
Osteology of the unenlagiid theropod Neuquenraptor argentinus from the Late Cretaceous of Patagonia
FEDERICO BRISSÓN EGLI, ALEXIS M. ARANCIAGA ROLANDO, FEDERICO L. AGNOLÍN, and FERNANDO E. NOVAS
Brissón Egli, F., Rolando Aranciaga, A.M., Agnolín, F.L., and Novas, F.E. 2017. Osteology of the unenlagiid theropod Neuquenraptor argentinus from the Late Cretaceous of Patagonia. Acta Palaeontologica Polonica 62 (3): 549–562.
Neuquenraptor argentinus was described as the first undoubted deinonychosaurian theropod from Gondwana. The only known specimen is represented by a fragmentary skeleton, including a nearly complete foot, coming from Late Cretaceous beds of Neuquén Province, Patagonia, Argentina. Neuquenraptor was later considered as belonging to the Southern Hemisphere paravian clade Unenlagiidae, and proposed as a junior synonym of Unenlagia comahuensis. The aim of the present paper is to include a detailed anatomical description of Neuquenraptor (up to the date only known by a preliminary description). Comparisons with other paravians, especially unenlagiids, resulted in the finding of characters that may be considered as diagnostic of Unenlagiidae (e.g., scar-like fourth trochanter of femur, metatarsal II with tongue-shaped process over the caudal surface of metatarsal III, well-developed convex longitudinal plantar crest of metatarsal IV). Furthermore, comparisons between Neuquenraptor and Unenlagia are carried out with the aim to evaluate the possible synonymy between these taxa. These comparisons indicate that at least two similar-sized unenlagiids were present on the Portezuelo Formation. However, in concordance with previous authors, we are not able to support nor dismiss the possible synonymy between Unenlagia and Neuquenraptor. Additional findings of unenlagiid fossils from the Portezulo Formation will help to clarify the taxonomic status of these taxa.
Key words: Dinosauria, Theropoda, Unenlagiidae, Neuquenraptor, Cretaceous, Argentina, Patagonia.
Federico Brissón Egli [fedebe@gmail.com], Alexis M. Aranciaga Rolando [mauro.a_guido@hotmail.com], and Fernando E. Novas [fernovas@yahoo.com.ar], CONICET and Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Av. Ángel Gallardo 470 (C1405DJR), Buenos Aires, Argentina.
Federico L. Agnolín [fedeAgnolín@yahoo.com.ar], CONICET, Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Av. Ángel Gallardo 470 (C1405DJR), Buenos Aires, Argentina and Fundación de Historia Natural “Félix de Azara”, Universidad Maimónides, Hidalgo 775 (C1405BDB), Buenos Aires, Argentina.
Received 2 February 2017, accepted 10 May 2017, available online 4 July 2017.
Copyright © 2017 F. Brissón Egli et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Novas and Pol (2005) described the first undoubted “deinonychosaur” from the Late Cretaceous of South America, on the basis of incomplete hindlimb material. They named this new taxon Neuquenraptor argentinus and included it within Dromaeosauridae. Later, Makovicky et al. (2005) recovered Neuquenraptor within the Gondwanan clade Unenlagiinae (Unenlagiidae herein) and suggested that it might be congeneric with Unenlagia, a criterion followed by other authors (Turner et al. 2007; Agnolín and Novas 2011, 2013). Previously, Novas (2009) indicated that based on the non-overlapping material Neuquenraptor may be retained as a valid taxon, and that new evidence is necessary in order to sustain the synonymy of both taxa. This criterion was endorsed by Porfiri et al. (2011), Gianechini and Apesteguía (2011), and Turner et al. (2012) who provisionaly considered Neuquenraptor as a valid taxon.
Neuquenraptor was briefly commented and figured by Novas and Pol (2005), Novas (2009), Gianechini and Apesteguía (2011) and Turner et al. (2012). The aim of the present contribution is to make a detailed and complete description and comparisons of available material of Neuquenraptor argentinus Novas and Pol, 2005.
Institutional abbreviations.—MACN, Museo Argentino de Ciencias Naturales, Bernardino Rivadavia, Ciudad de Buenos Aires, Argentina; MCF-PV, Museo Carmen Funes, Plaza Huincul, Neuquén, Argentina; MPCA, Museo Provincial Carlos Ameghino, Cipolletti, Río Negro, Argentina; MPCN, Museo Provincial de Ciencias Naturales, General Roca, Río Negro, Argentina.
Other abbreviations.—m., muscle.
Material and methods
The holotype specimen of Neuquenraptor argentinus (MCF PVPH 77) was found in Sierra del Portezuelo (Fig. 1), Neuquén Province, Argentina (Novas and Pol 2005) during the excavation of a partially articulated and associated titanosaur, lying within the rib cage (dorsal ribs and sternal plates) of this sauropod. The geological unit bearing these fossils, i.e., lower levels of the Portezuelo Formation (Coniacian; Novas and Pol 2005), is composed of continental sediments with frequent paleosol tops (Leanza et al. 2004). In the same area and coeval levels, abundant fishes, turtles, crocodyliforms, and dinosaurs including indeterminate titanosaurs, ornithopods, and theropods represented by Unenlagia comahuensis, Megaraptor namunhuaiquii, Patagonykus puertai, and a neornithine bird were found (Novas 1997, 1998; Novas and Puerta 1997; Agnolín et al. 2006).
We follow Hutchinson (2001) and Carrano and Hutchinson (2002) regarding the nomenclature of hindlimb muscles and tendons. Additionally, we follow Baumel and Witmer (1993) for bone orientation. In this way along the text the terms cranial and caudal are used instead of anterior and posterior, respectively. In the case of pedal phalanges the terms cranial and caudal are replaced by dorsal and ventral, respectively.
There is consensus that Buitreraptor, Unenlagia, Neuquenraptor, and Austroraptor are a monophyletic clade of paravian theropods (Makovicky et al. 2005; Novas et al. 2009), originally termed as Unenlagiidae by Bonaparte (1999). We follow the phylogenetic nomenclature by Agnolín and Novas (2013), considering unenlagiids as a family rank clade, in contrast with Makovicky et al. (2005) and Turner et al. (2012) who interpreted Unenlagiinae as a subfamily group within Dromaeosauridae.
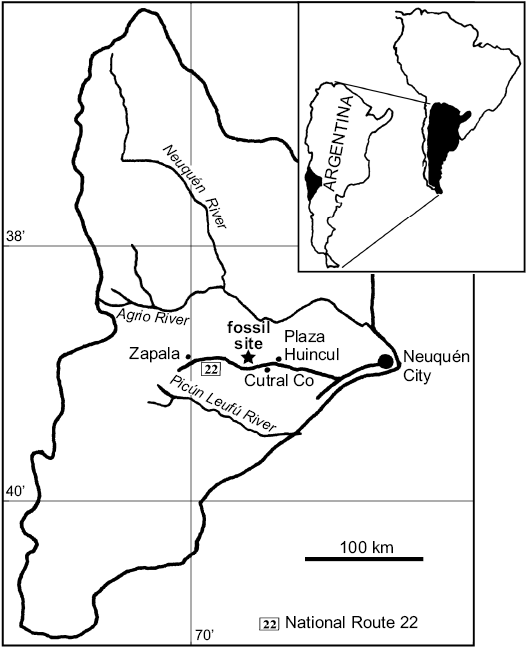
Fig. 1. Map of Neuquén Province (Argentina) showing the Sierra del Portezuelo fossil site, where the holotype of Neuquenraptor argentinus was found.
Systematic palaeontology
Theropoda Marsh, 1881
Tetanurae Gauthier, 1986
Coelurosauria Huene, 1920
Paraves Sereno, 1997
Unenlagiidae Bonaparte, 1999
Neuquenraptor argentinus Novas and Pol, 2005
Holotype: MCF PVPH 77, fragments of dorsal ribs, two proximal haemal arches, left proximal radius, incomplete right femur, distal end of right tibia, proximal tarsals, and most of the left foot.
Type locality: Sierra del Portezuelo (Fig. 1), Neuquén Province, Argentina.
Type horizon: Portezuelo Formation, Coniacian, Late Cretaceous.
Emended diagnosis.—Mid-sized unenlagiid theropod (Table 1) with the following combination of characters (autapomorphies marked with an asterisk): deep and wide extensor sulcus on proximal half of metatarsus; metatarsal III proximally pinched; metatarsal II with lateral expansion over the caudal surface of metatarsal III*; distal third of the medial surface of the metatarsal II is obliquely oriented and is delimited from the caudal surface of the metatarsal shaft by the subtle distal end of the plantar crest* distal third of metatarsal IV transversely wide*; pedal digit II with phalanges 1 and 2 sub-equal in length* and well-developed medial bump on pedal phalanx IV-3*.
