
Italy’s largest snake: Redescription of Palaeophis oweni from the Eocene of Monte Duello, near Verona
GEORGIOS L. GEORGALIS, LETIZIA DEL FAVERO, and MASSIMO DELFINO
Georgalis, G.L., Del Favero, L., and Delfino, M. 2020. Italy’s largest snake: Redescription of Palaeophis oweni from the Eocene of Monte Duello, near Verona. Acta Palaeontologica Polonica 65 (3): 523–533.
We here redescribe Palaeophis oweni, an almost neglected species of palaeophiid snakes from the Eocene of Monte Duello, northeastern Italy. Despite having been described since the 19th century and being the only known named species of its genus from Mediterranean Europe, P. oweni has so far received only minor and sporadic attention in the literature. The only as yet available information for this species was the original lithograph accompanying its brief, 1881 description. We here provide photographs and detailed description of the type and only known material for the first time, designate a lectotype, and clarify certain nomenclatural issues, as well as inaccuracies in the original lithograph. We distinguish certain features in its vertebral anatomy that allow us to treat P. oweni as a valid species and provide an emended diagnosis. Nevertheless, we highlight that a comprehensive reassessment of the nominal species of Palaeophis from the Eocene of the United Kingdom is essential in order to decipher with certainty the validity and distinctiveness of the Italian form. Palaeophis oweni represents the largest snake known from the Italian fossil record.
Key words: Squamata, Serpentes, Palaeophis, Paleogene, Europe, Italy.
Georgios L. Georgalis [georgios.georgalis@unito.it], Dipartimento di Scienze della Terra, Università di Torino, Via Valperga Caluso 35, 10125 Torino, Italy.
Letizia Del Favero [letizia.delfavero@unipd.it], Museo di Geologia e Paleontologia, Centro di Ateneo per i Musei, Via Giotto 1, 35121, Padua, Italy.
Massimo Delfino [massimo.delfino@unito.it], Dipartimento di Scienze della Terra, Università di Torino, Via Valperga Caluso 35, 10125 Torino, Italy; and Institut Català de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Edifici ICTAICP, Carrer de les Columnes s/n, Campus de la UAB, 08193 Cerdanyola del Vallès, Barcelona, Spain.
Received 3 December 2019, accepted 27 February 2020, available online 8 May 2020.
Copyright © 2020 G.L. Georgalis et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Palaeophiids, and more particularly species of the genus Palaeophis Owen, 1841, are among the first snakes to be described in the fossil record (Owen 1841, 1850). Among them rank some of the largest known snakes, extinct or extant, reaching truly gigantic sizes (Rage 1983a, b; McCartney et al. 2018). Two palaeophiid genera are recognized with certainty, i.e., Palaeophis Owen, 1841 and Pterosphenus Lucas, 1898, ranging from the Late Cretaceous up to the late Eocene and achieving a rather broad geographic distribution, encompassing Europe, Africa, Asia, and North and South America (Owen 1841, 1850; Cope 1869; Marsh 1869; Rochebrune 1880; Lydekker 1888a, b; Andrews 1901, 1906, 1924; Janensch 1906a; Lynn 1934; Arambourg 1952; Hoffstetter 1958, 1960; Holman 1982, 2000; Rage 1983a, b, 1984; Parmley and Case 1988; Averianov 1997; Parmley and Reed 2003; Rage et al. 2003, 2008; Parmley and DeVore 2005; Bajpai and Head 2007; Rage and Dutheil 2008; Houssaye et al. 2013). In addition, the genus Archaeophis Massalongo, 1859, from the Eocene of Italy and perhaps also Turkmenistan (Massalongo 1859; Janensch 1906b; Tatarinov 1963, 1988) has also been tentatively referred to palaeophiids (e.g., Rage 1983a, 1984; Parmley and Reed 2003; Rage et al. 2003; Houssaye et al. 2013; Wallach et al. 2014), though such taxonomic assignment is not fully resolved with certainty.
There is a practically ubiquitous consensus that palaeophiids had aquatic or at least semiaquatic habits (e.g., Hoffstetter 1955, 1958; Rage 1983a, b, 1984; Holman 2000; Rage et al. 2003, 2008; Parmley and DeVore 2005; Houssaye et al. 2013), a fact that had been almost immediately recognized since their first descriptions (Owen 1841, 1850; Cope 1869; Marsh 1869; Rochebrune 1880; Lydekker 1888a, b; Zittel 1887–1890). Their exact phylogenetic affinities within snakes, however, are still rather controversial: they were originally and for a long time considered as close to “booids”, and more particularly pythonids (Owen 1841, 1850; Marsh 1869; Rochebrune 1880; Zittel 1887–1890), a view that thoroughly persisted also during the 20th century (Gilmore 1938; Hoffstetter 1955; Kuhn 1963; Rage 1983a, 1984; Parmley and Case 1988), even as recently as less than a couple of decades ago (Holman 2000). Nevertheless, certain authors during the 20th century highlighted their distinctiveness from all other snakes, placed in their own clade, Cholophidia, along with the Cretaceous pachyophiids (Nopcsa 1923a, b; Hoffstetter 1939; Kuhn 1939; McDowell 1987), while Romer (1956) even casted doubt on their genuine snake affinities, suggesting that they simply might represent “snake-like reptiles”. Others have suggested affinities with the aquatic caenophidian lineage of acrochordids (Nessov 1995; Zvonok and Snetkov 2012; Wallach et al. 2014). Finally, other workers admitted that their exact affinities with other snake lineages cannot be resolved (Hoffstetter 1962), perhaps lying somewhere rather basally within alethinophidians (Rage and Werner 1999). This large uncertainty over their exact phylogenetic affinities undoubtedly rises from the unfortunate fact that both Palaeophis and Pterosphenus are known exclusively from postcranial material (mainly vertebrae); the much smaller-sized Archaeophis is represented by both cranial and postcranial material (Janensch 1906b; Tatarinov 1963, 1988), though the uncertainty about its palaeophiid affinities practically hinders any precise and definite knowledge of the palaeophiid skull.
Palaeophis oweni was established by Zigno (1881) on the basis of several vertebrae from the middle Eocene of Monte Duello, near the city of Verona, in northeastern Italy. Zigno (1881: figs. 9–12) briefly described the material and provided a lithograph of one of the vertebrae. Ever since then, the species was only briefly mentioned in ophidian literature (Kuhn 1939, 1963), and in the past decades, it was only briefly commented that it is not a valid taxon (e.g., Rage 1983b, 1984; Rage and Augé 1993; Delfino 2011; Wallach et al. 2014). We herein redescribe P. oweni and provide photographs of the material for the first time. We compare it with other palaeophiids, provide potential distinguishing features that could enable its distinction from other members of the genus, and discuss its validity.
