
Dipnoan from the Upper Triassic of East Greenland and remarks about palaeobiogeography of Ptychoceratodus
WOJCIECH PAWLAK, MATEUSZ TAŁANDA, TOMASZ SULEJ, and GRZEGORZ NIEDŹWIEDZKI
Pawlak, W., Tałanda, M., Sulej, T., and Niedźwiedzki, G. 2020. Dipnoan from the Upper Triassic of East Greenland and remarks about palaeobiogeography of Ptychoceratodus. Acta Palaeontologica Polonica 65 (3): 561–574.
Here we present a description of the dipnoan remains collected from the middle to upper Norian (Upper Triassic) of Jameson Land, East Greenland. The specimens consist of isolated tooth plates and skull bones of Ptychoceratodus, the most complete Late Triassic dipnoan material from Greenland. This genus is reported for the first time from the Upper Triassic of Greenland. The studied material belongs to Ptychoceratodus rectangulus previously known from the middle–upper Norian of Germany. It fills the biogeographical gap between the records of the Germanic and the Jameson Land basins. A reconstruction of the skull roof is provided, based on isolated bones collected from the same bone-bed. Their good preservation enables recognition of the sensory line pits, arranged similarly as in the extant Protopterus, suggesting a comparable mode of life. This finding has implications for our understanding of the disparity in Ptychoceratodus dipnoans, as well as the morphology between closely related dipnoans of the Late Triassic ecosystems.
Key words: Dipnoi, Ptychoceratodus, Triassic, Norian, Greenland, Carlsberg Fjord Beds.
Wojciech Pawlak [wojciech.pawlak@student.uw.edu.pl] and Mateusz Tałanda [m.talanda@biol.uw.edu.pl], Department of Palaeobiology and Evolution, Faculty of Biology, Biological and Chemical Research Centre, University of Warsaw, Żwirki i Wigury 101, 02-089 Warsaw, Poland.
Tomasz Sulej [sulej@twarda.pan.pl], Institute of Paleobiology, Polish Academy of Sciences, Twarda 51/55, 00-818 Warsaw, Poland.
Grzegorz Niedźwiedzki [grzegorz.niedzwiedzki@ebc.uu.s], Department of Organismal Biology, Evolutionary Biology Center, Uppsala University, Norbyvägen 18A, 752 36 Uppsala, Sweden.
Received 16 September 2019, accepted 24 January 2020, available online 15 May 2020.
Copyright © 2020 W. Pawlak et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The study of Late Triassic dipnoans (lungfish) has recently undergone renewed interest, improving our understanding of the taxonomy, diversity and palaeobiogeography of this important component of Triassic freshwater and marginal marine ecosystems (e.g., Skrzycki 2015, 2016; Agnolín et al. 2017, 2018; Skrzycki et al. 2018; Bhat and Ray 2020). The dipnoans are adapted to breathing with both gills and lungs (Wright 1974; Holmes 2008). They are characterized by crushing dental plates instead of true teeth. Despite this specialization to durophagy, the extant lungfishes are omnivorous (Kemp 1986; Greenwood 1986). Dipnoans are an old sarcopterygian fish lineage known from the Early Devonian to recent. Their massive and resistant to erosion dental plates are common remains in the Triassic fossil record.
The fossil record of the post-Devonian
lungfishes is biased in that bone elements from cranial or postcranial
skeleton are relatively rare, mainly preserved as isolated or fragmented
elements, and the most common remains of these fishes are their dental
plates. Only a few post-Palaeozoic species are known from well
preserved cranial or postcranial skeletal remains (Kemp
1998). This residual fossil record gives remarkably incomplete
knowledge about lungfish palaeoecology and evolution. Consequently,
previous phylogenetic analyses (Miles 1977; Marshall
1986b; Cavin et al. 2007; Kemp
2017) failed to offer a robust evolutionary scenario explaining
the major changes within the post-Palaeozoic dipnoans.
The Triassic was a golden age for dipnoans, at least when it comes to
the number of genera and species (Martin 1982; Marshall
1986a; Schultze 2004). Such a diversity is
probably related to a wide global distribution of warm and semi-dry
continental climate probably preferred by these fishes, similarly to
their Recent representatives. Comprehension of the Triassic evolutionary
radiation of dipnoans is crucial for the understanding of their later
Mesozoic and younger evolutionary history. The Triassic dipnoans are
known from freshwater and marginal marine (brackish) environments. There
are two genera, which are particularly common nearly worldwide: Arganodus
and Ptychoceratodus (Skrzycki
et al. 2018). Only two species of Ptychoceratodus
are known from a relatively complete cranial skeleton: Ptychoceratodus
serratus from the late Ladinian Erfurt Formation in Germany (Schultze 1981) and Ptychoceratodus
philippsi from the Cynognathus
Zone of the Bergersdorp Formation in the Karoo Basin, dated as late
Early Triassic–early Middle Triassic (Kemp 1996).
In East Greenland, fossiliferous Upper Triassic of Jameson Land Basin, such as the Malmros Klint Member or Ørsted Dal Member, are known for rich faunal assemblages (Jenkins et al. 1994; Clemmensen et al. 1998, 2016; Sulej et al. 2014). However, the remains of dipnoans are still poorly known from the Late Triassic record of Greenland. The oldest record of lungfishes from Greenland comes from the Middle and Late Devonian (Lehman 1959; Bendix-Almgreen 1976; Blom et al. 2007; Clack et al. 2018). In younger Carboniferous or Permian deposits of Greenland, dipnoan remains are unknown, but some remains have been collected from the Upper Triassic Fleming Fjord Formation (Jenkins et al. 1994; Clemmensen et al. 1998, 2016). Recently, dental plates and jaw bones belonging to a new species of Ceratodus (C. tunuensis) have been described from the upper Carlsberg Fjord Beds (Agnolín et al. 2018).
In this paper we present a detailed description of the newly collected ptychoceratodontid skull material and highlight its palaeoecological and palaeobiogeographic significances. We compare it with Ceratodus tunuensis reported from the same lithostratigraphical unit (Agnolín et al. 2018). The new material represents fossils which are much better preserved than the other dipnoan remains collected so far from the Ørsted Dal Member. For this reason, the study has required a brief re-study of the isolated elements described by Agnolín et al. (2018) and a comparison with other previously described ptychoceratodontid fossils. The Fleming Fjord Formation also yielded some aestivation burrows, which carry information about the Late Triassic climate of north Pangea and broadens our knowledge about the distribution of the aestivation behavior among the early Mesozoic dipnoans. These trace fossils will be a subject of an additional study.
Institutional abbreviations.—BRSUG, University of Bristol, Department of Geology, UK; KNK, Greenland National Museum & Archives, Nuuk, Greenland; NHMD, Natural History Museum of Denmark, Copenhagen, Denmark; SMNS, Staatliche Museum für Naturkunde, Stuttgart, Germany.
Other abbreviations.—BC1, distance between the mediolingual joint and the tip of the first ridge; C1C2, distance between the tips of the first and second ridges; BCp, the distance between mediolingual joint and the tip of the last ridge.
Geological setting
The Upper Triassic succession of the Jameson Land is composed of about 200–300 m thick continental sediments of the Fleming Fjord Formation (Clemmensen et al. 1998) represented mainly by red or greenish mudstone, siltstone, and sandstone beds. The formation is divided into three members: the lowermost Edderfugledal Member, the middle Malmros Klint Member and the uppermost Ørsted Dal Member (Fig. 1). Integrated magnetostratigraphical, cyclostratigraphical, and some biostratigraphical studies revealed that the Fleming Fjord Formation age is late Carnian–early Rhaetian (Clemmensen et al. 1998), Norian–Rhaetian (Clemmensen et al. 2016), or Norian–early Rhaetian (Andrews et al. 2014). Very rare invertebrate body fossils (e.g., conchostracans, bivalves), composition of the tetrapod faunas/ichnofaunas, and land-derived palynomorphs suggest that the Malmros Klint Member is of late Carnian–middle Norian age, and that the Ørsted Dal Member is of middle Norian–early Rhaetian age (Clemmensen 1980; Clemmensen et al. 1998, 2016; Sulej et al. 2014). A conspicuous feature of these deposits are sedimentation cycles reflecting climate changes, which are developed slightly differentially in every lithostratigraphical subunit (Clemmensen et al. 1998).