Table 1. Selected measurements (in mm) of Neuquenraptor argentinus (MCF-PVPH-77). Parentheses indicate preserved measurements.
| |
Radius left |
Femur right |
Tibia |
Astragalo- calcaneum left |
Metatarsal |
Digit I |
Digit II |
Digit III |
Digit IV |
||||||||||||
|
I |
II |
III |
IV |
1 |
2 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
5 |
||||||
|
Length |
(73) |
(232) |
|
|
(27) |
(140) |
(171) |
(146) |
25 |
(21) |
36 |
35 |
55 |
55 |
34 |
33 |
30 |
23 |
22 |
(26) |
|
|
Proximal end |
width |
15 |
22 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
height |
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mid-length |
width |
|
24 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
depth |
|
|
|
|
|
12 |
2 |
18 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Distal end |
width |
|
|
41 |
17 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
antero- posterior length |
|
|
16 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Height |
|
|
|
(22) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Description.—Haemal arches: Two proximal haemal arches have been preserved (Fig. 2). Based on comparisons with Velociraptor and Linheraptor they likely represent two of the three cranialmost haemal arches, because they lack a distal expansion (Norell and Makovicky 1997). The chevrons are distally curved, a feature more accentuated in the caudalmost preserved element. The cranial margin is convex in lateral view and the caudal one is concave, being deeper in the caudalmost haemal arch. The distal end of the blade is broken off in the cranial-most haemal arch, but appears to have been distally tapering. The distal end of the caudal element tapers to an acute margin. A cranial expansion is only present in the cranialmost haemal arch. It is formed by a well-developed hook-like process located at the dorsal half of the cranial surface of the bone. The caudal expansion seems to be absent.
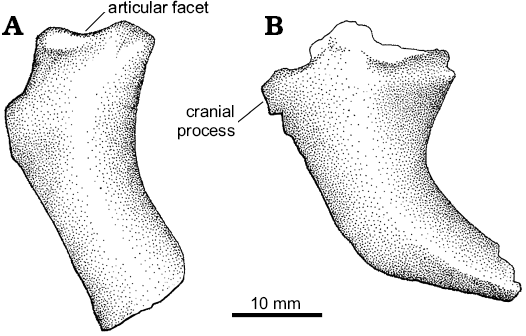
Fig. 2. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; two isolated proximal haemal arches in left lateral view.
Radius: A partial left radius was preserved (Fig. 3) lacking most of its distal half. It is long, gracile, and nearly straight. The proximal end shows a subtriangular-shaped articular surface. It is cup-shaped for articulation with the humeral ectepicondyle. The caudolateral and craniomedial margins of the proximal end of the radius are slightly upturned. The brachialis tubercle is represented by a poorly convex cranial surface of the shaft. The proximal end of the lateral surface of the bone shows a flattened caudal half that is decorated by ridges, indicating strong tendinal or muscular attachments, probably for the m. supinator (Remes 2007). The cranial half of the shaft is rounded, and lacks a surface for the anchoring of the m. adductor radialis, a condition shared with birds (Remes 2007). In medial view, the proximal end of the bone shows two distinct areas, the cranial one is relatively smaller and is craniolaterally oriented, whereas the caudal one is medially oriented. The cranial surface attaches the m. pronator teres and the caudal surface the m. pronator quadratus (Remes 2007).
A well-developed intermuscular line (ril2 in Langer et al. 2007) runs from the caudolateral surface of the proximal shaft to the caudomedial margin of mid-shaft and forms a rugose crest near its distal end. The shaft is lateromedially flattened distally, as in other paravians (e.g., Velociraptor, Deinonychus; Ostrom 1969; Norell and Makovicky 1999).
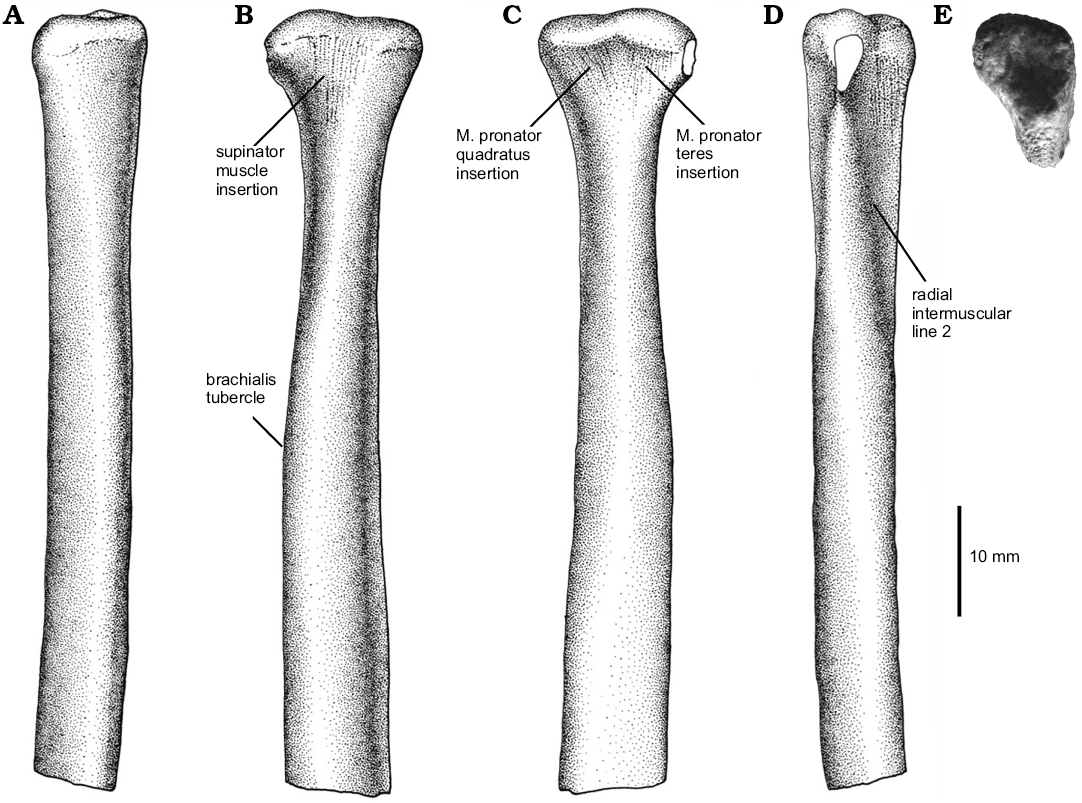
Fig. 3. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; partial left radius in cranial (A), lateral (B), medial (C), caudal (D), and proximal (E) views. A–D, drawings; E, photograph.
Femur: The right femur (Fig. 4) is represented by a highly damaged and distorted portion of the shaft, lacking both proximal and distal ends. As it is broken, the shaft appears to be robust, resembling Laurasian dromaeosaurids. However, comparisons with Unenlagia and Buitreraptor reveal that the shaft was long and slender when complete. The proximal end is damaged and the articular head is missing. In spite to that, the preserved portion of the femoral neck and the base of the proximal end of the bone indicate that the femoral head was probably caudomedially oriented, as occurs in Buitreraptor and Unenlagia. Only the base of the lesser trochanter is discernible on the lateral surface. Due to the lack of preservation, it is not possible to assure the presence or absence of a cleft separating the lesser and greater trochanters. Nevertheless, its occurrence in Unenlagia comahuensis (Novas and Puerta 1997) indicates that its presence is probable in Neuquenraptor.
In contrast to Buitreraptor, but similar to Unenlagia and other paravians, an obturator ridge (sensu Baumel and Witmer 1993) is absent. The fourth trochanter is highly reduced and represented by a subtle, scar-like groove that is bounded by two low oblique ridges. This condition is also verified in Unenlagia, Buitreraptor, and Austroraptor. The fourth trochanter is located in an unusual proximal position on the caudal surface of the bone. Distal to this trochanter, the shaft forms a flattened proximodistally extended surface that is delimited by a well-developed longitudinal lateral ridge, as also occurs in Velociraptor (Norell and Makovicky 1999). This ridge constitutes the caudal (= posterior) intermuscular line of Hutchinson (2001), that delimits the boundary between the m. femorotibialis externus (craniolaterally), and the m. adductor femoris (caudomedially).
