

The first ornithosuchid from Brazil and its macroevolutionary and phylogenetic implications for Late Triassic faunas in Gondwana
RODRIGO T. MÜLLER, M. BELÉN VON BACZKO, JULIA B. DESOJO, and STERLING J. NESBITT
Müller, R.T., Baczko, M.B. von, Desojo, J.B., and Nesbitt, S.J. 2020. The first ornithosuchid from Brazil and its macroevolutionary and phylogenetic implications for Late Triassic faunas in Gondwana. Acta Palaeontologica Polonica 65 (1): 1–10.
Ornithosuchidae is one of the most enigmatic clades of Triassic pseudosuchians. The group is composed by three carnivorous species that were excavated from Upper Triassic beds of Scotland and Argentina. We describe the first ornithosuchid from the Upper Triassic sediments of Brazil and explore its phylogenetic affinities and implications for the evolution of the group. Dynamosuchus collisensis gen. et sp. nov. was found as the sister taxon of the Argentinean form Venaticosuchus rusconii. These relationships reject a potential endemic radiation of ornithosuchids from the Ischigualasto-Villa Unión Basin and would better support multiple diversification events. Our findings with ornithosuchids is consistent with the pattern reported for proterochampsid and erpetosuchid archosauriforms from Ischigualasto-Villa Unión and the Paraná basins. In addition, the presence of an ornithosuchid in the Late Triassic of Brazil suggests that ornithosuchids were more widespread than previously thought in the southern hemisphere. The new ornithosuchid further demonstrates a faunistic link between the Argentinean and Brazilian basins during the Carnian. Finally, the discovery of the new species provides the first clue of a putative necrophagous vertebrate from the oldest dinosaur-bearing beds and expands our knowledge regarding the trophic structure of the Late Triassic of Brazil.
Key words: Archosauria, Pseudosuchia, Ornithosuchidae, biogeography, phylogeny, Carnian, Santa Maria Formation, South America.
Rodrigo T. Müller [rodrigotmuller@hotmail.com], Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia, Universidade Federal de Santa Maria, São João do Polêsine, RS 598, 97230-000, Brazil.
M. Belén Von Baczko [belen_vb13@yahoo.com.ar] and Julia B. Desojo [julideso2@gmail.com], División Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque s/n (B1900FWA), La Plata, Buenos Aires, Argentina; Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Godoy Cruz 2290 (C1425FQB), Ciudad Autónoma de Buenos Aires, Argentina.
Sterling J. Nesbitt [sjn2104@vt.edu], Department of Geosciences, Virginia Tech, Blacksburg, Virginia 24061, USA.
Received 2 July 2019, accepted 18 November 2019, available online 31 January 2020.
Copyright © 2020 R.T. Müller et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The radiation of pseudosuchian archosaurs during the Triassic Period is characterized by the origin and extinction of several peculiar and disparate clades, such as Aetosauria (Desojo et al. 2013; Parker 2016), Erpetosuchidae (Benton and Walker 2002; Nesbitt and Butler 2013; Ezcurra et al. 2017; Lacerda et al. 2018), Gracilisuchidae (Butler et al. 2014; Lecuona et al. 2017), and Ornithosuchidae (Walker 1964; Bonaparte 1970; Baczko et al. 2014; Baczko and Desojo 2016; Baczko 2018). The latter is one of the most enigmatic clades. Coined by Huene in 1908, Ornithosuchidae has a long and controversial taxonomic history. The clade has been found in different phylogenetic positions across Archosauria (e.g., Gauthier 1986; Sereno 1991; Nesbitt 2011; Ezcurra 2016). Nevertheless, most hypotheses following Sereno (1991) converged on a position near the base of Pseudosuchia. Three species form the clade: Ornithosuchus woodwardi (Newton, 1894), from the late Carnian–early Norian, the Lossiemouth Sandstone Formation, Scotland (Newton 1894; Walker 1964); Venaticosuchus rusconii (Bonaparte, 1970) from the late Carnian of the Ischigualasto Formation, Argentina (Baczko et al. 2014; Baczko 2018); and Riojasuchus tenuisceps (Bonaparte, 1967) from the Norian of the Los Colorados Formation, Argentina (Baczko and Desojo 2016; Baczko et al. 2020). All members of this group are carnivorous, putatively scavengers and facultative bipedal during fast gaits (Walker 1964; Baczko and Ezcurra 2013; Baczko 2018). Nevertheless, the fossil record of the group is geographically limited and so far, no remains outside Ischigualasto-Villa Union Basin (Argentina) or the Lossiemouth Sandstone Formation (Scotland) have been identified, thus leaving a large gap in their potential biogeographic distribution. Here, we describe the first partial skeleton of an ornithosuchid from the Upper Triassic sediments of Brazil (Fig. 1) and explore its phylogenetic affinities and implications for the evolution of the group.
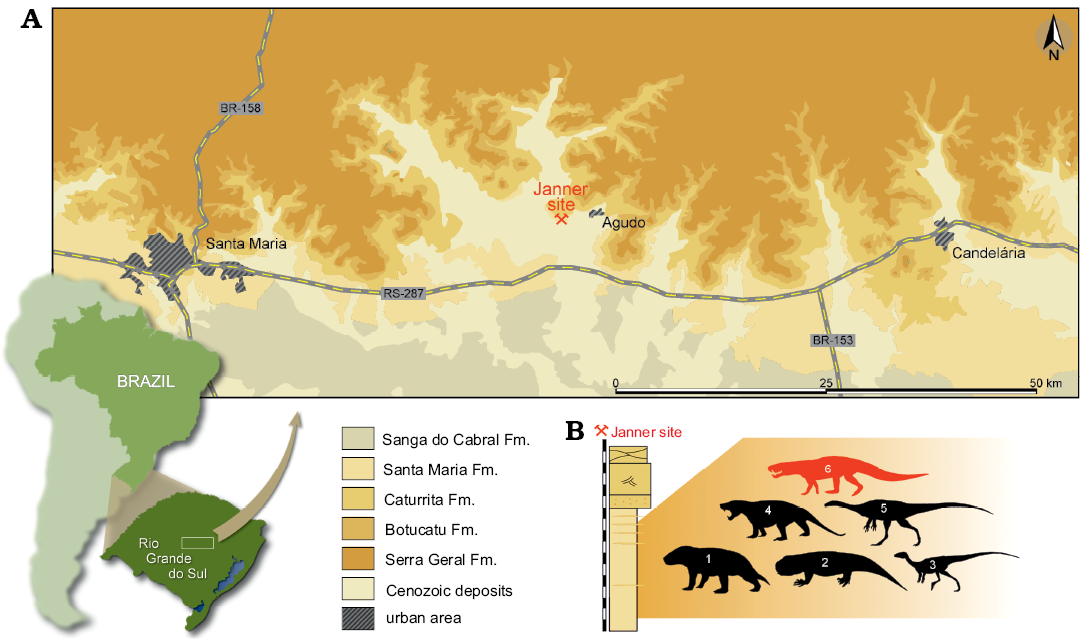
Fig. 1. A. Location map of the Janner site and the surface distribution of the geologic units in the area (modified from Müller et al. 2017). B. Stratigraphic column of the Janner site (modified from Da-Rosa 2015) depicting its fossiliferous content: 1, Exaeretodon; 2, Hyperodapedon; 3, Pampadromaeus; 4, Trucidocynodon; 5, Bagualosaurus; and 6, the new ornithosuchid.
