
Conulariids from the Lower Ordovician of the southern Montagne Noire, France
HEYO VAN ITEN and BERTRAND LEFEBVRE
Van Iten, H. and Lefebvre, B. 2020. Conulariids from the Lower Ordovician of the southern Montagne Noire, France. Acta Palaeontologica Polonica 65 (3): 629–639.
The Tremadocian–Floian (Lower Ordovician) Saint-Chinian, La Maurerie, and Landeyran formations of the southern Montagne Noire (France) collectively contain at least two species of conulariids, namely Archaeoconularia cf. insignis and Conularia azaisi, the latter herein designated as the type species of the new genus, Galliconularia. Archaeoconularia insignis may also occur in the Lower Ordovician Fezouata Shale of southern Morocco, and an indeterminate species of this genus probably occurs in the Lower Ordovician Tonggao Formation of South China. Galliconularia azaisi differs from all other conulariids in having a raised facial midline and very fine, trochoidal transverse ribs which cross the midline ridge without interruption or diminution. In specimens preserving the outermost peridermal lamellae, the transverse ribs bear sub-microscopic nodes, and the broad interspaces are crossed by very slender interspace ridges. Finally, even though the Montagne Noire was part of the western Gondwanan passive margin during Cambro-Ordovician times, G. azaisi remains unknown outside of France.
Key words: Cnidaria, Scyphozoa, Conulariida, Paleozoic, Gondwana, Europe.
Heyo Van Iten [vaniten@hanover.edu], Department of Geology, Hanover College, Hanover, Indiana 47243, USA and Cincinnati Museum Center, Department of Invertebrate Paleontology, 1301 Western Avenue, Cincinnati, Ohio 45203, USA.
Bertrand Lefebvre [bertrand.lefebvre@univ-lyon1.fr], CNRS, UMR 5276 LGLTPE, Université Claude Bernard Lyon 1, 2 rue Dubois, F-69622 Villeurbanne, France.
Received 23 January 2020, accepted 24 March 2020, available online 8 June 2020.
Copyright © 2020 H. Van Iten and B. Lefebvre. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Current knowledge of the fossil record of conulariids, coupled with the results of cladistic analyses of the phylogenetic relationships among genera within this extinct group of medusozoan cnidarians, imply that at least some of these genera diverged from each other during the Neoproterozoic Era (Van Iten et al. 2014, 2016a, b). However, the first appearance of diverse conulariids occurs within strata of the Lower Ordovician Series. In contrast to the underlying Cambrian System, which thus far has yielded only one genus of definite conulariid (Hughes et al. 2000; Van Iten et al. 2006; see however Guo et al. 2020 for a mention of Conulariella Bouček, 1928 from the Cambrian Fortunian Stage of China), Tremadocian–Floian strata collectively contain at least five genera, namely Archaeoconularia Bouček, 1928 (= Exoconularia Sinclair, 1952; Van Iten et al. 2020), Conularia Miller in Sowerby, 1821, Conulariella, Eoconularia Sinclair, 1943, and Teresconularia Leme, Heredia, Rodrigues, Simões, Aceñolaza, and Milana, 2003 (Van Iten et al. 2016b). Most of the currently known occurrences of these genera are in Core Gondwana (Argentina and Morocco) or in peri-Gondwanan areas (Bohemia, South China, and Wales). Additionally, Lower Ordovician strata in the southern Montagne Noire (France; Fig. 1), which was then part of the Gondwanan passive margin at high southern latitudes (Matte 2001; Nysaether et al. 2002; Ballèvre et al. 2009; Álvaro et al. 2013), have yielded rare specimens of a conulariid originally named Conularia azaisi Thoral, 1935. Subsequently, this species was variously assigned to the genera Eoconularia (e.g., Capéra et al. 1978; Vizcaïno et al. 2001) or Paraconularia Sinclair, 1940 (Leme et al. 2003). Most recently, Van Iten et al. (2016b) argued that C. azaisi probably belongs to a separate yet undescribed genus.
In the present paper we erect the new, monotypic genus, Galliconularia, with the type species Conularia azaisi, to accommodate specimens previously named C. azaisi, Eoconularia azaisi, or Paraconularia azaisi. Along with this material our paradigm also includes specimens originally labelled as Conularia sp. indeterminate or ?Eoconularia sp. and reposited in the palaeontological collections of Lyon and Montpellier universities, France (see below). In addition to erecting a new taxon, we describe Archaeoconularia cf. insignis (Barrande, 1867) from the late Floian Landeyran Formation, which has also yielded the medusozoan genus Sphenothallus (Van Iten et al. 2016b). This is the first report of Archaeoconularia in the Montagne Noire. This genus was previously known from the Lower Ordovician of Core Gondwana, Avalonia, and Perunica (Van Iten et al. 2016b).
Institutional abbreviations.—MNHN, Muséum National d’Histoire Naturelle, Paris, France; UCBL-FSL, Lyon 1 University, Villeurbanne, France; UM.ACI, Montpellier University, France.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:D166F2F7-BAFB-4F13-A8D7-5E971383B610
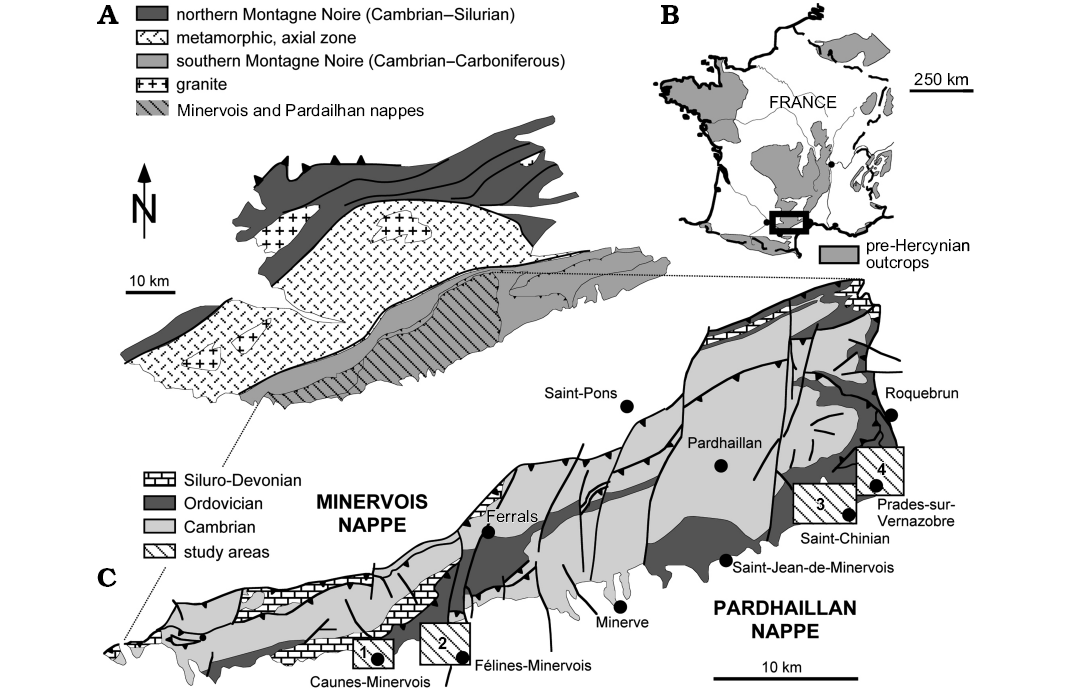
Fig. 1. Geology of the Montagne Noire with conulariid localities. A. Geologic sketch of the Montagne Noire. B. Geologic sketch of France showing the pre-Hercynian rocks and the location of the Montagne Noire (rectangle). C. Simplified geological map of the Minervois and Pardailhan nappes, southern Montagne Noire, with the four main areas yielding Lower Ordovician conulariids (dashed square boxes); 1, western Minervois area, Aude (ruisseau des Lavandières, vallée de l’Ourdivieille); 2, eastern Minervois area, Aude (Brama, vallée du Merlaux); 3, western Saint-Chinianais area, Hérault (Canimals, Donnadieu, Saint-Cels); 4, eastern Saint-Chinianais area, Hérault (Pech Tavernier, les Rocs Nègres, la Rouvelane, les Sources du Foulon). Maps redrawn and modified from Guérangé-Lozes and Burg (1990) and Álvaro and Vizcaïno (2001).
