
Tracking Late Jurassic ornithopods in the Lusitanian Basin of Portugal: Ichnotaxonomic implications
DIEGO CASTANERA, BRUNO C. SILVA, VANDA F. SANTOS, ELISABETE MALAFAIA, and MATTEO BELVEDERE
Castanera, D., Silva, B.C., Santos, V.F., Malafaia, E., and Belvedere, M. 2020. Tracking Late Jurassic ornithopods in the Lusitanian Basin of Portugal: Ichnotaxonomic implications. Acta Palaeontologica Polonica 65 (2): 399–412.
The Sociedade de História Natural in Torres Vedras, Portugal houses an extensive collection of as yet undescribed dinosaur tracks with ornithopod affinities. They have been collected from different Late Jurassic (Kimmeridgian–Tithonian) geological formations (Praia de Amoreira-Porto Novo, Alcobaça, Sobral, and Freixial) that outcrop along the Portuguese coast, and belong to two different sub-basins of the Lusitanian Basin (the Consolação and Turcifal sub-basins). Three main morphotypes can be distinguished on the basis of size, mesaxony and the morphology of the metatarsophalangeal pad impression. The minute to small-sized morphotype is similar to the Anomoepus-like tracks identified in other Late Jurassic areas. The small to medium-sized morphotype resembles the Late Jurassic–Early Cretaceous ichnotaxon Dinehichnus, already known in the Lusitanian Basin. Interestingly, these two morphotypes can be distinguished qualitatively (slightly different size, metatarsophalangeal pad impression and digit morphology) but are nevertheless difficult to discriminate by quantitatively analysing their length-width ratio and mesaxony. The third morphotype is considered a large ornithopod footprint belonging to the ichnofamily Iguanodontipodidae. This ichnofamily is typical for Cretaceous tracksites but the new material suggests that it might also be present in the Late Jurassic. The three morphotypes show a negative correlation between size and mesaxony, so the smaller tracks show the stronger mesaxony, and the larger ones weaker mesaxony. The Upper Jurassic ornithopod record from the Lusitanian Basin has yielded both small and medium-sized ornithopod remains, mainly iguanodontians such as dryosaurids and ankylopollexians, which are the main candidates to be the trackmakers.
Key words: Iguanodontipodidae, Dinehichnus, Anomoepus-like, Kimmeridgian, Tithonian, Europe.
Diego Castanera [diego.castanera@icp.cat; dcastanera@hotmail.es], Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, c/ Escola Industrial 23, 08201 Sabadell, Barcelona, Spain; Bayerische Staatssammlung für Paläontologie und Geologie and GeoBioCenter, Ludwig-Maximilians-Universität, Richard-Wagner-Str. 10, 80333 Múnich, Germany.
Bruno C. Silva [laboratorio@alt-shn.org], Laboratório de Paleontologia e Paleoecologia, Sociedade de História Natural, Apartado 2564-909 Torres Vedras, Portugal; European Centre of Paleontology, Institute of Biology, Laboratory of Paleobiology, University of Opole, ul. Oleska 48, 45-052 Opole, Poland.
Vanda F. Santos [vafsantos@fc.ul.pt], Departamento de Geologia, Faculdade de Ciências da Universidade de Lisboa, Edifício C6, Piso 3, Campo Grande, 1749-016 Lisboa, Portugal; Instituto Dom Luiz (IDL), Faculdade de Ciências da Universidade de Lisboa, Campo Grande, Lisboa, Portugal.
Elisabete Malafaia [emalafaia@gmail.com], Instituto Dom Luiz (IDL), Faculdade de Ciências da Universidade de Lisboa, Campo Grande, Lisboa, Portugal; Grupo de Biología Evolutiva. Facultad de Ciencias. Universidad Nacional de Educación a Distancia. C/ Senda del Rey, 9, 28040 Madrid, Spain.
Matteo Belvedere [matteo.belvedere@hotmail.com]; Institute for Studies in Landscape and Human Evolution, Faculty of Science and Technology, Bournemouth University, Talbot Campus, Fern Barrow, Poole, BH12 5BB, UK.
Received 27 November 2019, accepted 3 February 2020, available online 15 April 2020.
Copyright © 2020 D. Castanera et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Small to medium-sized tridactyl tracks assigned to ornithopods are scarce in the Late Jurassic European record compared with other types of tridactyl dinosaur footprints such as those of theropods, which are quite abundant in areas such as the Jura platform (Marty et al. 2018; Mazin et al. 2017; Castanera et al. 2018), the Lusitanian Basin (Lockley and Santos 1993; Lockley et al. 1994; Mateus and Milàn 2010) or the Asturian Basin (Lockley et al. 2008; Piñuela Suarez 2015). This scarcity of ornithopod tracks is a consequence of an abundance of sauropod- and theropod-dominated ichnofaunas preserved in carbonate facies in the so-called Brontopodus ichnofacies, which are characterized by a rather low ichnodiversity (Hunt and Lucas 2007). On the other hand, siliciclastic units have provided higher ichnodiversity, and two or possibly three different types of ornithopod tracks have been described in the Late Jurassic of the Morrison Formation in the USA and the Asturian Basin (Spain), respectively (Lockley et al. 2008). Furthermore, in the last few years new ornithopod tracks have been described in several Late Jurassic units belonging to the Maestrazgo and Asturian basins in Spain (e.g., Castanera et al. 2013b; Alcalá et al. 2014; Piñuela et al. 2016). Another factor that may explain the apparent scarcity of ornithopod tracks in the Late Jurassic is the similarity in footprint morphology of theropod and ornithopod tracks in Late Jurassic times (e.g., Lockley et al. 2008; Castanera et al. 2013b).
The classic ornithopod ichnotaxon during the Late Jurassic is Dinehichnus Lockley, dos Santos, Meyer, and Hunt, 1998, which has been described in North America, Europe, Africa, and possibly Asia (Lockley et al. 1998; Gierliński et al. 2009; Belvedere et al. 2010; Xing et al. 2014). This ichnotaxon has also been identified in the Lower Cretaceous in Europe (Lockley et al. 1998; Hornung and Reich 2012). In addition, Anomoepus-like tracks have been identified in the Late Jurassic (Lockley et al. 2008). However, large ornithopod tracks (generally those with a footprint length > 25 cm, Thulborn 1990; see also Moreno et al. 2012 and Díaz-Martínez et al. 2015 for a wider definition) from the Late Jurassic are rare, and only a few examples have been described (Mateus and Milàn 2008; Diedrich 2011; Schulp and Al-Wosabi 2012; Piñuela et al. 2016). It is noteworthy that some of these examples come from the Iberian Peninsula.
Although significant Late Jurassic tracksites have been described in the Lusitanian Basin, few ornithopod tracks have been identified so far (Lockley et al. 1998; Antunes and Mateus 2003; Santos 2008; Mateus and Milàn 2008). Of all these records, only a slab from São Martinho do Porto has yielded tracks assigned to Dinehichnus (Lockley et al. 1998), but this is now lost because of the erosion of the cliff. By contrast, the Late Jurassic ornithopod bone record from the Lusitanian Basin has yielded a sizeable, rich ornithopod fauna (Galton 1980; Rauhut 2001; Mateus and Antunes 2001; Antunes and Mateus 2003; Escaso et al. 2014; Rotatori et al. 2020).
The Sociedade de História Natural in Torres Vedras, Portugal houses an extensive collection of as yet undescribed tridactyl ornithopod-like dinosaur tracks, which considerably extend our knowledge of the morphological features of ornithopod tracks in the Late Jurassic. The aim of this paper is to describe this new material and discuss the tracks’ ichnotaxonomic affinities, emphasizing the implications for the ornithopod ichnodiversity during the Late Jurassic and discussing the morphological variations among the morphotypes.
Institutional abbreviations.—SHN.(JJS).ICNO, Sociedade de História Natural, Torres Vedras, Portugal.
Other abbreviations.—AT, mesaxony; ATl/w, anterior triangle length/width; FL, footprint length; FW, footprint width; HA, “heel” (metatarsophalangeal) area; L/W II–IV, length/width of digits II–IV; MP, morphological preservation.
Geographical and geological setting
The studied footprints were collected from different outcrops in the cliffs and beaches along the central-west coastline of Portugal. The footprints were found in various localities in the municipalities of Alcobaça (São Martinho do Porto), Caldas da Rainha (Salir do Porto, Boavista do Bouro, Serra do Bouro, and Foz do Arelho), Peniche, Lourinhã, and Torres Vedras (see Fig. 1 and SOM: table S1 in Supplementary Online Material available at http://app.pan.pl/SOM/app65-Castanera_etal_SOM.pdf).
These outcrops are located in the central sector of the Lusitanian Basin. This basin was developed during the Mesozoic and is positioned in what is nowadays the west region of the Iberian Peninsula. During the Mesozoic evolution of the Lusitanian Basin, several sub-basins were formed as a consequence of the rifting linked to the opening of the North Atlantic Ocean. The basin was infilled by sediments from the Upper Triassic to the Upper Cretaceous (Kullberg et al. 2013; Taylor et al. 2014 and references therein). The localities where the footprints were found are situated in the Consolação and Turcifal sub-basins (sensu Taylor et al. 2014). Since the lithostratigraphic framework of the infilling of these sub-basins is complex due to synsedimentary tectonic activity and the general scarcity of reliable biostratigraphic markers (but see Schneider et al. 2009), different lithostratigraphic schemes have been proposed. Here we follow the one proposed by Manuppella et al. (1999) (see Fig. 1) because this is the nomenclature generally used by the Serviços Geológicos de Portugal (Portuguese Geological Survey) in stratigraphic studies of Upper Jurassic levels from Portugal, specifically in the most recent geological maps.
