
Old and new specimens of a poorly known glyptodont from the Miocene of Patagonia and their biochronological implications
LAUREANO RAÚL GONZÁLEZ RUIZ, AGUSTINA REATO, MARTÍN CANO, and OSCAR MARTÍNEZ
González Ruiz, L.R., Reato, A., Cano, M., and Martínez, O. 2017. Old and new specimens of a poorly known glyptodont from the Miocene of Patagonia and their biochronological implications. Acta Palaeontologica Polonica 62 (1): 181–194.
The Palaehoplophorini is a tribe of glyptodonts represented by fragmentary specimens for which the oldest records occur in the Pedregoso and Río Mayo formations (middle–late Miocene) in Patagonia (Argentina). The mammal fossils from those formations were the basis for the recognition of the Mayoan fauna, placed between the Friasian sensu stricto/Colloncuran–Laventan and the Chasicoan faunas. The only identified glyptodont from Río Mayo Formation is Palaehoplophorus meridionalis represented only by its type material (i.e., caudal tube) and the precise provenance is unknown. We describe a new specimen of Palaehoplophorus meridionalis collected in the Río Mayo Formation represented by associated osteoderms of the dorsal carapace and postcranial elements, plus part of the type and assigned specimens that remain unpublished from the old collection. This is the first description of osteoderms from the dorsal carapace of this species and postcranial elements (the first for a Palaehoplophorini), and the first glyptodont with precise provenance from Río Mayo Formation. As result of improving the description and emending the diagnosis, we agree with previous authors about the basal position of Palaehoplophorus meridionalis within the tribe. Finally, we review the Glyptodontidae association of the Mayoan fauna, which are exclusively represented by Palaehoplophorini (Palaehoplophorus meridionalis and Palaehoplophoroides rothi) and Glyptodontidae indet. This association supports a greater affinity of this fauna with the younger Chasicoan than with the older Friasian sensu stricto/Colloncuran–Laventan faunas.
Key words: Mammalia, Glyptodontidae, Palaehoplophorus, Miocene, Río Mayo Formation, Patagonia.
Laureano Raúl González Ruiz [gonzalezlaureano@yahoo.com.ar], Agustina Reato [agustinareato@yahoo.com.ar], and Oscar Martínez [oam1958@gmail.com], Centro de Investigación Esquel de Montaña y Estepa Patagónica (CIEMEP) (CONICET-UNPSJB), Roca 780, 9200, Esquel, Chubut, Argentina.
Martín Cano [martinbahia@hotmail.com], Universidad Nacional del Sur, Departamento de Geología (CONICET), San Juan 670, Bahía Blanca, Buenos Aires, Argentina.
Received 21 May 2016, accepted 23 September 2016, available online 8 December 2016.
Copyright © 2017 L.R. González Ruiz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Glyptodontidae (late Eocene–early Holocene) (Simpson 1948; Soibelzon et al. 2012), like other members of the order Cingulata (i.e., Dasypodidae, Peltephilidae, Pampatheriidae, Palaeopeltidae) (McKenna and Bell 1997), are mammals that have a carapace that covers much of the animal’s body, forming in glyptodonts, a cephalic shield, a rigid dorsal carapace without complete transverse movable bands (some basal species have incomplete lateral movable bands) and a caudal carapace (Ameghino 1889; Scott 1903; Hoffstetter 1958). Some specimens could have osteoderms in the limbs, cheeks and in the ventral region as well (Rinderknecht 2000; Tauber and Di Ronco 2000; Soibelzon et al. 2006).
The carapace consists of articulated individual osteoderms having in the dorsal surface a particular pattern due to the overlying epidermal scales (Lydekker 1895; Ameghino 1895, 1920; Holmes and Simpson 1931; Carlini et al. 2009). These osteoderms are usually abundant in the Cenozoic fossil record and their characters from the dorsal surface (e.g., foramina, figures, sulci), particularly those of the dorsal and caudal carapace, are used for the recognition of fossil and extant cingulates, especially when the skull of the animal has not been preserved (González Ruiz et al. 2011b; Gaudin and Croft 2015). While the exclusive use of osteoderms for species diagnosis may lead to artificial groupings (Hoffstetter 1958; Perea 2005) most of the time osteoderms are the only preserved elements. Furthermore, in the case of glyptodonts it has been proven that exoskeletal characters have levels of homoplasy similar to that of endoskeletal characters and there is no reason to exclude them from a phylogenetic analysis or to favor them as the only evidence (Fernicola and Porpino 2012).
In this contribution we describe a new specimen of Palaehoplophorus meridionalis Ameghino, 1904 (Glyptodontidae, Palaehoplophorini) from the Río Mayo Formation (middle–late Miocene) that includes osteoderms of the dorsal carapace and postcranial elements of one individual (there are no duplicate elements). This species of glyptodont was recognized by Florentino Ameghino (1904) based on a caudal tube and osteoderms from the dorsal carapace collected by his brother Carlos Ameghino in Lago Blanco region (formerly Laguna Blanca, southwest Chubut Province), but the osteoderms were never described nor new specimens added (González Ruiz 2010).
The fossil mammals recovered since the early twentieth century in the headwaters of the Fénix and Mayo rivers, and in Lago Blanco region in the southwest of Chubut and northwest of Santa Cruz provinces (Argentina) have been the basis for the recognition of the Mayoan fauna (Ameghino 1903, 1904, 1906; Roth 1908, 1920), characterized by an aggregate of mammals which place it as an intermediate fauna between the Friasian sensu stricto/Colloncuran and the Chasicoan faunas (Ortiz-Jaureguizar et al. 1993). While it is accepted that the sediments bearing this fauna correspond to the Río Mayo Formation (e.g., González 1967) in general there is little data on the precise geographic or stratigraphic provenance of the specimens that were used to identify this fauna, which is a common methodological issue in the paleontological studies during the early twentieth century (Tauber 1997).
According to that, and considering that in both Chilean and Argentine areas, have been recognized several fossiliferous continental formations that could bear this fauna (e.g., Pedregoso Formation sensu Scillato-Yané and Carlini 1998), new collections with stratigraphic provenance became important to corroborate the validity and position of this association of mammals (i.e., Mayoan).
The study of this new specimen of Palaehoplophorus meridionalis and the unpublished specimens from the Ameghino National Collection, led us to improve the knowledge of this species with the description for the first time of osteoderms from the carapace, postcranial elements (the first ones for a Palaehoplophorini), amend the diagnosis and confirm the presence of this species in the Río Mayo Formation (middle–late Miocene). Finally we review the status of all glyptodonts cited for fossil localities assigned to the Mayoan fauna.
Institutional abbreviations.—MACN A, Colección Nacional Ameghino del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina; MLP, Museo de La Plata, La Plata, Buenos Aires Province, Argentina; MPM-PV, Museo Regional “Padre Jesús Molina”, Río Gallegos, Argentina.
Material and methods
The anatomical description follows mostly Gillette and Ray (1981), Duarte (1997), and Porpino et al. (2014).
For the regions of the carapace we modified the terminology used by the mentioned authors because the scarcity of our specimens does not allow us to assign them to a more specific region/section of the carapace, according to that we used more general divisions: cephalic shield, dorsal carapace (anterior, middle and posterior regions) and caudal carapace composed completely of movable rings (each composed of a proximal row of movable osteoderms and a distal row of fixed osteoderms) or by movable rings plus caudal tube.
Osteoderms were grouped according their mobility following Góis et al. (2013) considering that perpendicular (to the dorsal and ventral surface of the osteoderm) articular surface indicates fixed articulations and oblique articular surfaces indicates movable articulations between osteoderms: (i) fixed osteoderms: with all articular surfaces (anterior, laterals, posterior) perpendicular; (ii) semi-movable osteoderms: with the lateral articular surfaces perpendicular and with an antero-dorsal articular surface or a postero-ventral articular surface oblique to the dorsal or ventral surface of the osteoderm respectively; and (iii) movable osteoderms: with the lateral articular surfaces perpendicular and with antero-dorsal and postero-ventral articular surfaces oblique.
The macrotaxonomy follows McKenna and Bell (1997) (but see Fernicola 2008; Porpino et al. 2010; Zamorano and Brandoni 2013; Zurita et al. 2013a).
We analyzed specimens by direct observation of Palaehoplophorus antiquus (Ameghino, 1883a) (MACN A 741, cast; MACN A 589, cast; MACN A 1373, cast), Protoglyptodon primiformis Ameghino, 1885 (MLP M 119, cast-type), Palaehoplophoroides rothi Scillato-Yané and Carlini, 1998 (MLP 92-XI-15-1, type) and Aspidocalyptus castroi Cabrera, 1939 (MLP 37-III-7-6, type) whereas characters of Protoglyptodon sanpedroi Calcaterra, 1978 and Protoglyptodon sagradoi Calcaterra, 1978 are from Calcaterra (1978).
For comparative purposes we included characters from the literature of Glyptatelus Ameghino, 1897 (Glyptatelinae), Propalaehoplophorus Ameghino, 1887a (Propalaehoplophorinae), Neuryurus Ameghino, 1889 (Neuryurini), Panochthus Burmeister, 1866 (Panochthini), Plohophorus Ameghino, 1887b (Plohophorini), Lomaphorus Ameghino, 1889 (Lomaphorini), Neosclerocalyptus Paula Couto, 1957 (Hoplophorini), Doedicurus Burmeister, 1874 (Doedicurinae), and Glyptodon Owen, 1839 (Glyptodontinae) (Ameghino 1889, 1897; Scott 1903; Castellanos 1940, 1941, 1942; Hoffstetter 1958; Zurita et al. 2005, 2009, 2013b, 2016; Fernicola and Porpino 2012; Zamorano et al. 2014).
The geochronologic scheme follows Flynn and Swisher (1995), Madden et al. (1997), and Cohen et al. (2013). We use the faunal concept proposed by Woodburne (2004: 12): “For paleontology, this is an assemblage of vertebrate fossils of similar taxonomic composition obtained from a small number sites considered to have a limited temporal range. A fauna is commonly composed of a number of local faunas. See Tedford (1970). Depending on historical context and author intent, stratigraphic limits of a fauna may be supplied.”
Geological settting
Old specimens.—The precise provenance (especially geographic) of the Mayoan fossils collected by Carlos Ameghino and Santiago Roth were not identified in the original works, however, the available information restricts the provenance to the bluish grey sandstone levels cropping out at Guenguel and Chalía tablelands (Fig. 1).
