
Applications and limitations of micro-XCT imaging in the studies of Permian radiolarians: A new genus with bi-polar main spines
YIFAN XIAO, NORITOSHI SUZUKI, and WEIHONG HE
Xiao, Y., Suzuki, N., and He, W. 2017. Applications and limitations of micro-XCT imaging in the studies of Permian radiolarians: A new genus with bi-polar main spines. Acta Palaeontologica Polonica 62 (3): 647–656.
Microfocus X-ray Computed Tomography (micro-XCT) has been employed recently in radiolarian studies, though so far primarily to generate high quality tomographic images. Although micro-XCT technique cannot always produce high-quality tomographic images, it frequently can provide valuable information on the internal structure of spongy polycystines. Here we employ micro-XCT to understand internal skeletal structures of several Permian specimens of polycystine radiolarians. Structural inferences from micro-XCT images are compared to images of the same specimens made with SEM and transmitted light microscopy (TLM). The utility of micro-XCT for imaging internal structures is first confirmed by examining the spongy, flat, four-spined species Tetraspongodiscus stauracanthus. Micro-XCT method is then used to examine the internal structures of a spherical to elliptical polycystine with two bi-polar main spines, Dalongicaepa bipolaris Xiao and Suzuki gen. et sp. nov., from the Dalong Formation (Changhsingian) of South China. The new genus is characterized by four to seven densely concentric shells with a large spherical hollow in the center and two cylindrical spines at both poles of the cortical shell, and belongs to the family Spongotortilispinidae. Spherical to elliptical polycystines with bi-polar main spines are similar in external appearance, and their phylogenetic relationships are only determinable by examination of the internal structures. We therefore analyzed all Permian and Mesozoic spherical to elliptical polycystines with bi-polar main spines showing internal structures, using cluster analysis to measure similarity. The results show distinctive differences in internal structures and suggest that family level relationships should be revised in the future.
Key words: Radiolaria, taxonomy, micro-XCT, Permian, Changhsingian, China.
Yifan Xiao [yifanxiao@cug.edu.cn], State Key Laboratory of Biogeology and Environmental Geology, School of Earth Science, China University of Geosciences, Wuhan 430074, China; Department of Earth Science, Graduate School of Science, Tohoku University, Sendai 980-8578, Japan.
Noritoshi Suzuki [norinori@m.tohoku.ac.jp], Department of Earth Science, Graduate School of Science, Tohoku University, Sendai 980-8578, Japan.
Weihong He [whzhang@cug.edu.cn] (corresponding author), State Key Laboratory of Biogeology and Environmental Geology, School of Earth Science, China University of Geosciences, Wuhan 430074, China.
Received 30 March 2017, accepted 22 May 2017, available online 14 August 2017.
Copyright © 2017 Y. Xiao et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Spongy spherical to elliptical polycystines with bi-polar main spines are common radiolarians since the Ordovician (Aitchison et al. 1998; Won and Iams 2011), but their phylogenetic taxonomic position, in most cases, has been poorly understood because higher taxonomic classification of polycystines, at both family and genus level, is often determinable only by observation of the inner structure of their tests (De Wever et al. 2001) and the inner structure of the shells is difficult to investigate. This has particularly affected Paleozoic and Mesozoic radiolarian taxonomy, where polycystine specimens are illustrated and identified mostly by surface skeletal structures imaged by SEM. Such images are sufficient for species identification, but may not for higher level classification. Naturally or artificially broken specimens can be used to examine internal structures but may provide limited morphological information. In the case of spongy spherical to elliptical polycystines with bi-polar main spines, only few reports showed the internal structure by SEM (Permian: Feng et al. 2006; Triassic: Kozur and Mostler 1981; Gorican and Buser 1990; Sugiyama 1997; Moix et al. 2007; Jurassic: Dumitrica and Zügel 2008). Several papers also illustrated TLM images of spongy spherical to elliptical polycystines with bi-polar main spines for Permian (Noble and Jin 2010), Triassic (Dumitrica et al. 1980; Bragin 1991; Sugiyama 1992), Jurassic (Baumgartner and Bernoulli 1976; Aita 1987), and Cretaceous (Foreman 1971; Renz 1974; Schaaf 1981; Nakaseko and Nishimura 1982; Bragina 1991) in age. Using similarity in external features only, without knowledge of internal structures, may result in dubious taxonomic assignments, such as accommodating the late Permian species Archaeospongoprunum mengi Feng in Feng et al., 2006 in the Campanian genus Archaeospongoprunum Pessagno, 1973.
Destructive sectioning methods to examine radiolarian internal structures have been used since the nineteenth century (see the review in Ogane and Suzuki 2007), but the number of specimens examinable is limited, and the specimens are destroyed in the procedure. Non-destructive methods have also been used to observe internal structures: diascopic lighting with a normal light microscope (Danelian and Popov 2003), confocal laser scanning microscopy (CLSM) (O’Connor 1996), differential interference contrast microscopy (DIC) or Nomarski system (Ling and Stadum 1970), and composite focal depth transmitted light microscopy (Jackett and Baumgartner 2007). Recently micro-XCT technique has been developed for radiolarian studies, but only a few results have been published so far (Matsuoka et al. 2012; Yoshino et al. 2015; Ikenoue et al. 2016).
Most of the above papers have chosen to use and impressively illustrate particularly well-preserved specimens. However, extremely well-preserved fossil radiolarians, especially the Permian ones, are rare. In this study we focus more on the routine extraction of taxonomic information using micro-XCT imaging, and less on specimen illustration, using several Permian polycystine specimens, deliberately chosen to have variable preservation to know whether micro-XCT still works well on them. The internal skeletal features identified are in particular used to clarify the structure and taxonomy of a spherical to elliptical polycystine species with two bi-polar main spines. We also compare our results from the latter species via cluster analysis to related genera.
Institutional abbreviations.—LGFEC, Laboratory of Geobiology, Faculty of Earth Sciences, China University of Geosciences, Wuhan, China.
Other abbreviations.—CLSM, confocal laser scanning microscopy; DIC, differential interference contrast microscopy; micro-XCT, microfocus X-ray computed tomography; TLM, transmitted light microscopy.
Material and methods
The specimens examined here were collected from the Changhsingian (late Lopingian, Permian) Dalong Formation, from the basinal facies, in the Rencunping section located in Rencunping Village, Sangzhi County, northwest Hunan Province, South China (110°6′2′′ E, 29°34′49′′ N) and housed in LGFEC. The Dalong Formation in the Rencunping section consists of thin-bedded chert and manganese-rich mudstone with intercalations of siliceous limestone and carbonaceous shale. This section is rich in macrofossils such as ammonoids (Zhang et al. 2009) and small brachiopods (He et al. 2014); and microfossils like conodonts (Tian 1993a, b; Yue 2014). Deposition of Dalong Formation started from the upper part of Wuchiapingian and ended in the uppermost Changhsingian. Details of the stratigraphy and radiolarian biostratigraphy will be published in a separate paper.
Specimens were obtained from samples 33, 36, and 69 by a hydrofluoric acid (HF) extraction method and imaged with a SEM (Hitachi SU8010 in State Key Laboratory of Biogeography and Environmental Geology, China University of Geosciences, Wuhan). The same specimens were also scanned by micro-XCT (SkyScan 1172 X-ray in the same laboratory), and 3-dimensional images were rendered with a free software CTVox (Bruker microCT, http://bruker-microct.com/home.htm).
