Bajo de La Palangana. The mammals found are more closely related to those from Itaboraian SALMA of Las Flores Formation (Gelfo and Lorente 2010; Gelfo et al. 2010) or even to those from Eocene of Paso del Sapo in the west side of Patagonia, Argentina (Tejedor et al. 2009). A reinterpretation of the upper level of Rio Chico Group at Las Violetas Farm allows to assign the fossil mammal level to the Las Flores Formation (Raigemborn et al. 2010), which is in agreement with the faunistic inferences.
Despite the fossil mammal similitudes, the main difficulty in order to establish a litostratigraphic correlation between the localities of Las Flores and Las Violetas Farm rests in their different lithology and different diagenetic alteration of the sediments. In the main locality, Las Flores and the nearby outcrops, fossil mammals appeared in a succession of grey to pink and yellowish, sands to sabulitic sediments, strongly meteorized and where the typical badlands dominated the landscapes. In contrast, in Las Violetas Farm, fossil mammals appeared in medium and thick psammitic up to conglomeratic rocks. These differences were interpreted as a deep facies change of the Las Flores Formation from east to west (Raigemborn et al. 2010; Fig. 3). These differences are related to the paleoenvironmental reconstruction. The western area of Las Flores Formation was interpreted as a low energy environment with shallow lakes, while in the locality of Las Violetas Farm the unit represents a fluvial system with moderate to high sinuosity and extended floodplains (Raigemborn et al. 2010).
The fossil mammals from Las Flores Formation (i.e., Kibenikhoria Zone of Simpson 1935) have been considered as the austral representation of the Itaboraian SALMA (Bond et al. 1995), a biochronologic unit defined in relation to the specimens found in some of the fissure fills from São José de Itaboraí Basin, Brazil. The age interpretation of the Itaboraian SALMA was changing accordingly to the improvements of new isotopic dates and correlations. Originally it was considered as early Paleocene (Marshall 1985), then as late Paleocene (Marshall et al. 1997), late Paleocene–early Eocene (Gelfo et al. 2009), and finally as early Eocene (Woodburne et al. 2014).
Several isotopic dates in Patagonia allowed for a better temporal resolution of the SALMAs and a relocation of them in the geochronologic chart (Clyde et al. 2014; Krause et al. 2017). The new SALMAs calibration evidenced an important gap in the Paleocene (Selandian and Thanetian ages) fossil record. As a consequence, Paleocene mammalian records in Patagonia are Danian in age, and constrained to the Peligran SALMA (Bonaparte et al. 1993; Gelfo et al. 2009) and the Carodnia Zone (Simpson 1935). Although there is a very rich faunistic mammalian diversity in the Peligran SALMA, composed by Dryolestida, Gondwanatheria, Monotremata, Metatheria, Litopterna and archaic ungulates (Gelfo et al. 2009: table 1), no notoungulate was recorded there. The younger levels assigned to the Carodnia Zone, which include the lower levels with mammals in Bajo de La Palangana and Cerro Redondo localities, include xenungulates for the first locality, and polydolopids and probably borhyaenids marsupials, plus litopterns for the last one (Simpson 1935). The oldest records of notoungulates from Patagonia came from those levels related to of the Kibenikhoria Zone equivalent to the Itaboraian SALMA, and from outcrops assigned to Las Flores Formation.
In this context the new taxon here described improves our knowledge of notoungulate diversity by the time this group is first recorded in Patagonia and starting its diversification. Without dismissing the important gap of around 5 million years between the end of the early Paleocene and the Itaboraian SALMA records from the early Eocene, by now, the delayed presence of notoungulates in the Patagonian fossil record could be related with progressive change in the climatic conditions and particularly the EECO context in South America (Goin et al. 2012).