Meseta de Guenguel and Pampa de Chalía: Ameghino (1904) published fossil mammals collected by Carlos Ameghino during 1901–1902 in the Arroyo Chalía, Río Fénix, and Lago Blanco regions, at different levels of the lower and middle part or the outcrops corresponding to the bluish grey sandstone (Ameghino 1906).
Later, Roth (1908) recognized two sandstone horizons in the Lago Blanco region and the headwaters of the Río Mayo, the lower with abundant mammal fossils and the upper with scarce mammals but abundant gastropods and mollusks, and he later called these levels of bluish grey sandstone “Piso Mayoense” (Roth 1920).
Kraglievich (1930) based on the mammals collected by Santiago Roth included in the Mayoan horizon part of the sediments of Huemúles, Guenguel, Fénix, and Mayo rivers and Lago Blanco. This author also includes the lower levels of Lago Blanco in the Friasian horizon by the sharing record of Megathericulus Ameghino, 1904, however, this record was not confirmed for the Río Frías Formation that bears the Friasian sensu stricto (De Iuliis et al. 2008), plus Ameghino (1906) mentioned that Megathericulus was collected in the bluish grey sandstone levels of Lago Blanco, meaning in the levels that bears the Mayoan fauna.
Feruglio (1949) restricted the region where Carlos Ameghino and Santiago Roth collected the Mayoan fossils to the region between Río Fénix at south and Río Mayo at north. Between both rivers are located the Meseta de Guenguel (crossed by Arroyo Guenguel) and the Pampa de Chalía (crossed by Río Chalía), and between them is located the Lago Blanco.
González (1967, 1978) formally recognized in the area (Meseta de Guenguel and Pampa de Chalía) two consecutive sedimentary formations, Río Mayo and Pedregoso (but see Dal Molín and Franchi 1996), and limited the stratigraphic provenance of the fossils to the upper levels of Río Mayo Formation, and at the same time he mentioned that Carlos Ameghino and Santiago Roth collected the fossils at Lago Blanco (formerly Laguna Blanca).
Finally, new works mentioned and/or described new mammal fossils from Pedregoso Formation (or equivalent) and Río Mayo Formation at Cerro Guenguel and Arroyo Pedregoso, and considered the fossils from both formations as Mayoan (Dal Molín and Franchi 1996; Scillato-Yané and Carlini 1998; Escosteguy et al. 2003; De Iuliis et al. 2008).
Río Huemúles: Roth (1920) collected fossil mammals from a level of sandstone at Río/Arroyo Huemúles/Huemúl near its junction with Río Corcovado, according to Feruglio (1949) near Avilés farm, so they did not mention the Río/Arroyo Huemúles or Río/Arroyo Huemúl located at Lago Blanco nor Lago Fontana located further south. In the area indicated by Roth (1920) outcrops of the Mimosa Formation are composed of sandstone and tuffs, where the fossils could be collected, although the specific locality has not been identified (Turner 1982; Haller et al. 2010).
Those mammals were assigned to the Friasian by Roth (1920) and to the Mayoan by Kraglievich (1930), and according to Haller et al. (2010) the Mimosa Formation—where the fossil were supposed to be collected—to a post-Paleocene and pre-Pleistocene age, although there are no detailed paleontological or geological studies about this formation.
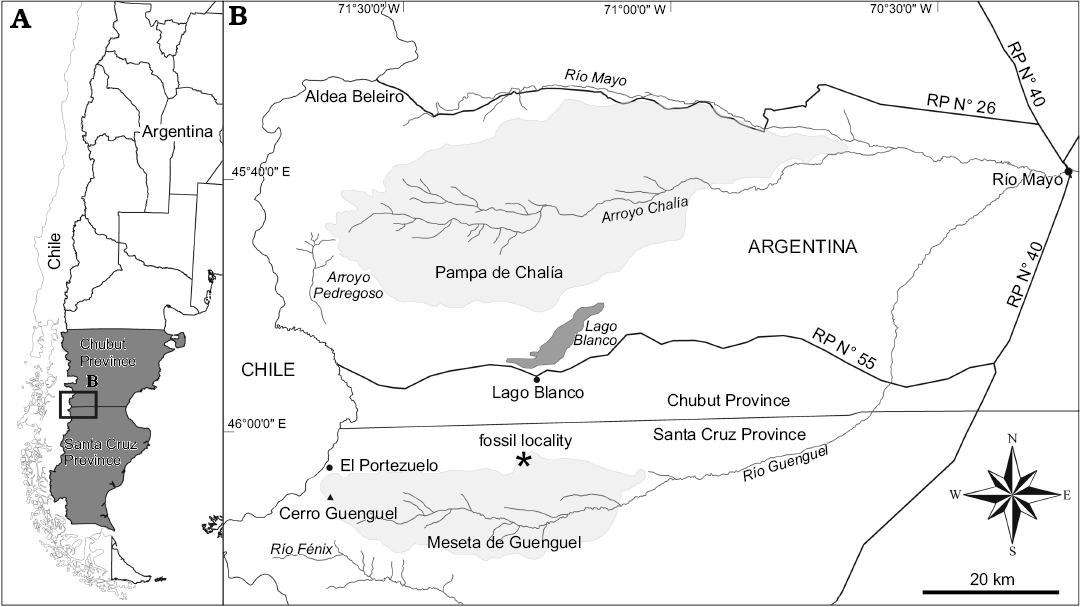
Fig. 1. Map of South Argentina showing location of the studied area (A) and the fossil locality mentioned in the text (B), tablelands marked in light gray.
New specimen.—The new specimen (MPM-PV 17410) was collected from outcrops located on the northern slope of Meseta de Guenguel (Fig. 1), corresponding to the northern boundary of the 4772-II Lago Buenos Aires Geologic Chart (Escosteguy et al. 2003). The specimen was found disarticulated, in a 2 m level of brown grayish sandstone, of coarse grain sized, poorly consolidate, well sorted and with very smooth horizontal bedding (Fig. 2). Above this fossiliferous level, there is a 1.5 m level of whitish gray claystone with bioturbation, then a 2 m level of bluish grey sandstone of medium to coarse grain, with moderately horizontal bedding at the base that changes to cross bedding on the top. Above and below the described section, are outcrop banks of sandstone and siltstone with occasional more clayey levels, some of these corresponding to paleosoils. The section is truncated by an erosional unconformity, and is overlain by stratified gravel covered by till, with a thickness of more than 10 m, preliminary assigned to Pliocene glaciations. We follow Escosteguy et al. (2003) and assign the fossiliferous section to the Río Mayo Formation, based on the presence of cross-bedding bluish grey sandstone plus paleosoils that characterize this formation.

Fig. 2. Stratigraphic profile of the fossil
locality (photograph by LRGR, 2015). Abbreviations: c, coarse; f,
fine; m, medium; vc, very coarse; vf, very fine.
In this region of Argentina, Cenozoic outcrops composed of four formations have been identified, from the base: (i) Río Frías Formation; (ii) El Portezuelo Formation; (iii) Río Mayo Formation; and (iv) Chalía Formation. Further north, on the Pampa de Chalía, on the 4572-IV Alto Río Senguerr Geologic Chart (Dal Molín 1998), the Pedregoso Formation has been proposed instead as equivalent of El Portezuelo Formation (González 1967; Escosteguy et al. 2003). In the equivalent region of Chile, above Río Frías Formation is the Galera Formation in which mammals have not yet been recorded (Skarmeta 1976; Skarmeta and Charrier 1976) and which is conceptually equivalent to Pedregoso, Río Mayo, and Chalía formations. Another remarkable correlation is the one between the upper levels of Oscuro and El Portezuelo formations (Dal Molin and Franchi 1996; de la Cruz et al. 2003; Escosteguy et al. 2003; Quiroz and Bruce 2010; de la Cruz and Cortés 2011; Rivas et al. 2015).
The absolute Ar40/Ar39 dates indicate an age of 12.18± 0.15–12.2 Ma for Pedregoso Formation at the margins of Arroyo Pedregoso, and an age of about 11.8 Ma for a level just below the fossil level of Río Mayo Formation at El Portezuelo on Cerro Guenguel (Dal Molin and Franchi 1996; Madden et al. 1997; De Iuliis et al. 2008). Dunn et al. (2015) mentioned an age of 11.78 for Río Mayo Formation (referring to the age of Madden et al. 1997) and an age of 12.2–13.3 for the underlying formation (El Portezuelo Formation according to Escosteguy et al. 2003), both at Cerro Guenguel. Finally, Rivas et al. (2015) obtained ages of 12.3±0.16 Ma, and 12.18±0.4 Ma for the upper levels of Oscuro Formation.
These ages place the Pedregoso/El Portezuelo, and Río Mayo formations around the Serravallian–Tortonian boundary (middle–late Miocene, 11.63 Ma; Cohen et al. 2013), between the older Collón Curá Formation (14.86±0.13 Ma; Bilmes et al. 2013) and Río Frías Formation (14.83±0.17 Ma; de la Cruz and Cortés 2011), and the younger Arroyo Chasicó Formation (9.23±0.09 Ma; Schultz et al. 2004). These ages partially overlap with La Victoria and Villavieja formations (13.5–11.8 Ma; Madden et al. 1997).
Systematic palaeontology
Xenarthra Cope, 1889
Cingulata Illiger, 1811
Glyptodontidae Gray, 1869
Palaehoplophorini Hoffstetter, 1958
Genus Palaehoplophorus Ameghino, 1883b
Type species: Palaehoplophorus antiquus (Ameghino, 1883a) (= Glyptodon? antiquus Ameghino, 1883a, = Palaehoplophorus scalabrini Ameghino, 1883b); Barrancas del río Paraná (Entre Ríos Province, Argentina), Ituzaingó Formation, Late Miocene (Ameghino 1883a; Scillato-Yané et al. 2013).
Palaehoplophorus meridionalis Ameghino, 1904
Figs. 3–7.
Type material: MACN A 11737, caudal tube (Fig. 4A); MACN A 11137, three fragments of 4, 2, and 2 articulated osteoderms and 12 disarticulated osteoderms of the dorsal carapace, and one osteoderm of the caudal ring (Fig. 3).