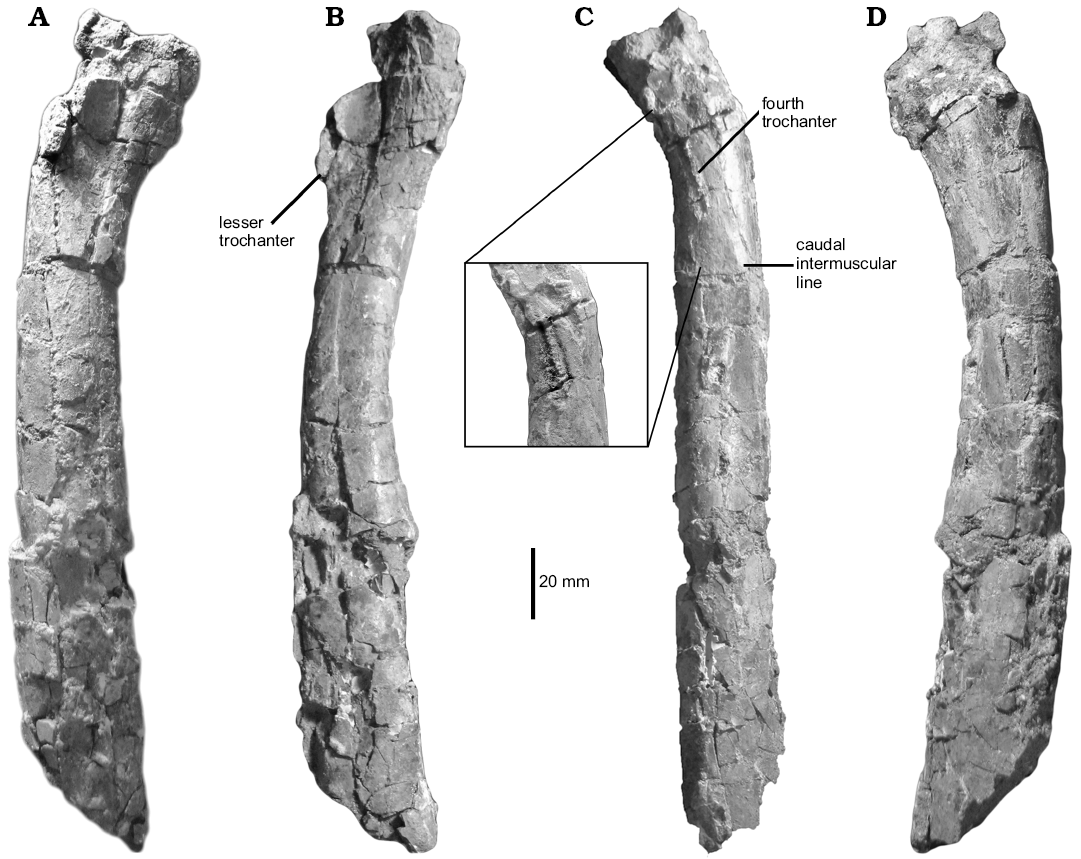
Fig. 4. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; right femur in cranial (A), medial (B), caudal (C), and lateral (D) views. The inset in C is an enhanced image displaying the fourth trochanter morphology.
Tibia and fibula: The left tibia is represented by its distal end (Fig. 5). A fragment of distal fibular shaft is still adhered on the craniolateral margin of the tibia. The latter bone exhibits a craniocaudally compressed and transversely expanded shaft. The distal end is transversely expanded. The outer malleolus is more distally projected than the medial. The outer malleolus is subrectangular in contour and is laterally delimited by a subtle proximodistally oriented rugose ridge. The medial malleolus bears a sharply marked medial margin.
The cranial surface of distal end of the tibia is nearly flat. The facet for the ascending astragalar process is extensive, with the medial surface obliquely oriented. Along the central portion of this surface there is a low proximodistally oriented mound-like ridge that separates the articular surface for the ascending astragalar process into two slightly concave areas.
The caudal surface of the distal tibia is transversely concave, forming an incipient trochlea cartilaginis tibialis (sensu Baumel and Witmer 1993). In caudal view, there are two distinct flattened surfaces separated by a strong sub-vertical ridge. The lateral surface is transversely wide and slightly caudolaterally oriented, whereas the medial one is notably narrow and caudomedially faced.
In distal view, the tibia is saddle-shaped in contour with the medial portion thicker than the lateral one. Furthermore, the cranial margin of the bone is gently convex, whereas the caudal one is concave.
The fibular shaft is splint-like, representing approximately 12% of the tibial transverse diameter. It likely contacted the proximal end of the calcaneum, as in other basal paravians.
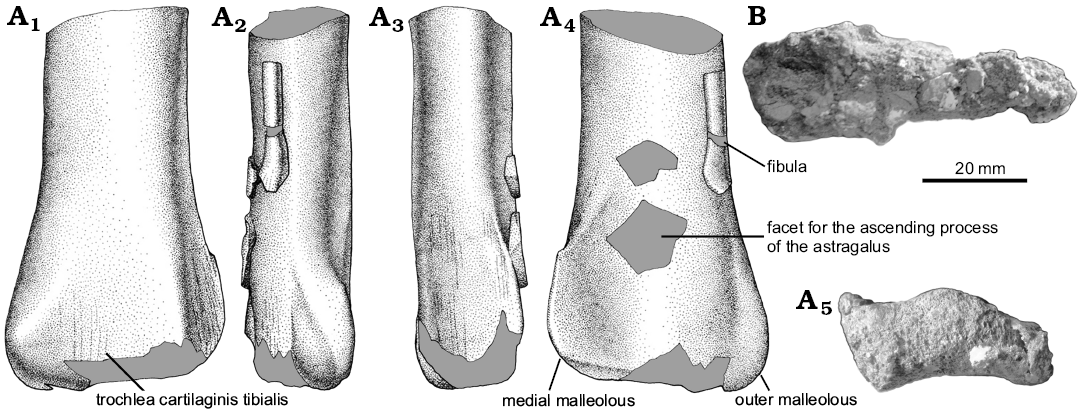
Fig. 5. A. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; distal end of the left tibia and fibula in caudal (A1), lateral (A2), medial (A3), cranial (A4), and distal (A5) views. B. Unenlagiid theropod Unenlagia comahuensis Novas and Puerta, 1997 (MCF PVPH 78) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; tibia in distal view. A1–A4, drawings; A5, B photographs.
Astragalocalcaneum: The astragalus and calcaneum (Fig. 6) are incomplete, but are anatomically informative. The astragalar ascending process is incompletely preserved, but fragments of it are attached to the cranial surface of distal tibia. The large and triangular facet of tibia accommodated the wide, laminar and proximodistally high ascending process of the astragalus. The base of the ascending process shows that the medial margin of the ascending process was obliquely oriented, whereas the lateral one is almost straight and vertical. At the base of the ascending process, there is a poorly defined and shallow subcircular fossa. Its base is separated from the astragalar body through a transversally oriented groove (Welles and Long 1974) and delimits the “cranial platform of the ascending process” (Langer and Benton 2006: 344). This groove probably served for anchoring of the m. tibialis cranialis (Carrano and Hutchinson 2002). The main astragalar body is incomplete, lacking its lateral portion, and the calcaneum is slightly dorsally displaced. Nevertheless, the astragalar body seems to have been sub-cylindrical in contour. The astragalus is constricted at mid-length. The asymmetry of the condyles in distal view cannot be confirmed due to the incomplete nature of the lateral portion of the astragalar body. In medial view, the astragalus is ellipsoidal, whereas in Deinonychus (Ostrom 1969) and Troodon (Russell 1969) it is sub-circular in shape. The medial surface has two shallow concavities, the ventral one being larger than the dorsal one. These concavities probably represent the “depressio epicondylaris medialis” (Baumel and Witmer 1993: 111). In Neuquenraptor the caudal half of the astragalar body is proximodistally compressed and craniocaudally shortened, so that it does not “wrap” around the tibia.
The calcaneum is poorly preserved and most anatomical details are not available. Only the lateral and dorsal portions of the bone are preserved. It is transversely compressed and it is not fused to the astragalus. The medial surface is nearly flat and devoid of a well-defined concavity. The cranialmost margin of the calcaneum in proximal and distal views is slightly laterally tilted.
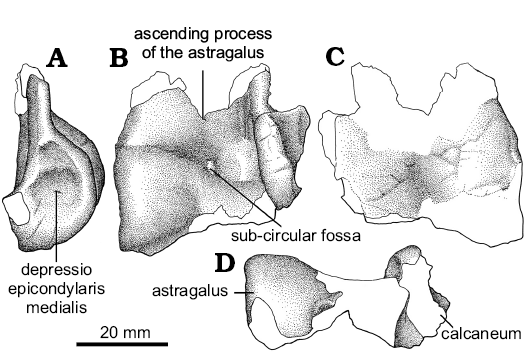
Fig. 6. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; partial left astragalocalcaneum in lateral (A), cranial (B), caudal (C), and distal (D) views.
Pes: An almost complete left pes is preserved. It is represented by metatarsals I, II, III, and IV, and most of their pedal phalanges. Metatarsals II, III, and IV lack their proximal part and the distal end of metatarsal IV is missing.
The metatarsus (Fig. 7) is long and slender. Metatarsals II–IV are tightly apressed each other. As is typical of most theropods, metatarsal III is the longest of the pes, with metatarsal IV and II being sub-equal in length. Metatarsal III is partially covered proximally by metatarsals II and IV, but still being exposed along most of the metapodium in cranial and caudal views, constituting a subarctometatarsalian pes (Xu and Wang 2000; Snively et al. 2004; White 2009 ).
Metatarsal I (Fig. 7E–H) lacks its proximal end. When found, metatarsal I was lying on the caudal surface of metatarsal II. The cranial surface of the metatarsal I was caudomedially oriented. The medial pit faces caudally. In spite to that, it is uncertain if the metatarsal I occupied that position when the animal was alive. The absence on metatarsal II of a clear facet for articulation with metatarsal I results in that the orientation of the latter is difficult to assess.
Metatarsal I lacks medial or lateral torsion. The shaft is proximodistally short, representing less than 16% of metatarsal II length, and tapers proximally. In cross-section the shaft is strongly craniocaudally compressed, having a transversely expanded shaft. The distal trochlea is asymmetrical and medially twisted with respect to the main shaft. Its external condyle is large and it has a transversely expanded trochlear ring. It is more distally projected than the medial condyle. The lateral ligament pit is wide and deep, which shows a bump on its proximal margin. The medial trochlear ring is barely developed, being proximodistally short. It lacks a well-developed collateral extensor pit.
