
A new occurrence of Dakotasuchus kingi from the Late Cretaceous of Utah, USA, and the diagnostic utility of postcranial characters in Crocodyliformes
JOSEPH A. FREDERICKSON, JOSHUA E. COHEN, TYLER C. HUNT, and RICHARD L. CIFELLI
Frederickson, J.A., Cohen, J.E., Hunt, T.C., and Cifelli, R.L. 2017. A new occurrence of Dakotasuchus kingi from the Late Cretaceous of Utah, USA, and the diagnostic utility of postcranial characters in Crocodyliformes. Acta Palaeontologica Polonica 62 (2): 279–286.
Cenomanian mesoeucrocodylians from North America are known primarily from isolated teeth and scutes; any associated remains of this age are noteworthy and represent welcome additions to knowledge. Herein, we describe postcranial elements belonging to a single individual goniopholidid from the Mussentuchit Member of the Cedar Mountain Formation. We argue that this individual represents only the second fossil referable to Dakotasuchus kingi, based on overlapping elements, such as the coracoid, dorsal vertebrae, and scutes, which are strikingly similar to their counterparts in the holotype. The coracoid, in particular, is readily distinguished from those belonging to other closely-related crocodilian taxa; enough so to warrant detailed comparisons in the absence of diagnostic cranial material. The new D. kingi specimen is nearly 20% larger than the holotype, with body length and mass estimates comparable to modern American alligators (Alligator mississippiensis), making it one of the largest aquatic predators in the Mussentuchit ecosystem.
Key words: Crocodylomorpha, Goniopholididae, Dakotasuchus, Cretaceous, Cenomanian, Utah, Mussentuchit.
Joseph Frederickson [joseph.a.frederickson-1@ou.edu], Joshua E. Cohen [jecohen@ou.edu], Tyler C. Hunt [tyler.c.hunt-1@ou.edu], and Richard L. Cifelli [rlc@ou.edu], Sam Noble Museum, 2401 Chautauqua Avenue, Norman, Oklahoma, 73072 USA; Department of Biology, University of Oklahoma, Norman, Oklahoma, 73019 USA.
Received 29 December 2016, accepted 11 February 2017, available online 11 April 2017.
Copyright © 2017 J. Frederickson et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Crocodyliform teeth and scutes are some of the most commonly encountered fossils in medial Cretaceous deposits of North America (e.g., Garrison et al. 2007); paradoxically, however, knowledge of these animals is severely limited and is based on a few associated specimens comprising a handful of species. Of named species, Dakotasuchus kingi Mehl, 1941 ranks among the most enigmatic and poorly known. Mehl (1941) named Dakotasuchus kingi based on a single relatively complete specimen (KWU uncatalogued), found encased in a large iron-cemented sandstone concretion. Though not collected by Maurice G. Mehl, this specimen was believed to have been discovered just outside of Salina, Kansas at the beginning of the 20th century, originating from the Upper Cretaceous Dakota Formation (Fig. 1A; see Scott 1970 for an alternative interpretation as originating from the Lower Cretaceous Kiowa Shale). The specimen itself is extraordinarily preserved, containing much of the axial column and both limb girdles, as well as impressions for most of the dorsal and ventral shields in full articulation, but as the specimen is encased within a concretion, the anatomy for the majority of these elements is obscured. The greater part of the descriptions of the KWU specimen is heavily based upon natural molds, wherein the concretion was broken and the original bone had weathered away. At the time, Mehl (1941) interpreted the fossil as a mesosuchian based on the shape of the dermal armor and the presence of amphicoelous vertebrae. Further, he estimated that the holotype specimen was from an individual between ten and twelve feet (3.0–3.7 m) in length and, owing to its short and robust scapula, he suggested that Dakotasuchus was a predominately terrestrial crocodilian. Since this description, only meager isolated material loosely identified to D. kingi has been reported from Kansas (Vaughn 1956; Everhart 2005). Outside of the state, varied specimens have been attributed to D. kingi, mainly on the basis of size and similar geologic age, ranging from isolated scutes and teeth from the Comanchean of Texas (Langston 1974) to large crocodilian tracks from the Dakota Group of Colorado (Lockley 2010; Kukihara and Lockley 2012). In most cases, however, the material is either limited in diagnostic utility (e.g., isolated scutes) or unknown in the holotype (e.g., teeth and the morphology of the manus and pes). Here we describe the first specimen, found outside the state of Kansas, which can be confidently attributed to D. kingi. This new specimen allows for the verification of anatomical descriptions based on the natural molds of the KWU specimen and provides a basis for comparisons with other mesoeucrocodylians from the medial Cretaceous of North America. Additional 3D files of coracoid, radius and dorsal vertebra are available as Supplementary Online Material at http://app.pan.pl/SOM/app62-Frederickson_etal_SOM.pdf.
Institutional abbreviations.—IRSNB, Institut Royal des Sciences Naturelles de Bruxelles, Brussels, Belgium; KWU, Kansas Wesleyan University, Salina, Kansas; OMNH, Sam Noble Oklahoma Museum of Natural History, Norman, Oklahoma; SMNH, Royal Saskatchewan Museum, Regina; SMU, Southern Methodist University Shuler Museum of Paleontology, Dallas, Texas.
