

A new subdisarticulated machaeridian from the Middle Devonian of China: Insights into taphonomy and taxonomy using X-ray microtomography and 3D-analysis
BENJAMIN GÜGEL, KENNETH DE BAETS, IWAN JERJEN, PHILIPP SCHUETZ, and CHRISTIAN KLUG
Gügel, B., De Baets, K., Jerjen, I., Schuetz, P., and Klug, C. 2017. A new subdisarticulated machaeridian from the Middle Devonian of China: Insights into taphonomy and taxonomy using X-ray microtomography and 3D-analysis. Acta Palaeontologica Polonica 62 (2): 237–247.
Machaeridians are an extinct group of armoured annelids, which are mainly known from isolated sclerites present from the Ordovician to the Permian. Based on articulated specimens with preserved soft-tissues and trace fossils, derived machaeridians are interpreted to have an infaunal burrowing mode of life. However, the taphonomy of sclerite associations is still largely unstudied. We herein investigated associated sclerites from the Middle Devonian of China using micro-computer tomography and 3D-analysis. These sclerites belong to a single individual and lie in close proximity. The absence of indications for current alignment, major bioturbation or other processes causing a disarticulation as reflected in the randomly arranged dacryoconarids suggest that the sclerites became disarticulated in the course of the normal decay processes, perhaps aided by scavenging and incomplete burial. The unique morphology of the sclerites indicates that the specimen presented here belongs to a previously undescribed species, which we describe herein as Lepidocoleus kuangguoduni sp. nov.
Keywords: Annelida, Lepidocoleidae, tomography, SPIERS, taphonomy, Devonian, China, Guangxi.
Benjamin Gügel [benjamin.guegel@gmail.com] and Kenneth De Baets [kenneth.debaets@fau.de] (corresponding author), Geozentrum Nordbayern, Friedrich-Alexander Universität Erlangen-Nürnberg, Loewenichstraße 28, 91054 Erlangen, Germany. Iwan Jerjen [iwan.jerjen@psi.ch], Department of Information Technology and Electrical Engineering, ETH Zürich, Gloriastrasse 35, 8092 Zürich, Switzerland.
Philipp Schuetz [philipp.schuetz@hslu.ch], Technik & Architektur, Hochschule Luzern, Technikumstrasse 21, 6048 Horw, Switzerland.
Christian Klug [chklug@pim.uzh.ch], Paläontologisches Institut und Museum, Universität Zürich, Karl-Schmid-Strasse 4, 8006 Zürich, Switzerland.
Received 30 January 2017, accepted 11 April 2017, available online 24 May 2017.
Copyright © 2017 B. Gügel et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Machaeridians are an enigmatic group of worm-like, multi-sclerite bearing benthic marine invertebrates. Their calcitic sclerites are widely distributed in marine rocks of Early Ordovician to middle Permian age (Dzik 1986; Caron 2008; Högström et al. 2009). Their affinities were long debated and they have been variously assigned to mollusks (Dzik 1986; Klug et al. 2008), arthropods (Barrande 1872), echinoderms (Withers 1926), and annelids (Bengtson 1978, 1979). An exceptionally well-preserved plumulitid machaeridian with preserved parapodia and chaetae placed them firmly within the annelids (Vinther et al. 2008; Parry et al. 2014). It is, however, still unclear if they are outside the annelid crown group (Caron 2008) or inside the annelid crown as stem aphroditaceans (Vinther and Briggs 2009; Vinther and Rudkin 2010; Parry et al. 2014).
Furthermore, they are the only known annelids with an armour of calcitic plates arranged in two or four rows. Mostly their isolated sclerites are found (Dzik 1986; Ekleris and Radzevičius 2014), while articulated skeletons (Jell 1979; Adrain et al. 1991; Högström and Taylor 2001; Klug et al. 2008; Vinther and Briggs 2009; De Baets et al. 2010; Vinther and Rudkin 2010) and particularly exceptionally preserved specimens are more rare (Vinther et al. 2008; Högström et al. 2009). Therefore, machaeridians have commonly been described and classified into three families (Turrilepadidae, Lepidocoleidae, Plumulitidae) based on the morphology of their plates. Adrain (1992) grouped the Turrilepadidae and Plumulitidae within the Turrilepadomorpha (Adrain 1992; Hints et al. 2004), largely due to their quadriseriate skeleton as opposed to the Lepidocoleidae, which was placed in the Lepidocoleomorpha, which he considered as biseriate—although Dzik (1986) and Högström (1997) clearly demonstrated that some lepidocoleids are quadriseriate. Parry et al. (2014) introduced the clade Cuniculepadida to unite the family Turrilepadidae and Lepidocoleidae based on the shared lateral compression of the shell plates, which enclose the body, and the lateral displacement of the outer shell plates. Turrilepadids and lepidocoleids do not only share the lateral compression of the shell plates, but also share the scars for muscle attachment and thicker shell plates. Cuniculepadida are more streamlined in their form than basal plumulitids, so that most of these characters have been interpreted as adaptations to an infaunal or burrowing mode of life (Dzik 1986; Hints et al. 2004). The infaunal mode of life is also supported by sinuous trace fossils associated with lepidocoleids (Vinther and Briggs 2009; but compare, Seilacher and Gishlick 2014). Högström et al. (2009) demonstrated the absence of parapodial lobes using a 3D-reconstruction of a lepidocoleid, which can also be inferred to be absent in turrilepadids (Vinther and Briggs 2009) and further corroborate the infaunal lifestyle of this group. Reduction of such structures is commonplace among infaunal polychaete taxa and the current evidence from the stem group suggests that all annelids evolved from an epibenthic ancestor with prominent parapodia (Parry et al. 2016) as in Plumulites bengtsoni (Vinther et al. 2008).
During field work in China, a set of associated sclerites was found in Eifelian rocks of the Nandan Formation. Herein, we use imagery and 3D-prints based on a micro-CT scan to determine the affinity of these sclerites and to assess the underlying reason for the association of these sclerites. We also discuss the depositional conditions and taphonomy of this fossil assemblage. The sclerites are associated with dacryoconarids, which can provide important information on the depositional environment (e.g., current alignment and degree of bioturbation).
Institutional abbreviations.—PIMUZ, Paleontological Institute and Museum, University of Zürich, Switzerland.

