

The sauropodomorph biostratigraphy of the Elliot Formation of southern Africa: Tracking the evolution of Sauropodomorpha across the Triassic–Jurassic boundary
BLAIR W. MCPHEE, EMESE M. BORDY, LARA SCISCIO, and JONAH N. CHOINIERE
McPhee, B.W., Bordy, E.M., Sciscio, L., and Choiniere, J.N. 2017. The sauropodomorph biostratigraphy of the Elliot Formation of southern Africa: Tracking the evolution of Sauropodomorpha across the Triassic–Jurassic boundary. Acta Palaeontologica Polonica 62 (3): 441–465.
The latest Triassic is notable for coinciding with the dramatic decline of many previously dominant groups, followed by the rapid radiation of Dinosauria in the Early Jurassic. Among the most common terrestrial vertebrates from this time, sauropodomorph dinosaurs provide an important insight into the changing dynamics of the biota across the Triassic–Jurassic boundary. The Elliot Formation of South Africa and Lesotho preserves the richest assemblage of sauropodomorphs known from this age, and is a key index assemblage for biostratigraphic correlations with other similarly-aged global terrestrial deposits. Past assessments of Elliot Formation biostratigraphy were hampered by an overly simplistic biozonation scheme which divided it into a lower “Euskelosaurus” Range Zone and an upper Massospondylus Range Zone. Here we revise the zonation of the Elliot Formation by: (i) synthesizing the last three decades’ worth of fossil discoveries, taxonomic revision, and lithostratigraphic investigation; and (ii) systematically reappraising the stratigraphic provenance of important fossil locations. We then use our revised stratigraphic information in conjunction with phylogenetic character data to assess morphological disparity between Late Triassic and Early Jurassic sauropodomorph taxa. Our results demonstrate that the Early Jurassic upper Elliot Formation is considerably more taxonomically and morphologically diverse than previously thought. In contrast, the sauropodomorph fauna of the Late Triassic lower Elliot Formation remains relatively poorly understood due to the pervasive incompleteness of many key specimens, as well as the relative homogeneity of their diagnostic character suites. Our metrics indicate that both Elliot Formation and global sauropodomorph assemblages had greater morphological disparity within the Early Jurassic than the Late Triassic. This result is discussed in the context of changing palaeoclimatic conditions, as well as macroevolutionary events associated with the end-Triassic extinction.
Key words: Dinosauria, Sauropodomorpha, disparity, Triassic–Jurassic boundary, South Africa.
Blair W. McPhee [blair.mcphee@gmail.com], Departamento de Biologia, FFCLRP, Universidade de São Paulo, Ribeirão Preto, São Paulo, 14040-901, Brazil; and Evolutionary Studies Institute, University of the Witwatersrand, Private Bag 3, Johannesburg, Gauteng, 2050, South Africa.
Emese M. Bordy [emese.bordy@uct.ac.za] and Lara Sciscio [lara.sciscio@uct.ac.za], Department of Geological Sciences, University of Cape Town, Western Cape, 7701, South Africa.
Jonah N. Choiniere [Jonah.Choiniere@wits.ac.za], Evolutionary Studies Institute, University of the Witwatersrand, Private Bag 3, Johannesburg, Gauteng, 2050, South Africa.
Received 27 April 2017, accepted 19 June 2017, available online 24 August 2017.
Copyright © 2017 B.W. McPhee et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The transition from the Triassic into the Jurassic was a critical period in the history of life on Earth. Representing a faunal decline of 50–80% of known species across the combined marine and terrestrial realms (Raup and Sepkoski 1982; Benton 1995; Hallam and Wignall 1999; Pálfy et al. 2000; Schaltegger et al. 2008; McGhee et al. 2013), the end-Triassic extinction event (ETE: 201.6 Ma, Blackburn et al. 2013) is equally remarkable for prefacing the rapid diversification of the dinosaurs during the Early Jurassic. Our understanding of the terrestrial effects of the ETE is restricted to a handful of continental formations that are dated (confidently or otherwise) to either side of this boundary (e.g., Newark-Hartford astrochronostratigraphic polarity time scale, Olsen et al. 2011; Lucas et al. 2005; Martinez et al. 2015). Although well-studied strata such as those of the Newark Supergroup from the Newark and Hartford basins of eastern North America preserve fine-scale depositional successions across the Triassic–Jurassic boundary (TJB), body fossils in these areas are either extremely rare (Olsen et al. 2011) or low in generic diversity (Sues and Olsen 2015) (see also the Moenave Formation, Glen Canyon Group, Lucas et al. 2011). The Elliot Formation, with a temporal range spanning the late Norian–?Sinemurian, is one of the few continental formations to preserve both successive phases of deposition across the ETE and highly fossiliferous rocks (Olsen and Galton 1984; Smith and Kitching 1997; Lucas and Hancox 2001; Knoll 2004, 2005). These strata, primarily accessible in South Africa and Lesotho (Fig. 1), preserve a broad range of vertebrate taxa, and are particularly abundant in fossils of sauropodomorph dinosaurs.
Given both the importance of the Elliot Formation for understanding faunal change across the ETE and the ubiquity of sauropodomorph fossils within this formation, a systematic account of this most common faunal constituent is long overdue. Although the Elliot faunal assemblage has already been the subject of a focused biostratigraphy (Kitching and Raath 1984) and a faunal revision (Knoll 2004, 2005), fossil sampling and taxonomic revision over the past three decades (and particularly in the last decade) has dramatically altered our understanding of the temporal and phylogenetic relationships of the sauropodomorph component of this assemblage (e.g., Yates 2003a, 2007a, b; Yates et al. 2010; McPhee et al. 2014, 2015a, b). Sauropodomorphs are worldwide biostratigraphic indicators in the latest Triassic and Early Jurassic (e.g., Young 1941, 1942; Sertich and Loewen 2010; Apaldetti et al. 2011; Martinez et al. 2015), and a systematic account of the stratigraphic relationships of Elliot Formation Sauropodomorpha has direct bearing on the faunal and temporal correlation of similarly aged global deposits. Here we synthesize the last 30 years of palaeontological, lithostratigraphic, and sedimentological investigations of the Late Triassic–Early Jurassic Elliot Formation, and present new information from our ongoing fieldwork programme with a goal of defining a revised, accurate and precise biostratigraphy of its sauropodomorph fauna. We follow this with an exploration of changing patterns of sauropodomorph morphological and taxonomic diversity across the TJB.
