
A new neolepadid cirripede from a Pleistocene cold seep, Krishna-Godavari Basin, offshore India
ANDY S. GALE, CRISPIN T.S. LITTLE, JOEL E. JOHNSON, and LIVIU GIOSAN
Gale, A.S., Little, C.T.S, Johnson, J.E., and Giosan, L. 2020. A new neolepadid cirripede from a Pleistocene cold seep, Krishna-Godavari Basin, offshore India. Acta Palaeontologica Polonica 65 (2): 351–362.
Valves of a thoracican cirripede belonging to a new species of the Neolepadidae, Ashinkailepas indica Gale sp. nov. are described from a Late Pleistocene cold seep (52.6 ka), cored in the Krishna-Godavari Basin, offshore from the eastern coast of India. This constitutes the first fossil record of the genus, and its first occurrence in the Indian Ocean. Other fossil records of the Neolepadidae (here elevated to full family status) are discussed, and it is concluded that only Stipilepas molerensis from the Eocene of Denmark, is correctly referred to the family. Cladistic analysis of the Neolepadidae supports a basal position for Ashinkailepas, as deduced independently from molecular studies, and the Lower Cretaceous brachylepadid genus Pedupycnolepas is identified as sister taxon to Neolepadidae. Neolepadids are not Mesozoic relics as claimed, preserved in association with the highly specialised environments of cold seeps and hydrothermal vents, but are rather an early Cenozoic offshoot from the clade which also gave rise to the sessile cirripedes.
Key words: Cirripedia, Thoracica, Neolepadidae, cold seep, Pleistocene, Indian Ocean.
Andy S. Gale [andy.gale@port.ac.uk], School of Earth, Environmental and Geographical Sciences, University of Portsmouth, Burnaby Building, Burnaby Road, Portsmouth PO13QL, UK.
Crispin T.S. Little [C.T.S.Little@earth.leeds.ac.uk], School of Earth and Environment, University of Leeds, Leeds LS2 9JT, UK.
Joel E. Johnson [joel.johnson@unh.edu], Department of Earth Sciences, University of New Hampshire, Durham, NH 03824, USA.
Liviu Giosan [lgiosan@whoi.edu], Geology & Geophysics, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USA.
Received 23 November 2019, accepted 14 February 2020, available online 22 April 2020.
Copyright © 2020 A.S. Gale et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Amongst the remarkable range of invertebrates associated with deep sea hydrothermal vents and cold seeps are thoracican cirripedes referred to the subfamily Neolepadinae Newman, 1985. Species belonging to this taxon are currently known from the South Atlantic, Indian, Pacific, and Southern oceans (Herrera et al. 2015; Watanabe et al. 2018). It is likely that many of these species obtain carbon through endosymbiotic chemoautotrophic bacteria (Southward and Newman 1998). A total of eight species of living neolepadines have been described, and assigned to the genera Neolepas, Ashinkailepas, Vulcanolepas, and Leucolepas. Neolepadids have been identified as relic Mesozoic forms which have survived in highly specialised habitats and considered to be primitive scalpellomorphs, most closely allied to the Jurassic Archaeolepas (Newman 1979; Yamaguchi et al. 2004). Three fossil taxa have been assigned to the Neolepadinae; ?Neolepas augurata Buckeridge and Grant-Mackie, 1985, from the Lower Jurassic (Sinemurian–Pliensbachian) of New Caledonia, Litholepas klausreschi Nagler, Haug, Glenner, and Buckeridge, 2017 from the Upper Jurassic (Tithonian) of Germany, and Stipilepas molerensis Carriol in Carriol et al., 2016, from the Eocene (Ypresian) of Denmark. None of these are associated with hydrothermal vents or cold seeps.
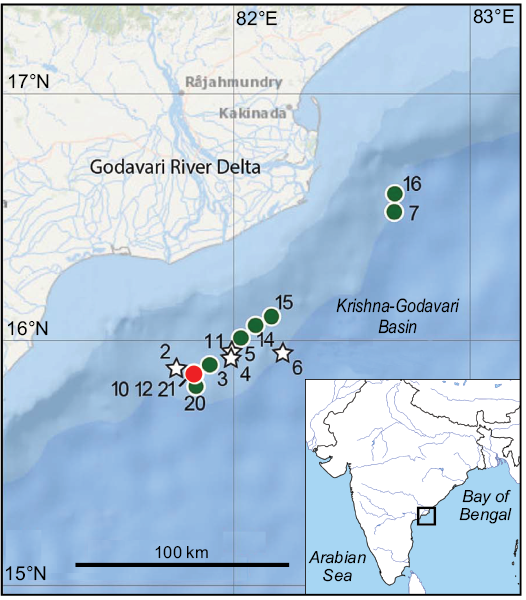
Fig. 1. NGHP-01 core sites (circles) and logging sites (stars) in the offshore Krishna-Godavari Basin, modified from Lorenson and Collett (2018). Site 12 is co-located with sites 10 and 21 (red circle). Inset shows the study area in Krishna-Godavari Basin is located on the eastern margin of peninsular India in the western Bay of Bengal.
In this paper, we describe a new species of Ashinkailepas from the Late Pleistocene (52.6 ka) found in a sediment core from the Krishna-Godavari (K-G) Basin, offshore India, which was associated with a contemporary methane hydrate system and methane seepage. On the basis of this new species we review the taxonomy, possible origin and fossil record of the Neolepadinae, and undertake a cladistic analysis of the subfamily.
Institutional abbreviations.—MJML, Museum of Jurassic Marine Life, Kimmeridge, Dorset, UK; NHMUK, Natural History Museum, London, UK; SMNS, Stuttgart Museum of Natural History, Stuttgart, Germany; UIWA, National Institute of Water and Atmospheric Research, Wellington, New Zealand; SNM, Smithsonian Museum, Washington DC, USA; UoA, University of Auckland, Geology Department, New Zealand.
Other abbreviations.—AOM, anaerobic oxidation of methane; K-G, Krishna-Godavari; mbsf, metres below sea floor; MDAC, methane derived authigenic carbonates; SMTZ, sulfate-methane transition zone; VPDB, Vienna PeeDee Belemnite.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:7D8A63AC-765E-4C95-8610-B8ED6D7B7605.