Geological setting
The small amount of available material of Galliconularia azaisi (just 32 specimens collected since the mid 19th century) suggests that it is a relatively rare fossil in the Lower Ordovician of the Montagne Noire. The locality details and stratigraphic positions of most specimens collected between 1850 and 1930 (Azaïs, Lignières, and Villebrun collections) are not known precisely (“Lower Ordovician of Saint-Chinian area”; see Appendix 1). However, the lithologic features of the rocks in which they are preserved make it possible to assign all of them relatively confidently either to the late Tremadocian Saint-Chinian Formation or to the lower part of the early Floian La Maurerie Formation (i.e., the Euloma filacovi and Taihungshania miqueli trilobite zones; Fig. 2). In contrast, the localities and stratigraphic positions of all specimens of G. azaisi collected between 1980 and 2015 (Monceret and Vizcaïno collections) are known precisely (see Appendix 1; Fig. 1). This new material confirms that G. azaisi occurs mainly in the Saint-Chinian Formation (mostly in the Euloma filacovi Zone) and that it is far less common in the overlying La Maurerie Formation (Fig. 2). In both rock units, specimens of G. azaisi are preserved in dark shale or siliceous concretions.
The stratigraphy and palaeontology of the two formations yielding G. azaisi have been thoroughly investigated and regularly updated since the early 20th century (see e.g., Miquel 1912; Thoral 1935; Capéra et al. 1978; Courtessole et al. 1981, 1983, 1985; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003; Tortello et al. 2006; Serpagli et al. 2007). The Saint-Chinian Formation is ~600 m thick and consists mostly of claystone and siltstone with thin beds of sandstone, abundant trace fossils, and numerous fossiliferous concretions. This formation has yielded particularly abundant and diverse invertebrate assemblages dominated by trilobites but also containing various brachiopods, echinoderms, graptolites, hyolithids, machaeridians, and molluscs (Thoral 1935; Capéra et al. 1978; Babin et al. 1982; Courtessole et al. 1983; Vidal 1996; Vizcaïno and Lefebvre 1999; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003; Kröger and Evans 2011). The Saint-Chinian Formation is generally interpreted as consisting of relatively deep shelf deposits that were formed at or slightly below storm-wave base (Vidal 1996; Vizcaïno and Lefebvre 1999; Vizcaïno et al. 2001; Lefebvre 2007).
The overlying La Maurerie Formation is ~400 m thick and consists of numerous thick sandstone beds intercalated within shales bearing fossiliferous concretions. This unit has yielded abundant fossil remains, though these are less diverse than those in the Saint-Chinian Formation. Assemblages are dominated by molluscs and trilobites associated with various brachiopods, echinoderms, graptolites, and machaeridians (Thoral 1935; Capéra et al. 1978; Babin et al. 1982; Courtessole et al. 1983; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003; Kröger and Evans 2011). Strata of the La Maurerie Formation are generally interpreted as shallower in origin than those of the Saint-Chinian Formation, having been deposited at or slightly above storm-wave base (Noffke and Nitsch 1994; Vizcaïno and Lefebvre 1999; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003; Kröger and Evans 2011). No conulariid remains have been reported so far from the overlying Setso, Cluse de l‘Orb, and Foulon formations (Fig. 2; Vizcaïno et al. 2001), all characterized by shallower, storm-dominated deposits (Courtessole et al. 1985).
The occurrence of conulariids referred to Conularia cf. azaisi in the lower part of the Landeyran Formation (Apatokephalus incisus Zone, late Floian) has been regularly mentioned in the literature since its first report by Dean (1966) in a faunal list (see e.g., Capéra et al. 1978; Vizcaïno et al. 2001; Van Iten et al. 2016b). New specimens collected in the 2000s (Kundura collection) confirm the occurrence of conulariids in the lower part of the Landeyran Formation and also extend their stratigraphic range into the upper part of the same unit (Hangchunolithus primitivus Zone, late Floian). Their geographic origin and stratigraphic position are known precisely (see Appendix 1). However, all specimens collected in the Landeyran Formation are assigned herein to Archaeoconularia cf. insignis. Their limited number, in spite of intensive quarrying in Dean’s original sections during the past 50 years, suggests that they were originally relatively uncommon members of the faunal assemblages.
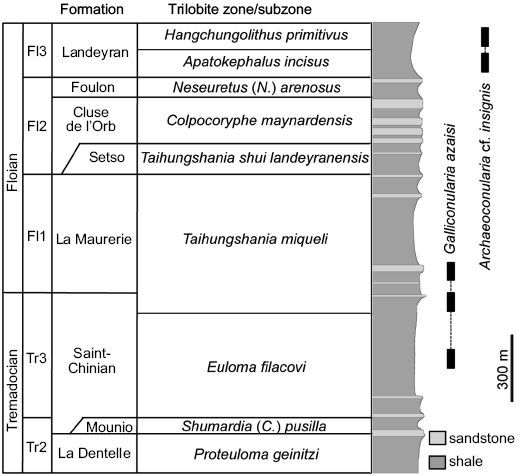
Fig. 2. Stratigraphic range of conulariids in the Lower Ordovician of the southern Montagne Noire. Stratigraphic column redrawn and modified from Courtessole et al. (1983). Stratigraphy based on Vizcaïno et al. (2001), Álvaro and Vizcaïno (2003), Tortello et al. (2006), and Serpagli et al. (2007).
The Landeyran Formation is a 200–400 m thick lithostratigraphic unit consisting of concretion-bearing dark shale conformably overlying the sandstone and shaly sandstone of the underlying Foulon Formation (Capéra et al. 1978; Vizcaïno et al. 2001). The Landeyran Formation has yielded particularly abundant and diverse faunal assemblages dominated by trilobites but also comprising annelids, brachiopods, echinoderms, graptolites, hyolithids, molluscs, and ostracods (Dean 1966; Capéra et al. 1978; Babin et al. 1982; Courtessole et al. 1983; Vizcaïno and Lefebvre 1999; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003). These assemblages are characterized by the re-occurrence of various taxa (e.g., raphiophorid trilobites and stylophoran echinoderms) which are absent in the underlying Cluse de l‘Orb and Foulon formations but are present in the Saint-Chinian and La Maurerie formations (Courtessole et al. 1991; Henry and Vizcaïno 1996; Vizcaïno and Lefebvre 1999). This pattern, which is also exhibited by the conulariids, is in good agreement with the interpretation of the Landeyran Formation as a transgressive unit consisting of relatively distal facies deposited at or below storm-wave base and thus comparable in origin to the Saint-Chinian and La Maurerie formations (Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003; Bonin et al. 2007).