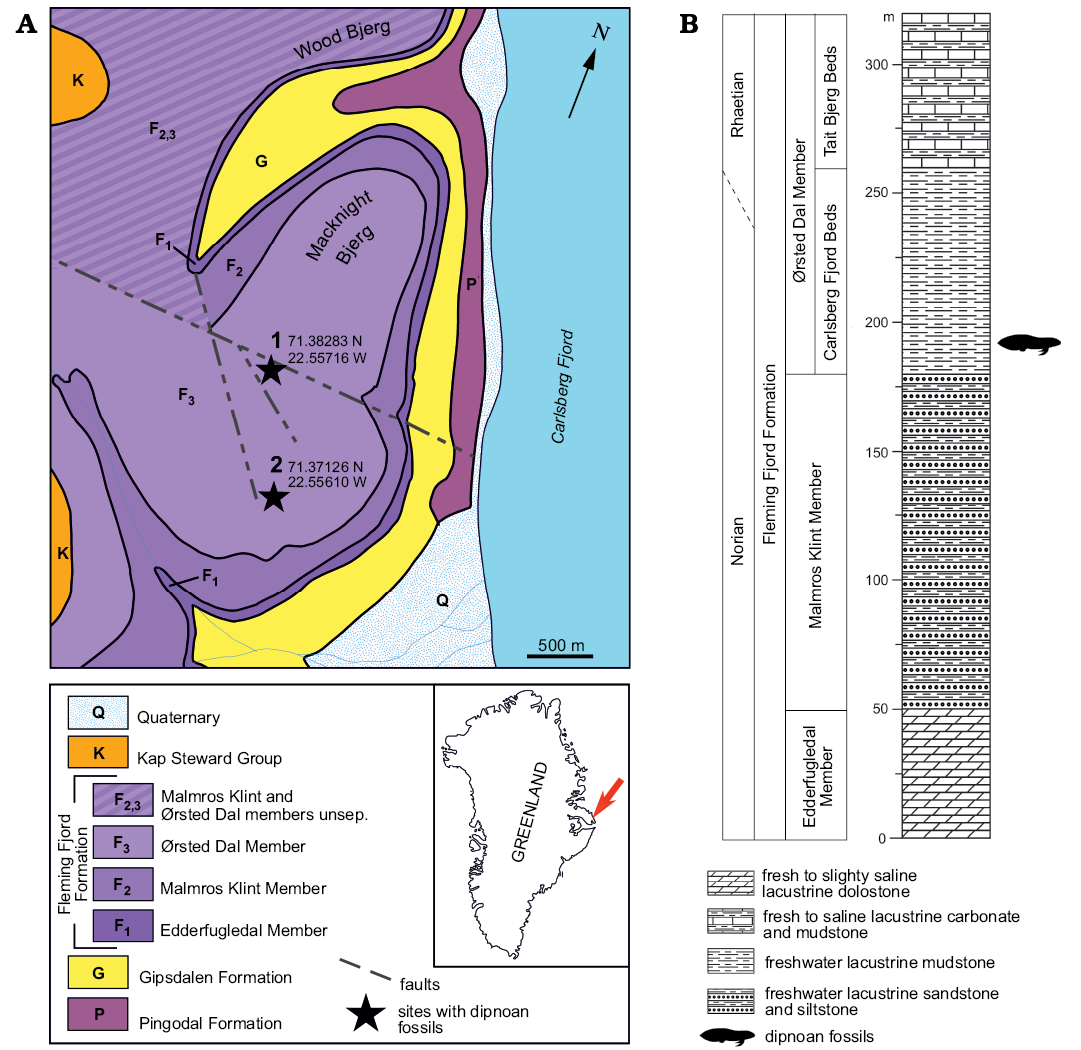
Fig. 1. Study area map (A) and stratigraphic column (B) with marked outcrops and age of the stratigraphic position of examined material. Based on Bengaard and Henriksen (1984) and Clemmensen et al. (1998).
The fossiliferous lower part of the Ørsted Dal Member, named the Carlsberg Fjord Beds, is composed of intraformational conglomerate, massive mudstone, mudstone with wave ripple cross-lamination, and sandstone with cross-bedding. Desiccation cracks are common in this interval within both the mudstone and the sandstone beds but there are no signs of pedogenesis (Clemmensen et al. 1998). The Ørsted Dal Member contains also two other lithostratigraphic subunits. In the western and central part of the Basin, the Carlsberg Fjord Beds are replaced by fluvial sediments of the Bjergkronerne Beds, and both units are overlaid by carbonate-rich mudstone, dolostone, and clay-rich mudstone of the Tait Bjerg Beds (Jenkins et al. 1994).
Both the Malmros Klint Member and Ørsted
Dal Member are abundant in skeletal and trace fossils of terrestrial and
aquatic vertebrates (Jenkins et al. 1994; Clemmensen
et al. 1998; Sulej et al. 2014; Clemmensen
et al. 2016). The existing outcrops indicate that the Fleming
Fjord Lakeland had about 3000 km2,
therefore it was a large but predominantly shallow reservoir with
frequent desiccation episodes (Clemmensen et al. 1998).
The periodic shallow lake conditions favored bone accumulation, thus the
Fleming Fjord Formation infill of the Jameson Land is an interesting
taphonomical window of the Late Triassic fauna.
These fossiliferous deposits have been searched for vertebrate remains
since the late 1980s (e.g., Jenkins et al. 1994; Clemmensen et al. 1998, 2016; Shapiro
and Jenkins 2001; Milan et al. 2004; Sulej
et al. 2014). The vertebrate fauna of the Fleming Fjord Formation
is relatively abundant and quite diverse, and includes actinopterygian
and dipnoan fishes, temnospondyls, sauropodomorph and theropod
dinosaurs, basal turtles, phytosaurs, aetosaurs, pterosaurs, cynodonts,
and mammaliaforms (Jenkins et al. 1994, 1997,
2001, 2008; Sulej et
al. 2014; Clemmensen et al. 2016; Niedźwiedzki
and Sulej 2017; Marzola et al. 2017; Agnolín
et al. 2018). Numerous tetrapod tracks have been found in several
horizons inside the Malmros Klint and Ørsted Dal members (e.g., Jenkins
et al. 1994; Sulej et al. 2014; Klein
et al. 2016; Lallensack et al. 2017; Niedźwiedzki
2017).
Material and methods
The examined specimens were collected during the Polish-Danish expedition to Greenland in 2014 (Sulej et al. 2014). Eleven skeletal elements and one imprint were found, which belonged to different individuals of various sizes. All specimens are disarticulated but three-dimensionally preserved without any sings of compaction and additional deformations. Skull roof bones have a characteristic radial pattern of points and lines diverging from the ossification center. Specimens KNK 4154–4161, 4163–4165 were found in the locality 1 (71.38283 N; 22.55716 W), and KNK 4162 in the locality 2 (71.37126 N; 22.55610 W). Both localities have very similar lithostratigraphic position: section exposed at locality 1 represents transitional layers between the Malmros Klint and the Ørsted Dal members, it is a new fossiliferous spot with poorly exposed bone-bed enriched in remains of freshwater fauna; section exposed at locality 2 shows lowermost part of the Carlsberg Fjord Beds and is rich in disarticulated elements of temnospondyls (Jenkins et al. 1994; Marzola et al. 2017) (Fig. 1). In previous studies this site was named Macknight Bjerg Quarry (see Jenkins et al. 1994). There are no signs of postmortal mechanical wearing on the studied elements. Surfaces, cusps, or crests are well preserved. Some observed changes on the bone surfaces are the effects of recent weathering. Almost all of the skeletal elements were enveloped by a slightly heterogeneous mudstone matrix, which was subjected to dissolving and softening in 10% solution of the acetic acid. Some of the specimens were prepared mechanically when the chemical method did not work.
Collected specimens are catalogued in both NHMD and KNK collections. Correspondence between KNK and NHMD numbers is as follows: KNK 4154 = NHMD 658404, KNK 4155 = NHMD 658405, KNK 4156 = NHMD 657408, KNK 4157 = NHMD 657409, KNK 4158 = NHMD 657410, KNK 4159 = NHMD 657411, KNK 4160 = NHMD 657412, KNK 4161 = NHMD 657413, KNK 4162 = NHMD 657414, KNK 4163 = NHMD 657415, KNK 4164 = NHMD 657416, KNK 4165 = NHMD 657417. In thie maintext of this article we use only the KNK numbers.
Several terminologies of lungfish skull bones exist in the literature. The reason is unclear homology between skull roof bones of lungfish and other vertebrates. In this paper we use nomenclature of bones and sensory lines after Kemp (1998).
Dental plates morphometry was based on the method proposed by Vorobyeva and Minikh (1968). Measured dimensions are explained on the Fig. 2. Biometric method based on the angles between ridges and sections between characteristic points was criticized by Kemp (1997), however, it is used in this paper only as a support (Kemp 1997; Skrzycki 2015). All the measurements of the specimens originally described herein were taken directly from original fossils. Measurements of Ceratodus tunuensis are based on the figures presented in Agnolín et al. (2018). All measurements collected in this study are presented in Table 1. This set of biometric measurements of dental plates was confronted with the measurements from Skrzycki (2015).