Both hemicondyles are slightly medially oriented, and in cranial view they are in contact each other, lacking a deep intercondylar groove and thus resulting in a “ball-like” distal end. In caudal view, the intercondylar groove is wide and deep, and it is slightly obliquely oriented.
Metatarsal II (Fig. 7) lacks its proximal end. The shaft of metatarsal II is transversely wider than that of metatarsal IV. It is nearly straight along all of its length, but flares slightly medially on its distal end. In cranial view, the distal portion of metatarsal II is covered by a well-developed medial expansion of metatarsal III. In caudal view, the distal portion of metatarsal II has a lateral expansion over the caudal surface of metatarsal III forming a flat and wide surface. Additionally, metatarsal II shows a moderately developed plantar crest that is relatively low but transversally thick. This low plantar crest gives the metatarsal shaft a subtle sub-triangular cross-section on its proximal portion.
In medial view, the proximal half of metatarsal II is craniocaudally wider than its distal half. On its distal third the medial surface of the bone is obliquely oriented and is delimited from the caudal surface by the subtle distal end of the plantar crest. This condition was not observed in Buitreraptor (MPCN-Pv-598) or Pamparaptor, in which this crest is notably more extended. Metatarsal II ends in a well-developed ginglymoid articulation with a well-developed intercondylar groove that extends along cranial and caudal surfaces. The lateral trochlear ring is incomplete. The medial one is deep and well-developed and bears a strong lateral ligament pit.
The third metatarsal (Fig. 7) lacks its proximal end. Although broken, it can be observed that the proximal portion of the shaft is compressed between metatarsals II and IV. The proximal half of metatarsal III is notably craniocaudally thin. The cranial surface of the shaft shows a long extensor groove, which could represent the area of origin of the m. digitorum brevis (Hutchinson et al. 2005). Near its distal end, the shaft has a medial expansion that partially covers the distal portion of metatarsal II. This expansion is gently concave cranially. In caudal view, the shaft of metatarsal III is ridge-like and is almost completely covered by metatarsals II and IV. It is subtriangular in cross-section.
Proximally, metatarsal III is more caudally located with respect to the shaft of metatarsals II and IV. Towards the distal end, this metatarsal is notably cranially oriented. The distal articulation is moderately ginglymoid, with a weak cranial groove. Both lateral ligament pits are well-developed and sub-equal in size and shape, being deeper than those of metatarsal II.
Metatarsal IV (Fig. 7) has shaft with a proximally convex and a distally slightly concave cranial surface. The lateral surface of the metatarsal is longitudinally concave, and shows a faint longitudinal groove. This surface is deeper at mid-length as a result of the caudal extension of the plantar crest. This crest is notably deeper than that of metatarsal II. In caudal view the shaft transversely broadens towards its distal end. The caudal surface of the shaft is slightly convex and shows a longitudinal plantar crest. It is slightly medially tilted on its caudal end and delimits a concavity on the medial surface of the plantar crest. The distal end of this crest is laterally oriented, and forms the lateral margin of the metatarsal. The distal articulation was not preserved, although its base indicates that it was transversely expanded.
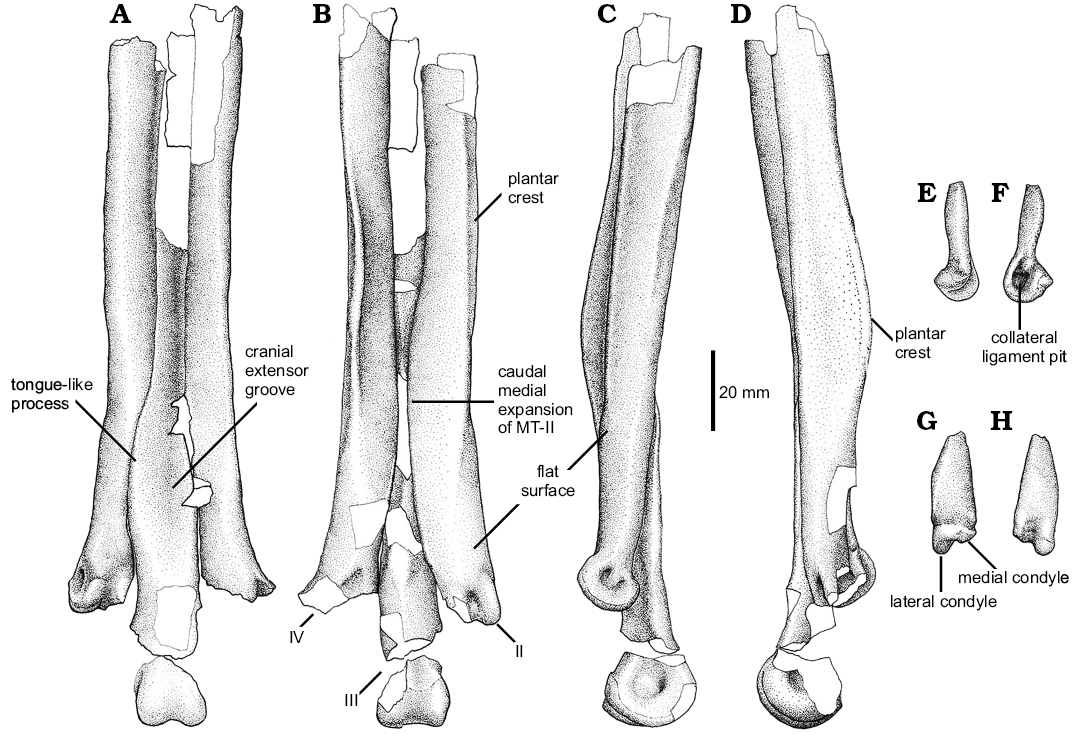
Fig. 7. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous. A–D. Left metatarsals II–IV in cranial (A), caudal (B), medial (C), and lateral (D) views. E–H. Metatarsal I in medial (E), lateral (F), caudal (G), and cranial (H) views.
The first digit (Fig. 8) is represented by phalanges I-1 and I-2. Phalanx I-1 is nearly completely preserved. It is relatively robust and is sub-equal in length to metatarsal I. In lateral view, the phalanx shows a dorsal concave profile. The shaft has a very narrow dorsal surface that is transversely thinner than the ventral surface. The proximal articular facet is relatively large and sub-circular in contour, lacking any sign of a median keel. This morphology is due to its articulation with the “ball-like” distal articular surface of metatarsal I, and probably it allowed a wide range of motion of this digit. The distal end of the phalanx is more transversely compressed than the proximal portion, and it is slightly asymmetrical, since the medial collateral ligament pit is smaller than the lateral one. In ventral view, the proximal end of the bone is notably expanded and shows two shallow concavities for the anchoring of flexor tendons, that are separated by a poorly developed posteroventral process.
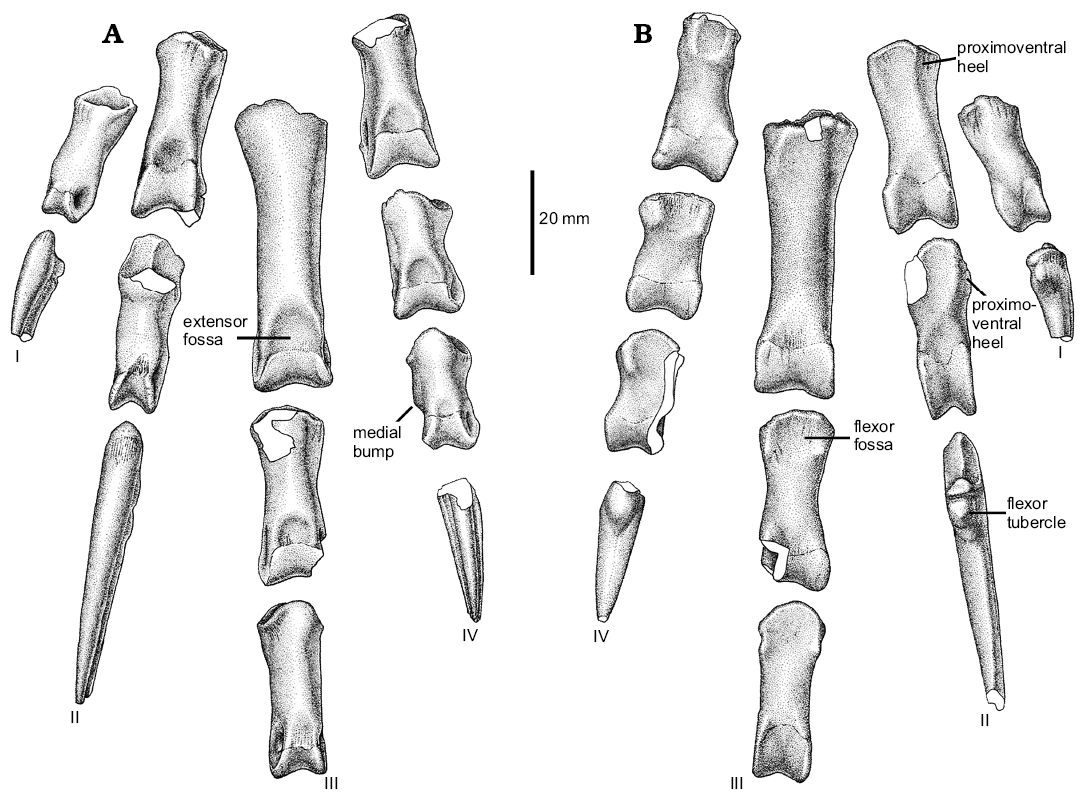
Fig. 8. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; left pedal phalanges in dorsal (A) and ventral (B) views.
