
A new hermit crab out of its shell from the Eocene Arguis Formation, Huesca, Spain
FERNANDO A. FERRATGES, SAMUEL ZAMORA, and MARCOS AURELL
Semi-articulated paguroids are rare fossils, and there are only few records from Cenozoic strata. Here we present a new and exceptionally preserved hermit crab (Diogenidae) from the Eocene of Huesca (Spain) that preserves the anterior part of the carapace, together with appendages. Diogenes augustinus sp. nov. represents one of most completely preserved hermit crabs known to date, providing crucial information to understand the evolution of the family Diogenidae. It is characterized by poorly marked regions of shield, absence of Y-line and markedly unequal and robust chelipeds. The specimen is preserved out of its host shell suggesting rapid burial in siliciclastic strata of a prodeltaic environment.
Introduction
Hermit crabs (superfamily Paguroidea Latreille, 1802) are an iconic group of decapod crustaceans with very specialized characteristics that usually include a poorly mineralized and asymmetric abdomen. This is the result of a life confined to empty gastropod shells (e.g., Walker 1992), or in cavities of sponges, corals and plant remains (e.g., De Forges et al. 2001). Due to the weakly mineralised body and articulations, the fossil record of paguroids is poor and often dominated by isolated chelipeds and other dissociated remains (e.g., Hyžný et al. 2016; Fraaije et al. 2020, and references therein).
Examples of articulated hermit crabs in the fossil record are rare, and only few nearly complete specimens are known; in most cases preserved in situ, inside gastropods or other shells (see Fraaije and Polkowsky 2016 and Pasini et al. 2020, who summarized main records). Because they hide naturally inside shells, hermit crabs usually reveal only a small part of their anatomy (for instance, chelipeds and sectioned pereiopods) or occur as incomplete remains with articulated chelipeds (e.g., Schweitzer and Feldmann 2001; Fraaije et al. 2011). So far, for only seven specimens, the anterior part of the carapace (shield), in association with, chelipeds have been recorded (Jagt et al. 2006; Garassino and Schweigert 2006; Fraaije et al. 2008; Garassino et al. 2009; Schweigert et al. 2013; Fraaije and Polkowsky 2016; Pasini et al. 2020). This information allows a better understanding of how chelipeds are associated with body fossils. Here we describe an extraordinary example of a paguroid from the Eocene Arguis Formation (South Pyrenean Basin, northeast Spain) that is preserved out its host shell and retains a partial carapace, plus chelipeds and walking legs.
Institutional abbreviations.—MPZ, Museo de Ciencias Naturales de la Universidad de Zaragoza, Spain.
Nomenclatural acts.—This published work and the nomenclatural acts it contains, have been registered in Zoobank: urn:lsid:zoobank.org:pub:1B4F2880-02E2-4D94-958F-4E30C973536F
Geological setting
The present material comes from sandstone and marly beds in the lower part of the Arguis Formation (middle–upper Eocene), as exposed along the southern margin of the Jaca-Pamplona Basin, between the villages of Belsué and Lúsera (Huesca) (co-ordinates 42º18’54” N, 0º20’41” W). This basin, located in the south-central Pyrenean zone (Sierras Exteriores), formed in consequence of the propagation of the Pyrenean tectonic structures during the Paleogene (Millán et al. 1994; Huyghe et al. 2009). This propagation during the middle and late Eocene resulted in the formation of reliefs that became part of sediment source areas for deltaic complexes (Dreyer et al. 1999).
The Arguis Formation (latest Lutetian–early Priabonian) consists of four deepening-shallowing third-order sequences, I to IV (Millán et al. 1994). Specifically, the paguroids described come from prodeltaic sandstone found in the lower part of Sequence II as described by Millán et al. (1994). The late Bartonian age of Sequence II is well constrained by magnetostratigraphical and palaeontological data (Pueyo et al. 2002; Silva-Casal et al. 2019). The Arguis Formation includes highly fossiliferous intervals, with benthic foraminifera, decapod crustaceans, echinoderms, and molluscs (e.g., Ferratges et al. 2020).