Fig. 1. Machaeridian annelid Lepidocoleus kuangguoduni sp. nov., Nandan Formation, Eifelian, near Napiao, Guangxi (China). A. The main plate containing most machaeridian sclerites. B. The counterplate of the same specimen (it was glued back onto the slab prior to CT-scanning); note the limonitic filling of the rugae and the chaotic arrangement of the plates.
Material and methods
The specimen (Fig. 1) was discovered in the Nandan Formation near Napiao, Guangxi, China (Fig. 2; coordinates: 24°58’40.6’’ N, 107°23’50.3’’ E) in Eifelian sediments associated with dacryoconarids and trilobites. The calcitic plates are currently preserved as iron oxide (which was probably oxidized from pyrite) and partially hollow cavities in claystone. The fossil specimen consisting of a main part and its fragmented counterpart is housed at the Paleontological Institute and Museum of the University of Zürich (PIMUZ 32127) and the tomograms can be obtained upon request. A video of the the three-dimensional model is provided as SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app62-Gugel_etal_SOM.pdf.
The sample (PIMUZ 32127) was scanned on the 8th of December 2011 at the Swiss Federal Laboratories for Materials Science & Technology (EMPA) by PS and IJ with a Viscom XT9160-TXD micro-CT-scanner at 150 kV and 160 µA—a Perkin Elmer detector was used. A beam hardening correction algorithm was used with a parameter value of 1.5. The CT-reconstructed volume was 2000×2000×700 voxels. Three-dimensional reconstructions and an animation were produced using these seven hundred images (tomograms) in the x-z-plane by using the free software SPIERS (Sutton et al. 2012; http://spiers-software.org). A fixed threshold value was manually chosen to separate sclerites and dacryoconarids from the matrix, because the objects of focus are partially limonitic and partially hollow in the claystone. These regions of interest were defined using the masking system in SPIERS (Sutton et al. 2014). Different coloured masks and arbitrary numbering were used in the final representation to distinguish individual plates and a uniform colour was used to mark dacryoconarids. We selected 70 specimens which could be clearly determined as dacryoconarids and where the apices were clearly visible. The orientations of the selected dacryoconarids were measured using the free 3D-Creation Suite Blender (blender.org) and these data were plotted using the software Stereonet 9 (Allmendinger et al. 2011; Cardozo and Allmendinger 2013). Moreover, two shell-plate-models were enlarged by factor 25 and printed using a custom-made “Fused Deposition Modeling” 3D-printer.
The terminology of machaeridians employed in this article is based on Adrain et al. (1991), Adrain (1992), Högström (1997), Högström and Taylor (2001), and Högström et al. (2009).
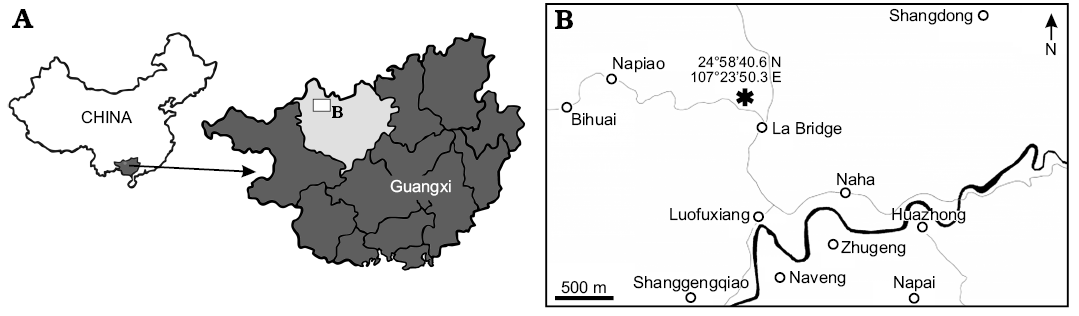
Fig. 2. Maps of China and Guangxi (A) adapted from “Croquant” on Wikimedia (licensed under CC BY 3.0). B. Map showing the region of machaeridian locality (asterisk); adapted from Google Maps.
Results
Machaeridian sclerites.—The 3D-analysis (Fig. 3) resulted in a total of sixteen objects which were attributed to be machaeridian sclerites or parts of sclerites. Only one object could not be determined (object 3) with certainty. Seven of these sixteen objects (Fig. 4) are virtually complete sclerites with the typical morphology of “dorsal flanges” which are a characteristic trait of lepidocoleids. In spite of visible indications for surface-structures on the raw sample (e.g., rugae, muscle scars, inner grooves, etc.), the 3D-objects did not show any usable texture for further analysis. Some surface structures are here considered artefacts, which are a result of the moderate scanning-resolution and 3D-reconstrution process. It is, however, possible to discern some of the objects by morphology only. Sclerites 1, 4, and 6 (group I for future reference) show a similar shape and profile. Because of the course of the rugae and in comparison with Lepidocoleus sarlei (Högström and Taylor 2001), we interpret the sclerites of group I as belonging to the right side of the animal. Sclerites 2, 5, 7, and 10 (group II) share a resemblance in slightly different shape and profile and thus are sclerites of the left series of the machaeridian. The less complete sclerites (Fig. 5) were not sorted into one of these two groups because of the lack of morphological features, but are considered in regard of taphonomic and systematic analysis, and are described here.