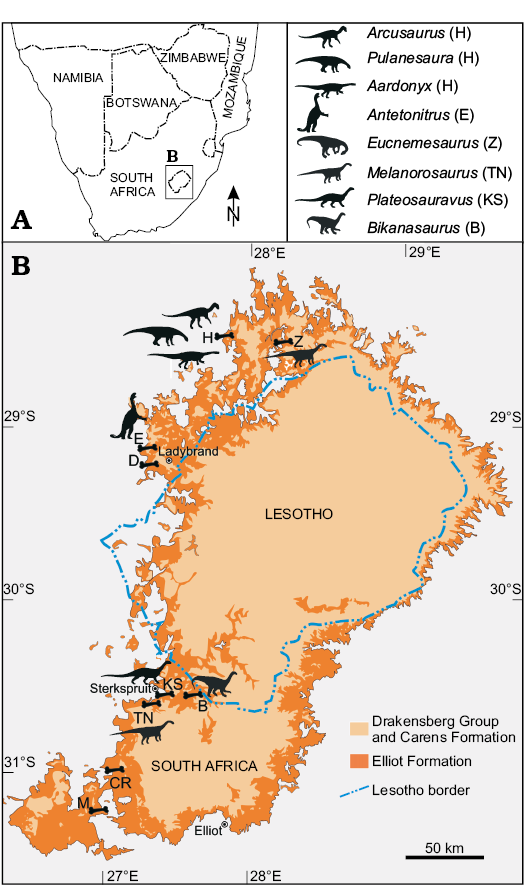
Fig. 1. Overview geological map of the Elliot Formation showing the spatial distribution of the fossil localities discussed in the text. Fossil locality abbreviations: B, Blikana; CR, Cannon Rock Farm; D, Damplaats 55 Farm; E, Edelweiss 698 Farm; H, Heelbo (Spion Kop 932) Farm; KS, Kromme Spruit; M, Milner Farm; TN, Thaba ’Nyama; Z, Zonderhout 291 Farm.
Institutional abbreviations.—AM, Albany Museum, Grahamstown, South Arica; BP, Evolutionary Studies Institute (previously the Bernard Price Institute), University of the Witwatersrand, Johannesburg, South Africa; NMQR, National Museum, Bloemfontein, South Africa; SAM-PK, Iziko-South African Museum, Cape Town, South Africa; TM, Ditsong Museum of Natural History, Pretoria, South Africa (previously the Transvaal Museum).
Other abbreviations.—ETE, end-Triassic extinction event; GER, Gap Excess Ratio; LEF, lower Elliot Formation; MIG, Minimum Implied Gaps; MSM, Manhattan Stratigraphic Measure; SCI, Stratigraphic Consistency Index; TJB, Triassic–Jurassic boundary; UEF, upper Elliot Formation.
Geological setting
The fluvio-lacustrine Elliot Formation is an Upper Triassic (lower Elliot Formation, LEF) to Lower Jurassic (upper Elliot Formation, UEF) succession that crops out around the Drakensberg Plateau (Fig. 1), with substantial exposures in the western side of the Lesotho/Free State border. It thickens generally to the south, with its thickest point (~460 m) in the Eastern Cape (Barkly Pass, near Elliot) and thinnest point (<30 m) in the north eastern Free State Province of South Africa (Fig. 1; Bordy and Eriksson 2015). Together with the underlying Molteno and overlying Clarens formations, it forms part of the Stormberg Group and represents the final depositional stages of the Karoo Supergroup in the main Karoo Basin (Haughton 1924; Catuneanu et al. 1998; Bordy et al. 2004a, b, 2005).
Traditionally, the lower part of the Elliot Formation was thought to be transitional with that of the underlying Molteno Formation, with the lowermost deposits of the Elliot accordingly referred to as the “Passage Beds” (Charig et al. 1965; Cole 1992; Anderson et al. 1998). However, more recent investigation into the nature of the contact between the Molteno and LEF suggests that the boundary is unconformable (Bordy et al. 2005). Although some lithological distinctiveness had long been recognised between various “red bed” outcrops throughout the basin (Haughton 1924), a formalized stratigraphic subdivision was only introduced by Kitching and Raath in 1984 (and later embellished by Smith and Kitching 1997), based primarily on differential faunal compositions between the lower, “middle”, and upper sections of the formation (see below). More recently, a detailed lithostratigraphic framework was established, based on a regional sedimentary facies contrast that subdivides the unit into the LEF and UEF throughout the basin (Bordy et al. 2004a, b; Bordy and Eriksson 2015). This boundary has been interpreted as an unconformity (Bordy et al. 2004a, b; 2005) and coincides with the contact of the lower and “middle” Elliot Formation of Kitching and Raath (1984).
Lithologically, the LEF is generally characterised by olive-grey to bluish and purplish mudstone units upwards of ~30 m thick that are interbedded with sandstone units of maximum of ~20 m thick. The latter have geometries evincing multi-storey, asymmetrical channel-fills with well-developed lateral accretion surfaces and irregular, erosive basal bounding surfaces. The depositional environment of the LEF is interpreted as one of perennial meandering river systems with extensive floodplains and overbank areas with riparian forests in a humid to semi-arid climatic setting (Bordy et al. 2004a, 2005). The LEF, while upwards of 300 m thick in the south of the basin, thins dramatically within the northern outcrop area, potentially due to a strengthening of the underlying lithosphere wherein it parallels the southern edge of the Kaapvaal Craton (Bordy et al. 2004a).
Following the stratigraphic gap at the LEF–UEF contact proposed by Bordy et al. (2004a), during which time the area embodied by the Elliot Formation became elevated and erosion took place, the UEF documents a markedly different fluvial environment than the LEF, and is characterised by mostly tabular, multi-storey sheet sandstone that are appreciably finer and thinner than those contained within the latter. The lack of significant down-cutting at the base of the sandstone units and extensive pedogenic modification of the UEF mudstone suggest an ephemeral, flash flood-dominated fluvial system whereby lower rates of sediment accumulation enabled more significant pedogenic alterations of the floodplain areas (Bordy et al. 2004b). This is consistent with the the progressive aridification of the Stormberg Group generally, with aeolian influences becoming increasingly dominant and culminating in the overlying wet sand-dune systems of the Clarens Formation. In contrast to the relatively heterogeneous olive-grey through purple mudstone units diagnostic of the LEF, UEF mudstone are more uniformly maroon to brick-red.
Due to the lack of absolute age determinations, dating of the Elliot Formation has generally relied on faunal correlations with other global Late Triassic–Early Jurassic deposits. Hence, the assemblage of large “prosauropods”, cynodonts, putative “rauisuchians”, and ichnofossils within the LEF are generally thought to correlate with later Norian or Rhaetian deposits elsewhere in the world (e.g., the Los Colorados Formation, Argentina; Trossingen Formation, Germany; e.g., Olsen and Galton 1984; Lucas and Hancox 2001; Knoll 2004). In contrast, a rich fauna of gracile “prosauropods”, heterodontosaurid, and “fabrosaurid” basal ornithischian dinosaurs, cynodonts, basal crocodylomorphs, basal turtles, and at least two species of theropod dinosaur suggests a Hettangian–Sinemurian age for the UEF based upon faunal congruence with other early Jurassic deposits (e.g., the Glen Canyon Group of the south-western USA and the Lower Lufeng Formation of China; Olsen and Galton 1984; Smith and Kitching 1997; Yates 2005; Knoll 2005; Butler 2005; Irmis 2011; McPhee et al. 2015a).
More recently, magnetostratigraphic investigation (Sciscio et al. 2017) of the Elliot Formation suggests that the TJB potentially lies within the lowermost UEF. These magnetostratigraphic results give an estimated age range for the formation that is tentatively constrained to ~213–190 Ma (Sciscio et al. 2017). Further refinement of this work would benefit from numeric ages drawn from several newly discovered volcanogenic layers within the Elliot Formation (Choiniere et al. 2014).