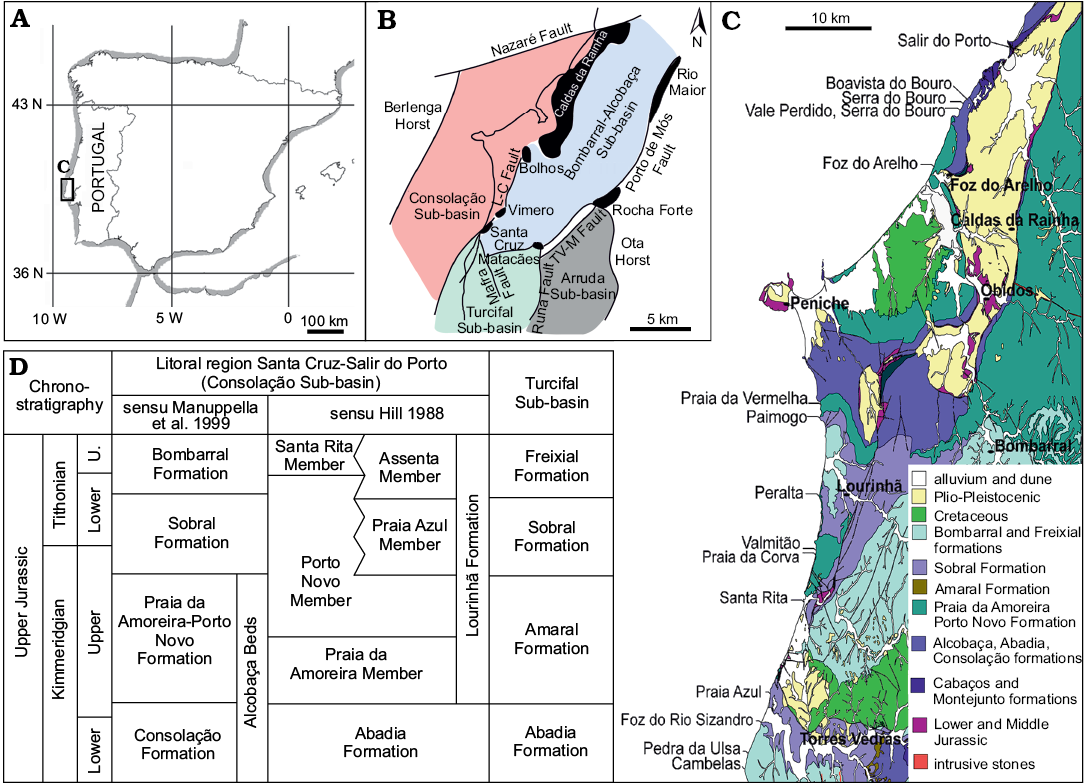
Fig. 1. Geographical and geological setting of the outcrops where the studied footprints were collected. A. Map of the Iberian Peninsula showing the location of the Lusitanian Basin. B. Structural setting of the Late Jurassic sub-basins in the western and central sector of the Lusitanian Basin (modified after Taylor et al. 2014). C. Geological map of the western and central sector of the Lusitanian Basin showing the localities where the described specimens were collected (adapted from Oertel et al. 1960; Oliveira et al. 1992). D. Stratigraphic correlation of Upper Jurassic units of the Consolação and Turcifal sub-basins. Abbreviations: L-C, Caldas da Rainha; TV-M, Torres Vedras-Montejunto; U, Upper.
Within the Consolação sub-basin the specimens belong mainly to three different geological formations: the Praia da Amoreira-Porto Novo (upper Kimmeridgian), the Sobral (upper Kimmeridgian–lower Tithonian) and the Alcobaça (Kimmeridgian–lower Tithonian) formations. The Praia da Amoreira-Porto Novo Formation (upper Kimmeridgian; Manuppella et al. 1999) is composed of siliciclastic deposits, mainly sandstone and mudstone deposited in alluvial fans or meandering river systems (Praia da Amoreira Member; Hill 1989; Taylor et al. 2014), in its lower part. The overlying sediments (Porto Novo Member sensu Hill 1989) are also siliciclastic in nature, mainly sandstone channel bodies, often with lateral accretion surfaces and cross-bedded lamination plus floodplain mudstone deposits and well-developed levels of calcrete palaeosoils. These sediments were deposited in fluvial meander systems grading laterally to upper deltaic deposits (Mateus et al. 2013). The Sobral Formation (upper Kimmeridgian–lower Tithonian; Manuppella et al. 1999; Schneider et al. 2009) is mainly composed of siliciclastic deposits, silty or sandy marl, siltstone or sandstone, plus some calcareous levels. This formation was deposited in a deltaic estuarine system (Schneider et al. 2009; Kullberg et al. 2013). The Alcobaça Formation (mostly Kimmeridgian, but some authors suggest that it may extend to the lower Tithonian; Manuppella et al. 1999; Schneider et al. 2009; Kullberg et al. 2013) is composed of an alternation of sandy marl and calcareous limestone, marl and sandstone, deposited in a shallow carbonate platform with frequent siliciclastic inputs (Manuppella et al. 1999; Schneider et al. 2009; Kullberg et al. 2013). It is interesting to note that these three formations are partially equivalent in age although they outcrop in different areas of the Consolação sub-basin. The Praia da Amoreira-Porto Novo Formation is overlapped by the Sobral Formation, and both outcrop in the littoral region from Santa Cruz to Salir do Porto whereas the Alcobaça Formation outcrops in the Alcobaça-Caldas da Rainha region (see Fig. 1). Within the Turcifal sub-basin the specimens belong to the Sobral Formation (upper Kimmeridgian–lower Tithonian) and the overlapping Freixial Formation. The Freixial Formation is composed of an alternation of limestones, marls and siliciclastic material deposited in a shallow ramp. The age of this formation is middle–upper Tithonian (Schneider et al. 2009; Kullberg et al. 2013).
Material and methods
The tracks (SOM: table S1) are housed in the collection of the Sociedade de História Natural in Torres Vedras. The tracks are labelled as: SHN.(JJS).ICNO.#, # being the entry number in the collection. Some tracks have the same number, so a letter has been added to distinguish them (e.g., SHN.(JJS).ICNO.29A, SHN.(JJS).ICNO.29B). Additionally, m and p refer to manus and pes, respectively.
Most of the tracks are preserved as natural casts (convex hyporelief, H) although some true tracks (concave epireliefs, E) are also present (see SOM: table S1). Analysis of track morphology was performed independently for each track, by evaluating the morphological preservation (MP) according to the scale of Marchetti et al. (2019). A total of 57 individual tracks were analysed (SOM: table S1), but only those tracks that have an MP grade of 2 or higher (n = 18, see SOM: tables S1, S2) were used for proper systematic assignments (see Marchetti et al. 2019). The outlines of only these tracks are drawn and figured in the present paper. Those with a grade of less than 2 (n = 39, see SOM: table S3) are described but the systematic assignment is only an attempt to link these track morphologies to a putative ornithopod trackmaker. Open nomenclature follows the recommendations of Bengston (1998). Several parameters such as the footprint length (FL), footprint width (FW), the length and width of digits II (LII, WII), III (LIII, WIII), and IV (LIV, WIV), the “heel” (metatarsophalangeal) area (HA), and the divarication angles (II–III, III–IV) were measured (see SOM: fig. S1, tables S2, S3). Subsequently, the FL/FW ratio and the mesaxony (AT) were calculated. The latter was calculated on the basis of the anterior triangle length (ATl)/width (ATw) ratio following Lockley (2009). The tracks were classified according to different size classes (Marty 2008) on the basis of pes length (FL) as: (i) minute, FL < 10 cm; (ii) small, 10 cm < FL < 20 cm; (iii) medium, 20 cm < FL < 30 cm; and (iv) large, 30 cm < FL < 50 cm. Due to the symmetry of the tracks it was difficult to distinguish between digit II and IV. The criterion for distinguishing them was the presence of a small medial notch in the proximal part of digit II, which gives some of the tracks a certain asymmetry.
All the measurements were taken from the false-colour depth maps exported from the 3D-photogrammetric models and measured with the software ImageJ. The 3D models were generated from pictures taken with a Sony Alpha 5100, using Agisoft Photoscan (v. 1.3.2, www.agisoft.com) following the procedures of Mallison and Wings (2014) and Matthews et al. (2016). The scaled meshes were exported as Stanford PLY files (.ply) and then processed in CloudCompare (v.2.7.0) in order to obtain false-colour depth maps. All photogrammetric meshes used in this study are available for download in the SOM, following the recommendations of Falkingham et al. (2018). The morphometric data of the studied tracks were compared in a bivariate plot (length/width ratio vs. mesaxony) using the software PAST (Hammer et al. 2001). The data for mesaxony and the length-width ratio were taken from Lockley (2009) for the holotypes of Anomoepus, Neoanomoepus, Dinehichnus, and Iguanodontipus.
Systematic palaeoichnology
Ichnofamily Iguanodontipodidae Vialov, 1988 emended Díaz-Martínez et al. 2015
Diagnosis.—“Mesaxonic, tridactyl, subsymmetrical pes tracks that are as wide as or wider than long; one pad impression in each digit and one in the heel; digit pads longer than wide; well-developed notches in the proximal part of the digit II and IV impressions; manus tracks occasionally present and much smaller than the pes tracks” (Díaz-Martínez et al. 2015).
Iguanodontipodidae indet.
Fig. 2.
Material.—SHN.(JJS).ICNO.03, 48, 54, 75, 110; from Valmitão Norte, Valmitão Sul, Praia Azul, Foz Do Rio Sizandro; Amoreira-Porto Novo (upper Kimmeridgian) and Sobral (upper Kimmeridgian–lower Tithonian) formations (for details see SOM: table S1).
Description.—Medium to large-sized (pes length 28–32.5 cm) tridactyl tracks (Fig. 2 and SOM: table S2), generally slightly wider than long or almost as wide as long (length/width ratio 0.82–1.03) with low mesaxony (AT ratio 0.32–0.42). The tracks are relatively symmetrical, with a lateral and medial notch and a rounded to quadrangular metatarsophalangeal pad impression. The medial notch looks slightly more developed in some specimens (SHN.(JJS).ICNO.03, 75), so these tracks are slightly asymmetric. The digits are considerably robust. Digit III is longer than digits II and IV, digit IV being slightly longer than digit II. The hypices are fairly symmetrical. No discrete phalangeal pad impressions can be discerned, possibly suggesting one pad impression in each digit. The distal end of the digit impressions is rounded, possibly indicating blunt claw marks. Only SHN.(JJS).ICNO.03 shows evidence of blunt but acuminate claw marks. The interdigital angle II–IV is medium (65–85°), interdigital angle II–III being slightly greater than interdigital angle III–IV.