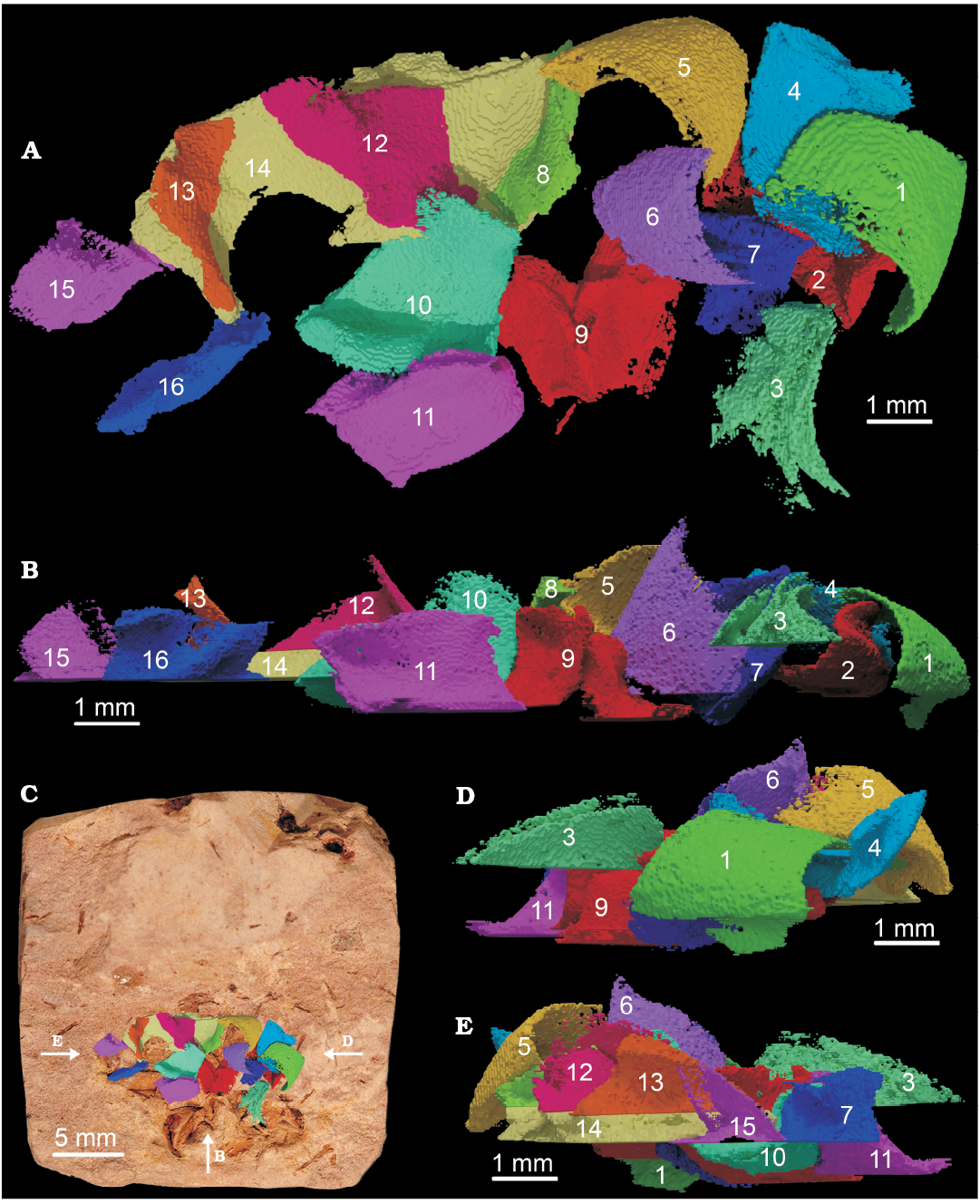
Fig. 3. Overview over the assemblage of the sixteen 3D-objects which were created from different viewpoints. Orthographic top (A) and front (B) views. Orthographic top view (C), projected on the sample to show the position of the 3D-model in the correct position on the x-y-plane and corresponding viewing directions. Orthographic left side (D) and right side (E) views.
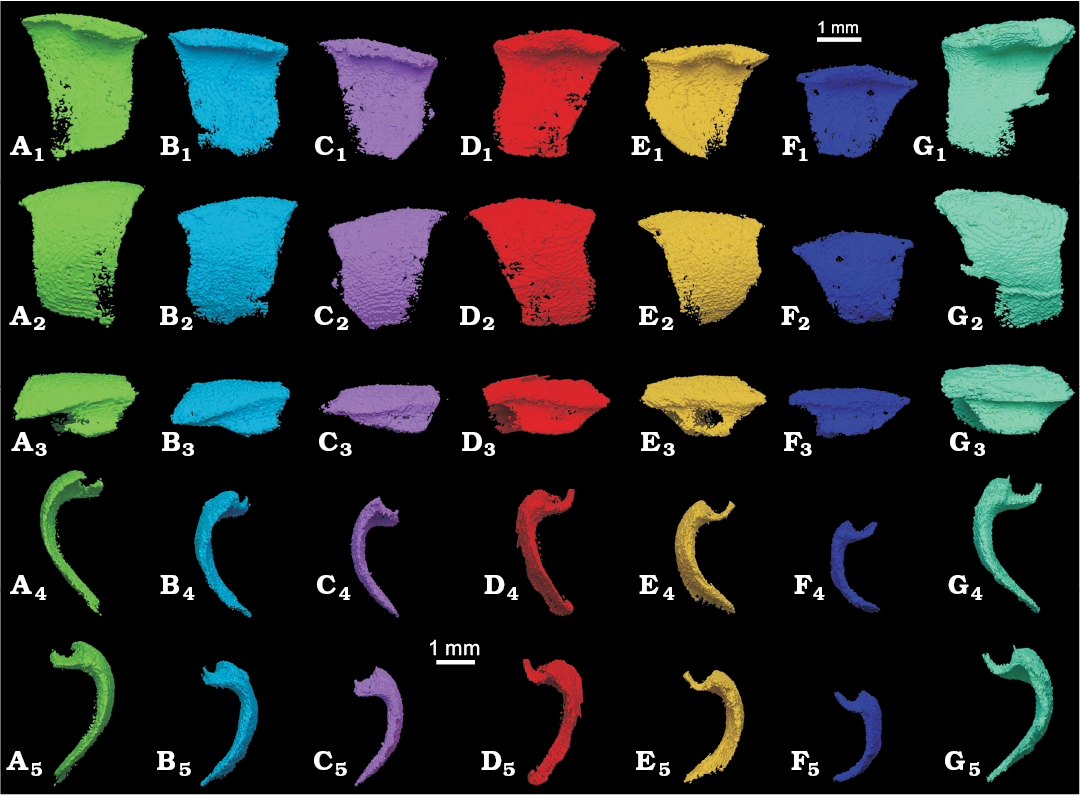
Fig. 4. Overview of the almost complete sclerites from the 3D-analysis (orthographic perspective). Group I: sclerites 1, 4, 6 (A–C) and group II: sclerites 2, 5, 7, 10 (D–G). Internal (A1–G1), lateral (A2–G2), dorsal (A3–G3), posterior (A4–G4), and anterior (A5–G5) views.