Hierarchical cluster analysis was employed to investigate the correspondence of nine spherical to elliptical polycystine genera by using the statistical software “R” and the EZR (Kanda 2013) and FactoMineR packages (Lê et al. 2008).
Systematic palaeontology
Order Spumellaria Ehrenberg, 1876
Family Spongotortilispinidae Kozur and Mostler in Moix et al., 2007, sensu emend. Kozur et al. 2009 sensu emend. herein
Remarks.—The definition of this family was once revised to include genera having “polar spines with two or three terminal wings (Angulocircus and Dumitricasphaera), propeller-like ridges on the spines (Zhamojdasphaera), no modifications of the ridges on the spines (Spongotortilispinus and Vinassaspongus) or presence of two polar sines or of three spines in triangular arrangement.” (Kozur et al. 2009: 44). Kozur et al. (2009) also noted twisted spines are one of common characteristics in this family. However, as pointed by char-style-override-3 et al. (2012), twisted main spines appear in many Triassic spumellarians among phylogenetically distant families, but absent in particular genus or species. In consideration with this point, it is inappropriate for the twisting of main spines to be regarded as a characteristic feature at the family level. Consequently we omit presence of twisted main spines from the definition and expand the definition to include cylindrical main spines as well as tri-bladed spines.
Genus Dalongicaepa Xiao and Suzuki nov.
Type species: Dalongicaepa bipolaris Xiao and Suzuki gen. et sp. nov.; see below.
Species included: Pseudospongoprunum fontainei Sashida in Sashida et al., 2000.
Etymology: The combination of the strata name “Dalong” and the Latin caepa(-ae), onion (female noun).
Diagnosis.—Spongy cortical shell of spherical to fusiform shape. Spongy tissue comprised with four to seven densely concentric shells. Central part of the cortical shell large spherical hollow without a latticed spherical shell. Two significant bi-polar spines are present: cylindrical and no blades. The proximal end of the spine is embedded into the densely concentric shell to connect with a thin cylindrical radial beam. The radial beam pierces through the inner large spherical hollow.
Remarks.—It is practically difficult to distinguish species of Dalongicaepa from any similar Permian species by the superficial appearance of specimens so that diagnosis characters can be examined in only broken specimens to an extent. All the known genera belonging to the Spongotortilispinidae are easily distinguished from this new genus in having needle-shaped bi-polar spines of circular cross section. Based on a breakage specimen of Dalongicaepa fontainei (Sashida in Sashida et al. 2000) illustrated by Feng et al. (2006: pl. 3: 6), this species belongs to Dalongicaepa Xiao and Suzuki gen. nov. because of the presence of a hollow space in the shell central part.
Stratigraphic and geographic range.—Upper Permian, South China.
Dalongicaepa bipolaris Xiao and Suzuki sp. nov.
Fig. 1.
Etymology: From Latin bi- and polaris, two polars; in reference to the bipolar arrangement of the two polar spines.
Type material: Holotype (LGFEC SR 69-057), Fig. 1H. Paratype (LGFEC SR 69-028), Fig. 1A.
Type locality: Hunan Province, South China.
Type horizon: Upper Dalong Formation; upper Changhsingian.
Material.—Nine specimens (LGFEC SR 69-028, 69-029, 69-035, 69-057, 69-058, 69-062, 69-069, 69-090, 69-091).
Diagnosis.—Spongy spherical shell of two long bi-polar spines with uneven length.
Description.—Test spherical and spongy, composed of four to seven concentric shells. Two rod-like spines are thin, gradually decreasing in width distally, but quite different in length and arrangement: shorter or longer, straight or curving, opposite or obliquely opposed. In some specimens, the spines are particularly long and the length of spines can be twice longer than the diameter of the shell.
Measurements (in μm).—Diameter of test 159–179, length of spines 209–300, width of spines near base 14–29 (based on nine specimens).
Remarks.—The superficial appearance of this new species may be confused with two late Permian species Dalongicaepa fontainei and Archaeospongoprunum mengi. D. bipolaris Xiao and Suzuki gen. et sp. nov. is different from D. fontainei by having more concentric shells, spherical cortical shell, and longer thin polar main spines. Besides, the polar main spines of the new species distally decrease in width and sometimes curve. The new species is easily distinguished from A. mengi in the outline of its shell outline and three-bladed polar main spines.
Stratigraphic and geographic range.—Type locality and horizon only.
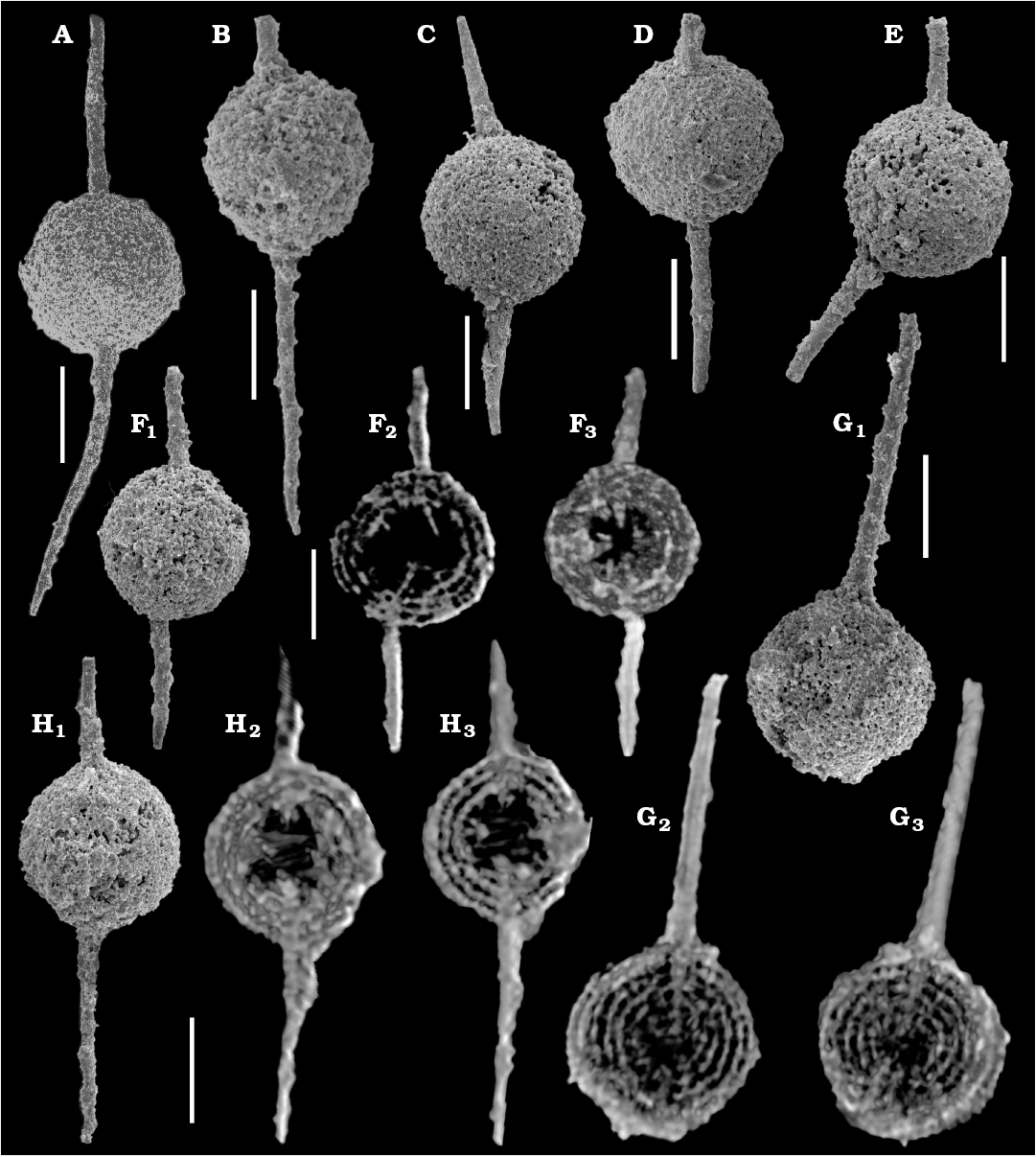
Fig. 1. SEM micrographs (A–E, F1, G1, H1) and micro-XCT cross-sectional images (F2, F3, G2, G3, H2, H3) of the polycystine radiolarian Dalongicaepa bipolaris Xiao and Suzuki gen. et sp. nov. from Changhsingian of the Rencunping section, South China. A. LGFEC SR 69-028 (paratype). B. LGFEC SR 69-090. C. LGFEC SR 69-058. D. LGFEC SR 69-091. E. LGFEC SR 69-029. F. LGFEC SR 69-062. G. LGFEC SR 69-069. H. LGFEC SR 69-057 (holotype). Different cross sections show clear concentric shells which are connected with radial beams (F2, F3, G2, G3, H2, H3). Scale bars 100 μm.