Geological setting
The Mussentuchit Member is the uppermost unit of the Barremian–Cenomanian Cedar Mountain Formation, which underlies the Naturita Formation (formerly known as and contemporaneous to the Dakota Formation; Carpenter 2014). Lithologically, the Mussentuchit Member is composed primarily of terrestrial mudstone, sandstone, and altered volcanic ashes (Kirkland et al. 1997), which were fluvially deposited in the foreland basin approximately 98 Ma during the Pavast thrust event (Cifelli et al. 1997). Unlike the other members of the Cedar Mountain Formation, localities within the Mussentuchit Member yield a highly diverse fauna composed of freshwater elasmobranchs, actinopterygians, lungfishes, lissamphibians, squamates, crocodilians, dinosaurs, and mammals (Cifelli et al. 1999; Fiorillo 1999; Garrison et al. 2007; Frederickson and Cifelli 2017). The specimen described here comes from OMNH locality V828 (Fig. 1B), discovered and excavated during the summer of 1994. The exposed fossiliferous layer was in a highly weathered sandy mudstone containing abundant rip-up clasts and small pebbles. Sediments up to ~250 mm stratigraphically above V828 were carbonaceous with abundant bituminous coals and unidentifiable carbonized plants. The main fossil layer was found as a single concentration of associated bones, with a few minor elements randomly strewn on the periphery, approximately a meter from the largest block. The specimen was largely disarticulated, but the preserved elements show a strong bias toward the right side of the body. The association, large size, and similar preservation leaves no doubt that this specimen comes from a single individual.
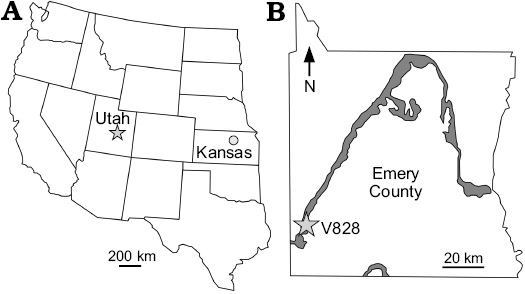 |
Fig. 1. Map of the western United States (A) with the approximate locations of the holotype in Salina, Kansas (KWU uncatalogued; circle) and referred specimen in Emery County, Utah (OMNH 34500; star) and map of Emery County (B) with the approximate location of V868 (star) and the distribution of the Mussentuchit Member (grey area) (modified from Cifelli et al. 1999). |
Systematic palaeontology
Crocodylomorpha Walker, 1970
Crocodyliformes Hay, 1930
Mesoeucrocodylia Whetstone and Whybrow, 1983
Coelognathosuchia Martin, Lauprasert, Buffetaut, Liard, and Suteethorn, 2014
Goniopholididae Cope, 1875
Genus Dakotasuchus Mehl, 1941
Type species: Dakotasuchus kingi Mehl, 1941; monotypic, see below.
Diagnosis.—As for the type and only species.
Dakotasuchus kingi Mehl, 1941
Figs. 2, 3A, E; SOM.
Holotype: KWU uncatalogued, an articulated partial skeleton in a concretion missing the skull and limb elements
Type locality: West of Salina, Kansas, USA.
Type horizon: Dakota Formation, Cenomanian (Late Cretaceous).
Material.—OMNH 34500, a disarticulated partial skeleton, consisting of a right coracoid, right radius, four partial to complete dorsal vertebrae, six caudal vertebrae, three cervical ribs, four nearly complete and many partial dorsal ribs, and multiple ventral and dorsal scutes. From OMNH locality V828, ca. 26 km S of Emery, Emery County, Utah; Cenomanian (Late Cretaceous), Mussentuchit Member of the Cedar Mountain Formation. OMNH V828 is on Federal land administered by the US Bureau of Land Management, which maintains and restricts access to specific locality coordinates.
Diagnosis.—Mehl (1941) never provided a diagnosis for Dakotasuchus kingi. This taxon resembles other North American medial Cretaceous coelognathosuchians such as Woodbinesuchus byersmauricei and Terminonaris robusta, but can be differentiated based on a unique combination of postcranial characters not uniformly present in either taxon: (i) the presence of triangular processes on the caudodorsal surface of the cervical ribs; (ii) dorsal vertebrae with a supporting buttress which forms at the caudal margin of the neural arch and continues through the ventral side of the transverse processes; (iii) the presence of vertebrae with a deep groove on the dorsal surface of the centra, giving the neural canal a heart-shaped appearance; (iv) a scapula with a deeply concave cranial margin with a thickened and expanded distal end that terminates in a ventrally-directed hook; (v) coracoid with dorsal and ventral heads that are rounded and relatively expanded caudally; (vi) a relatively robust radius that possesses a large hook-shaped tubercle on the medial side of the proximal condyle; (vii) ischium possessing a broad pubic process that is oriented more perpendicular to its shaft; and (viii) dorsal scutes with a rounded and concave caudolateral edge. Characters (i), (ii), (iii), (iv), (v), (vi), and (vii) are not shared with Woodbinesuchus byersmauricei, whereas characters (i), (ii), (iv), (v), and (vii) are not found in Terminonaris robusta.
Description.—OMNH 34500 preserves multiple elements on the right side of the body, including a coracoid and radius, as well as cervical ribs, together with vertebrae and osteoderms. In addition, at least 12 teeth were found in association with the specimen, with the largest possessing a maximum height of 18.3 mm; though many of these appear too small (< 10 mm in height) to belong to this individual. The bones are relatively well-preserved; however some elements, such as the radius and coracoid, possess fragmented bone surfaces. Broken edges of the bones are relatively sharp, with little smoothing evident on any of the larger elements, implying that taphonomic transport was minimal. A single theropod tooth and a possible mammal canine were also found in close proximity to OMNH 34500.