Systematic palaeontology
Systematic classification follows McLaughlin (2003) and carapace terminology follows Fraaije et al. (2019).
Order Decapoda Latreille, 1802
Infraorder Anomura MacLeay, 1838
Superfamily Paguroidea Latreille, 1802
Family Diogenidae Latreille, 1802
Genus Diogenes Dana, 1851
Type species: Pagurus miles Fabricius, 1787, by subsequent designation of Stimpson (1859); Eocene–Recent.
Diogenes augustinus sp. nov.
Figs. 1, 2.
Zoobank LSID: urn:lsid:zoobank.org:act:FFF586B9-DBC5-455B-A 38A-DD3011628FB9
Etymology: In honour of Agustín Gálvez (Zaragoza, Spain) who found and donated the specimen.
Type material: Holotype: MPZ2020/54, articulated specimen, partial carapace, plus chelipeds and walking legs. Paratypes: MPZ2020/55 and MPZ2020/56, isolated chelipeds. From the type locality and horizon.
Type locality: Village of Lúsera, Huesca Province, Spain.
Type horizon: Arguis Formation, upper Bartonian, middle–upper Eocene.
Material.—Type material only.
Diagnosis.—Shield longer than wide, length/width ratio 1.32, slightly convex transversely and longitudinally; rostrum not prominent. Orbital cavity concave, shallow antennal cavities. Regions of shield poorly marked; massetic region elongated, with perpendicular striate. Anterior branchial area globose, subtriangular. Eyestalks smooth, shorter than shield, diameter 1.42 mm. Chelipeds markedly unequal, left largest; palm quadrate, without stridulatory mechanism on mesial face of palm. Carpus triangular; axis of carpal-propodal articulation oblique in relation to sagittal plane. P2 merus depressed on outer side. Dorsolateral faces of propodus covered with small granules. Dactyls of P2 and P3 about same length as propodus, margin smooth.
Measurements (in mm).—MPZ2020/54: shield length 10.74; shield width 8.08; eyestalk length 7.63; left manus length 15.12; left manus height 14.29; right manus length 8.01; right manus height 7.05. MPZ2020/55: major cheliped height 19.35; length 17.74. MPZ2020/56: major cheliped height 17.33.
Description.—Shield longer than wide, slightly domed; surface roughened by shallow grooves, except for most of gastric region which is smooth with a subtle longitudinal ridge; rostrum weakly developed; anterior margin weakly convex; orbital cavity shallow; post-ocular projection acute; post-antenal projection slightly rounded; central gastric furrow present, but weakly developed; massetic groove weakly marked; massetic region elongated, with perpendicular stretch marks; keraial region poorly differentiated; gastric regions flat; U-shaped cervical groove; posterior carapace not well preserved (Fig. 1A). Eyestalks slightly longer than half length of shield. Chelipeds markedly unequal, with the left considerably larger; without stridulatory mechanism developed on the mesial face of the palm. Major cheliped: palm longer than broad, lateral surface of carpus covered with densely small granules; upper margin with pointed conical granules; weakly granulated dorso-ventrally (Fig. 1). Minor cheliped: palm covered by small and conic wide-spaced granules, upper margin with five pointed teeth. Merus and carpus of both chelipeds less granulated than palm; subtriangular fixed finger broad at base, with rounded distal extremity; fingers curved ventromesially; moveable finger longer than mesial margin of palm. Ambulatory legs symmetrical; merus subrectangular and elongate, upper margin convex, less ornamented than carpus and propodus; outer side of P2 merus depressed to accommodate P3; carpus subtriangular; carpus and propodus of P2–P3 with slightly serrated dorsal margins. Dactylus of ambulatory legs as long as propodus, broadly curved (Figs. 1, 2).