Results
A total of seven specimens of three morphospecies (two specimens of Klaengspongus spinosus Sashida in Sashida et al., 2000, two specimens of Tetraspongodiscus stauracanthus Feng in Feng et al., 2006, and three specimens of Dalongicaepa bipolaris Xiao and Suzuki gen. et sp. nov.) were scanned by micro-XCT. The Klaengspongus spinosus specimens were completely filled with a material (probably recrystallized quartz) on which the X-rays did not perform well. T. stauracanthus was selected to evaluate the quality of micro-XCT images since its internal structures have been illustrated previously with SEM (Feng et al. 2006: pl. 9: 13) and TLM (Noble and Jin 2010: pl. 7: 11). As shown in Fig. 2, the micro-XCT images displayed part of internal spicular system in the center, five concentric shells, the radial beam connecting with four main radial spines or embedded in the proximal part of them, and blades of the main spines. The specimens of D. bipolaris Xiao and Suzuki gen. et sp. nov. look like a simple spongy sphere with two rod-like bi-polar main spines, but their micro-XCT images (Fig. 1) show four to seven concentric structures with a large central hollow where some spiny structures are visible, and the radial beam which is directly connected with bi-polar main spines.
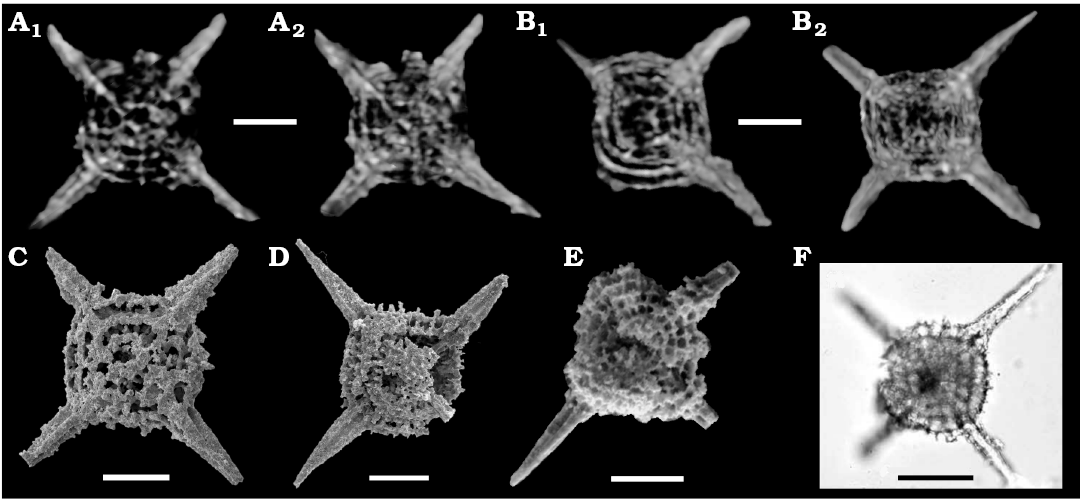
Fig. 2. Micro-XCT cross-sectional images (A, B), SEM micrographs (C–E), and TLM micrographs (F) of the spumellarian Tetraspongodiscus stauracanthus Feng in Feng et al., 2006 from Changhsingian of the Rencunping section (A–D) and Dongpan (E), South China and upper Capitanian of West Texas, USA (F). A. LGFEC SR 33-201. B. LGFEC SR 33-202. C. LGFEC SR 36-011. D. LGFEC SR 33-022. E. DP 4/4860 (from Feng et al. 2006; copyright is permitted from Elsevier). F. S3-2A (from Noble and Jin 2010). Different cross sections show clear hoops with few primary beams and a possible median bar (A1 and B1), the centrical internal spicule (A2), and the embedded proximal part of the main spines (B2). Scale bars 50 μm.
All Permian and Mesozoic genera which are spherical to elliptical spumellarians with bi-polar main spines and for which internal structures have been reported are illustrated in Fig. 3. Cluster analysis of the matrix (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app62-Xiao_etal_SOM.pdf) generated from a total of nine genera and nine characters under the corresponding analysis were analyzed (see SOM 2) and resulted in three major clusters (Fig. 4). Cluster 1 is composed of two genera: Spongotortilispinus Kozur and Mostler in Moix et al., 2007 and Dalongicaepa Xiao and Suzuki gen. nov. Cluster 2 consists of two genera: Archaeospongoprunum and Palaeospongurus Kozur and Mostler, 1981. Cluster 3 includes five genera: Spongoxystris Sugiyama, 1997, Spongopallium Dumitrica, Kozur, and Mostler, 1980, Falcispongus Dumitrica, 1982, Protopsium Pessagno and Poisson, 1981, and Pegoxystris Sugiyama, 1992.
