
Marine-like isopod Heterosphaeroma priscum from the Late Paleocene freshwater system in Sézanne, France, revisited
NINON ROBIN and RONALD VONK
Well-known for its Palaeocene terrestrial flora, the travertine deposit of Sézanne (Marne, Champagne, France) has also yielded a small number of well-preserved crustaceans, including isopods. In 1868, a single specimen of these isopods has been described by the French zoologist Alphonse Milne-Edwards who remarked on its resemblance to the common benthic isopods from the marine littoral (Sphaeromatidae). A few years later, more material of that same species from Sézanne allowed Charles P.E. Munier-Chalmas to place it in his new genus Heterosphaeroma, but without figuring the species or proposing many diagnostic features. Recently, more material of this species turned up in the Sorbonne collection allowing for a more complete description. Here, we provide a redescription of the Sézanne isopod species based on rediscovered original material. Systematic features are discussed that confirm its sphaeromid affinity. In light of recent knowledge on incursions of the Thanetian Sea in this area, and associated travertine palaeofauna and flora, we assume a high likelihood for this species to have lived in karstic spring waters and freshwater calcareous streams during the end of the Palaeocene. We finally discuss the ability of some Recent sphaeromatids to live in freshwater conditions, deep inland, and we compare their ecology to those found in the Paris Basin travertine palaeoenvironments.
Introduction
Nowadays, the large and predominantly marine isopod genus Sphaeroma consists of over a hundred species, of which around half have been placed in other genera over the years (Boyko et al. 2020). A very small portion of those species, however, are true freshwater inhabitants (Wetzer et al. 2018). The terrestrial plants (mostly Paleocene leaves) of Sézanne have been extensively described by De Saporta (1868), who attributed them to undergrowth in a forest-like environment that harboured a calcareous spring. In his prodrome, De Saporta (1868) also reports mollusks, previously identified by D’Orbigny in Duval and Meillet (1842) and also, for the first time, a small number of well-preserved crustaceans. It is probable that a single specimen was given to Alphonse Milne-Edwards when he described Sphaeroma priscum. When Charles P.E. Munier-Chalmas proposed in 1872 the new combination Heterosphaeroma priscum, he did not specify the number of specimens he examined. Here we re-examined all material now available to elucidate the systematics and the paleoecology of this Paleocene isopod.
Institutional abbreviations.—MNHN.F, Muséum national d’Histoire naturelle de Paris, collection de paléontologie, Paris, France; SU PAL, Sorbonne Université, collection de paléontologie, Paris, France.
Material
The Munier-Chalmas collection of fossil isopods is housed in the Jussieu collection of the Sorbonne University (ten specimens), and Invertebrate Palaeontology collection of the Museum national d’Histoire naturelle (one cast) in Paris, France; it includes original body fossils/imprints embedded in rock matrix and gypsum casts.
Geological setting
The locality of the Montagne des Crottes (or Grottes) northeast of the village of Sézanne (Marne department, Grand-Est region), in the Champagne Valley of France, was reported for the first time in 1842 for the profusion of Paleocene plant imprints it contains (Duval and Meillet 1842; Wegmann 1842). This horizon, consisting of travertine, relates to the fossil-rich lacustrine marl and limestone of Rilly-La-Montagne, 60 km northeast of Sézanne, because of its similar assemblage of molluscs, previously identified by d’Orbigny in Duval and Meillet (1842). The Sézanne travertine was formed in watersprings and running water systems rather than in lakes or rivers (De Saporta 1868). The travertine cascade type of stratification in Sézanne (De Lapparent 1964) reflects a truly freshwater, small stream ecosystem.