Institutional abbreviations.—CAPPA/UFSM, Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia/Universidade Federal de Santa Maria, São João do Polêsine, Rio Grande do Sul, Brazil; CRILAR-Pv, Centro Regional de Investigaciones y Transferencia Tecnológica de La Rioja, Paleontología de Vertebrados, Anillaco, La Rioja, Argentina; NHMUK PV, Natural History Museum, London, UK; PIN, Paleontological Institute of the Russian Academy of Sciences, Moscow, Russia; PIMUZ, Paläontologisches Institut und Museum der Universität Zurich, Switzerland; PULR, Paleontología, Universidad Nacional de La Rioja, La Rioja, Argentina; PVL, Paleontología de Vertebrados, Instituto Miguel Lillo, Tucumán, Argentina; PVSJ, División de Paleontología de Vertebrados del Museo de Ciencias Naturales y Universidad Nacional de San Juan, San Juan, Argentina; SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany; TTUP, Museum of Texas Tech University, Lubbock, Texas, USA; UFRGS, Universidade Federal do Rio Grande do Sul, Porto Alegre, Rio Grande do Sul, Brazil.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in ZooBank: urn:lsid:zoobank.org:pub:A90DC693-B65F-472D-AA40-3C608C00 B8D5
Material and methods
In order to test its phylogenetic affinities and potential implications for the understanding of pseudosuchian evolution, the new taxon was scored in the data matrix of Ezcurra et al. (2017). This data matrix was employed because of the large sample of erpetosuchids included, whose importance is crucial for this study because they were proposed as sister group of ornithosuchids (Ezcurra et al. 2017; Lacerda et al. 2018). The Brazilian erpetosuchid Pagosvenator candelariensis (Lacerda et al. 2018) which was published after Ezcurra et al. (2017) was also included here. Therefore, the final data matrix includes 676 characters and 109 terminal taxa. Our data matrix was subjected to an equally weighted parsimony analysis in TNT v. 1.1 (Goloboff et al. 2008). The characters treated by Ezcurra et al. (2017) as additive (1, 2, 7, 10, 17, 19, 20, 21, 28, 29, 36, 40, 42, 50, 54, 66, 71, 75, 76, 122, 127, 146, 153, 156, 157, 171, 176, 177, 187, 202, 221, 227, 263, 266, 279, 283, 324, 327, 331, 337, 345, 351, 352, 354, 361, 365, 370, 377, 379, 398, 410, 424, 430, 435, 446, 448, 454, 458, 460, 463, 472, 478, 482, 483, 489, 490, 504, 510, 516, 529, 537, 546, 552, 556, 557, 567, 569, 571, 574, 581, 582, 588, 648, 652, and 662) were also treated as such in our analysis. Petrolacosaurus kansensis was used to root the most parsimonious trees (MPTs), which were recovered with a “Traditional search” (random addition sequence + tree bisection reconnection) with 1000 replicates of Wagner trees (with random seed = 0), and using tree bisection reconnection and branch swapping (holding 10 trees save per replicate).
Systematic palaeontology
Archosauria Cope, 1869
(sensu Gauthier and Padian 1985)
Pseudosuchia Zittel, 1887–1890
(sensu Gauthier and Padian 1985)
Ornithosuchidae Huene, 1908
(sensu Sereno 1991)
Genus Dynamosuchus nov.
ZooBank LSID: urn:lsid:zoobank.org:act:B3DA19CB-0750-466F-A6C1-97A14114A41B
Type species: Dynamosuchus collisensis gen. et sp. nov., monotypic, see below.
Etymology: From Greek (transliterated) dynamis, power and soukhos, crocodile, referring to the strong bite force inferred to ornithosuchids.
Diagnosis.—Same as for the only known species.
Dynamosuchus collisensis sp. nov.
Fig. 2.
ZooBank LSID: urn:lsid:zoobank.org:act:4B1FFAFB-6287-4448-9E34-B7976D307C80
Etymology: From Latin collis, hill, reflecting the fact that the holotype was collected from the Janner site, which is located at the base of the Agudo Hill (Morro Agudo in Portuguese).
Holotype: CAPPA/UFSM 0248, a partial skeleton including the premaxillae, left maxilla, frontals, right postfrontal, right parietal, squamosals, quadratojugals, quadrates, parabasisphenoid, both hemimandibles, some cervical, dorsal, sacral and caudal vertebrae, several ribs, gastralia, osteoderms, left scapula, both forelimbs, left ilium, right pubis, left femur, right tibia, and left fibula.
Type locality: Janner site (29°39′10.89″ S, 53°17′34.20″ W), Agudo, Rio Grande do Sul, Brazil (Fig. 1).
Type horizon: Santa Maria Formation; Candelária Sequence, Paraná Basin. The presence of the genus Hyperodapedon and Exaeretodon at the Janner site (Langer et al. 2007) supports a Carnian age based on correlation with radioisotope dating of another Brazilian site (Langer et al. 2018) and Argentine beds from the La Peña Member of the Ischigualasto Formation (Martínez et al. 2011) with the same taxa.
Diagnosis.—Dynamosuchus collisensis differs from all other known ornithosuchids based on a unique combination of character states (* autapomorphy): dorsal process of the quadratojugal mostly dorsally oriented and with the lateral surface densely covered by rugosities; additional foramen on the medial wall of the quadrate foramen*; parabasisphenoid component of the basal tubera ventrally separated from the basipterygoid process by a slight notch at the midline; main axis of the parabasisphenoid process posteroventrally directed; surangular with a conspicuous lateral shelf and an anteroposteriorly wide surangular foramen; anterior dorsal vertebrae with anterior and posterior centrodiapophyseal laminae and without prezygodiapophyseal and postzygodiapophyseal laminae; dorsal body osteoderms with an external pattern of dense crests and grooves; and iliofibularis tubercle of the fibula located within the proximal half of the bone.
Ornithosuchus woodwardi differs from Dynamosuchus collisensis in that O. woodwardi has (for comparative illustrations see the SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app65-Muller_etal_SOM.pdf): dorsal process of the quadratojugal is more anteriorly projected (O. woodwardi, 45°; D. collisensis, 90°); no additional foramen on the medial wall of the quadrate foramen; the lateral surface of the angular has ornamentation; less laterally extended surangular shelf; presence of prezygodiapophyseal and postzygodiapophyseal lamina in the anterior dorsal vertebrae; presence of a fossa ventral to the neurocentral suture on the lateral side of the dorsal centra; presence of an accessory process on the anterior margin of the neural arch of the middle caudal vertebrae; and a less expanded distal end of the radius.