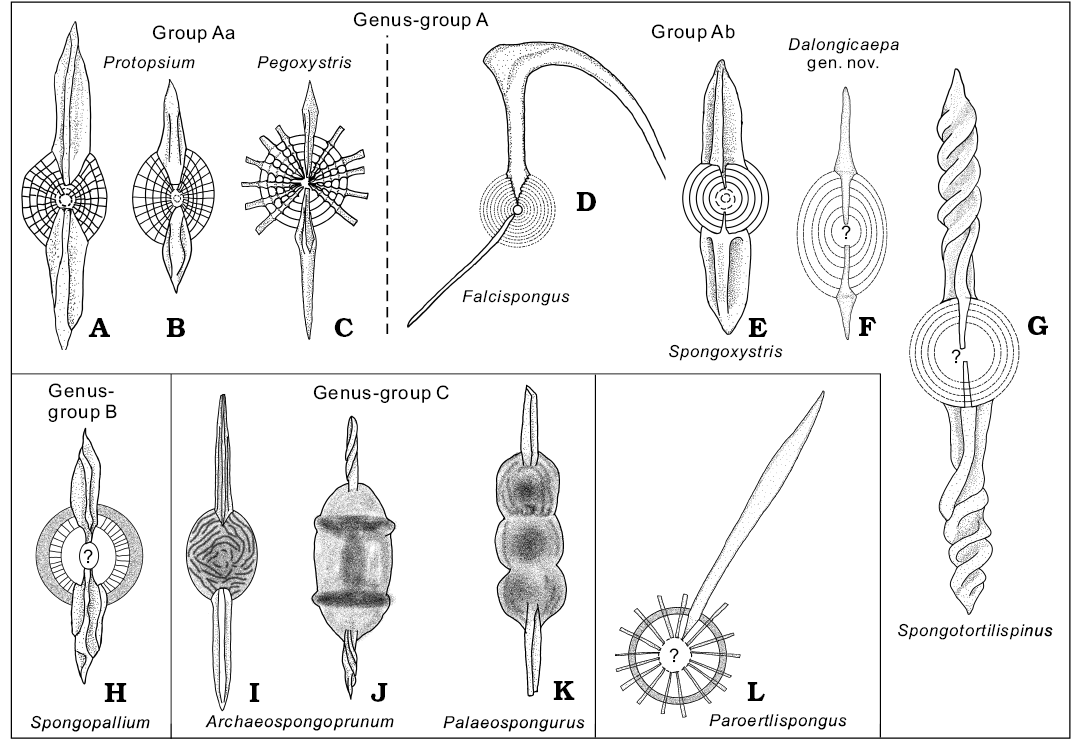
Fig. 3. Schematic drawings on internal structure of Permian and Mesozoic spumellarians with bi-polar main spines (detailed information including source is summarized in Table 2). A, B. Protopsium. C. Pegoxystris. D. Falcispongus. E. Spongoxystris. F. Dalongicaepa Xiao and Suzuki gen. nov. G. Spongotortilispinus. H. Spongopallium. I, J. Archaeospongoprunum. K. Palaeospongurus. L. Paroertlispongus.

Fig. 4. Dendrogram of the cluster analysis under the corresponding analysis based on nine genera (performed by software “R” and the EZR ).
Discussion
The usefulness and limitations of “ordinary” micro-XCT in detecting structures in spongy flat polycystines with four main spines.—Tetraspongodiscus stauracanthus is a common species in rocks of the upper Changhsingian of South China (Feng et al. 2006; He et al. 2008, 2011) and southwestern China (Yao and Kuwahara 2000), the Changhsingian of central Japan (Sashida and Tonishi 1985) and the upper Capitanian of North America (Noble and Jin 2010). A full synonym list of this species will be published in a separate paper. Morphological terminology here follows Ogane and Suzuki (2006). This species is characterized by a flat, subcircular and spongy crust (outer shell) with four three-bladed main spines that are arranged in the shape of a cross in the equatorial plane. SEM images of our broken specimens (Fig. 2C, D) display five or more concentric to spiral hoops. The broken surface of the specimen illustrated in Feng et al. (2006; Fig. 2E herein) is exposed on the equatorial plane, and shows concentric hoops that are connected with primary beams. The primary beam is defined as a radial beam that penetrates the paleas and hoops (Ogane and Suzuki 2006). However, the relationship between main spines and hoops are not clear in these SEM images. An informative TLM was published in Noble and Jin (2010; Fig. 2F herein). This image also shows relevant structures which are observable in the SEM images to an extent, but it is not possible to confirm the type of radial beams such as primary, secondary interconnecting beams of Ogane and Suzuki (2006). The central structure and the relationship between main spines and hoops are also unclear. By way of comparison, our micro-XCT images (Fig. 2A, B) clearly show five hoops with few primary beams, the embedded proximal part of the main spines, and a connecting bar in the center of the shell. It is noted that the connecting bar appears to join one of the main spines at least (Fig. 2A1, B1), suggesting a possible median bar. The presence of median bar makes it possible to suggest their positions in the Entactinaria (median bar is not present in Spumellaria) pending further research. By contrast, primary beams are not shown in our micro-XCT images.
The comparison of SEM, TLM, and micro-XCT images shows that all the internal structures except for fine radial beams can be well recognized by micro-XCT imagery, while SEM images are useful only to broken specimens. TLM images also give a sight of internal structures, but less clear (Table 1). Micro-XCT images even suggest or show the possible presence of median bar. Nevertheless, the limitation of the “ordinary” micro-XCT analysis is obvious in that it is not applicable to those specimens whose entrails are filled with recrystallized quartz even if they are superficially well preserved.
Table 1. Features observed among different photos taken by micro computed tomography (micro-XCT), scanning electron microscopy (SEM), and transmitted light microscopy (TLM).
|
Method |
Outer shell |
Spine shape |
Inner shells |
Innermost structure |
Internal rays |
|
Micro-XCT |
yes |
yes |
yes |
yes |
occasionally, if rays are strong |
|
SEM |
yes |
yes |
occasionally, if broken |
occasionally, if broken |
occasionally, if broken |
|
TLM |
no |
no |
yes, but not very good |
yes, but not very good |
occasionally, if beams are strong |
Recognition of internal structure of spongy spherical polycystines with bi-polar main spines using by micro-XCT images.—This section is based on images of Dalongicaepa bipolaris Xiao and Suzuki gen. et sp. nov. Terminology follows Petrushevskaya (1984) and Suzuki (2006). The SEM images (Fig. 1A–E, F1, G1, H1) show a thick, spongy, spherical outer shell with two bi-polar main spines. The spines are thin, long, rod-like, and variable in length and arrangement: shorter or longer, straight or curving, opposite or obliquely opposed. Similar features would be visible on TLM images although we have not produced them. We made an attempt to observe the internal structures, but appropriate (broken) specimens were not found. The overall appearance is similar to Pararchaeospongoprunum hermi Lahm, 1984, Dalongicaepa fontainei (Sashida in Sashida et al., 2000), Paroertlispongus obliquus Kozur and Mostler, 1994, and Yangia chinensis Feng in Feng and Liu, 1993 in having rod-like bi-polar main spines. As these externally similar species belong to different genera, it is very hard to classify our species not only into the appropriate genus but also even to the family due to the lack of internal structure information. Feng et al. (2006) show a hollow space in D. fontainei like that of D. bipolaris, and Sano et al. (2010: fig. 10.22) show a TLM image of this species, but it is unable to determine the presence of a hollow space in their figure. This illustrates the difficulty in recognition of hollow spaces in TLM images. Micro-XCT by contrast shows four to seven concentric shells which are connected with radial beams (Fig. 1F2, F3, G2, G3, H2, H3). A hollow space with or without some fine rods always exists in the innermost central part of all examined specimens, indicating a common morphological character in this species. The size of the hollow space is variable, probably caused by growth or damage. Smaller hollow spaces are seen in the specimens with seven concentric shells while larger hollow spaces are observed in those with a smaller number of concentric shells. It is noteworthy that two radial beams extend inward from the proximal end of the main spines towards the hollow space. The specimen illustrated in Fig. 1F has offset bi-polar main spines (Fig. 1F1) which join to a radial beam which is obliquely placed in the hollow space (Fig. 1F2). Several other beams on the inner wall of the hollow space are not connected with two main spines. The existence of many isolated radial beams inside the hollow space implies the presence of other fine inner structures which are however not detected in our images, and might have been dissolved by preservational alteration.