Important global changes in the climate and ocean temperatures occurred during the Paleogene (Zachos et al. 2001) with a great worldwide impact on biotic evolution (Woodburne et al. 2014). In South America and West Antarctica, land mammal development and evolution seem to be directly conditioned by the global climatic changes, floral diversity (Woodburne et al. 2014) and paleogeographic constraints (Reguero et al. 2014). The EECO, which approximately occurred between 53 and 50 Ma (Tsukui and Clyde 2012), was the warmest period during the Cenozoic and was characterized by a very low thermal gradient between the poles and the equator (Keating-Bitonti et al. 2011). According to different lines of evidence, Patagonia experienced several changes in the paleoclimatic conditions from the Paleocene to the early Eocene. The variation in the clay mineralogy of paleosols of the Rio Chico Group, particularly in the late Paleocene, was interpreted as a seasonality increase, and the Paleocene–Eocene transition considered as a shift from warm-temperate and humid climate with seasonal precipitation to a wetter subtropical climate with year-around rainfall (Raigemborn et al. 2009; Woodburne et al. 2014). The Golfo San Jorge Basin localities (e.g., Las Flores and Las Violetas Farm) and the Rio Chico Group, where notoungulates first appeared in the Patagonian fossil record, are mainly evidence of coastal lowland environment (Raigemborn et al. 2009). There, the floras experienced a change in their composition, from podocarp and araucarian conifer forest, and mangrove swamps and woodlands in the lowlands, to tropical to subtropical lowland forest along with grass understory. This last setting is particularly represented in the lower part of Las Flores Formation, with phytoliths of palm (Arecaceae), Mimosoideae (Acacia-like), Chrysobalanaceae (Coco plum), and Lauraceae (Brea et al. 2008). Particularly at Las Violetas Farm locality, the phytoliths found at the same levels of Las Flores Formation where some of the specimens of Orome deepi sp. nov. here described were found, indicated a predominance of palms and a few elements assigned to grasses (Erra et al. 2014). In the upper part of the Las Flores unit, there is a predominance of more arboreal elements as Magnoliaceae, Annonaceae, Burseraceae, Boraginaceae, and an herbaceous component of tropical Zingeberales and Poaceae (Woodburne et al. 2014 and literature therein).
Conclusions
The main characteristics of the notoungulate Orome deepi gen. and sp. nov., namely the presence of metacone column in the ectoloph, crochet and M3 triangular shaped, allow us to consider this new taxon as part of the family Henricosborniidae, and closely related to Peripantostylops. Although our cladistic analysis (Fig. 5) indicates the monophyly of the family, we considered this analysis as not robust enough to reinforce the hypothesis of henricosbornids as a natural group. Indeed, a second analysis, excluding the only qualitative character that define O. deepi, suggests the paraphyly of the family (see SOM). The presence of O. deepi in Las Flores and Las Violetas Farm localities, indicates that the notoungulate radiation in the early Eocene of Patagonia was more important than what has been recorded until now.
Notoungulate fossil record in southern South America, and particularly in Patagonia, seems to start concomitantly to the pre- and syn-EECO climatic conditions and the related floristic development. Since the Itaboraian SALMA the mammalian fossil record was enriched by the first xenarthran record, the radiation of basal “Ameridelphian” metatherians and polydolopimorphians (Goin et al. 2016), the increase of ungulate diversity, with the “explosive” appearance in Patagonia of Astrapotheria, Litopterna and notoungulates. In contrast to the post-EECO ungulates, for which a trend to hypsodonty is clearly identified (Scarano 2009; Madden 2014), the mentioned radiation of SAnu in Patagonia could be characterized by groups with brachydont dentitions, with different types and sizes, a bunodont pattern among didolodontids, and a bunolophodont up to bunoselenodont pattern among other litopterns, xenungulates, and notoungulates.
Acknowledgements
We would like to thank Darin Croft (Department of Anatomy Case Western Reserve University School of Medicine, Cleveland, OH, USA) and Ana Maria Ribeiro (Fundação Zoobotânica do Rio Grande do Sul, Porto Alegre, Brazil) for the suggestions made to an early version of this paper, to Bruno Pianzola (Museo de La Plata, La Plata, Argentina) for the photographs utilized in this work, and Olivier Lambert and the editorial team of Acta Palaeontologica Polonica. This work was partially financed by PIP 0489 (CONICET) to JNG and by the project 11/N876 (Universidad Nacional de La Plata, La Plata, Argentina) to GML.
