
Morphology and taxonomic position of the bizarre Permian pachydomid bivalve Leinzia from Western Gondwana
MARCELLO G. SIMÕES, VITOR B. GUERRINI, SUZANA A. MATOS, and ROSEMARIE ROHN
Simões, M.G., Guerrini, V.B., Matos, S.A., and Rohn, R. 2020. Morphology and taxonomic position of the bizarre Permian pachydomid bivalve Leinzia from Western Gondwana. Acta Palaeontologica Polonica 65 (2): 291–303.
The genus Leinzia is a typical member of the renowned Artinskian–Wuchiapingian (Permian) endemic bivalve fauna of the Passa Dois Group, Paraná Basin, Brazil. The extraordinary shells of Leinzia, characterized by a rostrum extending from the anterior cardinal margin led certain authors to regard them as bivalved arthropods (Spinicaudata). Due to the unusual morphology and typically poor preservation of the available specimens, the taxonomic position of Leinzia still remains obscure. Leinzia has been variously referred either to the Pterioida, the Crassatelloidea, the Sanguinolitidae, or the Megadesmidae, or to the Pholadomyida. Herein, based on a detailed review of the topotype material and description of newly found specimens of Leinzia from the Serrinha Member, Rio do Rasto Formation, southern Brazil, we shed light on the taxonomic position of this genus. The hinge of the right valve with its large, blunt, anteriorly inclined subumbonal tooth and corresponding socket in the left valve coupled with the absence of true lateral teeth indicate close affinities to Pyramus and Cowperesia. Thus, the data here strongly suggest a Pachydomidae (Edmondioidea) rather than a Crassatelloidea affinity for Leinzia. Conversely, Leinzia differs from all other known Pachydomidae due to its anteriorly rostrate and posteriorly elongated shell. Finally, detailed stratigraphic data indicate that the vertical distribution of Leinzia is constrained to the middle part of the Guadalupian Serrinha Member of the Rio do Rasto Formation.
Key words: Bivalvia, Pachydomidae, Megadesmidae, endemism, Guadalupian, Paraná Basin, Brazil.
Marcello G. Simões [profmgsimoes@gmail.com] and Suzana A. Matos [sumatos.s@gmail.com], Institute of Biosciences, São Paulo State University, Botucatu campus, Botucatu, P.O. Box 510, 18618-689, Brazil.
Vitor B. Guerrini [vbguerrini@gmail.com] and Rosemarie Rohn [rosemarie.rohn@unesp.br], Institute of Geosciences and Exact Sciences, São Paulo State University, Rio Claro campus, Rio Claro, 13506-900, Brazil.
Received 14 August 2019, accepted 10 January 2020, available online 22 April 2020.
Copyright © 2020 M.G. Simões et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
During most of the Permian (c. 278–252 Ma), vast areas of the South American continent were flooded by an epeiric sea with restricted connections to the Panthalassic Ocean. Nearly 1 100 000 km2 of the Brazilian territory area is represented by the intracratonic Paraná Basin. From the Cisuralian (Artinskian) to Lopingian (Wuchiapingian) stages this basin was filled with a thick (~ 1400 m) succession of mixed carbonate-siliciclastic rocks (i.e., Passa Dois Group) (Zalán et al. 1990, 1991; Milani et al. 2007; Holz et al. 2010). This unit is divided from the base to the top into the Irati, Serra Alta, Teresina/Corumbataí and Rio do Rasto formations (Milani et al. 2007; Rohn 1994; Holz et al. 2010; Fig. 1). Throughout the deposition of the Passa Dois Group the basin experienced variable salinity (hypersaline, brackish, and hyposaline) and dissolved oxygen (dysoxic, anoxic) regimes, mixed siliciclastic and carbonate sedimentation, under climatic changes (humid to arid conditions) (Milani et al. 2007; Rohn 1994; Holz et al. 2010). Given these environmental conditions, the shallow benthic environments of the Paraná epeiric sea were probably stressful for most of the fully marine macroinvertebrates and were mainly colonized by endemic bivalves. These mollusks radiated in the basin especially after the Artinskian Irati anoxic event (Mendes 1952; Beurlen 1954; Runnegar and Newell 1971; Simões et al. 1997, 1998; Wesselingh 2007; Matos et al. 2017b). Distinct bivalve assemblages are known in all units of the Passa Dois Group reaching their maximum abundance and diversity in the Teresina Formation and in the basal part of the Rio do Rasto Formation (Fig. 1). A unique feature of this molluscan-bearing long-lived fauna is the dominance of bivalves in all assemblages (Wesselingh 2007), whereas gastropods are rare or even absent in almost all fossil assemblages. The unique exception are some benthic fossil assemblages in the Rio do Rasto Formation (see Rohn 1994) where some hydrobiid gastropods (see Solem and Yochelson 1979) are recorded.
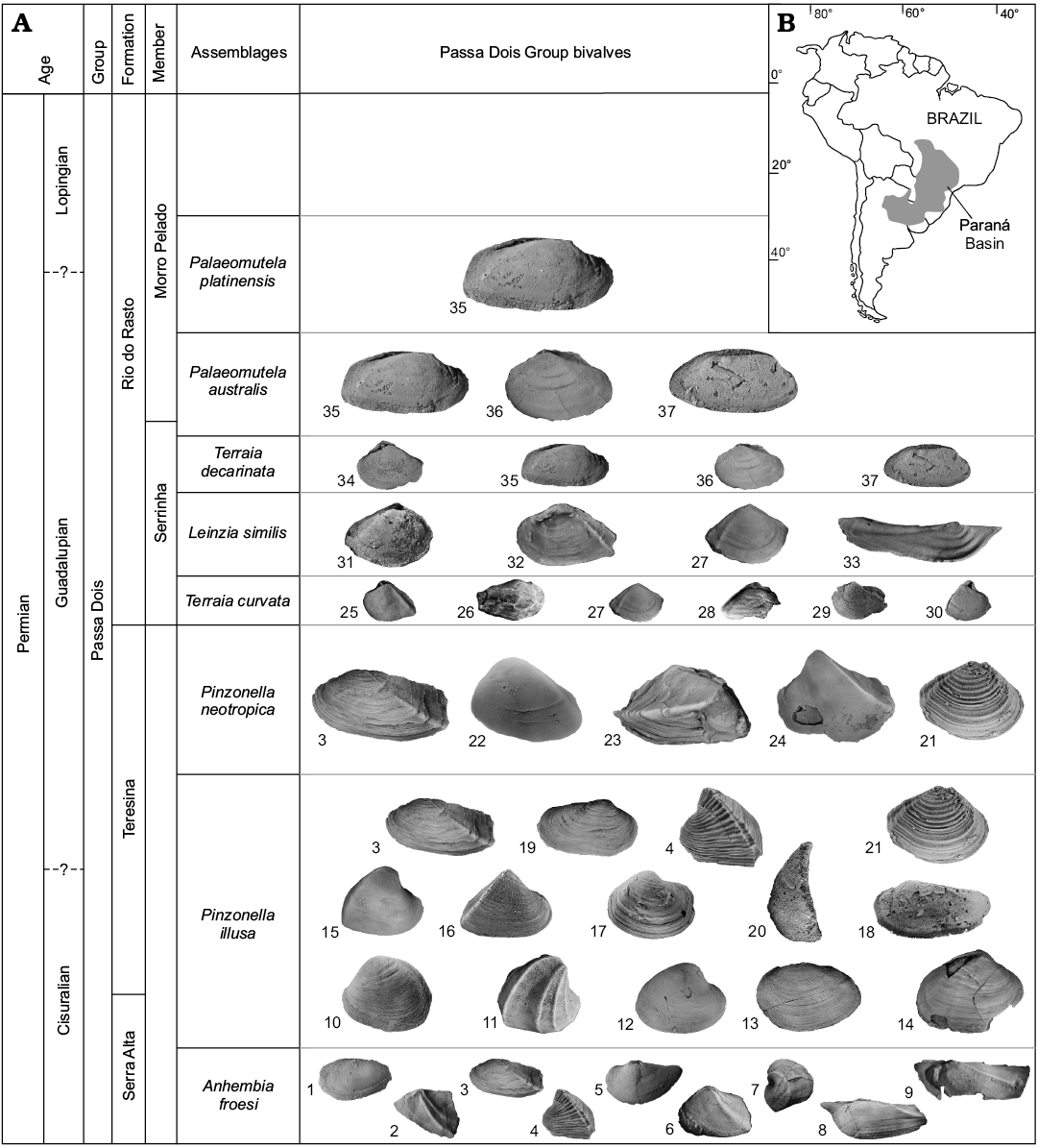
Fig. 1. Chart showing the main bivalve assemblages and constituent species recorded in the Permian Passa Dois Group, except the Irati Formation (A), and location of Paraná Basin, Brazil (B). Species composition (not to scale): 1, Kidodia cf. K. stockleyi; 2, Barbosaia angulata; 3, Holdhausiella elongata; 4, Tambaquyra camargoi; 5, Rioclaroa lefevrei; 6, Maackia contorta; 7, Ferrazia simplicicarinata; 8, Anhembia gigantea; 9, Anhembia froesi; 10, Pinzonella illusa; 11, Ferrazia cardinalis; 12, Casterella gratiosa; 13, Angatubia cowperesioides; 14, Othonella araguaiana, 15, Plesiocyprinella carinata; 16, Terraia aequilateralis; 17, Runnegariella fragilis; 18, Favalia arcuata; 19, Roxoa corumbataiensis; 20, Coxesia mezzalirai; 21, Cowperesia anceps; 22, Pinzonella neotropica; 23, Naiadopsis lamellosus; 24, Jaquesia brasiliensis; 25, Terraia curvata, 26, Beurlenella elongatella; 27, Cowperesia emerita; 28, “Leinzia” curta; 29, Terraia bipleura; 30, Astartellopsis prosoclina; 31, Oliveiraia pristina; 32, Terraia altissima; 33, Leinzia similis (sensu stricto); 34, Terraia decarinata; 35, Palaeomutela platinensis; 36, Relogiincola delicata; 37, Palaeomutela australis. Data from Mendes (1952), Runnegar and Newell (1971), Simões et al. (1998), Matos et al. (2017), and Guerrini et al. (2019).
