
On a dinosaur axis from one of the oldest dinosaur-bearing sites worldwide
RODRIGO T. MÜLLER, FLÁVIO A. PRETTO, MICHELI STEFANELLO, EDUARDO SILVA-NEVES, and SÉRGIO DIAS-DA-SILVA
The axial skeleton is proportionally underrepresented in the fossil record of early dinosaurs, when compared to other skeletal parts (e.g., pelvic girdle and hindlimb). For instance, the axis is poorly known in early dinosaurs, which precludes a better understanding of this important anatomical structure. Therefore, the present contribution fills an important gap with a description of the axis of a new early dinosaur (CAPPA/UFSM 0179). The specimen was collected at the Buriol outcrop, a Triassic fossiliferous locality from southern Brazil (Candelária Sequence, Santa Maria Supersequence) biostratigraphically correlated to Carnian units, placing this specimen among the oldest dinosaurs worldwide. Notable features include the combination of a neural spine that bears an almost straight dorsal margin along its length and presence of an epipophysis. This axis arrangement is unique among Carnian dinosaurs, representing a new morphotype, though a similar morphology is observed in some early theropods. Indeed, a phylogenetic analysis nested the specimen within Theropoda. However, this outcome is probably biased by the large amount of missing data in CAPPA/UFSM 0179 and also due to the limited sampling of the axis in early dinosaurs, particularly among sauropodomorphs. As the specimen comes from the site that includes Buriolestes schultzi (an early sauropodomorph), it is quite plausible that CAPPA/UFSM 0179 might be referable to that taxon. If so, the specimen improves the anatomical knowledge of Buriolestes schultzi, given its axis is yet unknown. An alternative possibility to be considered is that the specimen would belong to a dinosaur not yet known in the Candelária Sequence, which would increase its dinosaur diversity for the outcrop, improving the Triassic dinosaurian record from Southern Brazil.
Introduction
The Buriol (Fig. 1A) and surrounding outcrops in the municipality of São João do Polêsine, Rio Grande do Sul, Brazil has yielded many fossil vertebrates. Its fossiliferous content includes fishes (Perez and Malabarba 2002), temnospondyls (Dias-da-Silva et al. 2011, 2012), aetosaurs and rhynchosaurs (Roberto-da-Silva et al. 2014), dinosauromorphs (Cabreira et al. 2016), and cynodonts (Pacheco et al. 2017). Its dinosauromorph fossil record is particularly interesting because it encompasses the oldest unequivocal members of Dinosauria, as well as exceptionally well preserved dinosaur relatives (Cabreira et al. 2016). Indeed, the presence of the hyperodapedontid rhynchosaurs supports a Carnian age (Martínez et al. 2011) for the outcrop. As widely known, so far Carnian strata record the oldest unambiguous dinosaurs worldwide (e.g., Sereno et al. 1993; Langer et al. 1999; Martínez and Alcober 2009). Dinosauromorph taxa from the Buriol outcrop include the lagerpetid Ixalerpeton polesinensis and the early sauropodomorph Buriolestes schultzi (Cabreira et al. 2016). These records have crucially enhanced the knowledge regarding both the anatomy and phylogenetic affinities of early dinosaurs and their close relatives. In spite of this, many skeletal parts of both taxa remain unknown. Indeed, this is the current condition of most Carnian dinosaurs, as a number of skeletal portions are far more represented than other ones in the fossil record. For instance, pelvic and/or hind limb elements are known from all described Carnian dinosaurs. In contrast, the axial skeleton is still poorly sampled, usually represented by a few isolated vertebrae (e.g., Chromogisaurus novasi, Pampadromaeus barberenai, Panphagia protos). The axis is poorly known in early dinosaurs, being preserved in a few Carnian specimens (e.g., Sereno and Novas 1993; Alcober and Martínez 2010; Sereno et al. 2013). The axis is an important anatomical structure as it provides the attachment site for muscles connecting the skull with the postcranial skeleton (e.g., Snively and Russell 2007). Hence, considering the scarce knowledge regarding this structure in early dinosaurs and their anatomical importance, herein we describe the anatomy of an early dinosaur axis from the abovementioned site, providing its comparison with some other Triassic dinosauromorphs.