Nomenclatorial remarks: Ameghino (1904) in the original description mentioned as type material a caudal tube and osteoderms of the dorsal carapace, but described only the caudal tube (subsequently figured by Rovereto 1914: pl. 2: 3). The only evidence about the association of the type material (caudal tube plus osteoderms) is that they all were mentioned in the original description, and the osteoderms (MACN A 11137) are the only osteoderms assigned to Palaehoplophorus meridionalis in the collection. Accordingly, we suppose that they belong to the same individual or at least they are the osteoderms mentioned by Ameghino (1904) and collectively they should be considered as the holotype.
Type locality: Lower and middle levels of Río Fénix and Lago Blanco (formerly Laguna Blanca) (Ameghino 1906).
Type horizon: Río Mayo Formation, middle–late Miocene.

Fig. 3. Osteoderms of old specimens (MACN A 11137, syntype) of glyptodont Palaehoplophorus meridionalis Ameghino, 1904 from Río Fénix and Lago Blanco, Patagonia, Serravallian–Tortonian, middle–late Miocene. Osteoderms from posterior region (A–N) and caudal ring (O). Abbreviations: apf, accessory periferal figure; cf, central figure; f, foramina; s, sulci; pf, periferal figure.
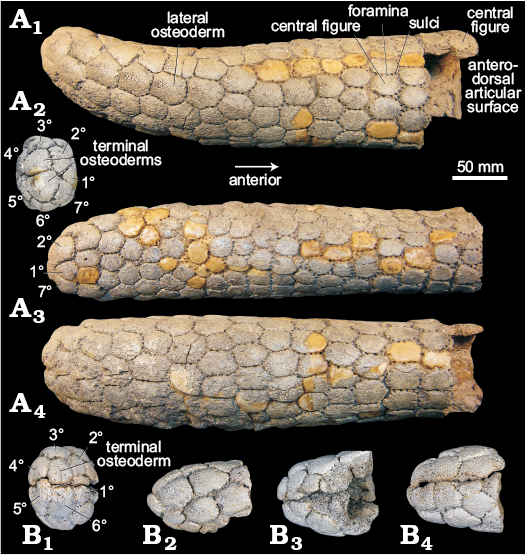
Fig. 4. Caudal tubes of old specimens, syntype and assigned specimens of Palaehoplophorus meridionalis Ameghino, 1904 from Río Fénix, Lago Blanco, and “Rionegrense”, Patagonia, Serravallian–Tortonian, middle–late Miocene. A. MACN A 11737, syntype, in lateral (A1), distal (A2), dorsal (A3), and ventral (A4) views. B. MACN A 11132, in distal (B1), lateral (B2), dorsal (B3), and ventral (B4) views.
Material.—MPM-PV 17410, 250 complete and 29 fragments of osteoderms, left half of the atlas, anterior portion of a caudal vertebra, medial and distal portion of right unla, right cuboid, right ectocuneiform, 1st right ungual phalanx, 12 indeterminate postcranial fragments (Figs. 5–7); from northern slope of Meseta de Guenguel (Santa Cruz Province), Río Mayo Formation. MACN A 11132, distal portion of the caudal tube (Fig. 4B) according to MACN database from “Rionegrense”. Although, the provenance of MACN A 11132 is lower and middle levels of Río Fénix and Lago Blanco (Río Mayo Formation, late–middle Miocene) because some mammals from this area of old collections were labeled “Rionegrense” (see Ameghino 1904, 1906).
Emended diagnosis (of Ameghino 1904: 286–287).—Osteoderms of the dorsal carapace with the rosette pattern (similarly to Propalaehoplophorus, Neosclerocalyptus, and Glyptodon, and absent in Neuryurus, Doedicurus, and Panochthus), with a circular or ellipsoidal flat central figure (concave in Palaehoplophorus antiquus, convex in Protoglyptodon sagradoi), surrounded by one row of 8–12 angular peripheral figures (one of rounded figures in Aspidocalyptus castroi, two plus one incomplete in Palaehoplophorus antiquus, one of small plus one of larger figures in Palaehoplophoroides rothi), defined by deep and narrow sulci (shallow and narrower in Glyptatelus, Propalaehoplophorus, and Neosclerocalyptus, wide and deeper in Glyptodon and Palaehoplophoroides rothi, obscurely defined in Palaehoplophorus antiquus and Protoglyptodon primiformis). Cylindrical caudal tube (absent in Propalaehoplophorus and Glyptodon) distally curved dorsally (straight in Palaehoplophorus antiquus, totally curved Palaehoplophoroides rothi). Composed by flat or slightly convex osteoderms (elevated cone-like in Palaehoplophoroides rothi), without peripheral figures (with peripheral figures in Palaehoplophorus antiquus and Protoglyptodon), separated by narrow sulci (wider in Protoglyptodon and Aspidocalyptus castroi) with conspicuous and numerous foramina (more than 15). The distal portion is formed by 1–2 terminal osteoderms surrounded by a ring of 6–7 small osteoderms (absent in Palaehoplophorus antiquus). The distal lateral osteoderms are the larger of the caudal tube (similar to Palaehoplophorus antiquus) but the big terminal osteoderms (of Plohophorus, Lomaphorus, Neosclerocalyptus) or the large excavated figures of Panochthus and Doedicurus are absent.
Table 1. Average measurements (in mm) of Palaehoplophorus meridionalis osteoderms (MPM-PV 17410, MACN A 11137).
| Diameter | ||||
| antero-posterior | transversal | dorso-ventral | ||
| Dorsal carapace | ||||
| Anterior region | ||||
| Fixed | 25.2 | 20.6 | 13.8 | |
|
Movable |
39.2 | 23.8 | 11.9 | |
| Semi-movable | 34.3 | 21.5 | 13.2 | |
| Middle region | ||||
| Fixed | anterior | 31.5 | 24.3 | 12.8 |
| posterior | 38.9 | 31.2 | 12.2 | |
| Lateral margin | 18.4 | 32.6 | 12.1 | |
| Posterior region | ||||
| Fixed | 42.0 | 34.0 | 14.1 | |
| Posterior margin | 33.9 | 26.1 | 14.1 | |
| Caudal carapace | ||||
| Caudal rings | ||||
| 1º proximal row | 25.1 | 22.1 | 15.1 | |
| Proximal row | 40.8 | 24.6 | 11.5 | |
| Distal row | 25.6 | 25.4 | 10.4 | |
| Intederminate region | ||||
| Semi- movable |
large | 27.8 | 23.5 | 12.7 |
| small | 22.8 | 22.2 | 11.05 | |
Description.—Dorsal carapace (anterior region): Fixed osteoderms (MPM-PV 17410), small sized (Table 1), quadrangular, rectangular, trapezoidal or pentagonal osteoderms (Fig. 5A). They have a large circular, sub-circular or ellipsoidal central figure, which occupies most of the dorsal surface of the osteoderm, and is surrounded by foramina. The peripheral figures are reduced, especially the lateral and posterior figures. Ventrally the osteoderms are flat or slightly convex.
Movable and semi-movable osteoderms (MPM-PV 17410), rectangular, quadrangular or exceptionally trapezoidal osteoderms. The movable osteoderms (Fig. 5C), larger than the semi-movable osteoderms (Fig. 5B; Table 1), have beveled sagittal section due to the opposite antero-dorsal and postero-ventral articular surfaces (Fig. 5C2). In a single row, movable osteoderms are placed laterally close to the lateral margin of the carapace, then internally the semi-movable osteoderms, and finally the fixed osteoderms, going from postero-ventral articular surface oblique (= movable) in the movable osteoderms to posterior perpendicular (= fixed) articulations in the semi-movable and fixed osteoderms, keeping the semi-movable osteoderms a reduced antero-dorsal articular surface oblique. Those are incomplete movable rows of osteoderms that are placed ventrally at the antero-lateral region that become fixed to the dorsal region of the carapace. Both movable as semi-movable osteoderms have a circular central figure surrounded lateral and posteriorly with foramina, while peripheral figures are absent in movable osteoderms and reduced in semi-movable osteoderms. Ventrally are slightly concave, especially in the posterior-most region.
Dorsal carapace (middle region): Fixed osteoderms (MPM-PV 17410), the anteriormost osteoderms are generally rectangular or pentagonal, and the posteriormost ones quadrangular, pentagonal or hexagonal. Osteoderms progressively increase in size from the anterior to posterior (Table 1). The anteriormost osteoderms have circular or elliptical central figure with the major axis coincident with the antero-posterior axis of the carapace (Fig. 5D), and the posteriormost osteoderms have a circular central figure (Fig. 5E). From the anteriormost to the posteriormost and from the lateral to the dorsal region of the carapace, the peripheral figures progressively increase size; in the anteriormost osteoderms the lateral figures are reduced or absent and in posteriormost osteoderms all (anterior, laterals and posterior) have 8–12 peripheral figures, but where there are more than ten, usually two of them are much smaller. In the intersections of the sulci there are well developed foramina. Accessory peripheral figures are in general absent. Ventrally the osteoderms are flat or slightly convex.
Lateral margin osteoderms (MPM-PV 17410), rectangular osteoderms with sharp ends (Fig. 5F) have an ellipsoidal central figure anteriorly that is laterally surrounded by a row of small foramina, and by poorly defined and generally reduced peripheral figures. The anterior margin presents a perpendicular articular surface while the posterior is sharp and does not articulate with any other osteoderm. The ventral surfaces are slightly concave or convex.
Dorsal carapace (posterior region): Fixed osteoderms (MACN A 11137, type; MPM-PV 17410), quadrangular, pentagonal, or hexagonal osteoderms, some could be rectangular with the major axis perpendicular or parallel to the antero-posterior axis of the carapace, generally of large sizes (Figs. 3A–N, 5G, H; Table 1). Fixed osteoderms have a circular central figure that becomes an ellipse posteriorly with the major axis transversal to the antero-posterior axis of the carapace, usually surrounded by 8–11 peripheral figures, and with foramina at the intersections of the sulci that delimit the figures. The presence of more than one or two accessory peripheral figures in the anterior or posterior region (outside of the row surrounding the central figure) is common, in some cases forming a complete anterior and/or posterior row of up to six figures. Towards the margins of the carapace fixed osteoderms become smaller but keep the described pattern. Generally, they are ventrally concave.