The Passa Dois Group bivalve fauna is taxonomically diverse, and in some clades (e.g., Pachydomidae Fischer, 1887; Astartidae d’Orbigny, 1844) the extensive radiations were accompanied by high degrees of specialization of life habits (Runnegar and Newell 1971; Simões and Anelli 1995; Simões et al. 1998; Wesselingh 2007; Matos et al. 2017a). These resulted in relatively high genus and species diversity, with some forms showing a puzzling array of morphologies (see also Wesselingh 2007, for other similar Mesozoic and Cenozoic examples). This is the case, for example, of the pachydomid genera Anhembia Mezzalira, Mendes, and Maranhão, 1990, Maackia Mendes, 1954, Ferrazia Reed, 1932, and also Leinzia Mendes, 1949. Despite of the careful systematic revision of Runnegar and Newell (1971), and subsequent systematic studies of Simões et al. (1997, 2000, 2010, 2015, 2017), Mello (1999), David (2014), and Guerrini et al. (2019), the taxonomic position of some genera remains unclear, mainly due to the poor knowledge of their internal morphology. This is the case of Leinzia, a genus erected by Mendes (1949) based on specimens found in the Rio do Rasto Formation cropping out eight kilometers north of Mallet County, State of Paraná, southern Brazil. The genus is monotypic, with Solenomorpha similis Holdhaus, 1918 as the type species. Similar forms may occur in coeval Permian strata in Uruguay, Yaguari Formation (Morton and Herbst 1990), and Paraguay, Independencia Formation (see Morton and Herbst 1990). Due to its unusual morphology and the poor preservational quality of available specimens, the taxonomic position of Leinzia still remains obscure. Therefore, based on a detailed review of the topotype specimens described by Mendes (1949) and description of newly found specimens of Leinzia from the Serrinha Member, Rio do Rasto Formation, we shed light on the taxonomic position of this genus. Contrary to previous authors, the data gathered here strongly suggest a Pachydomidae affinity for Leinzia.
Institutional abbreviations.—DGP, palaeontological collection, Institute of Geosciences, University of São Paulo, São Paulo, Brazil; DZP, palaeontological collection, Department of Zoology, Institute of Biosciences, São Paulo State University, Botucatu campus, Botucatu, São Paulo, Brazil.
Historical background
A bivalve rather than spinicaudatan.—As noted by Runnegar and Newell (1971: 54), “Leinzia has such an extraordinary shape that we have toyed with the possibility that it may have been an unusual bivalved arthropod rather than a mollusc.” Indeed, the external ornamentation of Leinzia resembles that of some spinicaudatan crustaceans, leading some authors (Almeida 1950) to describe tiny specimens of Leinzia (and other bivalves) as true Spinicaudata (i.e., Acantholeaia Almeida, 1950; see Ferreira-Oliveira 2007 for a detailed discussion on this issue). Nevertheless, Runnegar and Newell (1971: 54) rejected that possibility because Leinzia shells have fine growth striae, small ligamental nymphs, well-developed teeth, and a non-rectilinear hinge. Consequently, Runnegar and Newell (1971) synonimized Acantholeaia in favor of Leinzia. Unfortunately, however, Tasch (1982) and Chen and Shen (1985) retained Acantholeaia as a valid genus of Branchiopoda Latreille, 1817, ignoring the data in Runnegar and Newell (1971).
Ferreira-Oliveira (2007: 72) re-examined the original material described by Almeida (1950) and Cardoso (1965), reposited in the National Department of Mineral Production (DNPM-RJ 4074-4077), Rio de Janeiro, Brazil. Ferreira-Oliveira (2007) noted that in addition to the observations made by Runnegar and Newell (1971), the irregularly spaced growth lines typically observed in bivalve mollusk shells are also visible between the comarginal striae of the shells ascribed to Acantholeaia. Yet, the gross morphology of the shells studied by Almeida (1950) resemble that of Holdhausiella Mendes, 1952 (DNPM-RJ 4074-4076) and Leinzia (DNPM-RJ 4648, 4075). In other words, the tiny bivalve shells in Almeida (1950) and Cardoso (1965) belong to bivalve mollusks (see Ferreira-Oliveira 2007) rather than Spinicaudata.
The uncertain taxonomic position of Leinzia.—Although the benchmark study of Runnegar and Newell (1971) provided increased understanding of the Permian endemic bivalves of the Passa Dois Group, the systematic relationships of Leinzia remained obscure (Table 1). This is due to the poor knowledge of the external and internal morphology of Leinzia shells, and the fact that the specimens illustrated by these and other authors differ greatly from the original material described by Mendes (1949), here referred as Leinzia sensu stricto. It should be noted here that both Holdhaus (1918) and Reed (1929) were unaware of the curious rostrum present in the anterior cardinal margin of Leinzia shells (see Runnegar and Newell 1971: 55). Holdhaus (1918) placed the shells in genus Solenomorpha Cockerell, 1903 (Cardiidia Férussac, 1822, Grammysiidae Miller, 1877). These were subsequently assigned to genus Cuspidaria Dall, 1886 (Poromyida Ridewood, 1903; Cuspidariidae Dall, 1886) by Reed (1929). Mendes (1949) was the first author to describe the rostrum of the shells of Leinzia. Based on this character, Mendes (1949) referred them to Pterioidea Gray, 1847. However, five years later the same author (Mendes 1954) changed his opinion because Leinzia clearly lacks numerous pterioid shell characters (see also Runnegar and Newell 1971: 55). Since then Mendes (1954) considered the familial position of Leinzia as uncertain.