Material and methods
The present report is based on examination of 32 specimens of Galliconularia azaisi and four specimens of Archaeoconularia cf. insignis collectively reposited in the palaeontological collections of Lyon 1 University (France), Montpellier University (France), and the Muséum National d’Histoire Naturelle, Paris (France). The Montpellier specimens were collected in the late 19th and early 20th centuries (Azaïs and Villebrun collections) and correspond to the original material described by Thoral (1935). The Lyon material includes some samples from “old” collections (Lignières and Villebrun collections) but is mostly composed of specimens collected in the late 20th and early 21st centuries (Monceret and Vizcaïno collections). The Paris material consists entirely of specimens collected in the 2000s (Kundura collection).
All of the specimen photographs in this paper are of high-resolution latex casts whitened with ammonium chloride. The photographs were taken with a Canon 5DSR camera mounted with an MP-E 65 mm or 100 mm macro lens.
In addition to examining the material from the southern Montagne Noire, the senior author also examined specimens of Archaeoconularia from the Ordovician of Bohemia and reposited in the paleontological collections of the National Museum of Natural History of the Czech Republic in Prague.
Systematic palaeontology
Phylum Cnidaria Verrill, 1865
Subphylum Medusozoa Peterson, 1979
Class Scyphozoa Götte, 1887
Order Conulariida Miller and Gurley, 1896
Genus Archaeoconularia Bouček, 1939
Type species: Archaeoconularia insignis (Barrande, 1867), Bohemia (Czech Republic), Lower–Upper Ordovician.
Archaeoconularia cf. insignis (Barrande, 1867)
Fig. 3.
1966 Eoconularia? cf. azaisi (Thoral, 1935); Dean 1966: 271.
1978 Eoconularia cf. azaisi (Thoral, 1935); Capéra et al. 1978: 75.
2001 Eoconularia cf. azaisi (Thoral, 1935); Vizcaïno et al. 2001: 219, fig. 3.
2016 Conularia cf. azaisi Thoral, 1935; Van Iten et al. 2016b: 171, table 1.
Material.—Four specimens from the late Floian Landeyran Formation: NHN.F.A71848a, b, A71849, and A71850 from the Apatokephalus incisus Zone; les Sources du Foulon, Cessenon, Hérault; MNHN.F.A71851a, b, from the Hangchungolithus primitivus Zone; les Rocs Nègres, Causses-et-Veyran, Hérault. All Montagne Noire, France.
Description.—Most complete specimen (MNHN.F. A71851a, b; Fig. 3A, 3B, respectively) flattened, lacking the apertural margin but terminating at the apical end in a crumpled schott (apical wall; Fig. 3A2), measuring approximately 31 mm in length, with a maximum single face width of approximately 10 mm and a minimum single face width of approximately 0.5 mm. Faces approximately equal in width, with the corners bounding a given face diverging from each other at approximately 20°. Midline sulcus appears to be deeper than the corner sulcus; both the midline sulcus and the corner sulcus simple (without internal carina or other thickening). Minute, circular to subcircular nodes (Fig. 3B2) on the faces arranged in gently arching, bell-curve-shaped transverse rows and less distinct longitudinal files, with approximately 15–20 transverse rows per mm and approximately 15 longitudinal files per mm.
Remarks.—Of all previously described species of Archaeoconularia (Barrande 1867; Slater 1907; Bouček 1928, 1939; Reed 1933; Sinclair 1943; Sayar 1964; Van Iten et al. 2020), the four incomplete specimens here described from the Landeyran Formation appear to be most similar to A. insignis, a species with relatively small nodes (12–30 per mm; Bouček 1928) that occurs in the late Tremadocian (Lower Ordovician) Mílina Formation of Perunica (Czech Republic; Van Iten et al. 2016b). Both the French specimens and A. insignis are also similar to specimens identified as Archaeoconularia sp. from the late Tremadocian–middle Floian (Lower Ordovician) Fezouata Shale of southern Morocco (Van Iten et al. 2016b) and may be conspecific with them. Like the material from the Landeyran Formation, the Fezouata Shale specimens occur in pale green-brown or pale gray-green mudstone. Additionally, a single specimen illustrated by Van Iten et al. (2013: fig. 3b) from pale green-grey shale in the early Floian (Lower Ordovician) Tonggao Formation of South China, and identified by these authors as “Conulariid sp. and gen. indeterminate” (Van Iten et al. 2013: 719), closely resembles deeply exfoliated specimens of A. exquisita from the Middle or Upper Ordovician of Bohemia (see in particular Bouček 1928: pl. 2: 7); therefore it is probably a specimen of Archaeoconularia. In sum, then, A. insignis or a species closely related to it appears to be present both in the Lower Ordovician of Morocco and the southern Montagne Noire (Gondwana), and Archaeoconularia is also present in the Lower Ordovician of Perunica, Avalonia, and, probably, South China. Collectively, these five Early Ordovician areas spanned a broad range of palaeolatitudes extending from near the South Pole to the southern subtropics (see Van Iten et al. 2013: fig. 5).
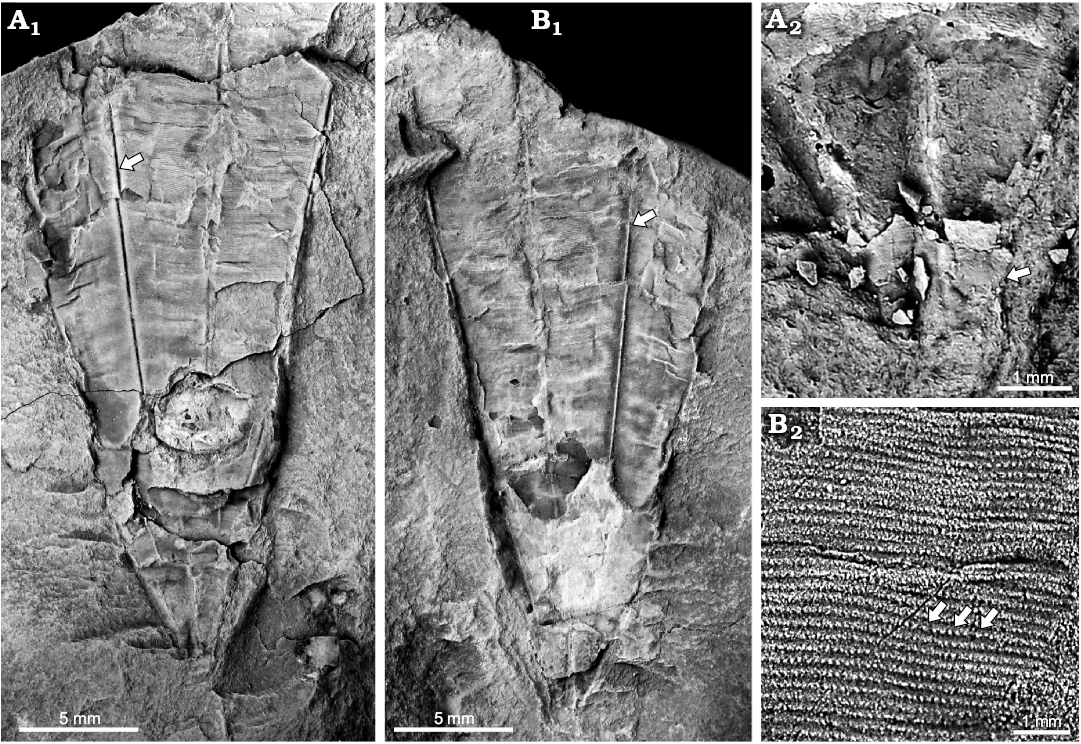
Fig. 3. Conulariid scyphozoan Archaeoconularia cf. insignis (Barrande, 1867), Landeyran Formation, upper Floian, Lower Ordovician; southern Montagne Noire, France. Specimens oriented with the apertural end directed toward the top of the page; arrows in A1 and B1 point to a facial midline. A. The two exposed faces of the part of the most complete specimen (MNHN.F.A71851a), which terminates in a crumpled schott (A1). Detail of the part, arrow pointing to the crumpled terminal schott (A2). B. The counterpart of the specimen in A (MNHN.F.A71851b) (B1). Detail of the apertural region of the counterpart, showing the fine ornament including some of the most clearly defined nodes (arrows) (B2).