Principal Component Analysis was performed in PAST 3.23 software. To normalize the data we transformed the measurements by dividing them by standard deviation. We excluded incomplete specimens. The analysis was performed in order to visualize variation occurring in dental plates of different Ptychoceratodus species. Lower and upper dental plates were analyzed separately.
Most of the specimens were imaged using Nikon Digital Camera D5300. KNK 4158 was imaged using Keyence Digital Microscope VHX-900F. Surface of the KNK 4165 is figured as untextured high-detailed surface scan. We used Shining 3D EinScan Pro 2X 3D scanner fixed on a tripod with EXScan Pro 3.2.0.2 software.
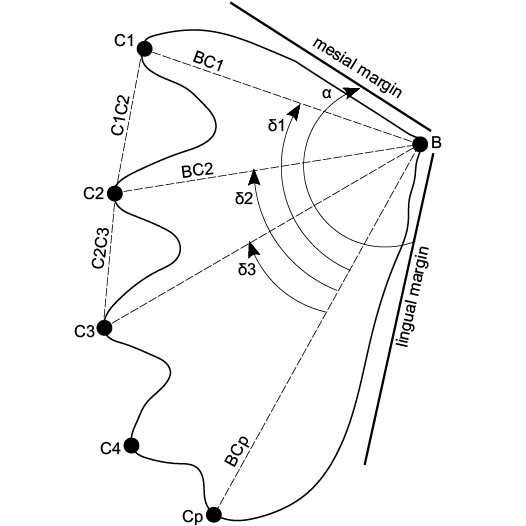
Fig. 2. Scheme of the measured dimensions of dental plates of Triassic dipnoans. Abbreviations: B, mediolingual junction; C1, C2, C3, C4, ends of corresponding ridges; Cp, end of the last ridge; α, angle between medial and lingual margin; ð1, ð2, ð3, angles between last ridge and corresponding ridges; BCp, BC1, BC2, C1C2, C2C3, C1Cp, C2Cp, distances between corresponding points. Based on Frederickson and Cifelli (2017).
Table 1. Biometric measurements (in mm; α, δ1, δ2, δ3 in degrees) of the chosen dental plates of Ptychoceratodus rectangulus from Jameson Land Basin (East Greenland).
|
Specimen |
BCp |
α |
δ1 |
δ2 |
δ3 |
BC1 |
BC2 |
C1C2 |
C2C3 |
C1Cp |
C2Cp |
|
KNK 4154 |
31.0 |
105.5 |
92.2 |
51.5 |
29.6 |
22.4 |
24.1 |
10.8 |
10.0 |
34.4 |
24.8 |
|
KNK 4155 |
24.9 |
109.1 |
79.5 |
46.4 |
27.0 |
14.4 |
16.1 |
8.8 |
7.1 |
26.4 |
18.1 |
|
KNK 4156 |
24.9 |
103.4 |
67.5 |
35.1 |
14.3 |
16.1 |
17.2 |
9.3 |
8.00 |
23.2 |
14.0 |
|
KNK 4157 |
15.1 |
– |
– |
– |
30.3 |
– |
– |
– |
– |
– |
– |
|
NHMD 115910 |
50.3 |
110.7 |
80.8 |
49.4 |
27.9 |
– |
36.5 |
– |
14.9 |
– |
38.4 |
Results
Examined dental plates have typical ptychoceratodont morphology. They are high crowned, with less than 7 ridges. Ridges originate anteriorly, which results in their fan-like arrangement. Crests meet in the medial margin. Number of the ridges in the examined specimens (5 in the upper and 4 in the lower dental plates) is reduced compared to Ptychoceratodus serratus (6 in the upper and 5 in the lower dental plates). Particularly distinctive feature is the flattening of the last ridge, present in Ptychoceratodus concinnus and Ptychoceratodus rectangulus (Linck 1936). However, dental plates of P. concinnus have a significantly different geometry (wider α angle and more robust first ridge expressed by generally higher BC1/BCp ratio in the upper and higher C1C2/BCp ratio in the lower dental plates) (see Skrzycki 2015; Fig. 3). In scatter plots presented on Fig. 3 the upper dental plates described herein group in close neighborhood of P. rectangulus dental plates, whereas the lower dental plates follow conspicuous ontogenetic trend (Fig. 3). However, the sample in both cases is very small.
Systematic palaeontology
Class Osteichthyes Huxley, 1880
Subclass Sarcopterygii Romer, 1955
Order Dipnoi Müller, 1845
Family incertae sedis
Genus Ptychoceratodus Jaekel, 1926
Type species: Ptychoceratodus serratus (Agassiz, 1838); Aargau, Switzerland; Keupersandstein.
Remarks.—Ptychoceratodus is not assigned to any family because of disagreement between researchers about taxonomy of Mesozoic lungfishes. Ptychoceratodontidae diagnosed on dental plates is commonly used by various authors (Kemp 1998; Martin et al. 1999; Richter and Toledo 2008). However, objections to taxonomy based on the dental plates are also present (Schultze 1992). Taxonomic difficulties are well outlined in Skrzycki (2015). We consider cranial features as a better taxonomic tool, however, because most dipnoan findings are isolated dental plates, there is no other and better possibility to distinguish the species or genera.
Ptychoceratodus rectangulus (Linck, 1936)
Figs. 3–5, 7.
1936 Ceratodus rectangulus sp. nov.; Linck 1936: 1–23, pl. 4: 1–5, pl. 5: 1–4.
1938 Ceratodus rectangulus Linck, 1936; Linck 1938: 1–11, text-figs. 1–3, 5.
1963 Ceratodus rectangulus Linck, 1936; Linck 1963: 1–9, pl. 20, text-fig. 1.
1981 Ceratodus rectangulus Linck, 1936; Martin 1981: 11, text-fig. 5a.
1981 Ptychoceratodus rectangulus (Linck, 1936); Schultze 1981: 21, text-fig. 13.
1998 Ptychoceratodus rectangulus (Linck, 1936); Cuny et al. 1998:7, text-fig. 5j.
2018 Ceratodus tunuensis sp. nov.; Agnolín et al. 2018: 1–6, text-figs. 3–5.
Syntypes: Upper dental plates: SMNS 17963–17965, 17970; lower dental plates: SMNS 17966, 17967, 17971, 17973.
Type locality: Ochsenbach region, Baden-Württemberg, Germany.
Type horizon: Löwenstein Formation (Stubensandstein), middle Keuper, middle–upper Norian.
Material.—Right upper dental plate with attached almost complete pterygopalatine bone: KNK 4154; right upper dental plate: KNK 4155; right lower dental plate: KNK 4156; right lower dental plate: KNK 4157; right KLMN+3 bone: KNK 4158; right XYZ bone: KNK 4159; right operculum: KNK 4160; incomplete IJ? bone: KNK 4161; E bone: KNK 4162; left XYZ bone: KNK 4163; incomplete dental plate: KNK 4164; KLMN+3 bone imprint: KNK 4165. All from middle–upper Norian, locality 1, despite KNK 4162 from locality 2: Macknight Bjerg, Greenland. Upper dental plates: NHMD 115910–115912; lower dental plates: NHMD 115913–115916; prearticular bone: NHMD 115917. All from upper Norian, Lepidopteriselv, Greenland. Unidentified dental plates: BRSUG 26352, from Norian, Grozon, France. KLMN+3 bones: SMNS 19705, 19706; XYZ bone: SMNS 17971; incomplete IJ? bones: SMNS 19707, E? bone: SMNS 19708. All from middle–upper Norian, Ochsenbach region, Germany.
Diagnosis.—Dental plates high-crowned, lingual margin about two times longer than the mesial. Angle between the medial and lingual margin about 90° or more. Upper dental plates with five ridges, the first ridge initially curved anteriorly anterolaterally, then slightly twisted posteriorly, the last ridge flattened. Lower dental plates with four ridges, the last ridge flattened on the lingual side. Prearticular sulcus is double (like in other ptychoceratodontids except P. serratus), but the posterior sulcus is deeper than the anterior. Pterygopalatine ascending process above the second ridge, slightly shifted toward the third ridge, similarly to P. serratus. Thick descending process of KLMN+3 bone with wide basement, confluent with relatively high and wide anterior crest.