The ungual of digit I (Fig. 9A) is relatively short when compared with the other pedal unguals. It is transversely compressed and ventrally curved. The proximal articular facet is dorsoventrally tall and asymmetrical, in order to match the distal articular surface of the preceeding phalanx. A flexor tubercle is present, which is moderately developed and is laterally located on the proximoventral corner of the ungual. The lateral and medial surfaces of the proximal portion of the ungual bears fine longitudinal striations. Single lateral and medial grooves are nearly symmetrical, being the lateral one slightly dorsally displaced.
Pedal digit II is completely preserved (Fig. 9B). Phalanx II-1 is sub-equal in length to phalanx II-2. Phalanx II-1 shows a straight ventral margin, whereas its dorsal margin is notably concave in lateral view. The posteroventral heel of pedal phalanx II-1 is transversally narrow, and medially offset. The proximal articular facet is relatively tall and asymmetrical, with the medial portion dorsoventrally taller and transversely wider than the lateral one. There is a well-developed median keel separating both articular facets. The shaft is dorsoventrally compressed near the distal end, where is formed a well developed neck. The distal end shows a well-developed ginglymoid articulation with both lateral and medial trochlear margins sub-equally extended. The distal end also presents dorsally extended condyles, a feature correlated with the hyperextensible nature of pedal digit II. On the dorsal surface, a well-developed extensor fossa is present, which is nearly confluent with the well-developed cranial intercondylar groove. The lateral collateral ligament pit is well-developed and deep, being located in the middle of the outer trochlear surface, whereas the medial one is slightly shallower, and proximally it shows an additional small pit.
Phalanx II-2 shows the typical basal paravian morphology. The phalanx shaft is dorsoventrally constricted at mid-length, forming a deep neck. The proximoventral heel is well-developed and low. It is medially offset, as in phalanx II-1. The proximal articular facet is asymmetrical and sub-divided by a vertical median keel. The distal end is highly symmetrical. The trochlear rings are large and bear dorsally displaced collateral ligament pits, as usual among paravians. These pits are ellipsoidal in shape, being dorsoventrally low and craniocaudally elongated. The dorsal margins of these pits can be seen in dorsal view, due to a dorsal constriction of the trochlear margins.
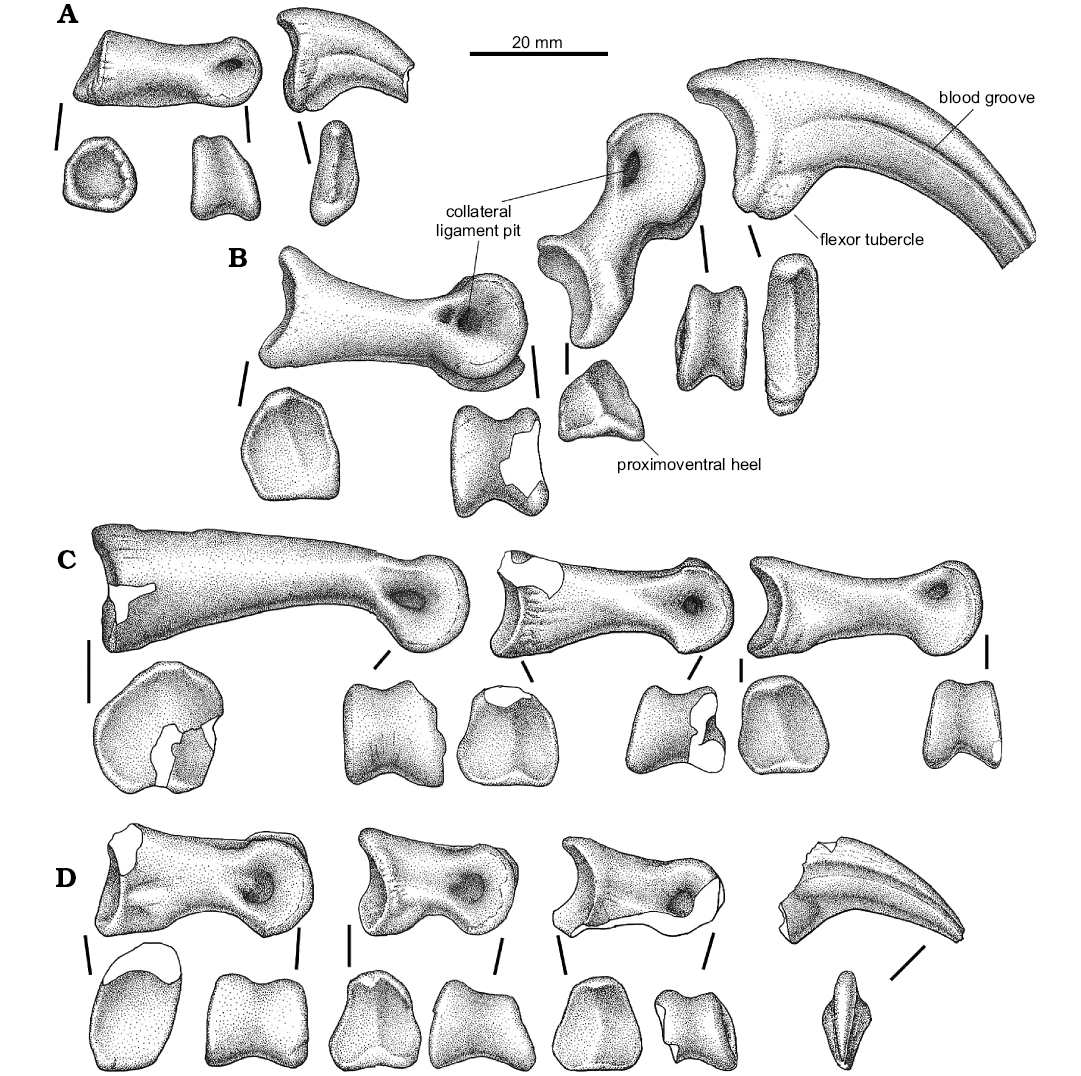
Fig. 9. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; left pedal phalanges: I (A), II (B), III (C), IV (D); in articular and medial views.
The ungual phalanx of digit II (Fig. 10B) is hypertrophied as in other paravians (Gauthier 1986). It is long, ventrally curved, and it is relatively low and narrow with a well-developed flexor tubercle. This tubercle is slightly rugose. Both lateral and medial sides of the claw bear single asymmetrical longitudinal grooves. The lateral one is more dorsally positioned than the medial one. The cutting edge of the ungual is medially displaced, and distally confluent with the medial surface of the claw, resulting in an apparently flat medial surface of the blade.
The first phalanx of digit III (Fig. 10C) is the largest phalanx of the pes. The ventral margin is strongly concave in lateral view, whereas the dorsal margin is gently convex. The proximal articular surface is damaged, but seems to be slightly asymmetrical and without a median keel. The shaft is dorsoventrally compressed just proximal to the distal articulation. The distal end is slightly ginglymoid. The collateral ligament pits are sub-equally developed, being deep and teardrop-shaped. The extensor fossa is crescent-shaped, notably wide, shallow, and with well-defined margins.
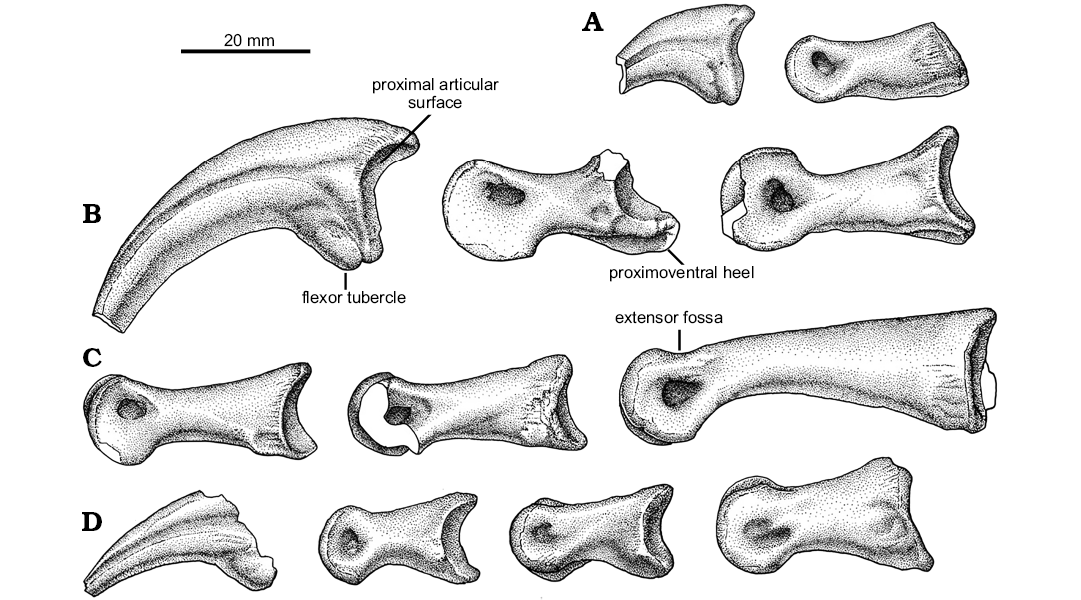
Fig. 10. Unenlagiid theropod Neuquenraptor argentinus Novas and Pol, 2005 (MCF PVPH 77) from Sierra del Portezuelo, Neuquén Province, Argentina; Portezuelo Formation, Coniacian, Late Cretaceous; left pedal phalanges: I (A), II (B), III (C), IV (D); in lateral views.