Table 1. Summary of Leinzia similis (Holdhaus, 1918) sensu stricto, classification and shell morphology according several authors. Abbreviations: AA, anterior adductor muscle scar; AR, anterior retractor muscle scar; AP, anterior protractor muscle scar; LV, left valve; RV, right valve.
|
Classification |
Morphology |
Comparisons |
Reference |
||
|
rostrum |
hinge |
muscle scars |
|||
|
Solenomorphidae |
– |
RV with a triangular tooth |
– |
general resemblance with solenomorphids |
|
|
Pterioida Pteriacea |
present |
RV with a triangular cardinal
tooth, corresponding socket in LV; |
– |
general resemblance with pterioids |
|
|
Pholadomyida Edmondiacea Grammysiidae |
present |
LV with a single cardinal socket; lateral teeth absent |
– |
– |
|
|
Veneroida Crassatellacea |
present, well-developed |
RV with inclined triangular tooth below beak; well-defined socket in LV; small elongate tooth on posterior side of socket, fitting a narrow groove behind tooth in RV |
undescribed |
hinge as in other Permian Passa Dois Group crassatellaceans |
|
|
?Orthonotacea Pholadomyacea Sanguinolitidae |
– |
– |
– |
– |
|
|
Pholadomyida |
– |
– |
– |
– |
|
|
Hiatellida Edmondioidea Pachydomidae |
present, well-developed |
RV with subumbonal blunt triangular tooth; lateral teeth absent |
AA small, reniform; AR well defined, separated from AA; AP attached to dorsal edge of AA. |
hinge/muscle scars as in other Passa Dois Group pachydomids |
this paper |
Based on the pronounced anterior elongation (= rostrum) of the Leinzia shells, Newell (1969: N820) tentatively assigned Leinzia to the Grammysiidae Miller, 1877. Subsequently, Runnegar and Newell (1971) referred Leinzia to an incertae familiae of Crassatellacea Férussac, 1822. Seemingly, Runnegar and Newell (1971) were influenced by comments in Holdhaus (1918), Leanza (1948), and Beurlen (1954) that noted the hinge similarities of Leinzia similis and Terraia altissima (Holdhaus, 1918). Runnegar and Newell (1971: 51) offered a detailed description of the hinge of Terraia altissima, which is very complex, as follows: “hinge with large triangular cardinal tooth in right valve and a second obscure tooth between large tooth and ligament nymph; left valve has, correspondingly, obvious triangular socket bordered posteriorly by narrow tooth and socket; anterior and posterior dorsal margins of left valve function as lateral teeth by fitting beneath edges of right valve; obscure posterior lateral tooth present in right valve but no corresponding tooth below anterior lateral socket.” These authors also mentioned that as “the affinities of Terraia are not well understood, Leinzia is even more difficult to classify” (Runnegar and Newell 1971: 55). The problem with the above comparisons is that they are mainly based on poorly preserved specimens, especially from Leinzia.
Subsequent authors have tentatively listed Leinzia in Sanguinolitidae Miller, 1877 (Morris et al. 1991), Megadesmidae Vokes, 1967 (Mello 1999; David 2010) or Pholadomyida Newell, 1965 (Sepkoski 2002). Unfortunately, Morris et al. (1991) and Sepkoski (2002) did not explicitly explain the reasons for assigning Leinzia to Sanguinolitidae or Pholadomyida. On the other hand, Mello (1999) and David (2010) reexamined and illustrated the original material described by Mendes (1949), showing in their studies that the hinge in Leinzia is clearly distinct of that of Terraia altissima (Table 1). This prompted us to reinterpret the hinge of Leinzia as well as to describe the preserved muscle scars, unknown characters until now.
In summary, the uncertainties about the taxonomic affinities of Leinzia are mainly related to poor knowledge of several shell characters, including the (i) rostrum, (ii) hinge architecture, and (iii) arrangement of the preserved muscle scars, which are all described in detail, illustrated and discussed in this contribution.
Material and methods
We examined Mendes’ (1949) topotype specimens from the middle part of the Serrinha Member of the Rio do Rasto Formation at a locality eight kilometers north of Mallet County, State of Paraná, southern Brazil (Figs. 2). Holdhaus (1918) and Reed (1929) obtained their specimens from the same location. The material of Mendes (1949) consists of three mudstone slabs (DGP/7-85, 86, 88) containing at least eight specimens of Leinzia similis. The specimens are not individually numbered in the slabs. These slabs should have been part of a shell pavement with densely packed valves. The slabs are stored in the palaeontological collection of the Institute of Geosciences, University of São Paulo, São Paulo, Brazil. In addition to Mendes’ (1949) material, four specimens of Leinzia similis were also collected in mudstone layers from the Serrinha Member, cropping out on BR-373 road, km 277, near Prudentópolis city, 25°15’17.87”S, 51°3’56.51”W, State of Paraná. These additional specimens are deposited in the palaeontological collection of the São Paulo State University, Botucatu campus, São Paulo State, Brazil. The Mendes’ (1949) and the new additional fossil material are both represented by molds, mainly composite ones. In some specimens the plastic deformation of the shell is clearly visible, but signs of shell breakage by compression are missing. The shells of Leinzia similis (sensu stricto) were almost certainly aragonitic originally and probably thin.
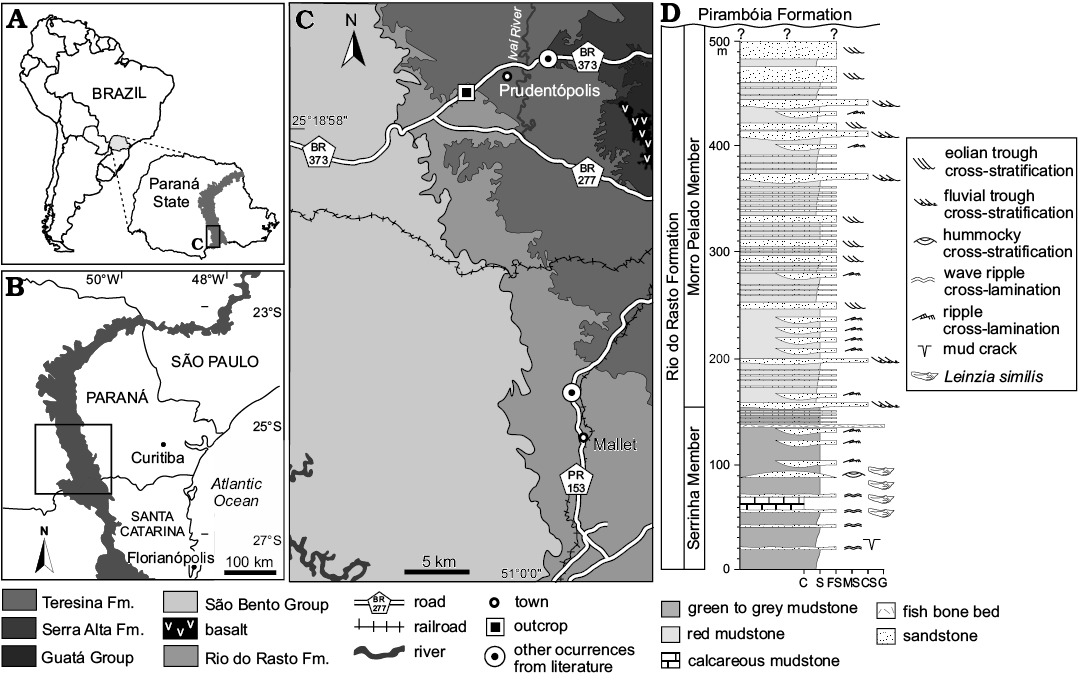
Fig. 2. A. Location of the studied area in the southern part of the Paraná State, south Brazil. The outcrop belt of the Passa Dois Group is in grey. B. Map of the Paraná State, Brazil (frame indicates the southern part where studied area is located) (modified from Rohn and Rösler 2000; Guerrini et al. 2019). C. Geological map showing the main geographic area of occurrence of Leinzia similis (sensu stricto). Cartographic base: Mineropar 2006 (Mapa geológico do Estado do Paraná, 1:250 000). D. Columnar section of the Rio do Rasto Formation; note the constrained stratigraphic distribution of Leinzia similis (sensu stricto) in the Serrinha Member. Abbreviations: C, calcareous mudstone; CS, coarse sandstone; G, gravel; FS, fine sandstone; MS, medium sandstone; S, siltstone.