Classification of spongy spherical polycystines with bi-polar main spines.—Similarity in outer appearances among many spherical to elliptical polycystines with bi-polar main spines are also well documented in the Cenozoic. Phylogenetic questions can in principle be more easily addressed due to better visibility of internal characters in the generally well preserved (primary transparent opal) of Cenozoic radiolarians, although such observations are still incomplete (Suzuki et al. 2009; Matsuzaki et al. 2015). The situation is more difficult in spongy spherical polycystines with bi-polar main spines in the Permian. It is so difficult to examine their internal structure that several Permian species with bi-polar main spines were tentatively assigned to the genera from geochronologically (and thus probably also biologically) distant material, e.g., the Silurian genus Pseudospongoprunum Wakamatsu, Sugiyama, and Furutani, 1990, the Triassic genus Paroertlispongus Kozur and Mostler, 1981, or the Cretaceous genus Archaeospongoprunum. To improve our phylogenetic understanding of these Permian forms, we collected all Permian and Mesozoic publications showing internal structures of these taxa from ca. 5300 papers on radiolarian taxonomy held by Tohoku University (including all geologic ages). We identified a total of 10 genera (12 species, 17 specimens) where the internal structures were illustrated (Table 2; Fig. 3), although details in the most central part of the shell (e.g., microsphere, the entactinarian spicular system) were usually unclear.
Table 2. Detailed information on schematic drawings on Fig. 3.
|
Figure |
Genus |
Currently assignable name |
Original name |
Age |
Family |
Status |
Source |
|
Fig. 3A |
Protopsium Pessagno and Poisson, 1981 |
Protopsium? sp. |
“Archaeospongoprunum
sp.” (Bragin, 1991); Spongopallium?
|
Middle Triassic |
Sponguridae (De Wever 1982), Angulobracchidae (De Wever et al. 2001), Archaeospongoprunidae (O’Dogherty et al. 2009a) |
– |
Bragin 1991: pl. 9: 15; Gorican and Buser 1990: pl. 4: 4 |
|
Fig. 3B |
Protopsium Pessagno and Poisson, 1981 |
Protopsium? ispartaense Pessagno and Poisson, 1981 |
Protopsium
ispartaense |
Pliensbachian |
Sponguridae (De Wever
1982), |
paratype |
Pessagno and Poisson 1981: pl. 13: 4 |
|
Fig. 3C |
Pegoxystris |
Pegoxystris mizutanii Sugiyama, 1992 |
Pegoxystris
mizutanii |
Olenekian |
Archaeospongoprunidae? |
holotype of the type species of the genus Pegoxystris |
Sugiyama 1992: fig. 17.8 |
|
Fig. 3D |
Falcispongus
|
Falcispongus
|
Falcispongus
calcaneum |
Anisian to Carnian |
Oertlispongidae |
holotype |
Dumitrica 1982: pl. 1: 1 and text-fig. 1 |
|
Fig. 3E |
Spongoxystris |
Spongoxystris hadra Sugiyama, 1997 |
Spongoxystris
hadra |
Olenekian |
Spongotortilispinidae |
holotype of the type species of the genus Spongoxystris |
Sugiyama 1997: figs. 46.15, 46.16 |
|
Fig. 3F |
Dalongicaepa Xiao and Suzuki gen. nov. |
Dalongicaepa
fontainei |
Paroertlispongus
fontainei |
Capitanian to Changhsingian |
Spongotortilispinidae |
– |
Feng et al. 2006: pl. 3: 6 |
|
Fig. 3G |
Spongotortilispinus
Kozur and Mostler |
Spongotortilispinus tortilis (Kozur and Mostler, 1979) |
Spongostylus
tortilis |
Ladinian to Norian |
Spongotortilispinidae |
topotype of this species |
Kozur and Mostler 1981: pl. 56: 3 |
|
Fig. 3H |
Spongopallium |
Spongopallium
|
Spongopallium
contortum |
Anisian to Ladinian |
Spongopallidae |
paratype of the type species of the genus Spongopallium |
Dumitrica
et al. 1980: pl. 11: 2; |
|
Fig. 3I |
Archaeospongoprunum
|
Archaeospongoprunum patricki Jud, 1994 |
Archaeospongoprunum tehamaensis Pessagno, 1973 (Schaaf 1981); Spongoprunum aff. probus (Rüst, 1888) (Renz 1974) |
Tithonian to Albian |
Archaeospongoprunidae |
– |
Schaaf 1981: pl. 10: 7; Renz 1974: pl. 10: 19 |
|
Fig. 3J |
Archaeospongoprunum
|
Archaeospongoprunum? nishiyamai Nakaseko and Nishimura, 1982 |
Spongoprunum
sp. (Foreman 1971); Archaeospongoprunum
nishiyamai Nakaseko and Nishimura, 1982 |
Santonian to Campanian |
Archaeospongoprunidae |
paratype |
Foreman 1971: pl. 1: 2; Nakaseko and Nishimura 1982: pl. 14: 4 |
|
Fig. 3K |
Palaeospongurus
|
Palaeospongurus triplus (Pessagno, 1973) |
Archaeospongoprunum triplum Pessagno, 1973 |
Coniasian to Campanian |
Sponguridae (Kozur and Mostler 1981), Archaeosponguridae (O’Dogherty 2009b) |
– |
Bragina 1991: pl. 12: 6 |
|
Fig. 3L |
Paroertligpongus Kozur and Moslter, 1981 |
Paroertlispongus
|
Paroertlispongus multispinosus Kozur and Mostler, 1981 |
Anisian to Ladinian |
Oertlispongidae |
paratype of the type species of the genus Paroertlispongus |
Kozur and Mostler 1981: pl. 45: 1a, b |
|
not shown but a similar to Fig. 3G |
Spongotortilispinus,
Kozur and Mostler |
Spongotortilispinus sp. |
Spongotortilispinus sp. |
Carnian |
Spongotortilispinidae (type genus) |
– |
Moix et al. 2007: pl. 2: 8 |
As shown in Fig. 3, they can be divided into three groups: (i) genus-group A with concentric shells (Fig. 3A–G), (ii) genus-group B with a probable double medullary shell (Fig. 3H), and (iii) genus-group C with fully spongy meshwork (Fig. 3I–K). The genus Paroertlispongus could not be assigned to any group due to a lack of information on its innermost structure. The first group is further subdivided into two subgroups either having distinctive radial beams (group Aa, Fig. 3A–C) or not (group Ab, Fig. 3D–G). Group Aa consists of Protopsium (Fig. 3A, B) and Pegoxystris (Fig. 3C). They have a common structure: the embedded proximal part of main spines in the shells. In group Ab, the combination of the central part with the shape of the embedded proximal part of main spines are characteristic. The genus Falcispongus has thorny connections with each concentric shell on the embedded proximal part of the main spines (Fig. 3D; see also Dumitrica 1982: text-fig. 1). Spongoxystris (Fig. 3E), Dalongicaepa fontainei (Fig. 3F), and Spongotortilispinus (Fig. 3G) have a common rod-like proximal part of the main spines, but the first one is different from the latter two in that Dalongicaepa fontainei and Spongotortilispinus have a large hollow in their center. Similar to these two radiolarians, the sole genus in genus-group B has a large hollow center (Spongopallium; Fig. 3H), but Spongopallium is characterized by the development of radial structures on the inner side of the spongy cortical shell and has different structure of the proximal part of the radial spines. Genus-group C comprises Archaeospongoprunum and Palaeospongurus, but the internal structure is unknown, hidden within the spongy outer structure. Our examined specimens of Dalongicaepa have an internal structure in common with both Dalongicaepa fontainei and Spongotortilispinus.