Object 9 (Fig. 5B) shows parts of two sclerites, which form a fused void. Parts of this 3D-model could be identified as a dorsal flange. Objects 15 and 16 (Fig. 5C) are parts of one sclerite, which is corroborated by the positioning of these objects in relation to each other. The missing middle part is explained through the positioning of this sclerite near the border area of the sample. Object 11 (Fig. 5D) is another part of a sclerite which features a dorsal flange.
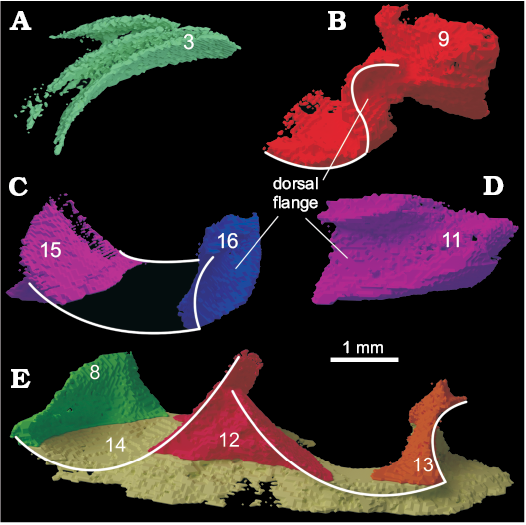 |
Fig. 5. Overview of the other objects found in the sample. A. Object 3 could not be identified with certainty, but is likely part of a sclerite from the flank. B. Sclerite 9 might be from the dorsal articulation. C. Objects 15 and 16 might belong to the same, incomplete sclerite. D. Sclerite 11 preserves only the dorsal flange. E. Objects 8, 12, 13, and 14 might actually be parts of two sclerites as indicated by the white lines. |
3D-printed sclerites: Two sclerites (1 and 2) were selected for 3D-printing (Fig. 6) because they were closely positioned in a subarticulated state and therefore seemed promising for further investigation pertaining lateral movement of the shell plates. The spatial arrangement suggests that the sclerites could be moved both outward, back and forth as well as rotated relative to each other. This has been corroborated before for other species, where articulated specimens, which are torted or dorsoventrally bent, have been found (Vinther and Briggs 2009). Future studies could potentially use such prints of more highly resolved scans to investigate the articulation constraints in order to figure out different relative positions of the sclerites in the pair.
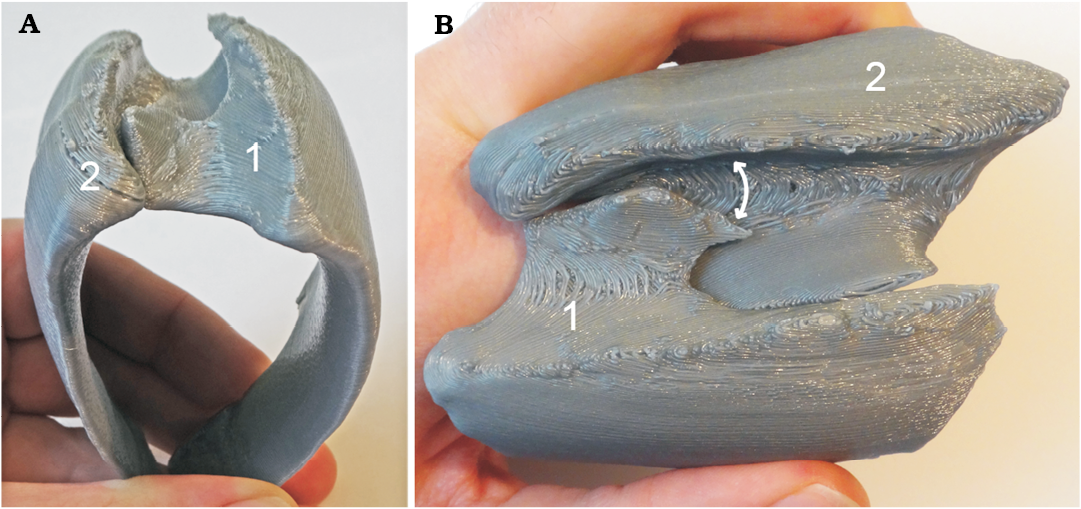
Fig. 6. View of a pair of 3D-prints of articulated right (1) and left (2) sclerites from obliquely posterior (A) and dorsal (B) views. Note the perfect fit of the sclerites. Within the hinge of sclerite 1 in B, the indentation on the right of the hinge flange is an artefact from tresholding (probably, the shell was too thin in that place). Sclerites 1 and 2 were enlarged 25 times (for original dimensions see Fig. 4).
Dacryoconarids.—Measuring the spatial position of the dacryoconarids (Fig. 7) showed that they are randomly arranged in the bedding plane (see Fig. 8B) and are orientated more or less (sub-)horizontally (the mean dip value is 8°; Fig. 8B, Table 1). However, a trend in the arrangement of dacryoconarids is evident; they are approximately positioned in the “north-south”-orientation of the sample (see Fig. 8B), if the positioning of the apices and “open ends” of the dacryoconarids is heeded. Dacryoconarids, which are orientated in relation to the sample between 310° and 30° are all deposited in a way, that the apices are higher than an imaginary x-y-plane, whereas dacryoconarids between 155° and 215° are deposited in a way, that the apices are below an imaginary x-y-plane. All other angles show a random distribution to the positioning of the apices.