Institutional abbreviations.—GMH, Geiseltalmuseum of Martin-Luther Universität Halle-Wittenberg, now referred to as the Geiseltalsammlung (part of the Zentralmagazin Naturwissenschaftlicher Sammlungen), Halle, Germany; HNHM, Hungarian Natural History Museum, Budapest, Hungary; MDHC, Massimo Delfino Herpetological Collection, University of Torino, Italy; MGP-PD, Museo di Geologia e Paleontologia dell’Università di Padova, Italy; MNCN, Museo Nacional de Ciencias Naturales, Madrid, Spain; MNHN, Muséum national d’Histoire naturelle, Paris, France; NHMUK, Natural History Museum, London, UK; NHMW, Naturhistorisches Museum, Vienna, Austria; PIMUZ, Paläontologisches Institut und Museum der Universität Zürich, Switzerland; UWr, University of Wrocław, Poland; ZZSiD, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Kraków, Poland.
Material and methods
The material described herein is permanently curated at the collections of MGP-PD. Comparative material of other palaeophiids specimens was studied at the collections of MNHN, NHMUK, and UWr. Comparative material of other extinct and extant snakes was studied at the collections of GMH, HNHM, MDHC, MNCN, MNHN, NHMUK, NHMW, PIMUZ, and ZZSiD.
Geological setting
Monte Duello (also known in the literature under the names Monte Zuello or Montezuello) is a middle Eocene fossil locality in northeastern Italy, near the city of Roncà, in the Province of Verona, Region of Veneto. Its age is considered to pertain to the middle Eocene; more particularly, it was once thought to be late Lutetian (Altichieri 1980; Rage 1983a) but is now considered to be younger, i.e., Bartonian (Frisone et al. 2014; Zorzin and Frisone 2015).
The locality is known already since the second half of the 19th century (Zigno 1875, 1880, 1881, 1889). The assemblage of Monte Duello has yielded a moderately diverse fossil fauna, comprising mostly invertebrates, but also fish, sirenians, and a large bird (Zigno 1875, 1881; Portis 1885). Among reptiles, only P. oweni, the crocodylian Megadontosuchus arduini (Zigno, 1880), and a trionychid turtle (Trionyx cf. marginatus; Zigno 1889), an indeterminate trionychid according to Georgalis and Joyce (2017) have been described so far from Monte Duello (Zigno 1880, 1881, 1889; Piras et al. 2007). The facies association of Monte Duello indicates a sedimentary succession mainly deposited in a shallow water, shoreface setting (Frisone et al. 2014).
Systematic palaeontology
Squamata Oppel, 1811
Serpentes Linnaeus, 1758
Palaeophiidae Lydekker, 1888b
Genus Palaeophis Owen, 1841
Type species: Palaeophis toliapicus Owen, 1841; Isle of Sheppey, United Kingdom; Ypresian, early Eocene.
Palaeophis oweni Zigno, 1881
Figs. 1–3.
Type material: Lectotype (herein designated): MGP-PD 6981Za, an anterior trunk vertebra (Figs. 1, 3). Paralectotypes: 23 trunk vertebrae (MGP-PD 6976Za, 6976Zb, 6977Za, 6977Zb, 6977Zc, 6978Z, 6978Za, 6978Zb, 6978Zc, 6978Zd, 6978Ze, 6978Zf, 6978Zg, 6978Zh, 6979Z (two vertebrae plus several tiny vertebral fragments), MGP-PD 6980Z, 6981Z, 6981Zb, 6981Zc, 6981Zd, 6981Ze, 6981Zf) (Fig. 2), from the type locality and horizon.
Type locality: Monte Duello, near Verona, Veneto, Italy.
Type horizon: Nummulitic limestone, Orizzonte di Roncà, Bartonian, late middle Eocene.
Emended diagnosis.—Palaeophis oweni can be assigned to Palaeophiidae on the basis of its vertebrae being tall and laterally compressed, the presence of pterapophyses, the horizontality of the cotyle-condyle axis, the rather large cotyle and condyle, the presence of a second, small hypapophysis in its anterior trunk vertebrae (i.e., anterior hypapophysis, right ventrally to the cotyle), the distinct ventral projection of the paradiapophyses, the reduced prezygapophyses, and the presence of compressed prezygapophyseal buttresses that form a ridge extending from the dorsal border of the paradiapophyses up to the prezygapophyseal articular facets (characters from Rage 1983a, 1984; Rage et al. 2003; Houssaye et al. 2013). Palaeophis oweni can be referred to the genus Palaeophis on the basis of the relatively small pterapophyses and the neural spine rising posteriorly from the level of the zygosphenal roof (characters from Rage 1984; Parmley and Case 1988; Rage et al. 2003, 2008; Parmley and DeVore 2005; Houssaye et al. 2013). Palaeophis oweni can be differentiated from all other species of the genus Palaeophis by the following combination of features: zygosphene thick and almost trapezoidal in shape when seen in anterior view, with its dorsal margin distinctly convex; cotyle large and slightly, in some cases, laterally compressed; small pterapophyses; massive and elongated paradiapophyses that extend ventrally, well below the ventral level of the cotyle; posterior hypapophysis massive; neural spine high and posteriorly inclined; centrum not much widened anteriorly.
Description.—The lectotype vertebra MGP-PD 6981Za is incomplete, missing most of its neural spine, the posterior portion of the neural arch with large part of both postzygapophyses, and most of its right prezygapophysis and right pterapophysis (Fig. 1). The vertebra is large, with a centrum length of 16.7 mm. In anterior view (Fig. 1A1), the vertebra is tall and distinctly laterally compressed. The zygosphene is thick with its dorsal roof being overall convex and almost triangular in shape. The neural canal is relatively small and trapezoidal. The prezygapophyses (only the left is partially preserved) do not seem to extend much above the ventral floor of the neural canal. There are prezygapophyseal buttresses, in the form of vertical ridges, which extend from the dorsal border of the paradiapophyses up to the prezygapophyseal articular facets. The cotyle is rather large and not particularly compressed. There are no paracotylar foramina, though large depressions are visible at each lateral side of the cotyle. The base of an anterior, moderately thick, hypapophysis is visible below the ventral level of the cotyle. The paradiapophyses are large and elongated, with their ventralmost tip reaching well below the ventral level of the cotyle. In posterior view (Fig. 1A2), the neural arch seems to be relatively vaulted, though this assumption can only be considered as tentative as this respective portion is damaged. The pterapophyses (only the left one is preserved) are relatively small and extend dorsolaterally forming an angle of around 45o with the neural arch. The condyle is large. The posterior hypapophysis is large, with its ventral tip ending well below the level of the condyle. This element is laterally compressed; it is relatively thick at its base but its thickness diminishes gradually towards the level of its ventralmost tip. In dorsal view (Fig. 1A4), the zygosphene is rather concave; there is no sign of a medial lobe. The base of the neural spine runs throughout most of the neural arch, though still it commences posteriorly to the level of the zygosphenal roof. The prezygapophyses extend more anteriorly than laterally. The prezygapophyseal articular facets are rather narrow and small. The interzygapophyseal constriction is relatively shallow. In ventral view (Fig. 1A5), the centrum does not significantly widen anteriorly. Two hypapophyses are present: one large one (posterior) that is situated at the posterior portion of the centrum but still prior to the level of the condyle, and one smaller (anterior), that is situated at the anteriormost portion of the centrum, right below the cotyle. These two hypapophyses are united with a relatively thin, longitudinal ridge. The paradiapophyses are broad and extend anterolaterally. In lateral view (Fig. 1A3, A6), the zygosphenal facets are elongated. The interzygapophyseal ridge is almost straight. The axis of the cotyle and condyle is horizontal. The paradiapophyses are not divided into diapophyseal and parapophyseal portions; they are large, elongated and they project anteroventrally. The posterior hypapophysis develops at the posterior portion of the centrum and projects much ventrally; its posterior border is not vertical with the centrum but rather forms an angle of around 45°. This vertebra is considered to originate from the anterior trunk vertebral column on the basis of the presence of a second (i.e., anterior) small hypapophysis below the cotyle, united with the first (i.e., posterior), large one with a thin longitudinal keel (see Rage et al. 2003).