Historical background
Owen (1854) named the first sauropodomorph (and dinosaur), Massospondylus, from the Elliot Formation, followed shortly after by Huxley’s (1866) erection of “Euskelosaurus”. Throughout the following century these two taxa were central to discussions of Elliot Formation biostratigraphy, with Haughton (1924: 474; see also Ellenberger et al. 1969; Ellenberger 1970) the first to observe the tendency for the lower part of the Elliot Formation to preserve large-bodied “Euskelosaurus”-type forms, whereas the upper sections were typified by smaller-bodied, gracile massospondylids and “thecodontosaurids”. This dichotomy was formalised by Kitching and Raath (1984; see also Bordy et al. 2004a), who established a biozonation of the Elliot Formation based on a similarly observed understanding of the stratigraphic distribution of Karoo “prosauropods”. Thus, the lower Elliot Formation was designated the “Euskelosaurus Range Zone”, while Kitching and Raath’s “middle” and upper Elliot Formation (UEF sensu Bordy et al. 2004a) was subsumed under the “Massospondylus Range Zone”.
Although several other sauropodomorph taxa had been named on material of varying quality throughout the late nineteenth and twentieth centuries (see reviews in Van Heerden 1979; Cooper 1981; Galton and Upchurch 2004), the taxonomic framework of Kitching and Raath (1984) followed contemporaneous suggestions that the generic content of both the lower and upper Elliot Formation was primarily restricted to “Euskelosaurus” (Van Heerden 1979; Cooper 1980) and Massospondylus (Cooper 1981), respectively. However, continued fossil sampling and taxonomic revision within the intervening decades has made it clear that the biozonation of Kitching and Raath (1984) is both taxonomically and stratigraphically inaccurate. Yates, in a series of influential studies (Yates 2003a, 2004, 2007a, b; Yates and Kitching 2003), convincingly argued that “Euskelosaurus” should be regarded as a nomen dubium, and showed the LEF to be represented by a more taxonomically diverse sauropodomorph fauna. Following Yates (Yates 2003a, 2004, 2007a, b; Yates and Kitching 2003), there are five valid sauropodomorph genera within the LEF: Plateosauravus, Eucnemesaurus, Blikanasaurus, Melanorosaurus, and Antetonitrus. Although the current study affects the stratigraphic (and in the case of Melanorosaurus, the taxonomic) relationships of the latter two taxa, the following brief review temporarily treats them as LEF taxa for the sake of clarity. The type localities of the specimens discussed in the text are shown in Fig. 2.
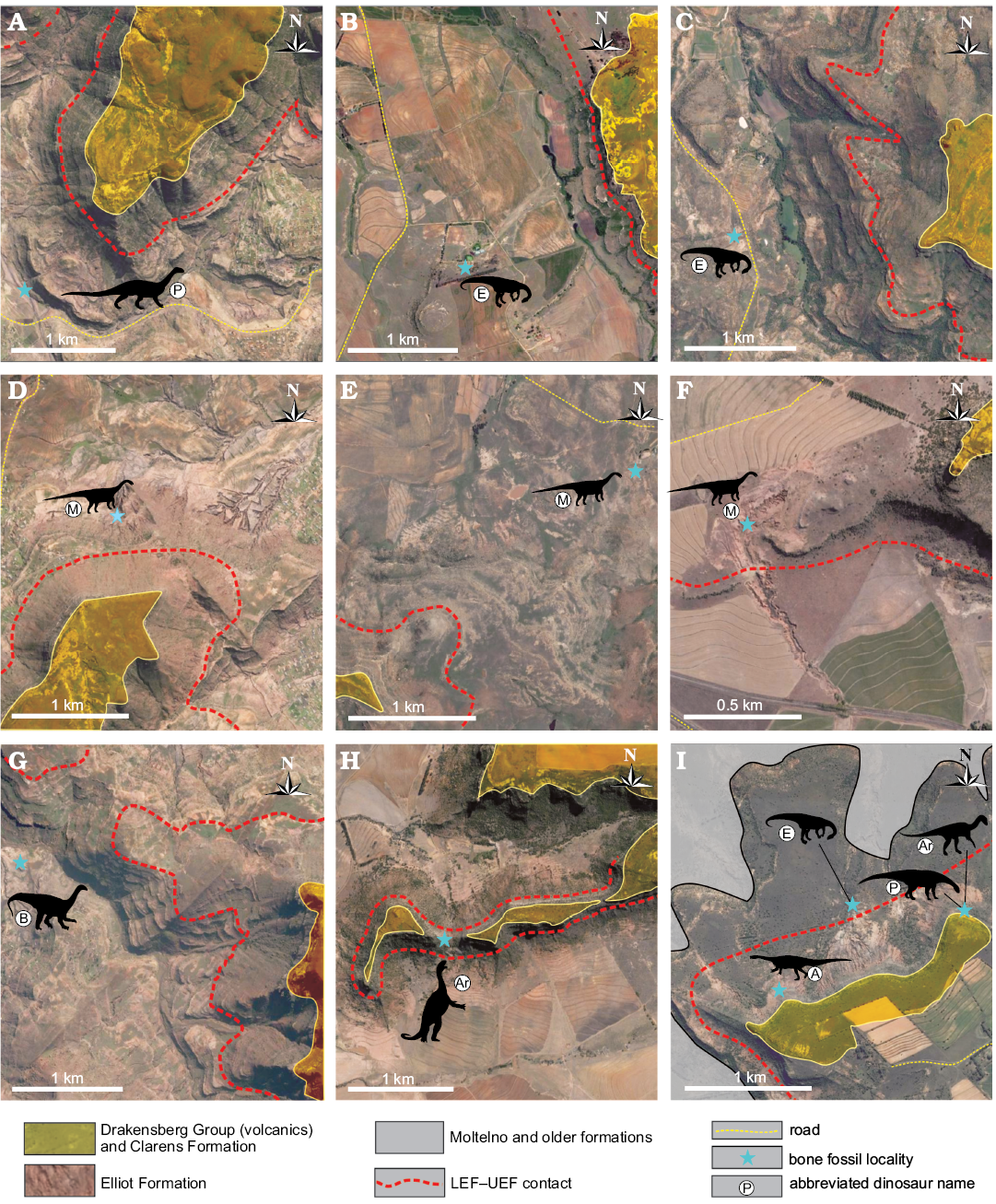
Fig. 2. Geological maps of the key sauropodomorph specimen localities discussed in the text. See Table 1 and Fig. 3 for GPS coordinates and details on stratigraphic positions, respectively. A. Kromme Spruit, ~6 km ESE of Sterkspruit, Eastern Cape Province (3027CB); the type locality of Plateosauravus cullingworthi. B. Zonderhout 291 Farm, ~108 km NE of Ladybrand, eastern Free State Province (2828AC); the type locality of Eucnemesaurus fortis. C. Cannon Rock (270) Farm, ~60 km SE of Sterkspruit, Eastern Cape Province (3127AA); the type locality of Eucnemesaurus entaxonis. D. Thaba ’Nyama, ~11 km SSW of Sterkspruit, Eastern Cape Province (3027CB); the type locality of Melanorosaurus readi. E. Milner (now Swempoort 65) Farm, ~86 km SSW of Sterkspruit, Eastern Cape Province (3126BB); the locality of NMQR 1551 (referred to Melanorosaurus). F. Damplaats 55 Farm, ~12 km WSW of Ladybrand, eastern Free State Province (2927AB); the locality of NMQR 3314 (referred to Melanorosaurus) and the type locality of the basal ornithischian Eucursor parvus (SAM 8025). A referred specimen of Blikanasaurus (BP/1/5271a) has also been documented from this site (see text). G. Blikana, ~25 km ESE of Sterkspruit, Eastern Cape (3027DA); the type locality of Blikanasaurus cromptoni. H. Edelweiss 698 Farm, ~16 km NW of Ladybrand, eastern Free State Province (2927AB); the type locality Antetonitrus ingenipes. I. Heelbo (Spion Kop 932) Farm, ~90 km NNE of Ladybrand, eastern Free State Province (2827BD); the type locality of Arcusaurus pereirabdalorum, Aardonyx celestae, and Pulanesaura eocollum. This locality is also produced a specimen of Eucnemesaurus fortis (Yates, 2007b) from about 30 m above the base of the LEF.
