
A reassessment of the purported ankylosaurian dinosaur Bienosaurus lufengensis from the Lower Lufeng Formation of Yunnan, China
THOMAS J. RAVEN, PAUL M. BARRETT, XING XU, and SUSANNAH C.R. MAIDMENT
Raven, T.J., Barrett, P.M., Xu, X., and Maidment, S.C.R. 2019. A reassessment of the purported ankylosaurian dinosaur Bienosaurus lufengensis from the Lower Lufeng Formation of Yunnan, China. Acta Palaeontologica Polonica 64 (2): 335–342.
The earliest definitive ornithischian dinosaurs are from the Early Jurassic and are rare components of early dinosaur faunas. The Lower Lufeng Formation (Hettangian–Sinemurian) of Yunnan Province, China, has yielded a diverse Early Jurassic terrestrial vertebrate fauna. This includes several incomplete specimens have been referred to Ornithischia, including the type specimen of the thyreophoran “Tatisaurus” and other generically indeterminate material. The highly fragmentary Lufeng ornithischian Bienosaurus lufengensis was described briefly in 2001 and identified as an ankylosaurian dinosaur. Recent studies have cast doubt on this hypothesis, however, and given that the referral of Bienosaurus to Ankylosauria would result in an extensive ghost-lineage extending between it and the first definitive eurypodans (ankylosaurs + stegosaurs) in the Middle Jurassic, the holotype specimen is re-examined and re-described. We identify Bienosaurus as a probable thyreophoran dinosaur, although the fragmentary nature of the material and the absence of autapomorphies means that the specimen should be regarded as a nomen dubium.
Key words: Dinosauria, Ornithischia, Thyreophora, anatomy, Jurassic, Lufeng Formation, Yunnan, China.
Thomas J. Raven [t.raven@nhm.ac.uk], Department of Earth Sciences, Natural History Museum, Cromwell Road, London SW7 5BD, UK; School of Environment and Technology, University of Brighton, Lewes Road, Brighton BN1 4JG, UK.
Paul M. Barrett [p.barrett@nhm.ac.uk] and Susannah C.R. Maidment [susannah.maidment@nhm.ac.uk], Department of Earth Sciences, Natural History Museum, Cromwell Road, London SW7 5BD, UK.
Xing Xu [xu.xing@ivpp.ac.cn], Key Laboratory of Evolutionary Systematics of Vertebrates, Institute of Vertebrate Paleontology & Paleoanthopology, Chinese Academy of Sciences, 100044 Beijing, People’s Republic of China; Center of Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, 100044 Beijing, People’s Republic of China.
Received 30 November 2018, accepted 15 January 2019, available online 19 March 2019.
Copyright © 2019 T.J. Raven et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Ornithischian dinosaurs exhibit exceptional diversity in the middle–late Mesozoic and have an excellent fossil record (Weishampel et al. 2004). Understanding the early evolution and radiation of the group is, however, hindered by the fact that early-diverging ornithischian dinosaurs are relatively rare (Weishampel et al. 2004). Until recently, it was thought that the earliest known ornithischians were from the Late Triassic and a review of purported Triassic ornithischian taxa concluded that only three specimens could be definitively assigned to the clade (Irmis et al. 2007). However, two of those specimens come from formations that are now considered to be Early Jurassic in age (Eocursor from the upper Elliot Formation of South Africa and an unnamed heterodontosaurid from the Laguna Colorada Formation of Argentina; Olsen et al. 2011). The third, Pisanosaurus from the Ischigualasto Formation of Argentina, has recently been recovered as a silesaurid in two independent phylogenetic analyses (Baron et al. 2017a; Agnolín and Rozadilla 2018). Consequently, the current earliest definitive ornithischians are from the earliest Jurassic (e.g., Laquintasaura; Barrett et al. 2014), and so any information that we can glean from this period will be vital for elucidating the early evolution of the clade.
The Lower Lufeng Formation of the Lufeng Basin, Yunnan, China, is known for its diverse, well-preserved Early Jurassic terrestrial vertebrate faunas. These include non-mammalian synapsids, mammals, lepidosaurs, crocodylomorphs, and non-avian dinosaurs (Young 1951; Sun et al. 1985; Luo and Wu 1994; Lucas 2001). Non-sauropod sauropodomorph dinosaurs, such as Lufengosaurus and Yunnanosaurus, dominate these faunas and theropod dinosaurs, such as Panguraptor and Eshanosaurus are also known. However, only three fragmentary specimens referable to Ornithischia have been recovered from Lower Lufeng deposits thus far (Barrett and Xu 2005; Irmis and Knoll 2008). These are a partial hind limb, referred to an indeterminate ornithischian (Irmis and Knoll 2008), the holotype of “Tatisaurus oehleri” a poorly preserved dentary of an indeterminate basal thyreophoran dinosaur (Norman et al. 2007), and Bienosaurus lufengensis, which was originally described as an ankylosaurian (Dong 2001). A further putative ornithischian, “Dianchungosaurus lufengensis”, which was originally referred to Heterodontosauridae (Yang 1982), has since been shown to represent a chimera of sauropodomorph dinosaur and crocodyliform remains (Barrett and Xu 2005).
Thyreophora comprises Ankylosauria, its sister taxon Stegosauria and several early-diverging forms. The earliest definitive ankylosaur is from the Callovian Oxford Clay Formation of the UK (Galton 1983), whilst the earliest stegosaur, Isaberrysaura, is from the early Bajocian Los Molles Formation of Argentina (Salgado et al. 2017; Han et al. 2018). This suggests that Ankylosauria and Stegosauria diverged during either the latest Early or early Middle Jurassic. The age of the Lower Lufeng Formation has been debated, although most studies suggest that it is Hettangian–Sinemurian in age (Luo and Wu 1994). Thus if Bienosaurus is an ankylosaur, it would represent the earliest record of the clade comprising Stegosauria + Ankylosauria (Eurypoda), invoking a ghost lineage of ~20 million years prior to their currently known minimum divergence time.