Apart from the lectotype, all other vertebrae are rather fragmentary (Fig. 2). All vertebrae pertain to the trunk region of the column, with no cloacal or caudal vertebrae known. MGP-PD 6976Za misses most of the neural spine, posterior portion of neural arch, prezygapophyses, postzygapophyses, and pterapophyses, and much of the zygosphene (Fig. 2A). MGP-PD 6976Zb preserves only the ventral portion of the centrum plus the right prezygapophysis (Fig. 2B). MGP-PD 6977Za misses the dorsal part of the neural spine, part of the pterapophyses, and the left prezygapophysis (Fig. 2F). MGP-PD 6977Zb and 6977Zc preserve only the ventral portion of the centrum (Fig. 2G). MGP-PD 6978Z misses the dorsal part of the neural spine and parts of prezygapophyses, postzygapophyses, and pterapophyses, as well as the ventralmost portion of the posterior hypapophysis (Fig. 2D). MGP-PD 6978Za preserves mostly the ventral portion of the centrum (Fig. 2C). MGP-PD 6978Zb misses the right prezygapophysis and pterapophysis, and part of the zygosphene and the neural spine (Fig. 2E). MGP-PD 6978Zc, 6978Zd, 6978Ze, 6978Zf, 6978Zg, and 6978Zh are only vertebral fragments. MGP-PD 6979Z consists of several large and tiny fragments pertaining to at least two vertebrae. MGP-PD 6980Z preserves only the ventral portion of the centrum. MGP-PD 6981Zb is a partial vertebra embedded in matrix, missing its posteroventral portion and pterapophyses (Fig. 2H). MGP-PD 6981Z misses most of the neural arch and neural spine, zygosphene, and zygantrum. MGP-PD 6981Zc is rather eroded specimen embedded on matrix, missing its anterior portion and the dorsal level of the neural spine (Fig. 2J). MGP-PD 6981Zd preserves solely the posteroventral portion of the vertebra along with the zygosphene, embedded on a matrix (Fig. 2I). MGP-PD 6981Ze preserves only the posteroventral portion of the centrum. MGP-PD 6981Zf preserves only remains of the ventral surface of the centrum, condyle, zygantrum, and the neural arch.
The neural spine is not fully preserved in most specimens. When it is partially preserved though, it seems that it develops in height gradually and always well posteriorly from the level of the zygosphenal roof (e.g., MGP-PD 6976Za, 6978Z). It is relatively thick in posterior view (e.g., MGP-PD 6981Zc, Fig. 2J2). The neural spine is most complete in MGP-PD 6981Zb, where it is obvious that it attains a considerable height and a slight posterior inclination (Fig. 2H). The same specimen possesses an unusual rugosity on its neural spine, probably due to some taphonomic/preservational factor. The zygosphene is situated rather high. The zygosphene is relatively thick and convex in anterior view (e.g., MGP-PD 6978Z, Fig. 2D2). Such convexity is prominent at around the mid-length of the zygosphenal roof in anterior view, taking the shape of a triangle (MGP-PD 6981Zd, Fig. 2I; MGP-PD 6981Ze). Zygantrum is damaged in practically most specimens—in fact, the best preserved zygantrum can be observed in MGP-PD 6981Zc, where its roof is rather thick (Fig. 2J2). Pterapophyses are always small. In MGP-PD 6981Zc, the left pterapophysis is almost complete—it is relatively short and moderately thick in posterior view, lying in parallel to the neural spine (Fig. 2J2). Prezygapophyses are small and are not much tilted dorsally; in fact they are almost horizontal with the ventral level of the neural canal in certain specimens (MGP-PD 6978Z, Fig. 2D2). Larger prezygapophyseal articular facets still exist among the material (e.g., MGP-PD 6976Zb, 6978Z), though still they are not considerably large. The postzygapophyses do not extend much laterally (e.g., MGP-PD 6981Zc, Fig. 2J2). The interzygapophyseal constriction is shallow in all specimens. Cotyle is always large, though the degree of its lateral compression may vary. Condyle is also large and its shape varies as well (e.g., Fig. 2B, G2). A (posterior) hypapophysis is present in all specimens. Similarly to the case of the lectotype described above, in anterior trunk vertebrae, there is also a small, second anterior hypapophysis, situated right below the ventral level of the cotyle (e.g., MGP-PD 6978Z, Fig. 2D1; MGP-PD 6978Za, Fig. 2C); specimens that pertain to the mid-trunk or posterior trunk region totally lack an anterior hypapophysis (e.g., MGP-PD 6977Za, Fig. 2F). The shape of the posterior hypapophysis also varies, apparently dependent on the intracolumnar position of the vertebrae. In some vertebrae, it is relatively small and/or does not protrude much ventrally (e.g., MGP-PD 6976Za, Fig. 2A1; MGP-PD 6978Z). In others, it is massive (e.g., MGP-PD 6977Zb, Fig. 2G). Its lateral surface is smooth in most specimens but in one vertebra it is rather rugose, perhaps due to preservation reasons (e.g., MGP-PD 6977Zb, Fig. 2G). Paradiapophyses are much ventrally projected in all specimens (when preserved). However, their shape and anteroventral expansion varies significantly, apparently dependent on the intracolumnar position of the vertebrae. Hence, paradiapophyses can be either relatively slender and anteroventrally directed (e.g., the lectotype MGP-PD 6981Za; MGP-PD 6977Zb, Fig. 2G1) or can be rather massive and face strictly ventrally (e.g., MGP-PD 6981Zb, Fig. 2H). The centra of all vertebrae are only rather slightly anteriorly widened.