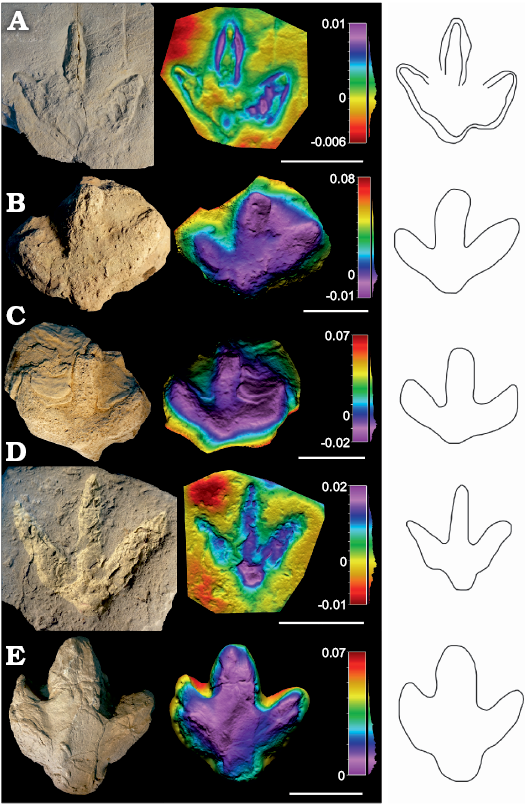
Fig. 2. Photographs, false-colour depth maps, and outline drawing of the tracks of high morphological quality (MP = 2) that belong to the Iguanodontipodidae indet. morphotype. Portugal, Praia de Amoreira-Porto Novo Formation, upper Kimmeridgian (A, B) and Sobral Formation, upper Kimmeridgian–lower Tithonian (C–E). A. SHN.(JJS).ICNO.03, Valmitão Norte. Note that the track still preserves part of the infill. B. SHN.(JJS).ICNO.54, Valmitão Sul. C. SHN.(JJS).ICNO.48, Praia Azul. D. SHN.(JJS).ICNO.75, Foz Do Rio Sizandro. Note that the track has been painted and the real morphology is not exactly the same as the drawing. E. SHN.(JJS).ICNO.110, Foz Do Rio Sizandro. Scale bars E, 15 cm; A–C, 20 cm; D, 25 cm.
Remarks.—Large ornithopod tracks, i.e., those larger than 25 cm and generally assigned to iguanodontians, are not very common during the Late Jurassic, although they are frequent subsequently during the Cretaceous (Lockley and Wright 2001; Lockley et al. 2014; Díaz-Martínez et al. 2015). On the basis of the metatarsophalangeal pad and digit impressions, the latter authors identified three main groups of large ornithopod tracks, which belong to the ichnofamily Iguanodontipodidae and are related to the ichnogenera Iguanodontipus, Caririchnium, and Hadrosauropodus. Interestingly, this ichnofamily is so far mainly restricted to the Cretaceous. The authors include some tracks from the literature in their so-called Group 1; these were included in Iguanodontipus. This ichnotaxon has been considered monospecific (I. burreyi), being characterized by a small, rounded metatarsophalangeal pad and elongate, narrow digit impressions (see also Castanera et al. 2013a), and its distribution is limited to the Berriasian–Valanginian of Europe. Recently, Piñuela et al. (2016) described medium to large-sized (28–30 cm) ornithopod tracks in the Late Jurassic of Asturias (Spain) and suggested that the tracks “are quite different from known ichnogenera, but they are not sufficiently well preserved to propose a new one”. Moreover, the authors questioned the validity of Iguanodontipus because of the preservation of the material and even called into question the ornithopod origin of the possible trackmaker. The tracks from the Lusitanian Basin studied here share some features with the tracks from Group 1 defined by Díaz-Martínez et al. (2015). Nonetheless, there are some features described in the diagnosis of I. burreyi (Sarjeant et al. 1998) that they do not share, such as the similar digit length in the three digits. Given the current status of the type material of Iguanodontipus burreyi, the poor preservation of whose holotype has been pointed out by several authors (Piñuela et al. 2016 and references therein), its distribution apparently limited to the Berriasian–Valanginian (Díaz-Martínez et al. 2015) and the possible presence of a different ornithopod ichnotaxon in the Late Jurassic (Piñuela et al. 2016), the tracks studied here are tentatively assigned to Iguanodontipodidae indet. because they perfectly fit the diagnosis proposed for the ichnofamily by Díaz-Martínez et al. (2015). These new specimens expand the record of the ichnofamily Iguanodontipodidae to the Late Jurassic.
Ichnofamily uncertain
Ichnogenus Dinehichnus Lockley, dos Santos, Meyer, and Hunt, 1998
Type ichnospecies: Dinehichnus socialis Lockley, dos Santos, Meyer, and Hunt, 1998, Boundary Butte, San Juan County, Utah; Salt Wash Member of the Morrison Formation (Upper Jurassic, Kimmeridgian)
Diagnosis.—“Trackway of small- to medium-sized biped with footprints about as wide as long. Tracks quadripartite, symmetric and tridactyl with distinctive circular heel pad impression. Digit impressions consisting of single elongate oval impressions sometimes with tapered claw impression. Digit divarication (II–IV) averaging about 90°. Trackway narrow with pace angulation of about 155°. Negative (inward) rotation of digit III about 10–15° from trackway midline. Step length averaging a little more than three times foot length” (Lockley et al. 1998).
Dinehichnus isp.
Fig. 3.
Material.—SHN.(JJS).ICNO.58, 60, 68, 109, 113; from Serra do Bouro, Boavista do Bouro, Pedra da Ulsa, Salir do Porto; Alcobaça (Kimmeridgian–lower Tithonian) and Freixial (middle–upper Tithonian) formations (for details see SOM: table S1).
Description.—Small to medium-sized (pes length 16.2–24 cm) tridactyl tracks (Fig. 3 and SOM: table S2), almost as wide as long (pes length/width ratio 0.89–1.08) with medium mesaxony (AT ratio 0.41–0.52). The tracks are robust and almost symmetrical, with a well-developed circular (to subquadrangular) metatarsophalangeal pad impression. One specimen (SHN.(JJS).ICNO.60) shows a quadripartite morphology, and no discrete phalangeal pad impressions have been recognized. A medial notch is slightly more developed in some specimens, so these tracks are slightly asymmetric. The digits are relatively robust; digit III is the longest, and digits II and IV are subequal in length, with variation in some specimens where digit IV is slightly longer than digit II. The hypices are quite symmetrical, with that of digit II located more proximally in some specimens. The distal end of the digit impressions is acuminated, showing slender and pointed claw marks in some specimens. The interdigital angle II–IV is medium (66–78°), interdigital angle II–III being generally greater than interdigital angle III–IV in the majority of the specimens.
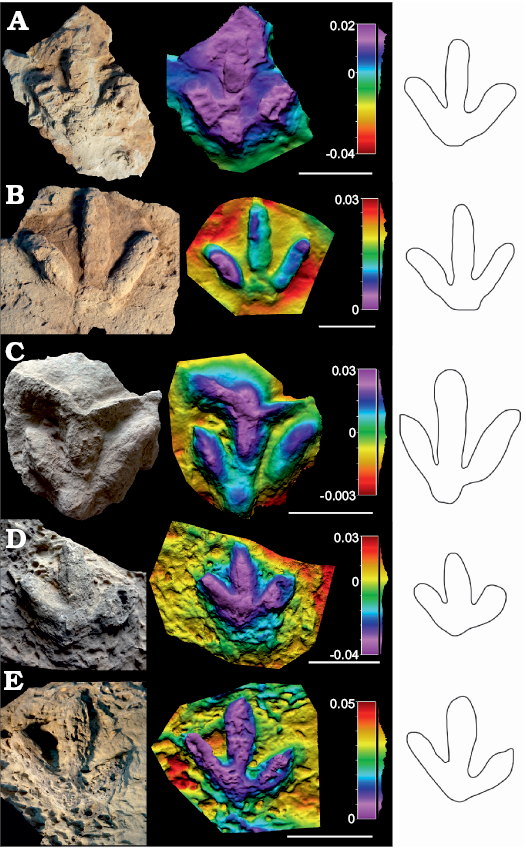
Fig. 3. Photographs, false-colour depth maps, and outline drawing of the tracks of high morphological quality (MP = 2) that belong to the Dinehichnus isp. morphotype. Portugal, Alcobaça Formation, Kimmeridgian–lower Tithonian (A, B, ?D, E) and Freixial Formation, middle–upper Tithonian (C). A. SHN.(JJS).ICNO.58, Serra do Bouro. B. SHN.(JJS).ICNO.60, Boavista do Bouro. C. SHN.(JJS).ICNO.68, Pedra da Ulsa. D. SHN.(JJS).ICNO.109, Salir do Porto. E. SHN.(JJS).ICNO.113, Serra do Bouro. Scale bars: A, B, 10 cm; C–E, 15 cm.
Remarks.—Dinehichnus is the main formally named Late Jurassic ichnogenus assigned to medium-sized ornithopods. It has been described in several places in Europe (including the Lusitanian Basin), the USA, Africa and possibly Asia (Lockley et al. 1998, 2008; Lockley 2009; Gierliński et al. 2009; Belvedere et al. 2010; Xing et al. 2014). Dinehichnus has also been identified in Early Cretaceous deposits in Europe (e.g., Lockley et al. 1998; Hornung and Reich 2012). The assignment of the studied tracks to this ichnotaxon is justified by the fact that the tracks have many of the features described in the diagnosis such as digit divarication, symmetry and especially the distinctive circular pad impression. The reason for not including the tracks in the type ichnospecies D. socialis is the fact that in our sample the quadripartite morphology described in the diagnosis of the ichnospecies is not clearly recognized.
Ichnofamily uncertain
Anomoepus? isp.
Figs. 4, 5.
Material.—SHN.(JJS).ICNO.18, 29, 34, 37, 56, 74, 103, 105C; from Serra do Bouro, Boavista do Bouro, Santa Rita, Foz Velha, Cambelas Sul, Praia da Corva; Alcobaça (Kimmeridgian–lower Tithonian) and Sobral (upper Kimmeridgian–lower Tithonian) formations (for details see SOM: table S1).
Description.—Minute to small-sized (pes length 9–15 cm) tridactyl tracks (Figs. 4, 5 and SOM: table S2), slightly longer than wide (pes length/width ratio 0.85–1.22) with medium mesaxony (AT ratio 0.36–0.61). The tracks are gracile and slightly asymmetrical. The metatarsophalangeal pad impression is subtriangular. One specimen (SHN.(JJS).ICNO.56) shows a constriction in the digits, suggesting possible phalangeal pads. Digit III is the longest, digit IV being slightly longer than II. The digits are considerably thin. The hypices are almost symmetrical. The distal end of the digit impressions is acuminated, showing tapered claw mark impressions in some specimens. The interdigital angle II–IV is medium to high (ranging from 68–102°); the interdigital angles II–III and III–IV are quite similar, with variations between the specimens. SHN.(JJS).ICNO.18 probably shows the evidence of a manus mark, being oval in morphology, but no digits can be recognized.