The age relationship of these travertines and lacustrine limestone within the encompassing stratigraphy has been subject to debate early on (see De Saporta 1868). A set of pisolitic limestone beds, which underlies the lacustrine layers in the rest of Champagne, is absent in Sézanne. These largely abundant pisolitic beds, covering the very top of the Paris Basin Cretaceous chalky unit, were dated as Dano-Selandian by Hébert (1848) in his exhaustive sedimentological study. In contrast, the Sézanne/Rilly-la-Montagne lacustrine sediments were more difficult to locate and date. They connect with Cenozoic marine and lagoon sandstone: the Bracheux (and other localities) sands. This relationship is important for dating and understanding the actual transgression/regression movements of the Thanetian sea towards Sézanne at the end of the Palaeocene. It has been suggested that these marine sands had been been deposited before the Sézanne/Rilly-la-Montagne lacustrine layer (Hébert 1848), but also right after (D’Archiac 1848; Prestwich 1853). Not much later, however, the latter authors agreed that Bracheux sandstone had been deposited prior to the Rilly lacustrine layers, but were actually not present, and therefore unobservable, in Sézanne outcrops.
These sands, present in many localities west and north of Sézanne, provide evidence of the transgression pulse of the Thanetian sea in the Paris Basin, which had not reached Sézanne. Sea movements followed the stages of the emersion of Paris Basin until the early Eocene when it started to gradually subside under the cover of Cenozoic deposits (Hébert 1848). In Sézanne, the spring and resurgence environment relates to a hot and humid climate (De Saporta 1868; De Lapparent 1964; Pentecost 2005) that acted by way of ample water supply on the underlying chalks to form this terrestrial karstic system at the end of the Thanetian.
The material of Heterosphaeroma priscum from Sézanne comes mostly from Munier-Chalmas (1872) and reportedly was associated with insects and one decapod crustacean of Astacus edwardsi Van Straelen, 1928a. Preserved in similar sediments, both arthropod types apparently show clear freshwater affinities: with (i) insects associated to trichopteran and dragonfly larvae (Nel and Blot 1990), and (ii) a decapod crustacean ascribed to a crayfish genus (Astacus, Astacoidea). The crayfish constitutes the sole report on the east side of the North-Atlantic in the early Paleogene (Cambarus primaevus is known from the Eocene of Wyoming). However, marine influences cannot be totally discarded at this stage. Indeed, it seems some stratigraphic relationships exist between weathering profiles, continental sediments and marine deposits during and in between Thanetian transgressions (see Laurain and Meyer 1986).
Systematic palaeontology
Order Isopoda Latreille, 1817
Suborder Sphaeromatidea Wägele, 1989
Superfamily Sphaeromatoidea Latreille, 1825
Family Sphaeromatidae Latreille, 1825
Genus Heterosphaeroma Munier-Chalmas, 1872
Type species: Sphaeroma priscum Milne-Edwards, 1868 by original designation, monotypic, La Montagne des Crottes, Sézanne, Marne, France, late Thanetian.
Heterosphaeroma priscum (Milne-Edwards, 1868)
Figs. 1–3.
1868 Sphaeroma priscum; Milne-Edwards 1868: 426, 2 figures.
1872 Heterosphaeroma priscum; Munier-Chalmas 1872: 166.
1928 Heterosphaeroma priscum; Van Straelen 1928b: 40.
1969 Heterosphaeroma priscum; Moore and Brooks 1969: 377.
1977 Heterosphaeroma priscum; Secrétan 1975: 319.
2014 Heterosphaeroma priscum; Giusberti et al. 2014: 81.
Type material: Holotype: SU.PAL.2017.0.2.61 (Munier-Chalmas old collection), outer mold (Fig. 1A). Despite the existence of several known specimens as early as 1868, it is clear that Alphonse Milne-Edwards had only seen a single specimen when he provided the species description. He writes: “The false abdominal legs of the last pair are lacking in the exemplar I could examine” (Milne-Edwards 1868: 427) leaving no doubt in that matter. Consequently, SU.PAL.2017.0.2.61 is the holotype, which originally consisted of both the outer and inner molds, the latter being now lost. The fossils in Sézanne are preserved as voids in the travertine. Munier-Chalmas recovered these fossils by pouring gypsum/stearine solution and subsequently dissolved the calcareous rock in acid leaving moulds for investigations. Consequently, the specimens SU.PAL.2017.0.0.4 and SU.PAL.2017.0.2.4, exist only in the form of casts and the original specimen has been dissolved.