Geological and geographic setting
The studied cirripede specimens come from sediment core NGHP-01-12A-03H-2, which was drilled in 2006 by the D/V JOIDES Resolution during the Indian National Gas Hydrate Program Expedition 01 (NGHP-01) in 1046 m of water depth at 15°51.6335’ N, 81°50.2274’ E in the K-G Basin, on the eastern Indian continental margin (Collett et al. 2015, 2014). The sediment in the core and throughout the offshore K-G Basin consists of variably coloured hemipelagic clay (Collett et al. 2015; Phillips et al. 2014) containing greater or lesser quantities of organic carbon (Johnson et al. 2014), and calcareous nannofossils and foraminifera (Flores et al. 2014). The cirripedes come from a level between 16 and 17 metres below sea floor (mbsf) in this core (Table 1) and they are associated with whole and fragmentary shells of seep-related bivalves and gastropods. In addition, there are numerous variably sized authigenic carbonate nodules formed of high magnesium calcite, two of which have δ13C values of -48.23‰ and -45.33‰ (VPDB), indicative the anaerobic oxidation of methane (AOM) near the sulfate-methane transition zone (SMTZ) (Teichert et al. 2014). The presence of the seep molluscs and methane derived authigenic carbonates (MDAC) defines a palaeo-seafloor SMTZ and thus palaeo-methane seepage. Similar shell material and MDAC have been found at the same ~16 mbsf level in two nearby sediment cores: NGHP-01-10D (15°51.8609’ N, 81°50.0749’ E, 1038 m water depth; Collett et al. 2015; Teichert et al. 2014), and a 29.9 m long sediment core collected by Giant Calypso piston corer water during cruise MD-161 of the R/V Marian Dufresne in May 2007 at 15°51.8624’ N, 81°50.0692’ E in 1033 m water depth (Mazumdar et al. 2009). An age model developed from a combined dataset of radiocarbon dates from foraminifera tests, bivalve shells and MDAC from the ~16 mbsf level from these three cores yields an extrapolated age of ~52.6 ka 14C years BP. (Mazumdar et al. 2009; Connolly et al. 2009), equating to the Late Pleistocene stage. Occurrences are listed in Table 1.
Table 1. Details of specimens obtained from NGHP-01 Site 12 Hole A, Core 3H, Section 2, offshore K-G Basin, India.
|
Sample interval (cm) |
Depth (mbsf) |
Number of specimens |
|
52–57 |
16.38 |
10 |
|
50–60 |
16.38 |
5 |
|
80–90 |
16.68 |
13 |
|
84–86 |
16.68 |
12 |
|
90–100 |
16.78 |
2 |
|
100–110 |
16.88 |
5 |
|
116–118 |
17 |
11 |
Systematic palaeontology
(by Andy S. Gale)
Infraclass Cirripedia Burmeister, 1834
Superorder Thoracica Darwin, 1854
Clade Thoracicalcarea Gale, 2015a
Diagnosis.—Thoracica in which the shell plates are composed of calcite.
Clade Unilatera Gale, 2019a
Diagnosis.—Thoracicalcarea in which only one lateral plate (the paired upper latus) is present; rostrum large, triangular, external to the scuta. Surface sculpture plesiomorphically composed of evenly spaced terracing.
Remarks.—Unilterans are a well-defined group, which appeared in the Lower Cretaceous, and include the sessile barnacles (Gale 2019a).
Family Neolepadidae Yamaguchi, Newman, and Hashimoto, 2004
Included genera: Neolepas Newman, 1979, Ashinkailepas Yamaguchi, Newman, and Hashimoto, 2004, Vulcanolepas Southward and Jones, 2003, Leucolepas Southward and Jones, 2003, Stipilepas Carriol in Carriol et al., 2016.
Diagnosis.—Pedunculate calcareous forms which possess an 8 plated capitulum comprising two scuta, two terga, two upper latera, a carina and a rostrum. The large rostrum articulates with the rostral margin of the scutum, and the carina with the lower carinal margin of the tergum.
Remarks.—As demonstrated by X-ray diffractometry analysis, Ashinkailepas has shell plates composed of calcite. The Neolepidinae Yamaguchi, Newman, and Hashimoto, 2004, is therefore raised to family status, because the family Eolepadidae Buckeridge, 1983, to which it was previously assigned, has been shown to have a phosphatic shell (Høeg et al. 1999; Gale 2014b; Gale and Schweigert 2015), and includes only the genera Eolepas and Toarcolepas. Living forms are exclusively associated with hydrothermal vents and methane seeps (Watanabe et al. 2018), but the Eocene Stipilepas is found in open marine clay sediments deposited under dysoxic conditions (Carriol et al. 2016).There have been three records of fossil neolepadids, of which only Stipilepas appears to me to belong to the family. The other records are described below.
?Neolepas augurata Buckeridge and Grant-Mackie, 1985
Fig. 2A, B.
Material.—UoA A291a, b, A292a, b; two mouldic valves from the Sinemurian–Pliensbachian (Lower Jurassic), New Caledonia.
Remarks.—This species is based upon a single scutum and a fragmentary tergum from the Lower Jurassic of New Caledonia (Fig. 2A, B). The scutum is asymmetrically triangular, with a broad basal margin and an acuminate umbo, and somewhat resembles in outline the scutum of Toarcolepas mutans Gale and Schweigert, 2015 (Fig. 2C). This phosphatic taxon from the Lower Jurassic of Germany is now known from articulated individuals (Gale et al. 2019: fig. 7A–C). However, the basal margin in ?N. augurata is more convex than in T. mutans, and the external surface of the valve appears to be swollen. The tergum of ?N. augurata appears similar in shape and sculpture (curved apicobasal line, moderately strong co-marginal ridges, weaker radial ridges) with those of Eolepas bathonica Withers, 1928 (compare Withers 1928: pl. 2: 7–9). The species is tentatively interpreted as a possible eolepadid, but the material requires further study.