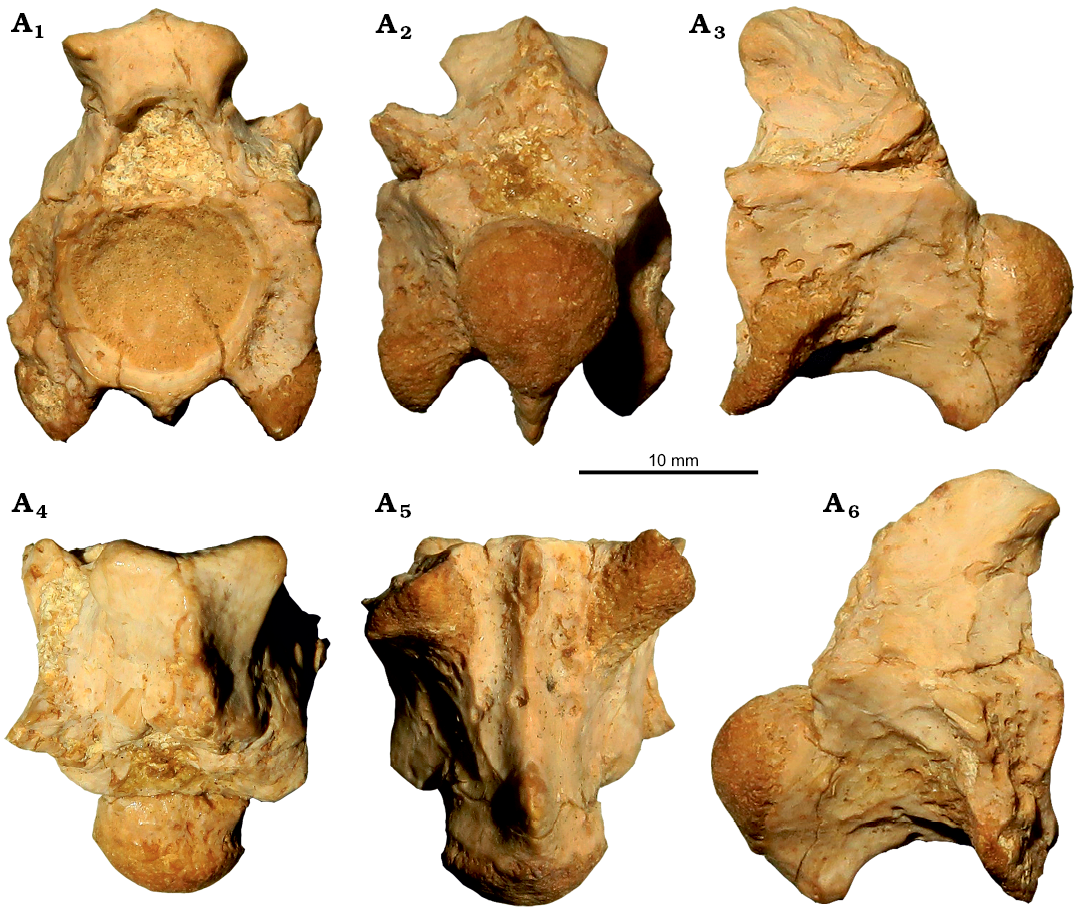
Fig. 1. Lectotype trunk vertebra MGP-PD 6981Za of palaeophiid snake Palaeophis oweni Zigno, 1881, from the late middle Eocene of Monte Duello, in anterior (A1), posterior (A2), left lateral (A3), right lateral (A4), ventral (A5), and dorsal (A6) views.

Fig. 2. Paralectotype trunk vertebrae of palaeophiid snake Palaeophis oweni Zigno, 1881, from the late middle Eocene of Monte Duello. A. MGP-PD 6976Za in left lateral (A1) and ventral (A2) views. B. MGP-PD 6976Zb in posterior view. C. MGP-PD 6978Za in ventral view. D. MGP-PD 6978Z in left lateral (D1), anterior (D2), and ventral (D3) views. E. MGP-PD 6978Zb in left lateral view. F. MGP-PD 6977Za in left lateral (F1), dorsal (F2), ventral (F3), anterior (F4), and posterior (F5) views. G. MGP-PD 6977Zb in left lateral (G1) and posterior (G2) views. H. MGP-PD 6981Zb embedded in matrix in left lateral view. I. MGP-PD 6981Zd in anterior view. J. MGP-PD 6981Zc in right lateral (J1) and posterior (J2) views.
Remarks.—As was a common case with establishments of new species during the 19th century, Zigno (1881) did not designate a holotype for Palaeophis oweni. Although Zigno (1881: figs. 9–12; Fig. 3) figured only one specimen, he clearly indicated that the available material from Monte Duello, consisted of 12 vertebrae; the vast majority of these vertebrae contained rock incretions and only two could be extracted (“Le vertebre di questo Ofidiano, trovate nel calcare nummulitico di m. Zuello, ammontano a dodici, ma la maggior parte di esse era talmente immedesimata nella roccia, che non fu possibile lo estrarne più di due discretamente conservate”; Zigno 1881: 8). It seems that Zigno (1881: 8) focused mostly on these two “better preserved” vertebrae, as he discussed their degree of completeness (“Le porzioni rimaste scoperte di queste due vertebre lascano vedere lo zigosfene, il foro neurico, le diapofisi anteriori, le ipapofisi ed il corpo della vertebra colla concavità anteriore e la protuberanza emisferica posteriore. La spina neurica od apofisi spinosa non è conservata ed il zigantro e le zigapofisi sono in gran parte nascosti dalla roccia”), however, we cannot be certain that he did not base part of his descriptions of P. oweni also on the other, incomplete vertebrae. As such, taking into consideration that Zigno (1881) had worked with more than one specimen upon the establishment of his new taxon, as well as the fact that there is no indication that all vertebrae pertained to the same individual (e.g., they were not articulated), then by definition these specimens have to be considered as syntypes (ICZN 1999: Article 73.2). We here designate the most complete specimen, MGP-PD 6981Za (i.e., the vertebra figured also by Zigno 1881), as the lectotype of the species. Accordingly, the rest of the 12 vertebrae are considered paralectotypes (ICZN 1999: Article 73.2.2). It is unclear to us which is the “second vertebra” (besides the lectotype) mentioned by Zigno (1881), as all of the remaining vertebrae are rather fragmentary—in any case, this specimen apparently is also one of the paralectotypes. The reason that we select to designate as lectotype one of the syntypes is in order to maintain nomenclatural stability that would secure that P. oweni is not a “chimaera” of different species (e.g., in [the unlikely] case that some of the fragmentary vertebrae eventually turn up to belong to a different palaeophiid taxon). We have to also note that in its current state of preservation, the palaeophiid material accompanying the labels with Zigno’s writings (Fig. 4) consisted of 23 fragmentary vertebrae and other tiny vertebral fragments; this number contradicts and surpasses the 12 vertebrae originally mentioned by Zigno (1881). We may tentatively interpret this slight inconsistency in the original number of vertebrae by the probable fact that certain of the incomplete vertebrae were broken into more than one vertebral fragments during the past (almost) 140 years since their original description, or that the smaller fragments were not taken into consideration by Zigno (1881). This view is also supported by the fact that no other palaeophiid material has been subsequently found in Monte Duello (or any other Italian locality) since that time.
The precise authorship date of this taxon is also another issue that needs to be settled. The authorship date of Palaeophis oweni has been continuously erroneously cited as “1882” (e.g., Kuhn 1939, 1963; Rage 1983a, 1984; Rage and Augé 1993; Wallach et al. 2014), however, it is clear from the original publication of Zigno that it was published in 1881. This is further testified also by the author of the species himself, who, in a subsequent work, firmly stated that the respective paper was published in 1881 (“...pubblicai la descrizione e la figura sotto il nome di Palaophis (sic!) Oweni in una memoria stampata nel 1881”; Zigno 1890: 4).
Finally, frustratingly, in his figure captions, Zigno (1881) used a second, erroneous, binomen for his species: Paleophis owenii; this has obviously to be considered as an incorrect spelling (lapsus calami) of both the genus name and the species epithet of Palaeophis oweni!