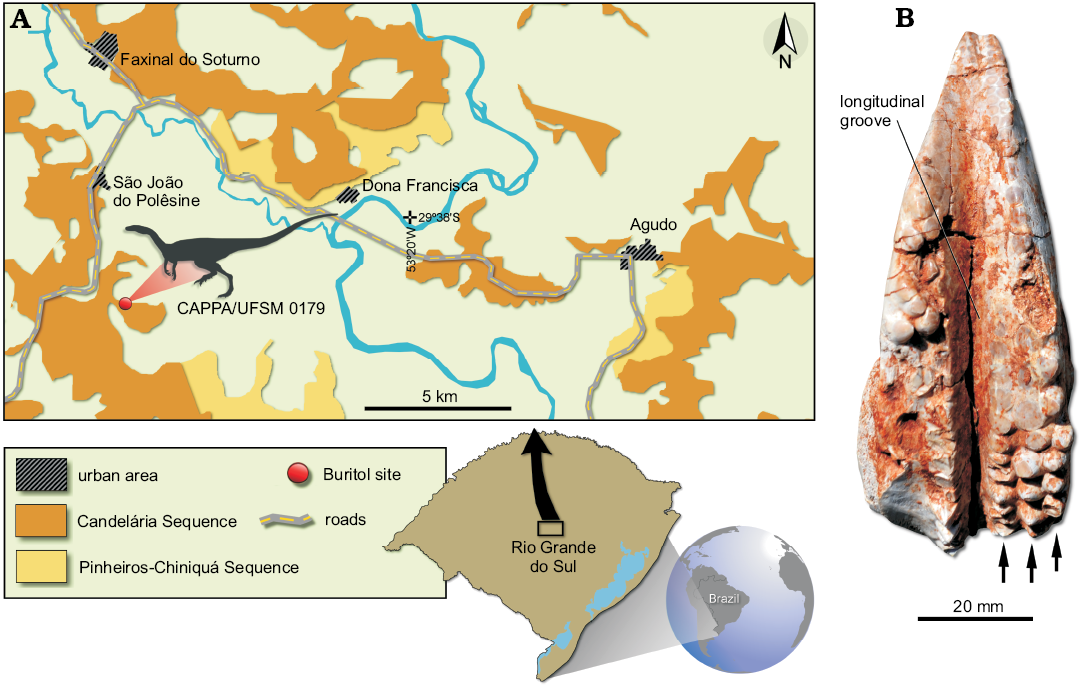
Fig. 1. A. Map of the São João do Polêsine área, Rio Grande do Sul, Brazil, showing the location of the Buriol outcrop (modified from Zerfass et al. 2007). B. CAPPA/UFSM 0044, a left maxilla of a Hyperodapedontinae from the Buriol ourcrop, Carnian, late Triassic, arrows point to the teeth rows.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; CAPPA/UFSM, Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia, São João do Polêsine, Rio Grande do Sul, Brazil; PULR, Universidad Nacional de La Rioja, La Rioja, Argentina; PVSJ, Instituto y Museo de Ciencias Naturales, San Juan, Argentina; SAM, Iziko South African Museum, Cape Town, South Africa; TMM, Texas Memorial Museum, Austin, USA; ULBRA, Universidade Luterana do Brasil, Coleção de Paleovertebrados, Canoas, Rio Grande do Sul, Brazil; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland.
Systematic palaeontology
Archosauria Cope, 1869
Dinosauromorpha Benton, 1985
Dinosauriformes Novas, 1992
Dinosauria Owen, 1842
Genus et species incertae sedis
Material.—CAPPA/UFSM 0179 (Fig. 2), an isolated and nearly complete axis including the caudal half of the centrum, the neural arch, and the neural spine. Bone surface is exceptionally well preserved. The specimen was recovered from the Buriol outcrop (Fig. 1A) (29°39’34.2” S; 53°25’47.4” W), in São João do Polêsine, Rio Grande do Sul, Brazil, in the same layer where the holotype (ULBRA-PVT280) of Buriolestes schultzi was excavated. This outcrop belongs to the Candelária Sequence (Horn et al. 2014), part of the Santa Maria Suspersequence of Zerfass et al. (2003). At least two rhynchosaur specimens collected at the site (Fig. 1B) may be referable to Hyperodapedontinae, and among these, more closely resemble Hyperodapedon sp. than other forms (Langer and Schultz 2000; Langer et al. 2000; Montefeltro et al. 2010; Schultz et al. 2016). They possess a characteristic longitudinal groove (Fig. 1B) between the lingual and buccal tooth rows. In both margins, the teeth cover a wide area, with as much as three rows of teeth that are more significantly worn towards the cranial portion of the series (Fig. 1B). Therefore, the fossil vertebrate content from this locality has been correlated with those from the Ischigualasto Formation, northwest Argentina (Langer et al. 2007), radioisotopically dated as 231.4 ± 0.3 Ma (Martínez et al. 2011).