The most cranially-located elements preserved in OMNH 34500 are three cervical ribs. Overall, the general morphology agrees with descriptions given by Mehl (1941); however, the cervical ribs of the KWU specimen are rather incomplete. Supplemental descriptions provided here are based on a complete, right cervical rib of unknown position (Fig. 2A). The ovate capitular articular surface is nearly three times larger than that of the nearly circular tubercular articular surface. In dorsal view, the capitulum is much thicker than the tuberculum, whereas the tuberculum is longer. A thin posteriorly-directed ridge descends off of the capitulum, ending adjacent to the ventral margin of a triangular process. This conspicuous triangular process (Fig. 2A2), on the caudodorsal surface, is oriented caudomedially and is either absent or incipient on other North American Cretaceous crocodilians, as well as modern crocodilians. The cranial process is thick, anterodorsally compressed with a rugose craniodorsal surface, and has a large groove on the ventral side. The posterior process is thin and is curled medially to accommodate the corresponding cranial process of the following cervical rib.
The dorsal vertebrae (Fig. 2C) are similar in size and shape to those described from the holotype specimen of Dakotasuchus kingi. The centra are amphicoelous, with a shallow central depression and flange-like processes on the margin of the articular surfaces, giving the centra a constricted appearance in lateral view (Fig. 2C3).r The cranial articular surface is circular (Fig. 2C1), while the caudal articular surface is dorsoventrally taller and subcircular in outline (Fig. 2C2). Mehl (1941) describes a conspicuous buttress that arises from the caudodorsal margin of the centrum and continues through the transverse process on the holotype specimen; this is also seen in the newly-referred specimen (Fig. 2C3). In OMNH 34500 this feature is present on all dorsal vertebrae, varying in thickness along the vertebral column from notably thickened in cranially-placed vertebrae to nearly flush with the rest of the ventral face of the transverse processes in more caudal vertebrae. The neural canal is heart-shaped in both Dakotasuchus specimens, with a deep groove incised into the dorsal face of the centrum (Fig. 2G). As described by Mehl (1941), the zygapophyses in OMNH 34500 are nearly continuous with the dorsal surface of the transverse processes. Both pre- and postzygapophyses are generally wide, elongate, and finger-like (Fig. 2C4). The neural spines are short and robust, with a mediolateral and craniocaudally expanded apex. Multiple nondescript fragmentary and incomplete dorsal ribs were recovered; these are uninformative and are ignored herein. The caudal vertebrae are largely incomplete, missing the majority of the neural arches; in known respects they are morphologically similar to caudals of other mesoeucrocodylians.
For the appendicular skeleton, only the right coracoid and radius are known in OMNH 34500. Mehl (1941) was able to make a cast of the right coracoid from a natural mold in the holotype specimen. Unfortunately, sometime in the following seven decades this section of the concretion was damaged; however, much of the left coracoid is still visible and descriptions (and figures) from Mehl (1941) appear to accurately reflect the morphology of the now broken right element. In OMNH 34500, the midline of the coracoid (taken in the coronal plane) measures 235 mm (greatest length along midline = 279 mm). The coracoid is long, with expanded dorsal (152 mm wide) and ventral (114 mm wide) ends (Fig. 2B). In profile, the coracoid is curved ventromedially and, like the holotype, is at its greatest curvature at about one third of the way from the dorsal head; the distal two-thirds of the shaft is straight (Fig. 2B2). The diameter of the shaft at the apex of the curve (34 mm) is the most constricted portion of the shaft, which despite its larger size, is the same as the holotype described by Mehl (1941). The outline of the dorsal head is mushroom-shaped in lateral view. The scapulocoracoid junction is straight, and gently curves cranially at the terminus of the junction. Cranial to the scapulocoracoid junction, the dorsalmost edge is gently rounded. The coracoid foramen is positioned cranial and dorsal to the glenoid (Fig. 2B3), similar to the position seen in other Cretaceous mesoeucrocodylians (Mehl 1941; Lee 1997; Wu et al. 2001). The articular surface of the glenoid on the coracoid is highly rounded (Fig. 2B2). The glenoid curves ventrally, forming a discrete ridge with a lip along the lateral perimeter of the glenoid. The ventral end of the coracoid flattens into a sharp ulu-like blade that approaches the ventral midline of the body. The caudal edge of the shaft is much straighter than the rostral edge, but both end in a noticeable pointed flare.
The right radius is nearly complete but bears multiple large, radial and longitudinal fractures. In profile, it is a robust, elongate element (248 mm long) with a flattened proximal condyle and a rounded distal condyle (Fig. 2D1). In lateral view (Fig. 2D2), the shaft has a sigmoidal shape and the lateral corner of the distal end is craniomedially rotated. As in modern crocodilians, the humeroarticular surface is kidney shaped with an enlarged proximal condyle, whereas the distal condyle is rounded with a central depression on the cranial face for the articulation with the ulna. There is a conspicuous hook-shaped tubercle for the insertion of the humeroantebrachialis inferior muscle on the medial side of the proximal condyle (Fig. 2D1). The insertion tubercle for the brachialis muscle, also present but expressed as a low mound rather than a tubercle, is located a quarter of the way down from the proximal end of the shaft.