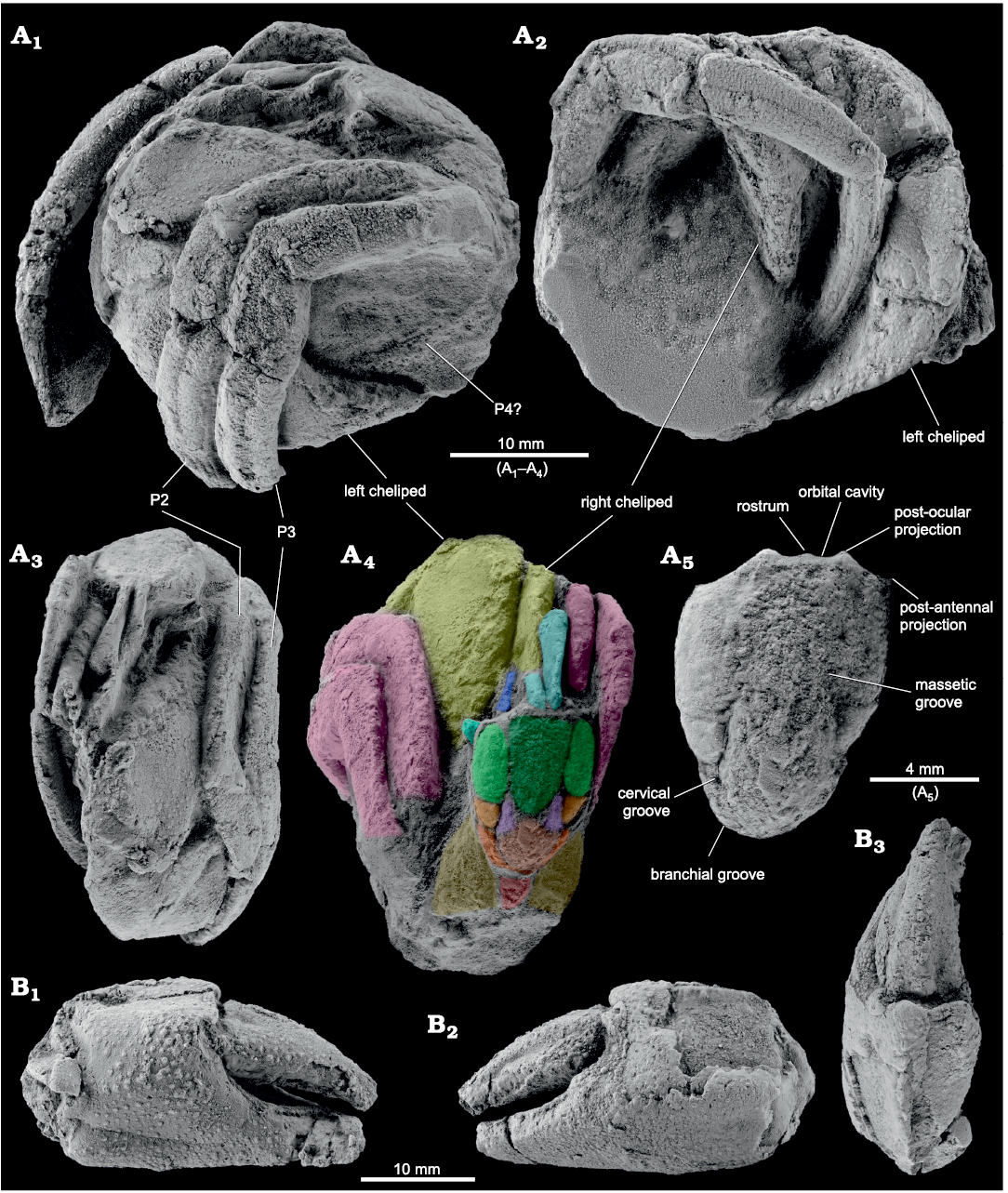
Fig. 1. Paguroid crab Diogenes augustinus sp. nov. from the upper Bartonian of Arguís Formation, Eocene, Lúsera, Huesca, Spain. A. Holotype (MPZ2020/54), in left lateral (A1), right lateral (A2), and frontal (A3) views. Dorsal region details (A4): light blue, eyestalk; dark blue, maxilliped?; turquoise, antenna; light green, massetic; dark green, anterior gastric; orange, lateral branchial; purple, keraial; light orange, anterior branchial; dark orange, mesobranchial; dark yellow, metabranchial; red, cardiac. Shield details (A5). P2, P3, P4?, pereiopods. B. MPZ2020/55, paratype, isolated left cheliped in mesial (B1), outer (B2), and dorsal (B3) views. Specimens have been photographed dry and coated with an ammonium chloride sublimated.