Bienosaurus lufengensis was found in the “Dark Red Beds” of the Lower Lufeng Formation in the late 1930s and consists of an incomplete right lower jaw with teeth and associated cranial fragments (Dong 2001). It was originally referred to Scelidosauridae, along with Emausaurus, “Lusitanosaurus”, Scutellosaurus, and Scelidosaurus, a family that Dong (2001) considered was within Ankylosauria. However, in the same publication, (Dong 2001: 241) also inconsistently described it a “neither a stegosaur nor ankylosaur, but as a primitive thyreophoran”. The validity and ankylosaurian affinities of Bienosaurus have been questioned on several occasions since the original description (Parish 2005; Irmis and Knoll 2008; Arbour 2014) and studies of ornithischian and thyreophoran phylogeny (Butler et al. 2008; Maidment et al. 2008; Boyd 2015; Raven and Maidment 2017) have independently suggested that Scutellosaurus, Emausaurus, and Scelidosaurus form a paraphyletic grade of successive sister-taxa to Eurypoda. The aim of this study is to re-examine the holotype and only known specimen of Bienosaurus and reassess its validity and systematic position.
Institutional abbreviations.—IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China; MNA, Museum of Northern Arizona, Flagstaff, USA; NHMUK, Natural History Museum, London, UK; SGWG, Sektion Geologische Wissenshaften Griefswald, Ernst-Moritz Universität, Griefswald, Germany.
Other abbreviations.—CI, consistency index; MPTs, most parsimonious trees; OTU, operational taxonomic unit; RI, retention index.
Systematic palaeontology
Dinosauria Owen, 1842
Ornithischia Seeley, 1887
Thyreophora Nopcsa, 1915 (sensu Norman 1984)
Thyreophora indet.
“Bienosaurus lufengensis” Dong, 2001 (nomen dubium)
Figs. 1, 2.
Holotype: IVPP V15311, partial right dentary with associated partial frontal and cranial fragments. Dong (2001) originally referred to this specimen using the catalogue number IVPP V9612, but this was already in use for the holotype specimen of Sinornithoides youngi, a troodontid theropod dinosaur from the Lower Cretaceous Ejinhoro Formation of Inner Mongolia, China (Russell and Dong 1993). Subsequently, the holotype of “Bienosaurus lufengensis” was assigned a new unique catalogue number, IVPP V15311, which should be used henceforth.
Type locality: Lufeng Basin, Yunnan, China.
Type horizon: Dark Red Beds, Lower Lufeng Formation, Hettangian–Sinemurian, Early Jurassic (Luo and Wu 1994).
Material.—Holotype only.
Description.—Dentary: A partial right dentary is preserved (Fig. 1). The anteroposterior length of the ramus is 53 mm and the maximum dorsoventral height is 12.5 mm, located at the posterior of the ramus. The transverse width of the anterior end is 3.5 mm and this thickens posteriorly to 9.5 mm posteriorly. This is a similar size to most basal thyreophorans such as “Tatisaurus” (length 64 mm; Norman et al. 2007), Scutellosaurus (MNA P1.175, length 57.5 mm; Colbert 1981) and Emausaurus (SGWG 85, length 68.6 mm; Haubold 1990), but it is smaller than Scelidosaurus (NHMUK PV R1111, length 111 mm) and more deeply-nested members of Thyreophora, such as the ankylosaur Euoplocephalus (length 163 mm; Vickaryous et al. 2004) or the early-diverging stegosaur Huayangosaurus (IVPP V6728, length 236 mm; Sereno and Dong 1992). Only the anterior portion of the dentary is preserved; there is no evidence of a predentary (contra Dong 2001), and the poorly preserved dentary symphysis suggests that the predentary would have been small. This condition is similar to that in the basal thyreophorans “Tatisaurus” (Norman et al. 2007), Scutellosaurus (MNA P1.175; Colbert 1981) and Emausaurus (SGWG 85; Haubold 1990). The dorsoventral depth of the dentary symphysis is less than half the maximum depth of the dentary ramus. There is a large fracture extending dorsoventrally at the mid-length of the ramus and the lateral surface is poorly preserved, so no foramina are visible. The lateral surface (Fig. 1A1) of the dentary is laterally convex but the medial surface (Fig. 1A2) is concave dorsoventrally at its anterior end and convex at its posterior end. Cortical remodelling, an area of rugose grooves and ridges that is present in many thyreophorans and which reaches maximum development in ankylosaurs where the entire skull roof is modified by cortical remodelling and the fusion of dermal ossifications (Maidment et al. 2008), is present at the posterior end of the dentary lateral surface but it does not extend past the midpoint of the ramus. A deeply incised Meckelian groove is present in medial view and extends for most of the length of the preserved ramus, a condition very similar to that of “Tatisaurus” (Norman et al. 2007), Emausaurus (SGWG 85; Haubold 1990) and Scutellosaurus (MNA P1.175; Colbert 1981). There is a tapering longitudinal bulge on the lateral surface of the dentary that forms the boundary of a buccal emargination and the posterior-most tooth is inset on the medial edge of the dentary. This is similar to the condition seen in the basal ornithischian Lesothosaurus (NHMUK PV RUB23) and the basal thyreophorans “Tatisaurus” (Norman et al. 2007), Scutellosaurus (MNA P1.175; Colbert 1981), and Scelidosaurus (NHMUK PV R1111), although this is not present in Emausaurus (SGWG 85; Haubold 1990). Eurypodans often have a deep buccal emargination that extends for the entire length of the tooth row (Vickaryous et al. 2004; Maidment et al. 2008). In Bienosaurus the tooth row is not obscured by a lamina and is visible in lateral view, in contrast to the condition present in some stegosaurs (Maidment et al. 2008). In lateral view, the tooth row of Bienosaurus is horizontal and straight, thereby differing from those of other thyreophorans, which possess a downturned anterior portion of the dentary (Norman et al. 2004). However, this is similar to the condition seen in Lesothosaurus (NHMUK PV RUB23). In dorsal view (Fig. 1A3), the tooth row is sinuous, again similar to Lesothosaurus (NHMUK PV R8501), the basal thyreophorans “Tatisaurus” (Norman et al. 2007), Scutellosaurus (MNA P1.175; Colbert 1981), Emausaurus (SGWG 85; Haubold 1990) and Scelidosaurus (NHMUK PV R1111), some ankylosaurs (e.g., Euoplocephalus; Arbour and Currie 2016) and stegosaurs (Maidment et al. 2008). In lateral view, the ventral margin of the dentary is horizontal and straight. The alveoli face dorsally, in contrast to those in some stegosaurs (e.g., Stegosaurus, NHMUK PV R36730) where the alveoli face dorsomedially, but similar to most other thyreophorans.