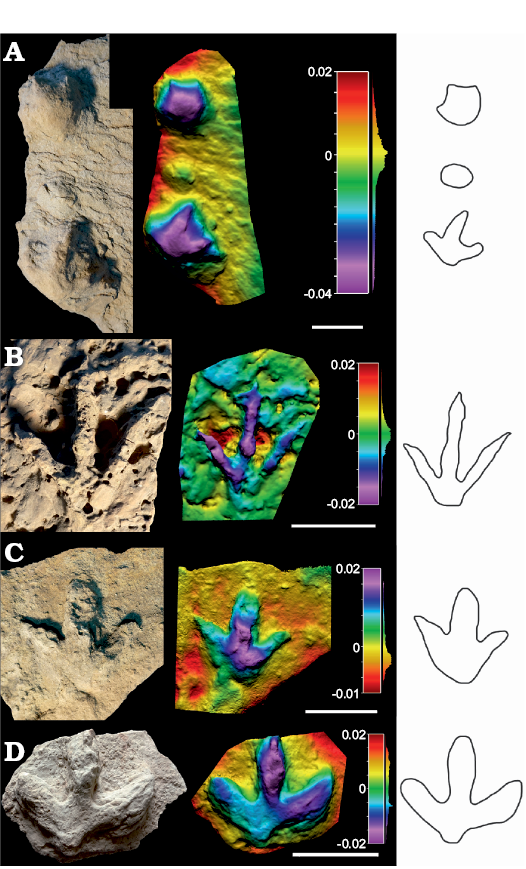
Fig. 4. Photographs, false-colour depth maps and outline drawing of the tracks of high morphological quality (MP = 2) that belong to the Anomoepus? isp. morphotype, Portugal, Alcobaça Formation, Kimmeridgian–lower Tithonian (A, B), Praia de Amoreira-Porto Novo Formation, upper Kimmeridgian (C), and Freixial Formation, middle–upper Tithonian (D). A. SHN.(JJS).ICNO.29, Serra do Bouro. Note the possible manus track. B. SHN.(JJS).ICNO.56, Boavista do Bouro. C. SHN.(JJS).ICNO.103, Santa Rita, Foz Velha. D. SHN.(JJS).ICNO.74, Cambelas Sul. Scale bars 10 cm.
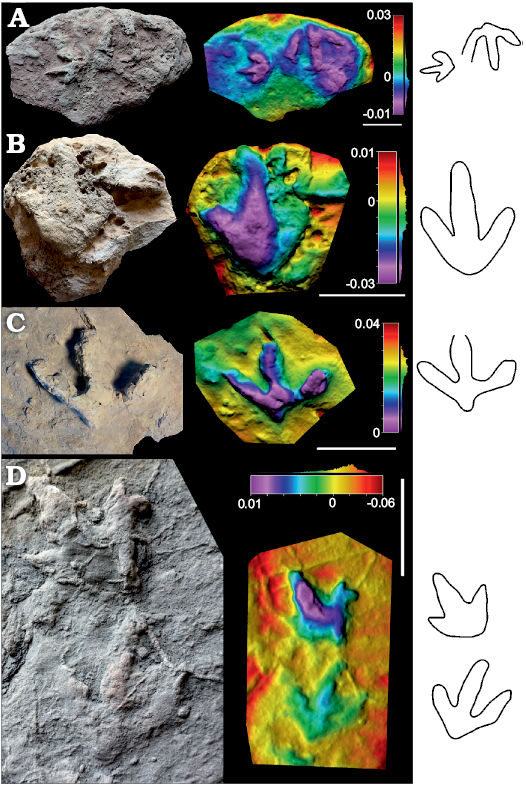
Fig. 5. Photographs, false-colour depth maps and outline drawing of the tracks of high morphological quality (MP = 2) that belong to the Anomoepus? isp. morphotype. Portugal, Alcobaça Formation, Kimmeridgian–lower Tithonian (A–C) and Praia de Amoreira-Porto Novo Formation, upper Kimmeridgian (D). A. SHN.(JJS).ICNO.34A (left) considered Anomoepus? isp. and SHN.(JJS).ICNO.34B, (right) considered Ornithopoda indet., Boavista do Bouro. B. SHN.(JJS).ICNO.37, Boavista do Bouro. C. SHN.(JJS).ICNO.105C, Serra do Bouro. D. SHN.(JJS).ICNO.18A (top) and SHN.(JJS).ICNO.18B (bottom), Praia da Corva. Scale bars 10 cm.
Remarks.—Anomoepus-like tracks have been known ever since the earliest days of dinosaur ichnology, thanks to the descriptions by Hitchcock (1848, 1863). The ichnofamily Anomoepodidae was defined by Lull (1904, 1953), and subsequently Gierliński (1991) emended the diagnosis. Anomoepus-like tracks are well known from Early Jurassic deposits all around the world (Díaz-Martínez et al. 2017 and references therein). Nonetheless, there are different views about how many ichnospecies should be considered valid (Olsen and Rainforth 2003; Lockley and Gierliński 2006). As suggested by Lockley et al. (2009), Anomoepus is hard to identify “unless both manus and pes tracks are found”, and in many cases hallux impressions may not be present when the trackmaker was progressing bipedally. The latter authors also pointed out the difficulties of distinguishing Anomoepus-like tracks from other tridactyl or tetradactyl dinosaur footprints, and suggested that trackway configuration may help in drawing this distinction. However, the absence of complete trackways in the studied sample precludes the use of trackway configuration to resolve the issue. Although notably abundant in Early Jurassic deposits, Anomoepus-like tracks are much scarcer in the context of younger deposits from the Late Jurassic. Lockley et al. (2009) suggested that “there is a significant gap in the distribution of Anomoepodidae between the early Jurassic and the early Cretaceous”. However, Anomoepus-like tracks have been described from the Late Jurassic of Asturias, Spain and the USA (Foster and Lockley 2006; Lockley et al. 2008; Piñuela Suarez 2015) and subsequently at the beginning of the Cretaceous with Neoanomoepus (Lockley et al. 2009). The studied tracks differ considerably with respect to the diagnosis of Neoanomoepus in that the pes is tridactyl instead of tetradactyl. The relation of the studied tracks to the ichnofamily Anomoepodidae should be taken with caution as they do not comply with one of the main features of the diagnosis, namely “pedal digits II, III, and IV tend to be subequal in length”. Nonetheless, we consider that the studied tracks resemble the group of tracks classified as Anomoepus found in Late Jurassic deposits from several areas, and this is why we have classified them with open nomenclature (see also discussion). It should be highlighted that there are some differences in digit width among the specimens. Notably, SHN.(JJS).ICNO.56 (Fig. 4B) and SHN.(JJS).ICNO.18B (Fig. 5D) look more gracile, with lower digit widths. These might just be extramorphological differences, as seen in the differences in the values of morphological quality (M-Preservation; Marchetti et al. 2019).
Ornithopoda indet. tracks
Figs. 6, 7.
Material.—SHN.(JJS).ICNO.02, 09, 11, 21, 43, 46, 53, 71, 72A, 76, 78C, E, 81, 83, 87, 89, 93, 94, 97, 99, 100, 101, 102, 105A–C, E, 106, 112, 120, 121; from Foz Arelho, Praia da Vermelha, Peralta Sul, Paimogo Sul, Salir do Porto, Boavista do Bouro, Serra do Bouro, Vale Perdido, Serra do Bouro; Amoreira-Porto Novo (upper Kimmeridgian), Alcobaça (Kimmeridgian–lower Tithonian), Sobral (upper Kimmeridgian–lower Tithonian) and Freixial (middle–upper Tithonian) formations (for details see SOM: table S1).
Remarks.—Within the collection there are a large number of footprints (n = 39) where the morphological quality is lower than 2, and whose ichnotaxonomic affinities should thus be regarded with caution (Marchetti et al. 2019). These specimens show ornithopod features, in that they are characterized by a low length/width ratio, low mesaxony, symmetry, broad digits, etc., which in many cases resemble the morphotypes described above. For instance, some specimens (SHN.(JJS).ICNO.21, 53, 87, 89, 97, 99, 112) are medium to large-sized (pes length 23–38.5 cm) tridactyl tracks (Figs. 6, 7 and SOM: table S3) with a low length/width ratio (0.84–1.21) and low mesaxony (0.29?–0.54). These specimens generally resemble in many features the tracks classified as Iguanodontipodidae indet. in the previous section. In addition, there are a large number (Figs. 6, 7 and SOM: table S3) of minute and small-sized specimens (pes length 9–21.5 cm), with a low to medium length/width ratio (0.79–1.35), low to medium mesaxony (0.3–0.86), broad digits, relatively high interdigital angle and quite symmetrical shape. These parameters generally fit with those described in the previous section for the tracks classified either as Dinehichnus isp. or Anomoepus? isp. Some specimens even show a rounded circular pad impression (e.g., SHN.(JJS).ICNO.78E, 94, 100) similar to that described in Dinehichnus isp. Nonetheless, given the low morphological quality of these tracks, and in the absence of other morphological features typical of the described ichnotaxa, these tracks are considered simply to be indeterminate ornithopod tracks.
