
The oldest representative of the bombyliid bee fly in the earliest Eocene French amber
AUDE MAHEU and ANDRÉ NEL
Eoconophorina delfineae gen. et sp. nov., the oldest representative of the bee fly subfamily Bombyliinae, is described from the earliest Eocene Oise amber (France). It is characterized by the presence of mid tibial spurs, cylindrical abdomen and a particular combination of wing venation characters. It is putatively attributed to the tribe Conophorini. This fossil is 53 Ma old, ca. 19 Ma older than the previous oldest records of the subfamily, known as compression fossils. There are very few older bee flies, one from the Paleocene of Menat (France), and six from the mid-Cretaceous Burmese amber, belonging to different subfamilies or of uncertain taxonomic position. Thus this fossil will be helpful for future calibration of phylogeny and dating of the bee flies.
Introduction
The Bombyliidae are a rather ancient clade of flies, known already from the mid-Cretaceous Burmese amber (Grimaldi 2016; Ye et al. 2019). The oldest known representatives of the subfamily Bombyliinae are from the late Eocene and the Oligocene (Wedmann and Yeates 2008). The earliest Eocene Oise amber has yielded several Bombyliidae (from subfamilies of Phthiriinae and Toxophorinae) and Mythicomyiidae (Nel and De Ploëg 2004; Nel 2006). Here we describe a new fossil bee fly from the same amber, belonging to a new genus and species which constitutes the oldest representative of the Bombyliinae.
Institutional abbreviations.—MNHN, Muséum National d’Histoire Naturelle, Paris, France.
Other abbreviations.—A, anal vein; br, basal radial cell; bm, basal medial cell; C, costal vein; Cu, cubital vein; cua, cubital cell; dm, discal cell; M, median vein; R, radial vein; r4+5, last radial cell; r-rm, radial-median crossvein; Sc, subcostal vein.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in urn:lsid:zoobank.org:pub:14724E8D-C04D-4A84-A642-935CCB0D6981
Material and methods
The type of Eoconophorina delfineae gen. et sp. nov. is embedded in a very small clear piece of amber, and was unfortunately broken into three fragments during preparation. It has been prepared using a diamond disk and examined using a Nikon binocular microscope SMZ 1500. Photographs have been taken with an Amscope camera MU900, and the images treated with Adobe Photoshop Element 12. For information on the age, outcrop, and fossils from the Oise amber, see Nel and Brasero (2010).
The list of extant Bombyliidae follows Evenhuis and Greathead (1999), the classification follows Li and Yeates (2019) and the morphological nomenclature follows Cumming and Wood (2017).
Systematic palaeontology
Order Diptera Linnaeus, 1758
Family Bombyliidae Latreille, 1802
Subfamily Bombyliinae Latreille, 1802
Tribe ?Conophorini Becker, 1913
Genus Eoconophorina nov.
ZooBank LSID: urn:lsid:zoobank.org:act:3D18D5B8-9BF4-4FAF-BDAE-A8EF1B42E498
Type species: Eoconophorina delfineae sp. nov. by monotypy, see below.
Etymology: Named after the Eocene period and the extant genus Conophorina.
Diagnosis.—As for the type species by monotypy.
Eoconophorina delfineae sp. nov.
Fig. 1.
Zoobank LSID: urn:lsid:zoobank.org:act:F8811EF3-6C33-4A77-8786- EB8AE5870605
Etymology: Named after Delphine Maheu, mother of first author.
Holotype: MNHN.F.A71306 (sex unknown, thorax, abdomen, legs and wings preserved but head missing).
Type locality: Farm Le Quesnoy, Chevrière, near Creil, Oise department, France.
Type horizon: Lowermost Eocene, in Oise amber, c. 53 Ma.
Diagnosis.—Mid tibial spurs present; R2+3 ending on C rather obliquely, not at right angle; R4 at right angle with R5 and strongly sigmoidal; cell r4+5 widely open with R5 and M1 slightly diverging; vein r-m at middle of cell dm; apex of R2+3 near apex of R1; cell cua open; abdomen cylindrical.