Remarks.—Reconciliation of fossils and neontologist classifications are problematic, because most of the taxonomic features to differentiate between modern genera are not preserved in extinct material (e.g., Jagt et al. 2006; Fraaije 2014; Fraaije et al. 2014). This drawback is exacerbated by the fact that the majority of fossil paguroids are found as disarticulated elements or, in the best cases, they are hidden inside gastropod shells; and this hampers our ability to provide a complete description of specimens. This explains why the systematic placement of fossil paguroids is fraught with difficulties (Fraaije and Polkowsky 2016). Fortunately, the material described herein includes both the chelipeds and the carapace and represents one of the few examples in which all of the anterior portion of the animal is available.
Despite these drawbacks, McLaughlin (2003) listed important morphological characters to distinguish between different modern taxa that can also be recognised in fossil species. For instance, Fraaije et al. (2019), among others, included detailed data on shields of extant taxa that can be seen in fossils.
The material studied retains delicate diagnostic characteristics, such as the morphology of the shield, cheliped ornamentation, ocular peduncles and ambulatory legs. This allows its definite assignment to the family Diogenidae. We include the new taxon in Diogenes on the basis of similarities in features of chelipeds, pereiopods and distribution of dorsal carapace regions. Our material also shows corrugated and poorly defined massetic regions, a gastric region without Y-line and eyestalks that are long and thin (Fig. 2). Diogenids of Eocene age include nine genera that have been described on the basis of isolated chelae only (Garassino et al. 2009; De Angeli and Caporiondo, 2017), making comparison with our material difficult.
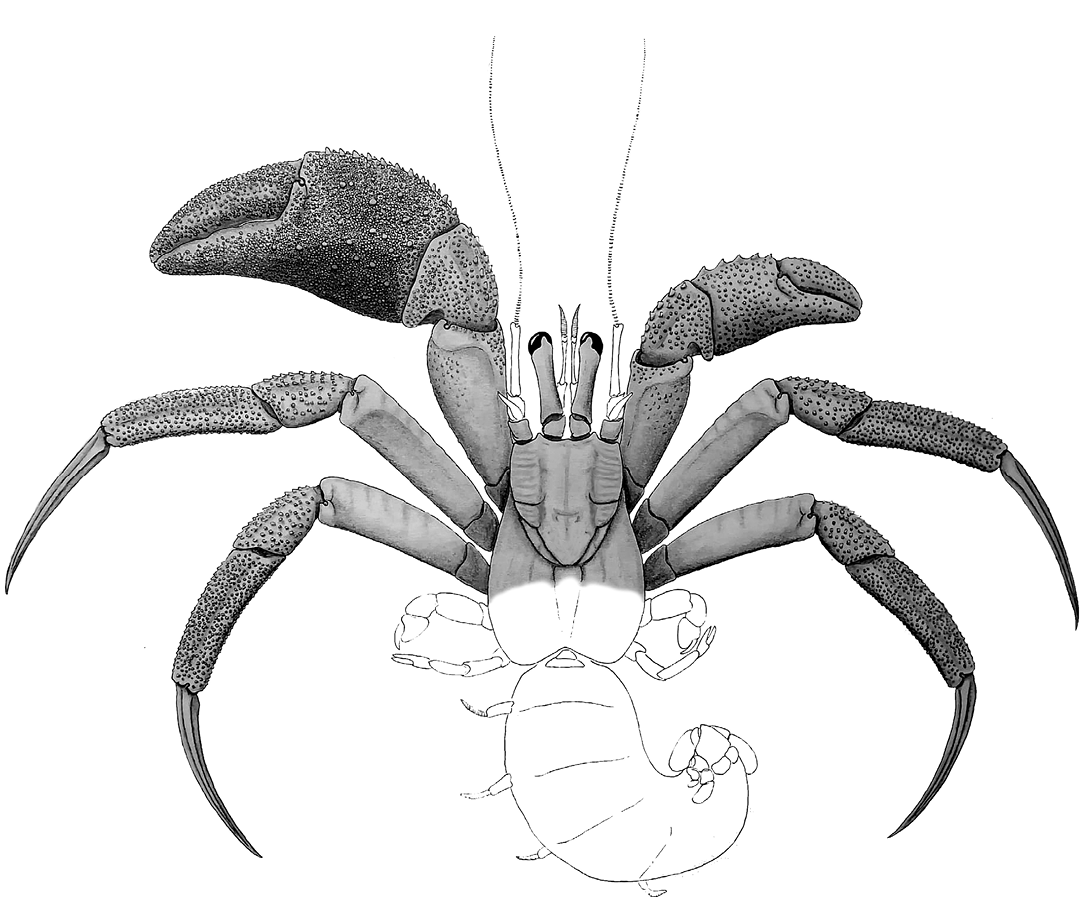
Fig. 2. Reconstruction of Diogenes augustinus sp. nov. Parts shown in grey are those preserved in the fossil material available. Other parts have been reconstructed from the most closely related modern taxa.