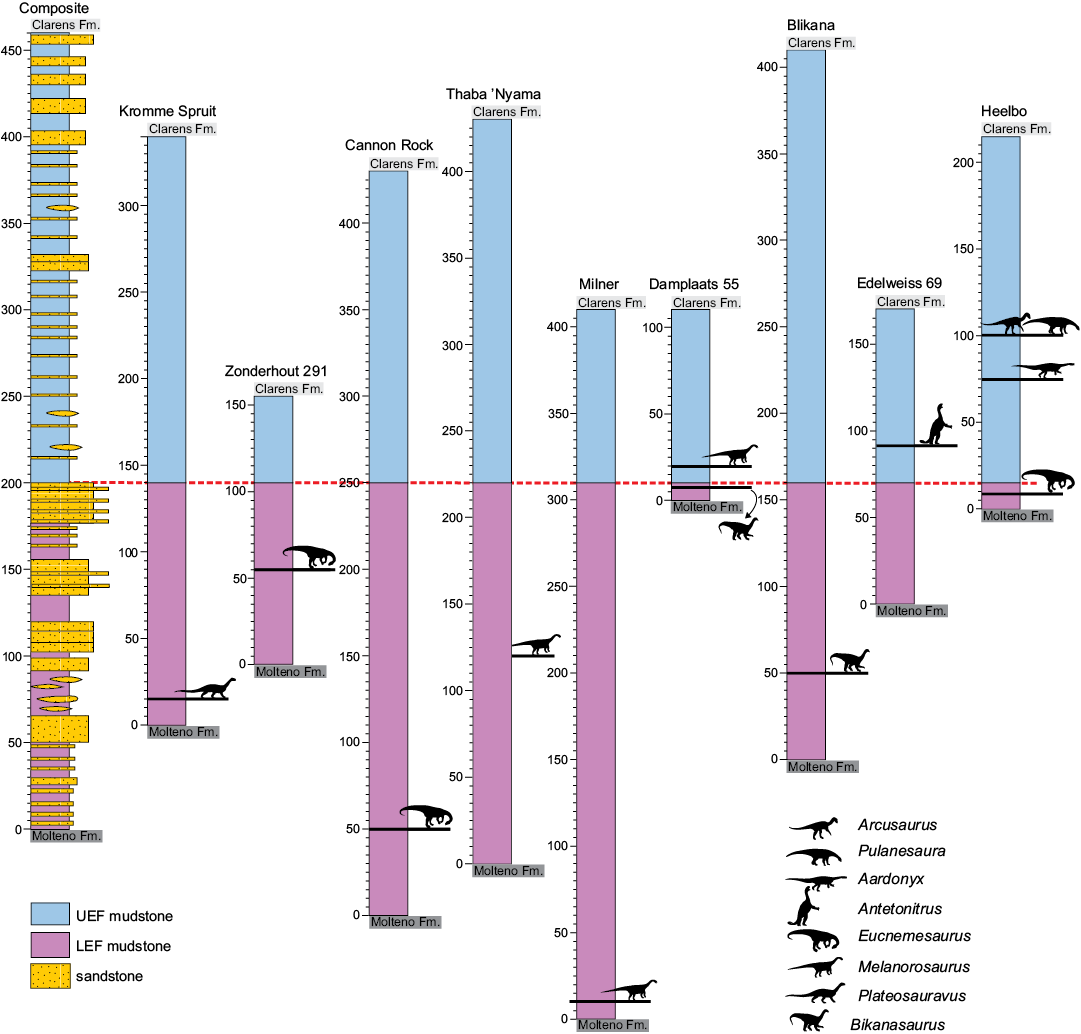
Fig. 3. Schematic sections of the sauropodomorph specimen localities discussed in the text. For each locality, the stratigraphic positon of (i) the fossil specimens, (ii) the lower and upper ranges of the Elliot Formation, and (iii) the LEF–UEF contact are shown. See text and Table 1 for details, and Figs. 1 and 2 for the geological maps of the locations. The composite stratigraphic section of the Elliot Formation is modified from Bordy and Eriksson (2015).
Lower Elliot Formation (former “Euskelosaurus Range Zone”) taxa
Plateosauravus cullingworthi Haughton, 1924 (SAM-PK 3341–3356, 3602–3603, 3607–3609; Huene 1932).—Plateosauravus is generally regarded as the phylogenetically basal-most (Yates 2007a, b; McPhee et al. 2014, 2015b) sauropodomorph within the Elliot Formation. Based on at least two partial skeletons collected in 1918 from a hillside (“koppie”) near Kromme Spruit (~6 km ESE of Sterkspruit, Eastern Cape, South Africa; Haughton 1924; Figs. 1–4, Table 1), this material was later referred to “Euskelosaurus” by Van Heerden (1979) and formed the primary anatomical content of that taxon prior to its invalidation by Yates (2003a, 2004; see also Gauffre 1993a). Although a formal diagnosis and comprehensive description of Plateosauravus is still pending, it is worth noting that some characters of the hypodigm are incongruent with its relatively basal phylogenetic position (e.g., high neural spines in the posterior dorsal vertebrae; relatively elongate humerus compared to the femur), suggesting either convergence with more derived Sauropodomorpha, or a potentially chimerical syntypic assemblage. This situation is further compounded by the lack of precise provenance information associated with the latter (see also Yates 2003a; McPhee et al. 2015b).
Eucnemesaurus Van Hoepen, 1920 (TM 119).—Eucnemesaurus fortis Van Hoepen, 1920 was originally named for material discovered on the farm Zonderhout 291 (~108 km NE of Ladybrand, eastern Free State; Figs. 1–3, Table 1). Effectively ignored prior to its synonymization with “Euskelosaurus” by Van Heerden (1979), Eucnemesaurus was later resurrected by Yates (2007a) as the senior synonym of the enigmatic “herrerasaurid” “Aliwalia rex” (Galton 1985b). Known primarily from fragmentary post-cranial material collected from throughout the basin over the past century, Eucnemesaurus remains a relatively problematic genus (Yates 2007a). Although the recent naming of a second species of Eucnemesaurus based on a partial, articulated skeleton (E. Entaxonis, McPhee et al. 2015b) further supports its validity, several points of similarity between the new species and aspects of the Melanorosaurus hypodigm (see below) point to ongoing taxonomic questions (McPhee et al. 2015b). These issues also have bearing on the validity of the “Riojasauridae” (Riojasaurus + Eucnemesaurus, Yates 2007a) and the observed palaeobiogeographic link between South Africa and other Late Triassic Gondwanan basins in South America (i.e., the Los Colorados Formation of Argentina).