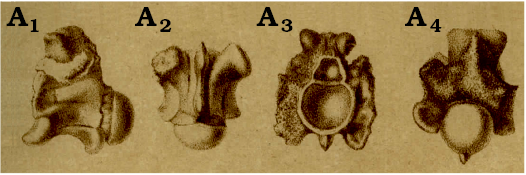
Fig. 3. Original lithograph of the lectotype trunk vertebra MGP-PD 6981Za of Palaeophis oweni as illustrated by Zigno (1881). Specimen illustrated in right lateral (A1), ventral (A2), anterior (A3), and posterodorsal (A4) views. Note the inaccuracies of the lithograph compared with the photographs of this specimen in Fig. 1. Note also that the specimen in (A1) seems like a left (and not right) lateral view of the vertebra, but it is in fact the reverse image, as is the common practice in lithography.

Fig. 4. Original labels accompanying the material of Palaeophis oweni, with the handwriting of Achille De Zigno. The label on the left writes in French: “Paleophis Oweni Zigno. 6976-6978. Éspéce voisine du P. porcatus Ow de Bracklesham. Vertébres trouvé au M. Zuello dan la zone a S. spirulea avec Halitherium Collines du Grumolo a l’ouest de Roncà Veronais”; the label on the right writes in Italian: “Paleophis Oweni Zigno. 6979-6981. Vertebre di un ofidiano trovate nello strato a Halitherium di M. Zuello nei colli di Grumolo all’ovest della V. di Roncà Veronese”.
Note on Zigno’s (1881) lithograph.—It is worth noting that our direct observation of the material allows us to confirm that the depiction of the lectotype vertebra in Zigno’s (1881) lithograph is not fully accurate; among others, most principally, the shape of the zygosphene, the broadness and length of the paradiapophyses, the shape and robustness of the anterior hypapophysis, the height of the zygosphene, and even the shortness of the centrum are depicted erroneously in the original lithograph of Zigno (1881: figs. 9–12; Fig. 3). Admittedly, such inaccuracies in lithographs of snake fossil vertebrae in 19th century’s publications appear to be in fact a rather common case and have been demonstrated for other extinct ophidian taxa named at that time (see discussion in Georgalis et al. 2016); besides, unfortunately, such phenomenon is not confined only to snake descriptions but it has been demonstrated that it is a general issue in lithographs of fossil specimens during the 19th century (e.g., Anquetin and Joyce 2014; Georgalis 2017; Georgalis and Joyce 2017).
Geographic and stratigraphic range.—Known exclusively from the type locality and horizon.
Discussion
Zigno (1881) originally described Palaeophis oweni on the basis of 12 vertebrae, of which he figured in a lithograph only the best preserved specimen (MGP-PD 6981Za), herein designated above as the lectotype of the species. Zigno (1881) provided a description of the material and considered that his new species had more resemblance and was more affiliated with Palaeophis toliapicus, the type species of the genus, from the early Eocene of England (Owen 1841, 1850), from which, nevertheless he distinguished it by its smaller absolute size and the different shape of the paradiapophyses. Zigno (1881) also noted some resemblance of P. oweni, though to a lesser degree, with Palaeophis porcatus Owen, 1850, from the early Eocene of England, a taxon that is currently considered a junior synonym of its sympatric P. typhaeus Owen, 1850, also from the early Eocene of England (Lydekker 1888a; Kuhn 1939, 1963; Rage 1984; Wallach et al. 2014).
Despite being the sole named species of Palaeophis from Mediterranean Europe, as well as one of only two documented occurrences of that genus from the area, rather unfairly and surprisingly, P. oweni has achieved only infrequent and rather sporadic mentions in ophidian literature (e.g., Zigno 1890; Kuhn 1939, 1963). Rage and Wouters (1979) were the first who casted doubt on the validity of this taxon, by considering it as a probable junior synonym of P. toliapicus. Later, Rage (1983a) regarded the Italian taxon as conspecific with Palaeophis typhaeus. This opinion was based on shared features among the two taxa, such as the overall dimensions, the reduced prezygapophyses, the size and shape of the zygosphene and the cotyle (Rage 1983a). That synonymy view was subsequently followed by the rather few workers that ever since mentioned this taxon again (Rage 1984; Rage and Augé 1993; Delfino 2011; Wallach et al. 2014).
We admit that there is resemblance among P. oweni and P. typhaeus. However, P. oweni can be still further differentiated from P. typhaeus by its shorter pterapophyses, more elongated centrum, different shape of the posterior hypapophysis (more robust, thicker, and more ventrally projected in the English species), more elongated paradiapophyses, less dorsally inclined prezygapophyses, more massive cotyle and condyle, thicker zygosphene that is situated rather high, more robust and prominent prezygapophyseal buttresses, less prominent subcentral ridges (see figures in Owen 1850 and Rage 1983; GLG, personal observation at NHMUK, 2017). Overall, it seems that the combination of the thickness, shape, convexity, and high position of the zygosphene, the low inclination of the prezygapophyses, relatively slender paradiapophyses that project much ventrally, and the large size and degree of lateral compression of the cotyle represent characters that could altogether distinguish the Italian form among English (Owen 1841, 1850), Eastern European (Zvonok and Snetkov 2012), and northern African (Arambourg 1952; Houssaye et al. 2013) species of Palaeophis. On the other hand, features such as the shape of the posterior hypapophysis, the thickness of the zygosphene, and the elongation and orientation of the paradiapophyses are intraspecifically and ontogenetically variable characters within palaeophiids and their taxonomic value should be treated with caution (e.g., Parmley and Reed 2003). As such, and pending a complete redescription of Palaeophis typhaeus and the other English species (GLG, in preparation), we tend to tentatively consider P. oweni as a valid, distinct taxon. Such taxonomic opinion may be further supported by the large geographic distance that separates the Italian P. oweni and the English P. typhaeus, as well as their stratigraphic separation (P. oweni is younger); however, it has to be highlighted that there are even cases of other species of Palaeophis that had supposedly achieved even broader distributions (e.g., Palaeophis africanus Andrews, 1924, that is known from both Africa and North America; Parmley and DeVore 2005), while admittedly certain extant aquatic snakes are also broadly distributed (e.g., certain hydrophiids; Hecht et al. 1974; Wallach et al. 2014). Interestingly also, other aquatic reptiles that usually occur sympatrically with palaeophiids are represented by related but still taxonomically distinct species in the Eocene of Italy and England (e.g., trionychid turtles: “Trionyx” capellinii Negri, 1892, in Italy and “Trionyx” henrici Owen in Owen and Bell, 1842, in England; see Georgalis and Joyce 2017).