Description.—The preserved portion of the axial centrum is 12.5 mm in length. It is constricted towards the mid-point (Fig. 2D), with 4 mm in width, while the caudal articular facet expands to 8.5 mm in width. This suggests that the centrum was spool-shaped. The dorsal surface of the centrum is excavated to form the floor of the neural canal, which also is transversely larger at the caudal extremity than in the mid-point. The lateral surface lacks any pneumatic features. The ventral surface bears a 1.85 mm wide longitudinal ventral keel that reaches the ventral margin of the caudal articular surface (Fig. 2A). A ventral keel is absent in some plateosaurian sauropodomorphs (e.g., Plateosaurus, Riojasaurus) but present in early dinosaurs and related groups (e.g., Herrerasaurus, Lewisuchus) (Yates 2003). The ventral extension of the keel is uncertain as the centrum is cranially fractured. The caudal articular surface is 8.8 mm in height and markedly concave (Fig. 2F). In lateral view, the ventral margin of this surface slightly extends more caudally than the dorsal margin (Fig. 2A, B). At the mid-point of the centrum, the bone wall is approximately 0.8 mm in width.
The preserved portion of the neural arch is completely fused to the centrum, and there is no sign of a neurocentral suture. The dorsoventral height of the neural canal is approximately 3.5 mm, whereas the space between the dorsal tip of the postzygapophysis and the ventral margin of the neural canal is 7.6 mm. The lateral surface, ventral to the cranial margin of the postzygapophysis, is gently excavated. The postzygapophysis extends laterally to form a convex margin in dorsal view (Fig. 2C). On the ventral surface of the postzygapophysis there is a subovoid facet with 6 mm in length, which would receive the dorsomedial oriented facet of the prezygapophysis of the third cervical vertebra. Indeed, the articular facet of the postzygapophysis is ventromedially oriented (Fig. 2F). The medial margin of such structure folds ventrally to form a faint hyposphene (Fig. 2F), an accessory articulation. Its caudal tip extends caudally as far the caudal margin of the dorsal portion of the centrum. The caudal extension of the postzygapophysis reaches the caudal extension of the ventral margin of the centrum, as in Silesaurus opolensis (ZPAL AbIII/1930; Piechowski and Dzik 2010), whereas in Herrerasaurus ischigualastensis (PVSJ 407; Sereno and Novas 1993) the postzygapophysis projects more caudally than the caudal articular facet of the axial centrum. On the dorsal surface of the postzygapophysis there is a caudodorsal process that exceeds its caudal extension (Fig. 2B, F). Its structure corresponds to the epipophysis but the caudal tip of both epipophyses is not preserved. Nonetheless, this corresponds to the maximum caudal extension of the axis. Some dinosauriforms lack axial epipophyses, such as, Silesaurus opolensis (ZPAL AbIII/1930), and Lewisuchus admixtus (PULR 01). A cleft separates the dorsal surface of the epipophysis from the caudal margin of the neural spine (Fig. 2A).
The 21.4 mm long neural spine is entirely preserved (Fig. 2A, B). The structure is 1.4 mm in width at the mid-point. Its height is constant along its length. Therefore, as in Lewisuchus admixtus (PULR 01) and Coelophysis bauri (AMNH FR 7224), the dorsal margin of the cranial half and the caudal half runs almost in the same plane, resulting in a straight margin, excepting the cranial and caudal extremities that are gently curved ventrally. In contrast, the dorsal margin of the neural spine is caudodorsally angled in most archosauriforms (Nesbitt 2011). The laterodorsal margin is densely covered by striations (Fig. 2A, B) which could be related to the origin of the m. splenius capitis (e.g., Snively and Russell 2007), whereas ventrally, close to the dorsal surface of the postzygapophysis, a depressed surface could be related to the attachment of the m. complexus. The cranial tip of the spine tappers to a point (Fig. 2A, B), whereas the caudal extremity of the spine bifurcates to form a Y-shaped caudal portion in dorsal view (Fig. 2C). This condition resembles that from Herrerasaurus ischigualastensis (PVSJ 407), whereas in some dinosauriforms, such as Silesaurus opolensis (ZPAL AbIII/1930; Piechowski and Dzik 2010) and Lewisuchus admixtus (PULR 01; Bittencourt et al. 2014), the caudodorsal tip of the neural spine projects further caudally, precluding the bifurcation and forming the roof of the postspinal chonos. In addition, distinct from the caudally extended neural spine of Heterodontosaurus tucki (e.g., SAM-PK-K1332), the caudal margin of the neural spine only slightly exceeds the caudal margin of the axial centrum in CAPPA/UFSM 0179. On the other hand, the neural spine is caudally short in some sauropodomorphs, such as Adeopapposaurus mognai (PVSJ568) and Sarahsaurus aurifontanalis (TMM 43646-2).