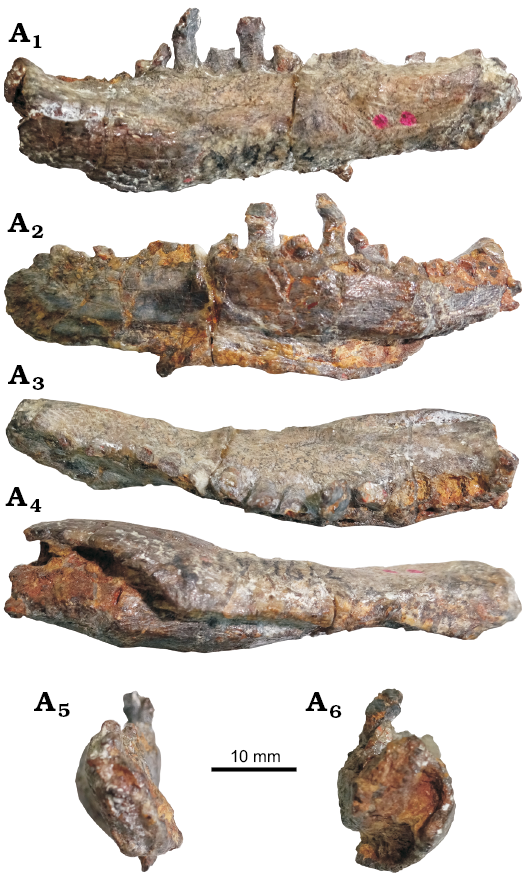
Fig. 1. Thyreophoran dinosaur Bienosaurus lufengensis Dong, 2001 (IVPP V15311, holotype), from the Hettangian–Sinemurian (Lower Jurassic), Lower Lufeng Formation of Yunnan, China; partial right dentary in lateral (A1), medial (A2), dorsal (A3), ventral (A4), anterior (A5), and posterior (A6) views.
Teeth: Ten dentary teeth are preserved, with an additional three alveoli. For the majority of the teeth, only the roots are preserved; two well-preserved tooth crowns are present, and in one tooth the crown is broken in half. The teeth are heavily eroded and no wear facets can be identified. The best-preserved tooth, which is the third from the posterior end, has a crown height of 2 mm and a minimum root length of 3.5 mm. As it is attached to the dentary; the total root length is presumably greater than this. Replacement teeth are associated with the second most and fourth most posterior teeth, where a cross-section of the tooth root can also be observed. As most of the teeth lack crowns, it is not possible to say whether tooth crown size changes along the tooth row, but of the two well-preserved tooth crowns, the posterior one is largest and is present at the mid-length of the ramus. The teeth are leaf-shaped and symmetrical in labial view, similar to those of many basal ornithischian dinosaurs such as Lesothosaurus (e.g., NHMUK PV R8501; Sereno 1991), the basal thyreophorans Scutellosaurus (MNA P1.175; Colbert 1981), Emausaurus (SGWG 85; Haubold 1990) and Scelidosaurus (NHMUK PV R1111; Barrett 2001), as well as ankylosaurs and stegosaurs (Barrett 2001; Vickaryous et al. 2004; Maidment et al. 2008). The crown surfaces are striated, but these striations are not confluent with marginal denticles, similar to the condition in basal thyreophorans such as Scutellosaurus (MNA P1.175; Colbert 1981) and Emausaurus (SGWG 85; Haubold 1990). This, however, contrasts with Scelidosaurus where the striations are confluent with the marginal denticles but they do not extend all the way down the tooth crown (NHMUK PV R1111; Barrett 2001). In some ankylosaurs (e.g., Pinacosaurus, IVPP V16346) the striations are confluent with marginal denticles and these extend all the way down the tooth crowns. In stegosaurs this is variable; the denticles are larger in the early-diverging stegosaur Huayangosaurus (IVPP V6728; Sereno and Dong 1992), but are smaller in the more deeply-nested taxon Stegosaurus (NHMUK PV R36730), with an intermediate size in Jiangjunosaurus (IVPP V14724), and the striations are not confluent with the marginal denticles except in Paranthodon (NHMUK OR47338; Raven and Maidment 2018). In Bienosaurus there is no central apical ridge, and although there is a slight swelling at the base of the crown that gives rise to cusps medially and distally, there is no well-developed cingulum. This is similar to the condition in the earliest-diverging ornithischians such as Lesothosaurus (e.g., NHMUK PV R8501), the early-diverging thyreophorans Scutellosaurus (MNA P1.175; Colbert 1981), Emausaurus (SGWG 85; Haubold 1990) and Scelidosaurus (NHMUK PV R1111; Barrett 2001) as well as the early-diverging stegosaur Huayangosaurus (IVPP V6728; Sereno and Dong 1992). In most stegosaurs and all ankylosaurs, however, a well-developed cingulum is present (Galton and Upchurch 2004; Vickaryous et al. 2004; Maidment et al. 2008). The best-preserved tooth crown has eight denticles present, but the crown is incomplete and it is likely there were originally more. This is more than Lesothosaurus (six, NHMUK PV R8501), Scelidosaurus (six, NHMUK PV R1111), Huayangosaurus (six, IVPP V6728), Stegosaurus (six, NHMUK PV R36730) and Kunbarrasaurus (seven, Leahey et al. 2015). The tooth roots are sub-cylindrical and expand mesiodistally at the base of the crown, and, based on the positioning of the tooth roots, the tooth crowns presumably overlapped slightly with an en echelon arrangement.