Description.—Skull roof: The largest specimen of the Greenland material is KNK 4158, interpreted as an incomplete right KLMN+3 bone because of characteristic lateral line grooves on the dorsal side, and descending process and crests on the ventral side (Fig. 4A). The bone is 36 mm long and 44 mm wide. The posterior part is damaged, whereas other edges are natural. The lateral part of the bone is clearly downturned. The lateral edge is concave. Posteriorly, the bone expands laterally and forms conspicuous lateral process, which gives KNK 4158 trapeze-like outline from dorsal view. The dorsal surface of the bone is relatively smooth, whereas the lateral and anterior margins display cancellous structure. The supraorbital sensory canal groove angle is not preserved. The anterior part of the groove is shallow, whereas the lateral part is deep and widens toward the bone’s margin. The indentation for the medial process of the XYZ bone is visible posterior to the lateral process. The ventral surface is predominantly cancellous, being smooth only posteriorly. Two crests are visible in the central region of the bone: anterior and posterior (Fig. 4A3, A4). The posterior crest is directed straight to the lateral margin of the bone, and forms a slight bulge, more conspicuous laterally. The anterior crest is higher and slenderer than the posterior one, and directed anteriorly. The angle between the crests is roughly right. The anterior crest passes smoothly to the descending process of the KLMN+3 bone. The process is partially broken. However, it is clearly leaned towards the medial side. The groove for the ascending process of pterygoid appears on the medial side of the descending process base. The protrusion for the E bone is visible on the medial margin of the KLMN+3 bone. The morphology of the posterior part of the KLMN+3 bone is visible on the imprint of the same individual KNK 4165 (Fig. 5A). The posterior margin has an indentation for the J bone, however, there are no visible differences between the indentation and rest of the posterior margin. It suggests that the I bone and the J bone were merged into a single IJ bone. Three shallow grooves begin from the inclined openings on the dorsal surface of both specimens. They probably represent blood vessel ducts. Shape of the complete KLMN+3 was roughly rectangular, which differs it from anteroposteriorly elongated KLMN+3 of Arganodus atlantis (see Kemp 1998). Two complete XYZ bones were found. KNK 4159 is 52 mm long and belonged to a large fish, whereas KNK 4163 is only 29 mm and represents evidently smaller individual. KNK 4159 is a flat bone, geometrically composed of three distinct regions: a flat posterior process, slightly convex but still relatively flat body, and curved articular process (Fig. 3B). The dorsal margin of XYZ has elongated protrusion for the KLMN+3 bone along the edge. The outer surface of the articular process differs from the other bone parts by its cancellous character. Radial marks cover the outer surface of the bone body and the posterior process, converging at the center of ossification. A temporal sensory line groove goes from the posterior to the anterior margin and is shallowing posteriorly. The split of the temporal line is absent. Thus, probably it was located among soft tissues. Five conspicuous elliptical openings appear in the equal distances inside the temporal line groove. The dorsal and anterodorsal margins articulated the XYZ with the KLMN+3 and IJ bones. The anterodorsal suture is relatively short, what is different from YZ bone in the skull of Ceratodus sturii (see Kemp 1998). The posteriormost part of the dorsal margin and the posterior process were joined with the IJ bone. The articular process, which was joined with the lower jaw is clearly thicker than other parts of the XYZ bone. The specimen KNK 4163 is slightly incomplete. Morphology of the bone surface is similar to the larger KNK 4159, but the distinctive feature of this specimen is bone geometry. Whole KNK 4163 is relatively flat, there is no curvature between the bone body and the articular process, unlike in KNK 4159. The delta-shaped bone (KNK 4162) is surrounded by hard sediment impossible to remove without damaging the specimen, thus only one side of the bone is visible (Fig. 5C). The bone is asymmetrical. A conspicuous convex outer side of the bone is probably its dorsal surface. There are visible openings for the blood vessels and a damaged, anterioposteriorly oriented crest along the entire bone. The inner surface is only partially exposed. There is visible an indentation matching the KLMN+3 protrusions. This is why KNK 4162 is interpreted as the E bone.
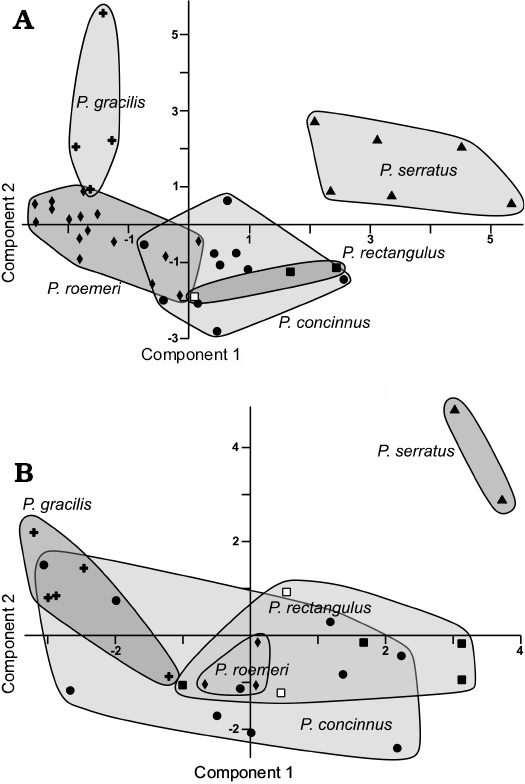
Fig. 3. Principal Component Analysis of species of Ptychoceratodus dental plates. A. Lower dental plates. B. Upper tooth plates. In the analysis we used α, δ1, δ2, δ3, BCp, C1C2, and C1Cp measurements sensu Vorobyeva and Minikh (1968). White rectangles mean specimens from Macknight Bjerg locality (East Greenland) described herein.

Fig. 4. Skull roof bones of dipnoan Ptychoceratodus rectangulus (Linck, 1936) from the Carlsberg Fjord Beds, Fleming Fjord Formation (Upper Triassic), Macknight Bjerg, East Greenland. A. KNK 4158, KLMN+3 bone in dorsal (A1, A2) and ventral (A3, A4) views. B. KNK 4159, XYZ bone in lateral (B1, B2) and medial (B3, B4) views. Arrows indicate sutures for corresponding bones.

Fig. 5. Skull roof bones and opercle of dipnoan Ptychoceratodus rectangulus (Linck, 1936) from the Carlsberg Fjord Beds, Fleming Fjord Formation (Upper Triassic), Macknight Bjerg, East Greenland. A. KNK 4165, KLMN+3 bone imprint, in dorsal view. B. KNK 4160, opercle in lateral (B1) and medial (B2) views. C. KNK 4162, E bone in dorsal view. Arrows indicate sutures for corresponding bones.
Operculum: The operculum (KNK 4160) is small, flattening posteriorly (Fig. 5B). Only the ventral half of the operculum is preserved. The posterior edge is very thin. Outer surface of KNK 4160 is significantly eroded, but the original surface was probably smooth. In contrast, the inner surface is cancellous. A conspicuous articulation with the XYZ bone is visible on the anterior margin. The cross section revealed a blood vessel canal that goes along the anterior margin.
Jaw bones and dental plates: One pterygopalatine bone was found with attached upper dental plate. KNK 4154 is an almost complete pterygopalatine missing only the ascending process (Fig. 6A). Surface of the pterygopalatine is fibrous above the dental plate, and smooth in the posterior part. The basis of the pterygopalatine process is located between the second and third ridge of the upper dental plate. Prearticular bones are represented only by fragments directly attached to the lower dental plates (KNK 4156, 4157). The ventral surface of the KNK 4156 prearticular bone is fibrous and has a distinctive double sulcus (Fig. 6B3, B4). The anterior sulcus is isometric and significantly smaller and shallower than the posterior, which is posteriorly elongated and deeper. Five dental plates were found. KNK 4154 and KNK 4155 are palatal plates of moderate size (31.3 and 22.1 mm long, respectively), whereas KNK 4156 and KNK 4157 are smaller prearticular plates (23.9 and 15.1 mm long, respectively). The dental plates are slightly elongated and high crowned with fan-like pattern of ridges (Fig. 6A–C). Crests of the three anterior ridges are meeting at the mediolingual junction. The specific feature of the dental plates is flattening of the last ridges.
The upper dental plates have five ridges, and slightly convex occlusal surfaces in the posterior part. The last ridges are almost completely flat. KNK 4155 has conspicuous cusps on the three last ridges. The fifth ridge of the smaller specimen (KNK4154) is more confluent with the fourth ridge than in the bigger (KNK4155).
The lower dental plates have four ridges and the occlusal surface is about flat. The last ridges of lower dental plates are composed of medial crest and flattened lingual part. Cusps are visible on all preserved ridges. KNK 4157 has slightly separated fifth ridge with low crest (Fig. 6C).