Genus Galliconularia nov.
Zoobank LSID: urn:lsid:zoobank.org:act:F7CBF965-787C-4414-89A3- 8CFE1F228C63
Type species: Conularia azaisi Thoral, 1935, by monotypy see below.
Etymology: From the Latin gallicus, meaning of or pertaining to Gaul or France, and Conularia.
Diagnosis.—As for the type species by monotypy.
Remarks.—Galliconularia is a highly distinctive genus, the most conspicuous characteristic of which is the presence of a broadly elevated facial midline together with very fine, trochoidal transverse ribs which are continuous across this feature. Additional notable features of the new genus are the presence on or within the faces of accessory longitudinal furrows or ridges (“filets”; Thoral 1935: 160) and (at least in some specimens) minute nodes and interspace ridges. Only three previously described genera, Climacoconus Sinclair, 1942, Pseudoconularia Bouček, 1928, and Reticulaconularia Babcock and Feldmann, 1986, share with Galliconularia the presence of an external midline ridge (see e.g., Van Iten et al. 2000, 2020). Unlike Galliconularia, however, Pseudoconularia lacks transverse ribs and its nodes are elongate and coarse, and the transverse ribs of Climacoconus and Reticulaconularia are coarse and usually alternate along the facial midline (see e.g., Sinclair 1942: pl. 3: 7). Moreover, no other genus or species exhibits trochoidal transverse ribs as fine as those of Galliconularia, though the pattern of arrangement of its tiny nodes and interspace ridges is similar to that of certain species in Conularia and Paraconularia. Superficially, specimens of G. azaisi exhibiting narrow longitudinal ridges or furrows coupled with adapical recurvature (scalloping) of the transverse ribs (Fig. 4E) resemble the faces of Glyptoconularia Sinclair, 1952, which bears a distinctive ornament consisting of longitudinal files of short, straight, or adaperturally arching transverse ridges collectively resembling chain mail (Van Iten 1994). The fine longitudinal ridges are also similar to the “accessory longitudinal lines” (Sinclair 1952: 139) commonly seen in specimens in the genus Archaeoconularia. Owing in part to deformation of available specimens of Galliconularia, it is difficult to discern details of the anatomy of the corner sulcus. Nevertheless, it appears that like Conularina Sinclair, 1942, Eoconularia, and Glyptoconularia, Galliconularia exhibits termination or severe diminution of the transverse facial ornament in or near the corner sulcus. Importantly, Van Iten et al. (2016b: 1) interpreted this similarity as a non-homoplasic synapomorphy uniting these four taxa in a single, monophyletic group that excludes other conulariids. G. azaisi (shown in the cladogram of Van Iten et al. 2016b as “Eoconularia” azaisi) is the basalmost taxon of this fully resolved clade. According to the phylogenetic analysis of Van Iten et al. (2016b), the midline ridge was derived independently in Galliconularia (coded as Conularia azaisi), Pseudoconularia, and the minor clade consisting of Climacoconus and Reticulaconularia. Trochoidal transverse ribs, a primitive feature present in Galliconularia, Conulariella, and many other conulariids, were lost in at least two lineages, including the lineage leading to the most recent common ancestor of Conularina and Eoconularia. However, Van Iten et al. (2016b) also scored G. azaisi as lacking nodes and interspace ridges, features which were detected in two specimens assigned to this species in the present study. Therefore, the phylogenetic relationships of G. azaisi to other conulariids should be reanalyzed based on the new anatomical data revealed by the present study.
Galliconularia azaisi (Thoral, 1935) comb. nov.
Fig. 4.
1912 Conularia nova sp.; Miquel 1912: 20.
1935 Conularia azaisi Thoral, 1935; Thoral 1935: 159, pl. 11: 7–9, pl. 15: 11a, b.
not 1966 Eoconularia? cf. azaisi (Thoral, 1935); Dean 1966: 271.
1978 Eoconularia azaisi (Thoral, 1935); Capéra et al. 1978: 73.
not 1978 Eoconularia cf. azaisi (Thoral, 1935); Capéra et al. 1978: 75.
2001 Eoconularia azaisi (Thoral, 1935); Vizcaïno et al. 2001: 219, fig. 3.
not 2001 Eoconularia cf. azaisi (Thoral, 1935); Vizcaïno et al. 2001: 219, fig. 3.
not 2001 Eoconularia azaisi (Thoral, 1935); Álvaro et al. 2001: 228.
2004 Eoconularia azaisi (Thoral, 1935); Van Iten and Vhylasová 2004: 122.
2003 Paraconularia azaisi (Thoral, 1935); Leme et al. 2003: 270, fig. 5.
2016 Conularia azaisi Thoral, 1935; Van Iten et al. 2016b: 171, table 1.
not 2016 Conularia cf. azaisi Thoral, 1935; Van Iten et al. 2016b: 171, table 1.
Type material: Thoral’s (1935) four syntypes: UM.ACI.416 (Thoral 1935: pl. 15: 11a, b), UM.ACI.645 (Thoral 1935: pl. 11: 7), UM.ACI.646 (Thoral 1935: pl. 11: 9), and UM.ACI.647 (Thoral 1935: pl. 11: 8). A specimen deposited in the palaeontological collections of western Brittany, Brest, France (LPB 10777) was erroneously mentioned as the holotype of Eoconularia azaisi by Álvaro et al. (2001). This specimen is instead a bivalve (Eopteria crassa [Thoral, 1935]) from the Lower Ordovician of the Montagne Noire (Muriel Vidal, personal communication 2018).
Type locality: Saint-Chinian, Hérault, France.
Type horizon: Euloma filacovi Trilobite Zone, Saint-Chinian Formation, upper Tremadocian, Lower Ordovician.