Venaticosuchus rusconii differs from D. collisensis in that V. rusconii has: absence of foramina on the lateral surface of the maxilla; no additional foramen on the medial wall of the quadrate foramen; parabasisphenoid component of the basal tubera ventrally separated from the basipterygoid process by a notch that forms an acute angle; basipterygoid processes more transversely separated from each other in comparison to D. collisensis; basipterygoid processes are ventrally directed with a semicircular shape in lateral view; and absence of the surangular foramen (autapomorphy of V. rusconii; Baczko et al. 2014).
Riojasuchus tenuisceps differs from D. collisensis in that R. tenuisceps has: absence of foramina on the lateral surface of the maxilla; anterior and ventral margins of the antorbital fossa almost meet with the anterior and ventral edges of the maxilla; lateral surface of the quadratojugal is smooth; dorsal process of the quadratojugal is more anteriorly projected (c. 45°); no additional foramen on the medial wall of the quadrate foramen; ventral process of the squamosal anteroposteriorly larger proportionally; absence of a semilunar depression on the posterolateral surface of the parabasisphenoid; absence of the anterior and posterior centrodiapophyseal laminae of the anterior dorsal vertebrae; osteoderms with smooth ornamentation; internal tuberosity of the humerus is more pronounced; postacetabular process of the ilium is proportionally anteroposteriorly longer; iliac blade with a concave dorsal margin in lateral view; and iliofibularis tubercle of the fibula more distally located.
Description.—The body of the premaxilla of Dynamosuchus collisensis is ventrally deflected at the anterior portion (= downturned) and bears three teeth, as typical of ornithosuchids (Baczko and Ezcurra 2013), and the anterior tip of the premaxilla extends further anteriorly than the anterior tip of the dentary (Fig. 2A–C). The elongated palatal process of the premaxilla of D. collisensis bears a ridge at its ventral margin, like in Venaticosuchus rusconii (PVL 2578; Baczko et al. 2014). There is a lateral edentulous constriction at the posterior process of the premaxilla, where two caniniform teeth from the dentary fit. The lateral surface of the maxilla is pierced by several foramina (see the Supplementary Online Material for additional figures), which are not recognized in V. rusconii (PVL 2578; Baczko et al. 2014) and R. tenuisceps (PVL 3827, PVL 3828; Baczko and Desojo 2016), but present in Ornithosuchus woodwardi (NHMUK PV R 3143; Walker 1964). The anterior and ventral portions of the antorbital fossa do not coincide with the anterior and ventral edges of the maxilla in D. collisensis, whereas in R. tenuisceps (PVL 3827, PVL 3828) these margins almost coincide with each other (Baczko and Desojo 2016). There are six tooth positions preserved; however, the total number of maxillary teeth of D. collisensis is uncertain, but probably does not exceeds seven or eight teeth based on the preserved length of the maxilla, the length of the hemimandible, and through comparisons with other ornithosuchids.
The right frontal, postfrontal, and anterior portion of the parietal were preserved in articulation showing a markedly constricted interorbital area and delimiting the medial half of the supratemporal fenestra (SOM: fig. S2). A depression for the olfactory bulbs occurs on the ventral surface of the anterior half of the frontal. Like in R. tenuisceps (PVL 3827), the supratemporal fenestra is anteroposteriorly short, approximately half of the orbit, whereas in Saurosuchus galilei (PVSJ 32; Alcober 2000), Prestosuchus chiniquensis (UFRGS-PV-0629-T; Mastrantonio et al. 2019), Tarjadia ruthae (CRILAR-Pv 495; Ezcurra et al. 2017) and in aetosaurs (e.g. Neoaetosauroides engaeus; PVL 5698; Desojo and Báez 2007), the supratemporal fenestra is equivalent to the length of the orbit. The ventral process of the squamosal of D. collisensis is elongated and slender (Fig. 2A), resembling that of O. woodwardi (NHMUK PV R 2409, NHMUK PV R 3562), whereas in R. tenuisceps (PVL 3827) it is proportionally wider anteroposteriorly. There is a longitudinally oriented sulcus bounded medially by a sharp crest on the dorsal surface of the squamosal of D. collisensis. The general morphology of the quadratojugal of D. collisensis resembles that of V. rusconii (PVL 2578), where the dorsal process is less anteriorly inclined than in R. tenuisceps (PVL 3827). Like in O. woodwardi (NHMUK PV R 3142), the lateral surface of the quadratojugal of D. collisensis bears a strongly rugose surface (Fig. 2D), whereas in other ornithosuchids the homologous surface is smooth (Baczko et al. 2018). The quadrate foramen lies between the medial margin of the quadratojugal and the lateral margin of the quadrate. In addition, there is a foramen piercing the wall that forms the medial margin of the quadrate foramen in the quadrate (Fig. 2D). Although present in some archosauriforms (e.g., Sarmatosuchus otschevi; PIN 2865/68; Ezcurra 2016), this trait was not reported previously for ornithosuchids.
There is a shallow semilunar depression on the posterolateral surface of the parabasisphenoid of D. collisensis (Fig. 2E), a trait shared with V. rusconii (PVL 2578; Baczko et al. 2014). Like in R. tenuisceps (PVL 3827), in lateral view, the parabasisphenoid component of the basal tubera of D. collisensis is ventrally separated from the basipterygoid process by a gentle notch, as in most of the pseudosuchians (Fig. 2E). On the other hand, in V. rusconii (PVL 2578) there is a more acute angle (about to 40°) formed by the main axis of the basal tubera and the basipterygoid process in lateral view, whereas in D. collisensis the angle is wider (about to 60°). The basipterygoid processes of V. rusconii (PVL 2578) are more transversely separated from each other when compared to D. collisensis. Moreover, in V. rusconii (PVL 2578) the basipterygoid processes are ventrally directed with a semicircular shape in lateral view (Baczko et al. 2014), whereas in D. collisensis, the processes lie posteroventrally, which results in a more elongated ventral margin in comparison. The basipterygoid recess of D. collisensis is well-excavated differing from the shallower recess seen in R. tenuisceps (PVL 3827) and in aetosaurs (e.g., Neoaetosauroides engaeus PVL 5698, Desmatosuchus spurensis TTUP 9024).
The lower jaws of D. collisensis are anteroposteriorly shorter than the skull (Fig. 2A–C). The symphysis is approximately 35% of the mandible length (synapomorphic character state of ornithosuchids; Sereno 1991) and the external mandibular fenestra is also anteroposteriorly longer than tall in comparison with other ornithosuchids. The dorsal margin of the anterior portion of the dentary is elevated relative to the more posterior portion. The splenial forms most of the medial portion of the lower jaw and lacks any foramen. The dorsoventrally slender angular is anteroposteriorly elongated, and delimits the posterior ¾ of the ventral margin of the external mandibular fenestra. Posteriorly, the angular meets the surangular, which delimits the posterodorsal and dorsal margin of the fenestra. There is a wide surangular foramen dorsal to the contact with the angular (Fig. 2A). Such structure occurs in the other ornithosuchids, except in V. rusconii (PVL 2578; Baczko et al. 2014). Dynamosuchus collisensis bears a well-developed surangular shelf, which resembles the sharp shelf of R. tenuisceps (PVL 3827; Baczko and Desojo 2016) and V. rusconii (PVL 2578; Baczko 2018), whereas in O. woodwardi (NHMUK PV R 2409) the shelf is less pronounced. This shelf is also present in proterochampsids, erpetosuchids, and gracilisuchids (Ezcurra 2016; Ezcurra et al. 2017).