?Frontal: In the original description of Bienosaurus a frontal is identified and figured (Dong 2001: fig. 17.2.D). This bone is longer anteroposteriorly than transversely wide (Fig. 2A). The transverse width is greater at the posterior end than the anterior end, and there is an anteroposteriorly straight medial edge for possible contact with the left frontal. It is relatively flat transversely, but curved anteroposteriorly in medial view. Dong (2001) also mentioned that osteoderms were fused to the frontal, but this is not clear. The element is fragmentary and does not conform to the morphology common for frontals in thyreophorans based on our observations; for example, there is no obvious site for articulation with the nasal or parietal and no orbital margin. Given the overall shape of the element and its incomplete nature, it is possible that this element represents either a partial supraorbital, a lacrimal or a postorbital, but confident identification is not possible.
Fragments: Seven other fragments are present. At least two of these (Fig. 2E, F) and possibly four (Fig. 2G, H) are teeth; comparisons with Dong (2001: fig. 17.2) suggest that since the original description, several tooth crowns have broken off from the dentary and are now isolated. The other three fragments are of unknown identity; Dong (2001) suggested one to be a maxilla and another to be a pterygoid. However, it is not possible to identify these elements, although the “maxilla” could also be a nasal and one fragment may be either a small osteoderm, a fragmentary prezygapophysis or an ectopterygoid. The supraorbital mentioned by Dong (2001: fig 17.2.E) could not be located by TJR at the IVPP in June 2018 and is presumed missing.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 2. Thyreophoran dinosaur Bienosaurus lufengensis Dong, 2001 (IVPP V15311, holotype), from the Hettangian–Sinemurian (Lower Jurassic), Lower Lufeng Formation of Yunnan, China. A. Possible frontal in dorsal view. B. Possible maxilla or nasal. C. Possible osteoderm, prezygapophysis or ectopterygoid. D. Possible pterygoid. E. Tooth, broken off since previous description of specimen. F. Poorly preserved tooth. G, H. Possible teeth.
Phylogenetic analysis
Bienosaurus lufengensis was added as an OTU to the most recent phylogenetic analyses of both Ankylosauria (Arbour and Evans 2017) and Stegosauria (Raven and Maidment 2017), as well as a recent analysis of early ornithischian phylogeny (Baron et al. 2017b). Due to its fragmentary nature, it could only be coded for nine out of 177, nine out of 115 and 16 out of 227 characters, respectively. It was not added into the ornithischian phylogeny of Boyd (2015) as this has previously been shown to be inadequate for resolving the phylogenetic positioning of thyreophorans (Raven and Maidment 2018). An equal-weights parsimony analysis was performed in TNT (Goloboff et al. 2008) with a New Technology search and Sect Search, Ratchet, Drift and Tree Fusing algorithms and 10 random addition sequences. Traditional TBR Branch-Swapping was then performed on the trees held in RAM. Support for groupings was then tested using symmetric resampling with a probability of 33% and 1000 replicates on a New Technology search of existing trees.
The analysis of the Arbour and Evans (2017) dataset produced two MPTs with lengths of 568 steps and a RI of 0.66 and a CI of 0.39. A strict consensus tree of these found Bienosaurus to be in a sister-taxon relationship with the early-diverging ankylosaurid Cedarpelta, from the Lower Cretaceous Cedar Mountain Formation of Utah (Arbour and Currie 2016). Analysis of the Baron et al. (2017b) dataset produced nine MPTs with lengths of 589 steps, a RI of 0.68 and a CI of 0.43. A strict consensus tree of these placed Bienosaurus in a large polytomy at the base of Ornithischia, and a 50% Majority Rule tree found it to be in a sister-taxon relationship with Pisanosaurus, a taxon that was previously thought to be the earliest diverging ornithischian but in fact may be a silesaurid (Baron et al. 2017a; Agnolín and Rozadilla 2018). Analysis of the Raven and Maidment (2017) dataset produced four MPTs with lengths of 260.61, a RI of 0.66 and a CI of 0.62 but neither a strict consensus tree nor a majority rule tree provided any resolution. All strict consensus trees can be found in the SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app64-Raven_etal_SOM.pdf.
Discussion
Systematic position.—Despite low resolution and conflicting results in the phylogenetic analyses, the morphological characters that are present in Bienosaurus can provide an indication of its systematic position. It is unquestionably an ornithischian, possessing a buccal emargination, apicobasally short tooth crowns, overlapping tooth crowns and mesiodistally expanded tooth crowns above the tooth root, all of which were considered unambiguous synapomorphies of Ornithischia by Butler et al. (2008). Additionally, the presence of a buccal emargination, the overlapping tooth crowns and medially inset teeth are synapomorphies of Ornithischia identified by the ornithischian phylogeny of Boyd (2015). Several of these features, namely a triangular tooth crown and the presence of serrated denticles, also occur in some other Late Triassic archosaurs, including aetosaurs, pseudosuchians and silesaurids (Irmis et al. 2007) and cannot, therefore, be used to assign isolated teeth to Ornithischia. However, the presence of a labiolingual swelling of the basal tooth crown is still considered to be an unambiguous synapomorphy of ornithischians (Irmis et al. 2007), as is the possession of a buccal emargination, meaning Bienosaurus can be regarded confidently as an ornithischian dinosaur.