Material.—28 specimens: UM.ACI.654–656 and 658; UCBL- FSL 133604–133610, 712799, and 713214–713229; from Lower Ordovician of Montagne Noire, France, for details see Appendix 1.
Diagnosis.—Raised facial midline marked by a low, rounded (transversely), continuous external ridge. Exterior-most lamellae of the faces exhibit very fine, adaperturally gently arching, trochoidal (longitudinally) transverse ribs bearing submicroscopic, circular to sub-circular nodes; transverse ribs cross the midline ridge without interruption or diminution, terminating or greatly diminished in height in the narrow corner sulcus. Exterior-most lamellae of the interspaces exhibit very slender interspace ridges that terminate at their apertural end in a node. Deeper lamellae exhibit fine longitudinal ridges or grooves.
Description.—Specimens incomplete, variably deformed and exfoliated, with the largest specimen measuring approximately 58 mm in length; apex unknown, schott (apical wall) not observed, apertural margin gently arcuate but without evidence of inward folding (apertural lappets). Transverse cross section slightly rectangular; corners and facial midline un-thickened and lacking internal carina(e). Angle of divergence of the corners bounding a given face ranges from approximately 10–50°. Corners sulcate; corner sulcus narrow (Fig. 4A1, C). Periderm along the facial midline outwardly folded to form a low, rounded (transversely), continuous ridge that is wider than the corner sulcus (Fig. 4A, C–E). On the faces, the exterior-most lamellae exhibit very fine, gently arched (adaperturally), trochoidal (longitudinally) transverse ribs, 2–11 per mm, which parallel the apertural margin; in two specimens, the transverse ribs exhibit closely spaced, sub-microscopic nodes (approximately 14–18 per mm; Fig. 4A2); transverse ribs cross the midline ridge without interruption or diminution (Fig. 4A2), terminating or greatly diminished in height in the corner sulcus. Interspaces wider than the transverse ribs, in the two node-bearing specimens exhibiting very slender interspace ridges that extend from a node on the transverse rib nearest the aperture to the gap between two nodes on the transverse rib nearest the apex (Fig. 4A2). Faces of many specimens exhibit coarse, irregular folds and/or broadly sinusoidal, gently arched transverse folds that may be artifacts of deformation (Fig. 4C, D). Transverse ribs may be scalloped or jagged, especially in completely flattened specimens preserved in shale (Fig. 4E). Many specimens show multiple fine, commonly discontinuous, longitudinal folds or grooves (“filets”; Thoral 1935: 160) that number about 2 per mm (transversely) and along which the transverse ribs may be recurved adapically.

Fig. 4. Conulariid scyphozoan Galliconularia azaisi (Thoral, 1935), Saint Chinian and La Maurerie formations, upper Tremadocian–lower Floian, Lower Ordovician; southern Montagne Noire, France. Specimens oriented with the apertural end directed toward the top of the page. A. The exterior surface of a single face of a relatively large specimen, parts of which preserve the sub-microscopic nodes and interspace ridges (UCBL-FSL 713229; midline ridge indicated by the arrow) (A1). Detail of the apertural region, showing some of the sub-microscopic nodes and interspace ridges, which cross the broad midline ridge (central part of the photograph) without diminution or disruption (A2). B. Close-up view of the central portion of a single face (corners bounding the face missing) (UM.ACI.645), showing at least six discontinuous, narrow longitudinal folds and furrows (arrows). C. Overall view of the external surface of the single fully exposed face and bounding corners of one the most complete specimens, which is preserved in a siliceous concretion (UCBL-FSL 712799A; midline ridge indicated by the white arrow, corner sulcus by the black arrow). D. View of the counterpart of the specimen in D (UCBL-FSL 712799B; midline ridge indicated by the arrow). E. View of the exterior surface of part of a single face of a flattened specimen, the very fine transverse ribs of which are slightly wavy (scalloped) owing, probably, to tectonic shearing and deformation (UM.ACI.646; midline ridge indicated by the arrow; specimen also figured in Thoral 1935: pl. 11: 9). Scale bars: A1, C, D, E, 10 mm; A2, B, 0.1 mm.
Remarks.—The foregoing description is based on Thoral’s (1935) original material (total of 12 specimens) plus 20 additional specimens. Though longer and more complete than that of Thoral (1935), the present description is generally in close agreement with equivalent portions of Thoral’s (1935) text. The most significant difference between the two descriptions is that Thoral (1935) did not note the presence of minute nodes and interspace ridges. Such features appear to be absent in Thoral’s (1935) material (e.g., Fig. 4C, D), but they are present in at least two of the additional specimens (UCBL-FSL 713215 and 713229) examined by the present authors. In all other respects, the node-bearing specimens are similar to those examined by Thoral (1935), exhibiting for example the same broadly rounded midline ridge and thread-like transverse ribs which cross the midline ridge without diminution or disruption.
As is true for many other conulariids, the appearance of a given specimen of G. azaisi depends in part on the depth or level within the finely lamellar periderm at which the specimen is being examined. This in turn depends on the preservation of the periderm (whether complete or exfoliated) as well as on whether the periderm was split along one or more internal lamellae during extraction from the host rock matrix. The very fine, nodose transverse ribs and interspace ridges appear to be restricted to the outermost lamellae, which in many specimens are either missing (owing, we believe, to exfoliation) or covered by rock matrix, which can only be removed through mechanical preparation (which may destroy delicate external surface features). The fine longitudinal ridges or furrows, which are similar to and possible homologous with those which occur within the periderm of many specimens of Archaeoconularia (see e.g., Bouček 1928: pl. 1: 17), occur in lamellae beneath those bearing the very fine transverse ornament and interspace ridges. In short, then, we do not believe that the two node-bearing specimens and the more numerous specimens that do not exhibit nodes should be assigned to separate species in the new genus. However, if future discoveries should prove this conclusion to be incorrect, then the diagnosis of Galliconularia will have to be emended accordingly.
Another factor affecting the appearance of specimens of G. azaisi is tectonic deformation, more specifically shearing of flattened specimens preserved in shale (Fig. 4E). In addition to being strongly compressed, such specimens differ from more or less three-dimensional specimens preserved in siliceous nodules in having the transverse ribs scalloped or jagged. But even in specimens preserved in siliceous nodules, deformation expressed as very fine wrinkling of the periderm may be present in the interspaces (Fig. 4A2).
Subsequent to the publication of Thoral’s (1935) description, Vizcaïno et al. (2001) listed G. azaisi as Eoconularia azaisi. This assignment apparently was based on the presence in certain specimens of relatively coarse, sinusoidal transverse folds that follow the course of the very fine, adaperturally arching transverse ribs. Such folds resemble the sinusoidal transverse ribs of species such as Eoconularia amoena Sinclair, 1944 and Eoconularia loculata (Wiman, 1895). However, because most specimens of G. azaisi are deformed, we are inclined to agree with Thoral’s (1935) suggestion that the transverse folds in this conulariid are artifacts of directed stress sustained after final burial. In this connection it should be noted that Thoral (1935: 160) grouped the 12 specimens of G. azaisi he examined in two “distinct series”. The first series consisted of specimens having broad faces, large apical angles, poorly defined transverse ribs, and numerous, clearly defined, narrow longitudinal furrows or ridges (“filets”). Thoral did not list the registration numbers of these specimens. The second series consisted of larger and narrower specimens, again unspecified, that were further characterized by well-defined transverse ribs but only a few, widely spaced and discontinuous longitudinal lines. Thoral (1935) provisionally interpreted his two series as consisting, respectively, of the apical (first series) and more distal regions (second series) of an originally more elongate periderm having high apical angles in the apical region and much lower apical angles closer to the apertural end of the periderm. Owing to distortion and incomplete preservation of the faces, we think that determination of apical angles in G. azaisi is more difficult than implied by Thoral (1935); however, at this point we cannot rule out his hypothesis that the two series represent different portions of the periderm of a single species.