First crest is differently oriented in the upper and lower plates. In the upper plates it follows mesial margin, but in the lower it is in distance from the margin (Fig. 6B1, B2, C). This pattern enables occlusion. KNK 4164 represents an incomplete dental plate of a large individual (Fig. 6D). Only first two ridges are preserved. The surface is heavily worn and the ratio of thickness to length is relatively small. It is difficult to establish if it is a palatal or prearticular dental plate because no identifiable bone part is attached to the specimen.
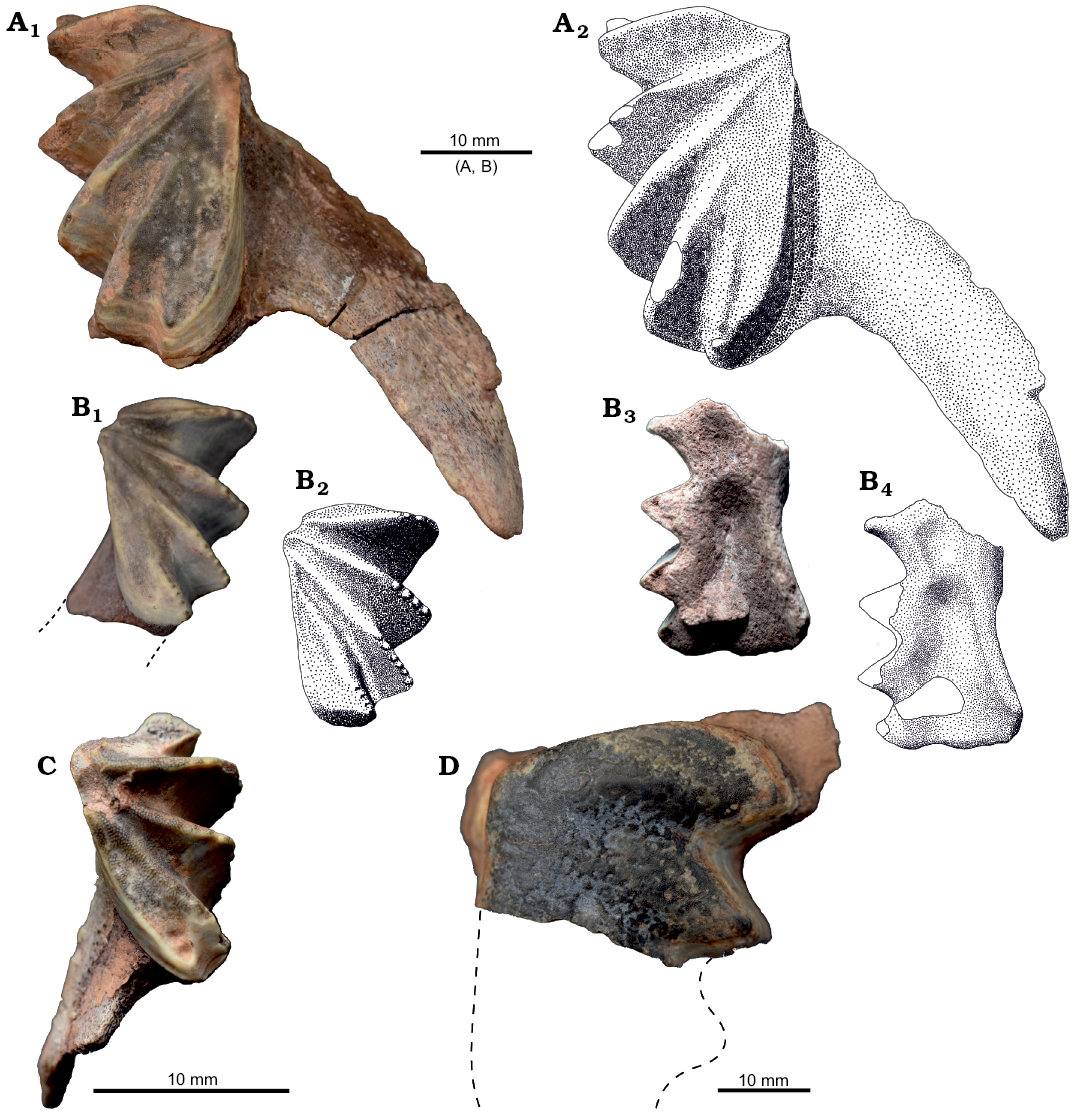
Fig. 6. Dental plates and jaw bones of dipnoan Ptychoceratodus rectangulus (Linck, 1936) from the Carlsberg Fjord Beds, Fleming Fjord Formation (Upper Triassic), Macknight Bjerg, East Greenland. A. KNK 4154, upper dental plate with pterygopalatine bone in occlusal view. B. KNK 4156, lower dental plate in occlusal (B1, B2) and ventral (B3, B4) views. C. KNK 4157, lower dental plate with fragmentary prearticular bone in occlusal view. D. KNK 4164, incomplete dental plate in occlusal view.
Remarks.—The first description of Ptychoceratodus rectangulus in Linck (1936) was based on eight poorly preserved dental plates from the middle Löwenstein Formation of Ochsenbach (middle Norian, Germany). It was initially assigned to genus Ceratodus, until Schultze (1981) transferred it to the genus Ptychoceratodus. Skull bones were also found in the same formation. Linck (1936, 1938, 1963) made biometrical studies, which were repeated in Skrzycki’s (2015) analysis according to current standards. Full morphological description or biometric measurements were not possible for some of the dental plates due to preservation (Skrzycki 2015). A holotype was never designed for P. rectangulus. Cuny et al. (1998) assigned to this species one minute dental plate from the Norian Marnes de Châlins Formation in eastern France. The tooth plates described here are not as numerous as the material from Ochsenbach but are better preserved, what makes more detailed morphological or histological studies possible.
The dental plates described by Agnolín et al. (2018) as Ceratodus tunuensis came also from the Carlsberg Fjord Beds. They were classified as ceratodontid due to robust and short ridges having broad occlusal surface with rounded outline. However, all described specimens are worn and incomplete (except the complete but worn NHMD 115912). Ptychoceratodus rectangulus specimens from Ochsenbach show similar degree of erosion, however, they are mostly complete. High degree of incompleteness of the C. tunuensis dental plates may indicate that they were subject of transport and their wear is partially result of post-mortem erosion. This makes the proper evaluation of features like roundness of the occlusal surface, ridges’ and crests’ length, or depth of the inter-ridge furrows, indicated as diagnostic features of C. tunuensis, difficult. Other features of C. tunuensis (less vulnerable to erosion) are common for P. rectangulus. These features are: the α angle dimension, curvature of the first ridge, dental plates elongation, number of ridges and prearticular sulcus shape. Measurements of the C. tunuensis holotype (NHMD 115910) show geometrical similarity to ptychoceratodontids (Fig. 3). On these bases we suppose that C. tunuensis is a younger synonym of P. rectangulus. It is impossible to determine if the differences in dimensions of the upper dental plates from Greenland and Germany (e.g., wider α angle and narrower ð2 angle in the German specimens) are caused by an intraspecific variability or evolutionary changes related to isolation in two distant basins (Fig. 7).
Stratigraphic and geographic range.—Germany, Ochsenbach region; Löwenstein Formation (Stubensandstein), Middle Keuper, middle–upper Norian. Greenland, Macknight Bjerg Quarry; lower part of the Carlsberg Fjord Beds (Ørsted Dal Member), Fleming Fjord Formation, middle–upper Norian. France, Grozon region; Marnes de Châlins Formation, Norian.
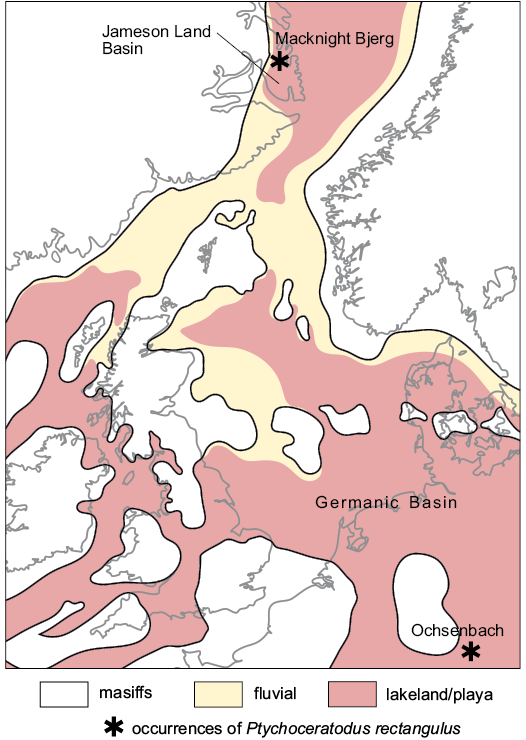
Fig. 7. Map of sedimentation environments in north-western Europe and south-eastern Greenland in Norian with marked occurrences of Ptychoceratodus rectangulus. Based on McKie and Williams (2009) and Ziegler (1998).