The axial neural spine is anteroposteriorly longer than the neural spine of the other preserved post-axial cervical vertebrae. At the ventral surface of the postzygapophysis of the cervical vertebrae there is a posterolaterally facing fossa (Fig. 2F), a trait unusual in pseudosuchians, except by Gracilisuchus stipanicicorum (PULR 08; Ezcurra 2016). The cervical neural spines are tall relative to the neural arch, straight, and possess transverse expansions at their dorsal end, forming “spine tables”. The cervical centra have a strongly developed ventral keel that extends longitudinally and project ventral to the margin of the centra (Fig. 2G). This condition is uncommon within pseudosuchians being present only in R. tenuisceps (PVL 3827), Erpetosuchus granti (NHMUK PV 3139), Stagonolepis robertsoni (NHMUK PV 4784), and Batrachotomus kupferzellensis (SMNS 80284). The anterior dorsal vertebrae (Fig. 2I) of D. collisensis have an anterior and posterior centrodiapophyseal lamina, absent in R. tenuisceps (PVL 3827) and unknown in V. rusconii, but lack prezygodiapophyseal and postzygodiapophyseal lamina. The dorsal vertebrae of D. collisensis lack hyposphene-hypantrum accessory intervertebral articulation similar to R. tenuisceps (PVL 3827). The condition is uncertain in other ornithosuchids. The centra of D. collisensis lack a fossa just ventral to the neurocentral suture like in R. tenuisceps (PVL 3827), whereas a poorly rimmed fossa occurs in O. woodwardi (NHMUK PV R 3916; Walker 1964). The preserved dorsal vertebrae D. collisensis do not preserve the distal tip of the neural spine. The neural spine of the caudal vertebra is posterodorsally directed and bears a pointed posterior projection. Dynamosuchus collisensis lacks an accessory laminar process on the anterior margin of the caudal vertebrae, differing from O. woodwardi (NHMUK PV R 3561).
The preserved paramedian osteoderms are square in dorsal view and strongly ornamented by radial ridges and elongated grooves (Fig. 2H), like in O. woodwardi (NHMUK PV R 3916; Walker 1964) whereas the same surface of the osteoderms of R. tenuisceps (PVL 3827) bear a gentle radial pattern (Baczko et al. 2020). Nevertheless, D. collisensis shares with the other ornithosuchids the presence of a prominence on the dorsolateral surface of the osteoderms. In D. collisensis, this dorsolateral prominence is marked by a rugose surface and, in some elements, it has a pointed dorsal tip. The dorsolateral prominence becomes less developed progressively in posterior osteoderms of the cervical and dorsal series. A longitudinally oriented ridge is absent on the dorsal surface of the osteoderms, such as in other ornithosuchids. The anterior margin of the osteoderms is straight and lacks an anterior process or an unornamented anterior articular lamina. The posterior margin is concave in dorsal view (Fig. 2H). The ventral surface is smooth and ventrally convex in anterior view. The length of the osteoderms indicate that there is a pair of osteoderms per vertebra, forming two paramedial rows along the dorsal surface of the body (unknown for the tail).
The acromion process of the scapula gently rises from the anterior margin of the scapular blade at an angle of approximately 120°, resembling the condition of O. woodwardi (NHMUK PV R 3916). In contrast, D. collisensis has a well-developed acromion process, which is nearly flat in O. woodwardi (NHMUK PV R 3916); the condition of D. collisensis resembles that of R. tenuisceps (PVL 3828). The humerus (Fig. 2L) is robust, about 0.6 times the total length of the femur (0.65–0.75 in R. tenuisceps), similar to the ration in some aetosaurs and Ticinosuchus ferox (PIMUZ 2817), among pseudosuchians (Ezcurra 2016). The proximal end of the humerus is widely expanded with a moderately developed internal tuberosity, which is less pronounced in D. collisensis than in R. tenuisceps (PVL 3826). The deltopectoral crest of D. collisensis is well-developed and restricted to the proximal third of the bone. Dynamosuchus collisensis lacks an ectepicondylar flange or it is poorly developed. The ulna (Fig. 2M) bears a moderately developed olecranon process and tappers towards its distal end, whereas the radius expands distally. Metacarpal I is the widest, whereas the metacarpal III is the longest of the metacarpals preserved (Fig. 2N).
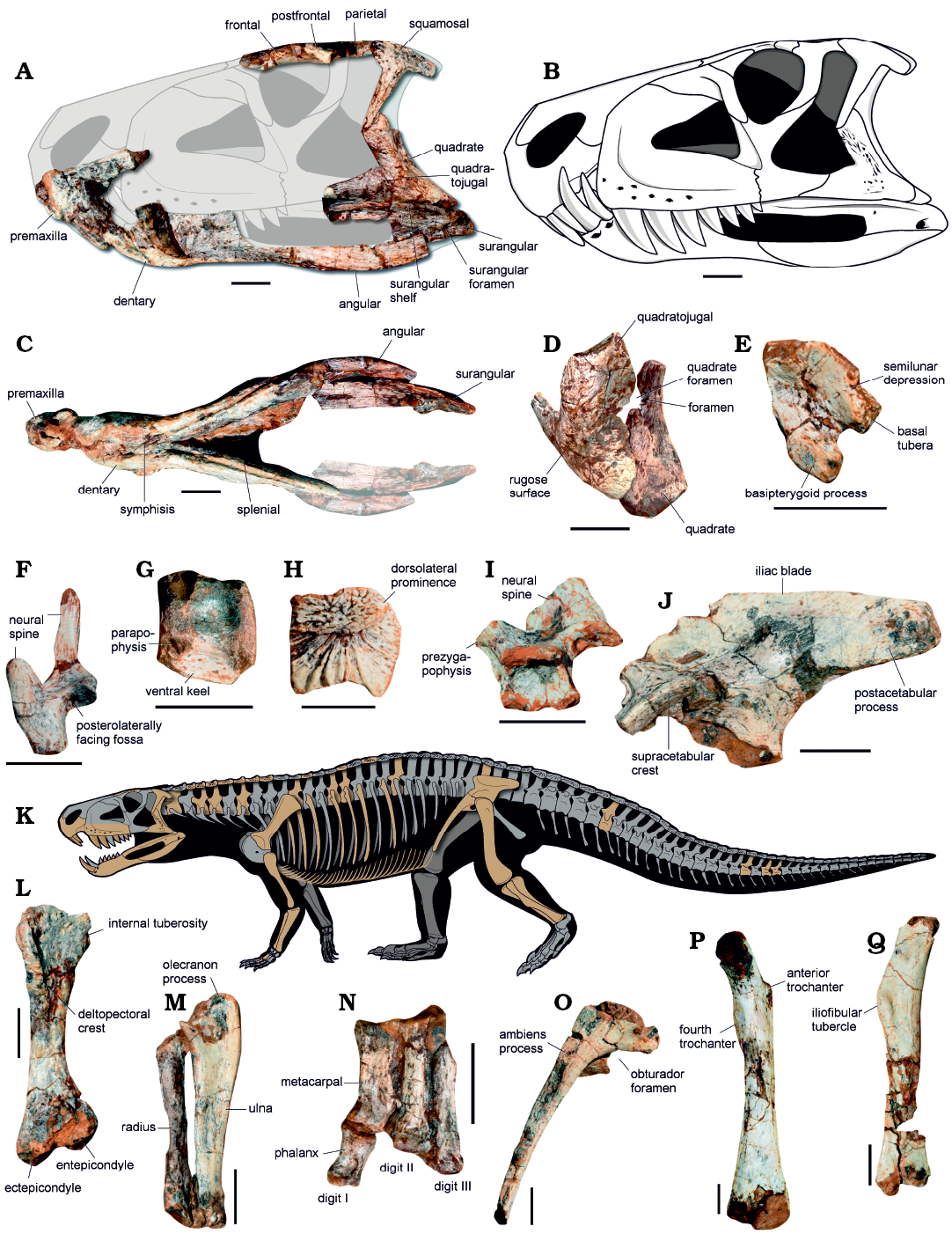
Fig. 2. Representative skeletal elements of ornithosuchid archosaur Dynamosuchus collisensis gen. et sp. nov. (CAPPA/UFSM 0248) from Janner outcrop, Carnian, Late Triassic. A. Selected skull bones in left lateral view. B. Reconstruction of the skull. C. Skull in ventral view. D. Left quadrate and quadratojugal in posterodorsal view. E. Parabasisphenoid in left lateral view. F. Neural arch of an anterior cervical vertebra in anterior view. G. Centrum of a cervical vertebra in left lateral view. H. Right osteoderm in dorsal view. I. Neural arch of an anterior dorsal vertebra in left lateral view. J. Left ilium in lateral view. L. Right humerus in anterior view. M. Right forearm in medial view. N. Left manus in dorsal view. O. Right (reversed) pubis in lateral view. P. Left femur in anterior view. Q. Left fibula in lateral view. Some unpreserved portions are modified from Baczko et al. in press, for the reconstruction of the skeleton of CAPPA/UFSM 0248 (preserved elements indicated in orange) (K). Scale bars 20 mm.