The modern genera Dardanus Paul’son, 1875 and Calcinus Dana, 1851, show similar characteristics to those observed in the studied specimen. However, Diogenes augustinus sp. nov. differs from both genera in having a near-straight frontal area, a less well-developed rostrum, a well-defined anterior branchial region (triangular in shape), much larger ambulatory dactyli; in addition, it lacks a Y-line on the posterior part of the shield. The genus Petrochirus Stimpson, 1858 has a characteristic squamose ornamentation; moreover, the anterior branchial region is not clearly defined.
Stratigraphic and geographic range.—Type locality and horizon only.
Concluding remarks
Articulated hermit crabs are rare fossils; mostly only isolated chelipeds are available that offer little information on the overall build of the animals. The depositional setting from which the present articulated specimen was recovered corresponds to a deltaic complex, some few kilometres away of the sediment source area (Millán et al. 1994; Dreyer et al. 1999). The matrix is a fine sandstone, with rare associated molluscs and irregular echinoids. Field observations suggest that bioturbation was rare at these levels except for vertical burrows assigned to Skolithos.
Based on taphonomic experiments, Klompmaker et al. (2017) have recently demonstrated that hermit crabs decay and disarticulate rapidly, only a few days after death. Articulated specimens will be preserved only under certain environmental conditions such as anoxic sea floors (e.g., Garassino et al. 2009) or in areas where sedimentation rates are high (e.g., Shives and Dunbar 2010). With this in mind, we consider the present specimens to have been transported and buried rapidly after the animals abandoned their host shells. High sedimentation rates in the study area were probably responsible for keeping at least one specimen intact by avoiding post-mortem interactions with scavengers. The specimen is preserved out of its host shell; there is no trace of shell material around the crab. This agrees with observations on modern representatives that suggest shell abandonment occurs in 46% of hermit crabs that are smothered after sudden episodic sedimentation (Shives and Dunbar 2010). In fact, episodic high-hydrodynamic pulses associated with a deltaic environment increase sedimentation rate and can bury benthic organisms even alive (e.g., Nichols et al. 1978; Hinchey et al. 2006; Shives and Dunbar 2010). In the study area this is also emphasised by the creation of tectonically induced, large spaces open to sedimentation (Millán et al. 1994).