Higher taxa used in this study are based on Bouchet et al. (2010), and the morphological terms used to describe internal and external shell characters of Leinzia followed Runnegar and Newell (1971), Runnegar (1974), Simões et al. (1997), and Carter et al. (2012). A key character in the description of Leinzia is the unique anterior rostrum of the shell. However, a varied terminology (i.e., prolongation, prong, anterior prolongation, rostrum, and beak) was applied to it in previous studies. The term rostrum is used here in order to indicate an elongate extension of the anterior margin of the shell (see also Carter et al. 2012: 155). Thus, the shells bearing this feature are here referred to as rostrate. Finally, the material was photographed in detail using a Nikon digital camera (D3200), with a macro lens (105 mm). Individual digital images were merged using the focus stacking technique in Adobe Photoshop CS6 software, and drawnings were prepared employing Corel Draw X8 software.
Systematic palaeontology
Order Pholadomyida Newell, 1965
[= Anomalodesmata Dall, 1889]
Superfamily Edmondioidea King, 1850
Family Pachydomidae Fischer, 1887
Subfamily Pachydominae Fischer, 1887
Genus Leinzia Mendes, 1949
= Acantholeaia Almeida, 1950
Type species: Solenomorpha similis Holdhaus, 1918, by original designation; see below.
Diagnosis.—As for the type species by monotypy, emended.
Leinzia similis (Holdhaus, 1918)
Figs. 3–6.
1918 Solenomorpha similis sp. nov.; Holdhaus 1918: 6, pl. 1: 1–5.
1929 Cuspidaria similis Holdhaus, 1918; Reed 1929: 45, pl. 4: 1–5, 11?
1949 Leinzia similis Holdhaus, 1918; Mendes 1949: 14, pl. 1: 1–5.
1971 Leinzia similis Holdhaus, 1918; Runnegar and Newell 1971: 53, pl. 24: D–G, J.
Type material: The specimens illustrated by Holdhaus (1918: pl. 1) from the Rio do Rasto Formation. Neotype: Since the whereabouts of Holdhaus (1918) type series are unknown and he provided only drawings in his original paper we consider it appropriate to designate a neotype (DGP/7-85) from Mendes (1949) topotypic collection.
Type locality: Outcrops of the Rio do Rasto Formation located about eight km NW of the town of Mallet (formerly São Pedro de Mallet), Paraná State, southern Brazil.
Type horizon: Basal interval of the Rio do Rasto Formation, Serrinha Member, Passa Dois Group, Paraná Basin, southern Brazil.
Diagnosis.—Posteriorly elongated (length to height ratio 2.22 to 4.16), compressed, and non-gaping shell. Anterior shell margin straight, with a well-defined rostrum. Hinge of the right valve with a large, anteriorly inclined, blunt triangular tooth beneath the umbo, and a corresponding socket in left valve; true lateral teeth absent. Shell ornamentation with regularly spaced comarginal rugae, also present in the rostrum, but absent in the posterior umbonal carina. Anterior adductor muscle scar small, reniform; anterior pedal retractor scars well defined and separated from anterior adductor; anterior pedal protractor scar attached to dorsal edge of anterior adductor scar. Other muscle scars and palial line unkown.
Measurements.—See Table 2 for Leinzia similis (sensu stricto) shell measurements.
Table 2. Measurements (in mm) of the valves of Leinzia similis (Holdhaus, 1918) sensu stricto.
|
Specimen |
Valve |
Length (L) |
Height (H) |
Elongation Index (L/H) |
|
DZP-20416 A |
right |
19 |
8 |
2.37 |
|
DZP-20416 B |
right |
20 |
9 |
2.22 |
|
DZP-20417 |
right |
28 |
9 |
3.11 |
|
DGP/7-85 |
left |
25 |
6 |
4.16 |
|
DGP/7-86 |
left |
36 |
11 |
3.27 |
|
DGP/7-88 |
left |
28 |
11 |
2.54 |
Description.—Small (maximum observed length 36 mm), thin-shelled, compressed bivalve, equivalve, posteriorly elongate (Figs. 3, 5B2), and with a well-defined rostrum (Fig. 5B). Dorsal and ventral margins are subparallel; posterior margin sligthly concave (Figs. 3A3, D, 5B2); cardinal margin straight in front of beak. Posterior carina well marked, running diagonally from beak to posterior-ventral shell margin (Fig. 3). Umbones low, prosogyrous; lunule absent, escutcheon long, well-defined. Ligament probably opisthodetic, parivincular, external. Hinge of right valve with large, anteriorly inclined triangular blunt tooth beneath the umbo and a corresponding socket in left valve; true lateral teeth absent (Fig. 4A1, A2, B). External valve ornamentation with regularly spaced comarginal rugae extending to the anterior prolongation but absent behind umbonal carina (Figs. 3, 5B). Fine growth lines visible between the regularly spaced comarginal rugae (Figs. 3A3, 5B2). The rugae are also evident on inner shell surfaces. Small lamellose or spinose projections along posterior dorsal shell margin are apparently absent. Anterior adductor muscle scar small, reniform; anterior pedal retractor scars well defined, separated from anterior adductor; anterior pedal protractor scar attached to dorsal edge of anterior adductor scar (Fig. 4A3, A4). Other muscle scars and pallial line unkown.

Fig. 3. Pachydomid bivalve Leinzia similis (Holdhaus, 1918) (sensu stricto), composite molds, Rio do Rasto Formation, Guadalupian (Permian), Paraná Basin, Brazil. A. DGP/7-88, left valve (A1) with well-defined, forwardly inclined, triangular socket (arrowed); right valve (A2), note the deformed anterior rostrum margin of the shell (arrowed); splayed open valves (A3). B. DZP-20417, right valve, note the well-developed triangular blunt tooth (arrowed). C. DZP-20416A, right valve showing well-defined escutcheon (arrowed). D. DGP/7-86, a slightly deformed specimen with articulated valves and broken anterior margin, note the straight slightly curved posterior margin of the shells. Scale bars 5 mm.