Regardless of the validity of the species, the identification of Palaeophis in the Eocene of northeastern Italy clearly marks the presence of palaeophiids in Mediterranean Europe, a broad area for which this taxon represents one of the very few known occurrences of its lineage. Practically, if we exclude archaeophiines, which may in fact pertain to a distantly related clade, the sole other known record of that lineage in Mediterranean Europe is represented by indeterminate material of Palaeophis from the middle Eocene locality of Ribaut in southern France (Doncieux and Leriche 1908). On the other side of the Mediterranean, palaeophiids appear to be more abundant and diverse in the Eocene fossil record of northern Africa (Andrews 1901, 1906; Janensch 1906a; Arambourg 1952; Hoffstetter 1960; Rage 1983a; Houssaye et al. 2013; McCartney and Seiffert 2016; Zouhri et al. 2018). Interestingly, both Palaeophis and Pterosphenus are known from northern African sediments, though the latter genus has never been recorded from Europe. Furthermore, P. oweni represents the sole record of palaeophiids from the Alpine region; a purported indeterminate record from the northern side of the Alps, i.e., the Eocene locality of Dielsdorf, Switzerland (Rosselet 1991) was recently shown to pertain in fact to “booids” (Georgalis and Scheyer 2019).
Like other species of its genus, P. oweni is envisaged to be a near shore/estuarine dweller, a life style fully supported by the accompanying marine fauna of the Monte Duello fossil assemblage, as well as sedimentological data (Frisone et al. 2014; see “Geological and palaeoecological setting” above).
Palaeophis oweni is the largest Italian snake. In fact, the rather large size of P. oweni was immediately recognized already in its original description by Zigno (1881: 3) who characterized his new species as “grande Ofidiano”. But how large was indeed P. oweni? Parmley and Reed (2003) suggested methods to assess size estimations for palaeophiid snakes on the basis of isolated vertebrae. Nevertheless, these authors used, as an extant analogue, vertebrae of “booids” (Parmley and Reed 2003), a lineage which is in fact much distantly related to palaeophiids and which has morphologically different vertebrae. More recently, McCartney et al. (2018) applied analytical methods to estimate total lengths of palaeophiids on the basis of vertebral dimensions. Nevertheless, the uncertainty over precise size estimations for palaeophiids is also hindered that the overall individual vertebral count of palaeophiids is as yet totally unknown, although it has been in the past suggested that it should have been rather large, probably surpassing 400 vertebrae (Rage 1983a). As such, we refrain from formally estimating a total length for P. oweni. In any case, the Monte Duello vertebrae clearly indicate a large animal. Indeed, all other snakes from Italy (extinct and extant) are significantly smaller: the largest, potential candidates could be certain “colubrines”, psammophiids, and “Oriental vipers” (Delfino 2002, 2004; Delfino et al. 2011; Colombero et al. 2017) from the Neogene and Quaternary of the country, though still always all their respective vertebral dimensions are far smaller than the Monte Duello giant serpent.
Conclusions
Palaeophis oweni, an almost neglected species of palaeophiid snakes from the Eocene of northeastern Italy, is the focus of this paper. The type and only known material is redescribed in detail and photographs are provided for the first time. A lectotype is designated and certain issues around the nomenclature and authorship of the species are clarified. Comparison with other palaeophiids lead us to distinguish certain vertebral features that can differentiate P. oweni from other species of the genus. An emended diagnosis for the Italian taxon is provided, however, a reassessment of the nominal species of Palaeophis from the Eocene of the United Kingdom is necessary before any definite conclusions can be made about the validity of any species of the genus. Palaeophis oweni is the largest snake documented from Italy; all other snakes from the country, extinct or extant, would certainly be no size match at all for the aquatic giant of Monte Duello.
Acknowledgements
We are grateful to Mariagabriella Fornasiero (MPG-PD) for access to material and providing useful information about De Zigno’s collection. We also thank Stefano Castelli (MPG-PD) for further preparation of specimen MGP-PD 6981Zd. Study of the material at MGP-PD was supported by a University of Torino grant to GLG. GLG acknowledges study of comparative material through the grants SYNTHESYS ES-TAF-5910 (MNCN), SYNTHESYS AT-TAF-5911 (NHMW), SYNTHESYS HU-TAF-6145 (HNHM), and SYNTHESYS GB-TAF-6591 (NHMUK), and the respective curators (Marta Calvo-Revuelta, Silke Schweiger, Judit Vörös, and Sandra Chapman and Mike Day) are highly thanked here. The quality of the manuscript was improved thanks to the useful comments provided by the Editor Daniel Barta (Oklahoma State University Center for Health Sciences, Tahlequah, USA) and the two reviewers Jacob McCartney (State University of New York College at Geneseo, USA) and Alexandra Houssaye (MNHN).
References
Altichieri, L. 1980. Il giacimento di Monte Duello (Verona). In: G. Parisi (ed.), I Vertebrati fossili italiani, catalogo della Mostra, 159–160. Tipografia la Grafica, Vago, Verona.
Andrews, C.W. 1901. Preliminary notes on some recently discovered extinct vertebrates from Egypt. (Part II). Geological Magazine 8: 436–444. Crossref
Andrews, C.W. 1906. A Descriptive Catalogue of the Tertiary Vertebrata of the Fayûm, Egypt. 324 pp. British Museum, London.
Andrews, C.W. 1924. Note on some Ophidian vertebrae from Nigeria. Geological Survey of Nigeria 7: 39–43.
Anquetin, J. and Joyce, W.J. 2014. A reassessment of the Late Jurassic turtle Eurysternum wagleri (Eucryptodira, Eurysternidae). Journal of Vertebrate Paleontology 34: 1317–1328. Crossref
Arambourg, C. 1952. Les vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Notes et Mémoires du Service Géologique du Maroc 92: 1–372.
Averianov, A.O. 1997. Paleogene sea snakes from the eastern part of Tethys. Russian Journal of Herpetology 4: 128–142. Crossref
Bajpai, S. and Head, J.J. 2007. An early Eocene palaeopheid snake from Vastan Lignite Mine, Gujarat, India. Gondwana Geological Magazine 22: 85–90.
Colombero, S., Alba, D.M., D’Amico, C., Delfino, M., Esu, D., Giuntelli, P., Harzhauser, M., Mazza, P.P.A., Mosca, M., Neubauer, T.A., Pavia, G., Pavia, M., Villa, A., and Carnevale, G. 2017. Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and paleoclimatological implications. Palaeontologia Electronica 20.1.10A: 1–66. Crossref
Cope, E.D. 1869. On some Cretaceous reptiles. Proceedings of the Academy of Natural Sciences of Philadelphia 20: 233–242. Crossref
Delfino, M. 2002. Erpetofaune italiane del Neogene e del Quaternario. 382 pp. Ph.D. dissertation, University of Modena and Reggio Emilia, Modena and Reggio Emilia, Italy.
Delfino, M. 2004. The Middle Pleistocene herpetofauna of Valdemino cave (Liguria, north-western Italy). Herpetological Journal 14: 113–128.
Delfino, M. 2011. I rettili fossili italiani. In: C. Corti, M. Capula, L. Luiselli, E. Razzetti, and R. Sindaco (eds.), Fauna d’Italia, Vol. XLV, Reptilia, 57–66. Calderini, Bologna.
Delfino, M., Bailon, S., and Pitruzzella, G. 2011. The late Pliocene amphibians and reptiles from ‘‘Capo Mannu D1 Local Fauna’’ (Mandriola, Sardinia, Italy). Geodiversitas 33: 357–382. Crossref
Doncieux, L. and Leriche, M. 1908. Catalogue descriptif des fossiles nummulitiques de l’Aude et de l’Hérault. Annales de l’Université de Lyon, Nouvelle Série 22: 1–250.