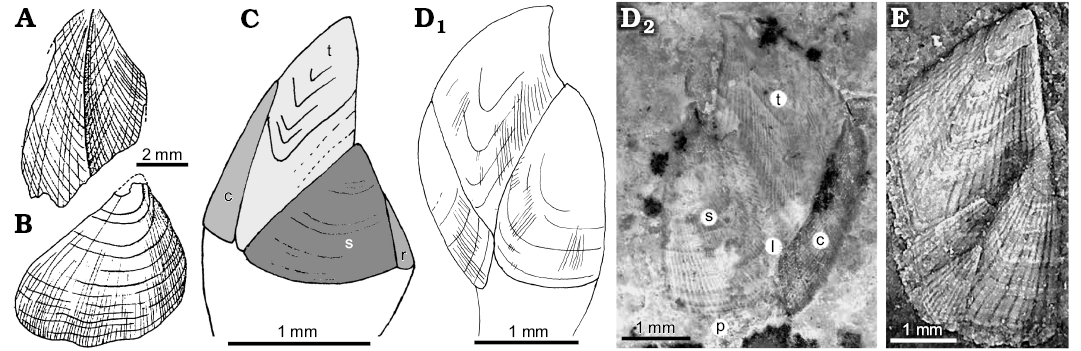
Fig. 2. Fossil taxa assigned to Neolepadidae. A, B. ?Neolepas augurata Buckeridge and Grant-Mackie, 1885 (original of Buckeridge and Grant-Mackie 1985: figs. 2, 3); Sinemurian–Pliensbachian (Lower Jurassic), New Caledonia. A. University of Auckland Geology Department UoA A291a, b, external mould of tergum. B. University of Auckland Geology Department UoA A292a, b, external mould of scutum. This is here tentatively interpreted as an eolepadid. C. Toarcolepas mutans Gale and Schweigert, 2015 (SMNS 26029); Toarcian (Jurassic), Harpoceras falciferum Zone, Zell u. Aichelberg, Germany; reconstruction based upon type material (Gale and Schweigert 2015: fig. 5). D. Litholepas klausreschi Nagler, Haug, Glenner, and Buckeridge, 2017; Tithonian (Upper Jurassic), Hybernoticeras hybernotum Zone, Eichstatt, Germany; D2, SMNS 70388/5 (decolourised original of Nagler et al. 2017: fig. 5B). I am unable to identify the upper latus, marked “l” in their figure; D1, reconstruction (mirrored for comparison with C). E. Concinnalepas costata (Withers, 1928) (NHMUK IC 1103); Kimmeridge Clay, 2 m beneath Freshwater Stone Band, Pectinatites pectinatus Zone, Tithonian (Upper Jurassic), Kimmeridge, Dorset, UK; fragmentary capitulum, showing scutum, tergum, and two lateral plates (based on original of Gale 2014: fig. 6l). Abbreviations: c, carina; l, upper latus; p, peduncle; r, rostrum; s, scutum; ; t, tergum.
Litholepas klausreschi Nagler, Haug, Glenner, and Buckeridge, 2017
Fig. 2D.
Material.—SMNS 70388/5; 13 individuals attached to a sponge, lower Tithonian (Jurassic), Solnhofen Limestone, Eichstätt, Germany.
Remarks.—This species is based upon 13 small (capitular length approximately 5 mm) specimens, attached to a sponge, from the Solnhofen Limestone of Eichstätt, Germany (Nagler et al. 2017). I am unable to agree with their published interpretation that an upper latus is present on the specimens, and it appears that the colourised regions marked “l” in their figures (e.g., Nagler et al. 2017: fig. 5B, reproduced here as Fig. 2D2) do not mark a distinct plate. I have therefore reconstructed the species without an upper latus (Fig. 2D1). The large number of growth-lines on the specimens can be taken as evidence that the individuals are fully grown.
The sculpture of the plates is cancellate, composed of very fine radial ribs which intersect with prominent growth lines and is similar to that found in zeugmatolepadids such as Concinnalepas (Fig. 2E). The shape of the terga and scuta are also similar to those of Concinnalepas (tergal apex inclined ventrally, gently convex carinal margins, rostral and basitergal angles of scuta nearly 90°). The genus is therefore tentatively interpreted as a paedomorphic zeugmatolepadid, in parallel with the titanolepadid genus Ivolepas Gale in Gale and Sorensen, 2015 and the scalpellid Virgilepas Gale, 2020. In these genera, lateral plates are absent, probably as a consequence of precocious (accelerated) development.
Stratigraphic and geographic range.—Lower Tithonian, southern Germany.
Genus Ashinkailepas Yamaguchi, Newman, and Hashimoto, 2004
Type species: Ashinkailepas seepiophila Yamaguchi, Newman, and Hashimoto, 2004; Recent, off Japan.
Species included: Ashinkailepas kermadecensis Buckeridge, 2009, Ashinkailepas indica Gale sp. nov.
Emended diagnosis.—Neolepadidae which possess short peduncles (height less than or equal to that of capitulum); peduncle armoured with 6 whorls of large, robust, triangular plates which terminate apically in a point; capitular plates bear sculpture of apicobasally radiating ribs, which intersect with well-marked growth increments.
Remarks.—Detailed morphological comparison of the fossil material with the two described living species of Ashinkailepas is difficult, because the figured material of complete individuals is alcohol-wet, and details of plate sculpture are quite clear. However, such differences as are visible are outlined below.
Ashinkailepas indica Gale sp. nov.
Figs. 3A–F, 4A–M.
Zoobank LSID: urn:lsid:zoobank.org:act:18AA3E40-1B2E-44EE-9588-109780A3A74C
Etymology: Indica, latinised form of India, the country adjacent to the discovery site.
Type material: Holotype: tergum (NHMUK IC1409). Paratypes: carinae, scuta, rostra, upper latera, peduncular plates (NHMUK IC 1402–1408; 1410–1420) from the type locality and horizon.
Type locality: Krishna-Godavari Basin, offshore east coast of India.
Type horizon: Late Pleistocene, 52.6 ka.
Material.—Over 100 capitular plates and peduncular scales (NHMUK) from the Late Pleistocene (52.6 ka) of the Krishna-Godavari Basin, offshore east coast of India.
Diagnosis.—Ashinkailepas in which radial ribbing is weakly developed, and restricted to the apical, occludent portion of the scutum; the terga possess a smooth, triangular region on the ventral side of the apicobasal ridge which bears only growth lines.
Description.—Terga (Figs. 3A–C, 4A–C) flat, rhomboidal, slightly asymmetrical, as the occludent margin is longer than the upper carinal margin. The umbo is slightly inclined towards the dorsal margin, and a weak, narrow ridge for scutal contact is present. A raised apicobasal ridge runs from the umbo to the basal angle; this is either straight, curved or slightly sinuous. A minor secondary ridge is present on the ventral side of the apicobasal ridge, and the two define a slightly raised, triangular region dominated by growth increments. The sculpture of the terga comprises evenly spaced, weakly terraced growth increments, and a variably developed radial element, which forms small nodes where ridges intersect with growth lines. The scuta (Figs. 3E, 4E, F) are rhomboidal and strongly dorsally inclined, with a concave tergal margin and a convex occludent margin; the rostral and inner margins are short and straight. A sharply defined, raised apicobasal ridge is present. The sculpture comprises evenly spaced terracing, parallel with growth increments, and weak radial ridges, concentrated in the apical region adjacent to the occludent margin. On small scuta, the radial sculpture forms discrete, short nodes on each growth increment.
The outline of the upper latera (Figs. 3C, 4I) forms an isosceles triangle, twice as tall as broad; the sculpture comprises irregular transverse grooves which are inflected towards the basal margin. The rostra (Figs. 3F, 4J) are triangular in ventral aspect, 2–3 times taller than broad, and carry a raised central ridge which strengthens apically, and the lateral margins are slightly swollen on larger plates. The carinae (Figs. 3D, 4D) are very tall (4 times width) and taper gradually to the apex; they carry a rounded central ridge, and raised lateral margins. Regularly spaced, terraced, growth increments flare slightly on the lateral margins and central ridge. A few (2–4) radial elements form irregular, narrow ridges, subparallel with the central axis. The peduncular plates (Fig. 4 G, H, K, L, M) have a slightly asymmetrically triangular form, and are robust. They possess two lateral processes and a tall, outwardly curved central spine. The interior face is flat to weakly concave and smooth; the external surface bears short, coarse ridges.