Most of the preacetabular process of the ilium is not preserved (Fig. 2J). Nevertheless, it is evident that the ilium of D. collisensis lacks a lateral crest dorsal to the supraacetabular crest, as in other ornithosuchids and many pseudosuchians (e.g., G. stipanicicorum PVL 4597; T. ruthae CRILAR-Pv 478; Ezcurra et al. 2017). As in O. woodwardi (NHMUK PV R 3561), the dorsal margin of the iliac blade of D. collisensis is straight to convex, whereas the homologous area in R. tenuisceps (PVL 3828) is concave. The supraacetabular crest merges into the main body of the bone approximately at the mid-length of the acetabulum in D. collisensis. The postacetabular process is proportionally shorter than in R. tenuisceps (PVL 3828) and lacks any evidence of a brevis fossa on the ventral surface. There are three articular surfaces for the sacral ribs on the medial surface of the ilium, as in other ornithosuchids and poposauroids, differing from the rest of pseudosuchians, which typically possess two sacral vertebrae. Although the distal end of the pubis is not preserved, the length of the pubis is estimated to be markedly elongated (~0.68 times the total length of the femur) compared to other pseudosuchians (Fig. 2O). The pubis is anteroventrally projected, but gently arched ventrally along its length. It has a large obturator foramen and a well-developed pubic apron present along the shaft. The distal end is not preserved and it is not possible to determine whether it has a pubic boot or any expansion.
The hindlimb is estimated to be about to 1.6 times longer than the forelimb. The femur (Fig. 2P) is sigmoidal with both ends expanded relative to the shaft. The proximal end preserves a well-developed posteromedial tuber, however, the anterolateral and anteromedial tubers as well as the greater trochanter have been eroded. There is a proximodistally oriented anterior trochanter at the proximal portion of the bone and a mound-like fourth trochanter more ventrally located at the opposite surface of the shaft. The presence of an anterior trochanter is a condition only registered in the ornithosuchids R. tenuisceps (PVL 3828) and O. woodwardi (NHMUK PV R 2410, NHMUK PV R 3561, NHMUK PV R 3916) within pseudosuchians (unknown in V. rusconii). The fourth trochanter of D. collisensis is less pronounced than in R. tenuisceps (PVL 3828), resembling the condition of O. woodwardi (NHMUK PV R 2410). The anterior surface of the distal end of D. collisensis is gently concave and the popliteal fossa, at the opposite surface, is proximodistally short. The distal condyles were eroded. The tibia is about 0.7 times of the length of the femur. The cnemial crest is poorly expanded from the shaft and has a straight anterior surface. The posterior edge of the proximal end is strongly projected. The fibula (Fig. 2Q) has an elliptical proximal end with an expanded posterior portion. There is a well-developed tubercle for the attachment of the iliofibularis muscle at the proximal half of the bone. In R. tenuisceps (PVL 3827), this tubercle is more distally located (Baczko and Ezcurra 2013). The specimen preserves slender gastralia (diameter 1.7 mm).
Geographic and strtigraphic range.—Type locality and horizon only.
Phylogenetic analysis
The phylogenetic analysis recovered 18 most parsimonious trees (MPTs) of 3356 steps each, with a consistency index of 0.261 and a retention index of 0.626. In all the MPTs, Dynamosuchus collisensis nests within Ornithosuchidae (Fig 3A). Ornithosuchus woodwardi lies as the basal-most member of Ornithosuchidae. The clade is supported by 19 synapomorphies (the complete list of synapomorphies is in Fig 3A), such as the alveolar margin of the premaxilla not reaching the contact with the maxilla [26 (0→1)], three premaxillary teeth [42 (2→3)], mandibular symphysis present along one-third of the lower jaw [265 (0→1)], three sacral vertebrae [370 (0→1)], perforated acetabulum [455 (0→1)], and presence of the anterior trochanter on the femur [502 (0→1)]. Dynamosuchus collisensis was found as the sister taxon of Venaticosuchus rusconii and this is supported by the semilunar depression on the posterolateral surface of the parabasisphenoid [238 (1)] and ascending process of the quadratojugal moderately anterodorsally to posteroventrally oriented in an angle higher than 40° [636 (0)]. The clade including both taxa is the sister group of Riojasuchus tenuisceps. This arrangement relies in the strongly downturned main body of the premaxilla [29 (2)] and the strongly anteroposteriorly expanded distal end of the radius [437 (1)]. Similar to the results of Ezcurra et al. (2017), Ornithosuchidae is found as the sister taxon of Erpetosuchidae, whereas this clade is found as the sister taxon of Aetosauria.