Frisone, V., Pisera, A., Hajdu, E., Preto, N., Zorzi, F., and Zorzin, R. 2014. Isolated spicules of Demospongiae from Mt. Duello (Eocene, Lessini Mts., northern Italy): preservation, taxonomy, and depositional environment. Facies 60: 883–904. Crossref
Georgalis, G.L. 2017. Necrosaurus or Palaeovaranus? Appropriate nomenclature and taxonomic content of an enigmatic fossil lizard clade (Squamata). Annales de Paléontologie 103: 293–303. Crossref
Georgalis, G.L. and Joyce, W.G. 2017. A review of the fossil record of Old World turtles of the clade Pan-Trionychidae. Bulletin of the Peabody Museum of Natural History 58: 115–208. Crossref
Georgalis, G.L. and Scheyer, T.M. 2019. A new species of Palaeopython (Serpentes) and other extinct squamates from the Eocene of Dielsdorf (Zurich, Switzerland). Swiss Journal of Geosciences 112: 383–417. Crossref
Georgalis, G.L., Szyndlar, Z., Kear, B.P., and Delfino, M. 2016. New material of Laophis crotaloides, an enigmatic giant snake from Greece, with an overview of the largest fossil European vipers. Swiss Journal of Geosciences 109: 103–116. Crossref
Gilmore, C.W. 1938. Fossil snakes of North America. Geological Society of North America, Special Papers 9: 1–96.Crossref
Hecht, M.K., Kropach, C., and Hecht, B.M. 1974. Distribution of the yellow-bellied sea snake, Pelamis platurus, and its significance in relation to the fossil record. Herpetologica 30: 387–396.
Hoffstetter, R. 1939. Contribution à l’étude des Elapidæ actuels et fossiles et de l’ostéologie des Ophidiens. Archives du Muséum d’Histoire naturelle de Lyon 15: 1–78.
Hoffstetter, R. 1955. Squamates de type moderne. In: J. Piveteau (ed.), Traité de Paléontologie, Vol. 5, 606–662. Masson, Paris.
Hoffstetter, R. 1958. Un serpent marin du genre Pterosphenus (Pt. sheppardi nov. sp.) dans l’Éocène supérieur de l’Equateur (Amérique du Sud). Bulletin de la Société geologique de France 8: 45–50. Crossref
Hoffstetter, R. 1960. Présence de Pterosphenus (serpent paléophidé) dans l’Eocène supèrieur du bord occidental du désert libyque. Comptes rendus sommaires de la Société géologique de France, Paris 1960: 41.
Hoffstetter, R. 1962. Revue des récentes acquisitions concernant l’histoire et la systématique des Squamates. In: J.-P. Lehman (ed.), Problèmes actuels de paléontologie (Évolution des vertébrés). Colloques internationaux du Centre national de la recherche scientifique, Paris 104: 243–279.
Holman, J.A. 1982. Palaeophis casei, new species, a tiny palaeophid snake from the early Eocene of Mississippi. Journal of Vertebrate Paleontology 2: 163–166. Crossref
Holman, J.A. 2000. Fossil Snakes of North America: Origin, Evolution, Distribution, Paleoecology. 357 pp. Indiana University Press, Bloomington.
Houssaye, A., Rage, J.-C., Bardet, N., Vincent, P., Amaghzaz, M., and Meslouh, S. 2013. New highlights about the enigmatic marine snake Palaeophis maghrebianus (Palaeophiidae; Palaeophiinae) from the Ypresian (Lower Eocene) phosphates of Morocco. Palaeontology 56: 647–661. Crossref
ICZN (International Commission on Zoological Nomenclature) 1999. International Code of Zoological Nomenclature. 4th ed. 306 pp. International Trust for Zoological Nomenclature, London.
Janensch, W. 1906a. Pterosphenus schweinfurthi Andrews und die Entwicklung der Palaeophiden. Archiv für Biontologie 1: 307–350.
Janensch, W. 1906b. Über Archaeophis proavus Mass., eine schlange aus dem Eocän des Monte Bolca. Beiträge zur Paläontologie und Geologie Oestreich-Ungarns und des Orients 19: 1–33.
Kuhn, O. 1939. Squamata: Lacertilia et Ophidia. Fossilium Catalogus. I: Animalia. Pars 86. 89 pp. [Lacertilia] + 33 pp. [Ophidia]. Verlag Gustav Feller, Neubrandenburg.
Kuhn, O. 1963. Serpentes (Supplementum I). Fossilium Catalogus, 1: Animalia. Pars 103. 45 pp. W. Junk, The Hague.
Linnaeus, C. 1758. Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. 824 pp. Laurentii Salvii, Holmiae. Crossref
Lucas, F.A. 1898. A new snake from the Eocene of Alabama. Proceedings of the United States National Museum 21: 637–638. Crossref
Lydekker, R. 1888a. Catalogue of Fossil Reptiles and Amphibians in the British Museum (Natural History). Part I. Containing the Orders Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria. 309 pp. British Museum (Natural History), London.
Lydekker, R. 1888b. Notes on Tertiary Lacertilia and Ophidia. Geological Magazine 5: 110–113. Crossref
Lynn, W.G. 1934. A new snake (Palaeophis virginianus) from the Eocene of Virginia. Johns Hopkins University Studies in Geology 11: 245–249.
Marsh, O.C. 1869. Descriptions of a new and gigantic fossil serpent (Dinophis grandis) from the Tertiary of New Jersey. American Journal of Science, 2nd Series 48: 397–400. Crossref
Massalongo, A. 1859. Specimen photographicum animalium quorumdam plantarumque fossilium agri Veronensis. 101 pp. Vicentinius et Franchinius Excudebant, Veronae.
McCartney, J.A. and Seiffert, E.R. 2016. A late Eocene snake fauna from the Fayum Depression, Egypt. Journal of Vertebrate Paleontology 36: e1029580. Crossref
McCartney, J.A., Roberts, E.M., Tapanila, L., and O’Leary, M.A. 2018. Large palaeophiid and nigerophiid snakes from Paleogene Trans-Saharan Seaway deposits of Mali. Acta Palaeontologica Polonica 63: 207–220. Crossref
McDowell, S.B. 1987. Systematics. In: R.A. Seigel, J.T. Collins, and S.S. Novak (eds.), Snakes: Ecology and Evolutionary Biology, 3–50. Macmillan, New York. Crossref
Negri, A. 1892. Trionici eocenici ed oligocenici del Veneto. Memorie della Società Italiana di Scienze 8: 1–53.
Nessov, L.A. 1995. Paleogene sea snakes as indicators of water mass peculiarities on the east of Tethys Ocean [in Russian]. Vestnik Sankt-Peterburgskogo universiteta, Series 7 2: 3–9.
Nopcsa, F. 1923a. Die Familien der Reptilien. Fortschritte der Geologie und Paläontologie 2: 1–210.
Nopcsa, F. 1923b. Eidolosaurus und Pachyophis. Zwei neue Neocom-Reptilien. Palaeontographica 65: 99–154.