Fig. 3. Capitular plates of neolepadid cirripede Ashinkailepas indica Gale sp. nov. from Late Pleistocene (52.6 ka), Krishna-Godavari Basin, offshore India (A–G) and living species of the genus (H, I). A–F. Capitular plates of paratypes. A. NHMUK IC 1402, tergum in external (A1) and internal (A2) views. B. NHMUK IC 1403 small tergum in external view. C. NHMUK IC 1404, upper latus in external view. D. NHMUK IC 1405, carina in dorsal (D1) and lateral (D2) views. E. NHMUK IC 1406, small scutum in internal (E1) and external (E2) views. F. NHMUK IC1407, rostrum in ventral view. G. Recent Ashinkailepas seepiophila Yamaguchi, Newman, and Hashimoto, 2004 from near Sagami Bay, Japan. USNM 1018131, paratype in lateral (right side) view (based on Yamaguchi et al. 2004: fig. 4). H. Recent Ashinkailepas kermadecensis Buckeridge, 2009 from Kermadec Ridge, northeast of North Island, New Zealand. NIWA 44722, holotype in lateral view (based on Buckeridge 2009: pl. 1: 5).

Fig. 4. Plates of neolepadid cirripede Ashinkailepas indica Gale sp. nov. from Late Pleistocene, 52.6 ka) Krishna-Godavari Basin, offshore India. A–C. External views of terga. A. NHMUK IC 1408. B. NHMUK IC 1409, holotype. C. NHMUK IC 1410. D. NHMUK IC 1411, carina in dorsal view. E. NHMUK IC 1412, scutum external (E1) and internal (E2) views. F. NHMUK IC 1413, small scutum in external view, with predatory gastropod drillhole Oichnus paraboloides Bromley, 1981. I. NHMUK IC 1416, upper latus in external view, with predatory boring made by gastropod. J. NHMUK IC 1417, rostrum in ventral view. G, H, K–M. NHMUK IC 1414, 1415, 1417–1419, respectively, peduncular plates in external (G, L, K) and internal (H, M) views.
Remarks.—The fossil material of dissociated plates can be referred confidently to Ashinkailepas, from comparison of the shape of the capitular and peduncular plates with illustrations of A. seepiophila (Yamaguchi et al. 2004) and A. kermadecensis Buckeridge, 2009. The capitular plates lack the numerous, fine radial ribs of A. seepiophila, which also lacks conspicuous growth increments, and are more closely comparable with those of A. kermadecensis. Detailed comparison is difficult, because the photographic illustrations of that species (Buckeridge 2009: pl. 1) are wet with alcohol and the surface sculpture is rather difficult to make out. Differences include the very narrow, sharply defined apicobasal ridge on the terga of A. kermadecensis, whereas this is blunter and broader, with an accessory ridge, in the material described herein as A. indica Gale sp. nov. Additionally, in A. kermadecensis, very fine radial ridges, interrupted by growth increments, cover the entire terga and scuta (e.g., Buckeridge 2009: pl. 1: 3); in A. indica Gale sp. nov. the radial sculpture is weakly developed, and concentrated in the apical region of the scuta and terga.
Stratigraphic and geographic occurrence.—Late Pleistocene of Krishna-Godavari Basin, offshore east coast of India.
Discussion
Palaeoecology.—Two of the plates, an upper latus (Fig. 4J) and a scutum (Fig. 4G) bear circular, bevelled borings ascribable to the trace Oichnus paraboloides Bromley, 1981. The boring in the base of the upper latus (Fig. 4J) is 1.4 mm in diameter, and incomplete, the missing portion of the boring must have penetrated the lower part of the tergum, or more probably, the scutum. The second boring is in the lower, dorsal part of a scutum and is 1.6 mm in diameter, and the missing portion must have penetrated the upper latus and tergum. The position of these two borings overlies the centre of the soma. Similar predatory borings in cirripedes were reviewed by Klompmaker et al. (2015), and recorded from Cenozoic pedunculate Zeugmatolepadidae and Scalpellidae. The probable predators responsible were identified as muricid gastropods. Predatory borings have not been recorded from living neolepadids.
After Ashinkailepas seepiophila Yamaguchi, Newman, and Hashimoto, 2004, A. indica Gale sp. nov. is the second species within the genus that is associated with active methane seepage. The benthic fauna associated with the disarticulated A. indica Gale sp. nov. specimens include vesicomyid and thyasirid clams, and bathymodiolin mussels. All of these bivalves house chemoautotrophic bacteria in their gills and are typical taxa of methane seep communities (e.g., Dubilier et al. 2008; Levin 2005). Modern cold seep communities have recently been reported from the Krishna-Godavari Basin in 900–1800 m water depth by Mazumdar et al. (2019), in the same general area as the material described here. The fauna from these active sites include vesicomyid, thyasirid and solemyid clams, bathymodiolin mussels, gastropods, munidopsid and galatheid squat lobsters, and neolepadid barnacles attached to mussel shells (Mazumdar et al. 2019: fig. 3q). The latter are suggested by Mazumdar et al. (2019) to be a species of Neolepas, but this identification awaits systematic confirmation.
Origin and phylogeny of the Neolepadidae.—Neolepadids have generally been identified as primitive scalpellomorphs, in which only a single lateral plate is present, and have been placed within the family Eolepadidae (Yamaguchi et al. 2004; see above) close to the base of the scalpellomorph clade. This scenario was supported by the discovery of a supposed neolepadid in the Lower Jurassic (Buckeridge and Grant-Mackie 1985). The discovery that Eolepas itself possessed a phosphatic shell (Høeg et al. 1999; Gale and Schweigert 2015) and thus represents a significantly more basal thoracican group (Phosphaticothoracica Gale, 2019b), means that the calcite-shelled Neolepadidae require assignment to the crown group Thoracicalcarea (Gale 2015b). They were placed in the clade Unilatera by Gale (2019a), a group characterised by possession (plesiomorphically) of a single lateral plate, the upper latus. Morphologically, the neolepadids are remarkably similar to the Cretaceous genera Pycnolepas and Pedupycnolepas, in that they all possess an 8-plated capitulum and a large, externally positioned rostrum (Gale 2014b: fig. 1D, E). The morphology of individual plates of these genera is also similar to that of neolepadid taxa (e.g., scuta; Fig. 5A, B). Additionally, a distinctive, evenly-spaced, terraced sculpture, present on the valves of unilaterans, including verrucids, Pycnolepas and Pedupycnolepas (Gale 2019b: fig. 11), is also found in Ashinkailepas (Fig. 5A–H), but not present in other neolepadids. The Upper Jurassic Etcheslepas durotrigensis (Gale 2014a, 2019a) shares this sculpture type and tall upper latus, but possesses numerous lateral plates (Fig. 5I–K).