The data provided by the present material reinforces the ideas previously documented that an increase in sedimentation rate, sudden sedimentary events and absence of scavengers favour the preservation of articulated hermit crabs. The fact that the new material is articulated and occurs outside of the host shells supports the fact that sedimentation rates were very high in this area, probably because it was close to the source area and local tectonics were conducive to such events.
Acknowledgements.—Isabel Pérez (University of Zaragoza, Spain) provided photographic assistance. We thank Agustín Gálvez (Voluntarios del Museo de Ciencias Naturales de la Universidad de Zaragoza) who found the specimen and donated it for study. We are also grateful to the three reviewers Javier Luque (Yale University, New Haven, USA), René Fraaije (Oertijduseum, Boxtel, The Netherlands), and John Jagt (Natuurhistorisch Museum Maastricht, The Netherlands) who greatly improved the resulting manuscript. The present work has been supported by CGL2017-85038-P, subsidised by the Spanish Ministry of Science and Innovation, the European Regional Development Fund and Project E18 “Aragosaurus: Recursos Geológicos y Paleoambientes” of the government of Aragón-FEDER.
References
Dana, J.D. 1851. Conspectus crustaceorum quae in orbis terrarum circumnavigatione, Carolo Wilkes e classe reipublicae foederatae duce, lexit et descripsit. Proceedings of the Academy of Natural Sciences, Philadelphia 5: 267–272.
De Angeli, A. and Caporiondo, F. 2017. I granchi eremiti (Crustacea, Decapoda, Anomura, Paguroidea) dell’Eocene medio di cava “Main” di Arzignano (Vicenza, Italia settentrionale). Studi Trentini di Scienze Naturali 96: 11–32.
De Forges, B.R., Chan, T.Y., Corbari, L., Lemaitre, R., Macpherson, E., Ahyong, S.T., and Ng, P.K. 2001. The MUSORSTOM-TDSB deep-sea benthos exploration programme (1976–2012): an overview of crustacean discoveries and new perspectives on deep-sea zoology and biogeography. Tropical Deep-sea Benthos 185: 1–13.
Dreyer, T., Corregidor, J., Arbues, P., and Puigdefabregas, C. 1999. Architecture of the tectonically influenced Sobrarbe deltaic complex in the Ainsa Basin, northern Spain. Sedimentary Geology 127: 127–169. Crossref
Ferratges, F.A., Zamora, S., and Aurell, M. 2020. Systematics and distribution of decapod crustaceans associated with late Eocene coral buildups from the southern Pyrenees (Spain). In: J.W.M. Jagt, R.H.B. Fraaije, B.W.M. van Bakel, S.K. Donovan, and C. Mellish (eds.), A Lifetime Amidst Fossil Crustaceans: A Tribute to Joseph S.H. Collins (1927–2019). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 296: 79–100. Crossref
Fraaije, R.H.B. 2014. Diverse Late Jurassic anomuran assemblages from the Swabian Alb and evolutionary history of paguroids based on carapace morphology. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 262: 247–255. Crossref
Fraaije, R.H.B. and Polkowsky, S. 2016. Dardanus colosseus, a new paguroid from the Eocene of Austria preserved in its host gastropod shell. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 279: 57–62. Crossref
Fraaije, R.H.B., Beschin, C., Busulini, A., Tessier, G., Jagt, J.W.M., and Van Bakel, B.W.M. 2020. Joecalcinus, a new hermit crab genus from the Eocene of northern Italy: the second Cenozoic representative of the extant family Calcinidae (Decapoda, Anomura). In: J.W.M. Jagt, R.H.B. Fraaije, B.W.M. van Bakel, S.K. Donovan, and C. Mellish (eds.), A Lifetime Amidst Fossil Crustaceans: A Tribute to Joseph S.H. Collins (1927–2019). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 296: 101–105. Crossref
Fraaije, R.H.B., Robins, C., Van Bakel, B.W.M., Jagt, J.W.M., and Bachmayer, F., 2019. Paguroid anomurans from the Tithonian Ernstbrunn Limestone, Austria—the most diverse extinct paguroid assemblage on record. Annalen des Naturhistorischen Museums in Wien 121: 257–289.