Discussion
The five skull bones, from the same strata of the German Triassic as the dental plates, were originally interpreted incorrectly by Linck (1963). They were reinterpreted by Schultze (1981) with a reconstruction of the skull roof, and mentioned by Martin (1981). However, preservation of these skull bones is comparably bad as that of the associated dental plates. All German specimens are imbedded in a rock matrix and only external surfaces of the bones are accessible for study. Most of them are incomplete, with broken or invisible edges, and one specimen was destroyed during preparation (Schultze 1981). The set of the Linck’s (1963) skull roof bones includes, according to Schultze (1981): the KLMN+3 bones left (SMNS 19705) and right (SMNS 19706), the incomplete right I bone (SMNS 19707), the E bone (destroyed) and an unidentified poorly preserved element (SMNS 19708).
In the German material the best preserved are the KLMN+3 bones, which resemble the Greenland specimens. Supraorbital line turning at roughly right angle, and deep indentation for the medial process of the XYZ on the lateral margin are particular similarities. Clear difference between the German and Greenland KLMN+3 bones is the lack of the indentation for the J bone on the posterior margin in SMNS 19705 and conspicuous indentation in KNK 4165 (in SMNS 19706 and KNK 4158 the posterior margins are not preserved or not visible). It implies that the J bone could be very small (Schultze 1981), or merged variably with neighboring bones (as suggested by Schultze 1981 concerning P. serratus). The J bone in P. serratus is placed between the KLMN+3, I, and AB bones. Both specimens interpreted as the I bone (SMNS 19707 and KNK 4161) are highly fragmentary, however SMNS 19707 is much more complete than KNK 4161. Schultze 1981:21 comments: “relation of the I bone and KLMN+3 bone is uncertain, if treated like in P. serratus or Arganodus, there is no space for the J bone, or the J bone is very small”. Indeed, the I bone has a protrusion interpreted in Schultze 1981 as the joint with the AB bone, thereby there is left only a little space for the J bone. Actually, the KLMN+3 imprint (KNK 4165) seems to confirm merging of the I and J bones into the IJ bone (Fig. 8) because of smooth transition between articulations with the I and J bones.
The delta-shaped KNK 4162 is interpreted as the E bone because its posterolateral edges match the KLMN+3 anteromedial margin. Badly preserved SMNS 19708 can represent the same skull element. In Schultze’s (1981) reconstruction, the destroyed bone was interpreted as the E bone (Linck 1963: fig. 3) on the basis of ragged edge in the posterior part, which matches the anteromedial margin the of the KLMN+3 (SMNS 19705, 19706). These interpretations are not contradictory because the destroyed specimen revealed only the ventral surface (Martin 1981), and KNK 4162 only the dorsal. Both specimens had similar size, therefore the shape differences are not an effect of allometric growth, but according to Schultze (1981), the E bone shape can be variable, or specimens are just incomplete. We used the delta-shaped bone as the E bone in the reconstruction, however, it needs confirmation by new material (Fig. 8). Morphology of the minute bone fragment, described in Linck (1936) as the operculum (SMNS 17973), indicates that it is rather the descending process of the XYZ bone.
The skull roof bone pattern of Ptychoceratodus rectangulus (Linck, 1936) is generally similar to other ptychoceratodontids with known skull anatomy: Ptychoceratodus serratus (see Schultze 1981), Ptychoceratodus philipsi (see Kemp 1996), Ferganoceratodus jurassicus Nessov and Kaznyshkin, 1985, and Ferganoceratodus martini Cavin, Suttethorn, Buffetaut, and Tong, 2007. Unique features of the P. rectangulus skull among other ptychoceratodontids are robust ventral crests of the KLMN+3 bone, similar rather to Arganodus atlantis (see Martin 1981), and possibly the delta-shaped E bone with dorsal ridge. Right angle curvature of the supraorbital sensory line is similar rather to Ferganoceratodus spp. than Ptychoceratodus spp. F. martini and F. jurassicus have reduced number of bones in the medial and mediolateral series in comparison with P. serratus. However, number of bones in the medial and mediolateral series in P. rectangulus is uncertain. Presence of only two bones at least in the mediolateral series is highly probable, as discussed above, and such a state is illustrated in the reconstruction (Fig. 8).
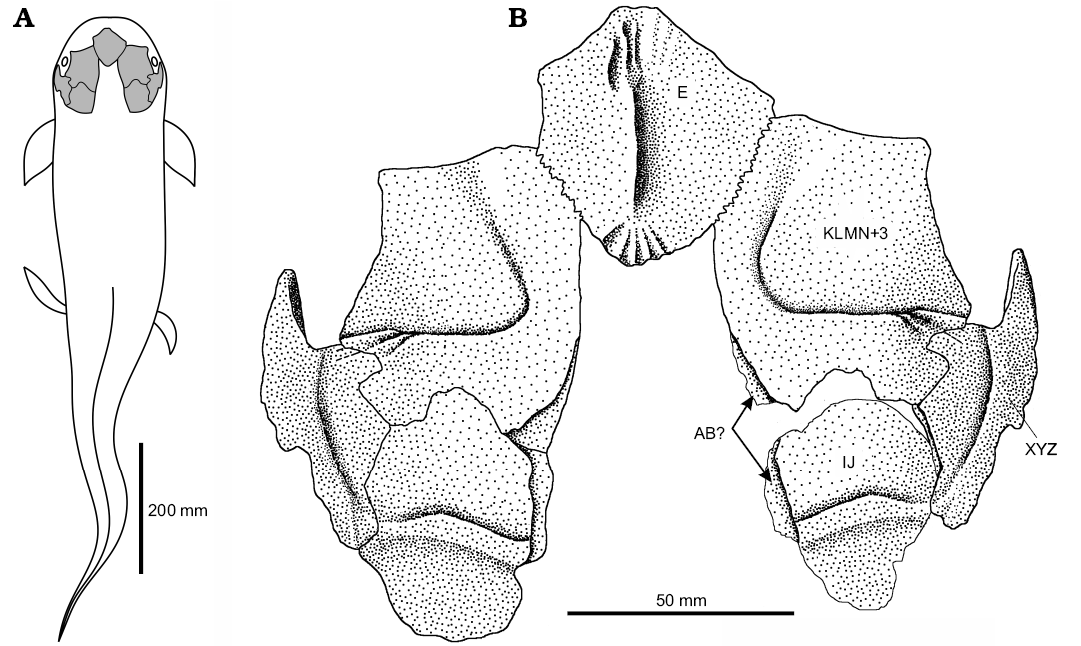
Fig. 8. Skull roof of dipnoan Ptychoceratodus rectangulus. A. Hypothetical fish silhouette with marked position of the skull roof. B. Skull roof reconstruction, originally preserved structures on the left side. Based on: SMNS 19707, KNK 4158, 4159, 4162, 4165. Arrows indicate sutures for corresponding bones.
Ptychoceratodus rectangulus has regularly spaced, relatively large openings in the grooves of temporal line. They correspond to localization of sensory canal pits in Recent lungfishes. Extant Protopterus has similarly developed pattern of these openings (compare Webb and Northcutt 1997). Importance of mechanoreception or electroreception for Protopterus (Bullock et al. 1983; Orlov et al. 2015) suggest similar mode of life for P. rectangulus.
Ptychoceratodus rectangulus coexisted in the Jameson Land Basin with large temnospondyls like the capitosaurid Cyclotosaurus naraserluki (see Marzola et al. 2017) and especially common plagiosaurid Gerrothorax pulcherrimus (see Jenkins et al. 1994, 2008; Schoch and Witzmann 2012; Sulej et al. 2014). Occurrence of the same dipnoan species in the middle–upper Norian sediments of Germanic and Jameson Land basins indicates that at least then the areas were connected. In the Late Triassic both basins had been situated relatively close (Fig. 7), until the opening of the northernmost part of Atlantic Ocean in the Cretaceous, which split Europe and Greenland (Rowley and Lottes 1988). The Mesozoic rifting favored origin of sedimentation basins, which could have been temporally connected with the Germanic Basin and enabled freshwater fauna exchange (McKie and Williams 2009; Ziegler 1998).