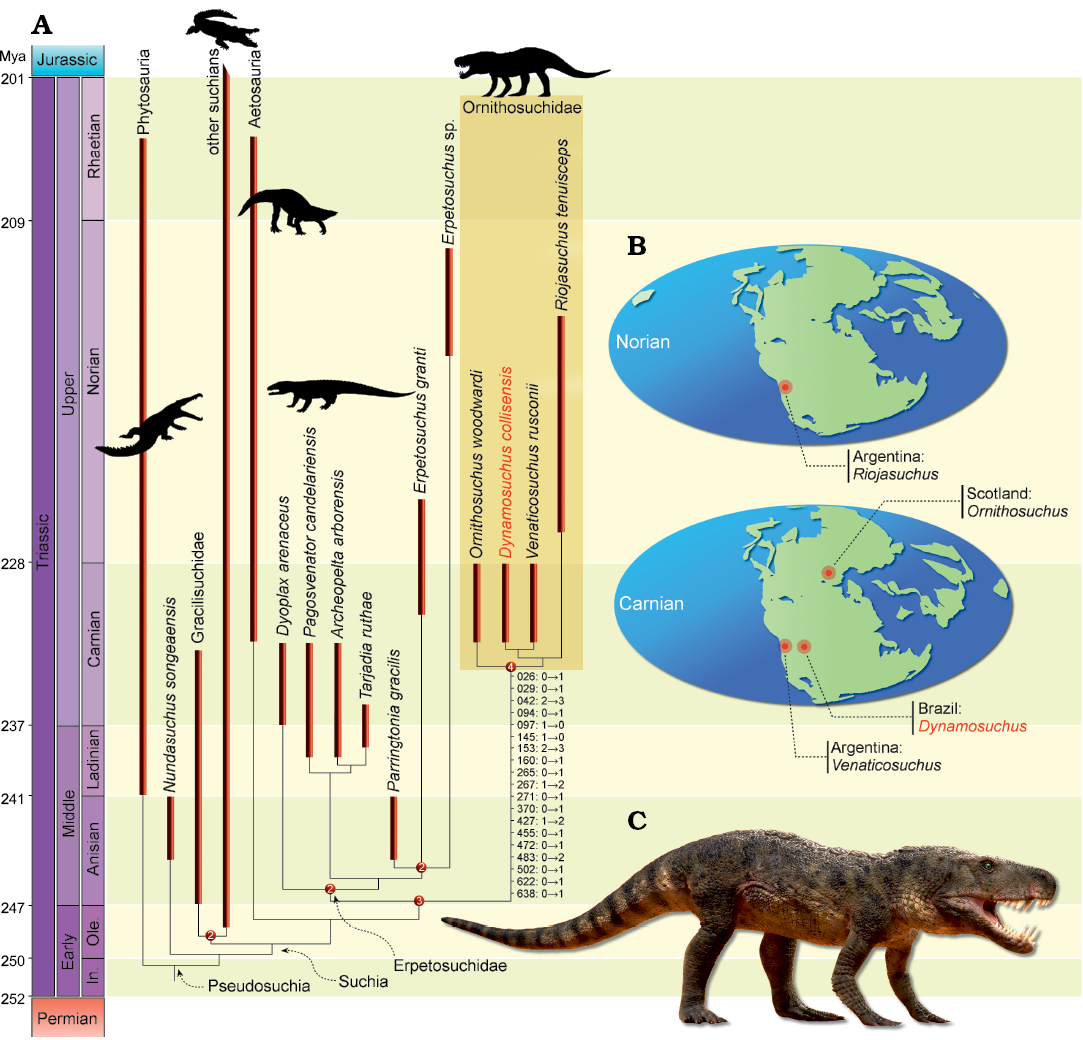
Fig. 3. Macroevolutionary patterns of Ornithosuchidae. A. Time-calibrated reduced strict consensus tree depicting the phylogenetic position of Dynamosuchus collisensis gen. et sp. nov. Numbers on nodes represent Bremer support values higher than one. Numbers associated with the ornithosuchid branch represent the characters and states that support the clade. B. Geographical distribution of ornithosuchids across the time. C. Life reconstruction of Dynamosuchus collisensis gen. et sp. nov. by Márcio L. Castro.
Discussion
Ornithosuchids, first described at the end of the 19th century (Newton 1894; see Baczko and Ezcurra 2016 for additional details], were important for deciphering the relationships among basal archosaurs because of their controversial anatomical features (Gauthier 1986; Sereno 1991; Nesbitt 2011; Ezcurra et al. 2017). However, no other ornithosuchids have been found since Venaticosuchus rusconii was discovered in Argentina almost a half century ago (Bonaparte 1970). Furthermore, Dynamosuchus collisensis is the first unambiguous Ornithosuchidae from Brazil.
The presence of ornithosuchids in Upper Triassic beds from Brazil is not a surprise, as the group is recorded in coeval strata of Argentina (i.e., V. rusconii). The age of the strata that yielded D. collisensis is similar to that of the levels in which Ornithosuchus woodwardi from Scotland and V. rusconii from Argentina were found, as these strata have also yielded remains of the rhynchosaur Hyperodapedon, which is extensively adopted as index fossil of Carnian age (Langer et al. 2007, 2018; Martínez et al. 2011; Desojo et al. 2018). The other known ornithosuchid, Riojasuchus tenuisceps, was exhumed from the Los Colorados Formation, Argentina, which is Norian in age (Kent et al. 2014). Therefore, D. collisensis falls within the stratigraphic range of the oldest known ornithosuchids (Fig. 3B).
The close relationship between Venaticosuchus rusconii and D. collisensis reinforces the previous proposed biostratigraphic hypotheses (Langer et al. 2007), indicating a similar faunal assemblage in both contemporaneous Paraná and Ischigualasto-Villa Unión basins. Moreover, the sister taxon affinity between V. rusconii and D. collisensis rejects a potential endemic radiation of ornithosuchids from the Ischigualasto-Villa Unión Basin and would better support multiple diversification events, at least at the Carnian. Indeed, according to Ezcurra et al. (2015, 2017), the phylogenetic affinities of proterochampsid archosauriforms and erpetosuchids would also suggest multiple dispersal events between the Ischigualasto-Villa Unión and the Paraná basins with the simultaneous appearance of several closely-related species in both basins (i.e., the Argentinean proterochampsids Proterochampsa barrionuevoi and Pseudochampsa ischigualastensis, and the Brazilian Rhadinosuchus gracilis and Proterochampsa nodosa; the Brazilian erpetosuchids Archeopelta arborensis and Pagosvenator candelariensis and the Argentinean Tarjadia ruthae). In addition, the presence of an ornithosuchid in Brazil expands the longitudinal distribution of the group, suggesting that ornithosuchids were more widespread than previously thought in the southern hemisphere. No records of the group have been found in lower latitudes close to the Paleo-Equator (Baczko and Ezcurra 2013), however, age equivalent formations or localities are very scarce (Irmis et al. 2011). Therefore, ornithosuchids are still restricted to higher latitudes of northern and southern Pangea.