Fig. 5. Comparative morphology of neolepadid cirripede Ashinkailepas indica Gale sp. nov. (A, D, G) from Late Pleistocene, Krishna-Godavari Basin, off shore India, and other cirripedes. A. NHMUK IC 1412, paratype, scutum. D. NHMUK IC 1410, paratype, tergum. G. NHMUK IC 1404, paratype, upper latus. B, C, F. Brachylepadid Pycnolepas rígida (J. de C. Sowerby, 1836) from Gault Clay, Upper Albian (Lower Cretaceous), Naccolt, Kent, UK (based on B, Gale 2014b: fig. 4S; C, Gale 2014b: fig. 4Q; F, Gale 2014b: fig. 4R). B. NHMUK IC 1034, scutum. C. NHMUK IC 1032, tergum. F. NHMUK IC 1033, upper latus. E. Brachylepadid Faxelepas bruennichi (Withers, 1914) from Paleocene, Danian, Faxe, Denmark (based on Gale 2014b: fig. 6A). NHMUK IC 1019, scutum. H. Brachylepadid Pedupycnolepas pulcher Gale, 2019 from Endemoceras amblygonium Zone, Hauterivian (Lower Cretaceous), Engelbostel, near Hannover, Germany (based on Gale 2019b: fig. 11B). NHMUK IC 1397, holotype, tergum. I–K. Zeugmatolepadid capitula of Etcheslepas durotrigensis Gale, 2014 from Pectinatites pectinatus Zone, Tithonian (Upper Jurassic), Freshwater Steps, Kimmeridge, Dorset, UK. I, J. MIJML coll. unnumbered (based on Gale 2019a: fig. 2C, D). K. MIJML K1261, holotype (based on Gale 2014a: fig. 1). All in external view. Abbreviations: r, rostrum; s, scutum; rl, rostrolatus; ul, upper latus.
Two molecular studies (based on cytochrome c oxidase subunit I and two nuclear markers histone H3 gene and the ribosomal large subunit 28S [639 bp in COI, 342 bp in 12S, 1782 bp in 18S, 328 bp in H3]; Lin et al. 2015; Herrera et al. 2015) placed the Neolepadidae as sister group to the Scalpellidae. The large number of taxa studied by Herrera et al. (2015: fig. 2) demonstrate a nested relationship within the neolepadids, with Ashinkailepas as basal to the other genera, and Neolepas, Leucolepas, and Vulcanolepas as more derived. Chan and Chang (2018: fig. 7, using the COI gene, cytochrome c oxidase subunit I) obtained a broadly similar pattern, with Ashinkailepas as the most basal genus of the family, and various species of Vulcanolepas and Neolepas as derived forms. Additionally, Vulcanolepas and Neolepas appear to be paraphyletic.
Here we provide a cladistic analysis of the morphology of neolepadids, including the Lower Cretaceous Pedupycnolepas and the Upper Jurassic Etcheslepas, based upon 16 characters (Appendix 1). Heuristic analysis, optimised to deltran, yields a consensus tree, using Etcheslepas as an outgroup (Fig. 6). This demonstrates: (i) Pedupycnolepas is sister taxon to the Neolepadidae, (ii) Ashinkailepas is basal to all other neolepadids, (iii) The relationships between Vulcanolepas, Stipilepas, Leucolepas, and Neolepas is an unresolved polytomy.
The topology of this tree (Fig. 7) is congruent with those provided by various molecular studies (e.g., Herrera et al. 2015: fig. 2; Chan and Cheng 2018: fig. 7; Chan et al. 2019: fig. 8), which consistently show Ashinkailepas as basal to all other neolepadids, and the polyphyly of species ascribed variously to Vulcanolepas and Neolepas. It seems probable that molecular information is needed to understand the true affinities and relationships of species ascribed to these two genera.
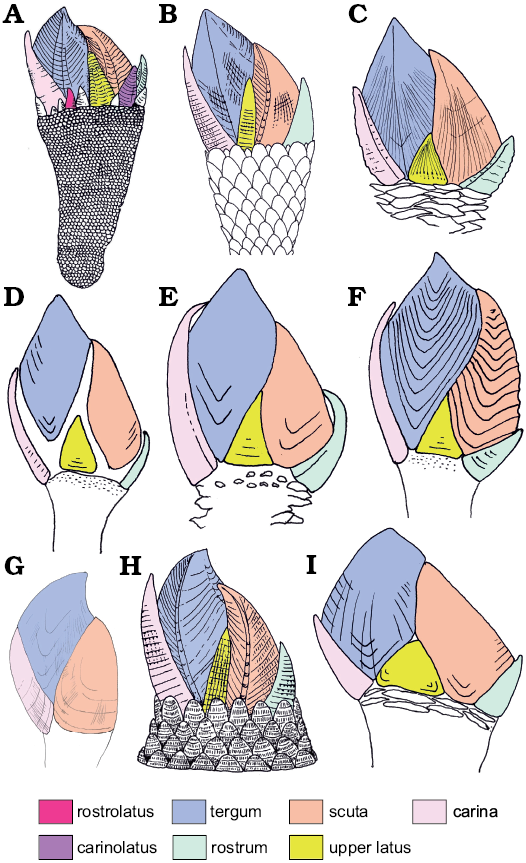
Fig. 6. Comparative morphology (capitula in lateral view) of neolepadids and other genera of pedunculated thoracicans. A. Etcheslepas durotrigensis Gale, 2014; Tithonian (Upper Jurassic), UK. B. Pedupycnolepas articulata (Collins, 1980); Aptian (Lower Cretacous), Antarctica. C. Ashinkailepas seepiophila Yamaguchi, Newman, and Hashimoto, 2004; Recent, Japan. D. Leucolepas longa Southward and Jones, 2003; Recent, Pacific. E. Neolepas zevinae Newman, 1979; Recent, Pacific. F. Vulcanolepas osheai Buckeridge, 2000; Recent, New Zealand. G. Litholepas klausreschi Nagler, Haug, Glenner, and Buckeridge, 2017; Tithonian (Upper Jurassic), Germany. H. Pycnolepas rigida (Sowerby, 1836); Albian (Lower Cretaceous), UK. I. Stipilepas molerensis Carriol in Carriol et al., 2016; Eocene of Denmark.