Posterior margin osteoderms (MPM-PV 17410), qadrangular osteoderms that become posteriorly thinner (Fig. 5I). They have a circular or sub-circular central figure contacting the posterior margin of the osteoderm and surrounded by foramina. They have an anterior roughened region, followed posteriorly by a region with three peripheral figures, and occasionally an accessory latero-anterior figure. They have no lateral or posterior peripheral figures. Foramina that are located at the intersections of the sulci that delimit the figures are larger. Posterior margin osteoderms are ventrally concave.
Caudal carapace (caudal rings): Osteoderms from the 1st proximal row (MACN A 11137, type), pentagonal osteoderms, with concave anterior margin concave in frontal view), two perpendicular lateral articular surfaces, and two perpendicular posterior articular surfaces (Fig. 3O). The posterior articular surfaces are at an oblique angle between them forming a vertex pointing posteriorly. A sub-circular central figure contacts the posterior margin of the osteoderm, with five very small anterior peripheral figures, and four foramina at the intersections of the sulci that delimits the figures. The ventral surface is concave.
Movable osteoderms from a proximal row (MPM-PV 17410), these pentagonal osteoderms have a large, oblique, antero-dorsal articular surface that covers ca. 40% of the surface and tubercle-like projections in the anterior margin (Fig. 5J). An intermediate region with 3–4 foramina and three peripheral figures is followed by an ellipsoidal central figure with its major axis transverse to the carapace surrounded by a row of foramina. The lateral and posterior articular surfaces are perpendicular. Two obliquely oriented, posterior articular surfaces form a vertex pointing posteriorly. The ventral surface is flat or slightly concave.
Fixed osteoderms from a distal row (MPM-PV 17410), two adjacent osteoderms of the proximal row articulate rigidly with one osteoderm of the distal row (Fig. 5K). These osteoderms are pentagonal with two anterior, obliquely oriented, perpendicular articular surfaces, forming an anteriorly oriented vertex. The lateral and posterior articular surfaces are perpendicular. These osteoderms have a sub-circular central figure reaching the posterior margin, and surrounded posteriorly and laterally by a row of foramina; they typically have no peripheral anterior figures (exceptionally they can have one). The ventral surface is slightly concave.
Caudal carapace (caudal tube): MACN A 11737 (type), the caudal tube is mostly cylindrical but somewhat depressed at its anterior and posterior portion (taphonomic deformation?) it is slightly narrowed and curves dorsally at its distal portion (Fig. 4A). The total length is 38.2 cm, the dorso-ventral diameter decreases from the anterior (10.6 cm) to the posterior region (6.86 cm) and the transverse diameter increases from the anterior (7.8 cm) to the middle region (9.1 cm), and decreases to the posterior region (8.7 cm). The caudal tube is made of osteoderms arranged in rings anteriorly that become disordered distally. The osteoderms from the 1st ring have an antero-dorsal articular surface. The osteoderms of the caudal tube have a rough dorsal surface without surface sculpturing, unlike the osteoderms from the carapace (i.e., figures, sulci), and are separated by sulci in which are located several foramina (ca. 15). Posteriorly, lateral osteoderms are larger than osteoderms in dorsal or ventral positions. Distally, a ring of seven osteoderms plus two terminal osteoderms “close” the caudal tube.
MACN A 11132, represented by a distal portion of caudal tube, has a length of 8.7 cm, and an antero-posterior and dorso-ventral diameters in the preserved region of 6.2 cm and 7.3 cm, respectively (Fig. 4B). The osteoderm pattern is the same as the type (MACN A 11737), although distally a ring of six osteoderms and one terminal osteoderm “close” the caudal tube.
Osteoderms from indeterminate region: Semi-movable osteoderms (MPM-PV 17410), pentagonal osteoderms (Fig. 5L). The larger pentagonal osteoderms have a large circular, sub-circular or elliptical central figure surrounded laterally and posteriorly by foramina, with no peripheral figures. The antero-dorsal articular surface has small tubercle-like projections in the anterior margin. In lateral view the antero-dorsal articular surface is slightly concave, and the transition between the osteoderm surface and the dorsal articulation is abrupt. The smaller pentagonal osteoderms have a reduced circular, sub-circular or ellipsoidal central figure and foramina, with no peripheral figures and the antero-dorsal articular surface is smaller. Osteoderms are ventrally flat or slightly convex. These osteoderms could belong to a nuchal band placed between the dorsal carapace and cephalic shield or to an anterior caudal ring, but they do not belong to the typical semi-movable osteoderms of the antero-lateral region of the dorsal carapace or to the typical caudal ring osteoderms.
Axial skeleton (vertebral column): Atlas (MPM-PV 17410), the dorsal arch (= neural arch) has a greater antero-posterior diameter (23.1 mm) than the ventral arch (= center) (15.2 mm). The vertebral foramen (= neural canal) has a dorso-ventral diameter of 27.4 mm and although only the left half of the atlas is preserved, the outline of the vertebral foramen indicates the ventral half is wider than the dorsal half. In anterior view (Fig. 6A), the major axis of the prezygapophysis is concave and dorso-ventrally oriented (25.1 mm × 18.0 mm), and the anterior intervertebral foramen is situated dorsally and internally. In posterior view (Fig. 6A2), the postzygapophysis with its concave major axis transversally oriented (13.4 mm × 22.2 mm); the vertebroarterial foramen is dorsally located in the midline. The ventral arch has an internal tuberosity (= tubercle) where the odontoid process of the axis articulates. In lateral view and externally (Fig. 6A3), only the base of the transverse process is preserved (23.2 mm × 9.0 mm), dorsally the intervertebral foramen is located within the alar foramen, and ventral to the transverse process another opening of the alar foramen could be observed.
Caudal vertebra (MPM-PV 17410), has preserved the anterior portion of the vertebral body (= centrum) and a portion of the transverse processes (Fig. 6B1). The anterior intervertebral articular surface has a dorso-ventral diameter of 37.1 mm and a transverse diameter of 54.2 mm. The preserved portion of the transverse processes are long (73.0 mm) and wide (22.4 mm), expanding even more distally (25.3 mm), while directed downward and forward slightly, which is the morphology of the first free caudal vertebrae.
Appendicular skeleton (anterior member): Right ulna (MPM-PV 17410), most of the proximal extremity (= olecranon) of the ulna is missing; only the anconeal process was preserved (Fig. 7A). Medially, the ulna has a large and wide trochlear notch (= semilunar notch, sigmoid cavity), a developed coronoid process for the articulation with the humerus, and the smaller radial notch (= fossa lunar) for the proximal articulation with the radius, all of them transverse to the longitudinal axis of the ulna. The diaphysis of the ulna is a straight, compressed triangular prism with a sharp and long interosseous crest. At the distal extremity there is a small styloid process, a large semicircular facet for the articulation with the cuneiform, an anterior facet from the contact with the radius, and a posterior smaller and rounded facet for the articulation with the pisiform. From the coronoid process to the styloid process the unla is 77.3 mm long and has a transverse diameter of 31.1 mm (not completely preserved).
Appendicular skeleton (posterior limb): Right cuboid (MPM-PV 17410), a pyramidal shape bone with a base of 38.5 mm and 21.4 mm height, having four articular facets and a posterior tuberosity. In the internal lateral position the navicular facet is triangular (Fig. 7B1). Dorso-laterally, a semi-circular facet for the articulation with the calcaneum (Fig. 7B3). Facets for articulation with the 4th and 5th metatarsal are situated antero-ventrally (Fig. 7B2); the facet for the 4th metatarsal is larger (22.4 mm × 13.0 mm) than the facet for the 5th metatarsal (15.8 mm × 9.8 mm). Separated from the cuboid’s body by a neck is a triangular plantar tuberosity.
Right ectocuneiform (MPM-PV 17410), the triangular ectocuneiform has a posteriorly oriented, rounded apex, with a base that measures is 28.8 mm, 35.2 mm height and 12.4 mm thickness. The smaller proximal articular facet (26.5 mm × 17.0 mm) is slightly concave, for articulation with the navicular (Fig. 7C1). The larger distal articular facet (33.8 mm × 31.2 mm) occupies the entire surface of the bone and articulates with the 3rd and 4th metatarsal (Fig. 7C3). A small facet (10.9 mm × 8.7 mm) at right angle to the distal facet for the 4th metatarsal contacts internally the cuboid and/or the 4th metatarsal (Fig. 7C4); another small facet (5.9 mm × 7.5 mm) at right angle of the distal facet of the 3rd metatarsal internally contacts the 2nd metatarsal (Fig. 7C2).
Right 1st ungual phalanx (MPM-PV 17410), has a proximal, concave, transversely oriented facet for the 2nd phalanx. Ventrally, the ungual crest (= tubercle) possesses a sub-ungual foramen in the center and a smaller foramen in the external margin (Fig. 7D2). Distally, the ungual process is compressed and internally curved (Fig. 7D1). The antero-posterior length is 29.6 mm and the transverse diameter at the base is 24.1 mm.
Stratigraphic and geographic range.—Middle–late Miocene; Patagonia, Argentina.
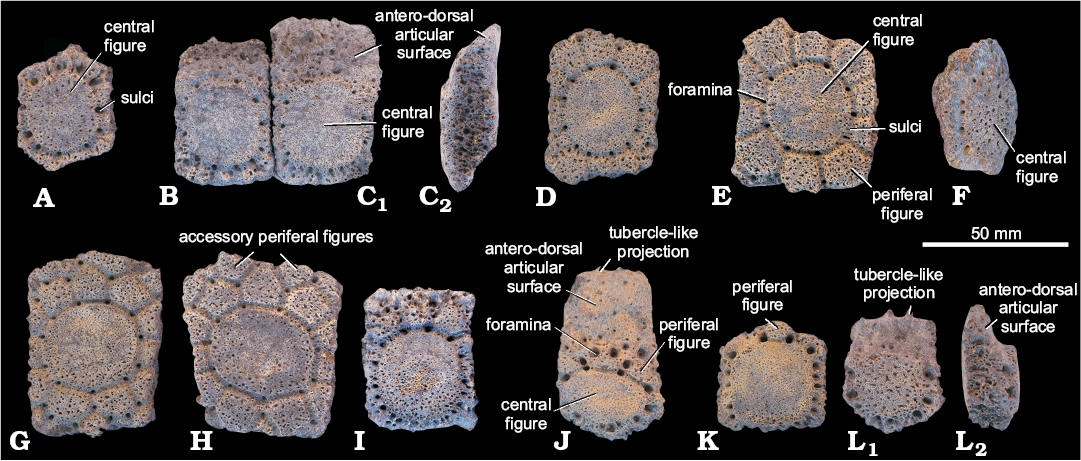
Fig. 5. Osteoderms of new specimen (MPM-PV 17410) of Palaehoplophorus meridionalis Ameghino, 1904 from Meseta de Guenguel, Patagonia, Río Mayo Formation, Serravallian–Tortonian, middle–late Miocene. Anterior region: fixed (A), semi-movable (B), movable (C). Middle region: fixed (D, E), lateral margin (F). Posterior region: fixed (G, H), posterior margin (I). Caudal rings: movable (J), fixed (K). Indeterminated region: semi-movable (L). Osteoderms in lateral view (C2, L2).