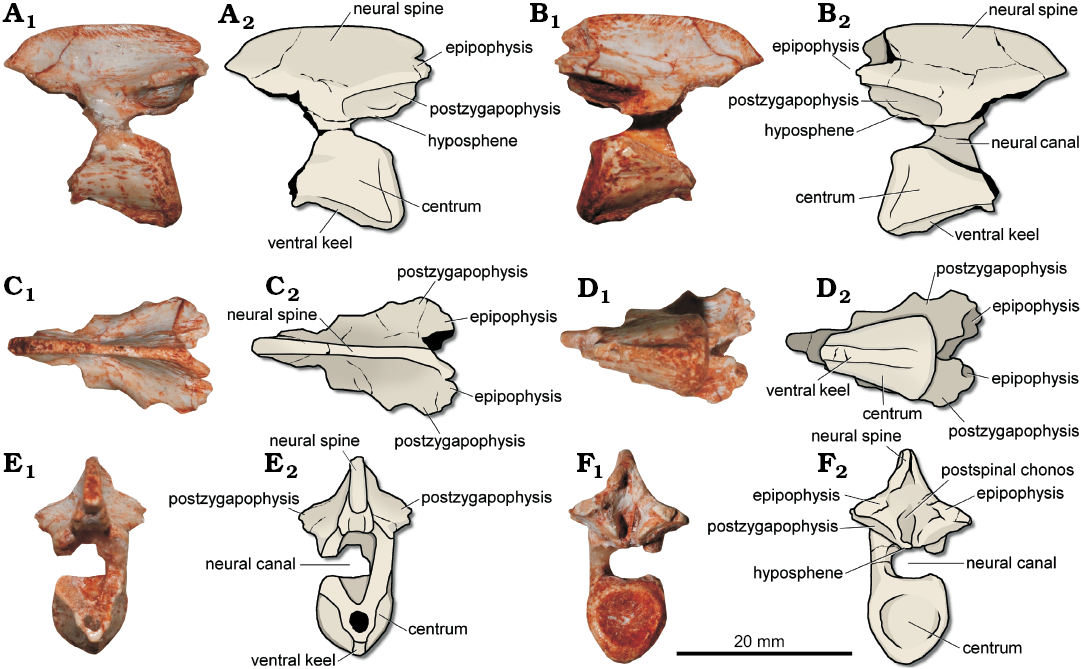
Fig. 2. Dinosauria gen. et sp. indet., CAPPA/UFSM 0179 from the Buriol outcrop, São João do Polêsine, Rio Grande do Sul, Brazil; Carnian, late Triassic. Photographs (A1–F1) and schematic drawings (A2–F2) of the axis in left (A) and right (B) lateral, dorsal (C), ventral (D), cranial (E), and caudal (F) views.
Phylogenetic analysis
CAPPA/UFSM 0179 was scored in the data matrix of Cabreira et al. (2016), in order to test its phylogenetic affinities (Appendix 1). The data matrix totalizes 256 morphological characters and 44 operational taxonomic units. It was processed with the software TNT v1.1 (Goloboff et al. 2008). All characters received the same weight and 31 (3, 4, 6, 11, 36, 60, 62, 64, 83, 115, 123, 139, 147, 148, 157, 160, 171, 173, 175, 178, 179, 182, 195, 200, 201, 202, 205, 216, 222, 240, and 248) were treated as ordered following the study of Cabreira et al. (2016). The most parsimonious trees (MPTs) were recovered via “traditional search” (random addition sequence + tree bisection reconnection) with 1000 replicates of Wagner trees (with random seed = 0), tree bisection reconnection (TBR) and branch swapping (holding 20 trees save per replicate).