References
Ameghino, F. 1901. Notices préliminaires sur des ongulés nouveaux des terrains crétacés de Patagonie. Boletín de la Academia Nacional de Ciencias en Córdoba 16: 349–426.
Ameghino, F. 1906. Les formations sedimentaires du cretace superieur et du tertiare de Patagonie avec un parallele entre leurs faunes mammalogiques et celles de L’ancien continent. Boletín de la Academia Nacional de Ciencias en Córdoba 8: 1–568.
Andreis, R.R., Mazzoni, M.M., and Spalletti, L.A. 1975. Estudio estratigráfico y paleoambiental de las sedimentitas terciarias entre Pico Salamanca y Bahía Bustamante, Provincia de Chubut, República Argentina. Revista de la Asociación Geológica Argentina 30: 85–103.
Andreis, R.R. 1977. Geología del área de Cañadón Hondo, Departamento Escalante, Provincia del Chubut, República Argentina. Obra del Centenario del Museo de La Plata 4: 77–102.
Babot, J., García-López, D., Deraco, V., Herrera, C.M., and del Papa, C. 2017. Mamíferos paleógenos del subtrópico de Argentina: síntesis de estudios estratigráficos, cronológicos y taxonómicos. In: C.M. Muruaga and P. Grosse (eds.), Ciencias de la Tierra y Recursos Naturales del NOA. Relatorio del XX Congreso Geológico Argentino, 730–753. Tucumán.
Billet, G. 2011. Phylogeny of the Notoungulata (Mammalia) based on cranial and dental characters. Journal of Systematic Palaeontology 9: 481–497. Crossref
Bonaparte, J.F., Van Valen, L.M., and Kramartz, A. 1993. La fauna local de Punta Peligro, Paleoceno inferior de la provincia del Chubut, Patagonia, Argentina. Evolutionary Monographus 14: 1–16.
Bond, M. and Vucetich, M. 1983. Indalecia grandensis sp. nov. del Eoceno temprano del noroeste Argentino, tipo de una nueva subfamilia de los Adianthidae (Mammalia, Litopterna). Revista de la Asociación Geológica Argentina 38: 107–117.
Bond, M., Carlini, A., Goin, F., Legarreta, L., Ortiz Jaureguizar, E., Pascual, R., and Uliana, M. 1995. Episodes in South Land Mammal evolution and sedimentation: testing their apparent concurrence in a Palaeocene succession form Central Patagonia. In: VI Congreso Argentino de Paleontología y Bioestratigrafía, Actas 1: 47–58.
Brea, M., Zucol, A.F., Raigemborn, M.S., and Matheos, S. 2008. Reconstruction of past vegetation through phytolith analysis of sediments from the upper Paleocene–Eocene? (Las Flores Formation), Chubut, Argentina. In: M.A. Korstanje, M. del P. Babot (eds.), Matices interdisciplinarios en estudios fitoliticos y otros microfósiles. BAR Internat Ser. S 1870: 91–108.
Cifelli, R.L., Schaff, C.R., and McKenna, M.C. 1989. The Relationships of the Arctostylopidae (Mammalia): New Data and Interpretation. Bulletin of the Museum of Comparative Zoology at Harvard College 152: 1–44.
Clyde, W.C., Wilf, P., Iglesias, A., Slingerland, R.L., Barnum, T., Bijl, P.K., Bralower, T.J., Brinkhuis, H., Comer, E.E., Huber, B.T., Ibañez-Mejia, M., Jicha, B.R., Krause, J.M., Schueth, J.D., Singer, B.S., Raigemborn, M.S., Schmitz, M.D., Sluijs, A., and Zamaloa, M. del C. 2014. New age constraints for the Salamanca Formation and lower Río Chico Group in the western San Jorge Basin, Patagonia, Argentina: Implications for Cretaceous–Paleogene extinction recovery and land mammal age correlations. GSA Bulletin 126: 289–306. Crossref
Erra, G., Strömberg, C.A., Gelfo, J.N., Madden, R.H., and Goin, F.J. 2014. Fitolitos del Grupo Río Chico (Paleógeno de la Patagonia central): análisis florístico e inferencias ambientales. In: XI Congreso Latinoamericano de Botánica – LXV Congresso Nacional de Botânica. Abstracts, 1. Salvador, Bahia.