Type locality: Sézanne, Marne department, Grand-Est region, France.
Type horizon: Upper Thanetian, Palaeocene, Cenozoic.
Material.—MNHN.F.B71849, plaster cast of lost SU.PAL.2017. 0.2.61 (inner mold, Munier-Chalmas old collection) made by André Tournouër in 1882; SU.PAL.2017.0.2.4–6, 0.2.11, 0.2.12, 0.3.9, 0.2.3.1, 0.2.3.2-1, 0.2.3.2-2 (Munier-Chalmas old collection).
Emended diagnosis.—Body oblong to ovoid in dorsal view, length 17 mm, width 10 mm, greatest width at pereonites 4 and 5; cephalon separate from pereonite 1, semi-enclosed by it laterally; pleonites narrow, four in number with first one mostly covered by seventh pereonite, fourth pleonite broadened mid-dorsally, covering or fusing with others at mid-dorsal part; pleotelson regularly rounded with an inflated mid-dorsal section, upper margin with S-form hinge locks on both sides making strong connection to fourth pleonite; uropods with broad and flat, not clearly surpassing in length lower rim of pleotelson, exopod on robust peduncle, much smaller than endopod with apparently smooth outer margin.
Description.—Cephalon semi-ovate and surrounded laterally by the first pereonite. Ophthalmic ridge visible. Eyes with lower half extended posteriorly to the anterior margin of first pereonite (Figs. 1C, 2B). Antennule with robust peduncle segments and much thinner flagellum of which separate segments cannot be seen, end covered by matrix. Antenna with peduncle partly visible, no separations between segments visible. Buccal mass (Fig. 1C) protruding out strongly from well-defined perimeter at the underside of cephalon. Frontal lamina not fused with clypeus but coming forward from mid-frontal margin of cephalon (Fig. 4A). Clypeus triangular with clear lateral lobes pointing downward and semi-enclosing labrum. Labrum semi-triangular and prominent, frontal margin slightly concave and downward part tapering toward blunt triangular point with rounded posterior margin. Mandible visible, showing palp on left side which is directed forward with geniculate last segment. Maxilla 1 (maxillule) (Fig. 1C), 2 (maxilla), and maxilliped of normal sphaeromatid form insofar their outlines can be seen. Basis of maxilliped L-shaped (with retinacula that lock opposing plate and visible endopodite). Pereopod 1 (second thoracopod) (Fig. 1C) with sharp triangular coxal plate and clear suture where plate connects to thoracic segment. Basis slender, broken off at ischium, rest of the leg not preserved. Pereopod 2 (Figs. 1C, 2A) with half of basis preserved. Coxa suture not as prominent as in pereopod 1. Pereopods 3 to 6 (Figs. 2A, 3A) slender, preserved partly on both sides, no protruding lobes of separate segments present, pereopod 7 without basal segments and with only long dactylus and claw present. Pleonites (Figs. 1A, B, 2C), four in number interrupted in middle on dorsal side by enlargement of fourth pleonite. Fourth pleonite has on both sides a hinging and locking process connected with pleotelson. Pleotelson (Fig. 1A, B) robust and bulbous in middle with one smooth surfaced bulge protruding mid-dorsally. Apex rounded without indentations, tunnel-shaped in fossil imprint. Uropods prominent (Fig. 1A), endopod broad and flat, exopod much smaller, not well preserved, condition of outer margin not discernible.

Fig. 1. Dorsal views of isopod Heterosphaeroma priscum (Milne-Edwards, 1868), late Thanetian of Sézanne, Marne, France. A. SU.PAL.2017.0.2.61 (outer mold, holotype). B. MNHN.F.B71849 (plaster cast of lost SU.PAL.2017.0.2.61, inner mold). C. SU.PAL.2017.0.2.3.1. D. SU.PAL.2017.0.2.3.2. Abbreviations: pln, pleonite. Scale bars 3 mm. Photographs Lilian Cazes (MNHN) and Rémi Brageu (SU).