Three dorsal osteoderms (two complete and one partial) are preserved in OMNH 34500. These are of typical coelognathosuchian (comprising Pholidosauridae and Goniopholididae; Martin et al. 2014) morphology, roughly rectangular in outline and paired along the midline of the body. The dorsal surface of each osteoderm is irregularly ornamented with deep oval pits varying in size from 1.0 to 13.0 mm in diameter (Fig. 2E). The cranial edge of each dorsal scute has a smooth margin that articulates with the caudal end of the preceding scute. In addition, a large, broadly triangular cranial dorsal process (Fig. 2E) is present on the craniolateral side of the scute. The lateral edge is ventrally deflected, creating a concavity to interlock with the aforementioned cranial dorsal process of the following dorsal scute. The caudolateral corner is also rounded, with a discrete notch for receiving this process (Fig. 2E2). The ventral surface is relatively smooth with a shallow groove running on the caudomedial and extending a third of the way along the length of the scute. Like the dorsal scutes, the ventral scutes compare favorably to the description provided by Mehl (1941) for Dakotasuchus kingi. The scutes range from pentagonal near the midline to hexagonal on the flanks. The ventral surface of each scute is only weakly pitted and is generally more flattened near the center of the body. On the dorsal surface, the bone fibers are arranged in a random orientation. The outer margins are covered in radial crenulations that are morphologically reminiscent of sutures on cranial bones (Fig. 2F).
The largest teeth associated with this specimen have an oval base, with two distinct carinae and multiple fine striae on the crown (Fig. 2H). Other large teeth from the Mussentuchit Member (31 mm, OMNH 32090; 32.9 mm, OMNH 34601) likely also come from this taxon as well, and are presumably synonymous with the “pholidosaurid” teeth described by Garrison et al. (2007: fig. 23E, F). Even so, these teeth are shorter than a sizeable crocodilian tooth (41.9 mm, OMNH 27825) that was discovered in the underlying, Lower Cretaceous Ruby Ranch Member of the Cedar Mountain Formation in Grand County (V857), suggesting that even larger crocodilians were present in the medial Cretaceous of Utah.
Stratigraphic and geographic range.—Cenomanian (Cretaceous), Dakota Formation of Kansas, USA and Mussentuchit Member of the Cedar Mountain Formation of Utah, USA.
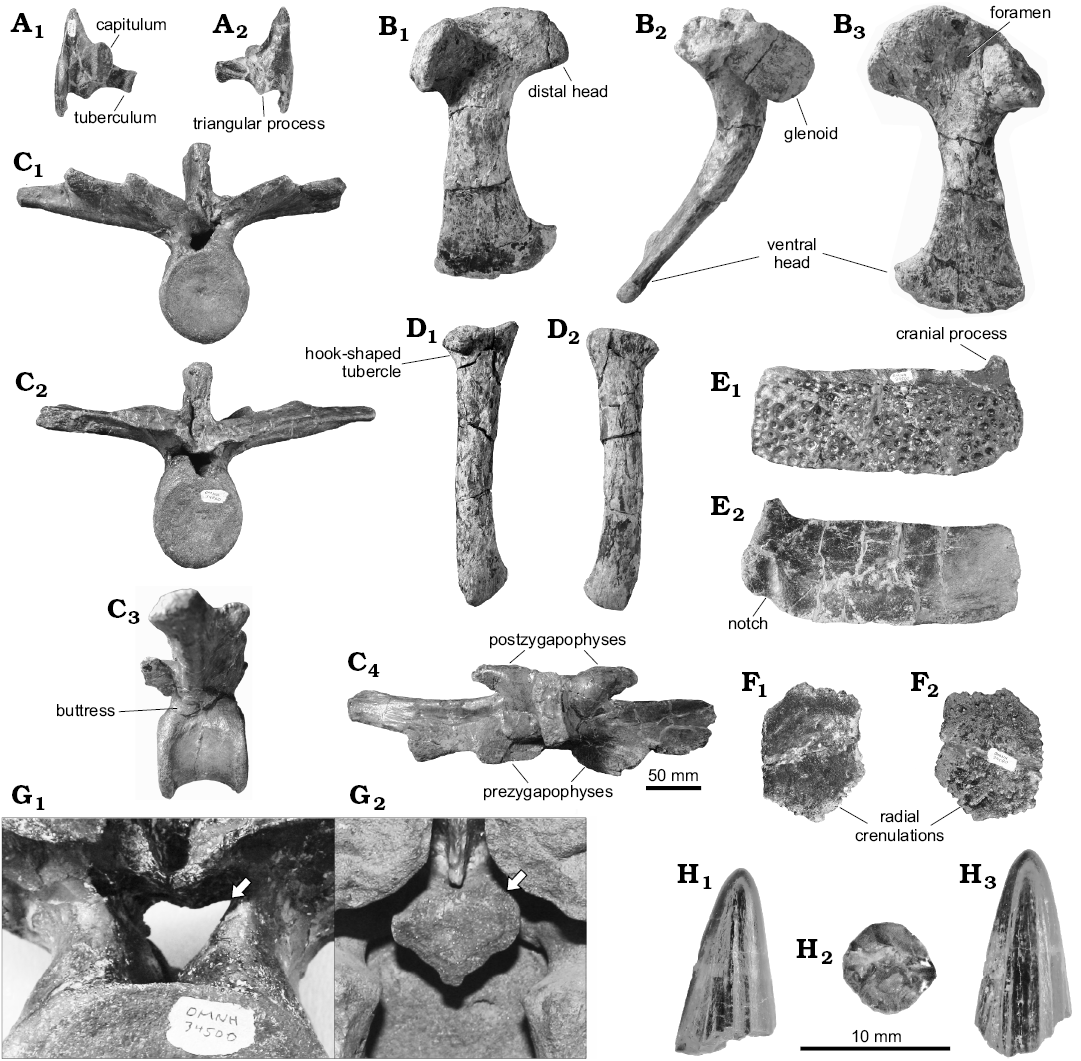
Fig. 2. Representative elements of goniopholidid crocodyliform Dakotasuchus kingi Mehl, 1941, OMNH 34500 from OMNH locality V828, Mussentuchit Member, Cedar Mountain Formation, Cenomanian. A. Right cervical rib in ventral (A1) and dorsal (A2) views. B. Right coracoid in lateral (B1), caudal (B2), and medial (B3) views. C. Dorsal vertebra in cranial (C1), caudal (C2), lateral (C3), and dorsal (C4) views. D. Right radius in medial (D1) and lateral (D2) views. E. Dorsal scute in dorsal (E1) and ventral (E2) views. F. Ventral scute in dorsal (F1) and ventral (F2) views. G. Close-up views of neural canal in dorsal vertebrae, illustrating distinctive heart shape (white arrows); G1, OMNH 34500 vertebra in caudal view; G2, D. kingi holotype vertebra mold in cranial view. H. Tooth in labiolingual (H1), basal (H2), and mesiodistal (H3) views.
Discussion