Melanorosaurus readi Haughton, 1924 (SAM-PK 3449, 3450).—Melanorosaurus is of particular importance as it forms the external taxonomic specifier in the current stem-based definition of Sauropoda (Yates 2007a, b; Pol et al. 2011). However, it is also the most problematic genus within the Elliot Formation. Although nominally one of the better known taxa from the LEF, only the skull of a referred specimen (NMQR 3314 recovered from farm Damplaats; see Figs. 1–3, Table 1) has thus far received an explicit diagnosis (Yates 2007b; see also Bonnan and Yates 2007). A recent review of LEF sauropodomorph diversity (McPhee et al. 2015b) noted a number of character conflicts between the post-crania of the two major referred assemblages of Melanorosaurus (NMQR 1551, Galton et al. 2005 and NMQR 3314, Yates 2007b; Bonnan and Yates 2007), especially between the sacra and pedes. This confusion is compounded by the Melanorosaurus syntype series (collected from Thaba Nyama, ~11 km SSW of Sterkspruit, Eastern Cape; Figs. 1–3, Table 1) which (i) lacks these important diagnostic parts of the skeleton, (ii) was collected from two neighbouring localities, and (iii) may have had non-syntype material added to it after initial collection. The anatomical differences between referred specimens of this taxon are suggestive of taxonomic and/or temporal differentiation and warrant more detailed morphological and stratigraphic investigation.
Blikanasaurus cromptoni Galton and Van Heerden, 1985 (SAM-PK 403).—Discovered near the village of Blikana (~25 km ESE of Sterkspruit, Eastern Cape; Figs. 1–3, Table 1) during the joint British-South African expedition of 1962 (Charig et al. 1965; Galton and Van Heerden 1985, 1998), this taxon is of particular interest given both its rarity and hyper-robust proportions. Known only from a holotype comprised of an articulated epipodium and pes, Blikanasaurus elicits a suggestive but poorly-understood insight into robusticity trends within basal Sauropodomorpha, as well as representing a rare morphological departure from the relatively conservative bauplan of the LEF taxa mentioned above (see also McPhee and Choiniere 2016). A second specimen (an isolated first metatarsal) of Blikanasaurus was reported by Yates (2008) from the farm Damplaats in the Ladybrand district of the Free State, and more recently an isolated ilium was suggested as being possibly referable to the genus (McPhee and Choiniere 2016). The former is discussed below with respect to our revised understanding of the stratigraphic relationships at Damplaats.
Antetonitrus ingenipes Yates and Kitching, 2003 (BP/1/ 4952).—Discovered on the farm Edelweiss 698 (~16 km NW of Ladybrand, eastern Free State; Figs. 1–3, Table 1), Antetonitrus was described as the basal-most sauropod known at the time (Yates and Kitching 2003). Although a more comprehensive analysis of the osteology of Antetonitrus highlighted several plesiomorphic features that question its inclusion within Sauropoda (McPhee et al. 2014; see also McPhee et al. 2015a), it nonetheless represents an intriguing morphological midpoint between the latter taxon and more typical basal sauropodomorph taxa such as Massospondylus and Plateosaurus. Originally assigned to the LEF (Kitching and Raath 1984: fig. 4; Yates and Kitching 2003), we present here the results of a comprehensive stratigraphic revision of the type locality.
Other possible LEF taxa.—Two additional taxa have been named from the Elliot Formation in recent years: Meroktenos (= “Melanorosaurus”) thabanensis Gauffre, 1993b (de Fabrègues and Allain 2016) and Sefapanosaurus zastronensis (Otero et al. 2015). The former, a fragmentary assemblage of post-cranial material collected from Lesotho in 1959, was explicitly referred to the LEF (having originally been assigned to the UEF; Gauffre 1993b) based on the recent reinterpretation of historical collection data (de Fabrègues and Allain 2016); the latter, a large assemblage of disarticulated post-cranial material from several individuals recovered from near Zastron (Free State) in the early twentieth century, is entirely lacking in collection information. Uncertainties pertaining to the stratigraphic provenance, inter-association, and highly fragmentary nature of Meroktenos render it of limited utility to the present analysis, and its validity is treated as provisional pending further fossil sampling. Likewise, the indeterminate stratigraphic relationships of the Sefapanosaurus material severely curtail its contribution to our understanding of Elliot biostratigraphy (although its manner of preservation is potentially consistent with the LEF; see McPhee and Choiniere 2016). The discovery of additional associated material is required in order to establish the stratigraphic provenance of this taxon, afford better comparison with the above LEF taxa, and confirm that certain features diagnosing the genus are not simply the result of diagenetic processes (e.g., the morphology of the proximal ulna has clearly been affected by post-mortem distortion, whereas the primary autapomorphy—the triradiate morphology of the astragalar ascending process—is highly unusual for Sauropodomorpha, also suggesting alteration by deformational processes).
Finally, Yates (2003a) described an unnamed “prosauropod” (BP/1/4953) that was found within “a horizon of large concretions near the top of the Lower Elliot Formation” on Hamelneuskop (= Nova Barletta, 28°58’49.42”S, 27°22’ 38.89”E). Here we suggest that this is potentially an UEF specimen (possibly of massospondylid affinity) based on the following: (i) horizons of large concretions are not common within the LEF (Smith and Kitching 1997; Bordy et al. 2004a, b) and (ii) Hamelneuskop does not present rock exposure below ~1700 m a.s.l., a level above which one of us (EMB) has mapped only sedimentary facies typical of the UEF in the immediate area.
Upper Elliot Formation (= Massospondylus Range Zone) taxa
The validity of Massospondylus is less questionable than that of “Euskelosaurus”, largely owing to the recent establishment of a neotype for Massospondylus carinatus based on a well-provenanced, articulated specimen with a skull (Yates and Barrett 2010). Recent discoveries nonetheless show that the UEF is more taxonomically diverse than previously thought (see also Barrett 2004). The majority of these discoveries have come from a series of excavations carried out at the Heelbo locality (Spion Kop 932 farm, ~90 km NNE of Ladybrand, eastern Free State; Figs. 1–3, Table 1). A brief review of all currently valid UEF taxa is given below.
Massospondylus carinatus Owen, 1854.—Massospondylus is ostensibly the most common genus within the UEF, lending its name to both the range zone as well as one of the more speciose (and geographically widespread) clades within the basal sauropodomorph stem—Massospondylidae (see e.g., Yates 2007b; Apaldetti et al. 2011, 2014). Recently, the assignation of a neotype (BP/1/4934) was deemed necessary due to the destruction of the holotype during WWII (Yates and Barrett 2010). However, a comprehensive treatment of both the crania and post-crania of the neotype skeleton of Massospondylus is still outstanding. Although Barrett (2009) named a second species of Massospondylus (M. kaalae) from an isolated partial skull found in the collections of the Iziko Museum (SAM-PK 1325; see also Gow et al. 1990), our current understanding of the postcranial morphology of the genus remains relatively informal, being typified primarily by “gracile” remains exhibiting a relatively plesiomorphic anatomy and proportionately elongate cervical vertebrae. Although Cooper (1981) provides an indepth account of the post-cranial anatomy of putative Massospondylus material from Zimbabwe, the conspecificity of this material with the South African assemblage has never been adequately demonstrated via e.g., synapomorphies or a unique combination of characters. The lack of an explicit diagnosis for the genus also has bearing on the status of historically referred taxa (see Cooper 1981), with some suggestion that the enigmatic “Gryponyx” (Broom 1911) may be taxonomically distinct from Massospondylus (Vasconcelos and Yates 2004; see also Galton and Upchurch 2004).