The analysis recovered 242 MPTs of 846 steps (CI = 0.345; RI = 0.637). CAPPA/UFSM 0179 nests as an early diverging theropod in all trees. In the strict consensus tree (Fig. 3), the specimen is placed in a polytomy at the base of Theropoda, together with Dilophosaurus wetherlli, Zupaysaurus rugeiri, and the “Petrified Forest theropod”. The only character state supporting CAPPA/UFSM 0179 as theropod is the dorsal margin of the axial neural spine that arcs dorsally, where the cranial portion height is equivalent to the caudal height (character 76, state 1). However, this character is not scored to other early diverging sauropodomorphs (e.g., Buriolestes schultzi, Eoraptor lunensis, Pampadromaeus barberenai). In addition, Lewisuchus admixtus presents the same condition of CAPPA/UFSM 0179. Therefore, this represents an ambiguous synapomorphy of theropods. So, the phylogenetic position of CAPPA/UFSM 0179 remains uncertain due to both the limited entries in the data matrix and the lack of the axis in other early diverging sauropodomorphs.
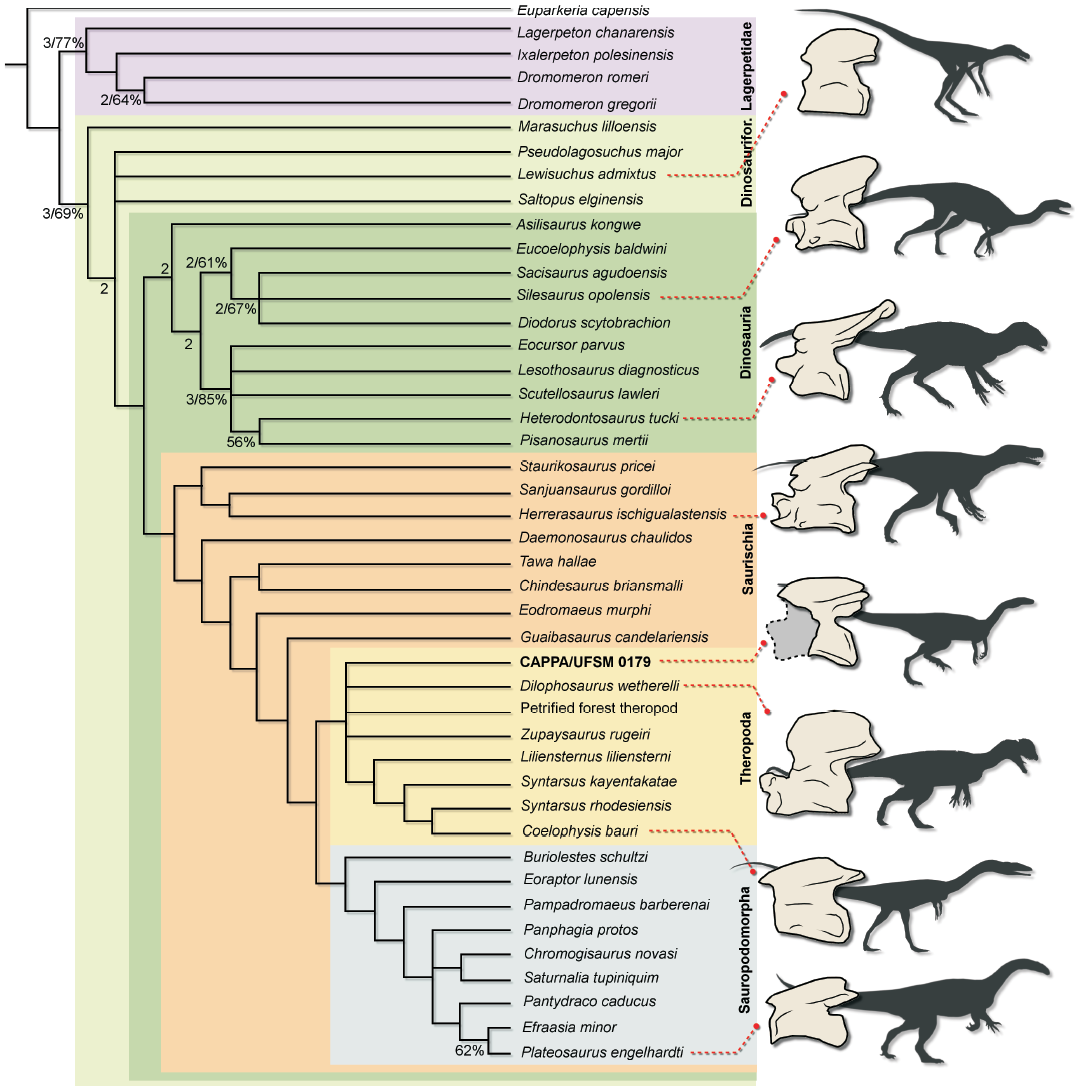
Fig. 3. Strict consensus tree depicting the phylogenetic position of CAPPA/UFSM 0179 and the schematic general morphology of the axis of some Triassic dinosauriforms. Numbers below nodes represent Bremer support values (left) higher than 1 and Bootstrap values (right) higher than 50%. Abbreviation: Dinosaurifor., Dinosauriformes.