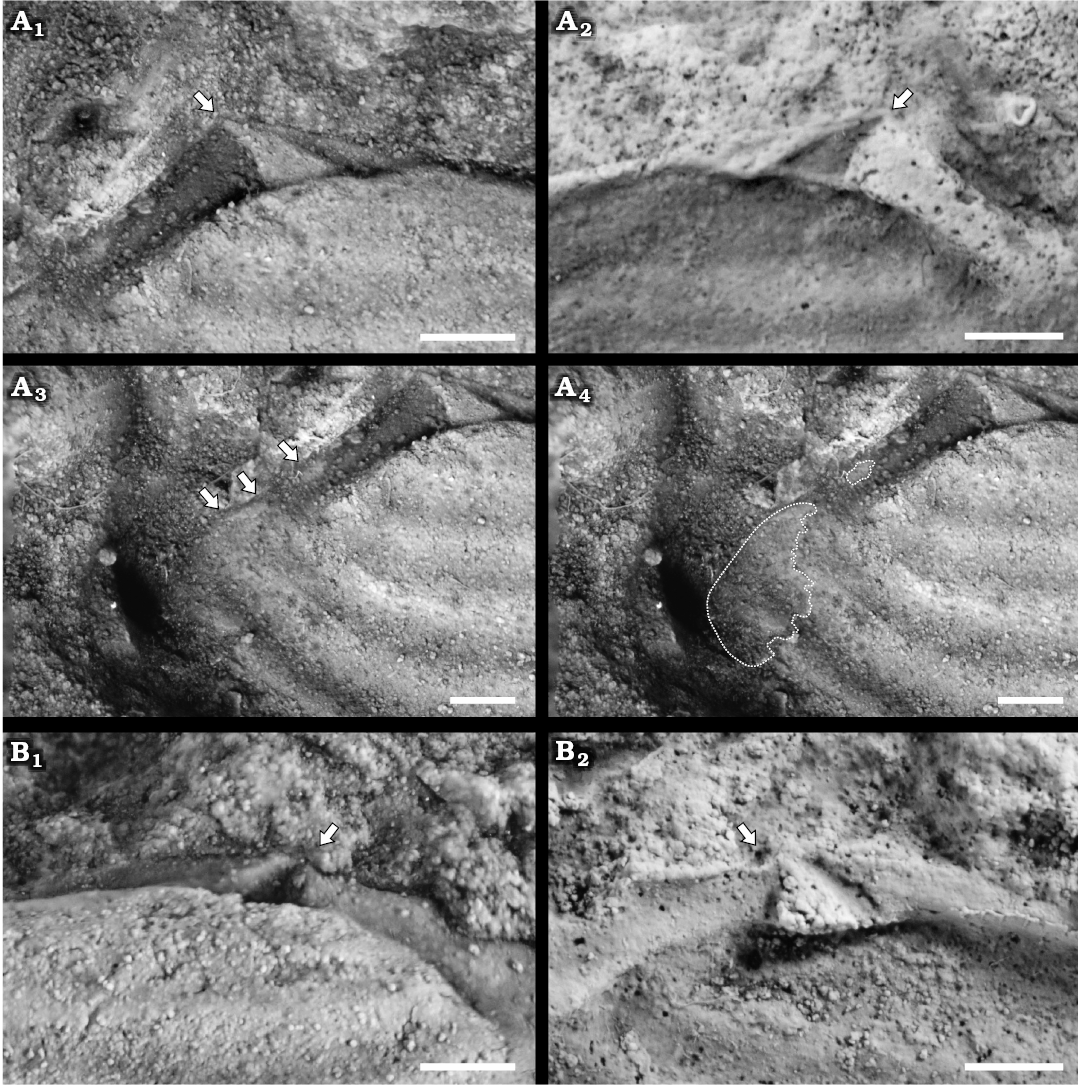
Fig. 4. Pachydomid bivalve Leinzia similis (Holdhaus, 1918) (sensu stricto), Rio do Rasto Formation, Guadalupian (Permian), Paraná Basin, Brazil. A. DGP/7-88, composite mold, left valve; A1, detail of the well-defined triangular socket (arrowed); A2, latex cast of specimen in A1, showing the hinge with well-defined triangular socket (arrowed); also note the absence of true lateral teeth; A3, details of the anterior adductor, pedal protractor, and pedal retractor muscle scars (arrows); A4, anterior muscle scars (outlined). B. DZP-20417, composite mold, right valve; B1, well-developed triangular, subcardinal, blunt tooth (arrowed); B2, latex cast showing the triangular socket (arrowed) and the hinge without true lateral teeth. Scale bars 1 mm.

Fig. 5. Pachydomid bivalve Leinzia similis (Holdhaus, 1918) (sensu stricto), Rio do Rasto Formation, Guadalupian (Permian), Paraná Basin, Brazil. A. DGP/7-88, right valve, detail of the cardinal margin showing a strongly deformed rostrum (arrowed). B. DGP/7-85, external mold, left valve; B1, the comarginal rugae on the anterior rostrum (arrow); B2, general view of the shell with well-preserved anterior rostrum. Scale bars A, B1, 1 mm; B2, 5 mm.

Fig. 6. Drawing of the pachydomid bivalve Leinzia similis (Holdhaus, 1918) (sensu stricto) showing the general morphology. The hinge area and anterior muscles scars are highlighted. Abbreviations: aa, anterior adductor muscle scar; app, anterior pedal protractor muscle scar; apr, anterior pedal retractor muscle scar; sck, socket; th, tooth.
Remarks.—The right valve hinge of Leinzia similis (sensu stricto) with its large, forwardly inclined triangular blunt tooth beneath the beak and corresponding socket in the left valve is similar to that of various Permian Pachydomidae bivalves. However, the gross morphology of the rostrate shell of Leinzia similis differs from all other known Pachydomidae. The general shape, hinge, and muscle scars of Anhembia Mezzalira, Mendes, and Maranhão, 1990 (Fig. 7B), the other rostrum-bearing Pachydomidae shell of the Passa Dois Group are distinct of Leinzia similis (see discussion in Mezzalira et al. 1990). The anterior muscle scars of Leinzia similis (sensu stricto) are also similar to those found in Pachydomidae (i.e., Cowperesia Mendes, 1952, Pyramus Dana, 1847, Astartilla Dana, 1847, Plesiocyprinella Holdhaus, 1918), as described in Runnegar (1965) and Runnegar and Newell (1971), for example. The hinge and external ornamentation of Leinzia similis resemble those of Cowperesia anceps (Reed, 1935) and, in addition, both shells are also laterally compressed. However, the general shell shape of Leinzia similis (sensu stricto) is completely distinct from that of Cowperesia anceps. Yet, the shells of Cowperesia anceps have a small, well-defined palial sinus and siphonal gape, which are absent in Leinzia similis. The posteriorly elongated shells of Leinzia similis (sensu stricto) with a well-marked posterior carina resemble that of Holdhausiella elongata (Holdhaus, 1918). Notably, the cardinal margin of some shells of this species have an anterior prolongation, but not a well-defined rostrum (Fig. 7A; see also Runnegar and Newell 1971: 41, fig. 16j–h). Yet, the external ornamentation of Holdhausiella elongata has irregularly spaced growth lines (Fig. 7A), which are clearly different from the regularly spaced comarginal rugae in Leinzia similis. Yet, the hinge of Holdhausiella elongata is edentelous and therefore different from that of Leinzia similis.
Although the external ornamentation of shells of Huabiella compressa David, Simões, Anelli, Rohn, and Holzförster, 2011, from the Permian Gai-As Formation, Huab Basin, Namibia, are similar to that of Leinzia similis (sensu stricto), small lamellose or spinose projections along the posterior dorsal margin are only recorded in the Namibian shells. Unfortunately, the anterior margin, hinge and muscle scars are still unknown in Huabiella compressa. Therefore, a more precise comparison between Leinzia and Huabiella is currently not feasible. Finally, the hinge and gross morphology of Terraia altissima and other Terraiinae Scarlato and Starobogatov, 1979 bivalves (Terraia aequilateralis Mendes, 1952, Terraia curvata [Reed, 1929], Terraia bipleura [Reed, 1929]) are completely distinct of Leinzia similis (sensu stricto) and, therefore, the comparison among them is groundless.
Stratigraphic and geographic range.—Middle part of the Guadalupian Serrinha Member of the Rio do Rasto Formation, Paraná State, southern Brazil (see Rohn 1994; Rohn and Rösler 2000).

Fig. 7. Pachydomid bivalves from the Guadalupian (Permian), Teresina and Serra Alta formations, respectively, Paraná Basin, Brazil. A. Holdhausiella elongata (Holdhaus, 1918), DZP-20441, silicified shell, left valve, note the pointed anterior margin of the shell. B. Anhembia gigantea (Mendes, 1949), DGP/7-91, composite mold, left valve; B1, detail of a well-developed rostrum, B2, general view. Scale bars 5 mm.
Discussion
Possible taphonomically-produced morphological shell character variants.—The shells of Leinzia similis (sensu stricto) were originally thin and the specimens are preserved as compressed composite molds. Consequently, the original gross morphology of the shells was subject to distortion and plastic deformation during diagenesis. Based on the examined collection and specimens in the literature three main characters are susceptible to distortion due to taphonomic processes, as follows: (i) the anterior rostrum, (ii) the dorsal margin, and (iii) the umbonal carina. Indeed, in the specimen DGP/7-88 the anterior rostrum is deeply slanted due to compressional plastic deformation of the umbonal area of the shell (Figs. 3A2, 5A). Therefore, a much pronounced taphonomically produced anterior rostrum was generated. In the specimen DZP-20417, the cardinal margin of the shell is also deformed and broken (Fig. 3B), causing substantial modification of the original shell outline. Thus, the anterior margin of the shell of these specimens is obviously distinct of that of non-distorted ones (see Fig. 5B; DGP/7-85). Yet, the rostrum is lacking (broken?) in most of the specimens available in the examined collections as well as in the individuals assigned to Leinzia similis by previous authors. The rostrum is also missing in the specimens of Holdhaus (1918) and Reed (1929), suggesting that this was thin and very fragile.