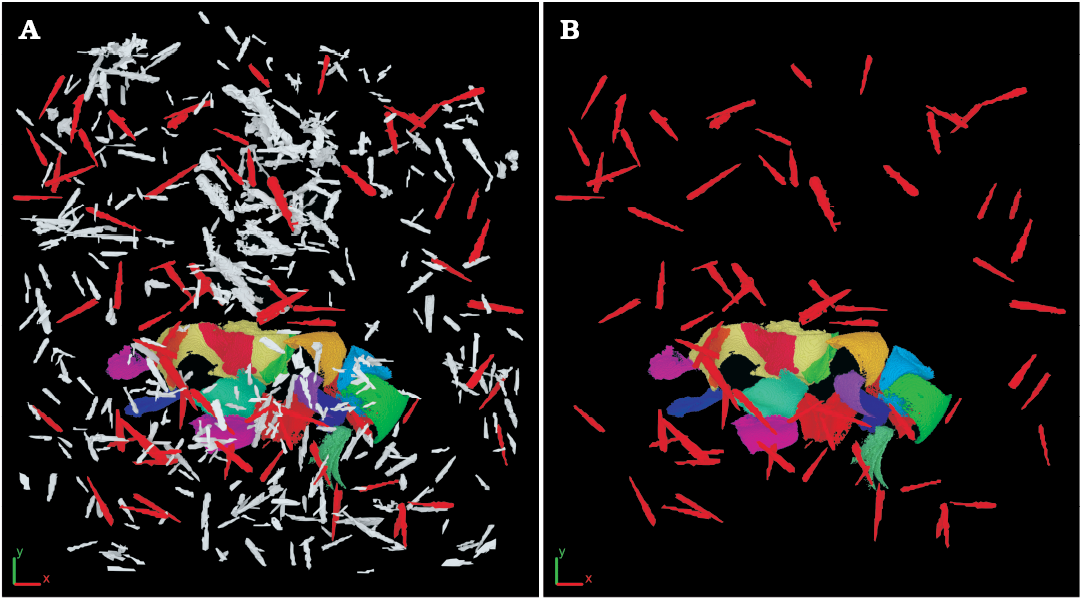
Fig. 7. Overview of objects interpreted as dacryoconarids surrounding the machaeridian sclerites. A. All objects including the ones discarded for further analysis (light grey). B. Dacryoconarids selected for measurements (red).
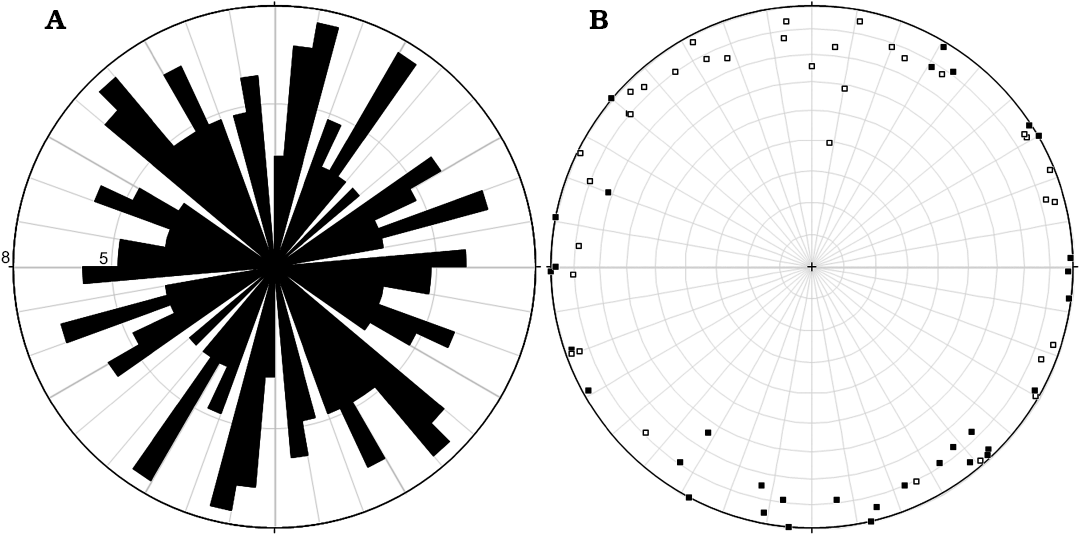
Fig. 8. Orientation of the 70 dacryoconarids in the sample. A. Rose diagram showing the lineations of the dacryoconarids (numbers 5 and 8 refer to dacryoconarid counts; note that both the tip and aperture where counted of each object resulting in double counts). B. Rose diagram showing dacryoconarids whose apices are higher (open rectangles) and lower (closed rectangles) positioned than their corresponding open ends in relation to an imagined x-y-plane.
Table 1. Measured orientations of the dacryoconarids.
|
Dacryoconarid |
Azimuth |
Dip |
Dacryoconarid |
Azimuth |
Dip |
|
1 |
57 |
1 |
36 |
225 |
-11 |
|
2 |
112 |
-6 |
37 |
68 |
-2 |
|
3 |
141 |
4 |
38 |
58 |
-5 |
|
4 |
147 |
11 |
39 |
268 |
-10 |
|
5 |
314 |
-4 |
40 |
0 |
-25 |
|
6 |
157 |
10 |
41 |
191 |
5 |
|
7 |
290 |
18 |
42 |
291 |
-10 |
|
8 |
136 |
3 |
43 |
137 |
2 |
|
9 |
108 |
-4 |
44 |
24 |
-14 |
|
10 |
310 |
-10 |
45 |
10 |
-32 |
|
11 |
269 |
0 |
46 |
325 |
-10 |
|
12 |
250 |
-6 |
47 |
185 |
0 |
|
13 |
8 |
-50 |
48 |
142 |
13 |
|
14 |
34 |
-12 |
49 |
354 |
-7 |
|
15 |
317 |
-7 |
50 |
214 |
11 |
|
16 |
59 |
-5 |
51 |
91 |
2 |
|
17 |
281 |
0 |
52 |
75 |
-4 |
|
18 |
154 |
10 |
53 |
154 |
-10 |
|
19 |
20 |
-12 |
54 |
6 |
-17 |
|
20 |
191 |
4 |
55 |
270 |
2 |
|
21 |
193 |
15 |
56 |
31 |
3 |
|
22 |
333 |
-12 |
57 |
36 |
9 |
|
23 |
60 |
0 |
58 |
74 |
-8 |
|
24 |
212 |
26 |
59 |
165 |
5 |
|
25 |
136 |
13 |
60 |
187 |
11 |
|
26 |
174 |
11 |
61 |
332 |
-3 |
|
27 |
338 |
-15 |
62 |
167 |
0 |
|
28 |
88 |
1 |
63 |
310 |
0 |
|
29 |
310 |
-10 |
64 |
120 |
-1 |
|
30 |
251 |
3 |
65 |
296 |
-2 |
|
31 |
97 |
1 |
66 |
11 |
-5 |
|
32 |
208 |
0 |
67 |
250 |
-3 |
|
33 |
275 |
-11 |
68 |
31 |
12 |
|
34 |
139 |
3 |
69 |
241 |
3 |
|
35 |
353 |
-13 |
70 |
119 |
3 |
Systematic palaeontology
Phylum Annelida Lamarck, 1809
Class Machaeridia Withers, 1926
Family Lepidocoleidae Clarke, 1896
Genus Lepidocoleus Faber, 1886
Type species: Lepidocoleus jamesi (Hall and Whitfield, 1875), Cincinnatian (Upper Ordovician) of Ohio, USA.