Arcusaurus pereirabdalorum Yates, Bonnan, and Neveling, 2011 (BP/1/6235).—Based on fragmentary material (including a partial skull) from the same quarry that produced Pulanesaura (on Heelbo; see below and Figs. 1–3, Table 1), Arcusaurus was hypothesised to represent either a basal sauropodiform closely related to South African taxa such as Aardonyx or, in the preferred scenario, a stem sauropodomorph more closely related to European taxa like Thecodontosaurus and Efraasia (and hence representing a possible relictual survivor from the Norian; Yates et al. 2011). This uncertainty is undoubtedly a reflection of the fragmentary nature of the known Arcusaurus material which, along with its clearly juvenile status, makes an assessment of its phyologenetic relationships difficult.
Ignavusaurus rachelis Knoll, 2010.—Named from an articulated, partial skeleton recovered from the Qacha’s Nek district of southern Lesotho, the validity of this taxon was promptly questioned by Yates et al. (2011) who suggested synonymy with Massospondylus. Although every analysis that has included Ignavusaurus has recovered it in a position basal to Massospondylidae sensu Yates (2007b) (Knoll 2010; Apaldetti et al. 2011; Pol and Powell 2013), this result should be tempered against the marked juvenility of the holotype and the known effects of ontogeny on phylogenetic reconstruction.
Aardonyx celestae Yates, Bonnan, Neveling, Chinsamy, and Blackbeard, 2010 (BP/1/6254).—The first new sauropodomorph genus named from the UEF of South Africa in several decades, Aardonyx was also the first of the three new genera recently described from the Free State farm Heelbo (Figs. 1–3, Table 1). Known from a disarticulated assemblage comprising both cranial and postcranial material from at least two individuals, Aardonyx is of primary interest because it provided the first unequivocal evidence that large-bodied, “near-sauropod” taxa were not exclusive to the LEF (Yates et al. 2010).
Pulanesaura eocollum McPhee, Bonnan, Yates, Neveling, and Choiniere, 2015a (BP/1/6982).—The most recently named taxon from the highly productive Free State farm Heelbo (Figs. 1–3, Table 1), Pulanesaura is currently the most derived sauropodomorph known in the Elliot Formation. Also known from the disarticulated remains of at least two individuals, it was argued by McPhee et al. (2015a) that specific apomorphic features of the neck and forelimb of Pulanesaura provided valuable insight into the palaeoecological circumstances underpinning the initial radiation of Sauropoda.
Although our taxonomic knowledge of Elliot Formation Sauropodomorpha has advanced dramatically since the pioneering investigations of Kitching and Raath (1984; see also Haughton 1924), our knowledge of the stratigraphic relationships of many other specimens has tended to uncritically follow the information given at the time of original collection (e.g., Yates and Kitching 2003; Yates 2003a, 2007b, 2008). Given that the majority of these discoveries occurred prior to our improved understanding of the lithostratigraphy of the Elliot Formation, an updated assessment of the stratigraphic relationships of their collection sites has the potential to yield important new insights. Our ultimate goal is to produce a high-resolution, basin-wide biostratigraphic model for the advancement and refinement of the original biostratigraphic framework of Kitching and Raath (1984), akin to the detailed models that have already been presented for older sections of the Karoo basin (Rubidge et al. 2016).
Table 1. Stratigraphic and geographic information of key sauropodomorph fossil localities within the Upper Triassic to Lower Jurassic Elliot Formation of South Africa. See text for discussion.
|
Locality |
Coordinate |
Specimen |
Catalogue number |
Elevation above sea level [m] |
Thickness [m] |
||||||
|
GPS |
1:50 000 map |
base LEF |
top UEF |
LEF–UEF |
fossil |
LEF |
UEF |
total EF |
|||
|
Kromme Spruit |
30°33’4.25”S 27°25’50.55”E |
3027CB |
Plateosauravus cullingworthi |
SAM-PK 3341–3356, 3602–3603, 3607–3609 |
1490 |
1830 |
1630 |
1505 |
140 |
200 |
340 |
|
Zonderhout 291 Farm |
28°29’39.04”S 28°13’46.70”E |
2828AC |
Eucnemesaurus fortis |
TM 119 |
1745 |
1900 |
1850 |
1800 |
105 |
50 |
155 |
|
Cannon Rock (270) Farm |
31° 0’3.92”S 27° 5’43.87”E |
3127AA |
Eucnemesaurus entaxonis |
BP/1/6234 |
1450 |
1880 |
1700 |
1500 |
250 |
180 |
430 |
|
Thaba ’Nyama |
30°36’45.56”S 27°20’45.59”E |
3027CB |
Melanorosaurus readi |
SAM-PK 3449, 3450 |
1480 |
1910 |
1700 |
1600 |
220 |
210 |
430 |
|
Milner (now Swempoort 65) Farm |
31°13’47.19”S 26°59’5.46”E |
3126BB |
Melanorosaurus sp. |
NMQR 1551 |
1540 |
1950 |
1800 |
1580 |
310 |
100 |
410 |
|
Damplaats 55 Farm |
29°13’21.86”S 27°20’13.46”E |
2927AB |
Melanorosaurus sp. |
NMQR 3314 |
1640 |
1750 |
1650 |
1660 |
10 |
100 |
110 |
|
Blikana |
30°33’29.81”S 27°37’20.38”E |
3027DA |
Blikanasaurus cromptoni |
SAM-PK 403 |
1500 |
1910 |
1660 |
1550 |
160 |
250 |
410 |
|
Edelweiss 698 Farm |
29° 6’36.14”S 27°19’16.38”E |
2927AB |
Antetonitrus
|
BP/1/4952 |
1650 |
1800 |
1720 |
1742 |
70 |
80 |
150 |
|
Heelbo (Spion Kop 932) Farm |
28°28’1.27”S 27°49’34.07”E |
2827BD |
Arcusaurus pereirabdalorum |
BP/1/6235 |
1645 |
1660 |
1760 |
1720 |
15 |
100 |
115 |
|
Aardonyx celestae |
BP/1/6254 (holotype) |
1745 |
|||||||||
|
Pulanesaura
|
BP/1/6982 (holotype) |
1745 |
|||||||||
Material and methods
We relocated sites from which taxonomically and/or stratigraphically significant sauropodomorph material has been recovered using historical literature, first-hand accounts, and specimen records at the Iziko Museum, National Museum, and Evolutionary Studies Institutes. We visited each of these sites and documented the stratigraphy and sedimentology of each using standard methods (e.g., jacob’s staff measurements of stratigraphic thickness, grain size comparison, palaeocurrent directions). On occasions when the precise location of fossil discoveries could not be identified to within a few meters (e.g., Plateosauravus), the immediate sedimentological, taphonomic, and stratigraphic context of the area nonetheless provided an informative contribution to the biostratigraphic question at hand.