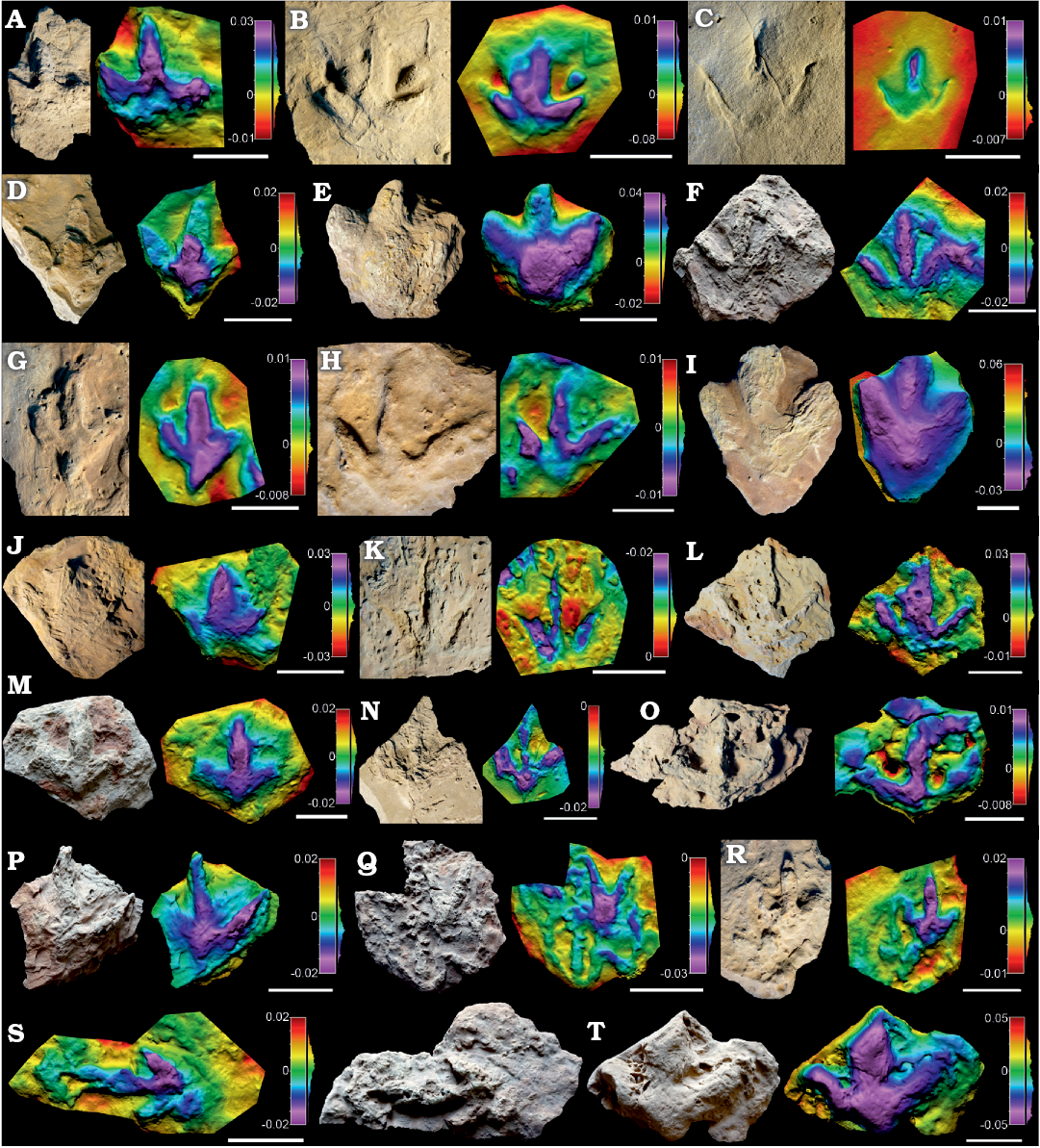
Fig. 6. Photographs, false-colour depth maps of the tracks of low morphological quality (MP < 2) of Ornithopoda indet. tracks, Portugal, Praia de Amoreira-Porto Novo Formation, upper Kimmeridgian (A–D), ?Sobral Formation, upper Kimmeridgian–lower Tithonian (E), Alcobaça Formation, Kimmeridgian–lower Tithonian (?F, G–T) A. SHN.(JJS).ICNO.11, Foz Arelho. B. SHN.(JJS).ICNO.43, Praia da Vermelha. C. SHN.(JJS).ICNO.46, Foz Arelho. D. SHN.(JJS).ICNO.83, Peralta Sul. E. SHN.(JJS).ICNO.89, Paimogo Sul. F. SHN.(JJS).ICNO.100, Salir do Porto. G. SHN.(JJS).ICNO.02, Boavista do Bouro. H. SHN.(JJS).ICNO.09, Serra do Bouro. I. SHN.(JJS).ICNO.53, Serra do Bouro. J. SHN.(JJS).ICNO.72A, Serra do Bouro. K. SHN.(JJS).ICNO.76, Serra do Bouro. L. SHN.(JJS).ICNO.81, Serra do Bouro. M. SHN.(JJS).ICNO.93, Serra do Bouro. N. SHN.(JJS).ICNO.94, Serra do Bouro. O. SHN.(JJS).ICNO.102, Vale Perdido, Serra do Bouro. P. SHN.(JJS).ICNO.105A, Serra do Bouro. Q. SHN.(JJS).ICNO.105B, Serra do Bouro. Note the three footprints ICNO.105B.1 (bottom left), ICNO.105B.2 (top) and ICNO.105B.3 (bottom left). R. SHN.(JJS).ICNO.105D, Serra do Bouro. S. SHN.(JJS).ICNO.105E, Serra do Bouro. T. SHN.(JJS).ICNO.120, Serra do Bouro. Scale bars O, 5 cm; A, B, D, G–I, L–N, P, R–T, 10 cm; C, E, F, J, K, Q, 15 cm.
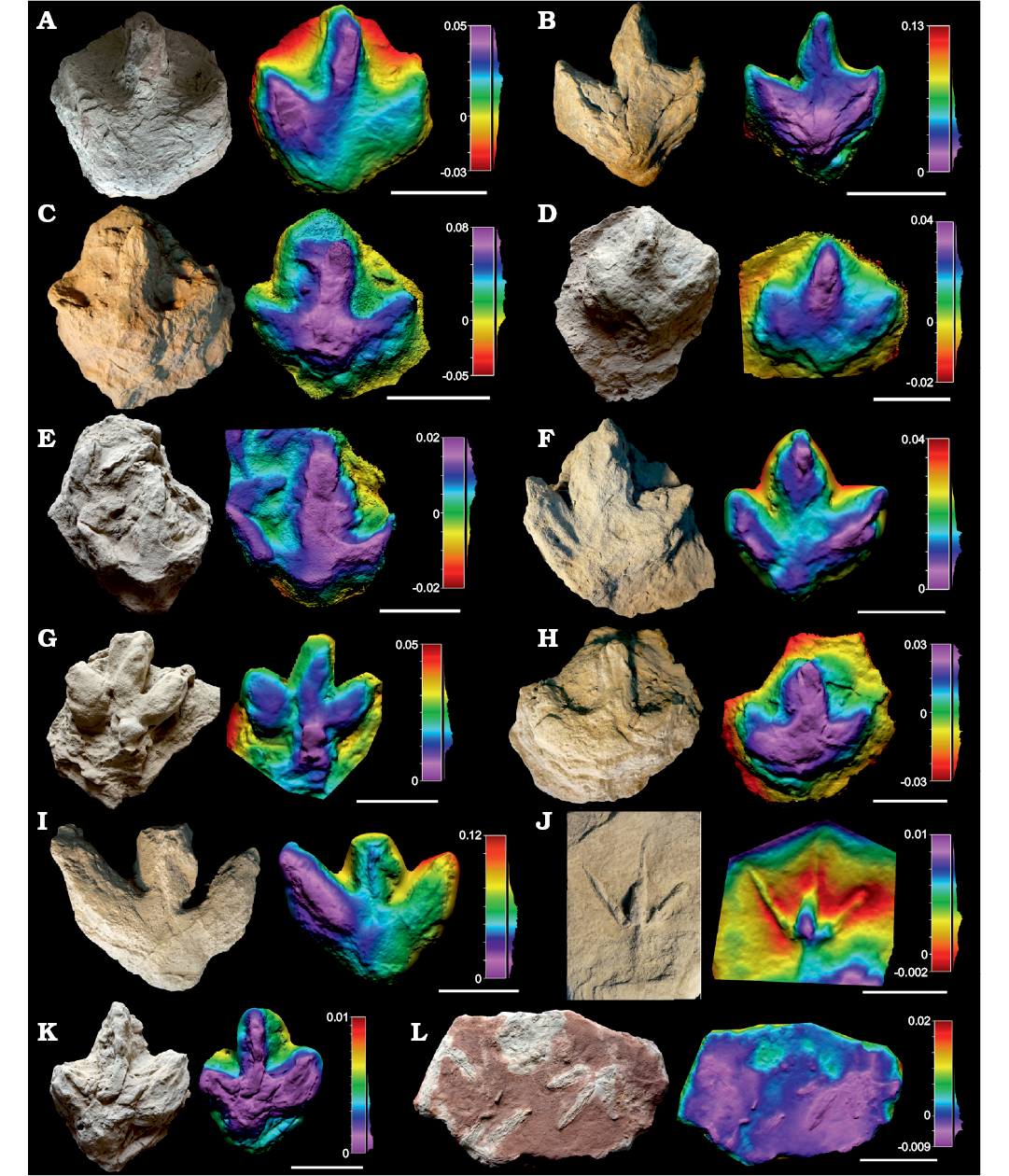
Fig. 7. Photographs, false-colour depth maps of the tracks of low morphological quality (MP < 2) of Ornithopoda indet. tracks, Portugal, Sobral Formation, upper Kimmeridgian–lower Tithonian (A–D), Freixial Formation, middle–upper Tithonian (E–K), and Alcobaça Formation, Kimmeridgian–lower Tithonian (L) A. SHN.(JJS).ICNO.21, Praia Azul. B. SHN.(JJS).ICNO.87, Praia Azul. C. SHN.(JJS).ICNO.97, Praia Azul. D. SHN.(JJS).ICNO.106, Foz do Rio Sizandro. E. SHN.(JJS).ICNO.71, Porto Chão, Cambelas. F. SHN.(JJS).ICNO.78C, Porto Chão, Cambelas. G. SHN.(JJS).ICNO.78E, Porto Chão, Cambelas. H. SHN.(JJS).ICNO.80, Cambelas Sul. I. SHN.(JJS).ICNO.99, Cambelas. J. SHN.(JJS).ICNO.101, Cambelas. K. SHN.(JJS).ICNO.112, Pedra da Ulsa Norte. L. SHN.(JJS).ICNO.121, Boavista do Bouro. Scale bars D–H, J, 10 cm; I, K, 15 cm; A–C, L, 20 cm.
Discussion
Despite the difficulties of distinguishing between theropod and ornithopod tracks in the Late Jurassic and Early Cretaceous (e.g., Lockley et al. 2008; Schulp and Al-Wosabi 2012; Castanera et al. 2013a, b; Hübner 2016), the features of the three morphotypes described herein fit well with the characters typical of small, medium to large-sized ornithopod tracks, namely: short, U-shaped digits, the absence of sharp claw impressions with generally blunt and rounded morphology of the digit tips, low length-width ratios, medium to high divarication, symmetry, and similar width among the digits (Thulborn 1990; Lockley et al. 2008). Hübner (2016) proposed that weak mesaxony and broad digits can be interpreted as diagnostic features of ornithischian tracks. Recently, Lallensack et al. (2019) proposed that for small-sized tridactyl tracks the interdigital angle and the projection of digit III are the best features for discriminating between theropods and ornithischians, the former having smaller interdigital angles and a larger projection of digit III. Here, we propose ornithopods, a group of ornithischians, as the best candidates to be the trackmakers according to the osteological record found in the Lusitanian Basin (see below). This is also consistent with the identification of gracile ornithopod tracks from the Late Jurassic in other sites of the Iberian Peninsula (Santisteban et al. 2003; Lockley et al. 2008; Castanera et al. 2013b; Alcalá et al. 2014).