Phalanx III-2 is somewhat damaged, lacking portions of its distal and proximodorsal ends. The proximal articular facet is slightly asymmetrical, with the medial facet larger than the lateral one. Both lateral and medial proximal articular facets are divided by a very low median keel. On the proximoventral corner of the phalanx, there is a shallow flexor fossa, which is medially delimited by a low and thick flexor ridge. The distal end is slightly ginglymoid. Both trochlear rings slightly converge dorsally. The extensor fossa is well-defined and relatively shallow. Both collateral ligamental pits are well-defined and moderately deep.
Phalanx III-3 is sub-equal in length to phalanx III-2. The proximal articular facet is nearly identical to that of phalanx III-2, being slightly asymmetrical, but more transversely compressed. The extensor fossa is not well defined. The distal end is ginglymoid, and the trochlear margins show dorsally displaced collateral ligamental pits.
Digit IV is almost complete (Fig. 9D). It lacks phalanx IV-4 and the proximal end of phalanx IV-5. All the phalanges of the fourth digit are relatively short and stout.
The phalanx identified by Novas and Pol (2005) as pedal IV-2 is re-interpreted here as pedal IV-1 because of the shape of the proximal articular surface. Phalanx IV-1 is medially tilted and its shaft narrows proximally. The dorsal surface of the shaft is transversely narrower than the ventral surface. The proximal end of the phalanx shows strong rugosities in lateral and medial views. On the proximoventral corner of the phalanx there is a well-defined and shallow flexor fossa. The proximal articular surface is dorsoventrally tall and slightly asymmetrical, with the lateral side ventrally projected. It lacks a median keel. The distal end is ginglymoid and is notably transversely expanded when compared with the proximal end of the phalanx. The trochlear margins are notably distally divergent and bear large and deep collateral ligament pits. At the caudal margin of the lateral pit there exists a small bump, as occurs on metatarsal I. The extensor fossa is shallow and sub-triangular in contour.
Phalanx IV-2 is shorter than phalanx IV-1. The shaft is slightly medially tilted in dorsal view. There is a weak median keel that divides the proximal articular facet in two distinct surfaces. This facet is asymmetrical, with the lateral cotyle sub-triangular in contour and laterally extended on its ventral half. Ventrally, a small flexor depression is bounded by two flexor heels on both lateral and medial sides. The shaft of the phalanx is transversely compressed on its dorsal half. The distal end is ginglymoid, with the trochlear margins bearing well-developed ligament pits. The extensor fossa is shallow and sub-triangular in contour. The medial collateral ligament pit is well developed and is caudally continuous with a slightly concave surface on the medial surface of the phalanx.
Phalanx IV-3 is nearly sub-equal in length to the preceding phalanx, but it is notably transversely narrower. It is slightly asymmetrical when viewed in dorsal view. It main axis is obliquely oriented. The proximal articular facet is divided by a very shallow median dorsoventral keel, and has a large and laterally expanded lateral cotyle. The shaft is short and highly transversely compressed on its dorsal surface. On the medial side, and located at mid-shaft length, there is a well-developed bump (Fig. 8A), a feature that seems to be unique of Neuquenraptor. The distal end is ginglymoid with the trochlear rings bearing well-developed collateral ligament pits that are located on the middle of the trochlea. In the dorsal surface, the extensor fossa is shallow and poorly defined.
The ungual of digit IV is incomplete, lacking its proximal end. The claw is weakly curved, transversely compressed and lacks a cutting ventral edge. Instead, its ventral edge is represented by a flattened surface. On both lateral and medial surfaces, single, symmetrical longitudinal grooves are present. A portion of the flexor tubercle is preserved. Although incomplete, it seems to be large and rounded in contour.
Stratigraphic and geographic range.—Type locality and horizon only.
Concluding remarks
Comparisons.—The proximal end of the radius in Neuquenraptor is notably gracile and shows a well-constricted shaft distal to the proximal articulation. This results in a roughly hourglass-shape when the radius is viewed from the side. This condition is shared with other paravians such as Deinonychus, Sinornithoides, and Bambiraptor (Ostrom 1969; Currie and Dong 2001; Burnham 2004). The radius in therizinosaurs and oviraptorosaurs such as Falcarius and Khaan (Zanno and Erickson 2006; Balanoff and Norell 2012), and more basal tetanurans (e.g., Tyrannosaurus, Allosaurus; Madsen 1976; Brochu 2003) is more robust, with subparallel margins that delimitate a poorly constricted radial neck. Thus, Neuquenraptor morphology may be viewed as a derived condition shared by most paravians.
The femur of Neuquenraptor is very incomplete, and thus, anatomical comparisons are limited. In paravians, including troodontids, dromaeosaurids, and birds the fourth trochanter is reduced (Ostrom 1990; Chiappe 1996; Xu 2002), with the single exception of Velociraptor in which it is represented by a relatively well-developed ridge (Norell and Makovicky 1999). In Neuquenraptor, as in Buitreraptor, Unenlagia, and Austroraptor the fourth trochanter is represented by a scar surrounded by two bony longitudinal ridges of located near the proximal end of the shaft. Moreover, in contrast to other theropods, including Velociraptor and Deinonychus (Ostrom 1976; Norell and Makovicky 1999) the fourth trochanter is notably proximally positioned, near to the level of the trochanteric shelf. The morphology of the fourth trochanter in Unenlagia and Neuquenraptor is very similar in being more proximally positioned than in Buitreraptor. In this way, the presence of a proximally positioned scar-like fourth trochanter may be considered as a shared derived character uniting Unenlagia, Neuquenraptor, Austroraptor, and Buitreraptor and may constitute a synapomorphy of Unenlagiidae.
In Neuquenraptor, Unenlagia, Buitreraptor, and Velociraptor (Norell and Makovicky 1999), the femoral shaft possesses a proximodistally flattened surface that extends distal from the fourth trochanter, which is delimited by a well-developed caudal intermuscular line (Hutchinson 2001).
In Neuquenraptor the distal articulation of metatarsal I is ball-shaped and non-ginglymous as it occurs in Buitreraptor (MPCN-PV-598), troodontids (Xu and Wang 2000), and basal avialans, such as Archaeopteryx (Mayr et al. 2007). In contrast, in dromaeosaurids (e.g., Deinonychus, Velociraptor), the distal end of metatarsal I is notably ginglymous (Fowler et al. 2011).
In Neuquenraptor, metatarsal II has an expanded tongue-shaped flange that overlaps most of the caudal surface of metatarsal III. This condition was regarded as autapomorphic for Neuquenraptor by Novas and Pol (2005), but later Porfiri et al. (2011) and Gianechini and Apesteguía (2011) mentioned its presence in Pamparaptor and Buitreraptor. However, in the latter two taxa, such expansion is narrower than in Neuquenraptor, in which it is notably transversely expanded and proximodistally elongate. Presence of a tongue-shaped process is unknown in other paravians, and it may represent a synapomorphic feature of Unenlagiidae.
As indicated above, Neuquenraptor presents a subarctometatarsalian pes, which shows the proximal shaft of metatarsal III constricted between metatarsals II and IV, but still being exposed along most of the metatarsus in dorsal and ventral surfaces (Xu and Wang 2000). In Neuquenraptor the proximal half of the shaft of metatarsal III is notably craniocaudally compressed, being sublaminar in cross section. Further, it shows a very deep and wide longitudinal extensor groove on its dorsal surface. This condition is shared with Buitreraptor and probably Pamparaptor (Novas and Pol 2004; Porfiri et al. 2011). The condition is unknown in Austroraptor, in which the metatarsal III is very poorly preserved (Novas et al. 2009).
In microraptorians (e.g., Sinornithosaurus, Graciliraptor), the proximal portion of the shaft of metatarsal III is transversely compressed and dorsoventrally deep (Xu 2002). In derived troodontids such as Talos and Sinornithoides the proximal end of metatarsal III is splint-like (Osmólska and Barsbold 1990; Currie and Dong 2001; Zanno et al. 2011). In sum, the sublaminar and cranially excavated proximal portion of metatarsal III might be considered as a synapomorphic trait of Unenlagiidae. Distally the shaft of metatarsal III is sub-triangular in cross-section and the ventral margin becomes ridge-like, a morphology present in Buitreraptor, Pamparaptor (Makovicky et al. 2005; Porfiri et al. 2011), troodontids (Xu 2002), and microraptorians (Xu and Wang 2000).