Morphological comparison with other coelognathosuchians.—Diagnostic cranial material is lacking from both OMNH 34500 and the holotype specimen of Dakotasuchus kingi, so that referral of the Mussentuchit specimen must rely entirely on postcranial elements. This lack of cranial material known for Dakotasuchus precludes phylogenetic treatment, as the majority of fossil crocodyliforms are known primarily by isolated skulls. Rectifying the imbalance of postcranial crocodyliform anatomy in phylogenentic analyses is beyond the scope of this paper. However, recent work by Martin et al. (2016a) has called for better descriptions of isolated postcrania for crocodyliforms, noting that without sufficient knowledge of the postcranial variability it is often difficult to associate material lacking a skull. Thus, even if a skull belonging to D. kingi were to be found, it would be difficult to make a clear association without better documentation of its postcranial variability. Here we argue that both specimens of D. kingi can be differentiated from other closely related North American species based on overlapping elements, specifically the coracoid and dorsal scutes.
In general, coelognathosuchians from the Cenomanian of North America are poorly represented, with only three named taxa: Woodbinesuchus byersmauricei Lee, 1997, Terminonaris cf. T. robusta (see Adams et al. 2011), and Dakotasuchus kingi (Mehl 1941; see also Vaughn 1956; Everhart 2005). Woodbinesuchus and Terminonaris both derive from the Woodbine Formation of north Texas, with Woodbinesuchus represented by at least one mostly-complete individual. Terminonaris is known from multiple fragmentary specimens, but is also well-documented elsewhere from the Turonian of North America (Osborn 1904; Mook 1934; Wu et al. 2001). Since the postcrania of the Terminonaris specimens from the Woodbine Formation are largely unknown (Adams et al. 2011), we base our comparisons on the better-known Turonian fossils (Wu et al. 2001). An additional unnamed taxon is known from the Woodbine Formation and awaits formal description (Main 2013). We believe this material is sufficiently different from Dakotasuchus (now including OMNH 34500) to warrant no further comparisons.
The coracoid of Dakotasuchus is complete in both specimens and contrasts with coracoids of both Woodbinesuchus and Terminonaris (Fig. 3A–D). The coracoid in the holotype of Woodbinesuchus byersmauricei (SMU 74626) is broken and displaced from life position at the midshaft; even so it is evidently much smaller and more compressed mediolaterally than either OMNH 34500 or the holotype specimen of Dakotasuchus kingi. The dorsal head in SMU 74626 is nearly complete and has a rostrodorsal edge that is flattened, giving the head a diamond-shaped appearance, strikingly different than the more rounded dorsal head in Dakotasuchus (Fig. 3A–C). In Woodbinesuchus the shaft of the coracoid has a rounded lateral ridge that is absent or very low in OMNH 34500. The ventral end of the coracoid differs in general shape between the two genera. In Woodbinesuchus the ventral end forms a rounded, medially directed boot that is decidedly more pronounced and less blade-like than in Dakotasuchus. Compared to both Dakotasuchus and Woodbinesuchus, the midshaft of the coracoid of Terminonaris is substantially less constricted (Wu et al. 2001: fig. 2D). Furthermore, Terminonaris possesses a knob-like process for the insertion of the caput coracoideum of the triceps brachii muscle, not seen in either of the other taxa. The proximal head in Terminonaris is also uniquely triangular, resulting from the dorsal placement of the rostral margin (Wu et al. 2001). The coracoid of OMNH 34500 also compares favorably to Goniopholis stovalli (OMNH 798 and OMNH 2382) from the Morrison Formation of Oklahoma: similarities include a curved rostral border, highly rounded articular surface of the glenoid, and similarly curved coracoid shaft; however, the dorsal head is more expanded and overall the coracoid is larger in Dakotasuchus. The coracoid resembles those of Anteophthalmosuchus hooleyi (IRSNB R47; Martin et al. 2016a) from the Lower Cretaceous of Bernissart, Belgium. Both species have a highly curved (medially) cranial edge, a wide glenoid, and a blade-like ventral head; they differ, however, in the shape of the dorsal head, specifically the rostromedial border, which extends more ventrally in Dakotasuchus.