Conclusions
The record of Ptychoceratodus from the Fleming Fjord Formation in East Greenland shows a close correlation between the Late Triassic Jameson Land Basin fish fauna and the fauna of the coeval Germanic Basin. Studied Ptychoceratodus rectangulus remains were found in the transitional layers between the Malmros Klint and the Ørsted Dal members and in the lower part of the Carlsberg Fjord Beds (middle–upper Norian). The species was previously known from the Löwenstein Formation (middle–upper Norian) in Germany. Geographical distribution of P. rectangulus shows at least temporary connection between the two basins during middle–late Norian. Recently described Ceratodus tunuensis from the uppermost part of the Carlsberg Fjord Beds is probably conspecific with P. rectangulus. Ptychoceratodus rectangulus had sensory line pit pattern at least similar to that of the extant Protopterus, which suggests a similar mode of life. The Late Triassic dipnoan freshwater fish faunas of the northern Pangea are characterized by a frequent occurrence of Ptychoceratodus.
Acknowledgments
We would like to thank the staff of the Constable Point (East Greenland) airport, and especially Stefan Schirmer (Air Greenland pilot), for logistic help and words of support. We thank to reviewers Jason Pardo (University of Calgary, Alberta, Canada) and Hans-Peter Schultze (Kansas University, Lawrence, USA) for all their help in improving this manuscript. We thank: Krzysztof Michalski for advices in organizing our expedition, other members of the 2014 team: Niels Bonde (University of Copenhagen, Denmark), Błażej Błażejowski (Institute of Paleobiology, PAS, Warsaw, Poland), and Andrzej Wolniewicz (University of Oxford, UK), for hard work in the field, camp and perseverance in unusual conditions. Piotr Skrzycki and Tomasz Szczygielski (both Institute of Paleobiology, PAS) for valuable advices. Łucja Fostowicz-Frelik and Justyna Słowiak (both Institute of Paleobiology, PAS) for sharing Keyence Digital Microscope VHX-900F. The funding for the expedition came from a grant awarded to TS (grant 2012/07/B/NZ8/02707). GN is currently funded by grant from the Swedish Research Council (2017-05248)
References
Agassiz, L. 1837–1843. Recherches sur les Poissons Fossiles, Tome III. viii + 390 pp. Imprimerie de Petitpierre, Neuchâtel.
Agnolín, F.L., Bogan, S., Egli, B.F., Novas, F.E., Isasi, M.P., Marsicano, C., Zavattieri, A., and Mancuso, A. 2017. A new lungfish (Dipnoi) from the Late Triassic of South America. Journal of Vertebrate Paleontology 37: e1245665. Crossref
Agnolín, F.L., Octávio, M., Milàn, J., Marzola, M., Wings, O., Adolfssen, J.S., and Clemmensen, L.B. 2018. Ceratodus tunuensis, sp. nov., a new lungfish (Sarcopterygii, Dipnoi) from the Upper Triassic of central East Greenland. Journal of Vertebrate Paleontology 38: e1439834. Crossref
Andrews, S.D., Kelly, S.R.A., Braham, W., and Kaye, M. 2014. Climatic and eustatic controls on the development of a Late Triassic source rock in the Jameson Land Basin, East Greenland. Journal of the Geological Society 171: 609–619. Crossref
Bendix-Almgreen, S.E. 1976. Palaeovertebrate faunas of Greenland. In: A. Escher and W.S. Watt (eds.), Geology of Greenland, 536–573. Geological Survey of Greenland, Copenhagen.
Bengaard, H.J. and Henriksen, N. 1984. Geological Map of Greenland, Sheet 12, Scoresby Sund, Scale 1: 500 000. Geological Survey of Greenland, Copenhagen.
Bhat, M.S. and Ray, S. 2020. A record of new lungfishes (Osteichthyes: Dipnoi) from the Carnian (Upper Triassic) of India. Historical Biology 32: 428–437. Crossref
Blom, H., Clack, J.A., Ahlberg, P.E., and Friedman, M. 2007. Devonian vertebrates from East Greenland: a review of faunal composition and distribution. Geodiversitas 29: 119–141.
Bullock, T.H., Bodznick, D.A., and Northcutt, R.G. 1983. The phylogenetic distribution of electroreception: evidence for convergent evolution of a primitive vertebrate sense modality. Brain Research 287: 25–46. Crossref
Cavin, L., Suttethorn, V., Buffetaut, E., and Tong, H. 2007. A new Thai Mesozoic lungfish (Sarcopterygii, Dipnoi) with an insight into post-Palaeozoic dipnoan evolution. Zoological Journal of Linnaean Society 149: 141–177. Crossref
Clack, J.A., Challands, T.J., Smithson, T.R., and Smithson, K.Z. 2018. Newly recognized Famennian lungfishes from East Greenland reveal tooth plate diversity and blur the Devonian/Carboniferous boundary. Papers in Palaeontology 5: 261–279. Crossref
Clemmensen, L.B. 1980. Triassic lithostratigraphy of East Greenland between Scoresby Sund and Kejser Franz Josephs Fjord. Geological Survey of Greenland Bulletin 139: 1–56.
Clemmensen, L.B., Kent, D.V., and Jenkins F.A. 1998. A Late Triassic lake system in East Greenland: facies, depositional cycles and palaeoclimate. Palaeogeography, Palaeoclimatology. Palaeoecology 140: 135–159. Crossref
Clemmensen, L.B., Milan, J., Adolfssen, J.S., Estrup, E.J., Frobøse, N., Klein, N., Mateus, O., and Wings O. 2016. The vertebrate-bearing Late Triassic Fleming Fjord Formation of central East Greenland revisited: stratigraphy, palaeoclimate and new palaeontological data. Geological Society, London. Special Publications 434: 31–47. Crossref
Cuny, G., Martin, M., Rauscher, R., and Mazin, J.M. 1998. A new neoselachian shark from the Upper Triassic of Grozon (Jura, France). Geological Magazine 135: 657–668. Crossref
Huxley, T.H. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Zoological Society of London 43: 649–662.
Frederickson, J.A. and Cifelli, R.L. 2017. New Cretaceous lungfishes (Dipnoi, Ceratodontidae) from western North America. Journal of Paleontology 91: 146–161. Crossref
Greenwood, P.H. 1986. The natural history of African lungfishes. Journal of Morphology 190 (Supplement 1): 163–179. Crossref
Holmes, T. 2008. March Onto Land: The Silurian Period to the Middle Triassic Epoch (Prehistoric Earth). 192 pp. Chelsea House, New York.
Jaekel, O. 1926. Zur Morphogenie der Gebisse und Zähne. Vierteljahresschrift Zahnheilkunde 42: 217–242.
Jenkins Jr., F.A., Shubin, N.H., Amaral, W.W., Gatesy, S.M., Schaff, C.R., Clemmensen, L.B., Downs, W.R., Davidson, A.R., Bonde, N., and Osbæck F. 1994. Late Triassic continental vertebrates and depositional environments of the Fleming Fjord Formation, Jameson Land, East Greenland. Meddelelser om Grønland, Geoscience 32: 3–25.
Jenkins Jr., F.A., Gatesy, S.M.,
Shubin, N.H., and
Amaral, W.W. 1997. Haramiyids and Triassic
mammalian evolution. Nature 385: 715–718. Crossref
Jenkins Jr, F.A, Shubin, N.H., Gatesy, S.M., and Padian, K. 2001. A diminutive species of pterosaur (Pterosauria: Eudimorphodontidae) from the Greenlandic Triassic. Bulletin of the Museum of Comparative Zoology, Harvard 156: 151–170.
Jenkins Jr, F.A., Shubin, N.H., Gatesy, S.M., and Warren, A. 2008. Gerrothorax pulcherrimus from the Upper Triassic Fleming Fjord Formation of East Greenland and a reassessment of head lifting in temnospondyl feeding. Journal of Vertebrate Paleontology 28: 935–950. Crossref
Kemp, A. 1986. The biology of the Australia lungfish, Neoceratodus forsteri (Krefft 1870). Journal of Morphology 190: 181–198. Crossref
Kemp, A. 1996. Triassic lungfish from Gondwana. In: G. Arratia and G. Viohl (eds.), Mesozoic Fishes—Systematics and Paleoecology, 409–416. Verlag Dr. Friedrich Pfeil, München.