The dorsal margin of the shell is the other character that is apparently distorted in compressed specimens. Indeed, in non-deformed individuals or less compacted ones the dorsal margin is slightly encurved, whereas in compressed examplars it is straight (Fig. 3A2, B, C). The same also occurs with the shape and width of the umbonal carina that is also slightly encurved in those specimens that were subject to lateral compression (Fig. 3A2).
Leinzia similis in the literature.—Aside the specimens of Leinzia similis described by Holdhaus (1918), Reed (1929), and Mendes (1949), other important mentions to this species appear in Beurlen (1954) and Morton and Herbst (1990), including specimens from Brazil and Uruguay, respectively. However, it was Runnegar and Newell (1971) that detailed described, illustrated and compared Leinzia similis with other Permian, endemic Passa Dois Group bivalves. These authors well figured five bivalve specimens of Leinzia similis from the Serrinha Member, Rio do Rasto Formation, from a road cut at km 109.7 between the towns of Prudentópolis to Guarapuava (USGS locality 22836), Paraná State, southern Brazil (Runnegar and Newell 1971: 54, fig. 24d–g, j). Three of these Leinzia specimens (Runnegar and Newell 1971: fig. 24e–g) are poorly preserved, broken shells, and the remaining are more complete valves, one splayed specimen and a single right valve (Runnegar and Newell 1971: fig. 24d, j, respectively). Notably, the anterior shell margin is missing in three specimens (Runnegar and Newell 1971: 54, fig. 24d, e, g), and is also poorly preserved in the articulated one (Runnegar and Newell 1971: 54, fig. 24j). The only specimen where this character is clearly evident is the one in their fig. 24f. Despite the rostrum, and the rugose external ornamentation, there are significant morphological differences in the anterior margin and hinge of the specimens in Runnegar and Newell (1971) and those in the topotype material of Leinzia by Mendes (1949) (Figs. 8, 9B; see also Runnegar and Newell 1971: fig. 24f, j). For example, the cardinal margins of the shells in Runnegar and Newell (1971) is not straight and, therefore, the shells are not as strongly beaked as those in the topotype material of Leinzia similis (sensu stricto) (Fig. 8). Indeed, the cardinal margin of the specimen in Runnegar and Newell (1971: fig. 24f) is concave and so distinctly different from the specimens of Leinzia similis in Mendes (1949) that its taphonomic origin cannot be ruled out.
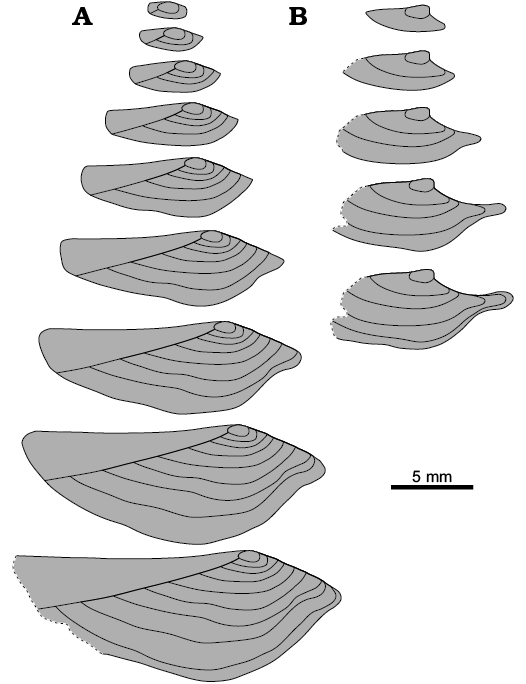
Fig. 8. Ontogenetic series of the pachydomid bivalve Leinzia sp. (sensu lato) from the Serrinha Member, Rio do Rasto Formation after Runnegar and Newell (1971: fig. 24). Sketches based on external comarginal ornamentation (rugae and growth lines) of individual shells. A, B, based on Runnegar and Newell 1971: fig. 24j, f, respectively. Note the differences in the shell shape of both specimens and those illustrated in Fig. 9; also, note the ubiquitous rostrum in the specimen in Fig. 9B.
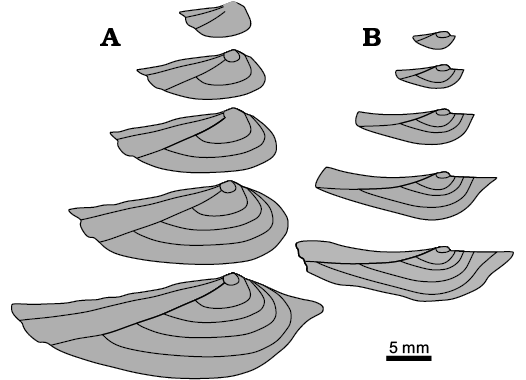
Fig. 9. Ontogenetic series of pachydomid bivalves Leinzia similis (sensu lato) (A) from the Yaguari Formation (after Morton and Herbst 1990) and Leinzia similis (sensu stricto) DGP/7-85 (B) from the Serrinha Member, Rio do Rasto Formation. Sketches based on external comarginal ornamentation (rugae and growth lines) of individual shells. Note the differences in the shell shape of both specimens: double posterior carina (A), the anterior rostrum developed since the early ontogenetic stages (B).
Additionally, Runnegar and Newell (1971) described the hinge of Leinzia similis in detail, since this character is well visible in the specimens in their fig. 24e–g. According to them, the hinge has an inclined triangular tooth below beak of the right valve, a well-defined socket in left valve, and a small elongate tooth on posterior side of the socket, fitting a narrow groove behind tooth in right valve. Therefore, the hinge of the Leinzia similis in Runnegar and Newell (1971) is slightly more complex of that in Leinzia similis (sensu stricto) of Mendes (1949), where a true lateral tooth is absent (Fig. 4A1, A2, B). Yet, the shape and size of the triangular tooth of Leinzia similis is distinct of that in the specimens of Runnegar and Newell (1971). Unfortunately, the musculature is not preserved in the material described by these authors and, thus, a more precise comparison between them is not permitted. In summary, based on the original material of Mendes (1949) and the additional specimens here described, the rostrate and rugose specimens in Runnegar and Newell (1971) are distinct of Leinzia similis (sensu stricto) and a revision of these is strongly advised.
Leinzia similis in Morton and Herbst (1990) from the Permian Yaguary Formation, Uruguay, is also distinct of those in Mendes (1949) topotype specimens. Unfortunatly, the hinge and musculature of the Uruguayan specimen is not preserved (Morton and Herbst 1990: 96, fig. 1). The unique resemblance with Mendes (1949) material is the posteriorly elongated nature of the shell and its rugose external ornamentation. As for other specimens in the literature, the rostrum is not fully preserved in the specimen figured by Morton and Herbst (1990), and the shape and size of this character is only inferred. Notably, the main difference is that the Uruguayan specimen has a double carina (Fig. 7A), which are missing in the specimens of Leinzia similis (sensu stricto).
Conclusions
During the late Permian distinct groups of endemic bivalves from the Passa Dois Group in Brazil and coeval strata in Uruguay as well as in the Huab Basin, Namibia, developed small, posteriorly elongated, rostrate shells with rugose ornamentation (see also David et al. 2011: 504, fig. 5). Traditionally, these shells (except those from Namibia) were all assigned to Leinzia. However, detailed examination of the original collection of Mendes (1949) indicates that the shells of Leinzia are distinct from those specimens described in Runnegar and Newell (1971), and Morton and Herbst (1990). Since we can rule out the hypothesis that taphonomic processes distorted the original gross morphology of the shells in Runnegar and Newell (1971), and Morton and Herbst (1990), we suggest a taxonomic revision before splitting them from Leinzia. However, it should be remembered here that the hinge of some specimens illustrated by Runnegar and Newell (1971: 54, fig. 24e, f) seems distinct from those in the type specimens of Leinzia similis.
The confusion with the tooth of Leinzia seems to have begun with Beurlen (1954: 128), when he commented (but not illustrated) that the hinge of this genus is the same as in Terraiopsis Beurlen, 1954 (= Terraia in Runnegar and Newell 1971: 50). According to our observations, the hinge of Leinzia similis (sensu stricto) has close resemblance with that described both in Mendes (1949) and Newell (1969). The hinge of the right valve of Leinzia similis (sensu stricto) with a subcardinal, triangular blunt tooth and corresponding socket in left valve, and no lateral teeth is extremely similar to that present in some Permian endemic Pachydomidae bivalves (i.e., Cowperesia) of the Passa Dois Group, Brazil. In this context, Leinzia similis (sensu stricto) with its unique set of curious shell characters may correspond to a highly derived Pachydomidae bivalve.