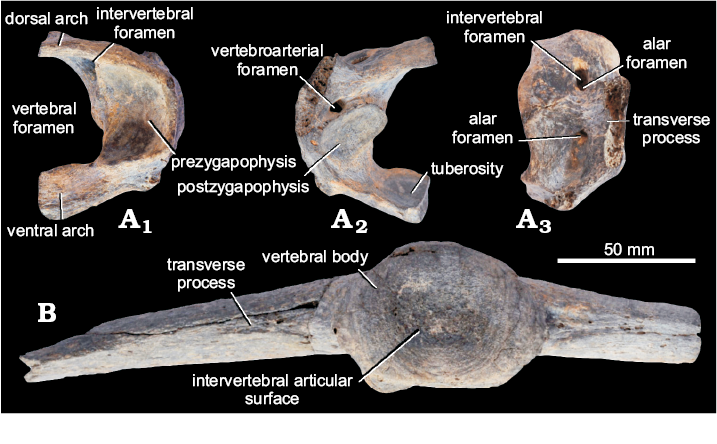
Fig. 6. Vertebrae of new specimen (MPM-PV 17410) of Palaehoplophorus meridionalis Ameghino, 1904 from Meseta de Guenguel, Patagonia, Río Mayo Formation, Serravallian–Tortonian, middle–late Miocene. A. Atlas in anterior (A1), posterior (A2), and latero-external (A3) views. B. Caudal vertebra in anterior view.

Fig. 7. Skeleton of new specimen (MPM-PV 17410) of Palaehoplophorus meridionalis Ameghino, 1904 from Meseta de Guenguel, Patagonia, Río Mayo Formation, Serravallian–Tortonian, middle-late Miocene. A. Ulna in anterior (A1), latero-external (A2), and distal (A3) views; B. Cuboid in latero-internal (B1), antero-ventral (B2), and dorso-lateral (B3) views. C. Ectocuneiform in proximal (C1), distal (C2), latero-internal (C3), and latero-external (C4) views. D. Ungual phalanx in dorsal (D1) and ventral (D2) views.
Discussion
Palaehoplophorus meridionalis in the Palaehoplophorini context.—The tribe Palaehoplophorini was created by Hoffstetter (1958) to include glyptodonts with osteoderms of the dorsal carapace with a pattern represented by a depressed central figure or a depressed cone-like central figure, surrounded by a rough area with large foramina, 1–3 disorderly rows of poorly defined tubercle-like or well defined rounded peripheral figures, and a conical-cylindrical caudal tube formed by osteoderms similar to each other and without modifications especially at its distal region. This author, however, pointed out that these taxa are represented by very fragmentary remains, which could correspond to different regions of the carapace of the same species.
There has not been an updated review of the tribe, and considering the most widespread taxonomic changes proposed would consist of two species of Palaehoplophorus (Palaehoplophorus antiquus, Palaehoplophorus meridionalis), four species of Protoglyptodon (Protoglyptodon primiformis, Protoglyptodon? solidus Ameghino, 1891, Protoglyptodon sagradoi, Protoglyptodon sanpedroi) and three monotypic genera (Aspidocalyptus castroi, Chlamyphractus pressulus [= Palaehoplophorus pressulus Ameghino, 1885] and Palaehoplophoroides rothi) (Hoffstetter 1958; Mones 1986; Scillato-Yané and Carlini 1998; Scillato-Yané et al. 2013). At the same time, Aspidocalyptus castroi has been included in Palaehoplophorus (i.e., Palaehoplophorus castroi) or in Urotherium Castellanos, 1926 (i.e., Urotherium castroi), Palaehoplophorus chapalmalensis Ameghino, 1908 was synonymized with Urotherium simplex Castellanos, 1926 and Palaehoplophorus disjunctus Ameghino, 1889 with Urotherium antiquus (Ameghino, 1888). Chlamyphractus pressulus and Pseudoeuryurus lelongianus Ameghino, 1889 are considered nomen dubium and Pseudoeuryurus lelongianus is sometimes included in the tribe Neuryurini. Finally Pseudohoplophorus Castellanos, 1926 and allied genera have been considered possibly to be Palaehoplophorini (Ameghino 1895, 1920; Castellanos 1926, 1940; Hoffstetter 1958; Paula Couto 1979; Scillato-Yané et al. 2013).
In this context, Palaehoplophorus meridionalis has been considered in pre-cladistic times a basal glyptodont within those having a caudal tube (i.e., Hoplophorinae and Doedicurinae sensu MacKenna and Bell 1997) because of the absence of differentiated osteoderms at its distal portion (e.g., large figures, depressions, projections) and because its carapace osteoderms were supposed to be similar to those of Palaehoplophorus antiquus (= Palaehoplophorus scalabrini) (i.e., without a clearly defined rosette pattern), although authors noted the inconsistency between this basal caudal tube and its derived osteoderms pattern (Castellanos 1925, 1926; Hoffstetter 1958).
Palaehoplophorus meridionalis (or any Palaehoplophorini) has not yet been included in a cladistic phylogenetic analysis, mainly due to the fragmentary specimens, but certain characters used in cladistic phylogenies of glyptodonts can be applied to this species.
Phylogenetic analyses of Porpino et al. (2010) and Fernicola and Porpino (2012) indicate that osteoderms with an approximately central figure (main figure for them) defined by a main sulcus, surrounded by a single row of well defined peripheral figures, is a potential synapomorphy of Glyptodontia (sensu Fernicola 2008), in addition to the fact that the number of rows of peripheral figures increases from the basal rosette pattern independently in Panochthus, Plohophorus, and Stromaphorus Castellanos, 1926. For these authors the presence of a caudal tube is a synapomorphy of Gliptodontoinei (sensu Fernicola 2008). The presence of modifications in osteoderms of the distal region of the caudal tube (e.g., large lateral figures, dorsal and lateral excavations) and morphological modifications (e.g., curved or depressed caudal tubes) are considered as derived characters (Scillato-Yané and Carlini 1998; Fernicola and Porpino 2012; Zamorano and Brandoni 2013; Zurita et al. 2013a).
The osteoderms of Palaehoplophorus meridionalis have the rosette pattern, a central figure surrounded by a row of angular peripheral figures and some accessory figures, all of them well defined by deep sulci, unlike other Palaehoplophorini in which the sulci that define the figures are not well marked and/or are much wider and shallow, which produces poorly defined peripheral figures. In addition, the osteoderms of other Palaehoplophorini are arranged in 1–3 disorderly rows (except Aspidocalyptus castroi which has a row of rounded figures). The caudal tube is formed by osteoderms of approximately the same size, without sculpturing on its exposed surface, unlike other Palaehoplophorini that have peripheral figures. Variation in the number of osteoderms forming the distal portion of the caudal tube, like that of these two specimens of Palaehoplophorus meridionalis, is common in cingulates (Perea 2005 and references cited herein). These characters from the osteoderms of the dorsal carapace and the caudal tube of Palaehoplophorus meridionalis indicate plesiomorphic characters within Palaehoplophorini, although more complete specimens are necessary for the inclusion of this taxon in a phylogenetic analysis in order to check its position within the Gliptodontoinei (sensu Fernicola 2008).
Mayoan fauna and Glyptodontidae record.—Since the first description of Mayoan mammals, this fauna was considered transitional between Santacrucian and “Araucoentrerrian” faunas (Kraglievich 1930). Later, new studies of these faunas demonstrated that the Mayoan fauna is intermediate between the Friasian sensu stricto/Colloncuran and the Chasicoan faunas (supported by geochronological studies, see geographic and geochronologic context), because of the presence of more derived taxa than in the Friasian sensu stricto/Colloncuran and more closely related to Chasicoan mammals (Bond 1993; Ortiz-Jaureguizar et al. 1993; Scillato-Yané et al. 1993; Vucetich et al. 1993; Flynn and Swisher 1995; Scillato-Yané and Carlini 1998). Between the Friasian sensu stricto/Colloncuran, and the Mayoan faunas, there is a faunal gap at high latitudes, maybe partially represented in Patagonia by the post-Collonuran/pre-Laventan fauna of El Petiso (Villafañe et al. 2008; Pérez 2010), represented in low and middle latitudes by the Laventan faunas of Colombia (La Venta), Perú (Fitzcarrald), and Bolivia (Quebrada Honda), although the affinities between them are still not resolved (Madden et al. 1997; Croft 2007; Croft et al. 2013; Tejada-Lara et al. 2015).
The record of glyptodonts for the Mayoan fauna includes Palaehoplophorus meridionalis and Palaehoplophoroides rothi, plus Glyptodontidae indet. (MLP, lost) for Río Guenguel (Kraglievich 1930) and Palaehoplophorus? for Lago Blanco (Bondesio et al. 1980), the last one probably corresponds to four phalanx of Glyptodontidae indet. (MLP, unnumbered). The mention of osteoderms of Propalaehoplophorus sp. by Kraglievich (1930) (Propalaehoplophorus? sensu Bondesio et al. 1980) for the Río Huemúles fauna is not included in the Mayoan record because the specimen was not figured, described or identified at the Museo de La Plata, and there is no certainty about the affinities of this fauna or the geographic provenance of the fossils (Roth 1920; Kraglievich 1930; Haller et al. 2010; Vucetich and Pérez 2011).