The descriptions of dorsal scutes of both
specimens of Dakotasuchus are based on at
least one complete dorsal plate in OMNH 34500, as well as impressions of
the entire dorsal series in the holotype. In general, the pits on the
dorsal face are more variable in size than those seen in Woodbinesuchus.
Most strikingly, however, is the caudolateral edge, which is rounded in
both Dakotasuchus and Terminonaris
(among other coelognathosuchians; Martin et al.
2016b), in contrast to the square edge seen in Woodbinesuchus
(Fig. 3E–H). The cranial dorsal process and corresponding groove
on the caudal side is angled craniolaterally, similar to the condition in
Woodbinesuchus and unlike the mostly straight
morphology of Terminonaris or the medially
curved processes of Anteophthalmosuchus hooleyi
(see Martin et al. 2016a) and
some specimens of Sunosuchus (Wu
et al. 1996).
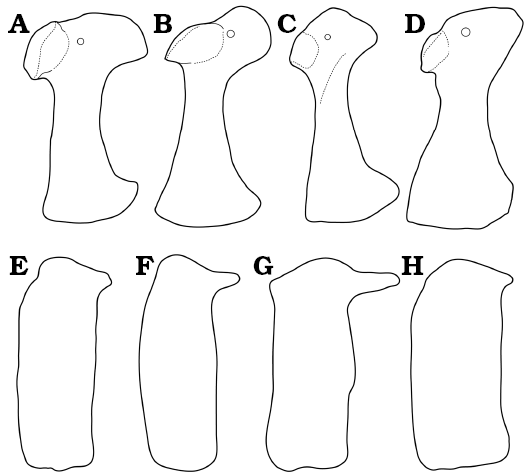 |
Fig. 3. Comparative morphology of coracoids (A–D) and dorsal scutes (E–H) in coelognathosuchians from the medial Cretaceous of North America. Right coracoids in lateral view and right dorsal scutes in ventral view. A, E. Dakotasuchus kingi Mehl, 1941, OMNH 34500, Mussentuchit Member of the Cedar Mountain Formation (Cenomanian), Utah, USA. B, F. Dakotasuchus kingi Mehl, 1941, KWU uncatalogued (holotype), Dakota Formation (Cenomanian), Kansas, USA. C, G. Woodbinesuchus byersmauricei Lee, 1997, SMU 74626 (holotype), Woodbine Formation (Cenomanian), Texas, USA. D, H. Terminonaris robusta Wu, Russell, and Cumbaa, 2001, SMNH P2411.1 (coracoid is inverted), Keld Member of the Favel Formation (Turonian), Saskatchewan, Canada. Images modified from Mehl (1941), Lee (1997), and Wu et al. (2001). Images are not to scale. |
Size reconstruction and ecological interpretations.—The paucity of large coelognathosuchians from the Cenomanian prompted us to calculate body size estimates, including both total length and body mass, for Dakotasuchus kingi, based on OMNH 34500. Body size analyses normally use femoral lengths, as femora tend to give the best indication of body size in crocodilians (Farlow et al. 2005), but unfortunately, the femur for Dakotasuchus is unknown. Two body size estimates were made: (i) by scaling up from Mehl’s (1941) measurements and body length estimate; and (ii) by calculating a femoral length, using the average radius/femur ratios calculated by Livingston et al. (2009: table 5). Once the femur length was calculated, we then estimated body length and body mass using the relevant equations from Farlow et al. (2005: table 3). Coracoid measurements were used to estimate relative size between OMNH 34500 and the KWU specimen, as it is the best represented bone. From the resulting coracoid length measurements (OMNH 34500 = 235 mm and KWU uncatalogued = 190 mm [Mehl 1941]), OMNH 34500 is nearly 20% larger than the KWU specimen. Assuming a direct relationship between coracoid length and body length and using Mehl’s (1941) length estimation, the resulting total body length for OMNH 34500 is estimated to be 3.7–4.5 m. Using the second body size estimate based on the length of the radius, the femur was estimated to be 375.3 mm long, resulting in a total body length of 5.4 m and a body mass of ~700 kg. Combining these techniques, a conservative body length estimate would be anywhere between 3.7 m and 5.4 m long. These estimates make Dakotasuchus slightly longer than the longest modern adult American Alligator (Alligator mississippiensis, 3.1–4.3 m measured by Woodward et al. 1995) and roughly equal to the maximum length of the Salt-Water Crocodile (Crocodylus porosus, 5.4–5.8 m estimated by Greer 1974). Garrison et al. (2007) hypothesized that the larger crocodilians from the Mussentuchit Member would have likely competed with predatory theropods for prey, such as juvenile or subadult ornithopods; much in the same way that modern Nile crocodiles (Crocodylus niloticus) steal kills from mammalian carnivores when the prey item takes to the water for refuge (Mitchell et al. 1965). While Mehl (1941) suggests that Dakotasuchus was more terrestrial than modern crocodilians based on its robust pectoral girdle, there is no reason to assume Dakotasuchus was not at home in the water as well. Even if Dakotasuchus was not in direct competition with terrestrial theropods, the most conservative length estimates would still make it one of the largest aquatic predators in the Mussentuchit ecosystem.