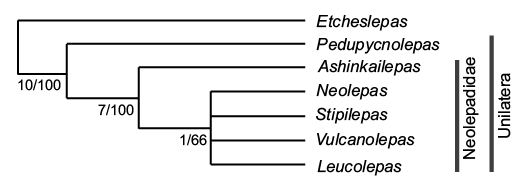
Fig. 7. Cladogram of relationships between Neolepadidae, Pedupycnolepas and Etcheslepas, using heuristic unconstrained analysis optimised to deltran and based on characters listed in Table 1. Values are for Bremer support/bootstrap.
Although the conclusions from morphological and molecular trees as to relationships within the Neolepadidae are consistent, the position of taxa inferred to be basal to the family from molecular data conflicts with that derived from morphology (Herrera et al. 2015: fig. 2). In their tree, Brachylepadidae (Neobrachylepas relica Newman and Yamaguchi, 1995) and Neoverricidae (Neoverruca brachylepadoformis Newman, 1979, Neoverruca sp.) are positioned between the Scalpellidae and Neolepadidae. This is inherently improbable, because N. relica is a basal sessile form which lacks a peduncle, and the tree of Herrera et al. (2015: fig. 2) infers that the peduncle was first lost in brachylepadids, then re-acquired by neolepadids. The relationships of thoracicans derived from molecular data are very inconsistent between different studies, and disagree profoundly with morphological data; for example, the tree of Herrera et al. (2015: fig. 2) places the pedunculate Pollicipedidae (Capitulum, Pollicipes) and Lithotryidae within the balanomorph clade, whereas the tree of Lin et al. (2015: fig. 3) places Lithotrya and Pollicipes in a group with Calanticidae, and Capitulum as basal to all other Thoracicalcarea.
From a morphological viewpoint, the best available evidence from fossil and extant taxa suggests a Late Jurassic split between the ubiquitously pedunculate (paraphyletic) Calamida and the Unilatera, characterised by possession of a single lateral, the upper latus (Gale 2019a: fig. 7). The latter group includes all the sessile thoracicans.
Conclusions
A new species of Ashinkailepas, A. indica Gale sp. nov. is described from a Late Pleistocene methane seep of the Krishna-Godavari Basin, offshore India. The Neolepadidae are raised to full family status, and removed from the phosphatic-shelled Eolepadidae, because they have calcite shells and are derived thoracicalcareans assigned to the Unilatera Gale, 2019a. Capitular morphology, including the presence of 8 plates (paired scuta, terga, upper latera, carina, rostrum), the shape and sculpture (regular terracing) of the plates, indicates a close relationship with the Cretaceous cirripede Pedupycnolepas (Gale 2014b). It is likely that the Neolepadidae were derived from a taxon close to Pedupycnolepas in the Late Cretaceous, during the major thoracican radiation which took place from the Albian to the Campanian (Gale 2020). Records of Jurassic Neolepadidae are here shown to be incorrect, and the oldest record of the family is Stipilepas Carriol in Carriol et al., 2016 from the Eocene of Denmark. Stipilepas is not associated with hydrothermal vents or methane seeps, and it is probable that this specialised habitat preference developed during the later Cenozoic. Thus, neolepadids are not relic primitive taxa providing “a glimpse of barnacle antiquity” (Newman 1985; Yamaguchi et al. 2004) which survived only in association with specialised vent (and methane seep) environments.
Acknowledgements
We would like to thank Claire Mellish (NHMUK) for assistance with registration of the material in the NHMUK. We would like to thank John Buckeridge (RMIT, Melbourne, Australia) and an anonymous reviewer for their most useful comments. We greatly appreciate the efforts of the NGHP-01 shipboard scientific party for the collection and initial interpretation of these cores. The authors wish to thank those that contributed to the success of the National Gas Hydrate Program Expedition 01 (NGHP-01). NGHP-01 was planned and managed through collaboration between the Directorate General of Hydrocarbons (DGH) under the Ministry of Petroleum and Natural Gas (India), the US Geological Survey (USGS), and the Consortium for Scientific Methane Hydrate Investigations (CSMHI) led by Overseas Drilling Limited (ODL) and FUGRO McClelland Marine Geosciences (FUGRO). The platform for the drilling operation was the research drill ship JOIDES Resolution, operated by ODL. Much of the drilling/coring equipment used was provided by the Integrated Ocean Drilling Program (IODP) through a loan agreement with the US National Science Foundation. Wireline pressure coring systems and supporting laboratories were provided by IODP/Texas A&M University (TAMU), FUGRO, USGS, US Department of Energy (USDOE) and HYACINTH/GeoTek. Downhole logging operational and technical support was provided by Lamont-Doherty Earth Observatory (LDEO) of Columbia University. The financial support for the NGHP-01, from the Oil Industry Development Board, Oil and Natural Gas Corporation Ltd., GAIL (India) Ltd. and Oil India Ltd. is gratefully acknowledged. We also acknowledge the support extended by all the participating organizations of the NGHP: MoP&NG, DGH, ONGC, GAIL, OIL, NIO, NIOT, and RIL.
References
Bromley, R.G. 1981. Concepts in ichnotaxonomy illustrated by small round holes in shells. Acta Geologica Hispanica 16: 55–64.
Buckeridge, J.S. 1983. The fossil barnacles (Cirripedia: Thoracica) of New Zealand and Australia. New Zealand Geological Survey Paleontological Bulletin 50: 1–152.
Buckeridge, J.S. 2000. Neolepas osheai sp. nov., a new deep-sea vent barnacle (Cirripedia: Pedunculata) from the Brothers Caldera, south-west Pacific Ocean. New Zealand Journal of Marine and Freshwater Research 34: 409–418. Crossref
Buckeridge, J.S. 2009. Ashinkailepas kermadecensis, a new species of deep-sea scalpelliform barnacle (Thoracica: Eolepadidae) from the Kermadec Islands, south-west Pacific. Zootaxa 2021: 57–65. Crossref
Buckeridge, J.S. and Grant-Mackie, J.A. 1985. A new scalpellid barnacle from the lower Jurassic of New Caledonia. Géologie de la France 1: 77–80.
Burmeister, H.B. 1834. The Organisation of Trilobites Deduced from Their Living Affinities and a Systematic Review of the Species Hitherto Described. 128 pp. Ray Society, London.