To assess the fit of our revised stratigraphic hypotheses to the current hypothesis for sauropodomorph phylogeny, we calculated the modified Manhattan Stratigraphic Measure (MSM; Pol and Norell 2001, 2006; Pol et al. 2004), modified Gap Excess Ratio (GER; Wills et al. 2008); and Minimum Implied Gaps (MIG; Benton 1994; Wills 1999) for our previous biostratigraphic hypothesis and the one presented here (see SOM: table S1, Supplementary Online Material available at http://app.pan.pl/SOM/app62-McPhee_etal_SOM.pdf). Because the UEF and LEF age ranges are uncertain, we used the following ranges in the randomization study (LEF, 208.5–201.3 Ma; UEF, 201.3–190.8 Ma; Cohen et al. 2013). All stratigraphic consistency values were calculated in the R statistical environment (R 3.3.3; R Core Team 2017) using the Strap package v2.0 (Bell and Lloyd 2015).
Using a broadly sampled discrete morphological character matrix, we investigated how sauropodomorph dinosaur disparity is partitioned between: (i) UEF and LEF taxa; and (ii) Late Triassic (Norian to Rhaetian) and Early Jurassic (Hettangian to Pliensbachian) taxa. These analyses were performed in R using the Claddis package (Lloyd 2016; scripts and supporting files available in the SOM). To assess the morphospace occupation across multidimensional ordinations of disparity, we employed the Functional Richness and Functional Evenness metrics from functional ecology studies (e.g., Villéger et al. 2008), using presence/absence “community” data structured to reflect UEF and LEF taxa and Late Triassic and Early Jurassic taxa, respectively (available in the SOM). These calculations were done in R using the FD package with default settings (Laliberté and Legendre 2010; Laliberté et al. 2014). In our usage, the Functional Richness metric is equivalent to the volume of a convex hull containing all community members (in this case, either UEF–LEF or Late Triassic–Early Jurassic taxa), and thus measures the comparative size of morphospace occupation. The Functional Evenness metric was proposed to assess how regularly taxa are placed and how evenly abundance is distributed in multidimensional ordination space (Villéger et al. 2008). In our usage, which includes only presence/absence data, it measures how evenly taxa are placed in the morphospace.
The analyses contained herein utilised a modified version of a data matrix used in recent analyses of sauropodomorph phylogeny (McPhee et al. 2015a, b; see SOM). As Claddis requires a fully resolved (i.e., without polytomies) phylogenetic tree, a single MPT from among 48 minimally differing MPTs was chosen that represents the current consensus of basal sauropodomorph relationships. All 48 MPTs were retained for the tests of stratigraphic congruence, which utilised the default settings within STRAP (i.e., 1000 resampling permutations of the original trees and 1000 randomly generated trees, see SOM). For the purposes of the current study, Melanorosaurus was treated as two OTUs representing the most informative of its referred specimens (NMQR 1551 and 3314), whereas all metataxa (e.g., Crurotarsi, Theropoda, Ornithischia) were removed from the analysis. Isanosaurus is here considered to be an Early Jurassic taxon following Racey and Goodall (2009).
Results
Stratigraphic provenance of key taxa.—The stratigraphic relationships of UEF taxa from Heelbo Farm (i.e., Aardonyx, Arcusaurus, and Pulanesaura), in addition to being uncontroversial, are given in some detail in McPhee et al. (2015a) and will not be reiterated here (although see Discussion below). Similarly, Massospondylus (or a massospondylid-like sauropodomorph(s)) is known from dozens of specimens at multiple localities throughout the UEF (e.g., Kitching and Raath 1984: tables 1–3; Gow et al. 1990; Barrett 2009; Knoll 2005; Yates and Vasconcelos 2005; Yates and Barrett 2010; Yates et al. 2011; Reisz et al. 2012; Butler et al. 2013; McPhee et al. 2015b), and hence a detailed assessment of the geographic and stratigraphic distribution of this commonly occurring taxon is beyond the scope of the current contribution. The results will therefore focus on the less well-known LEF and UEF locations. As the majority of current LEF taxa possess very limited occurrence data beyond their respective type localities, improved understanding of their stratigraphic provenances has greater potential to either corroborate or alter current hypotheses regarding the biostratigraphic distribution of Elliot Formation Sauropodomorpha.
Plateosauravus: The Plateosauravus syntype assemblage is disarticulated, but the majority of material agrees closely in its preservational quality (being also broadly consistent with other LEF fossil material), suggesting that it came from a single locality near Kromme Spruit in the Eastern Cape (Figs. 2–4; Table 1). Nonetheless, the information given in Haughton (1924: 407), which suggests some ex situ dislocation between “a number of… large Dinosaurian bones… found weathered out down one of the slopes of a steep kopje formed of the basal rocks of the Red Beds” and a “pocket” of in situ material excavated further up the same hillside, cautions against assumptions of monospecificity and/or association for this assemblage. Although the precise slope mentioned by Haughton (1924) is unknown, the strata in the immediate vicinity of Kromme Spruit expose rocks with multi-storey sandstone, up to 15–20 m thick interbedded with purple-red mudstone with limited pedogenic alteration features, typical of the LEF (Fig. 4). This contrasts with the rocks exposed farther to the north in the incised valley of the Kromme Spruit River (Fig. 2A), which are very coarse-grained, tabular and interbedded with grey to khaki mudstone (i.e., Molteno Formation). Within this general area, the LEF–UEF contact was located ~140 m above the base of the LEF, at the prominent break in the slope above which multi-storey sandstone are missing. If the Plateosauravus material had come from these latter strata it is unlikely that the ex situ material would have reached the bottom of the hill intact. We are therefore confident that the Plateosauravus syntype can be provenanced to the LEF, although its relative position (and association) within it is unknown.
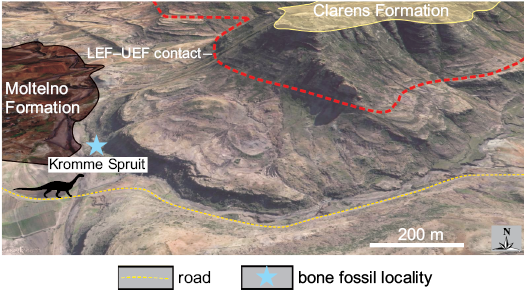
Fig. 4. Stratigraphic relationships at the type locality of Plateosauravus cullingworthi, Kromme Spruit, ~6 km ESE of Sterkspruit, Eastern Cape Province (3027CB). The key stratigraphic contacts have been mapped using the lithological characters of the various stratigraphic units. See Table 1 and text for details. Base image provided by Google Earth.
Melanorosaurus: Fossil material from three main assemblages has been referred to this genus: the syntype series (SAM-PK 3449 and 3450; Haughton 1924) and two referred assemblages, NMQR 3314 (Yates 2007b) and NMQR 1551 (Galton et al. 2005). Another collection of disarticulated post-cranial material (SAM-PK 3532) was referred to Melanorosaurus by Haughton (1924; see also Bonnan and Yates 2007) but, with the exception of a mostly complete ulna, ilium, humerus, and scapula, it is of relatively limited diagnostic utility. The syntype series is composed primarily of disarticulated vertebral and appendicular material that was found “under the first sandstone ridge west of the dolerite” dyke on the northern slope of the Thaba ‘Nyama (“Black Mountain”, ~11 km SSW of Sterkspruit, Eastern Cape Province; Haughton 1924; Galton et al. 2005: 5). Our investigation of the type locality (Figs. 2D, 3; Table 1) shows that all strata on the north slope, west of the dolerite dyke, and beneath the first sandstone ridge, are LEF. Sedimentological observations support this assignment, and include up to 10 m thick, multi-storey, cross-bedded sandstone with lateral accretion surfaces and upward-fining successions in which the grain-size ranges from very coarse-grained sand with rare, small quartz pebbles to very fine sand. These channel-shaped sandstone bodies are distinct from the tabular, laterally continuous, mostly very coarse grained sandstone of the underlying Molteno Formation, which crops out less than 4 km to the NE at elevations less than 1480 m. In addition, we observed that LEF rocks extend up to 1700 m elevation at the syntype locality.