Remarks.—The presence of a tongue-and-groove hinge, the absence of additional rows of sclerites, sclerite form, thickness and the presence of a dorsal groove corroborate the assignment of the here described material as Lepidocoleidae. It is worth stressing that this sclerite assemblage does not contain any obvious anterior outer sclerites as recognized in plumulitids (Vinther and Rudkin 2010) and possibly turrilepadids (Adrain et al. 1991) as they are never seen in lepidocoleids (so far at least). Furthermore, we did not find evidence for the presence of outer sclerites at all suggesting that we are dealing with a derived lepidocoleid (Parry et al. 2014). Unfortunately, muscle attachment structures cannot be distinguished with our scan-resolution. However, the shape of the rugae is consistent with the genus Lepidocoleus known from the Early Ordovician until the Givetian (Devonian).
In Fig. 9, we list species included in Lepidocoleus according to their relative age with some important characters and their occurrence. Devonian forms are listed at the bottom. Note that the here described specimen appears to be one of the youngest representatives of the genus. We did not include Aulakolepos elongatum Dzik, 1994 and Plicacoleus robustus Dzik, 1986 which might also belong to Lepidocoleus, but for the moment are still assigned to different genera pending a proper revision of the genus which falls outside the scope of our study.
Stratigraphic and geographic range.—Ordovician to Middle Devonian of northern America, Europe, southeast Asia, and northern Africa.

Fig. 9. Comparison between the known species of Lepidocoleus and L. kuangguoduni sp. nov. with number of sclerites, age, and geographic occurrence indicated; lateral (A) and dorsal (E) views of the fossils, images of sclerites (B), outlines of sclerites (C), cross sections, to show the proportions of the dorsal depression (D).
Lepidocoleus kuangguoduni sp. nov.
Figs. 1, 3–7, 9.
Etymology: Species named after Kuang Guodun, honouring his contributions to research on Paleozoic invertebrates.
Holotype: PIMUZ 32127, part and its fragmented counterpart, over 20 sclerites.
Type locality: 500 m E of the village Napiao and about 1 km NW of Luofuxiang, NW Guangxi, China.
Type horizon: Tentaculite claystone with the trilobite Ductina vietnamica, Eifelian, Nandan Formation, Middle Devonian.
Diagnosis.—Lepidocoleus with presumably a low number of sclerites (around 22). The slightly alternating sclerites are elongated dorsoventrally with pronounced anterior and posteriorly projecting points as well as an arched dorsal edge. The width of the dorsal depression is about one third of the entire width of the scleritome. Density of rugae is ca. 9–11 per mm with about 40 rugae per sclerite. Estimated length of the animal is around 20 mm.
Description.—The holotype and only specimen PIMUZ 32127 is a slightly disarticulated specimen with remains of over 20 sclerites. The sclerite-bearing patch is about 15 mm long, about 10 mm wide, and about 3 mm thick. The original skeletal material has been dissolved diagenetically and partially, the voids had been filled by limonite (likely primarily pyrite) and partially, the voids are empty, thus providing a reasonable density contrast for CT-scanning. The number of sclerites preserved indicates that each side had more than 10 sclerites per side.
In terms of number of sclerites per side, we presume that not many are missing due to the fact that so many sclerites are still in place and there are no signs for scattering due to currents. Additionally, no sclerites were seen around that patch. We thus suggest that this species had probably 22 sclerites.
The sclerites range between 2.5 and 2.9 mm in length (anterior-posterior), between 2.1 and 3.2 mm in height (dorsoventral) and between 1.0 and 1.5 mm in width (lateral). The depression in the dorsal hinge is 0.5 to 0.6 mm wide and 0.2 to 0.4 mm deep. The outline of the sclerites in lateral view ranges from subtrapezoidal (sclerite 7) via subtriangular (sclerite 6) to subrectangular (sclerite 1), but this might partially be due to incompleteness. The dorsal margins are quite strongly vaulted with the midpoint being 0.2 mm higher than the anterior and posterior edges in sclerite 2 (at a sclerite length of 2.7 mm). The anterior and posterior edges are slightly concave, with the edges being more strongly bent dorsally. This is linked with the presence of anterior and posterior processes at the margin of the dorsal depression.
The 3D-prints allowed the reconstruction of the dorsal depression, which has a width ratio of ca. 0.3 of depression width to articulated scleritome width when closed. The lateral edge of the dorsal depression is gently sinusoidally bent and visible in dorsal view (Fig. 4). Heeding this viewing direction, the overlapping part of the sclerite becomes wider anteriorly. The concave part of the dorsal depression of the sclerites is subtriangular to subtrapezoidal and tapering posteriorly (Fig. 4).
The rugae are discernible both optically and in the reconstructions based on the CT-image stack. At the ventral margin, sclerites 1 and 5 bear about 10 horizontal rugae on 1 mm, so the spacing appears to more or less uniform in the sclerites of both sides.
The exact position of the single sclerites is difficult to determine; none of the better preserved ones can be assigned unequivocally as being a terminal sclerite of either end. Nevertheless, the more subrectangular sclerites such as sclerites 1 and 2 likely belong rather to the middle of the tube while strongly tapering morphologies with pronounced differences in length between the anterior and posterior edges probably derive from one of the ends of the tube. Consequently, sclerite 5 was presumably positioned close to the posterior end of the tube. Due to its low overall height, we suggest that sclerite 7 might derive from near the anterior end.
Remarks.—Most members of the genus are clearly older, i.e., Silurian or Ordovician. Important characters apparently are the number of sclerites, the outline of the sclerites and the dimension of the dorsal furrow. Rugae spacing appears to be at least partially controlled by ecological factors as it is often quite irregular. The outline varies slightly through ontogeny as inferred from the course of rugae.
As discussed above, the number of sclerites of the new species might be as low as in Lepidocoleus britannicus, L. hohensteini, L. jamesi, and L. sarlei (see also Fig. 9). Its sclerite outline differs from most other known species in the elongate, often subtrapezoidal shape with anterior and posterior processes. Among all species, L. sarlei has the most similar sclerites, but even here, differences are evident such as the stronger dorsal arching and the longer anterior and posterior processes in the new species. As far as the dorsal furrow is concerned, it is again L. sarlei that is morphologically the closest: Both L. sarlei and L. kuangguoduni sp. nov. have a broad dorsal depression (width ratio of ca. 0.35 in L. sarlei and ca. 0.3 L. kuangguoduni sp. nov) with alternating left and right sclerites.