Kemp, A. 1997. A revision of Australian Mesozoic and Cenozoic lungfish of the family Neoceratodontidae (Osteichthyes: Dipnoi) with a description of four new species. Journal of Paleontology 71: 713–733. Crossref
Kemp, A. 1998. Skull structure in post-Paleozoic lungfish. Journal of Vertebrate Paleontology 18: 43–63. Crossref
Kemp, A. 2017. Evolutionary history of lungfishes with a new phylogeny of post-Devonian genera. Palaeogeography, Palaeoclimatology, Palaeoecology 471: 209–219. Crossref
Klein, H., Milàn, J., Clemmensen, L.B., Frobøse, N., Mateus, O., Klein, N., Adolfssen, J.S., Estrup, E.J., and Wings, O. 2016. Archosaur footprints (cf. Brachychirotherium) with unusual morphology from the Upper Triassic Fleming Fjord Formation (Norian–Rhaetian) of East Greenland. Geological Society, London, Special Publications 434: 71–85. Crossref
Lallensack, J.N., Klein, H.J., Wings, O., Mateus, O., and Clemmensen, L.B. 2017. Sauropodomorph dinosaur trackways from the Fleming Fjord Formation of East Greenland: evidence for Late Triassic sauropods. Acta Palaeontologica Polonica 62: 833–843. Crossref
Lehman, J.-P. 1959. Les dipneustes du Devonien Superieur du Groenland. Meddelelser om Grønland 160: 1–58.
Linck, O. 1936. Ein Lebensraum von Ceratodus im Stubensandstein des Strombergs mit Ceratodus rectangulus n. sp. und anderen Arten. Verein für vaterländische Naturkunde in Württemburg, Jahreshefte 92: 45–68. Crossref
Linck, O. 1938. Nachtrag zu: Ein Lebensraum von Ceratodus im Stubensandstein des Strombergs usw. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 94: 4–14.
Linck, O. 1963. Schädelknochen von Ceratodus Agassiz aus dem Stubensandstein (Trias, Mittl. Keuper 4) Württembergs und ihre Bedeutung. Paläontologische Zeitschrift 37: 268–276.
Marshall, C.R. 1986a. A list of fossil and extant Dipnoans. Journal of Morphology 190 (Supplement 1): 15–23. Crossref
Marshall, C.R. 1986b. Lungfish: phylogeny and parsimony. Journal of Morphology 190 (Supplement 1): 151–162. Crossref
Martin, M. 1981. Les Dipneustes et Actinistiens du Trias superieur continental marocain. Stuttgarter Beiträge zur Naturkunde Serie B 69: 1–30.
Martin, M. 1982. Nouvelles données sur la phylogénie et la systématique des Dipneustes postpaléozoïques, conséquences stratigraphiques et paléogéographiques. Geobios 15: 53–64. Crossref
Martin, M., Barbieri, L., and Cuny, G. 1999. The Madagascan Ptychoceratodontids (Dipnoi). Systematic relationships and paleobiogeographical significance. Oryctos 2: 3–16.
Marzola, M., Mateus, O., Shubin, N.H., and Clemmensen, L.B. 2017. Cyclotosaurus naraserluki, sp. nov., a new Late Triassic cyclotosaurid (Amphibia, Temnospondyli) from the Fleming Fjord Formation of the Jameson Land Basin (East Greenland). Journal of Vertebrate Paleontology 37: e1303501. Crossref
Mckie, T. and Williams B. 2009. Triassic palaeogeography and fluvial dispersal across the northwest European Basins. Geological Journal 44: 711–741. Crossref
Milàn, J., Clemmensen, B.L., and Bonde, N. 2004. Vertical sections through dinosaur tracks (Late Triassic lake deposits, East Greenland)—undertracks and other subsurface deformation structures revealed. Lethaia 37: 285–296. Crossref
Miles, R.S. 1977. Dipnoan (lungfish) skulls and the relationships of the group: a study based on new species from the Devonian of Australia. Zoological Journal of the Linnean Society 61: 1–328. Crossref
Müller, J. 1845. Über den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische. Abhandlungen der königlichen preussischen Akademie der Wissenschaften zu Berlin, Physikalisch-Mathematische Classe 11: 117–216.
Nessov, L.A. and Kaznyshkin, M.N. 1985. A lungfish and turtles from Upper Jurassic of Northern Fergana, Kirghiz SSR [in Russian]. Vestnik Zoologii 1: 33–39.
Niedźwiedzki, G. 2017. Tetrapod track assemblages from the Upper Triassic Fleming Fjord Formation, East Greenland. Palaeontologia Africana 52: 179.
Niedźwiedzki, G. and Sulej, T. 2017. A Norian coelophysoid theropod from Fleming Fjord Formation, East Greenland. In: A. Farke, A. MacKenzie, and J. Miller-Camp (eds.), 77th Annual Meeting of the Society of Vertebrate Paleontology, Calgary. Abstract Book, 169. Society of Vertebrate Paleontology, Calgary.
Orlov, A.A., Golubtsov, A.S., Baron, V.D., and Pavlov, D.S. 2015. Bioelectric fields of the African Marbled Lungfish Protopterus aethiopicus (Sarcopterygii: Protopteridae), African (Heterotis niloticus) and South American Silver (Osteoglossum bicirrhosum) Arowanas (Actinopterygii: Osteoglossidae): primitive electrogenesis? Journal of Ichthyology 55: 874–879. Crossref
Richter, M. and Toledo, C.E.V. 2008. The first Triassic lungfish from South America (Santa Maria Formation, Parana Basin) and its bearing on geological correlations within Pangaea. Geological Society, London, Special Publications 295: 43–54. Crossref
Romer, A. S. 1955. The Vertebrate Body. 643 pp. W.B. Saunders Company, Philadelphia.
Rowley, D.B. and Lottes, A.L. 1988. Plate-kinematic reconstructions of the North Atlantic and Arctic: Late Jurassic to Present. Tectonophysics 155: 73–120. Crossref
Schoch, R.R. and Witzmann, F. 2012. Cranial morphology of the plagiosaurid Gerrothorax pulcherrimus as an extreme example of evolutionary stasis. Lethaia 45: 371–385. Crossref
Schultze, H.-P. 1981. Das Schädeldach eines ceratodontiden Lungenfisches aus der Trias Süddeutschlands (Dipnoi, Pisces). Stuttgarter Beiträge zur Naturkunde. Serie B (Geologie und Paläontologie) 70: 1–31.
Schultze, H.-P. 1992. Dipnoi—Fossilium Catalogus. I: Animalia. Pars 131. 464 pp. Kugler Publications, Amsterdam.
Schultze, H.-P. 2004. Mesozoic sarcopterygians. In: G. Arratia and A. Tintori (eds.), Mesozoic Fishes—Systematics, Paleoenvironments and Biodiversity, 463–492. Dr. Friedrich Pfeil, München.
Shapiro, M.D. and Jenkins, F.A. 2001. Cynodont from the Upper Triassic of East Greenland: tooth replacement and double-rootedness. Bulletin of the Museum of Comparative Zoology at Harvard College 156: 49–58.
Skrzycki, P. 2015. New species of lungfish (Sarcopterygii, Dipnoi) from the Late Triassic Krasiejów site in Poland, with remarks on the ontogeny of Triassic dipnoan tooth plates. Journal of Vertebrate Paleontology 35: e964357. Crossref
Skrzycki, P. 2016. The westernmost occurrence of Gnathorhiza in the Triassic, with a discussion of the stratigraphic and palaeogeographic distribution of the genus. Fossil Record 19: 17–29. Crossref
Skrzycki, P., Niedźwiedzki, G., and Tałanda, M. 2018. Dipnoan remains from the Lower–Middle Triassic of the Holy Cross Mountains and northeastern Poland, with remarks on dipnoan palaeobiogeography. Palaeogeography, Palaeoclimatology, Palaeoecology 496: 332–345. Crossref
Sulej, T., Wolniewicz, A., Bonde, N., Błażejowski, B., Niedźwiedzki, G., and Tałanda, M. 2014. New perspectives on the Late Triassic vertebrates of East Greenland: preliminary results of a Polish-Danish palaeontological expedition. Polish Polar Research 35: 541–552. Crossref
Vorobyeva, E.I. and Minikh, M.G. 1968. Experimental application of biometry to the ceratodontid dental plates [in Russian]. Paleontologičeskij žurnal 2: 76–87.
Webb, J.F. and Northcutt, R.G. 1997. Morphology and distribution of pit organs and canal neuromasts in non-teleost bony fishes. Brain Behavior & Evolution 50: 139–151. Crossref
Wright, D.E. 1974. Morphology of the gill epithelium of the lungfish, Lepidosiren paradoxa. Cell and Tissue Research 153: 365–381. Crossref
Ziegler, P.A. 1998. Evolution of the Arctic-North Atlantic and the Western Tethys: A visual presentation of a series of Paleogeographic-Paleotectonic maps. AAPG Memoir 43: 164–196.
Acta Palaeontol. Pol. 65 (3): 561–574,
2020 https://doi.org/10.4202/app.00679.2019