Discussion and conclusions
The incomplete and fragmentary condition of the specimen prevents a reliable taxonomic ascription. Nevertheless, CAPPA/UFSM 0179 supports a dinosaurian assignation due to its combination of features. As observed by Nesbitt (2011), the axial neural spine of most archosauriforms is cranioventrally slanted, with the caudodorsal corner much taller than the craniodorsal corner. In contrast, the inverse condition occurs in some early diapsids like Araeoscelis gracilis and Petrolacosaurus kasensis (Nesbitt et al. 2015). In addition, Nesbitt (2011) also observed that the craniodorsal height is equivalent to the caudodorsal height in the axial neural spine of some ornithodirans, as in CAPPA/UFSM 0179. Among ornithodirans that share this condition, only dinosaurs bear an epipophysis on the axis, as it is absent in Lewisuchus admixtus (Bittencourt et al. 2014). Assignation of CAPPA/UFSM 0179 to a less inclusive group within Ornithodira is currently precluded, as it lacks diagnostic information in order to do so. As already stated, the theropod affinities expressed in the phylogenetic analysis could reflect the poor sampling of axis among early dinosaurs and relatives, and the large amount of missing data for CAPPA/UFSM 0179. Indeed, despite the sauropodomorph affinities of Buriolestes schultzi, it shares some particular traits with theropods (see Cabreira et al. 2016). So, it is plausible that CAPPA/UFSM 0179 could represent an axis from this species, as both CAPPA/UFSM 0179 and the holotype (ULBRA-PVT280) of Buriolestes schultzi were excavated in the exact same layer of the outcrop. Moreover, the hypothesis that CAPPA/UFSM 0179 could belong to the lagerpetid species Ixalerpeton polesinensis (ULBRA-PVT059, also from the same outcrop), is not plausible, as ULBRA-PVT059 lacks epipophyses on the preserved cervical vertebrae.
The new specimen increases the fossil record of dinosaurs from the Candelária Sequence and announces a new axial morphotype for Carnian dinosaurs, previously known only for younger taxa. The two most plausible hypotheses and implications related to the taxonomic ascription of CAPPA/UFSM 0179 are: (i) it belongs to Buriolestes schultzi; or (ii) it belongs to an as yet unknown dinosaur from the Buriol outcrop. In the first hypothesis, CAPPA/UFSM 0179 increases the anatomical knowledge related to Buriolestes schultzi, as the axis is currently unknown to the taxon. In the second hypothesis, the specimen increases the early dinosaur diversity for the outcrop, contributing to a better knowledge of the Carnian dinosaurian fauna in South America. As a putative new, still undescribed specimen (CAPPA/UFSM 0035) of Buriolestes schultzi was recently discovered, the possibility to assign CAPPA/UFSM 0179 to this taxon will be addressed in detail. At this point, it is possible to observe that both elements present an overall morphological similarity. However, the axis of CAPPA/UFSM 0035 is 12% smaller than CAPPA/UFSM 0179 and the caudal half of the dorsal margin of the neural spine of CAPPA/UFSM 0035 is broken. Therefore, it is uncertain if Buriolestes schultzi possesses the same straight margin of CAPPA/UFSM 0179. Concluding, only the discovery of a more complete specimen with the same morphology of CAPPA/UFSM 0179 would solve if it belongs to Buriolestes schultzi or to a still unknown new early dinosaur in South America.