In this context, the Glyptodontidae association of the Mayoan fauna (Pedregoso/El Portezuelo, and Río Mayo formations) includes the first confident record of Palaehoplophorini (i.e., Palaehoplophorus meridionalis and Palaehoplophoroides rothi), which are the only recorded group of glyptodonts in this fauna, while they are absent in the previous Miocene faunas of high and middle latitudes (Friasian sensu stricto/Colloncuran) (Scillato-Yané et al. 1993; González Ruiz et al. 2011a; Croft 2016), and middle and low latitudes (Laventan) (Frailey 1988; Carlini et al. 1997, 2008; Croft 2007; Croft et al. 2013; Tejada-Lara et al. 2015). The Palaehoplophorini are abundant and diverse in the posterior Miocene Chasicoan–Huayquerian faunas of middle and high latitudes (Bondesio et al. 1980; Dozo et al. 2010; González Ruiz et al. 2011a; Scillato-Yané et al. 2013), but absent in posterior Miocene–Pliocene low latitudes faunas (Acre, Urumaco, Nabón, Contamana) (Cozzuol 2006; Carlini et al. 2008, 2014; Antoine et al. 2016).
The mayoan glyptodonts supports the greater affinity (at least in middle and high latitudes) between the Mayoan with the subsequent Chasicoan–Huayquerian faunas, as indicated by the rodent and ungulate records as well (Bond 1993; Vucetich et al. 1993), than with the previous Friasian sensu stricto/Colloncuran–Laventan faunas. These affinities should be considered provisional because the glyptodonts from several Miocene–Pliocene faunas from Argentina, Bolivia, Ecuador and Brazil remain undescribed (Bondesio et al. 1980; Cozzuol 2006; Villafañe et al. 2008; Carlini et al. 2014; Croft 2016).
This pattern of taxonomic affinities between middle–late Miocene, and late Miocene–Pliocene faunas from middle and high latitudes shows the effects of the climatic changes produced by the West Antarctic glaciation and the Andean uplift, reflecting the primordial stages of those inhabitants of the drier, colder, and extended plains and pampas that developed throughout the rest of the Miocene, where most of the preceding mammal taxa related to subtropical woodlands became extinct, rare or absent (see Ortiz-Jaureguizar and Cladera 2006).
Conclusions
We have described a new specimen of Palaehoplophorus meridionalis represented by associated osteoderms of the dorsal carapace and postcranial elements, and part of the type (osteoderms from the dorsal carapace) and one assigned specimen (distal caudal tube) that were unpublished from old collections. The new specimens came from the Río Mayo Formation (Serravallian–Tortonian, late–middle Miocene, Mayoan fauna) and the old specimens probably as well.
This is the first description of osteoderms from the dorsal carapace of Palaehoplophorus meridionalis, which has a rosette pattern, and postcranial elements as well (the first for a Palaehoplophorini). Those new specimens allowed us to improve the original description and to amend the diagnosis of this taxon. As a result we agree with previous authors about a basal position of Palaehoplophorus meridionalis within the Palaehoplophorini.
We review the Glyptodontidae association of the Mayoan fauna, which consists exclusively of Palaehoplophorini (Palaehoplophorus meridionalis and Palaehoplophoroides rothi) and Glyptodontidae indet. This association supports a greater affinity of this fauna with the younger Chasicoan than with the older Friasian sensu stricto/Colloncuran–Laventan faunas.
Acknowledgements
We wish to thank the staff of Lago Blanco School Nº 922 for receiving us during the fieldwork, and to our colleagues Enrique Bostelmann (U-Chile, Valdivia, Chile), Diego Brandoni (CONICET—Consejo nacional de Investigaciones Científicas y Técnicas, Diamante, Argentina), Eli Amson (Humboldt-Universität, Berlin, Germany), Juan Fernicola (CONICET, Buenos Aires, Argentina), Fernando Galliari (CONICET, La Plata, Argentina), David Gillette (Museum of Northern Arizona, Flagstaff, USA), Álvaro Mones (Museo Nacional de Historia Natural, Montevideo, Uruguay), Nelson Novo (CONICET, Puerto Madryn, Argentina), Gabriel Martin (CONICET, Esquel, Argentina), and Alfredo Zurita (CONICET, Corrientes, Argentina), who selflessly collaborated with literature, stratigraphic, and anatomical information that improve the content of the manuscript. Finally, we acknowledge the reviewers Darin Croft (Case Western Reserve University, Cleveland, USA) and David Gillette (Museum of Northern Arizona, Flagstaff, USA) for their helpful suggestions that improved the manuscript.
References
Ameghino, F. 1883a. Sobre una colección de mamíferos fósiles del piso mesopotámico de la formación patagónica recogidos en las barrancas del paraná por el profesor Pedro Scalabrini. Boletín de la Academia Nacional de Ciencias en Córdoba 5: 101–116.
Ameghino, F. 1883b. Sobre una nueva colección de mamíferos fósiles recogidos por el Profesor Scalabrini en las barrancas del Paraná. Boletín de la Academia Nacional de Ciencias en Córdoba 5: 257–306.
Ameghino, F. 1885. Nuevos restos de mamíferos fósiles oligocenos recogidos por el Profesor Pedro Scalabrini y pertenecientes al Museo Provincial de la ciudad de Paraná. Boletín de la Academia Nacional de Ciencias en Córdoba 8: 5–207.
Ameghino, F. 1887a. Enumeración sistemática de las especies de mamíferos fósiles coleccionadas por Carlos Ameghino en los terrenos eocenos de Patagonia austral y depositados en el Museo La Plata. Boletín del Museo La Plata 1: 1–26.
Ameghino, F. 1887b. Apuntes preliminares sobre algunos mamíferos estinguidos del yacimiento de “Monte Hermoso” existentes en el “Museo La Plata”. Boletín del Museo La Plata 1: 1–20.
Ameghino, F. 1888. Lista de las especies de mamíferos fósiles del Mioceno superior de Monte-Hermoso, hasta ahora conocidas. 21 pp. P.E. Coni Press, Buenos Aires.
Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de Córdoba 6: 1–1027.
Ameghino, F. 1891. Mamíferos y aves fósiles argentinas. Especies nuevas, adiciones y correcciones. Revista Argentina de Historia Natural 1: 240–259.
Ameghino, F. 1895. Sur les édentés fossiles de l’Argentine: Examen critique, revisión et correction de l’ouvrage de R. Lydekker: The extinct edentates of Argentina. Revista del Jardín Zoológico de Buenos Aires 3: 97–192.
Ameghino, F. 1897. Mammifères crétacés de l’Argentine. Boletín del Instituto Geográfico Argentino 18: 406–521.
Ameghino, F. 1903. Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 56: 193–208.
Ameghino, F. 1904. Nuevas especies de mamíferos cretáceos y terciarios de la República Argentina. Anales de la Sociedad Científica Argentina 58: 241–291.
Ameghino, F. 1906. Les formations sédimentaires du Crétacé superieur et du Tertiaire de Patagonie, avec un parallèle entre leurs faunes mammalogiques et celles de l’ancient continent. Anales del Museo Nacional de Historia Natural de Buenos Aires 8: 1–568.
Ameghino, F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapalmalán. Anales del Museo Nacional de Buenos Aires 10: 343–428.
Ameghino, F. 1920. Sur les édentés fossiles de l’Argentine: Examen critique, revisión et correction de l’ouvrage de R. Lydekker: The extinct edentates of Argentina. (Ouvrage à démi-inédite avec des notes additionelles sur quelques ongulés et carnassiers). Obras Completas y Correspondencia Científica 11: 447–909.
Antoine, P.-O., Abellom M.A., Adnet, S., Altamirano Sierra, A.J., Baby, P., Billet, G., Boivin, M., Calderon, Y., Candela, A., Chabin, J., Corfu, F., Croft, D.A., Ganerød, M., Jaramillo, C., Klaus, S., Marivaux, L., Navarrete, R.E., Orliac, M.J., Parra, F., Pérez, M.E., Pujos, F., Rage, J.-C., Ravel, A., Robinet, C., Roddaz, M., Tejada-Lara, J.V., Vélez-Juarbe, J., Wesselingh, F.P., and Salas-Gismondi, R. 2016. A 60-million-year Cenozoic history of western Amazonian ecosystems in Contamana, eastern Peru. Gondwana Research 31: 30–59. [Crossref]
Bilmes, A., D’Elia, L., Franzese, J.R., Veiga, G.D., and Hernández, M. 2013. Miocene block uplift and basin formation in the Patagonian foreland: The Gastre Basin, Argentina. Tectonophysics 601: 98–111.[Crossref]
Bond, M. 1993. Comentarios sobre los Ungulados de la Edad Friasense de Patagonia, Argentina. Ameghiniana 30: 350R.
Bondesio, P., Laza, J.H., Scillato Yané, G.J., Tonni, E.P., and Vucetich, M.G. 1980. Estado actual del conocimiento de los Vertebrados de La Formación Arroyo Chasicó (Plioceno temprano) de La Provincia de Buenos Aires. II Congreso Argentino de Paleontología y Bioestratigrafía, y I Congreso Latinoamericano de Paleontología, Actas 3: 101–127.
Burmeister, G. 1866. Lista de los mamíferos fósiles del terreno diluviano. Anales del Museo Público de Buenos Aires 1: 121–232.
Burmeister, G. 1874. Monografía de los Glyptodontes en el Museo Público de Buenos Aires. Anales del Museo Público de Buenos Aires 2: 1–412.
Cabrera, A. 1939. Sobre vertebrados fósiles del Plioceno de Adolfo Alsina. Revista del Museo de La Plata nueva serie 2: 3–35.
Calcaterra, A. 1978. Dos nuevas especies de gliptodontes del Plioceno de Uruguay (Mammalia, Edentata). Comunicaciones paleontológicas del Museo Municipal Real de San Carlos 1: 1–7.
Carlini, A.A., Castro, M.C., Madden, R.H., and Scillato-Yané, G.J. 2014. A new species of Dasypodidae (Xenarthra: Cingulata) from the late Miocene of northwestern South America: implications implications in the Dasypodini phylogeny and diversity. Historical Biology 26: 728–736. [Crossref]
Carlini, A.A., Ciancio, M.R., Flynn, J.J., Scillato-Yané, G.J., and Wyss, A.R. 2009. The phylogenetic and biostratigraphic significance of new armadillos (Mammalia, Xenarthra, Dasypodidae, Euphractinae) from the Tinguirirican (Early Oligocene) of Chile. Journal of Systematic Palaeontology 7: 489–503. [Crossref]
Carlini, A.A., Vizcaíno, S.F., and Scillato-Yané, G.J. 1997. Armored Xenarthrans: A Unique Taxonomic and Ecologic Assemblage. In: R.F. Kay, R.H. Madden, R.L. Cifelli, and J.J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, 213–226. Smithsonian Institution Press, Washington.