Remarks.—The fourth pleonite extension overlapping the other three visible pleonites dorsally (Figs. 1A, B, 2C) seems a unique character not reported from other fossil Sphaeromatoidea. There are extant species with four pleonites visible dorsally that also have the sutures not completely spanning the entire mid-dorsal region such as Exosphaeroma waitemata Bruce, 2005 and species in the genus Gnorimosphaeroma Menzies, 1954. However, in extant species five pleopods are always present on the ventral side and no overlapping structure extending from the fourth pleonite is completely covering the others. Species of the enigmatic genus Parvireia Chilton, 1925, provisionally attributed to the Sphaeromatidae (Brökeland et al. 2001), also show four pleonites dorsally but have five functional pleopods. Comparisons to other, extant isopods living in travertine depositing conditions do not feature similar pleonite configurations. The presence of a medial tube-like folding (= tunnel) (Fig. 1A, B) on the posterior margin of the pleotelson is typical for sphaeromatoideans (Jones et al. 2014).
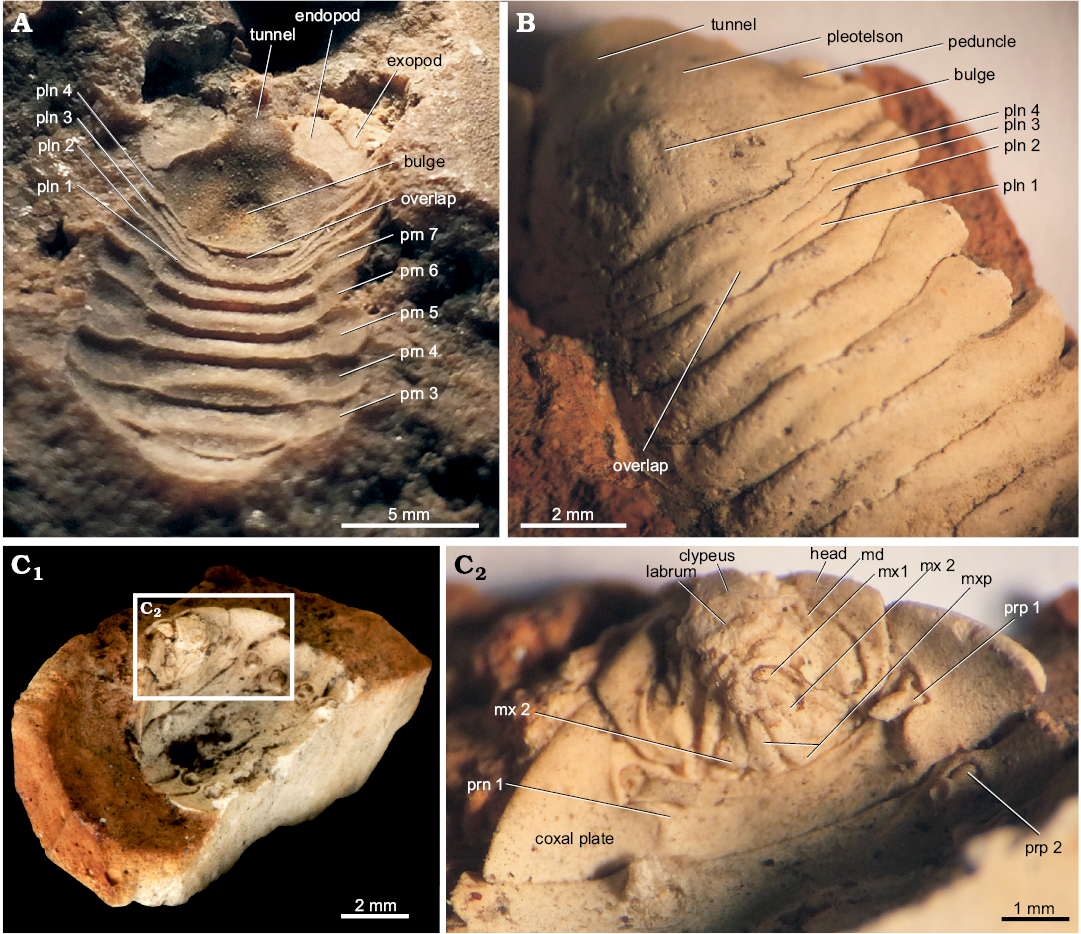
Fig. 2. Detailed anatomy of isopod Heterosphaeroma priscum (Milne-Edwards, 1868), late Thanetian of Sézanne, Marne, France. A. SU.PAL.2017.0.2.61 (inner mold, holotype), in dorsal view. B. SU.PAL.2017.0.0.4 (plaster cast), in dorsal view. C. SU.PAL.2017.0.2.4 (C1, body fossil anterior half; C2, head close-up), ventral side. Abbreviations: md, mandibular palp; mxp, maxilliped; mx, maxilla; pln, pleonite; prn, pereonite; prp, pereopod. Photographs RV and Rémi Brageu (SU).