Carriol, R.-P., Bonde, N., Jakobsen, S.L., and Hoeg, J.T. 2016. New stalked and sessile cirripedes from the Eocene Mo Clay, northwest Jutland (Denmark). Geodiversitas 38: 21–32. Crossref
Chan, B.K.K. and Chang, Y.W. 2018. A new deep-sea scalpelliform barnacle, Vulcanolepas buckeridgei sp. nov. (Eolepadidae: Neolepadinae) from hydrothermal vents in the Lau Basin. Zootaxa 4407: 117–129. Crossref
Chan, B.K.K., Ju, S.-J. and Kim, S.-J. 2019. A new species of hydrothermal vent stalked barnacle Vulcanolepas (Scalpelliformes: Eolepadidae) from the North Fiji Basin, Southwestern Pacific Ocean. Zootaxa 4563: 135–148. Crossref
Collett, T.S., Boswell, R., Cochran, J.R., Kumar, P., Lall, M., Mazumdar, A., Ramana, M.V., Ramprasad, T., Rieldel, M., Sain, K., Sathe, A.V., Cishwanath, K., and the NGHP Expedition 01 Scientific Party 2014. Geologic implications of gas hydrates in the offshore of India: results of the National Gas Hydrate Program Expedition 01. Journal of Marine and Petroleum Geology 58: 3–28. Crossref
Collett, T., Riedel, M., Cochran, J., Boswell, R., Presley, J., Kumar, P., Sathe, A., Sethi, A., Lall, M., and the NGHP Expedition Scientists 2015. Indian National Gas Hydrate Program Expedition 01 report. U.S. Geological Survey Scientific Investigations Report 2012-5054: 1442. Crossref
Connolly, E.C., Johnson, J.E., Torres, M.E., Teichert, B.M., Giosan, L., and Rose, K. 2009. New insights from sediment ages and carbon isotopes at a paleo-seep/chemosynthetic biological community in the Krishna-Godavari Basin, offshore India. Eos Transactions American Geophysical Union 90 (52), Fall Meeting Supplementary, Abstract OS31A-1188.
Connolly, E.C., Johnson, J.E., Torres, M.E., Teichert, B.M., Giosan, L. and Rose, K., 2009. New Insights from Sediment Ages and Carbon Isotopes at a Paleo-Seep/Chemosynthetic Biological Community in the Krishna-Godavari Basin, Offshore India. Eos Transactions AGU, 90 (52), Fall Meeting Supplementary, Abstract OS31A-1188
Darwin, C. 1854. A Monograph on the Subclass Cirripedia, With Figures of All the Species. 684 pp. Ray Society, London.
Dubilier, N., Bergin, C., and Lott, C. 2008. Symbiotic diversity in marine animals: the art of harnessing chemosynthesis. Nature Reviews Microbiology 6: 725–740. Crossref
Flores, J.A., Johnson, J.E., Mejía-Molina, A.E., Álvarez, M.C., Sierro, F.J., Singh, S.D., Mahanti, S., and Giosan, L. 2014. Sedimentation rates from calcareous nannofossil and planktonic foraminifera biostratigraphy in the Andaman Sea, northern Bay of Bengal, and eastern Arabian Sea. Marine and Petroleum Geology 58: 425–437. Crossref
Gale, A.S. 2014a. New thoracican cirripedes (Crustacea) from the Jurassic and Cretaceous of the UK. Proceedings of the Geologists’ Association 125: 406–418. Crossref
Gale, A.S. 2014b. Origin and phylogeny of the verrucomorph cirripedes (Crustacea; Thoracica). Journal of Systematic Palaeontology 13: 753–789. Crossref
Gale, A.S. 2015a. Origin and phylogeny of the thoracican cirripede family Stramentidae. Journal of Systematic Palaeontology 14: 653–702. Crossref
Gale, A.S. 2015b. Phylogeny of the deep sea cirripede family Scalpellidae based on capitular plate morphology. Zoological Journal of the Linnean Society 176: 1–34. Crossref
Gale, A.S. 2019a. Stalked barnacles (Cirripedia, Thoracica) from the Late Jurassic (Tithonian) Kimmeridge Clay of Dorset, UK; palaeoecology and bearing on the evolution of living forms. Proceedings of the Geologists’ Association 130: 355–365. Crossref
Gale, A.S. 2019b. Thoracican cirripedes (Crustacea) from the Hauterivian (Lower Cretaceous) of Hannover, northern Germany. Proceedings of the Geologists’ Association 130: 659–672. Crossref
Gale, A.S. 2020. New thoracican cirripedes (Crustacea) from the Cretaceous of Europe and north Africa. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 295: 243–282. Crossref
Gale, A.S. and Schweigert, G. 2015. A new phosphatic-shelled cirripede (Crustacea, Thoracica) from the Lower Jurassic (Toarcian) of Germany—the oldest epiplanktonic barnacle. Palaeontology 59: 59–70. Crossref
Gale, A.S. and Sørensen, A.M. 2015. Taxonomy and palaeoecology of thoracican cirripedes (Crustacea) from a Campanian rocky shoreline at Ivö Klack, southern Sweden. Cretaceous Research 54: 212–242. Crossref
Gale, A.S., Schweigert, G., Roper, M., and Keupp, R.A. 2019. A cirripede (Crustacea) from the Tithonian (Jurassic) of southern Germany. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 293: 1–17. Crossref
Herrera, S., Watanabe, H., and Shank, T.M. 2015. Evolutionary and biogeographical patterns of barnacles from deep-sea hydrothermal vents. Molecular Ecology 24: 673–689. Crossref
Høeg, J.T., Whyte, M.A., Glenner, H., and Schram, F.R. 1999. New evidence on the basic phylogeny of the Cirripedia Thoracica. In: F.R. Schram and J.C. von Vaupel Klein (eds.), Crustaceans and the Biodiversity Crisis. Proceedings of the Fourth International Crustacean Congress, Amsterdam, The Netherlands, July 20–24, 1998, Vol. 1, 101–114. Brill, Leiden.