Description.—Medium-sized bombyliine (preserved part of body length ca. 5.0 mm). sex unknown; head not preserved; scutum and scutellum clothed in hairs and scales; two prealar bristles; haired anepimeron and laterotergite; no peg on hind coxa; femora slender; empodium setiform half as long as claws; claw shorter than distitarsus; wing 4.0 mm long, 1.4 mm wide; hyaline; base of R2+3 very oblique and close to base of Rs, opposite basal angle of cell dm; basal angle of dm very acute; dm narrow; R2+3 ending on C rather obliquely, not at right angle, apex of R2+3 very close to apex of R1; R4 at right angle with R5 and strongly sigmoidal; cell r4+5 open with R5 and M1 diverging; crossvein r-m situated slightly distal middle of cell dm; crossvein m-m slightly longer than crossvein r-m; cell r5 widely open; cell cua open with CuA2+A1 and A2 approximating near posterior wing margin; cell br broader and shorter than cell bm; alula rather well developed; abdomen 3.5 mm long, 0.9 mm wide, cylindrical, tergites 3–7 broader than long, covered with dense black scales; terminalia not preserved.
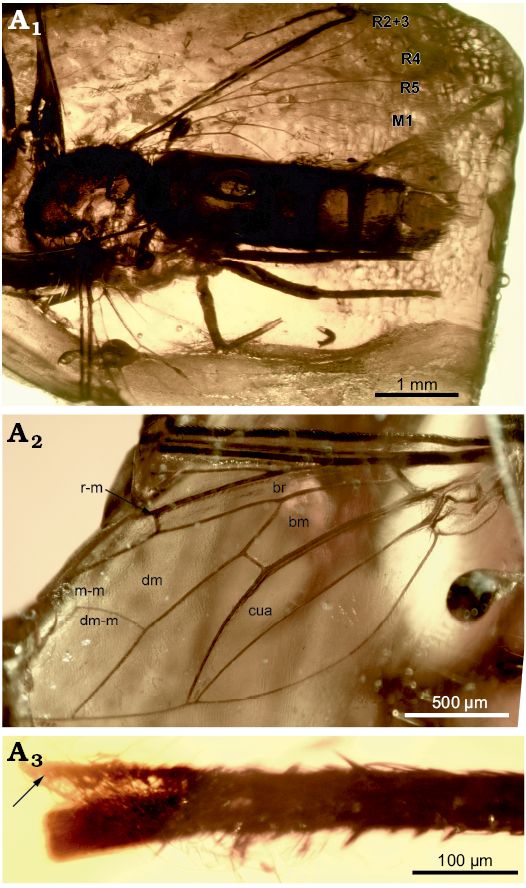
Fig. 1. Bombyliid bee fly Eoconophorina delfineae gen. et sp. nov., holotype MNHN.F.A71306; earliest Eocene, Oise amber, France. Habitus above (A1), left wing (A2), mid tibia (A3, arrow indicates spur). Abbreviations: bm, basal medial cell; br, basal radial cell; cua, cubital cell; dm discal cell; dm-m discal-medial crossvein; m-m medial crossvein; r-m radial-median crossvein; M1, first branch of median vein; R2+3, second + third branch of radial vein; R4, fourth branch; R5, fifth branch.