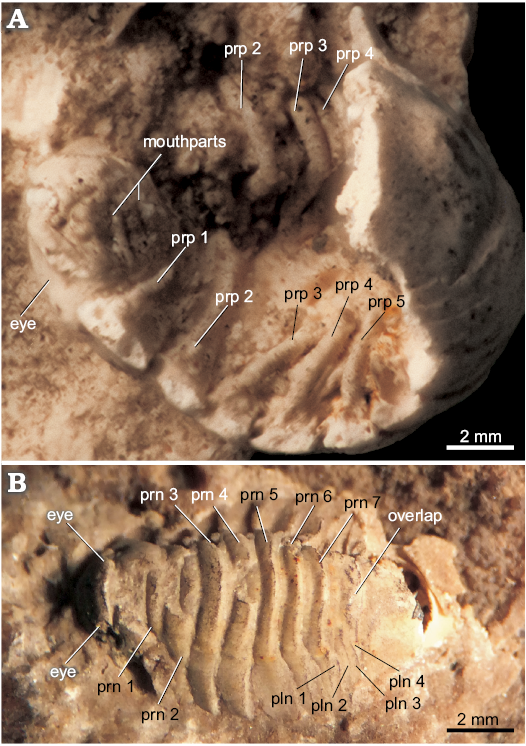
Fig. 3. Appendage views of isopod Heterosphaeroma priscum (Milne-Edwards, 1868), late Thanetian of Sézanne, Marne, France. A. SU.PAL.2017. 0.2.12 (plaster cast). B. SU.PAL.2017.0.2.3.2-2. Abbreviations: pln, pleonite; prn, pereonite; prp, pereopod. Photographs RV and Rémi Brageu (SU).
Concluding remarks
Sphaeromatid isopods occur nowadays mostly in marine habitats where they are benthic and fossorial detritus feeders. Already Milne-Edwards (1868) questioned the marine affinity of Heteropshaeroma when he compared it to species of Sphaeroma while making his short note on this fossil. He mentioned the resemblance of Sphaeroma priscum to common marine isopods. Heterosphaeroma priscum displays a morphology that nowadays is known only in marine taxa, therefore it remains disputable whether it was a true freshwater isopod or if it was a marine/euryhaline species that had been trapped in layers formed under mixed freshwater and marine influences. There are a few other fossil Sphaeromatidae that have unresolved marine—freshwater affinities (Quaggiotto and De Angeli 2016; Vonk et al. 2015), but also undoubted freshwater taxa such as Archaeosphaeroma frici Novák, 1872 from lower Miocene freshwater chalks in central Europe.
Crucial arguments against freshwater origin of Heterosphaeroma priscum come from better understanding of the Mezo-Cenozoic Champagne stratigraphy as presented by Laurain and Meyer (1986) who improved the resolution of the upper Thanetian transgression-regression cycles thanks to new key field observations. They identified the relationships between (i) altered chalk facies, (ii) concretion-type facies and (iii) detrital facies. They demonstrated that the Thanetian transgression had a larger extension towards the south-east (i.e., up to Sézanne, where beach pebbles cover the chalk surface) than previously known, and connect the travertine deposits to two different subfacies: terrestrial and stromatolitic. The latter usually display a thin stratification, encompassing darker filaments when seen at high magnification. Correlated to the activity of cyanobacteria, the formation of this part of the travertines in Champagne may be linked to shallow-water sea conditions, as confirmed by perforations ascribed to marine borers (gastrochaenid molluscs, Laurain and Meyer 1986: 122–123, pl. 2: 6). In Sézanne, no stromatolite travertine were reported by the authors: only terrestrial travertines laying over the reworked Cretaceous pebbles, marking the end of the transgression (Laurain and Meyer 1986).