Stratigraphic and geographic range.—Type locality and horizon only.
Discussion
Taphonomy.—The plates are quite similar in form and size. The two most complete reconstructable plates (sclerites 1 and 2) fit perfectly in one other and are positioned near each other in the sample, which corroborates the hypothesis that all plates belong to one individual. Still, the slightly disarticulated arrangement of the sclerites might speak against the preservation in infaunal life position. The question arises how the plates became rearranged and still stayed in close association. Possible explanations are bioturbation, scavenging, mild currents or in situ collapse with the disintegration of the soft parts.
The hypothesis of currents causing the disarticulation appears unlikely because the thin sclerites would have been distributed more widely under the influence of even slight currents. Additionally, the random arrangement and lack of telescoping of the dacryoconarids speaks against current alignment (Hladil et al. 1991, 2014; Stilkerich et al. 2016), though there appears to be a slight trend in the positioning of the apices on some angles—potentially related with uneven embedding of dacryoconarids around the machaeridian—but not enough to warrant a great influence of currents. This is further corroborated by the lithology of the sample—the claystone suggests a weak current regime.
If bioturbation was responsible for the rearrangement of the plates, one would expect the dacryoconarids to be arranged in more random angles in the vertical plane, although this also depends on the size of the infaunal organisms. Another explanation is that the carcass was lying on the seafloor and was partially exposed, allowing its slight disarticulation due to the decay of its soft-parts, but this might also have occurred within the sediment. No obvious damage to plates, which could be completely reconstructed was found, so that it appears less likely that the plates are part of a regurgitate or coprolite. It is well conceivable that the decay of the soft body, maybe in combination with a mild bioturbation, caused the collapse of the skeleton and the slight shift of the sclerites (potentially augmented by post-depositional compaction).
Conclusions
The shape and number of sclerites indicate that our specimen is a derived lepidocoleid. To our knowledge, it is the first machaeridian described from the Middle Devonian of China. Thus far, machaeridians have been described only from the Ordovician of China (Wu 1990). Due to the morphology of its sclerites, we introduce the new taxon Lepidocoleus kuangguoduni sp. nov. In its overall morphology, it was probably quite similar to the Siluran form L. sarlei.
The sizes and fitting of some sclerites, their morphological similarity and also the positioning of the plates suggest that all sclerites of this cluster belong to a single individual. There is no support for currents or bioturbation being responsible of the rearrangement of the sclerites found in this specimen. Thus, it appears more likely that the specimen was at least partially exposed and disarticulated due to the decay of its soft parts and potentially scavenging rather than the animal ending up on the sediment surface and the sclerites becoming dissociated by a predator or scavenger. The sclerites are embedded in the sediment and are not damaged. By contrast, if the sclerites had travelled through the digestive tract of a predator, it appears likely that the thin carbonatic plates became dissolved or at least visibly corroded. Thus, the good preservation of the sclerites speaks for either an in situ infaunal preservation or rapid burial. The latter is less likely in this clay-dominated pelagic sedimentary system, although sediment thickness appears to be moderately high in that basin (see Hunsrück Slate models for rapid burial by fine-grained turbidites, Sutcliffe et al. 1999; Sutcliffe et al. 2002).
Acknowledgements
We would have been completely lost in the field without our experienced field guide Kuang Guodun (Nanning, China). The Swiss Federal Laboratories for Materials Science & Technology (EMPA) is acknowledged for providing the micro-CT data. George Kähler (Siemens, Erlangen, Germany) kindly produced the 3D-prints. We would like to thank Luke Parry (University of Bristol, UK) and an anynomous reviewer for commenting on a previous version of this manuscript. CK and KDB greatly appreciate the financial support by the Swiss National Science foundation (project numbers 200020_132870, 200020_149120, 2011–2014); without this funding, our fieldwork in Guangxi would have been impossible.
References
Adrain, J.M. 1992. Machaeridian classification. Alcheringa 16: 15–32. Crossref
Adrain, J.M., Chatterton, B.D.E., and Cocks, L.R.M. 1991. A new species of machaeridian from the Silurian of Podolia, USSR, with a review of the Turrilepadidae. Palaeontology 34: 637–651.
Allmendinger, R.W., Cardozo, N., and Fisher, D.M. 2011. Structural Geology Algorithms: Vectors and Tensors. 302 pp. Cambridge University Press, Cambridge. Crossref
Barrande, J. 1872. Systême Silurien du centre de la Bohême: Ière Partie: Recherches Paléontologiques. Supplément au Vol. I. Trilobites, Crustacés divers et Poissons. xxx + 647 pp. Chez l’auteur et éditeur, Prague.
Bengtson, S. 1978. The Machaeridia—a square peg in a pentagonal hole. Thalassia Jugoslavica 12: 1–10.
Bengtson, S. 1979. Machaeridians. Sveriges Geologiska Undersökning C 762: 211–212.
Cardozo, N. and Allmendinger, R.W. 2013. Spherical projections with OSXStereonet. Computers & Geosciences 51: 193–205. Crossref
Caron, J.-B. 2008. Palaeontology: Ancient worms in armour. Nature 451: 133–134. Crossref
Clarke, J. 1896. The structure of certain Palaeozoic barnacles. American Geologist 17: 137–143.
De Baets, K., Klug, C., and Plusquellec, Y. 2010. Zlichovian faunas with early ammonoids from Morocco and their use for the correlation of the eastern Anti-Atlas and the western Dra Valley. Bulletin of Geosciences 85: 317–352.
Dzik, J. 1986. Turrilepadida and other Machaeridia. Problematic Fossil Taxa 5: 116–134.
Dzik, J. 1994. Machaeridians, chitons, and conchiferan molluscs of the Mójcza Limestone. Palaeontologia Polonica 53: 213–252.
Ekleris, A. and Radzevičius, S. 2014. The Upper Homerian (Silurian) machaerid sclerite from Lithuania. Geologija 55: 109–114. Crossref
Faber, C.L. 1886. Remarks on some fossils of the Cincinnati Group. Journal of the Cincinnati Society for Natural History 9: 14–20.