Acknowledgements
The authors are indebted to Ivone Cardoso Gonzales, and Juliana de Moraes Leme (both Institute of Geosciences, University of São Paulo, São Paulo, Brazil) for granting access to Josué C. Mendes original type collection. We thank an anonymous referee and Elizabeth M. Harper (University of Cambridge, UK) whose corrections, comments and suggestions substantially improved the final version of the manuscript. Partial financial support for this research was provided by CAPES and CNPq to VBG and MGS, respectively.
References
Almeida, F.F.M. 1950. Acantholeaia, um novo gênero de Leaiadidae. Notas Preliminares e Estudos da Divisão de Geologia e Mineralogia 51: 1–10.
Beurlen, K. 1954. As faunas de lamelibrânquios do sistema gonduânico no Paraná. In: F.W. Lange (ed.), Volume Comemorativo do 1° Centenário do Estado do Paraná, 107–136. Governo do Estado do Paraná, Curitiba.
Bouchet, P., Rocroi, J.-P., Bieler, R., Carter, J.G. and Coan, E.V. 2010. Nomenclator of bivalve families with a classification of bivalve families. Malacologia 52: 1–184. Crossref
Cardoso, R.N. 1965. Sobre a ocorrência no Brasil de Monoleiolophinae e Afrograptidae, conchostráceos carenados. Boletim da Divisão de Geologia e Mineralogia 221: 1–35.
Carter, J.G., Harries, P.J., Malchus, N., Sartori, A.F., Anderson, L.C., Bieler, R., Bogan, A.E., Coan, E.V., Cope, J.C.W., Cragg, S.M., García-March, J.R., Hylleberg, J., Kelley, P., Kleemann, K., Kříž, J., McRoberts, C., Mikkelsen, P.M., Pojeta Jr., J., Tëmkin, I., Yancey, T. and Zieritz, A. 2012. Illustrated Glossary of the Bivalvia. Treatise Online: Treatise on Invertebrate Paleontology, Part N, Revised, Volume 1, Chapter 31 48: 1–209.
Chen, P.J. and Shen Y.B. 1985. An Introduction to Fossil Conchostracans. 241 pp. Science Press, Beijing.
Cockerell, T.D.A. 1903. The name Solenopsis. Nature 67: 5. Crossref
Dall, W.H. 1886. Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877–1878) and in the Caribbean Sea (1879–1880), by the U.S. Coast Survey steamer “Blake” XXIX. Report on the Mollusca. Part 2, Gastropoda and Scaphopoda. Bulletin of the Museum of Comparative Zoology, Harvard University 18: 1–492.
Dall, W.H. 1889. On the hinge of pelecypods and its development, with an attempt toward a better subdivision of the group. American Journal of Science and Arts 228: 445–462. Crossref
Dana, J.D. 1847. Descriptions of fossil shells of the collections of the exploring expedition under the command of Charles Wilkes, U.S.N., obteined in Australia, from lower layers of the coal formation in Illawarra, and from a deposit probably of nearly the same age at Harper’s Hill, Valley of the Hunter. American Journal of Science 54: 151–160.
David, J.M. 2010. Bivalves Permianos da fase de continentalização das bacias do gondwana ocidental: Sistemática, Paleogeografia e Bioestratigrafia. 99 pp. M.Sc Thesis, Universidade Estadual Paulista, Rio Claro. Available at https://repositorio.unesp.br/handle/11449/92880. Crossref
David, J.M. 2014. Bivalves permianos do Grupo Passa Dois, Bacia do Paraná, Brasil e do Grupo Ecca, Bacia do Karoo, África do Sul: Implicações bioestratigráficas e paleoambientais. 165 pp. Ph.D Dissertation, Universidade de São Paulo, São Paulo.
David, J.M., Simões, M.G., Anelli, L.E., Rohn, R., and Holzförster, F. 2011. Permian bivalve molluscs from the Gai-As Formation, northern Namibia: systematics, taphonomy and biostatigraphy. Alcheringa 35: 497–516. Crossref
Ferreira-Oliveira, L.G. 2007. Conchostráceos permianos da Bacia do Paraná: taxonomia, evolução, bioestratigrafia e paleogeografia. 241 pp. Ph.D Dissertation, Universidade Estadual Paulista, Rio Claro. Available at https://repositorio.unesp.br/handle/11449/102996. Crossref
Férussac, A.E. 1822. Tableaux systématiques des animaux mollusques. 190 pp. A. Bertrand, Paris.
Fischer, P.H. 1887. Manuel de Conchyliologie et de Paléontologie Conchyliologique, ou Histoire Naturelle des Mollusques Vivants et Fossiles, 1009–1369. F. Savy, Paris. Crossref
Gray, J.E. 1847. A list of the genera of Recent Mollusca, their synonyma and types. Zoological Society of London, Proceedings 15: 129–219.
Guerrini, V.B., Matos, S.A., Rohn, R., Varejão, F.G., Warren, L.V., and Simões, M.G. 2019. The last chapter of 30 million years of molluscan evolution: Permian non-marine bivalves of the Rio do Rasto Formation, Paraná Basin, Brazil. Paläontologische Zeitschrift 23: 1–26. Crossref
Holdhaus, K. 1918. Sobre alguns lamelibrânquios fósseis do sul do Brasil. Serviço Geológico e Mineralógico 2: 1–24.
Holz, M., França, A.B., Souza, P.A., Iannuzzi, R., and Rohn, R. 2010. A stratigraphic chart of the Late Carboniferous/Permian succession of the eastern border of the Paraná Basin, Brazil, South America. Journal of South American Earth Sciences 29: 381–399. Crossref
King, W. 1850. Monograph of the Permian fossils of England. Palaeontographical Society of London 3: 1–258. Crossref
Latreille, P.A. 1817. Les crustacés, les arachnides, et les insectes. In: G.L.C.F.D. Cuvier (ed.), Le Regne Animal, distribue d’apres son organisation, pour servrir de base a l’histoire naturelle des animaux et d’introduction a l’anatomie comparee, Volume 3, 584 pp. Chez Déterville, Paris.
Leanza, A.F. 1948. El llamado Triasico marino de Brasil, Paraguay, Uruguay y la Argentina. Revista de la Asociación Geológica Argentina 3: 219–244.
Matos, S.A., Warren L.V., Fürsich, F.T., Alessandretti, L., Assine M.L., Riccomini, C., and Simões, M.G. 2017a. Paleoecology and paleoenvironments of Permian bivalves of the Serra Alta Formation, Brazil: Ordinary suspension feeders or Late Paleozoic Gondwana seep organisms? Journal of South American Earth Sciences 77: 21–41. Crossref
Matos, S.A., Warren, L.V., Varejão, F.G., Assine, M.L., and Simões, M. G. 2017b. Permian endemic bivalves of the “Irati anoxic event”, Paraná Basin, Brazil: Taphonomical, paleogeographical and evolutionary implications. Paleogeography, Paleoclimatology, Paleoecology 469: 18–33. Crossref
Mello, L.H.C. 1999. Análise cladística dos bivalves do Grupo Passa Dois (Neopermiano), Bacia do Paraná, Brasil: implicações taxonômicas, evolutivas e paleobiogeográficas. 160 pp. M.Sc Thesis, Universidade São Paulo, São Paulo. Available at https://10.11606/D.44.1999.tde-05102015-131815. Crossref
Mendes, J.C. 1949. Novos Lamelibrânquios fósseis da Série Passa Dois, sul do Brasil. Divisão Geológica e Mineralógica do Brasil, Boletim da Divisão de Geologia e Mineralogia 133: 1–40.