Fig. 4. Detailed anatomy of isopod Heterosphaeroma priscum (Milne-Edwards, 1868), late Thanetian of Sézanne, Marne, France. A. SU.PAL.2017. 0.2.3.1, ventral view. B. SU.PAL.2017.0.2.3.2-1, dorsal view and leaf co-occurrence (dashed), isopod individuals (arrows). C. SU.PAL.2017.0.2.3.2-2, dorsal view. Abbreviations: pln, pleonite; plp, pleopod; prn, pereonite; prp, pereopod. Photographs RV and Rémi Brageu (SU).
We compared the preservation of Heterosphaeroma priscum and the sedimentary matrix it is hosted in, with that of the other arthropods described from Sézanne. The travertine pieces display a high concretional density with dot-like structures, which are in all aspects similar to the matrix hosting insects figured by Nel and Blot (1990). These travertines, bearing adult true bugs (Hemiptera) but mostly dragonfly/trichopteran larvae, had to form close to or in the same freshwater streams. Isopod preservation is also comparable to that of Astacus edwardsi Munier-Chalmas, 1872 likely originating from the same locality; and described by Van Straelen (1928a) as an European representative of freshwater Astacoidea. SU.PAL.2017.0.2.3.2-1 (Fig. 4B) evidences the synlocation of the isopods with the plant/leaf-rich travertine of Sézanne, showing on the same surface a complete isopod and the leaf of the woody vine Cissus cissoides (De Saporta, 1868) Hollick, 1936. Considering the above arguments, it is plausible to propose a freshwater/waterspring environment for Heterosphaeroma priscum in the Thanetian of Champagne rather than any admixture of marine/brackish waters from a tidal creek in the karstic surface system.
Freshwater species related to Sphaeroma form probably independent lineage that made the transition from the marine environment to inland waters on several times, as was concluded by Wetzer et al. (2018) based on molecular studies of Gnorimosphaeroma Menzies, 1954, Lekanesphaera Verhoeff, 1942, and Thermosphaeroma Cole and Bane, 1978. Recent sphaeromids living in hot springs that provide calcifying conditions are known from the genus Thermosphaeroma, with seven species. Most of these forms are omnivorous, as implied by the scarce (vegetation) food supply of hot spring environments. The species of Thermosphaeroma have never been observed to prey on aquatic insects living in the same springs (Shuster 1981). Some species are even known to have cannibalistic behaviour. On the other hand, the springwaters usually completely exclude fish and avian predation risks, possibly permitting high densities of these isopods and decreasing the risks associated with male mate-searching behaviour of reproduction (Shuster 1981). The absence of modern sphaeromatids in more common epigean freshwater systems, such as streams, ponds, rivers and lakes, has been attributed to failure in escaping predaceous aquatic insects and competition with several species of larval insects (Bowman 1981; Shuster 1981). In Sézanne, the preservation and lithology of the fossil predaceous insects (adults and larvae) indicate spring/streams habitats shared with Heterosphaeroma priscum.
Acknowledgements.—We thank Stéphane Jouve (SU) and co-workers for help and access to the Sorbonne Université, collection of palaeontology. We thank Jean-Michel Pacaud (MNHN) and Sylvain Charbonnier (Centre de Recherche en Paléontologie, Paris, France) for support in finding correspondences between SU and MNHN collections, as well as Denis Audo (Yunnan University, Kunming, China) for discussion about material locations. We are grateful to Rémi Brageu (Laboratoire d’Archéologie Moléculaire et Structurale, SU) and Lilian Cazes (Centre de Recherche en Paléontologie, Paris, France) for providing part of the fossil pictures. We finally thank Rodney Feldmann (Kent State University, Ohio, USA) and Christopher Boyko (Hofstra University, Hempstead, USA) for reviewing our manuscript and contributing to its improvement through their respective inputs.