Interestingly, the holotype specimen and a referred specimen (Vaughn 1956) were likely from a brackish water, river delta (Hattin 1967), implying that the species lived close to the paleo-shoreline. Conversely, the newly described specimen here comes from a more terrestrial, freshwater lithology. Given that other coelognathosuchians from the Cenomanian of North America are known from marine deposits (e.g., Terminonaris robusta; Adams et al. 2011), and that Dakotasuchus is now know from both freshwater and brackish environments, there is good evidence Dakotasuchus was euryhaline, capable of inhabiting a wide-range of salinities. This could, in-part, explain the relatively wide geographic distribution for this species.
Conclusions
While the holotype for Dakotasuchus kingi is exceptionally preserved within a concretion, key anatomical features are obscured or represented by natural molds, making detailed comparisons to mesoeucrocodylians from the medial Cretaceous difficult. A new specimen of D. kingi prompts a full description of obscured elements, as well as new elements unknown in the holotype and comparisons to medial Cretaceous mesoeucrocodylians. These comparisons confirm the taxonomic uniqueness of Dakotasuchus, despite the absence of cranial material, based largely on the coracoid and dorsal scutes. OMNH 34500 is nearly 20% larger than the holotype of D. kingi, with a body length of 3.7 m to 5.4 m, resulting in a predator as large or larger than the modern American Alligator in the Mussentuchit ecosystem.
Acknowledgements
We thank Stephanie Welter (Kansas Wesleyen University, Salina, KS, USA) and Dale Winkler (Shuler Museum of Paleontology, Dallas, TX, USA) for access to specimens in their care. We thank Michael Everhart and Jeremy Martin (both Université Lyon, France) for their thoughtful and constructive reviews of the manuscript. JAF thanks Michael Everhart and Jonathan Tennant (both Université Lyon) for additional advice and assistance, which greatly benefited this paper. It is a pleasure to acknowledge the help, cooperation, and support of Scott Madsen (Salt Lake City, Utah, USA), who found the specimen described herein; the Judd family of Castle Dale, Utah; and Blaine Miller and Michael Leschin of the Price Field Office, Bureau of Land Management, Price, Utah. Partial support for this research was provided by grants from the National Geographic Society (4761-91, 5021-92) and the National Science Foundation (BSR 8906992, DEB 9401994).
References
Adams, T.L., Polcyn, M.J., Mateus, O., Winkler, D.A., and Jacobs, L.L. 2011. First occurrence of the long-snouted crocodyliform Terminonaris (Pholidosauridae) from the Woodbine Formation (Cenomanian) of Texas. Journal of Vertebrate Paleontology 31: 712–716. Crossref
Carpenter, K. 2014. Where the sea meets the land—the unresolved Dakota problem in Utah. Utah Geological Association Publication 43: 357–372.
Cifelli, R.L., Kirkland, J.I., Weil, A., Deino, A.L., and Kowallis, B.J. 1997. High-precision 40Ar/39Ar geochronology and the advent of North America’s Late Cretaceous terrestrial fauna. Proceedings of the National Academy of Sciences USA 94: 11163–11167. Crossref
Cifelli, R.L., Nydam, R.L., Weil, A., Gardner, J.D., Kirkland, J.I., Eaton, J.G., and Madsen, S.K. 1999. Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County, Utah: the Mussentuchit local fauna. In: D.D. Gillette (ed.), Vertebrate Fossils of Utah, 219–242. Utah Geological Survey, Salt Lake City.
Cope, E.D. 1875. Check-list of North American Batrachia and Reptilia with a systematic list of the higher groups and an essay on geographical distribution based on the specimens in the US National Museum. Bulletin of the United States National Museum 1: 1–104.
Everhart, M.J. 2005. Oceans of Kansas—A Natural History of the Western Interior Sea. 322 pp. Indiana University Press, Bloomington.
Farlow, J.O., Hurlburt, G.R., Elsey, R.M., Britton, A.R.C., and Langston, W. 2005. Femoral dimensions and body size of Alligator mississippiensis: estimating the size of extinct mesoeucrocodylians. Journal of Vertebrate Paleontology 25: 354–369. Crossref
Fiorillo, A.R. 1999. Non-mammalian microvertebrate remains from the Robison Eggshell Site, Cedar Mountain Formation (Early Cretaceous), Emery County, Utah. In: D.D. Gillette (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication 99-1: 259–268.
Frederickson, J.A. and Cifelli, R.L. 2017. New Cretaceous lungfishes (Dipnoi, Ceratodontidae) from western North America. Journal of Paleontology [published online]. Crossref
Garrison, J.R., Jr., Brinkman, D.B., Nichols, D.J., Layer, P., Burge, D.L., and Thayn, D. 2007. A multidisciplinary study of the Lower Cretaceous Cedar Mountain Formation, Mussentuchit Wash, Utah: a determination of the paleoenvironment and paleoecology of the Eolambia caroljonesa dinosaur quarry. Cretaceous Research 28: 461–494. Crossref
Greer, A.E. 1974. On the maximum total length of the salt-water crocodile (Crocodylus porosus). Journal of Herpetology 8: 381–384. Crossref
Hattin, D.E. 1967. Stratigraphic and paleoecologic significance of macroinvertebrate fossils in the Dakota Formation (Upper Cretaceous) of Kansas. In: C. Teichert and E.L. Yochelson (eds.), Essays in Paleontology and Stratigraphy—R.C. Moore Commemorative Volume. University of Kansas Special Publication 2: 570–589.