In dorsal view the metatarsal III has a notable tongue-shaped flange that overlaps metatarsal II. On the other hand, metatarsal III do not overlaps the dorsal surface of metatarsal IV. In this way, the metatarsal IV is notably wide on its distal third, being sub-equal in width to the exposed portion of metatarsal II. Conversely in Buitreraptor and Pamparaptor, metatarsal IV is notably transversely compressed towards its distal end (Makovicky et al. 2005; Porfiri et al. 2011).
The distal third of metatarsal III in Neuquenraptor lies in a dorsal plane with respect to metatarsals II and IV. This is in concordance with the ventral displacement of the proximal half of this element results in a proximodistally obliquely oriented metatarsal III. This condition is also present in Buitreraptor (MPCA PV-598; Porfiri et al. 2011) and appears to be present also in Archaeopteryx (Thermopolis specimen, WDC-CSG-100; see Mayr et al. 2007).
In Neuquenraptor and other unenlagiids, on the caudal surface of metatarsal IV, there is a very well-developed longitudinal plantar crest that is present in a wide array of basal paravians (e.g., Archaeopteryx, Microraptor, Sinovenator, Deinonychus, Velociraptor, Rahonavis, Enantiornithes; Novas and Pol 2005; Agnolín and Novas 2011). However, in Neuquenraptor, Buitreraptor, and Pamparaptor this crest is notably convex in lateral view, especially at mid-length of metatarsal shaft. Furthermore, the caudal flange of Pamparaptor and Neuquenraptor are more expanded than in Buitreraptor, which might indicate a closer relationship between those taxa, as previously mentioned by Porfiri and colleagues (2011).
In Neuquenraptor the pedal phalanx IV-1 proximal end is transversely compressed and dorsoventrally tall, whereas the distal end is notably transversely expanded, a condition also present in Austroraptor among unenlagiids (Novas et al. 2009; Currie and Paulina Carabajal 2012). On the other hand in other paravians, including Buitreraptor (MPCN-PV-598), the proximal and distal ends are subequals in transverse width.
The phalanx III-1 is strongly ventrally arched, which is a feature present in other unenlagiids such as Pamparaptor (Porfiri et al. 2011) and Buitreraptor (MPCN-PV-598); but not in Austroraptor (Currie and Paulina Carabajal 2012). This condition is not present in other basal paravians and may result synapomorphic of Unenlagiidae.
The digit II in Neuquenraptor is modified into the typical paravian hyperextensible raptorial digit (Gauthier 1986; Rauhut and Werner 1995). As in unenlagiids and microraptorians the proximoventral heel of phalanx II-2 is transversely narrow and medially located (Makovicky et al. 2005). As in other paravians the phalanx II-1 is notably modified, having a much expanded distal articulation with trochlear margins caudally extended and separated from the rest of the phalanx by means of a notably deep extensor fossa (Ostrom 1969; Turner et al. 2007). The dorsal surface of the phalanx is also very concave. In Neuquenraptor the phalanx II-1 is sub-equal in length to phalanx II-2, whereas in “Unenlagia paynemili” and Buitreraptor the phalanx II-2 is shorter than II-1, a condition widespread in paravians (Ostrom 1969).
The metatarsus of Neuquenraptor shows well-developed and expanded plantar crests that delimit a notably deep and wide flexor groove. In birds of prey these is correlated with the accommodation of the bundle of tendons for the flexor muscles of the digits (Baumel and Witmer 1993). Furthermore, the expanded plantar crest constitutes enlarged anchoring of the flexor musculature of the foot (Baumel 1979). In the same way, the presence of a very deep and well-defined cranial extensor groove is related to the accommodation of the intrinsic extensor muscles of the digits (Baumel and Witmer 1993). These features, also present in other unenlagiids, are related to enhance raptorial capabilities.
Comments on the taxonomic status of Neuquenraptor argentinus.—In 1997, Novas and Puerta described Unenlagia comahuensis coming from the Portezuelo Formation (late Turonian–early Coniacian) of Sierra del Portezuelo, Neuquén Province. The holotype of this species consists on an incomplete skeleton. Later, Calvo and colleagues (2004) described isolated and incomplete elements as a new species of Unenlagia (U. paynemili). The specimens referred to this taxon come from Portezuelo Formation at Futalognko fossiliferous site. They distinguish both species on the basis of anatomical details.
Later, Novas and Pol (2005) described Neuquenraptor argentinus on the basis of hindlimb material coming from the Portezuelo Formation (late Turonian–early Coniacian) of Sierra del Portezuelo. Novas and Pol (2005) distinguished Neuquenraptor from Unenlagia both taxa on the basis of femoral proportions (the femur is the only overlapping element between both specimens). However, Makovicky et al. (2005) concluded that both femora are nearly identical and are similar in size, a criterion endorsed by Turner et al. (2012). On the contrary, Gianechini and Apesteguía (2011) indicate the femur in both taxa is heavily distorted, limiting comparisons between Neuquenraptor and U. comahuensis. Makovicky et al. (2005) and Turner et al. (2007, 2012) indicate that the co-occurrence and size similarities between the overlapping skeletal elements of Neuquenraptor and Unenlagia was suggestive evidence for a possible synonymy between each other. However, Novas (2009) criticized the later view, and considered that based on absence of informative overlapping material, there is no enough evidence to dismiss or support the synonymy of Neuquenraptor and Unenlagia (see also Gianechini and Apesteguía 2011). On the other hand, Porfiri et al. (2011) sustained that Neuquenraptor and Unenlagia may be valid taxa on the basis of pedal morphology.
In spite of the scarce overlapping material between Neuquenraptor and Unenlagia the proximal location of the fourth trochanter, as well as its morphology is nearly identical between both taxa.
However, the distal end of the tibia in Neuquenraptor argentinus and Unenlagia comahuensis show some anatomical differences. In distal view, the tibia of Neuquenraptor is transversely narrower (41 mm) than that of Unenlagia comahuensis (70 mm), and proportionaly craniocaudally thicker (Fig. 5E). Further, the preserved portion of the femur indicates that this element in Neuquenraptor was more gracile and slender than in Unenlagia comahuensis. On the other hand, pedal phalanx II-2 is sub-equal to II-1 in Neuquenraptor, whereas in materials referred to Unenlagia paynemili (Calvo et al. 2004; Porfiri et al. 2011; Gianechini and Apesteguía 2011; but see Novas 2009) phalanx II-2 is longer than phalanx II-1. Anyway, since the pedal phalanges described by Calvo et al. (2004) belong to Unenlagia (Porfiri et al. 2011) or to an indeterminate taxon (Novas 2009), they are different from those of Neuquenraptor. This indicates that at least two similar-sized unenlagiids were present on the Portezuelo Formation. In sum, in concordance with previous authors, we are not able to support nor dismiss the possible synonymy between Unenlagia and Neuquenraptor.
Acknowledgements
We thank Jorge Calvo (Centro Paleontológico Lago Barreales, Universidad Nacional del Comahue, Argentina), Juan Domingo Porfiri (Universidad Nacional del Comahue, Neuquén, Argentina), and Federico Gianechini (Universidad Nacional de San Luis, San Luis, Argentina) for discussions and comments on discussion on unenlagiid systematics and anatomy. We also acknowledge Sebastian Rozadilla and Matias Motta (both Laboratorio de Anatomía comparada y Evolución de los vertebrados -MACN), for their help during the confection of the present manuscript. We thank Rodolfo Coria (Museo Carmen Funes, Neuquén, Argentina) for allowing studying material under his care. Illustrations were skillfully made by Jorge Antonio Gonzalez (Argentina). We especially thank an anonymous reviewer and Alan Turner (Department of Anatomical Sciences, Stony Brook University, USA) who made valuable comments that greatly improved the quality of the manuscript.
References
Agnolín, F.L. and Novas, F.E. 2011. Unenlagiid theropods: are they members of Dromaeosauridae (Theropoda, Maniraptora). Anais da Academia Brasileira Ciencias 83: 117–162. Crossref
Agnolín, F.L. and Novas, F.E. 2013. Avian Ancestors: A Review of the Phylogenetic Relationships of the Theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. 96 pp. Springer, Dordrecht. Crossref
Agnolín, F.L., Novas, F.E., and Lio, G.L. 2006. Neornithine bird coracoid from the Upper Cretaceous of Patagonia. Ameghiniana 43: 245–248.
Balanoff, A.M. and Norell, M.A. 2012. Osteology of Khaan mckennai (Oviraptorosauria: Theropoda). Bulletin of the American Museum of Natural History 372: 1–77. Crossref
Baumel, A. 1979. Nomina Anatomica Avium. 779 pp. Academic Press, London.
Baumel, J.J. and Witmer, L.M. 1993. Osteologia. In: J.J. Baumel, A.S. King, J.E. Brazile, H.E. Evans, and J.C. Vanden Berge (eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium, 45–132. Nuttall Ornithological Society, Cambridge.
Bonaparte, J.F. 1999. Tetrapod faunas from South America and India: A palaeobiogeographic interpretation. Proceedings of the Indian National Science Academy 65: 427–437.
Brochu, C.R. 2003. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Society of Vertebrate Paleontology Memoirs 7: 1–138. Crossref
Burnham, D.A. 2004. New information on Bambiraptor feinbergi (Theropoda: Dromaeosauridae) from the Cretaceous of Montana. In: P.J. Currie, E.B. Koppelhus, M.A. Shugar, and J.L. Wright. (eds.),. Feathered Dinosaurs, 67–111. Indiana University Press, Bloomington.