Acknowledgements.—We are grateful to the Buriol family (São João do Polêsine, Rio Grande do Sul, Brazil) for allowing access to their property to excavate the specimen herein described. The comments and suggestions of Matthew G. Baron (Department of Earth Sciences, University of Cambridge, UK) and an anonymous reviewer greatly improved the quality of this manuscript. We also thank the Coordenação de Aperfeiçoamento de Pessoal de Nivel Superior (CAPES) for the scholarship to RTM, ESN, and ME; the Conselho Nacional de Desenvolvimento Científco e Tecnológico (CNPq) for the research grant to SDS (process 306352/2016-8). We thank the Willi Henning Society, for the gratuity of TNT software.
References
Alcober, O.A. and Martínez, R.N. 2010. A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina. ZooKeys 63: 55–81. Crossref
Benton, M.J. 1985. Classification and phylogeny of diapsid reptiles. Zoological Journal of the Linnean Society 84: 97–164. Crossref
Bittencourt, J.S., Arcucci, A.B., Maricano, C.A., and Langer, M.C. 2014. Osteology of the Middle Triassic archosaur Lewisuchus admixtus Romer (Chañares Formation, Argentina), its inclusivity, and relationships amongst early dinosauromorphs. Journal of Systematic Palaeontology 13: 189–219. Crossref
Cabreira, S.F., Kellner, A.W.A., Dias-da-Silva, S., Roberto-da-Silva, L., Bronzati, M., Marsola, J.C., Müller, R.T., Bittencourt, J.S., Batista, B.J., Raugust. T., Carrilho, R., and Langer, M.C. 2016. A unique Late Triassic dinosauromorph assemblage reveals dinosaur ancestral anatomy and diet. Current Biology 26: 3090–3095. Crossref
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia, and Aves of North America. Transactions of the American Philosophical Society (New Series) 14: 1–252. Crossref
Dias-da-Silva, S., Cabreira, S.F., and Roberto-da-Silva, L. 2011. Occurrence of giant stereospondyl remains in the Santa Maria Formation (Middle⁄Upper Triassic of southern Brazil). Alcheringa 35: 11–19. Crossref
Dias-da-Silva, S., Sengupta, D.P., Cabreira, S.F., and Roberto-da-Silva, L. 2012. The presence of Compsocerops (Brachyopoidea: Chigutisauridae) (Late Triassic) in southern Brazil with comments on chigutisaurid palaeobiogeography. Paleontology 55: 163–172. Crossref
Goloboff, P.A., Farris, J.S., Nixon, and K.C.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Horn, B.L.D., Melo, T.M., Schultz, C.L., Philipp, R.P., Kloss, H.P., and Goldberg, K. 2014. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. Journal of South American Earth Sciences 55: 123–132. Crossref
Langer, M.C. and Schultz, C.L. 2000. A new species of the Late Triassic rhynchosaur Hyperodapedon from the Santa Maria Formation of South Brazil. Palaeontology 43: 633–652. Crossref
Langer, M.C., Abdala, F., Richter, M., and Benton, M.J. 1999. A sauropodomorph dinosaur from the Upper Triassic (Carnian) of Southern Brazil. Comptes Rendus de l’Academie des Sciences IIA 329: 511–517.
Langer, M.C., Boniface, M., Cuny, G., and Barbieri, L. 2000. The phylogenetic position of Isalorhynchus genovefae, a Late Triassic rhynchosaur from Madagascar. Annales de Paléontologie 86: 101–127. Crossref
Langer, M.C., Ribeiro, A.M., Schultz, C.L., and Ferigolo, J. 2007. The continental tetrapod-bearing Triassic of South Brazil. Bulletin of the American Museum of Natural History 41: 201–218.
Martínez, R.N. and Alcober, O.A. 2009. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS One 4 (2): e4397. Crossref
Martínez, R.N., Sereno, P.C., Alcober, O.A., Colombi, C.E., Renne, P.R., Montañez, I.P., and Currie, B.S. 2011. A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science 331: 201–210. Crossref
Montefeltro, F.C., Langer, M.C., and Schultz, C.L. 2010. Cranial anatomy of a new genus of hyperodapedontine rhynchosaur (Diapsida, Archosauromorpha) from the Upper Triassic of southern Brazil. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 27–52. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: Relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Nesbitt, S.J., Flynn, J.J., Pritchard, A.C., Parrish, J.M., Ranivoharimanana, L., and Wyss, A.R. 2015. Postcranial anatomy and relationships of Azendohsaurus madagaskarensis. Bulletin of the American Museum of Natural History 398: 1–126. Crossref
Novas, F.E. 1992. Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae. Palaeontology 35: 51–62.