This qualitative observation can be statistically supported by the cluster analysis under the corresponding analysis on nine genera, indicating three major clusters (Fig. 4). Cluster 1 is composed of two genera: Spongotortilispinus and Dalongicaepa. Spongotortilispinus belongs to the family Spongotortilispinidae Kozur and Mostler in Moix et al., 2007, sensu emended by Kozur et al. (2009), which contain the Triassic genera Dumitricasphaera Kozur and Mostler, 1979, Vinassaspongus Kozur and Mostler, 1979, Zhamojdasphaera Kozur and Mostler, 1979, and Angulocircus Lahm, 1984; but these genera have nothing in common except for an internal structure. Spongotortilispinus essentially has twisted bi-polar main spines but such spines have not been recognized in any Lopingian spherical polycystines. As discussed in the systematic paleontology above, our examined species belongs to the same new genus with Dalongicaepa fontainei. Combined with the similar internal structure, the new genus Dalongicaepa undoubtedly belongs to the same family as Spongotortilispinus. Cluster 2 consists of two genera: Archaeospongoprunum and Palaeospongurus. This cluster corresponds to the above mentioned genus-group C, thus the opinion of O’Dogherty (2009b) is accepted that they both belong to the family Archaeospongoprunidae Pessagno, 1973. Cluster 3 includes five genera: Spongoxystris, Spongopallium, Falcispongus, Protopsium, and Pegoxystris. There seems to be an inconsistency to the qualitative examination to an extent at the level of family. According to O’Dogherty et al. (2009a, b), Spongoxystris belongs to Spongotortilispinidae, Spongopallium to Spongopalliidae Kozur, Krainer, and Mostler, 1996, Falcispongus to Oertlispongidae, and both Protopsium and Pegoxystris to Archaeospongoprunidae, respectively. Both qualitative and statistical analysis do not support the assigned families to Spongoxystris, Protopsium, and Pegoxystris at least. The dendrogram makes clear that a revision is needed of these genera at the level of family.
Conclusions
Although micro-XCT technique not always produces high-quality tomographic images because of the specimens preservation, it frequently can provide valuable information on the internal structure of spongy polycystines. We use micro-XCT images to clarify what skeletal parts can or cannot be recognized after comparing the detectable and undetectable structures between micro-XCT, SEM, and TLM images of a spongy flat polycystine with four main spines.
A new genus and species Dalongicaepa bipolaris Xiao and Suzuki are proposed after carefully examining the inner structures using micro-XCT. The new species has four to seven concentric shells connected by multiple thin internal rays and a hollow space in the innermost central part. Two rays extend inward from the main spines towards the innermost space. All these characteristics suggest the new genus belongs to the family Spongotortilispinidae.
Since it is quite difficult to differentiate spherical to elliptical polycystines with bi-polar main spines because of their similarity in superficial appearance, we analyzed all Permian and Mesozoic taxa with published internal structures (Archaeospongoprunum, Dalongicaepa Xiao and Suzuki gen. nov., Falcispongus, Palaeospongurus, Paroertlispongus, Pegoxystris, Protopsium, Spongopallium, Spongotortilispinus, and Spongoxystris) for better phylogenetic understanding. The schematic illustration of the internal structure and dendrogram of the cluster analysis show distinctive differences in their internal structure which suggests that their taxonomic position at the level of family should be revised in future. Going further, “ordinary” micro-XCT images for spherical polycystines could play an important role in differeniation between Spumellaria and Entactinaria because the Entactinaria differ from Spumellaria in having an internal spicular system including median bar.
Although the images produced by micro-XCT are not always appealing, we believe, they should be produced and published, regardless of their esthetic value, as they undoubtedly augment the information on the investigated objects and above all the knowledge of their internal structure.
Acknowledgements
We thank Songzhu Gu and Muhui Zhang (both China University of Geosciences, Wuhan, China) for their technical assistance in micro-XCT images. We gratefully acknowledge David Lazarus (Museum für Naturkunde, Berlin, Germany) for proofing our English. We also thank Nikita Bragin (Geological Institute RAS, Moscow, Russia) and an anonymous reviewer for their valuable comments. Financial support by NSFC (Grant Nos. 41372030, 40872008 and 41602024), the Ministry of Education of China (B08030 of 111 Project), the State Key Laboratory of Biogeology and Environmental Geology (GBL11605) and the Fundamental Research Funds for the Central University of CUG (CUG 160842) are greatly acknowledged.
References
Aita, Y. 1987. Middle Jurassic to Lower Cretaceous radiolarian biostratigraphy of Shikoku with reference to selected sections in Lombardy Basin and Sicily. Science Reports of the Tohoku University. Second Series, Geology 58: 1–91.
Aitchison, J.C., Flood, P.G., and Malpas, J. 1998. Lowermost Ordovician (basal Tremadoc) radiolarians from the Little Port Complex, western Newfoundland. Geological Magazine 135: 413–419. Crossref
Baumgartner, P.O. and Bernoulli, D. 1976. Stratigraphy and radiolarian fauna in a Late Jurassic–Early Cretaceous section near Achladi (Evvoia, eastern Greece). Eclogae Geologicae Helvetiae 69: 601–626.
Bragin, N.Yu. [Bragin, N.Û.] 1991. Radiolârii i Nižnemezozojskie Tolŝi Vostoka SSSR. 125 pp. Nauka, Moskva.
Bragina, L.G. 1991. Radiolarians in Santonian–Campanian deposits of the Bystrinskaya Formation of the Northwestern Kamchatka [in Russian]. Izvestiâ Akademi Nauk SSSR, Seria Geologičeskaâ 7: 129–136.
Danelian, T. and Popov, L. 2003. Ordovician radiolarian biodiversity: insights based on new and revised data from Kazakhstan. Bulletin de la Société Géologique de France 174: 325–335. Crossref
De Wever, P. 1982. Radiolaires du Trias et du Lias de la Téthys (Systématique, Stratigraphie). Société Géologique du Nord 7: 1–599.