The discovery of D. collisensis (Fig. 3C) in the Paraná Basin also provides novel paleoecological implications for this type locality (i.e., Janner site). Strictly carnivorous reptiles were previously absent from this site, as the early sauropodomorph Pampadromaeus barberenai (Cabreira et al. 2011) was more likely to be an omnivorous animal according to a recent ecomorphological analysis (Müller and Garcia 2019). On the other hand, Bagualosaurus agudoensis (Pretto et al. 2018) is more related to post-Carnian sauropodomorphs, which are considered herbivorous or omnivorous. Therefore, strictly carnivorous animals are solely represented in the site through the record of ecteniniid cynodonts (Oliveira et al. 2010; Stefanello et al. 2018). Indeed, ornithosuchids have been considered as top tier predators (Walker 1964). However, recently the feeding behavior of these animals have been explored through more rigorous techniques (i.e., quantitative approaches), indicating that ornithosuchids likely adopted scavenging feeding habits or preyed only on small vertebrates (Baczko 2018). Such statement relies on the elevated bite force together with the low bite speed and the morphology of their constricted snouts. Therefore, the discovery of D. collisensis provides the first clue of a putative necrophagous vertebrate from the Janner site and expands our knowledge regarding the trophic chain of Late Triassic of Brazil. This is particularly interesting because necrophagous animals are an important part of the extant terrestrial ecosystems, interacting with the carcasses before its incorporation within the lithosphere. Therefore, identifying the animals that played such role in ancient environments is crucial for the reconstruction of reliable pictures of these. Indeed, the Janner site has yielded evidences of osteophagic behavior by insects (Paes et al. 2016) and the putative action of necrophagous animals that produced an accumulation with predominance of cranial elements (Müller et al. 2015).
Conclusions
Dynamosuchus collisensis is the first unambiguous member of Ornithosuchidae from Brazil, and was recovered as sister taxa of the Argentinean taxon Venaticosuchus rusconii. This sister taxon relationship would support multiple radiation events between Paraná and Ischigualasto-Villa Unión basins in agreement with previous hypothesis based on other archosauriforms that are shared across these basins. The discovery of an ornithosuchid in Carnian continental outcrops of Brazil provides a new shared faunal component between the basins. The Carnian archosauromorph communities of Argentina and Brazil would therefore consist of rhynchosaurs, proterochampsids, ornithosuchids, aetosaurs, rauisuchians, erpetosuchids, sauropodomorphs, and theropods. In addition, the discovery Dynamosuchus collisensis provides the first clue of a putative necrophagous vertebrate from the oldest dinosaur-bearing beds and expands our knowledge regarding the trophic structure of the Late Triassic of Brazil.
Acknowledgements
We thank the colleagues of CAPPA/UFSM for their help during fieldwork. We also extend our gratitude to the editor Stephen Brusatte and Davide Foffa (bothe School of GeoSciences, University of Edinburgh, Edinburgh, UK) for comments and suggestions that improved the quality of this manuscript. We thank the Willi Henning Society, for the gratuity of TNT software. This work was partially supported by the Agencia Nacional de Promoción Científica y Tecnológica (PICT 2016-0159 to PB and JBD).
References
Alcober, O. 2000. Redescription of the skull of Saurosuchus galilei (Archosauria: Rauisuchidae). Journal of Vertebrate Paleontology 20: 302–316. Crossref
Baczko, M.B. von 2018. Rediscovered cranial material of Venaticosuchus rusconii enables the first jaw biomechanics in Ornithosuchidae (Archosauria: Pseudosuchia). Ameghiniana 55: 365–380. Crossref
Baczko, M.B. von and Desojo, J.B. 2016. Cranial anatomy and palaeoneurology of the archosaur Riojasuchus tenuisceps from the Los Colorados Formation, La Rioja, Argentina. PloS One 11: e0148575. Crossref
Baczko, M.B. von and Ezcurra, M.D. 2013. Ornithosuchidae: a group of Triassic archosaurs with a unique ankle joint. Geological Society, London, Special Publications 379: 187–202. Crossref
Baczko, M.B. von and Ezcurra, M.D. 2016. Taxonomy of the archosaur Ornithosuchus: reassessing Ornithosuchus woodwardi Newton, 1894 and Dasygnathoides longidens (Huxley 1877). Earth and Environmental Science Transactions of The Royal Society of Edinburgh 106: 199–205. Crossref
Baczko, M.B. von, Desojo, J.B., and Pol, D. 2014. Anatomy and phylogenetic position of Venaticosuchus rusconii Bonaparte, 1970 (Archosauria, Pseudosuchia), from the Ischigualasto Formation (Late Triassic), La Rioja, Argentina. Journal of Vertebrate Paleontology 34: 1342–1356. Crossref
Baczko, M.B. von, Desojo, J.B., and Ponce, D. 2020. Postcranial anatomy and osteoderm histology of Riojasuchus tenuisceps and a phylogenetic update on Ornithosuchidae (Archosauria, Pseudosuchia). Journal of Vertebrate Paleontology 39 (5): e1693396. Crossref
Benton, M.J. and Walker, A.D. 2002. Erpetosuchus, a crocodile-like basal archosaur from the Late Triassic of Elgin, Scotland. Zoological Journal of the Linnean Society 136: 25–47. Crossref
Bonaparte, J.F. 1967. Dos nuevas “faunas” de reptiles triásicos de Argentina. Gondwana Symposium Proceedings and Papers 1: 283–306.
Bonaparte, J.F. 1970. Annotated list of the South American Triassic tetrapods. Gondwana Symposium Proceedings and Papers 2: 665–682.
Butler, R.J., Sullivan, C., Ezcurra, M.D., Liu, J., Lecuona, A., and Sookias, R.B. 2014. New clade of enigmatic early archosaurs yields insights into early pseudosuchian phylogeny and the biogeography of the archosaur radiation. BMC Evolutionary Biology 14: 128. Crossref
Cabreira, S.F., Schultz, C.L., Bittencourt, J.S., Soares, M.B., Fortier, D.C., Silva, L.R., and Langer, M.C. 2011. New stem-sauropodomorph (Dinosauria, Saurischia) from the Triassic of Brazil. Naturwissenschaften 98: 1035–1040. Crossref
Cope, E.D. 1869. Synopsis of the extinct Batrachia and Reptilia of North America. Transactions of the American Philosophical Society 14: 1–252. Crossref
Da-Rosa, A.A.S. 2015. Geological context of the dinosauriform-bearing outcrops from the Triassic of Southern Brazil. Journal of South American Earth Sciences 61: Pages 108–119. Crossref
Desojo, J.B. and Báez, A.M. 2007. Cranial morphology of the Late Triassic South American archosaur Neoaetosauroides engaeus: evidence for aetosaurian diversity. Palaeontology 50: 267–276. Crossref
Desojo, J.B., Heckert, A.B., Martz, J.W., Parker, W.G., Schoch, R.R., Small, B.J., and Sulej, T. 2013. Aetosauria: a clade of armoured pseudosuchians from the Upper Triassic continental beds. Geological Society, London, Special Publications 379: 203–239. Crossref
Desojo, J.B., Langer, M.C., Da-Rosa, A.A.S., Ezpeleta, M., Ezcurra, M.D., Martinelli, A.G., Baczko, M.B. von, Fiorelli, L., Taborda, J.R.A., Trotteyn, M., Hechenleitner, M., Montefeltro, F., and Ramezani, J. 2018. Nuevos hallazgos y dataciones en la Formación Ischigualasto en Cerro Las Lajas (Cuenca Ischigualasto-Villa Unión), Provincia de La Rioja, Argentina. Publicación Electrónica de la Asociación Paleontológica Argentina 19: 46.