Remarks.—Following the key to fly families of Marshall et al. (2017), this fossil falls in the Bombyliidae because of the following characters: wing fully developed; body not strikingly modified; wing membrane without a pattern of folds between veins; cell cua very long; vein R4+5 forked; empodium bristle-like; no spurious vein; veins not conspicuously curved anteriorly before wing apex; basal median cell with veins arising from three corners; arculus present. Yeates (1994) proposed a series of synapomorphies for the Bombyliidae; unfortunately, all concern characters that are not visible in our fossil. The absence of the head structures renders its attribution difficult. The absence of peg on the hind coxa excludes affinities with the Oligodraninae and Crocidiinae. But, after Yeates (1994) the presence of prealar bristles is a synapomorphy of the Bombyliinae. The absence of the mid tibial spurs is a character of the Bombyliinae, except the majority of the Conophorini, which could suggest an attribution to this tribe, but it is certainly a symplesiomorphy not sufficient for such an attribution. Nevertheless, its wing venation is very close to those of some Bombyliinae. The characters “three branches of Rs; R2+3 arises at an acute angle and in a proximal position; M2 present; cell r5 open; dm-m present; costal vein circumambient” are also shared by Eoconophorina gen. nov. and the Bombyliinae. The open cell cua is shared by many Bombyliinae but also by representatives of some other subfamilies, e.g., some Lomatiinae. Some extant genera with wing venations similar to that of Eoconophorina gen. nov. belong to the Acrophthalmydini (Acrophthalmyda Bigot, 1858), the Bombyliini (Bombylisoma Rondani, 1856, Laurella Hull, 1971), the Marmasomini (Marmasoma White, 1917, Paratoxophora Engel, 1936), and the Conophorini (Conophorina Becker, 1920) (Hull 1973). The haired anepimeron and laterotergite is a character shared by Eoconophorina gen. nov. and the Bombyliini sensu Yeates (1994), while they are bare in Paratoxophora. Among these genera, only Conophorina shares with Eoconophorina gen. nov. the following venation characters: base of R2+3 very oblique; basal angle of dm very acute; R2+3 ending on C rather obliquely, not at right angle; cell r4+5 open with R5 and M1 diverging; cell cua open with CuA2+A1 and A2 approximate near posterior wing margin (Hull 1973). The unique species of Conophorina, C. bicellaris Becker, 1920, differs from Eoconophorina gen. nov. in the bare laterotergites (Becker 1920; Greathead and Evenhuis 2001), the apex of R2+3 well distal of apex of R1, instead of being very close to it, as in Eoconophorina gen. nov., the abdomen of Eoconophorina gen. nov. is cylindrical elongate, while it is as broad as thorax at base and distally tapered in Conophorina (Hull, 1973). Eoconophorina gen. nov. differs from all the other genera of the tribe Conophorini sensu Li and Yeates (2019) in the wing venation.
After the key of Li and Yeates (2019), Eoconophorina gen. nov. would fall in the Conophorini because of the following characters: vein dm-m slightly curved; mid tibial spurs present (absent in Conophorina). Eoconophorina gen. nov. has also the wing venation characters of the Conophorini as defined by Li and Yeates (2019), viz. cell br longer than cell bm; crossvein r-m located in basal part or at middle of cell dm; vein R5 nearly parallel to M1, cell r5 wide open.
Stratigraphic and geographic range.—Type locality and horizon only.
Concluding remarks
Eoconophorina gen. nov. is the oldest representative of the subfamily Bombyliinae. Its tribal attribution is relatively problematic, even if it is possibly related to the Conophorini. It demonstrates the presence of Bombyliinae during the earliest Eocene, in congruence with the unique proposal of dating of the bombyliid subfamilies of Lamas and Nihei (2007) who proposed a range of ages between 125 Ma and 70 Ma for this subfamily. Eoconophorina gen. nov. should help to precise an age for this clade in future phylogenetic analyses.
Acknowledgements.—We greatly thank Sonja Wedmann (Forschungsinstitut Senckenberg, Weimar, Germany) and an anonymous referee for their useful comments on the first version of this paper. We thank the Lafarge-Granulat company for help with the fossil sampling and the Langlois-Meurinne family for the authorization of working on its property.
References
Becker, T. 1920. Conophorina, novum genus Bombyliidarum. Entomologische Mitteilungen 9: 181–184. Crossref
Cumming, J.M. and Wood, D.M. 2017. Adult morphology and terminology. In: A.H. Kirk-Spriggs and B.J. Sinclair (eds.), Manual of Afrotropical Diptera, Volume 1. Introductory chapters and keys to Diptera families. Suricata 4: 89–134.