Gelfo, J.N. and Lorente, M. 2010. Asociaciones de elementos postcraneales en ungulados nativos del paleógeno. Ciências Morfológicas 12: 1–6.
Gelfo, J.N., Chornogubsky, L., López, G.M., Goin, F.J., and Ciancio, M. 2010. Biochronological relationships of the mammal fauna from the Paleogene of Las Violetas, Chubut Province, Argentina. In: S. Ballent, A. Artabe, and F. Tortello (eds.), X Congreso Argentino de Paleontología y Bioestratigrafía – VII Congreso Latinoamericano de Paleontología, 61–62. Museo de La Plata, La Plata.
Gelfo, J.N., Goin, F.J., Woodburne, M.O., and Muizon, C.D. 2009. Biochronological relationships of the earliest South American Paleogene mammalian faunas. Palaeontology 52: 251–269. Crossref
Goin, F.J., Gelfo, J.N., Chornogubsky, L., Woodburne, M.O., and Martin, T. 2012. Origins, radiations, and distribution of South American mammals: from greenhouse to icehouse worlds. In: B. Patersson (ed.), Bones, Clones, and Biomes: An 80-million Year History of Modern Neotropical Mammals, 20–49. University of Chicago Press, Chicago. Crossref
Goin F.J., Woodburne M.O., Zimicz A.N., Martin G.M., and Chornogubsky L. 2016. Dispersal of vertebrates from between the Americas, Antarctica, and Australia in the Late Cretaceous and Early Cenozoic. In: F.J. Goin, M.O. Woodburne, A.N. Zimicz, G.M. Martin, and L. Chornogubsky (eds.), A Brief History of South American Metatherians, 77–124. Springer Earth System Sciences, Dordrecht. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Keating-Bitonti, C.R., Ivany, L.C., Affek, H.P., Douglas, P., and Samson, S.D. 2011. Warm, not super-hot, temperatures in the early Eocene subtropics. Geology 39: 771–774. Crossref
Klingenberg, C.P. 2011. MorphoJ: an integrated software package for geometric morphometrics. Molecular Ecology Resources 11: 353–357. Crossref
Kondrashov, P.E. and Lucas, S.G. 2004. Palaeostylops iturus from the upper Paleocene of Mongolia and the status of Arctostylopida (Mammalia, Eutheria). In: S.G. Lucas, K.E. Zeigler, and P.E. Kondrashov (eds.), Paleogene Mammals. New Mexico Museum Natural History and Science Bulletin 26: 195–204.
Krause, J., Clyde, W., Ibañez-Mejia, M., Schmitz, M.D., Barnum, T., Bellosi, E., and Wilf, P. 2017. New age constraints for early Paleogene strata of central Patagonia, Argentina: Implications for the timing of South American Land Mammal Ages. Bulletin of the Geological Society of America 129: 886–903. Crossref
Legarreta, L. and Uliana, M.A. 1994. Asociación de fósiles y hiatos en el Supracretácico–Neógeno de Patagonia: una perspectiva estratigráfico-secuencial. Ameghiniana 31: 257–281.
López, G.M. 2008. Los ungulados de la Formación Divisadero Largo (Eoceno inferior?) de la provincia de Mendoza, Argentina: sistemática y consideraciones bioestratigráficas. 407 pp. Tesis doctoral, Universidad Nacional de La Plata, La Plata.
Lorente, M. 2015. Desarrollo de modelos de asociación y clasificaciones de restos postcraneanos aislados de ungulados nativos del Paleoceno–Eoceno de América del Sur. 588 pp. Tesis doctoral, Universidad Nacional de La Plata, La Plata.
Madden, R.H. 2014. Hypsodonty in Mammals. 423 pp. Cambridge University Press, Cambridge. Crossref
Marshall, L.G. 1985. Geochronology and land-mammal biochronology of the transamerican faunal interchange. In: F.G. Stehli and S.D. Webb (eds.), The Great American Biotic Interchange, 49–85. Springer, Boston. Crossref
Marshal, L.G., Sempere, T., and Butler, R.F. 1997. Chronostratigraphy of the Mammal-Bearing Paleocene of South America. Journal of South American Earth Sciences 10: 49–70. Crossref
McKenna, M.C. and Bell, S.K. 1997. Classification of Mammals: Above the Species Level. 631 pp. Columbia University Press, New York.