De Wever, P., Dumitrica, P., Caulet, J.P., Nigrini, C., and Caridroit, M. 2001. Radiolarians in the Sedimentary Record. 533 pp. Gordon and Breach Science Publishers, Amsterdam.
Dumitrica, P. 1982. Triassic Oertlisponginae (Radiolaria) from eastern Carpathians and Southern Alps. Dari de Seama ale Sedintelor 67: 57–74.
Dumitrica, P. and Zügel, P. 2008. Early Tithonian Saturnalidae (Radiolaria) from the Solenhofen area (southern Franconian Alb, southern Germany). Paläontologische Zeitschrift 82: 55–84. Crossref
Dumitrica, P., Kozur, H., and Mostler, H. 1980. Contribution to the radiolarian fauna of the Middle Triassic of the Southern Alps. Geologisch-Paläontologische Mitteilungen Innsbruck 10: 1–46.
Ehrenberg, C.G. 1876. Fortsetzung der mikrogeologischen Studien als Gesammt-Übersicht der mikroskopischen Paläontologie gleichartig analysirter Gebirgsarten der Erde, mit specieller Rücksicht auf den Polycystinen-Mergel von Barbados. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1875: 1–226.
Feng, Q.L. and Liu, B.P. 1993. Radiolaria from Late Permian and Early–Middle Triassic in southwest Yunnan. Earth Science, Journal of China University of Geosciences 18: 540–552.
Feng, Q.L., He, W.H., Gu, S.Z., Jin, Y.X., and Meng, Y.Y. 2006. Latest Permian Spumellaria and Entactinaria (Radiolaria) from South China. Revue de Micropaléontologie 49: 21–43. Crossref
Foreman, H.P. 1971. Cretaceous Radiolaria, Leg 7, DSDP. In: E.L. Winterer, W.R. Riedel, P. Brönnimann, E.L. Gealy, G.R. Heath, L. Kroenke, E. Martini, R. Moberly, Jr., J. Resig, and T. Worsley (eds.), Initial Reports of the Deep Sea Drilling Project, Volume 7, 1673–1693. U.S. Government Printing Office, Washington, D.C.
Gorican, S. and Buser, S. 1990. Middle Triassic radiolarians from Slovenia (Yugoslavia). Geologija 31/32: 267–304.
Guex, J., O’Dogherty, L., Carter, E.S., Goričan, Š., Dumitrica, P., and Bartolini, A. 2012. Geometrical transformations of selected Mesozoic radiolarians. Geobios 45: 541–554. Crossref
He, W.H, Shi, G.R., Zhang, Y., Yang, T.L., Zhang, K.X., Wu, S.B., Niu, Z.J., and Zhang, Z.Y. 2014. Changhsingian (latest Permian) deep-water brachiopod fauna from South China. Journal of Systematic Palaeontology 12: 907–960. Crossref
He, W.H., Zhang, Y., Zhang, Q., Zhang, K.X., Yuan, A., and Feng, Q.L. 2011. A latest Permian radiolarian fauna from Hushan, South China, and its geological implications. Alcheringa 35: 471–496. Crossref
He, W.H., Zhang, Y., Zheng, Y., Zhang, K.X., Gui, B., and Feng, Q.L. 2008. A late Changhsingian (latest Permian) radiolarian from Chaouhu, Anhui and a comparison with its contemporary faunas of South China. Alcheringa 32: 199–222. Crossref
Ikenoue, T., Bjørklund, K.R., Kimoto, K., Matsuno, K., and Harada, N. 2016. Two new living Entactinaria (Radiolaria) species from the Arctic province: Joergensenium arcticum n. sp. and Joergensenium clevei n. sp. Marine Micropaleontology 124: 75–94. Crossref
Jackett, S.J. and Baumgartner, P.O. 2007. New imaging techniques applied to Paleogene radiolaria. Micropaleontology 53: 239–247. Crossref
Kanda, Y. 2013. Investigation of the freely available easy-to-use software “EZR” for medical statistics. Bone Marrow Transplanation 48: 452–458. Crossref
Kozur, H. and Mostler, H. 1979. Beiträge zur Erforschung der mesozoischen Radiolarien. Teil III: Die Oberfamilien Actinommacea Haeckel 1862 emend., Artiscacea Haeckel 1882, Multiarcusellacea nov. der Spumellaria und triassische Nassellaria. Geologisch-Paläontologische Mitteilungen Innsbruck 9: 1–132.
Kozur, H. and Mostler, H. 1981. Beitrage zur Erforschung der mesozoischen Radiolarien. Teil IV: Thalassosphaeracea Haeckel 1862, Hexastylacea Haeckel 1862, emended Petrushevskaja 1979, Sponguracea Haeckel 1862, und weitere triassisch Lithocycliacea, Trematodiscea, Actinommacea und Nassellaria. Geologisch-Paläontologische Mitteilungen Innsbruck, Sonderband 1981: 1–208.
Kozur, H. and Mostler, H. 1994. Anisian to Middle Carnian radiolarian zonation and description of some stratigraphically important radiolarians: Geologisch-Paläontologische Mitteilungen Innsbruck, Sonderband 3: 39–255.
Kozur, H., Krainer, K., and Mostler, H. 1996. Radiolarians and facies of the Middle Triassic Loibl Formation, South Alpine Karawanken Mountains (Carinthia, Austria). Geologisch-Paläontologische Mitteilungen Innsbruck, Sonderband 4: 159–269.
Kozur, H.W., Moix, P., and Ozsvárt, P. 2009. New Spumellaria (Radiolaria) from the early Tuvalian Spongotortilispinus moixi Zone of southeastern Turkey, with some remarks on the age of this fauna. Jahrbuch der Geologischen Bundesanstalt 149: 25–59.
Lahm, B. 1984. Spumellarienfaunen (Radiolaria) aus den mitteltriassischen Buchensteiner-Schichten von Recoaro (Norditalien) und den obertriasschen Reifingerkalen von Großreifling (Österreich) – Systematik – Stratigraphie. Münchner Geowissenschaftliche Abhandlungen A 1: 1–161.
Lê, S., Josse, J., and Husson, F. 2008. FactoMineR: An R package for multivariate analysis. Journal of Statistical Software 25: 1–18. Crossref
Ling, H.-Y. and Stadum, C.J. 1970. An evaluation of the Nomarski system for the study of siliceous microskeletons. Journal of Paleontology 44: 781–784.
Matsuoka, A., Yoshino, T., Kishimoto, N., Ishida, N., Kurihara, T., Kimoto, K., and Matsuura, S. 2012. Exact number of pore frames and their configuration in the Mesozoic radiolarian Pantanellium: An application of X-ray micro-CT and layered manufacturing technology to micropaleontology. Marine Micropaleontology 88–89: 36–40. Crossref
Matsuzaki, K.M., Suzuki, N., and Nishi, H. 2015. Middle to Upper Pleistocene polycystine radiolarians from Hole 902-C9001C, northwestern Pacific. Paleontological Research 19 (Supplement): 1–77. Crossref
Moix, P., Kozur, H.M., Stampfli, G.M., and Mostler, H. 2007. New paleontological, biostratigraphic and paleogeographic results from the Triassic of the Mersin Melange, SE Turkey. New Mexico Museum of Natural History and Science, Bulletin 41: 282–311.