Bienosaurus was considered a thyreophoran by Dong (2001) and although it does not exhibit any of the synapomorphies of Thyreophora identified by Butler et al. (2008), it does possess one synapomorphy of the node that unites all thyreophorans except Lesothosaurus: the presence of cortical remodelling on skull bones (Butler et al. 2008). Additionally, the medially inset tooth row is a synapomorphy of Thyreophora in Boyd (2015) and this is also a synapomorphy, along with the presence of cortical remodelling, of all Thyreophora excluding Lesothosaurus in Raven and Maidment (2017). The sinuous tooth row in dorsal view is present in the majority of thyreophoran taxa, but this has not been included as a character in any previous phylogenetic analysis. None of the unambiguous thyreophoran or eurypodan synapomorphies proposed in the ankylosaurian analyses of Thompson et al. (2012) or Arbour and Currie (2016) can be evaluated in Bienosaurus due to a lack of material. If a supraorbital is present in Bienosaurus, as suggested by Dong (2001), then this could indicate an affinity to Thyreophoroidea (Eurypoda + Scelidosaurus) as this is a synapomorphy of the group (Butler et al. 2008). It would at least indicate that the specimen is more deeply-nested than Lesothosaurus and Emausaurus as these taxa possess “free” palpebrals, rather than homologous supraorbitals that are incorporated into the dorsal margin of the orbit (Maidment and Porro 2010).
The retention of thyreophoran symplesiomorphies, such as a Meckelian groove extending for most of the length of the dentary ramus, a tapering buccal emargination, teeth with striations that are not confluent with the marginal denticles, and a small swelling present on the tooth crowns rather than a well-developed cingulum, suggests that Bienosaurus is likely an early-diverging member of Thyreophora, which diverged prior to the origin of Eurypoda. There are, however, no eurypodan or thyreophoroidean synapomorphies in the material and so further phylogenetic placement is difficult. Bienosaurus lacks the ventral deflection of the anterior end of the dentary tooth row, which was identified as a synapomorphy of Thyreophora by Norman et al. (2004) and as a synapomorphy of Thyreophora excluding Lesothosaurus by Butler et al. (2008) and Raven and Maidment (2017). Lesothosaurus, which itself has been suggested to be a thyreophoran (Butler et al. 2008; Boyd 2015), also lacks this synapomorphy. It is possible, therefore, that Bienosaurus represents a taxon that would have been in some ways intermediate in morphology between Lesothosaurus and more deeply nested thyreophoran taxa such as Scutellosaurus. This scenario would indicate a step-wise acquisition of thyreophoran features, with a ventral deflection of the dentary appearing later than features such as a swelling at the base of the tooth crowns or a sinuous tooth row in dorsal view. The ventral deflection of the anterior end of the dentary tooth row was identified as a synapomorphy of Thyreophoroidea, by Boyd (2015) and this would indicate that Bienosaurus lies within Thyreophora but outside Thyreophoroidea.
It is also possible, however, that the anterior dentary is not sufficiently complete to fully evaluate this character. The dentary of Bienosaurus is strikingly similar to that of “Tatisaurus” and as they are from the same formation (Norman et al. 2007), it is possible that “Tatisaurus” represents the same taxon but simply has more of the anterior dentary preserved. The two specimens are of almost the same size and are comparably incomplete, with only a partial dentary present, although in Bienosaurus it is a right dentary and in “Tatisaurus” a left dentary (Norman et al. 2007). They both have a sinuous tooth row in dorsal view, although this is more pronounced in Bienosaurus. The posterior end of the dentary rami in both specimens curves slightly dorsally, and it appears that the tooth row also curves slightly along the anteroposterior length of the rami, although in “Tatisaurus” the anterior end of the dentary curves ventrally and this is not the case in Bienosaurus. Additionally, there is no evidence of cortical remodelling in “Tatisaurus”. Consequently, although the two specimens are superficially similar in morphology, they do differ in several respects. It is possible that they represent the same taxon, but the lack of comparative material between them means this cannot be determined satisfactorily, especially when the possibility if individual and taphonomic variation is taken into account.
The incompleteness of Bienosaurus also hinders the interpretations that can be drawn from the phylogenetic analyses. Although analyses of the Arbour and Evans (2017) dataset suggests that it is more parsimonious for Bienosaurus to be an ankylosaur, this is poorly supported by symmetric resampling and the sister-taxon relationship with Cedarpelta is supported by only one morphological character: the absence of a well-developed cingulum on the teeth. This feature is considered a symplesiomorphy in analyses that include more early-diverging taxa (Butler et al. 2008; Boyd 2015). Additionally, it seems unlikely that Bienosaurus is the sister-taxon of Cedarpelta given the large temporal and geographical distances that separate these two taxa, and we believe the recovery of this relationship is likely an artefact of the phylogenetic analysis. Analyses of the Raven and Maidment (2017) and Baron et al. (2017b) datasets produce little resolution, suggesting there is too much missing data or noise for a successful analysis (Kearney and Clark 2003). A phylogenetic dataset of whole-group Thyreophora may help in alleviating this problem, but the lack of scorable characters for Bienosaurus is a larger concern. A satisfactory quantitative phylogenetic analysis of the taxon will not be possible until more specimens are found that represent more of the skeleton.
Despite its fragmentary nature, it is likely that the holotype of Bienosaurus represents an early-diverging thyreophoran dinosaur and a taxon that is more deeply-nested than Lesothosaurus, but earlier diverging than Scelidosaurus and eurypodans.
Is Bienosaurus valid?—The original diagnosis proposed by Dong (2001) was based on four characters: (i) predentary short and wide; (ii) frontal thick with small bony scutes fused to dorsal surface; (iii) dentary wide as typical of ankylosaurs with ornamented lateral surface; and, (iv) teeth small and leaf-shaped with symmetrical crown and a developed cingulum. However, no predentary is preserved (see Description) and thus the validity of character (i) cannot be determined. The anterior portion of the dentary is poorly preserved and there is a small crack on the lateral surface of the anterior end and so it is possible this was misidentified as a dentary-predentary suture by Dong (2001).