References
Bowman, T.E. 1981. Thermosphaeroma milleri and T. smithi, new sphaeromatid isopod crustaceans from hot springs in Chihuahua, Mexico, with a review of the genus. Journal of Crustacean Biology 1: 105–122. Crossref
Boyko, C.B., Bruce, N.L., Hadfield, K.A., Merrin, K.L., Ota, Y., Poore, G.C.B., Taiti, S., Schotte, M., and Wilson, G.D.F. (eds.) 2020. World Marine, Freshwater and Terrestrial Isopod Crustaceans Database. Accessed through: World Register of Marine Species at: http://www.marinespecies.org/aphia.php?p=taxdetails&id=257071 on 2020-02-10
Brökeland, W., Wägele, J.W., and Bruce, N. L. 2001. Paravireia holdichi n. sp., an enigmatic isopod crustacean from the Canary Islands with affinities to species from New Zealand. Organisms Diversity & Evolution 1: 83–98. Crossref
Bruce, N. 2005. New sphaeromatids (Crustacea: Isopoda: Sphaeromatidea) from coastal and freshwater habitats in New Zealand. Zootaxa 1002: 1–20. Crossref
Chilton, C. 1925. Some Amphipoda and Isopoda from the Chatham islands. Records of the Canterbury Museum 2: 317–320.
Cole, G.A. and Bane, C.A. 1978. Thermosphaeroma subequalum, n. gen., n. sp. (Crustacea: Isopoda) from Big Bend National Park, Texas. Hydrobiologia 59: 223–228. Crossref
D’Archiac, E.J.A. 1848. Histoire des progrès de la géologie. Tome II, Part 2. 600 pp. Société Géologique de France, Paris.
De Saporta, G. 1868. Prodrome d’une flore fossile des travertins anciens de Sézanne. Mémoires de la Société Géologique de France 3: 1–436.
De Lapparent, A.F. 1964. Région de Paris. Excursions géologiques et voyages pédagogiques. 2eme edition. 196 pp. Hermann, Paris.
Duval, G. and Meillet, C. 1842. Coupe des terrains des environs de Sézanne. Bulletin de la Société Géologique de France 1: 100–104.
Hébert, E. 1848. Notice sur les dépôts situés dans le bassin de Paris, entre la craie blanche et le calcaire grossier. Bulletin de la Société Géologique de France 2: 388–410.
Hollick, C.A. 1936. The tertiary floras of Alaska. U.S. Geological Survey Professional Paper 182: 1–185.
Giusberti, L., Fornasiero, M., and Zorzin, R. 2014. The Pesciara-Monte Postale fossil-Lagerstätte: 4. the “minor fauna” of the laminites. Rendiconti della Società Paleontologica Italiana 4: 73–87.
Jones, W.T., Feldmann, R.M., and Garassino, A. 2014. Three new isopod species and a new occurrence of the tanaidacean Niveotanais brunnensis Polz, 2005 from the Jurassic Plattenkalk beds of Monte Fallano, Italy. Journal of Crustacean Biology 34: 739–753. Crossref
Latreille, P.A. 1817. Le règne animal distribue d’après son organisation, pour servir de base a l’histoire naturelle des animaux et d’introduction a l’anatomie comparée, pas M. le Cher. Cuvier. Tome III, contenant les crustacés, les arachnides et les insectes. 653 pp. Deterville, Paris.
Latreille, P.A. 1825. Familles naturelles du règne animal, exposé succinctement et dans un ordre analytique avec l’indication de leurs genres. 570 pp. J.B. Baillière, Paris. Crossref
Laurain, M. and Meyer, R. 1986. Stratigraphie et paléogéographie de la base du paléogène champenois. Géologie de la France 1: 103–123.
Menzies, R.J. 1954. The comparative biology of reproduction in the wood-boring isopod crustacean Limnoria. Bulletin of the Museum of Comparative Zoology 112 (5): 364–388.