Nakaseko, K. and Nishimura, A. 1982. Upper Jurassic and Cretaceous Radiolaria from the Shimanto Group in Southwest Japan. Science Reports, College of General Education, Osaka University 30: 133–203.
Noble, P.J. and Jin, Y.X. 2010. Radiolarians from the Lamar Limestone, Guadalupe Mountains, West Texas. Micropaleontology 56: 117–147.
O’Connor, B. 1996. Confocal Laser Scanning Microscopy: a new technique for investigating and illustrating fossil Radiolaria. Micropaleontology 42: 395–402. Crossref
O’Dogherty, L., Carter, E.S., Dumitrica, P., Gorican, S., De Wever, P., Bandini, A.N., Baumgartner, P.O., and Matsuoka, A. 2009a. Catalogue of Mesozoic radiolarian genera. Part 2. Jurassic–Cretaceous. Geodiversitas 31: 271–356. Crossref
O’Dogherty, L., Carter, E.S., Dumitrica, P., Gorican, S., De Wever, P., Hunberbühler, A., Bandini, A.N., and Takemura, A. 2009b. Catalogue of Mesozoic radiolarian genera. Part 1: Triassic. Geodiversitas 31: 213–270. Crossref
Ogane, K. and Suzuki, N. 2006. Morphological terms describing discoidal radiolarians. Revue de Micropaléontologie 49: 97–100. Crossref
Ogane, K. and Suzuki, N. 2007. A sectioning method using nail polish for observation of the internal structure of disk-shaped polycystine radiolarians. Journal of Paleontology 81: 216–219. Crossref
Pessagno, E.A. Jr. 1973. Upper Cretaceous Spumellarina from the Great Valley sequence, California Coast Ranges. Bulletins of American Paleontology 63: 48–102.
Pessagno, E.A. Jr. and Poisson, A. 1981. Lower Jurassic Radiolaria from the Gümüslü allochthon of southwestern Turkey (Taurides occidentales). Bulletin of the Mineral Research and Exploration Institute of Turkey 9: 47–69.
Petrushevskaya, M.G. [Petruševskaâ, M.G.] 1984. Terminji, Yporebkâemye pri Opisanii Skeletov Radiolârji. 72 pp. Ministerstvo Geologii SSSR, Vsesoûznyj Ordena Lenina Naučno-Issledovatel’skii Geologičeskij Institut Imeni A.P. Karpinskogo, Leningrad.
Renz, G.W. 1974. Radiolaria from Leg 27 of the DSDP. In: J.J. Veevers, J.R. Heirtlzer, H.M. Bolli, A.N. Carter, P.J. Cook, V. Krasheninnikov, B.K. McKnight, F.P. Decima, G.W. Renz, P.T. Robinson, K. Rocker, Jr., and P.A. Thayer (eds.), Initial Reports of the Deep Sea Drilling Project, Volume 27, 769–841. U.S. Government Printing Office, Washington, D.C.
Rüst, D. 1888. Beiträge zur Kenntniss der fossilen Radiolarien aus Gesteinen der Kreide. Palaeontographica 34: 181–213.
Sano, H., Kuwahara, K., Yao, A., and Agematsu, S. 2010. Pantahalassan seamount-associated Permian–Triassic boundary siliceous rocks, Mino terrane, central Japan. Paleontological Research 14: 293–314. Crossref
Sashida, K. and Tonishi, K. 1985. Permian radiolarians from the Kanto Mountains, central Japan—some Upper Permian Spumellaria from Itsukaichi, western part of Tokyo Prefecture. Science Reports of the Institute of Geoscience, University of Tsukuba, Section B (Geological Sciences) 6: 1–19.
Sashida, K., Salyapongse, S., and Nakornsri, N. 2000. Latest Permian radiolarian fauna from Klaeng, eastern Thailand. Micropaleontology 46: 245–263. Crossref
Schaaf, A. 1981. Late Early Cretaceous radiolaria from Deep Sea Drilling Project Leg 62. In: J. Thiede, T.L. Vallier, C.G. Adelseck, A. Boersma, P. Čepek, W.E. Dean, N. Fujii, V.I. Koporulin, D.K. Rea, C. Sancetta, W.O. Sayre, K. Seifert, A. Schaaf, R.R. Schmidt, K. Windom, and E. Vincent (eds.), Initial Reports of the Deep Sea Drilling Project, Volume 62, 419–470. U.S. Government Printing Office, Washington, D.C. Crossref
Sugiyama, K. 1992. Lower and Middle Triassic radiolarians from Mt. Kinkazan, Gifu Prefecture, central Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New, Series 167: 1180–1223.
Sugiyama, K. 1997. Triassic and Lower Jurassic radiolarian biostratigraphy in the siliceous claystone and bedded chert units of the southeastern Mino Terrane, central Japan. Bulletin of the Mizunami Fossil Museum 24: 79–193.
Suzuki, N. 2006. Ontogenetic growth and variation in the skeletal structure of two late Neogene Sphaeropyle species (Polycystina radiolarians). Journal of Paleontology 80: 849–866. Crossref
Suzuki, N., Ogane, K., and Chiba, K. 2009. Middle to late Eocene polycystine radiolarians from the Site 1172, Leg 189, Southwest Pacific. News of Osaka Micropaleontologists, Special Volume 14: 239–296.
Tian, S.G. 1993a. Late Permian–Triassic conodont palaeoecology in northwestern Hunan [in Chinese]. Acta Palaeontologica Sinica 32: 332–348.
Tian, S.G. 1993b. The Permo-Triassic boundary and conodont zones in northwestern Hunan Province [in Chinese]. Bulletin of the Chinese Academy of Geological Sciences 26: 134–150.
Wakamatsu, H., Sugiyama, K., and Furutani, H. 1990. Silurian and Devonian radiolarians from the Kurosegawa Tectonic Zone, Southwest Japan. Journal of Earth Sciences, Nagoya University 37: 157–192.
Won, M.Z. and Iams, W.J. 2011. Earliest Arenig radiolarians from the Cow Head Group, western Newfoundland. Journal of Paleontology 85: 156–177. Crossref
Yao, A. and Kuwahara, K. 2000. Permian and Triassic radiolarians from the Guizhou Province, China. Journal of Geosciences, Osaka City University 43: 1–19.
Yue, M.L. 2014. Study of Sea Level Changes in the Late Permian to the Earliest Triassic in Sangzhi, Hunan Province [in Chinese]. 39 pp. Unpublished master dissertation, China University of Geosciences, Wuhan.
Yoshino, T., Matsuoka, A., Kurihara, T., Ishida, N., Kishimoto, N., Kimoto, K., and Matsuura, S. 2015. Polyhedron geometry of skeletons of Mesozoic radiolarian Pantanellium. Revue de Micropaléontologie 58: 51–56. Crossref
Zhang, Z.Y., He, W.H., Zhang, Y., Yang, T.L., and Wu, S.B. 2009. Late Permian–earliest Triassic ammonoid sequences from the Rencunping section, Sangzhi County, Hunan Province, South China and their regional correlation [in Chinese]. Geological Science and Technology Information 28: 23–30.
Acta Palaeontol. Pol. 62 (3): 647–656, 2017
https://doi.org/10.4202/app.00367.2017