Owen, R. 1842. Report on British fossil reptiles. Part II. Reports of the British Association for the Advancement of Science 11: 60–204.
Pacheco, C.P., Martinelli, A.G., Pavanatto, A.E., Soares, M.B., and Dias-da-Silva, S. 2017. Prozostrodon brasiliensis, a probainognathian cynodont from the Late Triassic of Brazil: second record and improvements on its dental anatomy. Historical Biology [published online]. Crossref
Perez, P.A. and Malabarba, M.C. 2002. A Triassic freshwater fish fauna from the Paraná Basin in southern Brazil. Revista Brasileira de Paleontologia 4: 27–33.
Piechowski, R. and Dzik, J. 2010. The axial skeleton of Silesaurus opolensis. Journal of Vertebrate Paleontology 30: 1127–1141. Crossref
Roberto-da-Silva, L., Desojo, J.B., Cabreira, S.F., Aires, A.S.S., Müller, R.T., Pacheco, C.P., and Dias-da-Silva, S. 2014. A new aetosaur from the Upper Triassic of the Santa Maria Formation, southern Brasil. Zootaxa 3764: 240–278. Crossref
Schultz, C.L., Langer, M.C., and Montefeltro, F.C. 2016. A new rhynchosaur from south Brazil (Santa Maria Formation) and rhynchosaur diversity patterns across the Middle–Late Triassic boundary. Paläontologische Zeitschrift 90: 593–609. Crossref
Sereno, P.C. and Novas, F.E. 1993. The skull and neck of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology 13: 451–476. Crossref
Sereno, P.C., Forster, C.A., Rogers, R.R., and Monetta, A.M. 1993. Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria. Nature 361: 64–66. Crossref
Sereno, P.C., Martínez, R.N., and Alcober, O.A. 2013. Osteology of Eoraptor lunensis (Dinosauria, Sauropodomorpha). Journal of Vertebrate Paleontology 32: 83–179. Crossref
Snively, E. and Russell, A. P. 2007. Functional variation of neck muscles and their relation to feeding style in Tyrannosauridae and other large theropod dinosaurs. The Anatomical Record 290: 934–957. Crossref
Yates, A.M. 2003. A new species of the primitive dinosaur, Tecodontosaurus (Saurischia: Sauropodomorpha) and its implications for the systematics of early dinosaurs. Journal of Systematic Palaeontology 1: 1–42. Crossref
Zerfass, H., Lavina, E.L., Schultz, C.L., Garcia, A.J.V., Faccini, U.F., and Chemale, F. 2003. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: a contribution to Southwestern Gondwana palaeogeography and palaeoclimate. Sedimentary Geology 161: 85–105. Crossref
Zerfass, H., Sander, A., and Flores, A.F. 2007. Agudo, Folha SH.22-V.C.V, escala 1:100.000, Rio Grande do Sul. Serviço Geológico do Brasil (CPRM), Brasília.
Rodrigo Temp Müller [rodrigotmuller@hotmail.com], Micheli Stefanello [michelistefanello@hotmail.com], and Eduardo Silva-Neves [edusneves90@gmail.com], Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria, Av. Roraima, 1000, Bairro Camobi, 97105-900 Santa Maria, RS, Brasil.
Flávio Augusto Pretto [flavio_pretto@yahoo.com.br] and Sérgio Dias-da-Silva [paleosp@gmail.com], Centro de Apoio à Pesquisa da Quarta Colônia, Universidade Federal de Santa Maria, Rua Maximiliano Vizzotto, 598, 97230-000 São João do Polêsine, RS, Brasil.
Received 4 April 2017, accepted 11 May 2017, available online 12 July 2017.
Copyright © 2017 R.T. Müller et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Codification for CAPPA/UFSM 0179 of the 256 characters of Cabreira et al. (2016).
???????????????????????????????????????????????????????????????????????????
1?1????1???????????????????????????????????????????????????????????????????
???????????????????????????????????????????????????????????????????????????
???????????????????????????????
Acta Palaeontol. Pol. 62 (3): 543–548, 2017
https://doi.org/10.4202/app.00369.2017