Missiaen, P., Escarguel, G., Hartenberger, J.L., and Smith, T. 2012. A large new collection of Palaeostylops from the Paleocene of the Flaming Cliffs area (Ulan-Nur Basin, Gobi Desert, Mongolia) and an evaluation of the phylogenetic affinities of Arctostylopidae (Mammalia, Gliriformes). Geobios 45: 311–322. Crossref
Muizon, C. de 1991. La Fauna de mamíferos de Tiupampa (Paleoceno inferior, Formación Santa lucía), Bolivia. Revista Técnica de Yacimientos Petrolíferos Fiscales Bolivianos 12: 575–624.
Muizon, C. de, Marshall, L., and Sigé, B. 1984. The mammal fauna from the El Molino Formation (Late Cretaceous, Maastrichtian) at Tiupampa, Southcentral Bolivia. Bulletin du Muséum national d’histoire naturelle. Section C, Sciences de la terre, paléontologie, géologie, minéralogie 6 section C 4: 327–351.
Pascual, R., Vucetich, M.G., and Fernandez, J. 1978. Los primeros mamíferos (Notoungulata, Henricosborniidae) de la Formación Mealla (Grupo Salta, subgrupo Santa Bárbara). Sus implicancias filogenéticas, taxonómicas y cronológicas. Ameghiniana 15: 366–390.
Patterson, B. 1934. Cranial characters of Homalodotherium. Geological Series of Field Museum of Natural History 6 (7): 113–117. Crossref
Raigembron, M.S. 2007. Estudio Estratigráfico, Sedimentológico y Composicional de las Sedimentitas del Terciario Onferior (Grupo Río Chico) en el Sector Sudoriental del Chubut Extraandino. 367 pp. Tesis doctoral, Universidad Nacional de La Plata, La Plata.
Raigemborn, M.S., Brea, M., Zucol, A.F., and Matheos, S.D. 2009. Early Paleogene climate at mid latitude in South America: Mineralogical and paleobotanical proxies from continental sequences in Golfo San Jorge basin (Patagonia, Argentina). Geologica Acta 7: 125–145.
Raigemborn, M.S., Krause, J.M., and Bellosi, E. 2010. Redefinición Estratigráfica del Grupo Río Chico (Paleógeno Inferior), En El Norte De La Cuenca Del Golfo San Jorge, Chubut. Revista de la Asociación Geológica Argentina 67: 239–256.
Reguero, M.A., Gelfo, J.N., López, G.M., Bond, M., Abello, A., Santillana, S.N., and Marenssi, S.A. 2014. Final Gondwana breakup: the Paleogene South American native ungulates and the demise of the South America-Antarctica land connection. Global and Planetary Change 123: 400–413. Crossref
Rohlf, F.J. 2004. tpsUtil, File Utility Program, Version 1.26. Department of Ecology and Evolution, State University of New York, Stony Brook.
Rohlf, F.J. 2005. tpsDig, Digitize Landmarks and Outlines, Version 2.05. Department of Ecology and Evolution, State University of New York, Stony Brook.
Rohlf, F.J., and Slice, D. 1990. Extensions of the Procrustes method for the optimal superimposition of landmarks. Systematic Biology 39: 40–59. Crossref
Roth, S. 1903. Noticias preliminares sobre nuevos mamíferos fósiles del cretáceo superior y terciario inferior de la Patagonia. Revista del Museo de La Plata 11: 133–138.
Scarano, A.C. 2009. El proceso de desarrollo de la hipsodoncia durante la transición Eoceno–Oligoceno. 306 pp. Doctoral dissertation, Universidad Nacional de La Plata, La Plata.
Scott, W. B. 1912. Mammalia of the Santa Cruz beds. Part 2. Toxodonta of the Santa Cruz Beds. Reports of the Princeton University expeditions to Patagonia 6 (2): 111–238.