Fraaije, R.H.B., Van Bakel, B.W.M., Iserbyt, A., and Jagt, J.W.M. 2011. New extinct Paguroidea (Crustacea, Decapoda, Anomura), with the first example of capsulated setae from the fossil record. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 262: 247–255. Crossref
Fraaije R.H.B., Van Bakel, B.W.M., Jagt, J.W.M., and Artal, P. 2008. New decapod crustaceans (Anomura, Brachyura) from the mid-Cretaceous reefal deposits at Monte Orobe (Navarra, northern Spain), and comments on related type-Maastrichtian material. In: E. Steurbaut, J.W.M. Jagt, and E.A. Jagt-Yazykova (eds.), Annie V. Dhondt Memorial Volume. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre 78: 193–208.
Fraaije, R.H.B., Krzemiński, W., Van Bakel, B.W.M., Krzemińska, E., and Jagt, J.W.M. 2014. New Late Jurassic symmetrical hermit crabs from the southern Polish Uplands and early paguroid diversification. Acta Palaeontologica Polonica 59: 681–688.
Garassino, A. and Schweigert, G. 2006. The Upper Jurassic Solnhofen decapod crustacean fauna: review of the types from old descriptions. Part I. Infraorders Astacidea, Thalassinidea, and Palinura. Memorie della Società Italiana di Scienze Naturali e del Museo Civico di Storia naturale di Milano 34: 1–64.
Garassino, A., De Angeli, A., and Pasini, G. 2009. A new hermit crab (Crustacea, Anomura, Paguroidea) from the Late Cretaceous (Cenomanian) of Lebanon. Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale in Milano 150: 215–228.
Hinchey, E.K., Schaffner, L.C., Hoar, C.C., Vogt, B.W., and Batte, L.P. 2006. Response of estuarine benthic invertebrates to sediment burial: the importance of mobility and adaptation. Hydrobiologia 556: 85–98. Crossref
Huyghe, D., Mouthereau, F., Castelltort, S., Filleaudeau, P.Y., and Emmanuel, L. 2009. Paleogene propagation of the southern Pyrenean thrust wedge revealed by finite strain analysis in frontal thrust sheets: implications for mountain building. Earth and Planetary Science Letters 288: 421–433. Crossref
Hyžný, M., Fraaije, R.H.B., Martin, J.E., Perrier, V., and Sarr, R. 2016. Paracapsulapagurus poponguinensis, a new hermit crab (Decapoda, Anomura, Paguroidea) from the Maastrichtian of Senegal. Journal of Paleontology 90: 1133–1137. Crossref
Jagt, J.W.M., Van Bakel, B.W.M., Fraaije, R.H.B., and Neumann, C. 2006. In situ hermit crabs (Paguroidea) from northwest Europe and Russia. Preliminary data on new records. Revista Mexicana de Ciencias Geológicas 23: 364–369.
Klompmaker, A.A., Portell, R.W., and Frick, M.G. 2017. Comparative experimental taphonomy of eight marine arthropods indicates distinct differences in preservation potential. Palaeontology 60: 773–794. Crossref
Latreille, P.A. 1802. Histoire naturelle, générale at particulière, des Crustacés et des Insects, Familles Naturelles des Genres. F. Tome 3. 467 pp. Dufart, Paris.