Hall, J. and Whitfield, R. 1875. Descriptions of invertebrate fossils, mainly from the Silurian System. Report of the Geological Survey of Ohio, Palaeontology 2: 65–161.
Hints, O., Eriksson, M., Högström, A.E., Kraft, P., and Lehnert, O. 2004. Worms, wormlike and sclerite bearing taxa. In: B. Webby, M.L. Droser, F. Paris, and I. Percival (eds.), The Great Ordovician Biodiversification Event, 223–230. Columbia University Press, New York. Crossref
Hladil, J., Čejchan, P., and Beroušek, P. 1991. Orientation of the conical tests of tentaculites: internal waves in aqueous environment. Časopis pro mineralogii a geologii 36: 115–130.
Hladil, J., Simcik, M., Ruzicka, M.C., Kulaviak, L., and Lisy, P. 2014. Hydrodynamic experiments on dacryoconarid shell telescoping. Lethaia 47: 376–396. Crossref
Högström, A.E.S. 1997. Machaeridians from the Upper Wenlock (Silurian) of Gotland. Palaeontology 40: 817–831.
Högström, A.E.S. and Taylor, W.L. 2001. The Machaeridian Lepidocoleus Sarlei Clarke, from the Rochester Shale (Silurian) of New York state. Palaeontology 44: 113–130. Crossref
Högström, A.E.S., Briggs, D.E.G., and Bartels, C. 2009. A pyritized lepidocoleid machaeridian (Annelida) from the Lower Devonian Hunsruck Slate, Germany. Proceedings of the Royal Society B: Biological Sciences 276: 1981–1986. Crossref
Jell, P. 1979. Plumulites and the machaeridian problem. Alcheringa 3: 253–259. Crossref
Klug, C., Kröger, B., Korn, D., Rücklin, M., Schemm-Gregory, M., De Baets, K., and Mapes, R.H. 2008. Ecological change during the early Emsian (Devonian) in the Tafilalt (Morocco), the origin of the Ammonoidea, and the first African pyrgocystid edrioasteroids, machaerids and phyllocarids. Palaeontographica A 283: 83–176. Crossref
Lamarck, J.B. de 1809. Philosophie Zoologique. 475 pp. Dentu et L’Auteur, Paris.
Moberg, J.C. 1914. Om svenska Silurcirripedier. Lunds Universitets Arsskrift, NY Följd, Adfelning 2 11: 1–20.
Parry, L.A., Edgecombe, G.D., Eibye-Jacobsen, D., and Vinther, J. 2016. The impact of fossil data on annelid phylogeny inferred from discrete morphological characters. Proceedings of the Royal Society B: Biological Sciences 283: 20161837. Crossref
Parry, L., Tanner, A., and Vinther, J. 2014. The origin of annelids. Palaeontology 57: 1091–1103. Crossref
Reed, F.C. 1901. III.—Woodwardian Museum notes: Salter’s undescribed species. III. Geological Magazine (Decade IV) 8 (03): 106–110. Crossref
Ruedemann, R. 1925. Some Silurian (Ontarian) faunas of New York. New York State Museum Bulletin 265: 1–134.
Savage, T.E. 1913. Some interesting new species of arthropods from Devonian strata of Illinois. American Journal of Science 35: 149–152. Crossref
Seilacher, A. and Gishlick, A.D. 2014. Morphodynamics. 551 pp. CRC Press, Boca Raton.
Sieverts, H. 1935. Das erste machaerid aus Deutschland: Lepidocoleus eifeliensis n. sp. aus dem Rheinischen Mitteldevon. Zeitschrift der Deutschen Geologischen Gesellschaft 87: 683–687.
Sieverts-Doreck, H. 1952. Weitere Machaeridier-Funde aus Deutschland. Decheniana 105/106: 57–62.
Stilkerich, J., Smrecak, T.A., and De Baets, K. 2016. 3D-Analysis of a non-planispiral ammonoid from the Hunsrück Slate: natural or pathological variation? PeerJ Preprints 4: e2479v1.
Sutcliffe, O.E., Briggs, D.E.G., and Bartels, C. 1999. Ichnological evidence for the environmental setting of the Fossil-Lagerstatten in the Devonian Hunsruck Slate, Germany. Geology 27: 275–278. Crossref
Sutcliffe, O.E., Tibbs, S., and Briggs, D. 2002. Sedimentology and environmental interpretation of the fine-grained turbidites in the Kaub Formation of the Hunsrück Slate: analysis of a section excavated for Project Nahecaris. Metalla (Bochum) 9: 89–104.
Sutton, M.D., Garwood, R.J., Siveter, D.J., and Siveter, D.J. 2012. SPIERS and VAXML; a software toolkit for tomographic visualisation and a format for virtual specimen interchange. Palaeontologia Electronica 15 (2): 1–14.
Sutton, M., Rahman, I.A., and Garwood, R.J. 2014. Techniques for Virtual Palaeontology. 208 pp. Wiley-Blackwell, Hoboken.
Vinther, J. and Briggs, D.E.G. 2009. Machaeridian locomotion. Lethaia 42: 357–364. Crossref
Vinther, J. and Rudkin, D. 2010. The first articulated specimen of Plumulites canadensis (Woodward, 1889) from the Upper Ordovician of Ontario, with a review of the anterior region of Plumulitidae (Annelida: Machaeridia). Palaeontology 53: 327–334. Crossref
Vinther, J., Van Roy, P., and Briggs, D.E.G. 2008. Machaeridians are Palaeozoic armoured annelids. Nature 451: 185–188. Crossref
Withers, T.H. 1922. The “Cirripede” Lepidocoleus in the upper Ordovician rocks of Scotland. Journal of Natural History 9: 653–656. Crossref
Withers, T.H. 1926. Catalogue of the Machaeridia (Turrilepas and Its Allies) in the Department of Geology. 99 pp. British Museum, London.
Wolburg, J. 1938. Beitrag zum problem der Machaeridia. Paläontologische Zeitschrift 20: 289–298. Crossref
Wu, H.-J. 1990. Occurrence of Machaeridia in China. Acta Palaeontologica Sinica 29: 567–575.
Acta Palaeontol. Pol. 62 (2):
237–247, 2017
https://doi.org/10.4202/app.00346.2017