Johnson, J.E., Phillips, S.C., Torres, M.E., Piñero, E., Rose, K.K., and Giosan, L. 2014. Influence of total organic carbon deposition on the inventory of gas hydrate in the Indian continental margins, Marine and Petroleum Geology 58: 406–424. Crossref
Klompmaker, A.A., Portell, R.W., Lad, S.E., and Kowalewski, M. 2015. The fossil record of drilling predation on barnacles. Palaeogeography, Palaeoclimatology, Palaeoecology 426: 95–111. Crossref
Levin, L.A. 2005. Ecology of cold seep sediments: interactions of fauna with flow, chemistry and microbes. In: R.N. Gibson, R.J.A. Atkinson, and J.D.M. Gordon (eds.), Oceanography and Marine Biology: An Annual Review 43, 1–46. CRC Press-Taylor & Francis Group, Boca Raton. Crossref
Lin, H.-C., Høeg, J.T., Yusa, Y., and Chan, B.K. 2015. The origin and evolution of dwarf males and habitat use in thoracican barnacles. Molecular Phylogenetics and Evolution 91: 1–11. Crossref
Lorenson, T.D. and Collett, T.S. 2018. National Gas Hydrate Program Expedition 01 offshore India; gas hydrate systems as revealed by hydrocarbon gas geochemistry. Marine and Petroleum Geology 92: 477–492. Crossref
Mazumdar, A., Dewangan, P., Joäo, H.M., Peketi, A., Khosla, V., Kocherla, M., Badesab, F.K., Joshi, R.K., Roxanne, P., Ramamurty, P.B., Karisiddaiah, S.M., Patil, D.J., Dayal, A.M., Ramprasad, T., Hawkesworth, C.J., and Avanzenelli, R. 2009. Evidence of paleo-cold seep activity from the Bay of Bengal, offshore India. Geochemistry Geophysics Geosystems 10: Q06005. Crossref
Mazumdar, A., Dewangan, P., Peketi, A., Gullapalli, S., Kalpana, M.S., Naik, G.P., Shetty, D., Pujari, S., Pillutla, S.P.K., Gaikwad, V.V., Nazareth, D., Sangodkar, N.S., Dakara, G., Kumar, A., Mishra, C.K., Singha, P., and Reddy, R. 2019. The first record of active methane (cold) seep ecosystem associated with shallow methane hydrate from the Indian EEZ. Journal of Earth Systems Science 128: 18. Crossref
Nagler, C., Haug, J.T., Glenner, H., and Buckeridge, J. 2017. Litholepas klausreschi gen. et sp. nov. (Cirripedia, Thoracica) a new neolepadinine barnacle on a sponge from the Upper Jurassic lithographic limestones of southern Germany. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 284: 29–42. Crossref
Newman, W.A. 1979. A new scalpellid (Cirripedia); a Mesozoic relic living near an abyssal hydrothermal spring. Transactions of the San Diego Society of Natural History 19: 153–167.
Newman, W.A. 1985. The abyssal hydrothermal vent invertebrate fauna: a glimpse of antiquity? In: M.L. Jones (ed.), The Hydrothermal Vents of the Eastern Pacific: An Overview. Bulletin of the Biological Society of Washington 6: 231–242.
Newman, W.A., and Yamaguchi, T. 1995. A new sessile barnacle (Cirripedia: Brachylepadomorpha) from the Lau Back-Arc Basin, Tonga: first record of a living representative since the Miocene. Bulletin de la Muséum d’Histoire naturelle, Paris 4 (17): 221–243.
Phillips, S.C., Johnson, J.E., Underwood, M.B., Guo, J., Giosan, L., and Rose, K. 2014. Long-timescale variation in bulk and clay mineral composition of Indian continental margin sediments in the Bay of Bengal, Arabian Sea, and Andaman Sea. Marine and Petroleum Geology 58: 117–138. Crossref
Southward, A.J. and Jones, D.S. 2003. A revision of stalked barnacles (Cirripedia: Thoracica: Scalpellomorpha: Eolepadidae: Neolepadinae) associated with hydrothermalism, including a description of a new genus and species from a volcanic seamount off Papua New Guinea. Senckenbergiana Maritima 32: 77–93. Crossref
Southward, A.J. and Newman, W.A. 1998. Ectosymbiosis between filamentous sulphur bacteria and a stalked barnacle (Scalpellomorpha, Neolepadinae) from the Lau Back Arc Basin, Tonga. Cahiers de Biologie Marine 39: 259–262.
Sowerby, J. de C. 1836. Appendix A: Descriptive notes respecting shells figured in plates 11–22. In: W.H. Fitton (eds.), Observations on Some of the Strata Between the Chalk and the Oxford Oolite in the South-East of England. Transactions of the Geological Society of London, Second Series 4 (2): 103–388. Crossref
Teichert, B.M.A., Johnson, J.E., Solomon, E.A., Giosan, L., Rose, K., Kocherla, M., Connolly, E.C., and Torres, M.E. 2014. Composition and origin of authigenic carbonates in the Krishna-Godavari and Mahanadi Basins, eastern continental margin of India. Marine and Petroleum Geology 58: 438–460. Crossref
Watanabe, H.K., Chen, C., Marie, D.P., Takai, K., Fujikura, K., and Chan, B.K.K. 2018. Phylogeography of hydrothermal vent stalked barnacles: a new species fills a gap in the Indian Ocean “dispersal corridor” hypothesis. Royal Society Open Science 5: 172408. Crossref
Withers, T.H. 1928. British Museum (Natural History). Catalogue of the Fossil Cirripedia in the Department of Geology Vol. 1. Triassic and Jurassic. 154 pp. Dulau & Co., London.
Yamaguchi, T., Newman, W.A., and Hashimoto, J. 2004. A cold seep barnacle (Cirripedia: Neolepadinae) from Japan and the age of the vent/seep fauna. Journal of the Marine Biological Association of the United Kingdom 84: 111–120. Crossref
Character list of taxa in cladistics analysis
Lateral plates numerous (0); single lateral plate (upper latus)
Carina overlaps tergum (0); articulates with lower carinal margin of tergum (1)
Upper latus 2–3 times taller than broad (0); as tall as broad (1)
Scutum overlaps tergum (0); tergum overlaps scutum (1)
Rostrum overlaps base of scutum (0); rostrum articulates with basal margin of scutum (1)
Upper latus overlaps scutum and tergum (0); upper latus in V-shaped space between scutum and tergum (1)
Peduncular plates possess interiorly-directed basal shelf (0); shelf absent (1)
Peduncular plates large (0); small (1)
Peduncular plates bear spines (0) lack spines (1)
Apicobasal ridge present on scutum and tergum (0); ridge absent (1)
Valves bear evenly spaced, terraced sculpture (0); only growth lines on valves (1)
Radial striae present on tergum and scutum (0); striae absent (1)
Apex of tergum inclined ventrally (0); inclined dorsally (1)
Raised margin present on rostrum (0); raised margin absent (1)
Rostrum possesses raised central ridge (0); central ridge absent (1)
Subcarina present (0); absent (1)
Character matrix
| |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
|
Etcheslepas |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Pedupycnolepas |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0 |
0 |
1 |
0 |
0 |
1 |
|
Ashinkailepas |
1 |
1 |
1 |
0 |
1 |
1 |
1 |
1 |
1 |
0 |
0 |
0 |
1 |
1 |
1 |
1 |
|
Leucolepas |
1 |
1 |
1 |
? |
1 |
1 |
1 |
1 |
1 |
0 |
1 |
1 |
0 |
1 |
1 |
1 |
|
Stipilepas |
1 |
1 |
1 |
? |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
0 |
1 |
1 |
1 |
|
Vulcanolepas |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
0 |
1 |
1 |
0 |
1 |
1 |
1 |
|
Neolepas |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
0 |
1 |
1 |
0 |
1 |
1 |
1 |
Acta Palaeontol. Pol. 65 (2): 351–362, 2020
https://doi.org/10.4202/app.00705.2019