The three morphotypes described in the present paper have been assigned to three different ornithischian ichnotaxa, named Iguanodontipodidae indet., Dinehichnus isp. and Anomoepus? isp. These three ichnotaxa have generally been associated with iguanodontians such as dryosaurids and ankylopollexians and with basal ornithischians as candidate trackmakers (Lockley et al. 1998; Lockley and Gierliński 2006; Díaz-Martínez et al. 2015). Several features prevent us from considering birds as potential trackmakers of the smaller morphotype (Anomoepus? isp.): bird footprints are generally smaller; the hallux impressions (when present) are oriented posteriorly; and generally they have slender digits and wider divarication angles of about 100–150° (Belvedere et al. 2011). Moreover, given that the majority of the specimens are preserved as natural casts, possible morphological variations due to undertrack preservation should be kept in mind, as some theropod tracks might look ornithopod in origin when preserved as undertracks (Avanzini et al. 2012). Accordingly, we cannot confidently assert that all the studied specimens (especially those of low morphological quality) were produced by ornithopods, even though the parameters and the features (length-width ratio, mesaxony, interdigital angle, symmetry, broad digits) do indeed suggest an ornithopod origin.
Distinguishing between ornithopod morphotypes is not an easy task. In fact, Hornung et al. (2016) have recently pointed out the difficulties of discriminating in qualitative and quantitative terms between several ornithopod tracks from the Berriasian of Germany. In this regard, it is interesting to note that in our sample the three described morphotypes can be distinguished qualitatively and also by size, whereas quantitative discrimination analyzing mesaxony (AT) and length/width ratio is not significant (Fig. 8), especially between the smaller (Anomoepus? isp.) and the medium-sized (Dinehichnus isp.) morphotypes. This raises the question of how similar to one another or different from one another the three different morphotypes are. The fact that the footprints come from different locations and different geological formations rules out possible ontogenetic variations as the only reason for the size differences among the three morphotypes, so diversity and taxonomy are likely to have played a role in this distinction. Whatever the case, we cannot exclude the possibility that ontogeny may play a role in distinguishing some of the tracks included in the different morphotypes, and that the morphometric differences might be a consequence of differential allometric growth between small and large individuals (e.g., Olsen et al. 1998). Nonetheless, little is known about the influence of ontogenetic changes on the feet of ornithopod dinosaurs and thus possible footprint shape variations, so this assumption would be more a matter of speculation than a scientific fact based on currently available data.
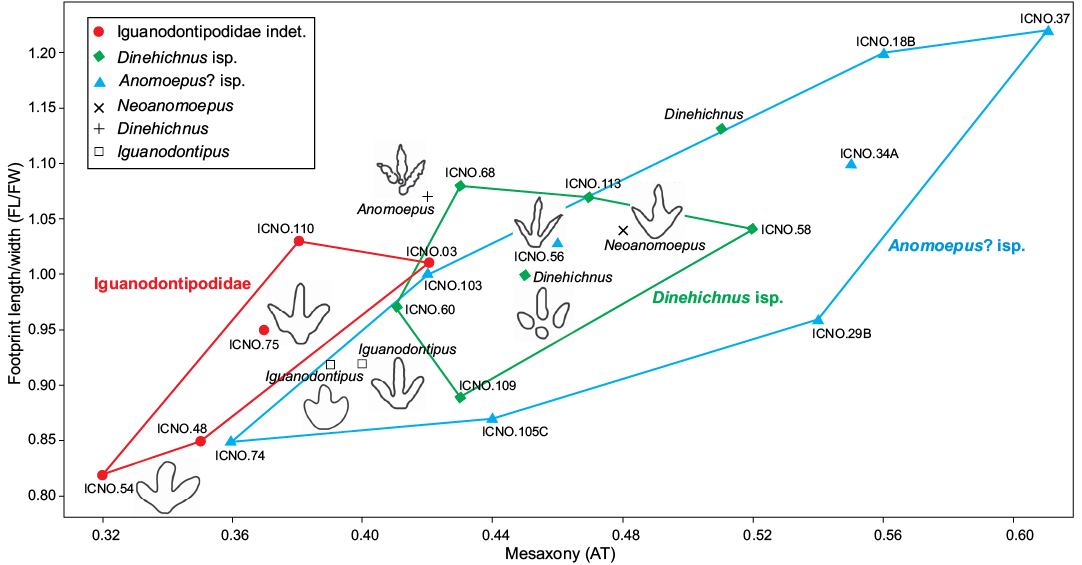
Fig. 8. Bivariate graph plotting the footprint length/footprint width ratio against the mesaxony (AT) of the studied tracks (bolded) compared with the ichnotaxa Anomoepus, Neoanomoepus, Dinehichnus, and Iguanodontipus (data from Lockley 2009). Outline drawings not to scale. For simplicity the acronym ICNO.# was used instead of SHN.(JJS).ICON.#.
It is noteworthy that the length/width ratio is very similar among the three different morphotypes in the studied sample, the main differences being in the mesaxony (Fig. 8). The studied sample shows the tendency noted by Lockley (2009) for smaller ornithopod tracks to be more elongated (stronger mesaxony) than large ornithopod tracks. In fact, some of the smaller tracks (Anomoepus? isp.) show the strongest mesaxony whereas tracks assigned to Iguanodontipodidae indet. display the weakest mesaxony. Nonetheless, there is some overlap between the three morphotypes. In this respect, Anomoepus? isp. and Iguanodontipodidae indet. can mainly be distinguished quantitatively, but the intermediate morphotype (Dinehichnus isp.) shows intermediate values. This raises the question whether the three morphotypes should be classified as three different ichnotaxa or not. For instance, some authors have suggested that Dinehichnus should be synonymized with Anomoepus at the ichnogeneric level (Dalman and Weems 2013; Piñuela Suarez 2015). Other authors (Gierliński et al. 2009) have also noted the similarities but emphasized the differences, stating that “Anomoepus footprints are often narrower, with less divergent and thinner digits, which usually show well defined phalangeal pads”. In our sample, Anomoepus? isp. tracks do not show clear phalangeal pads (with the possible exception of SHN.(JJS).ICNO.56, Fig. 4B), although the digits are considerably thinner and some specimens show higher divarication angles than in the Dinehichnus isp. morphotype. Lockley et al. (2008) studied Anomoepus-like tracks from the Late Jurassic of Asturias and suggested that “these tracks are enigmatic because they are unlike any previously described from the Upper Jurassic”. The authors pointed out the similarities with Anomoepus but added that “this label is tentative at best” and concluded that “very little is known of the morphology and spatial and temporal distribution of Anomoepus-like tracks”. According to Lockley et al. (2009), Anomoepus ichnospecies generally vary in size from 5 to 15 cm (without metatarsus impressions), so although size should not be taken into account in classifying footprints (Bertling et al. 2006), this threshold value of 15 cm is also a good criterion for distinguishing between the smaller (Anomoepus? isp.) and the intermediate (Dinehichnus isp.) morphotypes. Another issue of note is that these Anomoepus-like tracks from the Late Jurassic do not properly comply with the diagnosis of the ichnofamily Anomoepodidae because digit III is clearly the longest whereas in the emended diagnosis proposed by Gierliński (1991) “the pedal digits II, III, and IV tend to be subequal in length”. Thus, further work is needed in order to understand how these Late Jurassic Anomoepus-like tracks fit within Anomoepodidae, as well as whether Dinehichnus tracks might also fit within this ichnofamily.
The ornithopod record from the Lusitanian Basin is quite rich compared with that of other Late Jurassic areas in Europe (e.g., Campos-Soto et al. 2019), where fossils from this group are relatively scarce. The ornithopod fauna is mainly composed of medium-sized ornithopods: members of Ankylopollexia (Galton 1980; Mateus and Antunes 2001; Ortega et al. 2009; Rotatori et al. 2020) such as Draconyx loureiroi (Bombarral Formation, Tithonian), but also smaller forms such as dryosaurids (e.g., Rotatori et al. 2020), including Eousdryosaurus nanohallucis (Praia da Amoreira-Porto Novo Formation, Kimmeridgian, Escaso et al. 2014), and some materials assigned to possible “hypsilophodontids” (Galton 1980; Rauhut 2001; Antunes and Mateus 2003). None of the tracks has preserved hallux marks, which may suggest a “camptosaur” affinity rather than a dryosaurid one (Gierliński et al. 2009) for some of the morphotypes. These authors estimated 26 cm for the length of the track that Camptosaurus dispar would have produced, so this estimate would fit within the range of the tracks here classified as Iguanodontipodidae indet. Bakker (1996) proposed that small ornithopod dinosaurs such as Drinker or Othnielia would have been good candidates for producing Anomoepus-like tracks. Assigning the described morphotypes to a specific group of ornithopods is difficult since a synapomorphy-based technique (Carrano and Wilson 2001) cannot be applied with confidence, making it impossible to positively identify the best ornithopod group to be the candidate trackmaker. Nonetheless, it is interesting to note that considering their size and in the light of the previous discussion, ankylopollexians are the best candidates for having produced Iguanodontipodidae indet., whereas both dryosaurids and the possible “hypsilophodontids” are better candidates to be the producers of the Dinehichnus isp. and Anomoepus? isp. morphotypes.
Conclusions
Ornithopod footprints collected from different areas along the Portuguese coast within the Lusitanian Basin (Consolação and Turcifal sub-basins) are described for the first time. The footprints come from different geological formations (Praia de Amoreira-Porto Novo, Alcobaça, Sobral, and Freixial) that are Late Jurassic in age (Kimmeridgian–Tithonian). Three different morphotypes are identified: Anomoepus? isp., Dinehichnus isp., and Iguanodontipodidae indet. This new study highlights the difficulties of quantitatively distinguishing between ornithopod footprints although they can be differentiated qualitatively by morphological differences (robustness of the digits, metatarsophalangeal pad impression morphology and size). The tracks included in the largest morphotype (Iguanodontipodidae indet.) represent one of the oldest records of this ichnofamily. The descriptions of the smallest (Anomoepus? isp.) and the medium-sized (Dinehichnus isp.) morphotypes provide new data for identifying gracile ornithopod tracks in the Late Jurassic, although the ichnotaxonomic status of the former and the relationships of both within the ichnofamily Anomoepodidae cannot be completely resolved with the studied material. Thus, the study also highlights the great need for a systematic review to clarify the ichnotaxonomy of small to medium-sized ornithopod tracks in general, and help to ascertain the similarities/differences between Early Jurassic and Late Jurassic ichnotaxa. The new material adds to the ornithopod record of the Lusitanian Basin, where osteological remains have already been described but few ornithopod tracks had been identified. Although no direct correlations can be drawn between the ichnological and osteological records, the described footprints are consistent with the main ornithopod groups described so far (dryosaurids, ankylopollexians, and “hypsilophodontids”), which are excellent candidates to be the trackmakers.