Calvo, J.O., Porfiri, J.D., and Kellner, A.W. 2004. On a new maniraptoran dinosaur (Theropoda) from the Upper Cretaceous of Neuquén, Patagonia, Argentina. Archivos do Museu Nacional do Rio de Janeiro 62: 549–566.
Carrano, M.T. and Hutchinson, J.R. 2002. Pelvic and hindlimb musculature of Tyrannosaurus rex (Dinosauria: Theropoda). Journal of Morphology 253: 207–228. Crossref
Chiappe, 1996. Late Cretaceous birds of southern South America: Anatomy and systematics of Enantiornithes and Patagopteryx deferrariisi. In: G. Arratia (ed.), Contributions of Southern South America to Vertebrate Paleontology. Münchner Geowissenschaftliche Abhandlungen A 30: 203–244.
Currie, P.J. and Dong Z. 2001. New information on Cretaceous troodontids (Dinosauria, Theropoda) from the People’s Republic of China. In: P.J. Currie (ed.), Results from the Sino-Canadian Dinosaur Project. Part 3. Canadian Journal of Earth Sciences 38: 1753–1766. Crossref
Currie, P.J. and Paulina Carabajal, A. 2012. A new specimen of Austroraptor cabazai, Novas, Pol, Canale, Porfiri, and Calvo, 2008 (Dinosauria, Theropoda, Unenlagiidae) from the latest Cretaceous (Maastrichtian) of Río Negro, Argentina. Ameghiniana 49: 662–667. Crossref
Fowler, D.W., Freedman, E.A., Scannella, J.B., and Kambic, R.E. 2011. The predatory ecology of Deinonychus and the origin of flapping in birds. PLoS ONE 6 (12): e28964. Crossref
Gauthier, J.A. 1986. Saurischian monophyly and the origin of birds. Memoirs of the California Academy of Sciences 8: 1–55.
Gianechini, F.A. and Apesteguía, S. 2011. Unenlagiinae revisited: dromaeosaurid theropods from South America. Anais da Academia Brasileira de Ciencias 83 (1): 163–195. Crossref
Huene, F. von 1920. Bemerkungen zur Systematik und Stammesgeschichte einiger Reptilien. Zeitschrift für Induktive Abstammungs und Vererbungslehre 22: 209–212.
Hutchinson, J.R. 2001. The evolution of femoral osteology and soft tissues on the line to extant birds (Neornithes). Zoological Journal of the Linnean Society 131: 169–197. Crossref
Hutchinson, J.R., Anderson, F.C., Blemker, S.S., and Delp, S.L. 2005. Analysis of hindlimb muscle moment arms in Tyrannosaurus rex using a three-dimensional musculoskeletal computer model: implications for stance, gait, and speed. Paleobiology 31: 676. Crossref
Langer, M.C. and Benton, M.J. 2006. Early dinosaurs: a phylogenetic study. Journal of Systematic Palaeontology 4: 309–358. Crossref
Langer, M.C., Ribeiro, A.M., Schultz, C.L., Ferigolo, J. 2007. The continental tetrapod-bearing Triassic of south Brazil. In: S.G. Lucas and J.A. Spielmann (eds.), The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41: 201–218.
Leanza, H., Apesteguía, S., Novas, F.E., and de la Fuente, M.S. 2004. Cretaceous terrestrial beds from the Neuquèn Basin (Argentina) and their tetrapod assemblages. Cretaceous Research 25: 61–87. Crossref
Madsen, J.H., Jr. 1976. Allosaurus fragilis: a revised osteology. Utah Geological Survey Bulletin 109: 1–163.
Makovicky, P.J, Apesteguía, S., and Agnolín, F.L. 2005 The earliest dromaeosaurid theropod from South America. Nature 437: 1007–1011. Crossref
Marsh, 1881. A new order of extinct Jurassic reptiles (Coeluria). American Journal of Science (series 3) 21: 339–340.
Mayr, G., Pohl, B., Hartman, S., and Peters, D.S. 2007. The tenth skeletal specimen of Archaeopteryx. Zoological Journal of the Linnean Society 149: 97–116. Crossref
Novas, F.E. 1997. Anatomy of Patagonykus puertai (Theropoda, Avialae, Alvarezsauridae), from the Late Cretaceous of Patagonia. Journal of Vertebrate Paleontology 17: 137–166. Crossref
Novas, F.E. 1998. Megaraptor namunhuaiquii, gen. et sp. nov., a large-clawed, Late Cretaceous theropod from Patagonia. Journal of Vertebrate Paleontology 18: 4–9. Crossref
Novas, F.E. 2009. The Age of Dinosaurs in South America. 452 pp. Indiana University Press, Bloomington.
Novas, F.E. and Puerta, P. 1997. New evidence concerning avian origins from the Late Cretaceous of NW Patagonia. Nature 387: 390–392.Crossref
Novas, F.E. and Pol, D. 2005. New evidence on deinonychosaurian dinosaurs from the Late Cretaceous of Patagonia. Nature 433: 858–861. Crossref
Novas, F.E., Pol, D., Canale, J.I., Porfiri, J.D., and Calvo, J.O. 2009. A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids. Proceedings of the Royal Society of London B 276: 1101–1107. Crossref
Norell, M.A. and Makovicky, P.J. 1997. Important features of the dromaeosaur skeleton: information from a new specimen. American Museum Novitates 3215: 1–28.
Norell, M.A. and Makovicky, P.J. 1999. Important features of the dromaeosaur skeleton II: information from newly collected specimens of Velociraptor mongoliensis. American Museum Novitates 3282: 1–45.
Osmólska, H. and Barsbold, R. 1990. Troodontidae. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 259–268. University of California Press, Berkeley.
Ostrom, J.H. 1969. Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana. Bulletin of the Peabody Museum of Natural History 30: 1–165.
Ostrom, J.H. 1976. On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus. Breviora 439: 1–21.
Ostrom, J.H. 1990. Dromeosauridae. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 269–279. University of California Press, Berkeley.
Porfiri, J.D., Calvo, J.O., and Santos, D. 2011. A new small deinonychosaur (Dinosauria: Theropoda) from the Late Cretaceous of Patagonia, Argentina. Anais da Academia Brasileira de Ciencias 83: 109–116. Crossref
Rauhut, O.W.M. and Werner, C. 1995. First record of the family Dromaeosauridae (Dinosauria: Theropoda) in the Cretaceous of Gondwana (Wadi Milk Formation, northern Sudan). Paläontologische Zeitschrift 69: 475–489. Crossref
Remes, K. 2007. Evolution of the Pectoral Girdle and Forelimb in Sauropodomorpha (Dinosauria, Saurischia): Osteology, Myology, and Function. 355 pp. Ph.D. Thesis, Ludwig Maximilians-Universität, Munich.
Russell, D.A. 1969. A new specimen of Stenonychosaurus from the Oldman Formation (Cretaceous) of Alberta. Canadian Journal of Earth Sciences 6: 595–612. Crossref
Sereno, 1997. The origin and evolution of dinosaurs. Annual Review of Earth and Planetary Sciences 25: 435–489.
Snively, E., Russell, A.P., and Powell, L. 2004. Evolutionary morphology of the coelurosaurian arctometatarsus: descriptive, morphometric and phylogenetic approaches. Zoological Journal of the Linnean Society 142: 525–553. Crossref
Turner, A.H., Makovicky, P.J., and Norell, M. 2012. A review of dromaeosaurid systematics and paravian phylogeny. Bulletin of the American Museum of Natural History 371: 1–206. Crossref
Turner, A.H, Pol, D., Clarke, J.A., Erickson, G.M., and Norell, M.A. 2007. A basal dromaeosaurid and size evolution preceding avian flight. Science 317: 1378–1381. Crossref
Welles, S.P. and Long, R.A. 1974. The tarsus of theropod dinosaurs. Annals of the South African Museum 64: 191–218.
White, M.A. 2009. The subarctometatarsus: intermediate metatarsus architecture demonstrating the evolution of the arctometatarsus and advanced agility in theropod dinosaurs. Alcheringa 33: 1–21. Crossref
Xu, X. 2002. Deinonychosaurian Fossils from the Jehol Group of Western Liaoning and the Coelurosaurian Evolution. 322 pp. Ph.D. Thesis, Chinese Academy of Sciences, Beijing.
Xu, X. and Wang, X.-L. 2000. Troodontid-like pes in the dromaeosaurid Sinornithosaurus. Paleontology Society of Korea, Special Publication 2000 (4): 179–188.
Zanno, L.E. and Erickson, G.M., 2006, “Ontogeny and life history of Falcarius utahensis, a primitive therizinosauroid from the Early Cretaceous of Utah”. Journal of Vertebrate Paleontology 26: 143A.
Zanno, L.E., Varricchio, D.J., O’Connor, P.M., Titus, A.L., and Knell, M.J. 2011. A new troodontid theropod, Talos sampsoni gen. et sp. nov., from the Upper Cretaceous Western Interior Basin of North America. PLoS ONE 9 (6): e24487. Crossref
Acta Palaeontol. Pol. 62 (3): 549–562, 2017
https://doi.org/10.4202/app.00348.2017