Munier-Chalmas, C. 1872. Sur des crustacés, des fleurs et des insectes découverts dans les Travertins lacustres de Sézanne. Bulletin de la Société Géologique de France 2: 166.
Milne Edwards [sic!], A. 1868. Sur une nouvelle espèce de crustacé aquatique découvert à Sézanne. In: G. de Saporta (ed.), Prodrome d’une flore fossile des travertins anciens de Sézanne. Mémoires de la Société Géologique de France 2: 426–427.
Moore, R.C. and Brooks. H.K. 1969. Treatise of Invertebrate Palaeontology. Vol. Part R, Arthropoda 4, Volume 1. 398 pp. Geological Society of America, Boulder.
Nel, A. and Blot, P. 1990. Paléoentomologie de la paléotufiere écocene de Sézanne (Marne, France) (Insecta Odonata, Trichoptera, Hemiptera, Diptera). Entomologica Gallica 2: 26–31.
Novák, O. 1872. Über eine neue Isopoden-Gattung aus dem tertiären Süsswasser-Kalk von Waltsch. Sitzungsberichte der Königlich Böhmischen Gesellschaft der Wissenschaften, Mathematisch-naturwissenschaftliche Klasse 1872: 39–45.
Pentecost, A. 2005. Travertine. 446 pp. Springer Science & Business Media, Berlin.
Prestwich, J. 1853. Sur la position geologique des sables et du calcaire lacustre de Rilly. Bulletin de la Société Géologique de France 2: 300–310.
Quaggiotto, E. and De Angeli, A. 2016. Fauna delle marne fitolitiche dell’Oligocene della Valle del Ponte (Lusiana, Vicenza, Italia settentrionale). Natura Vicentina 19: 15–36.
Secrétan, S. 1975. Les crustacés du Monte Bolca. Studi e Ricerche sui Giacimenti Terziari di Bolca 2: 427–431.
Shuster, S.M. 1981. Life history characteristics of Thermosphaeroma thermophilum, the Socorro isopod (Crustacea: Peracarida). The Biological Bulletin 161: 291–302. Crossref
Van Straelen, V. 1928a. Astacus edwardsi Munier-Chalmas Ms., Astacidae du Paleocene de Sézanne (Champagne). Bulletin de la Société Géologique de France 2: 3–6.
Van Straelen, V. 1928b. Contribution à l’étude des isopodes méso- et cénozoïques. Mémoires de l’Académie Royale de Belgique. Classe scientifique Série 2 9: 1–66.
Verhoeff, K.W. 1942. Sphaeromiden – Studien und buchnerillo N.G. 83. Isopoden-aufsatz. Zeitschrift für Morphologie der Tiere und Ökologie der Tiere 39: 153–175. Crossref
Vonk, R., Latella, L., and Zorzin, R. 2015. Eocene isopods of Pesciara di Bolca (Italy) revisited. Journal of Crustacean Biology 35: 540–546. Crossref
Wägele, 1989. Evolution und phylogenetisches System der Isopoda: Stand der Forschung und neue Erkenntnisse. Zoologica 140: 1–262.
Wegmann, C.-E. 1842. Séance du 21 Novembre 1842. Bulletin de la Société Géologique de France 1 (14): 70.
Wetzer, R., Bruce, N., and Pérez-Losada, M. 2018. Relationships of the Sphaeromatidae genera (Peracarida: Isopoda) inferred from 18S rDNA and 16S rDNA genes. Arthropod Systematics and Phylogeny 76: 1–30.
Ninon Robin [ninonrobin23@gmail.com], School of Biological, Earth & Environmental Sciences, University College Cork, Butler Building, North Mall, T23 TK3 Cork, Ireland.
Ronald Vonk [ronald.vonk@naturalis.nl] (corresponding author), Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands; Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, P.O. Box 94248, 1090 GE Amsterdam, The Netherlands.
Received 10 May 2020, accepted 3 June 2020, available online 27 August 2020.
Copyright © 2020 N. Robin and R. Vonk. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 65 (4): 793–798,
2020 https://doi.org/10.4202/app.00767.2020