Acknowledgements
We acknowledge José Joaquim Santos and André Mano (both Sociedade de História Natural (SHN), Torres Vedras, Portugal) for field assistance. Rupert Glasgow (Zaragoza, Spain) revised the English grammar. The comments of the two reviewers Ignacio Díaz Martínez (CONICET-Universidad Nacional de Río Negro, Argentina) and Jahn Hornung (Landesmuseum Hannover, Germany), and the editor Olivier Lambert (Institut royal des Sciences naturelles de Belgique, Brussels, Belgium) have greatly improved the paper and are very much appreciated. This study was funded by the Alexander von Humboldt Foundation (DC and MB). DC is supported by the Beatriu de Pinós postdoctoral programme (BP2017-00195) of the Government of Catalonia’s Secretariat for Universities and Research of the Ministry of Economy and Knowledge. This project was partially supported by the Municipality of Torres Vedras (Portugal) through an institutional agreement with the Sociedade de História Natural. We also acknowledge the financial support provided by the FCT-project UID/GEO/50019/2019 Instituto Dom Luiz.
References
Alcalá, L., Mampel, L., Royo-Torres, R., and Cobos, A. 2014. On small quadrupedal ornithopod tracks in Jurassic–Cretaceous transition intertidal deposits (El Castellar, Teruel, Spain). Spanish Journal of Palaeontology 29: 83–190.
Antunes, M.T. and Mateus, O. 2003. Dinosaurs of Portugal. Comptes Rendus Palevol 2: 77–95. Crossref
Avanzini, M., Piñuela, L., and García-Ramos, J.C. 2012. Late Jurassic footprints reveal walking kinematics of theropod dinosaurs. Lethaia 45: 238–252. Crossref
Bakker, R.T. 1996. The real Jurassic park: dinosaurs and habitats at Como Bluff, Wyoming. In: M. Morales (ed.), The Continental Jurassic. Museum of Northern Arizona Bulletin 60: 35–49.
Bengston, P. 1998. Open nomenclature. Palaeontology 31: 223–227. Crossref
Belvedere, M., Dyke, G., Hadri, M., and Ishigaki, S. 2011. The oldest evidence for birds in Northern Gondwana? Small tridactyl footprints from the Middle Jurassic of Msemrir (Morocco). Gondwana Research 19: 542–549. Crossref
Belvedere, M., Mietto, P., and Ishigaki, S. 2010. A Late Jurassic diverse ichnocoenosis from the siliciclastic Iouaridène Formation (Central High Atlas, Morocco). Geological Quarterly 54: 367–380.
g, M., Braddy, S.J., Bromley, R.G., Demathieu, G.R., Genise, J., Mikuláš, R., Nielsen, J.K., Nielsen, K.S.S., Rindsberg, A.K., Schlirf, M., and Uchman, A. 2006. Names for trace fossils: a uniform approach. Lethaia 39: 265–286. Crossref
Campos-Soto, S., Benito, M.I., Cobos, A., Caus, E., Quijada, I.E., Suarez-Gonzalez, P., Mas, R., Royo-Torres, R., and Alcalá, L. 2019. Revisiting the age and palaeoenvironments of the Upper Jurassic–Lower Cretaceous. Dinosaur-bearing sedimentary record of eastern Spain: implications for Iberian palaeogeography. Journal of Iberian Geology 45: 471–510. Crossref
Carrano, M.T. and Wilson, J.A. 2001. Taxon distributions and the tetrapod track record. Paleobiology 27: 564–582. Crossref
Castanera, D., Belvedere, M., Marty, D., Paratte, G., Lapaire-Cattin, M., Lovis, C., and Meyer, C.A. 2018. A walk in the maze: variation in Late Jurassic tridactyl dinosaur tracks from the Swiss Jura Mountains (NW Switzerland). PeerJ 6: e4579. Crossref
Castanera, D., Pascual, C., Razzolini, N.L., Vila, B., Barco, J.L., and Canudo, J.I. 2013a. Discriminating between medium-sized tridactyl trackmakers: tracking ornithopod tracks in the base of the Cretaceous (Berriasian, Spain). PloS one: 8 (11): e81830. Crossref
Castanera, D., Vila, B., Razzolini, N.L., Falkingham, P.L., Canudo, J.I., Manning, P.L., and Galobart, À. 2013b. Manus track preservation bias as a key factor for assessing trackmaker identity and quadrupedalism in basal ornithopods. PloS one 8 (1): e54177. Crossref
Dalman, S.G. and Weems, R.E. 2013. A new look at morphological variation in the ichnogenus Anomoepus, with special reference to material from the Lower Jurassic Newark Supergroup: implications for ichnotaxonomy and ichnodiversity. Bulletin of the Peabody Museum of Natural History 54: 67–125. Crossref
Díaz-Martínez, I., González, S.N., and De Valais, S. 2017. Dinosaur footprints in the Early Jurassic of Patagonia (Marifil Volcanic Complex, Argentina): biochronological and palaeobiogeographical inferences. Geological Magazine 154: 914–922. Crossref
Díaz-Martínez, I., Pereda-Suberbiola, X., Perez-Lorente, F., and Canudo, J.I. 2015. Ichnotaxonomic review of large ornithopod dinosaur tracks: temporal and geographic implications. PloS one 10 (2): e0115477. Crossref
Diedrich, C. 2011. Upper Jurassic tidal flat megatracksites of Germany—coastal dinosaur migration highways between European islands, and a review of the dinosaur footprints. Palaeobiodiversity and Palaeoenvironments 91: 129–155. Crossref
Escaso, F., Ortega, F., Dantas, P., Malafaia, E., Silva, B., Gasulla, J.M., Mocho, P., Narváez, I., and Sanz, J.L. 2014. A new dryosaurid ornithopod (Dinosauria, Ornithischia) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology 34: 1102–1112. Crossref
Falkingham, P.L., Bates, K.T., Avanzini, M., Bennett, M., Bordy, E.M., Breithaupt, B. H., Castanera, D., Citton, P., Díaz-Martínez, I., Farlow, J., Fiorillo, A.R., Gatesy, S.M., Getty, P., Hatala, K.G., Hornung, J.J., Hyatt, J.A., Klein, H., Lallensack, J.N., Martin, A.J., Marty, D., Matthews, N.A., Meyer, C.A., Milàn, J., Minter, N.J., Razzolini, N.L., Romilio, A., Salisbury, S.W., Sciscio, L., Tanaka, I., Wiseman, A.L.A. Xing, L.D., and Belvedere, M. 2018. A standard protocol for documenting modern and fossil ichnological data. Palaeontology 61: 469–480. Crossref
Foster, J.R. and Lockley, M.G. 2006. The vertebrate ichnological record of the Morrison Formation (Upper Jurassic, North America). New Mexico Museum of Natural History and Science Bullletin 36: 203–216.
Galton, P.M. 1980. European Jurassic ornithopod dinosaurs of the families Hypsilophodontidae and Camptosauridae. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 160: 73–95.
Gierliński, G. 1991. New dinosaur ichnotaxa from the early Jurassic of the Holy cross Mountains, Poland. Palaeogeography, Palaeoclimatology, Palaeoecology 85: 137–148. Crossref
Gierliński, G., Niedźwiedzki, G., and Nowacki, P. 2009. Small theropod and ornithopod footprints in the Late Jurassic of Poland. Acta Geologica Polonica 59: 221–234.
Hammer, Ø., Harper, D.A., and Ryan, P.D. 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia electronica 4 (1): 1–9.
Hill, G. 1988. The Sedimentology and Lithostratigraphy of the Upper Jurassic Lourinhã Formation, Lusitanian Basin, Portugal. 300 pp. Ph.D. Thesis, Open University, Milton Keynes.
Hill, G. 1989. Distal alluvial fan sediments from the Upper Jurassic of Portugal: controls on their cyclicity and channel formation. Journal of the Geological Society of London 146: 539–555. Crossref
Hitchcock, E. 1848. An attempt to discriminate and describe the animals that made the fossil footmarks of the United States, and especially of New England. Memoirs of the American Academy of Arts and Sciences 3: 129–256. Crossref
Hitchcock, E. 1863. New facts and conclusions respecting the fossil footmarks of the Connecticut Valley. American Journal of Science and Arts 36: 47–57. Crossref
Hornung, J.J. and Reich, M. 2012. Dinosaur tracks from the Berriasian Obernkirchen Sandstone on exhibit at the Göttingen University Geopark. In: A. Richter and M. Reich (eds.), Dinosaur tracks 2011. An International Symposium, Obernkirchen, April 14–17, 2011. Abstract Volume and Field Guide to Excursions, 169–187. Universitätsverlag Göttingen, Göttingen.
Hornung, J.J., Böhme, A., Schlüter, N., and Reich, M. 2016. Diversity, ontogeny, or both? A morphometric approach to iguanodontian ornithopod (Dinosauria: Ornithischia) track assemblages from the Berriasian (Lower Cretaceous) of North Western Germany. In: P.L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks. The Next Steps, 202–225. Indiana University Press, Bloomington.
Hübner, T. 2016. Elusive Ornithischian Tracks in the Famous Berriasian (Lower Cretaceous) “Chicken Yard” Tracksite of Northern Germany: Quantitative Differentiation between Small Tridactyl Trackmakers. In: P.L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks. The Next Steps, 310–333. Indiana University Press, Bloomington.
Hunt, A.P. and Lucas, S.G. 2007. Tetrapod ichnofacies: a new paradigm. Ichnos 14: 59–68. Crossref
Kullberg, J.C., Rocha, R.B., Soares, A.F., Rey, J., Terrinha, P., Azerêdo, A.C., Callapez, P., Duarte, L.V., Kullberg, M.C., Martins, L., Miranda, J.R., Alves, C., Mata, J., Madeira, J., Mateus, O., Moreira, M., and Nogueira, C.R. 2013. A Bacia Lusitaniana: Estratigrafia, Paleogeografia e Tectónica. In: R. Dias, A. Araújo, P. Terrinha, and J.C. Kullberg (eds.), Geologia de Portugal. Vol. 2: Geologia Meso-cenozóica de Portugal, 195–347. Escolar Editora, Lisboa.