Ezcurra, M.D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ 4: e1778. Crossref
Ezcurra, M.D., Desojo, J.B., and Rauhut, O.W.M. 2015. Redescription and phylogenetic relationships of the proterochampsid Rhadinosuchus gracilis (Diapsida: Archosauriformes) from the early Late Triassic of southern Brazil. Ameghiniana 52: 391–417. Crossref
Ezcurra, M.D., Fiorelli, L.E., Martinelli, A.G., Rocher, S.M., Baczko, M.B. von, Ezpeleta, M., Taborda, J.R.A., Hechenleitner, E.M., Trotteyn, M.J., and Desojo, J.B. 2017. Deep faunistic changes preceded the raise of dinosaurs 1 in southwestern Pangaea. Nature Ecology & Evolution 1: 1477–1483. Crossref
Gauthier, J. and Padian, K. 1985. Phylogenetic, functional, and aerodynamic analyses of the origin of birds and their flight. In: M.K. Hecht, J.H. Ostrom, G. Viohl, P. Wellnhofer (eds.), The Beginning of Birds. Proceedings of the International Archaeopteryx Conference, Eichstatt, 1984, 185–97. Freunde des Jura-Museums, Eichstatt.
Gauthier, J.A. 1986. Saurischian monophyly and the origin of birds. Memoirs of the California Academy of Science 8: 1–55.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT: a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Huene, F. von 1908. Die Dinosaurier der europäischen Triasformation mit Berücksichtigung der aussereuropäischen Vorkommnisse. Geologische und Paläontologische Abhandlungen 1 (Supplement): 1–419.
Irmis, R.B., Mundil, R., Martz, J.W., and Parker, W.G. 2011. High-resolution U-Pb ages from the Upper Triassic Chinle Formation (New Mexico, USA) support a diachronous rise of dinosaurs. Earth and Planetary Science Letters 309: 258–267. Crossref
Kent, D.V., Santi Malnis, P., Colombi, C.E., Alcober, O.A., and Martínez, R.N. 2014. Age constraints on the dispersal of dinosaurs in the Late Triassic from magnetochronology of the Los Colorados Formation (Argentina). Proceedings of the National Academy of Sciences of the United States of America 111: 7958–7963. Crossref
Lacerda, M.B., de França, M.A.G., and Schultz, C.L. 2018. A new erpetosuchid (Pseudosuchia, Archosauria) from the middle–late Triassic of southern Brazil. Zoological Journal of the Linnean Society 184: 804–824. Crossref
Langer, M.C., Ramezani, J., and Da-Rosa, Á.A.S. 2018. U-Pb age constraints on dinosaur rise from south Brazil. Gondwana Research 57: 133–140. Crossref
Langer, M.C., Ribeiro, A.M., Schultz, C.L., and Ferigolo, J. 2007. The continental tetrapodbearing Triassic of south Brazil. New Mexico Museum of Natural History & Science Bulletin 41: 201–218.
Lecuona, A., Desojo, J.B., and Pol, D. 2017. New information on the postcranial skeleton of Gracilisuchus stipanicicorum (Archosauria: Suchia) and reappraisal of its phylogenetic position. Zoological Journal of the Linnean Society 181: 638–677. Crossref
Martinez, R.N., Sereno, P.C., Alcober, O.A., Colombi, C.E., Renne, P.R., Montañez, I.P., and Currie, B.S. 2011. A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science 331: 206–210. Crossref
Mastrantonio, B., Baczko, M.B. von, Desojo, J.B., and Schultz, C.L. 2019. The skull anatomy and cranial endocast of the pseudosuchid archosaur Prestosuchus chiniquensis from the Triassic of Brazil. Acta Palaeontologia Polonica 64: 171–198. Crossref
Müller, R.T. and Garcia, M.S. 2019. Rise of an empire: analysing the high diversity of the earliest sauropodomorph dinosaurs through distinct hypotheses. Historical Biology [published online, https://doi.org/10.1080/08912963.2019.1587754] Crossref
Müller, R.T., Araújo-Júnior, H.I., Aires, A.S.S., Silva Roberto-da-Silva, L., and Dias-da-Silva, S. 2015. Biogenic control on the origin of a vertebrate monotypic accumulation from the Late Triassic of southern Brazil. Geobios 48: 331–340. Crossref
Müller, R.T., Langer, M.C., and Dias-da-Silva, S. 2017. Biostratigraphic significance of a new early sauropodomorph specimen from the Upper Triassic of southern Brazil. Historical Biology 29: 187–202. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Nesbitt, S.J. and Butler, R.J. 2012. Redescription of the archosaur Parringtonia gracilis from the Middle Triassic Manda beds of Tanzania, and the antiquity of Erpetosuchidae. Geological Magazine 150: 225–238. Crossref
Newton, E.T. 1894. Reptiles from the Elgin Sandstone—Description of two new genera. Philosophical Transactions of the Royal Society of London B, Biological Sciences 185: 573–607. Crossref
Oliveira, T.V., Soares, M.B., and Schultz, C.L. 2010. Trucidocynodon riograndensis gen. nov. et sp. nov. (Eucynodontia), a new cynodont from the Brazilian Upper Triassic (Santa Maria Formation). Zootaxa 2382: 1–71. Crossref
Paes, V.D., Parkinson, A.H., Pretto, F.A., Soares, M.B., Schwanke, C., Schultz, C.L., and Kellner, A.W. 2019. Oldest evidence of osteophagic behavior by insects from the Triassic of Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology 453: 30–41. Crossref
Parker, W.G. 2016. Revised phylogenetic analysis of the Aetosauria (Archosauria: Pseudosuchia); assessing the effects of incongruent morphological character sets. PeerJ 4: e1583. Crossref
Pretto, F.A., Langer, M.C., and Schultz, C.L. 2018. A new dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Brazil provides insights on the evolution of sauropodomorph body plan. Zoological Journal of the Linnean Society 185: 388–416. Crossref
Sereno, P.C. 1991. Basal archosaurs: phylogenetic relationships and functional implications. Journal of Vertebrate Paleontology 11: 1–53. Crossref
Stefanello, M., Müller, R.T., Kerber, L., Martínez, R.N., and Dias-Da-Silva, S. 2018. Skull anatomy and phylogenetic assessment of a large specimen of Ecteniniidae (Eucynodontia: Probainognathia) from the Upper Triassic of southern Brazil. Zootaxa 4457: 351–378. Crossref
Walker, A.D. 1964. Triassic reptiles from the Elgin area Ornithosuchus and the origin of Carnosaurs. Philosophical Transactions of the Royal Society of London B, Biological Sciences 744: 53–134. Crossref
Zittel, K.A. von 1887–1890. Handbuch der Paläontologie. Abteilung 1: Paläozoologie Band III. Vertebrata (Pisces, Amphibia, Reptilia, Aves). 1890 pp. Oldenbourg, Munich.
Acta Palaeontol. Pol. 65 (1): 1–10, 2020
https://doi.org/10.4202/app.00652.2019