Stratigraphic and geographic range.—Late Tremadocian Saint Chinian Formation and early Floian La Maurerie Formation, Lower Ordovician of the southern Montagne Noire, France. See Appendix for detailed geographic and stratigraphic information on each specimen.
Concluding remarks
Archaeoconularia, previously known from the Lower Ordovician of Avalonia (Wales) and Perunica, is now documented from two distinct regions of Core Gondwana: the Anti-Atlas (Morocco) and the southern Montagne Noire (France). The same genus probably also occurs in the Lower Ordovician of South China. The new, monospecific genus Galliconularia is currently known only from the Lower Ordovician of the Montagne Noire. This fact is interesting given that during this time interval, this region was part of the passive margin of Gondwana and, based on palaeogeographic reconstructions (see e.g., Fortey and Cocks 2003; Torsvik and Cocks 2013), was situated close to several other peri-Gondwanan areas from which conulariids have been described (e.g., the Anti-Atlas, Argentina, Bohemia, and Wales; Leme et al. 2003; Van Iten and Vhylasová 2004; Van Iten et al. 2016a).
The diversity pattern observed for conulariids in the Montagne Noire (Fig. 2) is identical to those described for most other invertebrates, in particular echinoderms and trilobites (Vizcaïno and Lefebvre 1999; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003): thus, diversity peaks in the late Tremadocian (Saint-Chinian Formation) before decreasing in the early Floian (La Maurerie Formation) and even more dramatically during the middle Floian (Cluse de l’Orb and Foulon formations). This diversity trend mostly follows large-scale environmental changes, with the Saint-Chinian Formation corresponding to deeper settings (transgressive interval) and the early–middle Floian units recording shallowing conditions (regressive interval).
Not surprisingly, endemicity is particularly reduced during the late Tremadocian transgressive interval, with most invertebrate taxa reported from the Saint-Chinian Formation also occurring in other peri-Gondwanan areas, in particular Argentina and Morocco (Vizcaïno and Lefebvre 1999; Vizcaïno et al. 2001; Vizcaïno and Álvaro 2003; Lefebvre et al. 2016; Martin et al. 2016). In short, then, we predict that the genus Galliconularia probably also occurs in other peri-Gondwanan areas, possibly as one or more new species distinct from G. azaisi.
Acknowledgements
This paper is a contribution to the International Geoscience Programme (IGCP) Project 653—the onset of the Great Ordovician Biodiversification Event. The authors thank Jean-Paul and Marie-Hélène Kundura (Epinay-sous-Sénart, France), Eric Monceret, and Daniel Vizcaïno (Carcassonne, France) for making available and donating their specimens of conulariids from the Montagne Noire to public palaeontological collections (UCBL-FSL and MNHN), as well as for providing useful information on localities and stratigraphy. Suzanne Jiquel (UM.ACI) provided access to the original conulariid material of Thoral, and both Vincent Perrier and Emmanuel Robert (UCBL-FSL) are acknowledged for taking photographs of these specimens. Jean-Michel Pacaud and Peggy Vincent (MNHN) facilitated access to material deposited in their institution. The authors are also grateful to Muriel Vidal (Brest University, France) for her assistance with Montagne Noire specimens deposited in the collections of the University of Western Brittany, Brest, France. The constructive comments of reviewer Graham Young (Manitoba Museum, Winnipeg, Canada) are greatly appreciated. Finally, the senior author thanks Petr Kraft and Jana Bruthansová for assistance in examining specimens of Archaeoconularia in the paleontological collections of the National Museum of Natural History of the Czech Republic in Prague.
References
Álvaro, J.J. and Vizcaïno, D. 2001. The southern Montagne Noire: a key region for theresearch on Lower Paleozoic paleontology. Annales de la Société géologique du Nord 8: 185–189.
Álvaro, J.J., Lefebvre, B., and Vizcaïno, D. 2001. Catalogue of the fossil species definedin the Lower Paleozoic of the Southern Montagne Noire. Annales de la Société géologique du Nord 8: 227–232.
Álvaro, J.J., Zamora, S., Clausen, S., Vizcaïno, D., and Smith, A.B. 2013. The role ofabiotic factors in the Cambrian Substrate Revolution: benthic communityreplacements of West Gondwana. Earth-Science Reviews 118: 69–82. Crossref
Babcock, L.E. and Feldmann, R.M. 1986. Devonian and Mississippian conulariids of North America. Part B. Paraconularia, Reticulaconularia, new genus, andorganisms rejected from Conulariida. Annals of Carnegie Museum 55: 411–479.
Babin, C., Courtessole, R., Mélou, M., Pillet, J., Vizcaïno, D, and Yochelson, E.L. 1982. Brachiopodes (Articulés) et Mollusques (Bivalves, Rostroconches, Monoplacophores, Gastropodes) de l’Ordovicien inférieur (Trémadocien–Arenigien) de la Montagne Noire (France Méridionale). 63 pp. Société des Études Scientifiques de l’Aude, Carcassonne.
Ballèvre, M., Bosse, V., Ducassou, C., and Pitra, P. 2009. Palaeozoic history of the Armorican Massif: models for the tectonic evolution of the suture zones. Comptes-Rendus Geoscience 341: 174–201. Crossref
Barrande, J. 1867. Système Silurien du centre de la Bohême. Première Partie, Tome 3. Classe des Mollusques, Ordre des Ptéropodes. xv + 179 pp. Chez l’auteur et l’éditeur, Prague.
Bonin, A., Nardin, E., Vennin, E., and Vizcaïno, D. 2007. Brachiopod coquinas of the Upper Arenig, Montagne Noire, France. Publications Scientifiques du Muséum 195: 91–93.
Bouček, B. 1928. Révision des Conulaires paléozoïques de la Bohême. Palaeontographica Bohemiae 11: 1–108.
Bouček, B. 1939. Conularida. In: O.H. Schindewolf (ed.), Handbuch der Paläozoologie, Band 2, A113–A131. Gebruder Borntraeger, Berlin.
Capéra, J.C., Courtessole, R., and Pillet, J. 1978. Contribution à l’étude de l’Ordovicien inférieur de la Montagne Noire. Biostratigraphie et révision des Agnostida. Annales de la Société géologique du Nord 98: 67–88.
Courtessole, R., Henry, J.L., and Vizcaïno, D. 1991. Quelques Calymenidae (Trilobita) de l’Ordovicien inférieur (Arenig) de la Montagne Noire, France: systématique, évolution et paléoenvironnements. Palaeontographica Abteilung A 218: 1–15.