Hay, O.P. 1930. Second Bibliography and Catalogue of the Fossil Vertebrata of North America. 1074 pp. Carnegie Institute of Washington, Washington, DC.
Kirkland, J.I., Britt, B., Burge, D.L., Carpenter, K., Cifelli, R.L., DeCourten, F., Eaton, J.G., Hasiotis, S., and Lawton, T. 1997. Lower to middle Cretaceous dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution and biogeography. Brigham Young University Geology Studies 42 (2): 69–103.
Kukihara, R. and Lockley, M.G. 2012. Fossil footprints from the Dakota Group (Cretaceous) John Martin Reservoir, Bent County, Colorado: New insights into the paleoecology of the dinosaur freeway. Cretaceous Research 33: 165–182. Crossref
Langston, W., Jr. 1974. Nonmammalian Comanchean tetrapods. Geoscience and Man 8: 77–102.
Lee, Y.-N. 1997. The Archosauria from the Woodbine Formation (Cenomanian) in Texas. Journal of Paleontology 71: 1147–1156. Crossref
Livingston, V.J., Bonnan, M.F., Elsey, R.M., Sandrik, J.L., and Wilhite, D.R. 2009. Differential limb scaling in the American Alligator (Alligator mississippiensis) and its implications for archosaur locomotor evolution. Anatomical Record 292: 787–797. Crossref
Lockley, M.G. 2010. A solution to the Mehliella mystery: tracking, naming, identifying and measuring the first crocodylian trackway reported from the Cretaceous (Dakota Group, Colorado). New Mexico Museum of Natural History and Science Bulletin 51: 157–164.
Main, D.J. 2013. Appalachian Delta Plain Paleoecology of the Cretaceous Woodbine Formation at the Arlington Archosaur Site. 585 pp. Ph.D. Thesis, University of Texas at Arlington, Arlington.
Martin, J.E., Delfino, M., and Smith, T. 2016a. Osteology and affinities of Dollo’s goniopholidid (Mesoeucrocodylia) from the Early Cretaceous of Bernissart, Belgium. Journal of Vertebrate Paleontology [published online]. Crossref
Martin, J.E., Lauprasert, K., Buffetaut, E., Liard, R., and Suteethorn, V. 2014. A large pholidosaurid in the Phu Kradung Formation of north-eastern Thailand. Palaeontology 57: 757–769. Crossref
Martin, J.E., Raslan-Loubatié, J., and Mazin, J.M. 2016b. Cranial anatomy of Pholidosaurus purbeckensis from the Lower Cretaceous of France and its bearing on pholidosaurid affinities. Cretaceous Research 66: 43–59. Crossref
Mehl, M.G. 1941. Dakotasuchus kingi, a crocodile from the Dakota of Kansas. Denison University Journal of Sciences and Laboratories 36: 47–65.
Mitchell, B.L., Shenton, J.B., and Uys, J.C.M. 1965. Predation on large mammals in the Kafue National Park, Zambia. Zoologica Africana 1: 297–318. Crossref
Mook, C.C. 1934. A new species of Teleorhinus from the Benton Shales. American Museum Novitates 702: 1–11.
Osborn, H.F. 1904. Teleorhinus browni—a teleosaur in the Fort Benton. Bulletin of the American Museum of Natural History 20: 239–240.
Scott, R.W. 1970. Paleoecology and paleontology of the Lower Cretaceous Kiowa Formation, Kansas. University Kansas Paleontological Contributions 52: 1–94.
Vaughn, P.P. 1956. A second specimen of the Cretaceous crocodile Dakotasuchus from Kansas. Transactions of the Kansas Academy of Sciences 59: 379–381. Crossref
Walker, A.D. 1970. A revision of the Jurassic reptile Hallopus victor (Marsh), with remarks on the classification of crocodiles. Philisophical Transactions of the Royal Society of London B257: 323–372. Crossref
Whetstone, K. and Whybrow, P. 1983. A “cursorial” crocodilian from the Triassic of Lesotho (Basutoland), southern Africa. Occasional Papers of the Museum of Natural History of the University of Kansas 106: 1–37.
Woodward, A.R., White, J.H.,
and Linda, S.B. 1995. Maximum size of the alligator (Alligator
mississippiensis). Journal
of Herpetology 29: 507–513. Crossref
Wu, X.-C., Brinkman, D.B., and Russell, A.P. 1996. Sunosuchus junggarensis sp. nov. (Archosauria: Crocodyliformes) from the Upper Jurassic of Xinjiang, People’s Republic of China. Canadian Journal of Earth Sciences 33: 606–630. Crossref
Wu, X.-C., Russell, A.P., and Cumbaa, S. 2001. Terminonaris (Archosauria: Crocodyliformes): new material from Saskatchewan, Canada, and comments on its phylogenetic relationships. Journal of Vertebrate Paleontology 21: 492–514. Crossref
Acta Palaeontol. Pol. 62 (2):
279–286, 2017
https://doi.org/10.4202/app.00338.2016