Character (ii) would provide a clear indication of thyreophoran affinities, as cortical remodelling of the skull roof is a synapomorphy of Thyreophora (excluding Lesothosaurus) (Butler et al. 2008; Raven and Maidment 2017). However, given the wide distribution of this feature it cannot be autapomorphic for Bienosaurus. Moreover, the identification of the frontal is problematic; this element could be one of several different skull bones and cannot be identified confidently (see Description, above).
Character (iii), an ornamented lateral surface of the dentary, would also provide an indication of thyreophoran affinities, with Scutellosaurus (MNA P1.175; Colbert 1981) and many ankylosaurs (e.g., Pinacosaurus, IVPP V16283) possessing this feature (Vickaryous et al. 2004). As a result, this character cannot be considered autapomorphic for Bienosaurus.
Character (iv) is slightly inaccurate; a cingulum is not well-developed, as in ankylosaurs or stegosaurs, but instead is a slight swelling at the base of the crown. Assuming this to be the state originally described, character (iv) is widespread among early-diverging ornithischians (e.g., Lesothosaurus, NHMUK PV R8501) as well as the early-diverging thyreophorans Scutellosaurus (MNA P1.175; Colbert 1981), Emausaurus (SGWG 85; Haubold 1990) and Scelidosaurus (NHMUK PV R1111; Barrett 2001). This character could, therefore, be used to support the thyreophoran affinities of Bienosaurus, but cannot be used to diagnose the taxon.
None of the characters originally proposed to diagnose Bienosaurus are autapomorphic, and no further autapomorphic characters or unique character combinations could be identified given the fragmentary nature of the material. Bienosaurus is, therefore, considered a nomen dubium and the specimen represents an indeterminate thyreophoran.
Implications.—The confirmation of “Bienosaurus” as an early-diverging thyreophoran, and its removal from Ankylosauria, has important stratigraphic and biogeographic ramifications. Excluding this taxon from Eurypoda obliterates the extensive ghost lineage that would otherwise occur between “Bienosaurus” and Middle Jurassic eurypodans and a deep, cryptic split between Ankylosauria and Stegosauria during the earliest Jurassic is no longer implied. Instead, current data indicate that this split is more likely to have occurred in the late Early or early Middle Jurassic, prior to appearance of the earliest definitive ankylosaur (Callovian, Sarcolestes; Galton 1983) and stegosaur (Bajocian, Isaberrysaura; Salgado et al. 2017; Han et al. 2018).
Although eurypodans are currently unknown from Early Jurassic deposits, non-eurypodan (“basal”) thyreophorans were diverse and widespread at this time. These include: Scutellosaurus (Colbert 1981) and another indeterminate taxon (Padian 1989) from the Sinemurian–Pliensbachian Kayenta Formation of Arizona, USA; Scelidosaurus (Owen 1861) from the Sinemurian–lower Pliensbachian Charmouth Mudstone Formation of England; Emausaurus (Haubold 1990) from the Toarcian of Germany; an indeterminate thyreophoran (“Lusitanosaurus”) from the Sinemurian of Portugal (Norman et al. 2004); and Lesothosaurus from the upper Elliot and Clarens Formations of South Africa and Lesotho (Sereno 1991; Porro et al. 2015; Baron et al. 2017b), which is thought to be of Hettangian–Sinemurian age (Olsen and Galton 1984). In addition, the recently described Laquintasaura (Barrett et al. 2014), from the earliest Hettangian La Quinta Formation of Venezuela, has been identified as an early-diverging thyreophoran dinosaur in recent cladistic analyses (Baron et al. 2017b; Raven and Maidment 2017). Radioisotopic dating of the Laquintasaura bonebed suggests that this occurrence was within 0.5 Ma of the Triassic–Jurassic boundary (Barrett et al. 2014), making it the oldest known thyreophoran, and the oldest securely dated ornithischian dinosaur. An undescribed specimen from Niger has also been identified as an early-diverging thyreophoran but is awaiting full description (Nicole Ridgwell, personal communication; Ridgwell and Sereno 2010; Ridgwell 2011). This survey is completed by “Bienosaurus” and the indeterminate thyreophoran “Tatisaurus” (Norman et al. 2007) from the Lower Lufeng Formation of China, demonstrating that non-eurypodan thyreophorans achieved an almost worldwide distribution very early in their evolutionary history, although their remains are currently unknown from Antarctica, Australia, and India. This suggests the rapid diversification and dispersal of Thyreophora in either the latest Triassic or earliest Jurassic following its origin, which is likely to have been close to the Triassic–Jurassic boundary and potentially in southern Pangaea, given the early occurrences of Laquintasaura and Lesothosaurus within this region and their early divergences within the clade.
Conclusions
The fragmentary taxon “Bienosaurus lufengensis” from the Lower Jurassic Lower Lufeng Formation of Yunnan, China, is re-described and found to be an indeterminate non-eurypodan “basal” thyreophoran dinosaur, which might represent the same taxon as the also indeterminate “Tatisaurus” from the same formation. This implies the removal of a previously proposed deep, cryptic ghost lineage between “Bienosaurus” and the first definitive eurypodans (ankylosaurs or stegosaurs) in the Middle Jurassic. It also helps to demonstrate that thyreophorans achieved an almost global distribution early in their evolutionary history, following rapid diversification and dispersal in either the latest Triassic or earliest Jurassic.
Acknowledgements
We thank Zheng Fang (IVPP) for access to specimens and Richard Butler (University of Birmingham, UK) for photographs of Tatisaurus. We thank the editor, Stephen Brusatte (University of Edinburgh, UK), and the reviewers Lucy Leahey (University of Queensland, St Lucia Queensland, Australia) and Peter Galton (University of Bridgeport, USA) for comments that improved the manuscript. Funding for TJR was provided by a University of Brighton scholarship, the Geologist’s Association New Researchers’ Award, the Geological Society’s Daniel Pigeon Fund, the Jurassic Foundation and the Universities’ China Committee in London; PMB’s trips to China were funded by the Royal Society and the Earth Sciences Departmental Investment Fund (NHMUK); XX was supported by National Natural Sciences Foundation of China (41688103).