Lallensack, J.N., Engler, T., and Barthel, H.J. 2019. Shape variability in tridactyl dinosaur footprints: the significance of size and function. Palaeontology [published online, https://doi.org/10.1111/j.1502-3931.2011. 00276.x]. Crossref , Crossref
Lockley, M.G. 2009. New perspectives on morphological variation in tridactyl footprints: clues to widespread convergence in developmental dynamics. Geological Quarterly 53: 415–432.
Lockley, M.G. and Gierliński, G. D. 2006. Diverse vertebrate ichnofaunas containing Anomoepus and other unusual trace fossils from the Lower Jurassic of the western United States: implications for paleoecology palichnostratigraphy. The Triassic–Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin 37: 176–191.
Lockley, M.G. and Santos, V.F. 1993. A preliminary report on sauropod trackways from the Avelino site, Sesimbra region, Upper Jurassic, Portugal. Gaia 6: 38–42.
Lockley, M.G. and Wright, J.L. 2001. Trackways of large quadrupedal ornithopods from the Cretaceous: A review. In: D.H. Tanke and K. Carpenter (eds.), Mesozoic Vertebrate Life, 428–442. Indiana University Press, Bloomington.
Lockley, M.G., García-Ramos, J.C., Piñuela, L., and Avanzini, M. 2008. A review of vertebrate track assemblages from the Late Jurassic of Asturias, Spain with comparative notes on coeval ichnofaunas from the western USA: implications for faunal diversity in siliciclastic facies assemblages. Oryctos 8: 53–70.
Lockley, M.G., Mccrea, R.T., and Matsukawa, M. 2009. Ichnological evidence for small quadrupedal ornithischians from the basal Cretaceous of SE Asia and North America: implications for a global radiation. Geological Society, London, Special Publications 315: 255–269. Crossref
Lockley, M.G. Meyer, C.A., and Santos, V.F. 1994. Trackway evidence for a herd of juvenile sauropods from the late Jurassic of Portugal. Aspects of sauropod paleobiology. Gaia 10: 27–35.
Lockley, M.G., Santos, V.F., Meyer, C., and Hunt, A. 1998. A new dinosaur tracksite in the Morrison Formation, Boundary Butte, southeastern Utah. Modern Geology 23: 317–330.
Lockley, M.G., Xing, L., Lockwood, J.A., and Pond, S. 2014. A review of large Cretaceous ornithopod tracks, with special reference to their ichnotaxonomy. Biological Journal of the Linnean Society 113: 721–736. Crossref
Lull, R.S. 1904. Fossil footprints of the Jura–Trias of North America. Boston Society of Natural History 5: 461–557.
Lull, R.S. 1953. The Triassic life of the Connecticut Valley. Connecticut State Geological and Natural History Survey, Bulletin 81: 1–336.
Mallison, H. and Wings, O. 2014. Photogrammetry in paleontology—a practical guide. Journal of Paleontological Techniques 12: 1–31.
Manuppella, G., Antunes, M.T., Pais, J., Ramalho, M.M., and Rey, J. 1999. Notícia explicativa da Folha 30-A. 83 pp. Instituto Geológico e Mineiro de Lisboa, Lourinhã.
Marchetti, L., Belvedere, M., Voigt, S., Klein, H., Castanera, D., Díaz-Martínez, I., Marty, D., Xing, L., Feola, S., Melchor, R.N., and Farlow, J.O. 2019. Defining the morphological quality of fossil footprints. Problems and principles of preservation in tetrapod ichnology with examples from the Palaeozoic to the present. Earth-Science Reviews 193: 109–145. Crossref
Marty, D. 2008. Sedimentology, taphonomy, and ichnology of Late Jurassic dinosaur tracks from the Jura carbonate platform (Chevenez-Combe Ronde tracksite, NW Switzerland): insights into the tidal-flat palaeoenvironment and dinosaur diversity, locomotion, and palaeoecology. GeoFocus 21: 1–278.
Marty, D., Belvedere, M., Razzolini, N.L., Lockley, M.G., Paratte, G., Cattin, M., Lovis, C., and Meyer, C.A. 2018. The tracks of giant theropods (Jurabrontes curtedulensis ichnogen. & ichnosp. nov.) from the Late Jurassic of NW Switzerland: palaeoecological & palaeogeographical implications. Historical Biology 30: 928–956. Crossref
Mateus, O. and Antunes, M.T. 2001. Draconyx loureiroi, a new camptosauridae (Dinosauria, Ornithopoda) from the Late Jurassic of Lourinhã, Portugal. Annales de Paléontologie 87: 61–73. Crossref
Mateus, O. and Milàn, J. 2008. Ichnological evidence for giant ornithopod dinosaurs in the Upper Jurassic Lourinhã Formation, Portugal. Oryctos 8: 47–52.
Mateus, O. and Milàn, J. 2010. A diverse Upper Jurassic dinosaur ichnofauna from central-west Portugal. Lethaia 43: 245–257. Crossref
Mateus, O., Dinis, J., and Cunha, P.P. 2013. Upper Jurassic to Lowermost Cretaceous of the Lusitanian Basin, Portugal—landscapes where dinosaurs walked. Ciências da Terra 8: 1–37.
Matthews, N., Noble, T., and Breithaupt, B.H. 2016. Close-range photogrammetry for 3D ichnology: the basics of photogrammetric ichnology. In: P.L. Falkingham, D. Marty, and A. Richter (eds.), Dinosaur Tracks. The Next Steps, 29–55. Indiana University Press, Bloomington.
Mazin, J.M., Hantzpergue, P., and Olivier, N. 2017. The dinosaur tracksite of Plagne (early Tithonian, Late Jurassic; Jura Mountains, France): the longest known sauropod trackway. Geobios 50: 279–301. Crossref
Moreno, K., de Valais, S., Blanco, N., Tomlinson, A.J, Jacay. J., and Calvo, J.O. 2012. Large theropod dinosaur footprint associations in western Gondwana: Behavioural and palaeogeographic implications. Acta Palaeontologica Polonica 57: 73–83. Crossref
Oertel, G., Freire de Andrade, C., Zbyszewski, G., and Da Veiga Ferreira, O. 1960. Carta geológica de Portugal, escala 1:50 000, Folha 26-C, Peniche. Serviços Geológicos de Portugal, Lisboa.
Oliveira, J.T., Pereira, H., Ramalho, M. Antunes, M.T, and Monteiro, J.H. 1992. Carta Geológica de Portugal, Escala 1:500 000. Serviços Geológicos de Portugal, Lisboa.
Olsen, P.E. and Rainforth, E.C. 2003. The early Jurassic ornithischian dinosaurian ichnogenus Anomoepus. In: P.M. LeTourneau and P.E. Olsen (eds.), The Great Rift Valleys of Pangea in Eastern North America. Sedimentology, Stratigraphy, and Paleontology 2: 314–367.
Olsen, P.E., Smith, J.B., and Mcdonald, N.G. 1998. Type material of the type species of the classic theropod footprint genera Eubrontes, Anchisauripus, and Grallator (Early Jurassic, Hartford and Deerfield basins, Connecticut and Massachusetts, USA). Journal of Vertebrate Paleontology 18: 586–601. Crossref
Ortega, F., Malafaia, E., Escaso, F., Pérez-García, A., and Dantas, P. 2009. Faunas de répteis do Jurássico Superior de Portugal. Paleolusitana 1: 43–56.
Piñuela, L., García-Ramos, J.C., Romano, M., and Ruiz-Omeñaca, J.I. 2016. First record of gregarious behavior in robust medium-sized Jurassic ornithopods: evidence from the Kimmeridgian trackways of Asturias (N Spain) and some general considerations on other medium-large ornithopod tracks in the Mesozoic record. Ichnos 23: 298–311. Crossref
Piñuela Suarez, L. 2015. Huellas de dinosaurios y de otros reptiles del Jurásico Superior de Asturias. 326 pp. Unpublished Ph.D. Thesis, Oviedo University, Oviedo.
Rauhut, O.W. 2001. Herbivorous dinosaurs from the Late Jurassic (Kimmeridgian) of Guimarota, Portugal. Proceedings of the Geologists’ Association 112: 275–283. Crossref
Rotatori, F.M., Moreno-Azanza, M., and Mateus, O. 2020. New information on ornithopod dinosaurs from the Late Jurassic of Portugal. Acta Palaeontologica Polonica 65: 35–57. Crossref
Santisteban, C., Gaete, R., Galobart, A., and Suñer, M. 2003. Rastros de dinosaurios en el Jurásico terminal (facies Purbeck) de Corcolilla (Los Serranos, Valencia). In: F. Pérez-Lorente (ed.), Dinosaurios y otros reptiles mesozoicos en España, 33–40. Universidad de La Rioja, Logroño.
Santos, V.F. 2008. Pegadas de dinossáurios de Portugal. 123 pp. Museu Nacional de História Natural da Universidade de Lisboa, Lisboa.
Sarjeant, W.A., Delair, J.B., and Lockley, M.G. 1998. The footprints of Iguanodon: a history and taxonomic study. Ichnos 6 (3): 183–202. Crossref
Schneider, S., Fursich, F.T., and Werner, W. 2009. Sr-isotope stratigraphy of the Upper Jurassic of central Portugal (Lusitanian Basin) based on oyster shells. International Journal of Earth Sciences 98: 1949–1970. Crossref
Schulp, A.S. and Al-Wosabi, M. 2012. Telling apart ornithopod and theropod trackways: a closer look at a large, Late Jurassic tridactyl dinosaur trackway at Serwah, Republic of Yemen. Ichnos 19: 194–198. Crossref
Taylor, A.M., Gowland, S., Leary, S., Keogh, K.J., and Martinius, A.W. 2014. Stratigraphical correlation of the Lourinhã Formation in the Consolação Sub-basin (Lusitanian Basin), Portugal. Geological Journal 49: 143–162. Crossref
Thulborn, T. 1990. Dinosaur Tracks. 410 pp. Chapman and Hall, London. Crossref
Vialov, O.S. 1988. On the classification of dinosaurian traces [in Russian]. Ežegodnik Vsesoûznogo Paleontologičeskogo Obŝestva 31: 322–325.
Xing, L., Liu, Y.Q., Kuang, H.W., Klein, H., Zhang, J.P., Burns, M.E., Chen, J., Wang, M.W., and Hu, J. 2014. Theropod and possible ornithopod track assemblages from the Jurassic/Cretaceous boundary Houcheng Formation, Shangyi, northern Hebei, China. Palaeoworld 23: 200–208. Crossref
Acta Palaeontol. Pol. 65 (2): 399–412, 2020
https://doi.org/10.4202/app.00707.2019