Evenhuis, N.L. and Greathead, D.J. 1999. World Catalog of Bee Flies (Insecta: Diptera). xlviii + 756 pp. Backhuys Publishers, Leiden.
Greathead, D.J. and Evenhuis, N.L. 2001. Annotated keys to the genera of African Bombylioidea (Diptera: Bombyliidae; Mythicomyiidae). African Invertebrates 42: 105–224.
Grimaldi, D. 2016. Diverse orthorrhaphan flies (Insecta: Diptera: Brachycera) in amber from the Cretaceous of Myanmar: Brachycera in Cretaceous amber, Part VII. Bulletin of the American Museum of Natural History 408: 1–131. Crossref
Hull, F.M. 1973. Bee flies of the world. The genera of the family Bombyliidae. Bulletin of the United States National Museum 286: 1–687. Crossref
Lamas, C.J.E. and Nihei, S.S. 2007. Biogeographic analysis of Crocidiinae (Diptera, Bombyliidae): finding congruence among morphological, molecular, fossil and paleogeographical data. Revista Brasileira de Entomologia 51: 267–274. Crossref
Li, X. and Yeates, D.K. 2019. Phylogeny, classification and biogeography of bombyliine bee flies (Diptera, Bombyliidae). Systematic Entomology [published online, https://doi.org/10.1111/syen.12361] Crossref
Marshall, S.A., Kirk-Spruggs, A.H., Muller, B.S., Paiero, S.M., Yau, T., and Jackson, M.D. 2017. Chapter 12: Key to Diptera families—adults. In: A.H. Kirk-Spriggs and B.J. Sinclair (eds.), Manual of Afrotropical Diptera, Volume 1. Introductory chapters and keys to Diptera families. Suricata 4: 267–355.
Nel, A. 2006. Oldest records of Bombyliidae: Phthiriinae and Mythicomyiidae: Glabellulinae from the Lowermost Eocene amber of France (Diptera: Bombylioidea). European Journal of Entomology 103: 109–114. Crossref
Nel, A. and Brasero, N. 2010. Oise amber. In: D. Penney (ed.), Biodiversity of Fossils in Amber from the Major World Deposits, 137–148. Siri Scientific Press, Manchester.
Nel, A. and De Ploëg, G. 2004. New fossil bee flies in the Lowermost Eocene amber of the Paris basin (Diptera: Bombylioidea). Geologica Acta 2: 57–65.
Wedmann, S. and Yeates, D. 2008. Eocene records of bee flies (Insecta, Diptera, Bombyliidae, Comptosia): their paleobiogeographic implications and remarks on the evolutionary history of bombyliids. Palaeontology 51: 231–240. Crossref
Ye, X.-N., Yao, G., Shih, C.-K., Ren, D., and Wang, Y.-J. 2019. New bee flies from the mid-Cretaceous Myanmar amber (Brachycera: Asiloidea: Bombyliidae). Cretaceous Research 100: 5–13. Crossref
Yeates, D.K. 1994. The cladistics and classification of the Bombyliidae (Diptera: Asiloidea). Bulletin of the American Museum of Natural History 219: 1–191.
Aude Maheu [aude.maheu23@gmail.com], 5 rue Marie Davy, F-75014, Paris, France.
André Nel [anel@mnhn.fr] (corresponding author), Institut Systématique Evolution Biodiversité (ISYEB), Muséum national d’Histoire naturelle, CNRS, Sorbonne Université, Université des Antilles, EPHE, 57 rue Cuvier, CP 50, F-75005 Paris, France.
Received 23 July 2019, accepted 20 September 2019, available online 31 January 2020.
Copyright © 2019 A. Maheu and A. Nel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 65 (1): 81–83, 2020
https://doi.org/10.4202/app.00657.2019