References
Agnolín, F.L. and Rozadilla, S. 2018. Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina. Journal of Systematic Palaeontology 16: 853–879. Crossref
Arbour, V.M. 2014. Systematics, Evolution, and Biogeography of the Ankylosaurid Dinosaurs. 470 pp. Ph.D. Thesis, University of Alberta, Edmonton.
Arbour, V.M. and Currie, P.J. 2016. Systematics, phylogeny, and palaeobiogeography of the ankylosaurid dinosaurs. Journal of Systematic Palaeontology 14: 385–444. Crossref
Arbour, V.M. and Evans, D.C. 2017. A new ankylosaurine dinosaur from the Judith River Formation of Montana, USA, based on an exceptional skeleton with soft tissue preservation. Royal Society Open Science 4: [published online, https://doi.org/10.1098/rsos.161086] Crossref
Baron, M.G., Norman, D.B., and Barrett, P.M. 2017a. Baron et al. reply. Nature 551: E4–E5. Crossref
Baron, M.G., Norman, D.B., and Barrett, P.M. 2017b. Postcranial anatomy of Lesothosaurus diagnosticus (Dinosauria: Ornithischia) from the Lower Jurassic of southern Africa: implications for basal ornithischian taxonomy and systematics. Zoological Journal of the Linnean Society 179: 125–168. Crossref
Barrett, P.M. 2001. Tooth wear and possible jaw action of Scelidosaurus harrisonii Owen and a review of feeding mechanisms in other thyreophoran dinosaurs. In: K. Carpenter (ed.), The Armored Dinosaurs (Life of the Past), 25–54. Indiana University Press, Bloomington.
Barrett, P.M. and Xu, X. 2005. A reassessment of Dianchungosaurus lufengensis Yang, 1982a, an enigmatic reptile from the Lower Lufeng Formation (Lower Jurassic) of Yunnan Province, People’s Republic of China. Journal of Paleontology 79: 981–986. Crossref
Barrett, P.M., Butler, R.J., Mundil, R., Scheyer, T.M., Irmis, R.B., and Sanchez-Villagra, M.R. 2014. A palaeoequatorial ornithischian and new constraints on early dinosaur diversification. Proceedings of the Royal Society B 281: 2014.1147. Crossref
Boyd, C.A. 2015. The systematic relationships and biogeographic history of ornithischian dinosaurs. PeerJ 3: e1523. Crossref
Butler, R.J., Upchurch, P., and Norman, D.B. 2008. The phylogeny of the ornithischian dinosaurs. Journal of Systematic Palaeontology 6: 1–40. Crossref
Colbert, E.H. 1981. A primitive ornithischian dinosaur from the Kayenta Formation of Arizona. Museum of Northern Arizona Bulletin 53: 1–61.
Dong, Z. 2001. Primitive armored dinosaur from the Lufeng Basin, China. In: D.H. Tank and K. Carpenter (eds.), Mesozoic Vertebrate Life, 237–243. Indiana University Press, Bloomington.
Galton, P.M. 1983. Armored dinosaurs (Ornithischia: Ankylosauria) from the Middle and Upper Jurassic of Europe. Palaeontographica Abteilung A 182: 1–25.
Galton, P.M. and Upchurch, P. 2004. Stegosauria. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, Second Edition, 343–362. University of California Press, Berkeley. Crossref
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Han, F.L., Forster, C.A., Xu, X., and Clark, J.M. 2018. Postcranial anatomy of Yinlong downsi (Dinosauria: Ceratopsia) from the Upper Jurassic Shishugou Formation of China and the phylogeny of basal ornithischians. Journal of Systematic Palaeontology 16: 1159–1187. Crossref
Haubold, H. 1990. Ein neuer Dinosaurier (Ornithischia, Thyreophora) aus dem unteren Jura des nordlichen Mitteleuropa. Revue de paleobiologie 9: 1–149.
Irmis, R.B. and Knoll, F. 2008. New ornithischian dinosaur material from the Lower Jurassic Lufeng Formation of China. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 247: 117–128. Crossref
Irmis, R.B., Parker, W.G., Nesbitt, S.J., and Liu, J. 2007. Early ornithischian dinosaurs: the Triassic record. Historical Biology 19: 3–22. Crossref
Kearney, M. and Clark, J.M. 2003. Problems due to missing data in phylogenetic analyses including fossils: a critical review. Journal of Vertebrate Paleontology 23: 263–274. Crossref
Leahey, L.G., Molnar, R.E., Carpenter, K., Witmer, L.M., and Salisbury, S.W. 2015. Cranial osteology of the ankylosaurian dinosaur formerly known as Minmi sp. (Ornithischia: Thyreophora) from the Lower Cretaceous Allaru Mudstone of Richmond, Queensland, Australia. PeerJ 3: e1475. Crossref
Lucas, S.G. 2001. Chinese Fossil Vertebrates. 386 pp. Columbia University Press, New York. Crossref
Luo, Z.X. and Wu, X. 1994. The small vertebrate fauna of the Lower Lufeng Formation, Yunnan, China. In: N.C. Fraser (ed.), The Shadow of the Dinosaurs: Early Mesozoic Tetrapods, 251–270. Cambridge University Press, Cambridge.
Maidment, S.C.R. and Porro, L.B. 2010. Homology of the palpebral and origin of supraorbital ossifications in ornithischian dinosaurs. Lethaia 43: 95–111. Crossref
Maidment, S.C.R., Norman, D.B., Barrett, P.M., and Upchurch, P. 2008. Systematics and Phylogeny of Stegosauria (Dinosauria: Ornithischia). Journal of Systematic Palaeontology 6: 367–407. Crossref
Nopcsa, F. 1915. Die Dinosaurier der Siebenbürgischen Landesteile Ungarns. Mitteilungen aus dem Jahrbuche der Königlich ungarischen Geologischen Anstalt 23: 1–26.