Simpson, G.G. 1935. Occurrence and relationships of the Río Chico fauna of Patagonia. American Museum Novitates 818: 1–21.
Simpson, G.G. 1948. The beginning of the age of mammals in South America. Part 1: Introduction. Systematics: Marsupialia, Edentata, Condylarthra, Litopterna and Notioprogonia. Bulletin of the American Museum of Natural History 91: 1–232.
Tejedor, M.F., Goin, F.J., Gelfo, J.N., López, G., Bond, M., Carlini, A.A., Scillato-Yané, G.J., Woodburne, M.O., Chornogubsky, L., Aragón, E., Reguero, M.A., Czaplewski, N.J., Vincon, S., Martin, G.M., and Ciancio, M.S. 2009. New early Eocene mammalian fauna from western Patagonia, Argentina. American Museum Novitates 3638: 1–43. Crossref
Tsukui, K. and Clyde, W.C. 2012. Fine-tuning the calibration of the early to middle Eocene geomagnetic polarity time scale: Paleomagnetism of radioisotopically dated tuffs from Laramide foreland basins. Geological Society of America Bulletin 124: 870–885.Crossref
Vera, B. 2015. Phylogenetic revision of the South American notopithecines (Mammalia: Notoungulata). Journal of Systematic Palaeontology 14: 461–480. Crossref
Woodburne, M.O., Goin, F.J., Raigemborn, M.S., Heizler, M., Gelfo, J.N., and Oliveira, E.O. 2014. Revised timing of the South American early Paleogene land mammal ages. Journal of South American Earth Sciences 54: 109–119. Crossref
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292: 686–693. Crossref
Taxa used in this work:
Non-notoungulates
Zhelestes temirkazyk: CCMGE 10/12176, ZIN 88469 (outgroup); Arctostylops steini: MCZ 20004; Gashatostylops macrodon: AMNH 109521, AMNH 21741; Phenacodus spp.: Thewissen (1990).
Notoungulata
Henricosborniidae: Henricosbornia lophodonta: MACN 10808, MACN 10795; Peripantostylops minutus: AMNH 28494, MACN 10796; Othniemarshia lacunifera: MACN 10807; MACN 10803; Simpsonotus praecursor: MLP 73-VII-3-11; Simpsonotus major: MLP 73-VJI-3-12. Notostylopidae: Notostylops murinus: MACN 10509, MACN 10498. Oldfieldthomasiidae: Oldfieldthomasia debilitata: photographs of AMNH 28691, AMNH 28963; Colbertia magellanica, Colbertia lumbrerense: AMNH49873, 49867, 49868, 49874, 49871, and 49887; photographs of PVL-5-11 and PVL-5-2. Archaeophitecidae: Acropithecus rigidus: photographs of AMNH 28782. Interatheriidae: Protypotherium spp.: MNHN-SCZ 178, 179, and 184; MACN A-4001; FMNH P13002; AMNH 9534, photographs of AMNH 9500. Isotemnidae: Pleurostylodon modicus: AMNH 28880; Pleurostylodon similis: AMNH 28830. Homalodotheriidae: Homalodotherium spp.: MNHN-SCZ 2; FMNH P 13092; Patterson (1934). Notohippidae: Argyrohippus fraterculus: AMNH 29685. Toxodontidae: Adinotherium spp.: MNHN-SCZ 5, 13, and 14; FMNH P 13110; Scott (1912).
Characters added to the character-taxon matrix:
– Middle portion of the ectoloph (in occlusal view)
(0: curved outwards, 1: slightly curved outwards, 2: straight, 3: curved
inwards)
– Hypoconid (0: absent, 1: present)
– Landmark configurations: M1–M3 for Henricosbornia
lophodonta, Peripantostylops minutus,
Othniemarshia lacunifera (only M1), Simpsonotus
praecursor, Orome deepi, Notostylops
murinus, Acropithecus rigidus, Oldfieldthomasia debilitata,
Pleurostylodon modicus, Protypotherium, and Argyrohippus;
m1–m3 for H. lophodonta, P.
minutus, O. lacunifera (only m1),
S. praecursor, N.
murinus, A. rigidus, O.
debilitata, and P.
modicus.