Carlini, A.A., Zurita, A.E., Scillato-Yané, G.J., Sánchez, R., and Coro, O.A.A. 2008. New glyptodont from the Codore Formation (Pliocene), Falcón State, Venezuela, its relationship with the Asterostemma problem, and the paleobiogeography of the Glyptodontinae. Paläontologische Zeitschrift 82: 139–152. [Crossref]
Castellanos, A. 1925. Breves notas sobre Evolución de la coraza y del extremo caudal en los Loricata (Hicanodonta). Revista El Círculo 93–96.
Castellanos, A. 1926. Sobre un nuevo gliptodóntido chapadmalense, Urotherium simplex n. gen. et n. sp. y las formas afines. Anales del Museo Nacional de Historia Natural Bernardino Rivadavia 34: 263–278.
Castellanos, A. 1940. A propósito de los géneros Plohophorus, Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología 1: 1–279.
Castellanos, A. 1941. A propósito de los géneros Plohophorus, Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología 2: 279–418.
Castellanos, A. 1942. A propósito de los géneros Plohophorus, Nopachthus y Panochthus. Publicaciones del Instituto de Fisiografía y Geología 3: 418–592.
Cohen, K.M., Finney, S.C., Gibbard, P.L., and Fan, J.X. 2013. The ICS International Chronostratigraphic Chart (updated). Episodes 36: 199–204.
Cope, E.D. 1889. The Edentata of North America. American Naturalist 23: 657–664. [Crossref]
Croft, D.A. 2007. The middle Miocene (Laventan) Quebrada Honda Fauna, southern Bolivia, and a description of its notoungulates. Palaeontology 50: 277–303. [Crossref]
Croft, D. A. 2016. Horned Armadillos and Rafting Monkeys. The Fascinating Fossil Mammals of South America. 320 pp. Indiana University Press, Bloomington.
Croft, D.A., Anaya, F., Catena, A., Ciancio, M.R., and Engelman, R. 2013. New species, local faunas, and paleoenvironmental data for the middle Miocene Quebrada Honda Fauna, Bolivia. In: E. Maxwell, J. Miller-Camp, and R. Anemone (eds.), 73rd Annual Meeting SVP, Journal of Vertebrate Paleontology, Program and Abstracts Book, 109. Society of Vertebrate Paleontology, Bethesda.
Cozzuol, M.A. 2006. The Acre vertebrate fauna: age, diversity, and geography. Journal of South American Earth Sciences 21: 185–203. [Crossref]
Dal Molín, C.N. 1998. Hoja Geológica 4572-IV, Alto Río Senguerr, provincia del Chubut. SEGEMAR Boletín 255: 1–34.
Dal Molín, C.N. and Franchi, M. 1996. Reinterpretación estratigráfica de las sedimentitas terciarias del suroeste de Chubut. XIII Congreso Geológico Argentino y III Congreso de Exploración de Hidrocarburos, Actas 1: 473–478.
De Iuliis, G., Brandoni, D., and Scillato-Yané, G.J. 2008. New remains of Megathericulus patagonicus Ameghino, 1904 (Xenarthra, Megatheriidae): Information on primitive features of megatheriines. Journal of Vertebrate Paleontology 28: 181–196. [Crossref]
de la Cruz, R. and Cortés, J. 2011. Geología del área oriental de la Hoja Puerto Cisnes, Región Aysén del Gral. Carlos Ibáñez del Campo. Serie Geología Básica 127: 1–70.
de la Cruz, R., Suárez, M.D., Belmar, M.U., Quiroz, D.P., and Bell, M. 2003. Geología del área Coihaique-Balmaceda, Región Aysén del General Carlos Ibañez del Campo. Serie Geología Básica 80: 1–40.
Dozo, M.T., Bouza, P., Monti, A., Palazzesi, L., Barreda, V., Massaferro, G., Scasso, R.A., and Tambussi, C. 2010. Late Miocene continental biota in Northeastern Patagonia (Península Valdés, Chubut, Argentina). Palaeogeography, Palaeoclimatology, Palaeoecology 297: 100–109. [Crossref]
Duarte, G.R. 1997. Gliptodontes del Pleistoceno tardío de Agua de las Palomas, Campo del Pucará, Catamarca, Argentina. Variaciones morfológicas del caparazón de Glyptodon reticulatus. Ameghiniana 34: 345–355.
Dunn, R.E., Strömberg, C.A.E., Madden, R.H., Kohn, M.J., and Carlini, A.A. 2015. Linked canopy, climate, and faunal change in the Cenozoic of Patagonia. Science 347: 258–261. [Crossref]
Escosteguy, L., Dal Molín, C., Franchi, M., Geuna, S., and Lapido, O. 2003. Hoja Geológica 4772-II, Lago Buenos Aires, Provincia de Santa Cruz. SEGEMAR Boletín 339: 1–80.
Fernicola, J.C. 2008. Nuevos aportes para la sistemática de los Glyptodontia Ameghino 1889 (Mammalia, Xenarthra, Cingulata). Ameghiniana 45: 553–574.
Fernicola, J.C. and Porpino, K.O. 2012. Exoskeleton and systematics: a historical problem in the classification of glyptodonts. Journal of Mammal Evolution 19: 171–183. [Crossref]
Feruglio, E. 1949. Descripción Geológica de La Patagonia. Buenos Aires, Ministerio de Industria y comercio de La Nación. Dirección General de Yacimientos Petrolíferos Fiscales 2: 1–349.
Flynn, J.J. and Swisher III, C.C. 1995. Cenozoic South American land mammal ages: correlation to global geochronologies. In: W.A. Berggren, D.V. Kent, M.P. Aubry, and J. Hardenbol (eds.), Geochronology, Time Scales, and Global Stratigraphic Correlation. Society for Sedimentary Geology Special Publication 54: 317–333.
Frailey, C.D. 1988. The Miocene vertebrates of Quebrada Honda, Bolivia. Part II. Edentata. Occasional Papers of the Museum of Natural History, The University of Kansas 123: 1–13.
Gaudin, T.J. and Croft, D.A. 2015. Paleogene Xenarthra and the evolution of South American mammals. Journal of Mammalogy 96: 622–634. [Crossref]
Gillette, D.D. and Ray, C.E. 1981. Glyptodonts of North America. Smithsonian Contributions to Paleobiology 40: 1–255. [Crossref]
Góis, F., Scillato-Yané, G.J., Carlini, A.A., and Guilherme, E. 2013. A new species of Scirrotherium Edmund & Theodor, 1997 (Xenarthra, Cinculata, Pampatheriidae) from the late Miocene of South America. Alcheringa 37: 177–188. [Crossref]
González, R.R. 1967. Geología de la Región de Lago Blanco y Paso río Mayo (Chubut). Acta Geológica Lilloana 9: 45–65.
González, R.R. 1978. Descripción Geológica de las Hojas 49a, Lago Blanco y 49b, Paso Río Mayo. SEGEMAR Boletín 154–155: 1–45.
González Ruiz, L.R. 2010. Los Cingulata (Mammalia, Xenarthra) del Mioceno temprano y medio de Patagonia (edades Santacrucense y “Friasense”). Revisión sistemática y consideraciones bioestratigráficas. 471 pp. Unpublished Ph.D. dissertation, Universidad Nacional de La Plata, La Plata.
González Ruiz, L.R., Dozo, M.T., Scillato-Yané, G.J., and Zurita, A.A. 2011a. Los Glyptodontidae Palaehoplophorini (Mammalia, Xenarthra, Cingulata) de la Formación Puerto Madryn (Mioceno tardío) en Península Valdés (Chubut, Argentina). Ameghiniana 48 (4): R170.
González Ruiz, L.R., Zurita, A.E, Scillato-Yané, G.J., Zamorano, M., and Tejedor, M.F. 2011b. Un nuevo Glyptodontidae (Mammalia, Xenarthra, Cingulata) del Mioceno de Patagonia (Argentina) y comentarios acerca de la sistemática de los gliptodontes del SALMA “Friasense”. Revista Mexicana de Ciencias Geológicas 28: 566–579.
Gray, J.E. 1869. Catalogue of Carnivorous, Pachydermatous and Edentate Mammalia in the British Museum. 398 pp. British Museum (Natural History), London.
Haller, M.J., Lech, R.R., Martínez, O., Meister, C.M., and Poma, S. 2010. Hoja Geológica 4372-III/IV, Trevelin, Provincia del Chubut. SEGEMAR Boletín 322: 1–86.
Hoffstetter, R. 1958. Xenarthra. In: J. Piveteau (ed.), Traité de Paléontologie, 535–636. Masson et Cie, Paris.
Holmes, W.W. and Simpson, G.G. 1931. Pleistocene exploration and fossil Edentates in Florida. Bulletin of the American Museum of Natural History 59: 383–418.
Illiger, C. 1811. Prodromus systematis mammalium et avium. 301 pp. Sumptibus C. Salfeld, Berlin.
Kraglievich, L. 1930. La Formación Friaseana del río Frias, río Fénix, Laguna Blanca, y su fauna de mamíferos. Physis 10: 127–161.
Lydekker, R. 1895. Contribution to a knowledge of the fossil vertebrates of Argentina. 2. The extinct edentates of Argentina. Anales del Museo de La Plata 3: 1–118.
Madden, R.H., Guerrero, J., Kay, R.F., Flynn, J.J., Swisher, C.C. III, and Walton, A.H. 1997. The Laventan stage and age. In: R.F. Kay, R.H. Madden, R.L. Cifelli, and J.J. Flynn (eds.), Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, 499–519. Smithsonian Institution Press, Washington.
McKenna, M.C. and Bell, S.K. 1997. Classification of Mammals above the Species Level. 631 pp. Columbia University Press, New York.
Mones, A. 1986. Paleovertebrata Sudamericana. Catálogo sistemático de los vertebrados fósiles de América del Sur. Parte I: Lista preliminar y bibliografía. Courier Forschungsinstitut Senckemberg 82: 1–625.