Oppel, M. 1811. Die Ordnungen, Familien und Gattungen der Reptilien als Prodrom einer Naturgeschichte derselben. 87 pp. Joseph Lindauer, Munich. Crossref
Owen, R. 1841. Description of some ophidiolites (Palaeophis toliapicus) from the London Clay of Sheppey, indicating an extinct species of serpent. Transaction of the Geological Society Second Series 6: 209–210. Crossref
Owen, R. 1850. Part III. Ophidia (Palaeophis &c.). In: R. Owen (ed.), Monograph on the Fossil Reptilia of the London Clay and of the Bracklesham and Other Tertiary Beds, 51–63. Palaeontographical Society of London, London. Crossref
Owen, R. and Bell, T. 1849. Part I: Chelonia. In: R. Owen and T. Bell (eds.), Monograph on the Fossil Reptilia of the London Clay and of the Bracklesham and Other Tertiary Beds, 1–76. Palaeontographical Society of London, London. Crossref
Parmley, D. and Case, G.R. 1988. Palaeopheid snakes from the gulf coastal region of North America. Journal of Vertebrate Paleontology 8: 334–339. Crossref
Parmley, D. and DeVore, M. 2005. Palaeopheid snakes from the Late Eocene Hardie Mine local fauna of central Georgia. Southeastern Naturalist 4: 703–722. Crossref
Parmley, D. and Reed, H.W. 2003. Size and age class estimates of North American Eocene palaeopheid snakes. Georgia Journal of Science 61: 220–232.
Piras, P., Delfino, M., Del Favero, L., and Kotsakis, T. 2007. Phylogenetic position of the crocodylian Megadontosuchus arduini and tomistomine palaeobiogeography. Acta Palaeontologica Polonica 52: 315–328.
Portis, A. 1885. Contribuzioni alla Ornitologia Italiana. Memorie della Reale Accademia delle Scienze di Torino, Serie seconda 36: 361–383.
Rage, J.-C. 1983a. Les serpents aquatiques de l’Éocène européen. Définition des espèces et aspects stratigraphiques. Bulletin du Muséum national d’Histoire naturelle, Section C, Sciences de la terre, paléontologie, géologie, minéralogie 4: 213–241.
Rage, J.-C. 1983b. Palaeophis colossaeus nov. sp. (le plus grand Serpent connu?) de l’Eocène du Mali et le problème du genre chez les Palaeopheinae. Comptes Rendus des Séances de l’Académie des Sciences de Paris, II 296: 1741–1744.
Rage, J.-C. 1984. Serpentes. Encyclopedia of Paleoherpetology, Part 11. 80 pp. Gustav Fischer Verlag, Stuttgart.
Rage, J.-C. and Augé, M. 1993. Squamates from the Cenozoic of the western part of Europe. A review. Revue de paléobiologie, Volume spécial 7: 199–216.
Rage, J.-C. and Dutheil, D.B. 2008. Amphibians and squamates from the Cretaceous (Cenomanian) of Morocco. Palaeontographica Abteilung A 285: 1–22. Crossref
Rage, J.-C. and Werner, C. 1999. Mid-Cretaceous (Cenomanian) snakes from Wadi Abu Hashim, Sudan: the earliest snake assemblage. Palaeontologia Africana 35: 85–110.
Rage, J.-C. and Wouters, G. 1979. Découverte du plus ancien Palaeopheidaé (Reptilia, Serpentes) dans le Maestrichtien du Maroc. Geobios 12: 293–296. Crossref
Rage, J.-C., Bajpai, S., Thewissen, J.G.M., and Tiwari, B.N. 2003. Early Eocene snakes from Kutch, Western India, with a review of the Palaeophiidae. Geodiversitas 25: 695–716.
Rage, J.-C., Folie, A., Rana, R.S., Singh, H., Rose, K.D., and Smith, T. 2008. A diverse snake fauna from the early Eocene of Vastan Lignite Mine, Gujarat, India. Acta Palaeontologica Polonica 53: 391–403. Crossref
Rochebrune, A.T. de 1880. Revision des ophidiens fossiles du Museum d’Histoire Naturelle. Nouvelles Archives du Muséum d’Histoire Naturelle, 2ème Série 3: 271–296.
Romer, A.S. 1956. Osteology of the Reptiles. 772 pp. University Chicago Press, Chicago.
Rosselet, C. 1991. Fie fauna der Spaltenfullungen von Dielsdorf (Eozan, Kanton Zurich). Documenta Naturae 64: 1–177.
Tatarinov, L.P. 1963. First occurrence of ancient sea snakes in the USSR [in Russian]. Paleontologičeskij žurnal 1963 (2): 109–115.
Tatarinov, L.P. 1988. Structure of the skull in the sea snake, “Archaeophis” turkmenicus from the lower Eocene of Turkmeniya [in Russian]. Paleontologičeskij žurnal 1988 (1): 75–82.
Wallach, V., Williams, K.L., and Boundy, J. 2014. Snakes of the World: A Catalogue of Living and Extinct Species. 1237 pp. CRC Press, Boca Raton.
Zigno, A. de 1875. Sirenii fossili trovati nel Veneto. Memorie del Reale Instituto veneto di scienze, lettere ed arti 18: 427–456.
Zigno, A. de 1880. Sopra un cranio di coccodrillo scoperto nel terreno eoceno del Veronese. Atti della Reale Accademia Lincei, Memorie della Classe di Scienze Fisiche, Matematiche e Naturali, Serie 3 5: 65–72.
Zigno, A. de 1881. Nuove aggiunte alla fauna eocena del Veneto. Memorie del Reale Istituto Veneto di Scienze, Lettere ed Arti 21: 775–790.
Zigno, A. de 1889. Chelonii scoperti nei terreni Cenozoici delle Prealpi Venete: Memoria. Memorie del Regio Istituto Veneto di Scienze Lettere Arti 23:119–130.
Zittel, K.A. 1887–1890. Handbuch der Paläontologie. Palaeozoologie. III. Pisces, Amphibia, Reptilia, Aves. 900 pp. Druck und Verlag von R. Oldenbourg, Munchen.
Zorzin, R. and Frisone, V. 2015. Montecchia di Crosara, Monte Duello. Il giacimento ricco di spicole di spugna: indagini 2011–2013. Notizie di Archeologia del Veneto 2015: 99–103.
Zouhri, S., Khalloufi, B., Bourdon, E., Lapparent de Broin, F. de, Rage, J.-C., M’Haïdrat, L., Gingerich, P.D., and Elboudali, N. 2018. Marine vertebrate fauna from the late Eocene Samlat Formation of Ad-Dakhla, southwestern Morocco. Geological Magazine 155: 1596–1620. Crossref
Zvonok, E.A. and Snetkov, P.B. 2012. New findings of snakes of the genus Palaeophis Owen, 1841 (Acrochordoidea: Palaeophiidae) from the middle Eocene of Crimea. Proceedings of the Zoological Institute RAS 316: 392–400.
Acta Palaeontol. Pol. 65 (3): 523–533,
2020 https://doi.org/10.4202/app.00711.2019