MacLeay, W.S. 1838. On the brachyurous decapod Crustacea brought from the Cape by Dr. Smith. In: A. Smith (ed.), Illustrations of the Annulosa of South Africa; Being a Portion of the Objects of Natural History Chiefl y Collected During an Expedition into the Interior of South Africa, Under the Direction of Dr. Andrew Smith, In the years 1834, 1835, and 1836; Fitted out by “The Cape of Good Hope Association for Exploring Central Africa”, 53–71. Smith, Elder, and Co., London.
McLaughlin, P.A. 2003. Illustrated keys to families and genera of the superfamily Paguroidea (Crustacea: Decapoda: Anomura), with diagnoses of genera of Paguridae. Memoirs of Museum Victoria 60: 111–144. Crossref
Millán, H., Aurell, M., and Meléndez, A. 1994. Synchronous detachment folds and coeval sedimentation in the Prepyrenean External Sierras (Spain): a case study for a tectonic origin of sequences and systems tracts. Sedimentology 41: 1001–1024. Crossref
Nichols, J.A., Rowe, G.T., Clifford, C.H., and Young, R.A. 1978. In situ experiments on the burial of marine invertebrates. Journal of Sedimentary Petrology 48: 419–425. Crossref
Pasini, G., Garassino, A., Nyborg, T., Dunbar, S.G., and Fraaije, R.H.B. 2020. In situ hermit crab (Anomura, Paguroidea) from the Oligocene Pysht Formation, Washington, USA. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 295: 17–22. Crossref
Paul’son, O. 1875. Izsledovaniya rakoobraznykh krasnago moryas zametkami otnositel’no rakoobraznykh drugikh morei.Chast’ 1. Podophthalmata i Edriophthalmata (Cumacea). i–xiv + 144 pp. S.V. Kul’zhenko, Kiev. Crossref
Pueyo, E., Millán, H., and Pocoví, A. 2002. Rotation velocity of a thrust: a paleomagnetic study in the External Sierras (southern Pyrenees). Sedimentary Geology 146: 191–208. Crossref
Schweigert, G., Fraaije, R.H.B., Havlik, P., and Nützel, A. 2013. New Early Jurassic hermit crabs from Germany and France. Journal of Crustacean Biology 33: 802–817. Crossref
Schweitzer, C.E. and Feldmann, R.M. 2001. New Cretaceous and Tertiary decapod crustaceans from western North America. Bulletin of the Mizunami Fossil Museum 28: 173–210.
Shives, J.A. and Dunbar, S.G. 2010. Behavioral responses to burial in the hermit crab, Pagurus samuelis: Implications for the fossil record. Journal of Experimental Marine Biology and Ecology 388: 33–38. Crossref
Silva-Casal, R., Aurell, M., Payros, A., Pueyo, E.L., and Serra-Kiel, J. 2019. Carbonate ramp drowning caused by flexural subsidence: the South Pyrenean middle Eocene foreland basin. Sedimentary Geology 393: 1–23. Crossref
Stimpson, W. 1858. Prodromus descriptionis animalium evertebratorum, quae in expeditione ad oceanum Pacifi cum septentrionalem, a Republica Federate missa, Cadwaldaro Ringgold et Johanne Rodgers ducibus, obseravit et descripsit. Proceedings of the Academy of Natural Sciencs of Philadelphia 7: 225–252. Crossref
Walker, S.E. 1992. Criteria for recognizing marine hermit crabs in the fossil record using gastropod shells. Journal of Paleontology 66: 535–558. Crossref
Fernando A. Ferratges [fer.afk87@gmail.com] (corresponding author) and Marcos Aurell [maurell@unizar.es], Departamento de Ciencias de la Tierra-IUCA, Universidad de Zaragoza, Zaragoza 50009, Spain.
Samuel Zamora [s.zamora@igme.es], Instituto Geológico y Minero de España, C/Manuel Lasala, 44/9B, Zaragoza 50006, Spain.
Received 27 may 2020, accepted 20 July 2020, available online 18 September 2020.
Copyright © 2020 F.A. Ferratges et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 65 (4): 787–792,
2020 https://doi.org/10.4202/app.00779.2020