Ortiz-Jaureguizar, E. and Cladera, G.A. 2006. Paleoenvironmental evolution of southern South America during the Cenozoic. Journal of Arid Environments 66: 498–532. [Crossref]
Ortiz-Jaureguizar, E., Madden, R., Vucetich, G., Bond, M., Carlini, A., Goin, F., Scillato-Yané, G., and Vizcaíno, S. 1993. Un análisis de similitud entre las faunas de la “Edad-Mamífero Friasense”. Ameghiniana 30: 351R–352R.
Owen, R. 1839. Description of a tooth and part of the skeleton of the Glyptodon, a large quadruped of the edentate order, to which belongs the tessellated bony armour figured by Mr. Clift in his memoir on the Megatherium, brought to England by Sir Woodbine Parish, F.G.S. Proceedings of the Geological Society of London 3: 108–113.
Paula Couto, C. 1979. Tratado de Paleomastozoología. 590 pp. Academia Brasileira de Ciencias, Río de Janeiro.
Perea, D. 2005. Pseudoplohohorus absolutus n. sp. (Xenarthra, Glyptodontidae), variabilidad en Sclerocalyptinae y redefinición de una biozona del Mioceno Superior de Uruguay. Ameghiniana 42: 175–190.
Pérez, M.E. 2010. A new rodent (Cavioidea, Hystricognathi) from the middle Miocene of Patagonia, mandibular homologies, and the origin of the crown group Cavioidea sensu stricto. Journal of Vertebrate Paleontology 30: 1848–1859. [Crossref]
Porpino, K.O., Fernicola, J.C., and Bergqvist, L.P. 2010. Revisiting the intertropical brazilian species Hoplophorus euphractus (Cingulata, Glyptodontoidea) and the phylogenetic affinities of Hoplophorus. Journal of Vertebrate Paleontology 30: 911–927. [Crossref]
Porpino, K.O., Fernicola, J.C., Cruz, L.E., and Bergqvist, L.P. 2014. The intertropical Brazilian species of Panochthus (Xenarthra, Cingulata, Glyptodonoidea): a reappraisal of their taxonomy and phylogenetic affinities. Journal of Vertebrate Paleontology 34: 1165–1179. [Crossref]
Quiroz, D.P. and Bruce, Z. 2010. Geología del área Puerto Ingeniero Ibáñez-Villa Cerro Castillo. Serie Geología Básica 124: 1–48.
Rinderknecht, A. 2000. La presencia de osteodermos en las extremidades posteriores de Glyptodon clavipes (Mammalia, Cingulata). Ameghiniana 37: 369–373.
Rivas, H., Bostelmann, E., Le Roux, J., and Ugalde, R. 2015. Fluvial facies and architecture of the late middle Miocene, Mayoan, deposits of Chilean Patagonia. In: XIV Congreso Geológico Chileno, Actas 1, 812–815. Colegio de Geólogos de Chile, La Serena.
Roth, S. 1908. Beitrag zur Gliederung der Sedimentablegerungen in Patagonien und der Pampasregion. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 26: 92–150.
Roth, S. 1920. Investigaciones geológicas en la Llanura Pampeana. Revista del Museo de La Plata 25: 135–342.
Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo Nacional de Historia Natural de Buenos Aires 25: 1–247.
Scillato-Yané, G.J. and Carlini, A.A. 1998. Nuevos Xenarthra del Friasense (Mioceno medio) de Argentina. Studia Geologica Salmanticensia 34: 43–67.
Scillato-Yané, G.J., Carlini, A.A., and Vizcaíno, S.F. 1993. Los Xenarthra (Mammalia, Edentata) del Mioceno Medio de Argentina. Ameghiniana 30: 352R.
Scillato-Yané, G.J., Góis, F., Zurita, A.E., Carlini, A.E., González Ruiz, L.R., Krmpotic, C.M., Oliva, C., and Zamorano, M. 2013. Los Cingulata (Mammalia, Xenarthra) del “Conglomerado Osífero” (Mioceno tardío) de la Formación Ituzaingó de Entre Ríos, Argentina. In: D. Brandoni and J.I. Noriega (eds.), El Neógeno de la Mesopotamia Argentina. Asociación Paleontológica Argentina, Publicación Especial 14: 118–134.
Schultz, P.H., Zárate, M., Hames, B., Koeberl, C., Bunch, T., Storzer, D., Renne, P., and Wittke, J. 2004. The Quaternary impact record from the Pampas, Argentina. Earth and Planetary Science Letters 219: 221–238. [Crossref]
Scott, W.B. 1903. Edentata. In: W.B. Scott (ed.), Reports of the Princeton University Expeditions to Patagonia, 1896–1899, 5 (1–2), 1–277, Schweizerbart’sche Verlagshandlung, Stuttgart.
Simpson, G.G. 1948. The beginning of the age of mammals in South America. Part 1. Introduction. Systematics: Marsupialia, Edentata, Condylarthra, Litopterna, and Notioprogonia. Bulletin of the American Museum of Natural History 91: 1–227.
Skarmeta, J.M. 1976. Estratigrafía del terciario sedimentario continental de la región central de la provincia de Aysén, Chile. Revista de la Asociación Geológica Argentina 31: 73–82.
Skarmeta, J.M. and Charrier, R. 1976. Geología del sector fronterizo de Aysén entre los 45° y 46° de latitud sur, Chile. VI Congreso Geológico Argentino, Actas 1: 267–286.
Soibelzon, E., Zurita, A.E., and Carlini, A.A. 2006. Glyptodon munizi Ameghino (Mammalia, Cingulata, Glyptodontidae): redescripción y anatomía. Ameghiniana 43: 377–384.
Soibelzon, L.H., Zamorano, M., Scillato-Yané, G.J., Piazza, D., Rodríguez, S., Soibelzon, E., Toni, E.P., San Cristóbal, J., and Beilinson, E. 2012. Un Glyptodontidae de gran tamaño en el Holoceno temprano de la región Pampeana, Argentina. Revista Brasileira de Paleontologia 15: 105–112. [Crossref]
Tedford, R.H. 1970. Principles and practices of mammalian geochronology in North America. Proceedings of the North American Paleontological Convention 2: 666–703.
Tejada-Lara, J.V., Salas-Gismondi, R., Pujos, F., Baby, P., Benammi, M., Brusset, S., De Franceschi, D., Espurt, N., Urbina, M., and Antoine, P.O. 2015. Life in Proto-Amazonia: Middle Miocene mammals from the Fitzcarrald Arch (Peruvian Amazonia). Palaeontology 58: 341–378. [Crossref]
Tauber, A. 1997. Bioestratigrafía de la Formación Santa Cruz (Mioceno Inferior) en el extreme sudeste de la Patagonia. Ameghiniana 34: 413–426.
Tauber, A.A. and Di Ronco, J. 2000. Nuevo hallazgo de placas ventrales de Glyptodon (Mammalia, Cingulata, Glyptodontidae) en la provincia de Córdoba, República Argentina. Boletín de la Academia Nacional de Ciencias 64: 335–347.
Turner, J.C.M. 1982. Descripción Geológica de la Hoja 44c, Tecka. SEGEMAR Boletín 180: 1–92.
Villafañe, A.L., Pérez, M.E., Abello, M.A., Bedaton, E., and Bond, M. 2008. Nueva localidad fosilífera del Mioceno medio en el noroeste de la provincia del Chubut. In: III Congreso Latinoamericano de Paleonotología de Vertebrados, Actas Resúmenes, 265. Universidad Nacional del Comahue, Neuquén.
Vucetich, M.G. and Pérez, M.E. 2011. The putative cardiomyines (Rodentia, Cavioidea) of the middle Miocene of Patagonia (Argentina) and the differentiation of the family Hydrochoeridae. Journal of Vertebrate Paelontology 31: 1382–1386. [Crossref]
Vucetich, M.G., Massoni, M.M., and Pardiñas, U.F. 1993. Los roedores de la Formación Collón Curá (Mioceno medio) y la Ignimbrita Pilcaniyeu, Cañadón del Tordillo, Neuquén. Ameghiniana 30: 361–381.
Woodburne, M.O. 2004. Definitions. In: M.O. Woodburne (ed.), Late Cretaceous and Cenozoic Mammals of North America, 11–13, Columbia University Press, New York.
Zamorano, M. and Brandoni, D. 2013. Phylogenetic analysis of the Panochthini (Xenarthra, Glyptodontidae), with remarks on their temporal distribution. Alcheringa 37: 1–10.
Zamorano, M., Scillato-Yané, G.J., and Zurita, A.E. 2014. Revisión del género Panochthus (Xenarthra, Glyptodontidae). Revista del Museo de La Plata 14: 1–46.
Zurita, A.E., González Ruiz, L.R., Gómez-Cruz, A.J., and Arenas-Mosquera, J.E. 2013a. The most complete known Neogene Glyptodontidae (Mammalia, Xenarthra, Cingulata) from northern South America: taxonomic, paleobiogeographic and phylogenetic implications. Journal of Vertebrate Paleontology 33: 696–708.
Zurita, A.E., González Ruiz, L.R., Miño-Boilini, A., Herbst, R., Scillato-Yané, G.J., and Cuaranta, P. 2016. Paleogene Glyptodontidae Propalaehoplophorinae (Xenarthra, Cingulata) in extra-Patagonian areas. Andean Geology 43: 127–136.
Zurita, A.E., Scillato-Yané, G.J., and Carlini, A.A. 2005. Paleozoogeographic, bioestratigraphic, and systematic aspects of the Genus Sclerocalyptus Ameghino, 1981 (Xenarthra, Glyptodontidae) of Argentina. Journal of South American Earth Sciences 20: 121–129.
Zurita, A.E., Soibelzon, E., Scillato-Yané, G.J., and Cenizo, M. 2009. The earliest record of Neuryurus Ameghino (Mammalia, Glyptodontidae, Hoplophorinae). Alcheringa 33: 49–57.
Zurita, A.E., Taglioretti, M., Zamorano, M., Scillato-Yané, G.J., Luna, C., Boh, D., and Magnussen Saffer, M. 2013b. A new species of Neosclerocalyptus Paula Couto (Mammalia: Xenarthra: Cingulata): the oldest record of the genus and morphological and phylogenetic aspects. Zootaxa 372: 387–398.
Acta Palaeontol. Pol. 62 (1): 181–194, 2017
http://dx.doi.org/10.4202/app.00280.2016