Norman, D.B. 1984. A systematic reappraisal of the reptile order Ornithischia. In: W.-E. Reif and F. Westphal (eds.), Third Symposium on Mesozoic Terrestrial Ecosystems, short papers, 157–162. Attempto Verlag, Tubingen.
Norman, D.B., Butler, R.J., and Maidment, S.C.R. 2007. Reconsidering the status and affinities of the ornithischian dinosaur Tatisaurus oehleri Simmons, 1965. Zoological Journal of the Linnean Society 150: 865–874. Crossref
Norman, D.B., Witmer, L.M. and Weishampel, D.B. 2004. Basal Thyreophora. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, Second Edition, 335–342. University of California Press, Berkeley. Crossref
Olsen, P.E. and Galton, P.M. 1984. A review of the reptile and amphibian assemblages from the Stormberg of Southern Africa, with special emphasis on the footprints and the age of the Stormbeg. Palaeontologia Africana 25: 87–110.
Olsen, P.E., Kent, D.V., and Whiteside, J.H. 2011. Implications of the Newark Supergroup-based astrochronology and geomagnetic polarity time scale (Newark-APTS) for the tempo and mode of the early diversification of the Dinosauria. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 201–229. Crossref
Owen, R. 1842. Report on British fossil reptiles. Reports of the British Association for the Advancement of Science 11: 60–204.
Owen, R. 1861. Monograph of the fossil Reptilia of the Liassic formations. Part 1. A monograph of a fossil dinosaur (Scelidosaurus harrisonii Owen) of the Lower Lias. Palaeontographical Society Monographs 13: 1–13. Crossref
Padian, K. 1989. Presence of the dinosaur Scelidosaurus indicates Jurassic age for the Kayenta Formation (Glen Canyon Group, northern Arizona). Geology 17 (5): 438. Crossref
Parish, J.C. 2005. The Evolution and Palaeobiology of the Armoured Dinosaurs. 471 pp. Ph.D. Thesis, University of Oxford, Oxford.
Porro, L.B., Witmer, L.M., and Barrett, P.M. 2015. Digital preparation and osteology of the skull of Lesothosaurus diagnosticus (Ornithischia: Dinosauria). PeerJ 3: e1494. Crossref
Raven, T.J. and Maidment, S.C.R. 2017. A new phylogeny of Stegosauria (Dinosauria, Ornithischia). Palaeontology 60: 401–408. Crossref
Raven, T.J. and Maidment, S.C.R. 2018. The systematic position of the enigmatic thyreophoran dinosaur Paranthodon africanus, and the use of basal exemplifiers in phylogenetic analysis. PeerJ 6: e4529. Crossref
Ridgwell, N. 2011. A basal thyreophoran (Dinosauria, Ornithischia) from Africa clarifies the early evolution of armored dinosaurs. In: Program and Abstracts, 71st Annual Meeting of the Society of Vertebrate Paleontology, Las Vegas, NV, Program and Abstracts. Journal of Vertebrate Paleontology 31 (Supplement 2): 179. Crossref
Ridgwell, N. and Sereno, P.C. 2010. A basal thyreophoran (Dinosauria, Ornithischia) from the Tioraren Formation of Niger. In: 70th Annual Meeting of the Society of Vertebrate Paleontology, Pittsburgh, PA, Program and Abstracts, 150A. Society of Vertebrate Paleontology, Pittsburgh.
Russell, D. and Dong, Z. 1993. A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People’s Republic of China. Canadian Journal of Earth Sciences 30: 2163–2173. Crossref
Salgado, L., Canudo, J.I., Garrido, A.C., Moreno-Azanza, M., Martínez, L.C.A., Coria, R.A., and Gasca, J.M. 2017. A new primitive Neornithischian dinosaur from the Jurassic of Patagonia with gut contents. Scientific Reports 7: 42778. Crossref
Seeley, H.G. 1887. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London 43: 165–171. Crossref
Sereno, P.C. 1991. Lesothosaurus, “fabrosaurids”, and the early evolution of ornithischia. Journal of Vertebrate Paleontology 11: 168–197. Crossref
Sereno, P.C. and Dong, Z. 1992. The skull of the basal stegosaur Huayangosaurus taibaii and a cladistic diagnosis of stegosauria. Journal of Vertebrate Paleontology 12: 318–343. Crossref
Sun, A., Cui, G., Li, Y., and Wu, X. 1985. A verified list of the lufeng saurischian fauna. Vertebrata PalAsiatica 23: 1–12.
Thompson, R.S., Parish, J.C., Maidment, S.C.R., and Barrett, P.M. 2012. Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora). Journal of Systematic Palaeontology 10: 301–312. Crossref
Vickaryous, M.K., Maryańska, T., and Weishampel, D.B. 2004. Ankylosauria. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, Second Edition, 363–392. University of California Press, Berkeley. Crossref
Weishampel, D.B., Barrett, P.M., Coria, R.A., Le Loeuff, J., Xu, X., Zhao, X.-J., Sahni, A., Gomani, E.M.P., and Noto, C.R. 2004. Dinosaur distribution. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, Second Edition, 536–605. University of California Press, Berkeley. Crossref
Yang, Z.J. 1982. A new genus of dinosaur from Lufeng County, Yunnan Province [in Chinese]. In: M.Z. Zhou (ed.), Collected Works of Yang Zhongjian, 38–42. Academia Sinica, Beijing.
Young, C.C. 1951. The Lufeng Saurischian fauna in China. Palaeontologica Sinica NS C13: 1–96.
Acta Palaeontol. Pol. 64 (2): 335–342, 2019
https://doi.org/10.4202/app.00577.2018