Courtessole, R., Marek, L., Pillet, J., Ubaghs, G., and Vizcaïno, D. 1983. Calymenina, Echinodermata et Hyolitha de l’Ordovicien Inférieur de la Montagne Noire (France Méridionale). 162 pp. Société des Études Scientifiques de l’Aude, Carcassonne.
Courtessole, R., Pillet, J., and Vizcaïno, D. 1981. Nouvelles Données sur la Biostratigraphie de l’Ordovicien Inférieur de la Montagne Noire. Révision des Taihungshaniidae, de Megistaspis (Ekeraspis) et d’Asaphopsoides (Trilobites). 32 pp. Société des Études Scientifiques de l’Aude, Carcassonne.
Courtessole, R., Pillet, J., Vizcaïno, D., and Eschard, R. 1985. Étude Biostratigraphique et Sédimentologique des Formations Arénacées de l’Arenigien du Saint-Chinianais Oriental (Hérault) Versant Sud de la Montagne Noire (France Méridionale). 99 pp. Société des Études Scientifiques de l’Aude, Carcassonne.
Dean, W.T. 1966. The Lower Ordovician stratigraphy and trilobites of the Landeyran valley and the neighbouring district of the Montagne Noire, south-western France. Bulletin of the British Museum (Natural History) Geology 12: 247–353.
Fortey, R.A. and Cocks, L.R.M. 2003. Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions. Earth-Sciences Reviews 61: 245–307. Crossref
Götte, A. 1887. Entwicklungs Geschichte der Aurelia aurita und Cotylorhiza tuberculata. Abhandlungen der Entwicklungsgeschichte der Tiere 4: 1–79.
Guérangé-Lozes, J. and Burg, J.M. 1990. Les nappes varisques du sud-ouest du Massif Central. Géologie de la France 1990 (3/4): 71–106.
Guo, J.F., Han, J., Van Iten, H., Song, Z.-C., Qiang, Y.Q., Wang, W.Z., Zhang, Z.F., Li, G.X., Sun, Y.F. and Sun, J. 2020. A new tetraradial olivooid (Medusozoa) from the lower Cambrian (Stage 2) Yanjiahe Formation, South China. Journal of Paleontology 94: 457–466. Crossref
Henry, J.L. and Vizcaïno, D. 1996. Le genre Prionocheilus Rouault 1847 (Trilobita) dans l’Ordovicien inférieur de la Montagne Noire, France: évolution et paléoenvironnements. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 199: 49–64. Crossref
Hughes, N.C., Gunderson, G.O., and Weedon, M.J. 2000. Late Cambrian conulariids from Wisconsin and Minnesota. Journal of Paleontology 74: 828–838. Crossref
Kröger, B. and Evans, D.H. 2011. Review and palaeoecological analysis of the late Tremadocian–early Floian (Early Ordovician) cephalopod fauna of the Montagne Noire, France. Fossil Record 14: 5–34. Crossref
Lefebvre, B. 2007. Early Palaeozoic palaeobiogeography and palaeoecology of stylophoran echinoderms. Palaeogeography, Palaeoclimatology, Palaeoecology 245: 156–199. Crossref
Lefebvre, B., Allaire, N., Guensburg, T.E., Hunter, A.W., Kouraïss, K., Martin, E.L.O., Nardin, E., Noailles, F., Pittet, B., Sumrall, C.D., and Zamora, S. 2016. Palaeoecological aspects of the diversification of echinoderms in the Lower Ordovician of central Anti-Atlas, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 97–121. Crossref
Leme, J.M., Heredia, S., Rodrigues, S.C., Simões, M.G., Aceñolaza, G.F., and Milana, J.P. 2003. Teresconularia gen. nov. from the Lower Ordovician of the Cordillera Oriental of Salta (NW Argentina): the oldest conulariid (Cnidaria) from South America. Revista Española de Micropaleontologia 35: 265–273.
Martin, E.L.O., Vidal, M., Vizcaïno, D., Vaucher, R., Sansjofre, P., Lefebvre, B., and Destombes, J. 2016. Biostratigraphic and palaeoenvironmental controls on the trilobite associations from the Lower Ordovician Fezouata Shale of the central Anti-Atlas, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 142–154. Crossref
Matte, P. 2001. The Variscan collage and orogeny (480–290 Ma) and the tectonic definition of the Armorica microplate: a review. Terra Nova 13: 122–128. Crossref
Miller, S.A. and Gurley, W.F.E. 1896. New species of Palaeozoic invertebrates from Illinois and other states. Illinois State Museum of Natural History, Bulletin 11: 1–50.
Miquel, J. 1912. Nouvel essai sur les terrains primaires du département de l’Hérault. Classification des terrains siluriens. 48 pp. Imprimerie du Commerce, Béziers.
Noffke, N. and Nitsch, E. 1994. Sedimentology of Lower Ordovician clastic shelf deposits, Montagne Noire (France). Géologie de la France 4: 3–19.
Nysaether, E., Torsvik, T.H., Feist, R., Walderhaug, H.J., and Eife, E.A. 2002. Ordovician palaeogeography with new palaeomagnetic data from the Montagne Noire (Southern France). Earth and Planetary Science Letters 203: 329–341. Crossref
Peterson, K.W. 1979. Development of coloniality in Hydrozoa. In: G. Larwood and B.R. Rosen (eds.), Biology and Systematics of Colonial Organisms, 105–139. Academic Press, New York.
Reed, F.R.C. 1933. Some new Ordovician species of Conularia from Girvan. Geological Magazine 70: 354–358. Crossref
Sayar, C. 1964. Ordovician conulariids from the Bosphorus area, Turkey. Geological Magazine 101: 193–197. Crossref
Serpagli, E., Ferretti, A., Vizcaïno, D., and Álvaro, J.J. 2007. A new Early Ordovician conodont genus from the southern Montagne Noire, France. Palaeontology 50: 1447–1457. Crossref
Sinclair, G.W. 1940. The genotype of Conularia. Canadian Field-Naturalist 54: 72–74.
Sinclair, G.W. 1942. The Chazy conulariids and their congeners. Annals of Carnegie Museum 29: 219–240.
Sinclair, G.W. 1943. A new genus of conulariids. Canadian Field-Naturalist 57: 123.
Sinclair, G.W. 1944. Notes on the genera Archaeoconularia and Eoconularia. Royal Society of Canada, Section IV, Transactions, series 3, 38: 87–95.
Sinclair, G.W. 1952. A classification of the Conularida. Fieldiana Geology 10: 135–145.
Slater, I.L. 1907. A monograph of British Conulariae. Palaeontographical Society 61: 1–41. Crossref
Sowerby, J. 1821. The Mineral Conchology of Great Britain; or Coloured Figures and Descriptions of those Remains of Testaceous Animals or Shells, Which Have Been Preserved at Various Times, and Depths in the Earth. 194 pp. W. Arding Co., London.
Thoral, M. 1935. Contribution à l’Étude paléontologique de l’Ordovicien inférieur de la Montagne Noire et Révision Sommaire de la Faune Cambrienne de la Montagne Noire. 362 pp. Imprimerie de la Charité, Montpellier.