Mendes, J.C. 1952. A Formação Corumbataí na região do Rio Corumbataí. (Estratigrafia e descrição dos lamelibrânquios). Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo 8: 1–119. Crossref
Mendes, J.C. 1954. Contribuição à estratigrafia da Série Passa Dois no Estado do Paraná. Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo 10: 1–119. Crossref
Mezzalira, S., Mendes, J.C., and Maranhão, M.S.M.A. 1990. Anhembia: um novo gênero de bivalves do Grupo Passa Dois-Permiano. Revista do Instituto Geológico 11: 51–54. Crossref
Milani, E.J., Melo, J.H.G., Souza, P.A., Fernandes, L.A., and França, A.B. 2007. Bacia do Paraná. Boletim de Geociências da Petrobras 15: 265–287.
Miller, S.A. 1877. The American Palaeozoic Fossils, a Catalog of the Genera and Species. 253 pp. Published by author, Cincinnati.
Morris, N.J., Dickins, J.M., and Astafieva-Urbajtis, K. 1991. Upper Paleozoic anomalodesmatan bivalves. Bulletin of the British Museum of Natural History 47: 51–100.
Morton, L.S. and Herbst, R. 1990. Leinzia similis (Holdhaus), (Pelecypoda) del permico superior (Formacion Yaguari) del Uruguay. Revista de la Associación de Ciencias Naturales del Litoral 21: 95–97. Crossref
Newell, N.D. 1965. Classification of the Bivalvia. American Museum Novitates 2206: 1–25.
Newell, N.D. 1969. Grammysiidae Miller, 1877. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part N, Mollusca 6, Bivalvia Vol. 2, N819–823. The Geological Society of America, Universidade do Kansas, Lawrence.
d’Orbigny, A. 1844–1848. Lamellibranches. In: A. d’Orbigny, A. Cotteau, G.H. Mr. Piette, Eudes-Deslongchamps, E. De Loriol-Le Fort, P. Saporta, G. De Fromentel, E., and Mr. Ferry (eds.), 1840–1894, Paléontologie Française. Description Zoologique et Géologique de Tous le Animaux Mollusques et Rayonnés Fossiles de France: Comprenant leur Application à la Reconnaissance des Couches, pls. 237–489. Published by the authors, Paris. Crossref
Reed, F.R.C. 1929. Faunas Trássicas do Brasil. Serviço Geológico e Mineralógico do Brasil, Monographia 1: 4–87.
Reed, F.R.C. 1932. Some new Triassic fossils from Brazil. Annals and Magazine of Natural History 10: 479–487. Crossref
Reed, F.R.C. 1935. Some Triassic lamellibranchs from Brazil and Paraguay. Geological Magazine 72: 33–42. Crossref
Ridewood, W.G. 1903. On the structure of the gills of the Lamellibranchia. Royal Society of London, Philosophical Transactions (series B) Biological Sciences 195: 147–284. Crossref
Rohn, R. 1994. Evolução ambiental da Bacia do Paraná durante o Neopermiano no leste de Santa Catarina e do Paraná. 480 pp. Ph.D. Dissertation, Universidade de São Paulo, São Paulo. Available at https://10.11606/T.44.1995.tde-22062015-145002 Crossref
Rohn, R. and Rösler, O. 2000. Middle to upper Permian phytostratigraphy of the eastern Paraná Basin. Revista da Universidade de Guarulhos-Geociências 5 (Número Especial): 69–73.
Runnegar, B. 1965. The bivalves Megadesmus Sowerby and Astartila Dana from the Permian of eastern Australia. Journal of the Geological Society of Australia, 12: 227–252. Crossref
Runnegar, B. 1974. Evolutionary history of the bivalve Subclass Anomalodesmata. Journal of Paleontology 48: 904–939.
Runnegar, B. and Newell, N.D. 1971. Caspian-like relict molluscan fauna in the South American Permian. Bulletin of the American Museum of Natural History 146: 1–66.
Scarlato, O. and Starobogatov, Y.I. 1979. General evolutionary patterns and the system of the Class Bivalvia [in Russian]. Transactions of the Zoological Institute 80: 5–38.
Sepkoski, J.J. 2002. A compendium of fossil marine genera. Bulletins of American Paleontology 353: 560.
Simões, M.G. and Anelli, L.E. 1995. Runnegariella, um novo gênero de Megadesmidae (Pelecypoda) da Formação Corumbataí (Neopermiano), Bacia do Paraná, Brasil. Geociências 14: 161–173.
Simões, M.G., Anelli, L.E., and David, J.M. 2010. Othonella araguaiana Mendes (Bivalvia, Megadesmidae), Corumbataí Formation (Upper Permian), eastern margin of the Paraná Basin: systematical, evolutionary and biostratigraphical significances. Geologia USP, Série Científica 10: 45–55. Crossref
Simões, M.G., David, J.M., Anelli, L.E., Klein, C., Matos, S.A., Guerrini, V.B., and Warren, L.V. 2017. The Permian Tiaraju bivalve assemblage, Passa Dois Group, southern Brazil: biostratigraphic and paleobiogeographic significance. Brazilian Journal of Geology 47: 209–224. Crossref
Simões, M.G., Marques, A.C., Mello, L.H.C., and Anelli, L.E. 1997. Phylogenetic analysis of the genera of the extinct Family Megadesmidae (Pelecypoda, Anomalodesmata), with remarks on its paleoecology and taxonomy. Journal of Comparative Biology 2: 75–90.
Simões, M.G., Matos, S.A., Anelli, L.E., Rohn, R., Warren, L.V., and David, J.M. 2015. A new Permian bivalve-dominated assemblage in the Rio do Rasto Formation, Paraná Basin, Brazil: faunal turnover driven by regional-scale environmental changes in a vast epeiric sea. Journal of South American Earth Sciences 64: 14–26. Crossref
Simões, M.G., Mello, L.H., Torello, F.F., and Ghilardi, R.P. 2000. Tambaquyra gen. n. (Bivalvia, Anomalodesmata), Formação Serra Alta (Neopermiano), Grupo Passa Dois, Bacia do Paraná, Brasil. Revista UNG, Serie Geociências 5 (6): 11–19.
Simões, M.G., Rocha-Campos, A.C., and Anelli, L.E. 1998. Paleoecology and evolution of Permian pelecypod assemblages (Paraná Basin) from Brazil. In: P.A. Johnston, and J. W. Haggart (eds.), Bivalves—An Eon of Evolution: Paleobiological Studies Honoring Norman D. Newell, 443–452. University of Calgary Press, Calgary.
Solem, A. and Yochelson, E.L. 1979. North American Paleozoic Land Snails, with a summary of other Paleozoic Nonmarine Snails. Geological Survey Professional Paper 1072: 1–67. Crossref
Tasch, P. 1982. Conchostracan dispersal between South America, Africa and Antarctica. Antarctic Journal 17 (5): 45–46.
Vokes, H.E. 1967. Genera of the Bivalvia: A systematic and bibliographic catalogue. Bulletins of American Paleontology 51: 111–393.
Wesselingh, F.P. 2007. Long-lived lake molluscs as island faunas: a bivalve perspective. In: W. Remena (ed.), Biogeography, Time and Place: Distributions, Barriers and Islands, 275–314. Springer, Dordrecht. Crossref
Zalán, P.V., Wolf, S., Conceição, J.C., Astolfí, A.M., Vieira, I.S., Appi, V.T., Zanotto, O.A., and Marques, A. 1991. Tectonics and sedimentation of the Paraná Basin. In: H.H.G.J. Ulbrich and A.C. Rocha-Campos (eds.), Proceedings of the Gondwana Seven Meeting, São Paulo, 1991, 83–117. Instituto de Geociências da Universidade de São Paulo, São Paulo.
Zalán, P.V., Wolff, S., Conceição, J.C.J., Marques, A., Astolfi, M.A.M., Vieira, I.S., Appi, V.T., and Zanotto, O.A. 1990. Bacia do Paraná. In: G.P. Raja Gabiglia and E.J. Milani (eds.), Origem e Evolução de Bacias Sedimentares, 681–708. Petrobras, Rio de Janeiro.
Acta Palaeontol. Pol. 65 (2): 291–303, 2020
https://doi.org/10.4202/app.00665.2019