Torsvik, T.H. and Cocks, L.R.M. 2013. New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society of London, Memoirs 38: 5–24. Crossref
Tortello, M.F., Vizcaïno, D., and Álvaro, J.J. 2006. Early Ordovician agnostoid trilobites from the southern Montagne Noire, France. Journal of Paleontology 80: 477–495. Crossref
Van Iten, H. 1994. Redescription of Glyptoconularia Sinclair, an Ordovician conulariidfrom North America. In: E. Landing (ed.), Studies in Honor of Donald W. Fisher. New York State Museum & Geological Survey, Bulletin 481: 363–366.
Van Iten, H. and Vhylasová, Z. 2004. Conulariids. In: B.D. Webby, M.L. Droser, M.L., and F. Paris (eds), The Great Ordovician Biodiversification Event, 119–123. Columbia University Press, New York. Crossref
Van Iten, H., Gutiérrez-Marco, J.C., Muir, L.A., Simões, M.G., and Leme, J.M. 2020. Ordovician conulariids (Scyphozoa), from the Upper Tiouririne Formation (Katian), eastern Anti-Atlas Mountains, southern Morocco. Geological Society Special Publications 485 [published online, https://doi.org/10.1016/j.palaeo.2016.03.008]. Crossref
Van Iten, H., Leme, J.M.L., Pacheco, M.L.A.F., Simões, M.G., Fairchild, T.R., Rodrigues, F., Galante, D., Boggiani, P.C., and Marques, A.C. 2016a. Origin and early diversification of phylum Cnidaria: key macrofossils from the Ediacaran System of North and South America. In: S. Goffredo and Z. Dubinsky (eds.), The Cnidaria, Past, Present and Future, 31–40. Springer International Publishing, Switzerland. Crossref
Van Iten, H., Leme, J.M.L., and Simões, M.G. 2006. Additional observations on the gross morphology and microstructure of Baccaconularia Hughes, Gunderson et Weedon, 2000, a Cambrian (Furongian) conulariid from the north-central USA. Palaeoworld 15: 294–306. Crossref
Van Iten, H., Marques, A.C., Leme. J.M.L., and Simões, M.G. 2014. Origin and early evolution of the phylum Cnidaria Verrill: major developments in the analysis of the taxon’s Proterozoic–Cambrian history. Palaeontology 3:1–14.
Van Iten, H., Muir, L.A., Botting, J.P., Zhang Y.-D., and Li, J.-P. 2013. Conulariids and Sphenothallus (Cnidaria, Medusozoa) from the Tonggao Formation (Lower Ordovician, China). Bulletin of Geosciences 88:713–722. Crossref
Van Iten, H., Muir, L., Simões, M.G., Leme. J.M.L., Marques, A.C., and Yoder, N. 2016b. Palaeobiogeography, palaeoecology and evolution of Lower Ordovician conulariids and Sphenothallus (Medusozoa, Cnidaria), with emphasis on the Fezouata Shale of southeastern Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 170–178. Crossref
Van Iten, H., Zhu, Z.-K., and Zhu, M.-Y. 2000. Anatomy and systematics of the Devonian conulariids Changshaconus carinata Zhu, 1985 and Reticulaconularia Babcock and Feldmann, 1986. Acta Palaeontologica Sinica 39: 466–475.
Verrill, A.E. 1865. Classification of polyps (extract condensed from Synopsis of the Polyps and Corals of the North Pacific Exploring Expedition under Commodore C. Ringgold and Captain John Rogers, U.S.N.). Communications of the Essex Institute 4: 145–152.
Vidal, M. 1996. Quelques Asaphidae (Trilobita) de la Formation de Saint-Chinian, Ordovicien inférieur, Montagne Noire (France): systématique et paléoenvironnements. Geobios 29: 725–744. Crossref
Vizcaïno, D. and Álvaro, J.J. 2003. Adequacy of the Early Ordovician trilobite record in the southern Montagne Noire (France): biases for biodiversity documentation. Transactions of the Royal Society of Edinburgh: Earth Sciences 93: 393–401. Crossref
Vizcaïno, D. and Lefebvre, B. 1999. Les échinoderms du Paléozoïque inférieur de la Montagne Noire: biostratigraphie et paléodiversité. Geobios 32: 353–364. Crossref
Vizcaïno, D., Álvaro, J.J., and Lefebvre, B. 2001. The Lower Ordovician of the southern Montagne Noire. Annales de la Société géologique du Nord 8: 213–220.
Wiman, C. 1895. Paleontologische Notizen 1–2. Conularia loculata n. sp. University of Uppsala, Geological Institution, Bulletin 2: 113–117.
Appendix 1
Locality and stratigraphic information for all available specimens of conulariids registered in the collections of Lyon 1 University (UCBL-FSL), Montpellier University (UM), and the Muséum National d’Histoire Naturelle, Paris (MNHN), France:
Saint-Chinian Formation (trilobite zone unknown, upper Tremadocian); locality unknown, Saint-Chinian area (Hérault): UM.ACI. 416, 645, 658 (Azaïs coll.), and UM.ACI.646–647, 654–656, UCBL-FSL 133605–133606 (Villebrun coll.).
Saint-Chinian Formation (Euloma filacovi Zone, upper Tremadocian); Donnadieu, Babeau-Bouldoux (Hérault): UCBL-FSL 713218 (Monceret coll.), and UCBL-FSL 713224–713225 (Vizcaïno coll.); locality unknown, Félines-Minervois area (Hérault): UCBL-FSL 133609 (Lignières coll.); vallée du Merlaux, Félines-Minervois (Hérault): UCBL-FSL 133610 (Lignières coll.); Pech Tavernier, Prades-sur-Vernazobre (Hérault): UCBL-FSL 713216–713217, 713220 (Monceret coll.), and UCBL-FSL713221, 713228 (Vizcaïno coll.); SW of Prades-sur-Vernazobre (Hérault): UCBL-FSL 713222 (Vizcaïno coll.); locality unknown, Saint-Chinian area (Hérault): UCBL-FSL 133608 (Villebrun coll.); Canimals, Saint-Chinian (Hérault): UCBL-FSL 133604 (Villebrun coll.), and UCBL-FSL 712799, 713219 (Monceret coll.); Brama, Trausse (Aude): UCBL-FSL 713223 (Vizcaïno coll.).
Saint-Chinian Formation (lower part of the Taihungshania miqueli Zone, late Tremadocian); ruisseau des Lavandières, Caunes-Minervois (Aude): UCBL-FSL 713226 (Vizcaïno coll.); vallée de l’Ourdivieille, Caunes-Minervois (Aude): UCBL-FSL 713227 (Vizcaïno coll.).
La Maurerie Formation (upper part of the Taihungshania miqueli Zone, lower Floian); la Rouvelane, Prades-sur-Vernazobre (Hérault): UCBL-FSL 713214 and 713215 (Monceret coll.), and UCBL-FSL 713229 (Vizcaïno coll.); Saint-Cels, Saint-Chinian (Hérault): UCBL- FSL 133607 (Villebrun coll.).
Landeyran Formation (Apatokephalus incisus Zone, upper Floian); les sources du Foulon, Cessenon (Hérault): MNHN.F.A71848–A71850 (Kundura coll.).
Landeyran Formation (Hangchunolithus primitivus Zone, upper Floian); les Rocs Nègres, Causses-et-Veyran (Hérault): MNHN.F. A71851 (Kundura coll.).
Acta Palaeontol. Pol. 65 (3): 629–639, 2020
https://doi.org/10.4202/app.00728.2020