NMQR 1551 represents a large assemblage of post-cranial material (at least two individuals) collected between 1967 and 1971 from the “base of the Elliot Formation, Milner Farm, Wodehouse (Dordrecht) District” (Galton et al. 2005: 6). The material was apparently excavated from the “banks of a narrow furrow” (Galton et al. 2005: 6) over an area of approximately 6 meters. Our visit to Milner Farm (~86 km SSW of Sterkspruit, Eastern Cape Province; Figs. 2E, 3; Table 1) confirmed the presence of a small trench (or “donga”) that extends from the base of the hill on which the main farm buildings are situated (to the NE of the site). Although no additional fossil material was uncovered, our investigation of the locality revealed sedimentary features congruent with the LEF as observed at the Melanorosaurus type locality at Thaba ‘Nyama (see above). Furthermore, the channel-shaped LEF sandstone are distinct from the tabular, laterally continuous, mostly very coarse grained sandstone of the underlying Molteno Formation, which in turn are mapped <1.5 km from the fossil site at elevations <1540 m (Fig. 3; Table 1).
NMQR3314 was recovered in 1994 from the farm Damplaats 55 (~12 km WSW of Ladybrand, eastern Free State Province; Figs. 2F, 3; Table 1). This locality is remarkable for also having produced the basal ornithischian Eocursor (Butler et al. 2007). Although recorded in relatively fine detail in the supplementary information of Butler et al. (2007), the lithostratigraphy of Damplaats remains somewhat unclear. Yates (2008: 40) described a second specimen of Blikanasaurus from this locality, asserting that it had to have come from the upper 20 m of the LEF, “because that is all that is exposed” at Damplaats. However, this ignores an additional ~20 m of UEF documented by Butler et al. (2007: fig. S1) who cite as the boundary between the UEF and LEF a semi-continuous large carbonate nodule horizon that is approximately in the middle of the Elliot succession at Damplaats. Eocursor was reported as coming from 5.5 m below this large carbonate nodule horizon, and 10.5 m above where NMQR 3314 was discovered (Butler et al. 2007: S1; see also Olsen et al. 2011). Butler et al. (2007: S1) characterise the taphonomic style of Eocursor as that of a desiccated animal covered by “windblown silt”, which is directly overlain by a sandstone that probably originated in a flash-flooding event. With respect to the former, no supporting sedimentological field or petrographic data were provided, and were not found during our investigations either (see below). Additionally, the large carbonate nodule horizons are not diagnostic of the boundary between the LEF–UEF because the UEF itself contains several large carbonate nodule horizons, this site included.
In our high-resolution sedimentological investigation of Damplaats (Fig. 5), the fossiliferous, pedogenically altered zone, recorded by Butler et al. (2007), is considered to fall ~15 m above the LEF–UEF contact and within the UEF (see Figs. 2F, 3; Table 1). We consider that the contact of the LEF–UEF is conjunctive with the regional trend, i.e., a change in sandstone architecture from ~5–6 m thick channel-shaped bodies to < 2–3 m thick, laterally extensive, tabular or sheet-like sandstone. Our study shows (Figs. 2F, 3; Table 1) that except for the lowermost ~10 m of outcrops, which expose the LEF, the outcrops at this site are dominated by pedogenically altered mudstone that are characterised by numerous, well-developed desiccation cracks, rhizocretions, discolouration features (mottles), root halos, slickenlines and in situ carbonate nodules. Up-section at Damplaats, characteristic UEF sedimentological features persist, including multiple thin sandstone and massive-to-laminated mudstone layers with increasingly better-developed pedogenic alteration features (Fig. 5). Therefore, the contact of the LEF–UEF can be placed just a few metres above the only thick sandstone in the lower part of the exposures (top ~1556 m), below which the distinctly purple-red mudstone show features typical of the LEF regionally, e.g., very limited pedogenic overprinting (rare, isolated blotches; very sparse nodules; Fig. 5).

Fig. 5. Lithostratigraphic details of the erosional gully (donga) located on Damplaats Farm. Abbreviations: Fl, horizontal laminated mudstone; Fm, massive mudstone; Gcm, clast-supported conglomerate; Gmm, matrix-supported conglomerate; P, paleosol; Sl, low-angle cross-bedded sandstone; Sm, massive or faintly laminated sandstone; Sp, cross-bedded sandstone; Sr, ripple cross-laminated sandstone.
Unfortunately, the original field description of NMQR 3314 is not available, making placement of this specimen difficult. However, the taphonomic appearance of NMQR 3314 and the sedimentary rock matrix still adhering to it are highly characteristic of the UEF. The former is typical of fossil bones found within the UEF, showing the explosive deformation of bone surfaces (supposedly as an effect of prolonged subaerial exposure—a preservational condition that is comparatively rare with respect to LEF fossils; McPhee and Choiniere 2016). The matrix adhering to the bone of NMQR 3314, a light pink, competent, pedogenically pervasively altered siltstone, is hard to explain if the specimen was indeed located 16 m below the lowermost large-nodule horizon (and 10.5 m below Eocursor), because at that stratigraphic position the outcrops expose the purple-red mudstone with very limited pedogenic alteration (see Figs. 2F, 3, 4). Finally, several eye-witness accounts from the time of the excavation also provide support for a position higher within the section (e.g., by Bruce Rubidge [personal communication 2016], who viewed the specimen in situ and recalls it being excavated well above the thick sandstone mentioned above).
In summary, the current placement of NMQR 3314 some 10.5 meters below Eocursor is considered doubtful, not only because Butler et al. (2007) provide no further reasoning for it, but also because of the above sedimentological observations. Furthermore, a revision of the placement of the Eocursor specimen means that it comfortably falls within the lowermost UEF (see Figs. 2F, 3; Table 1), and this would imply that this iconic taxon is Early Jurassic. These observations show that NMQR 3314 is located within the UEF—incongruent with the stratigraphic position of the other Melanorosaurus material. Furthermore, reassessment of the postcranial anatomy of NMQR 3314 shows character conflict with NMQR 1551 (McPhee et al. 2015b). The implications of this for the taxonomic assignment of NMQR 3314 are discussed further below, as are the possible stratigraphic relationships of the referred Blikanasaurus specimen (BP/1/5271a).
Eucnemesaurus: The type locality of Eucnemesaurus fortis is the farm Zounderhout 291 Farm (~108 km NE of Ladybrand; Figs. 2B, 3; Table 1) in the eastern Free State Province (Van Hoepen 1920), although Kitching and Raath (1984: table 1) noted the absence of more specific locality information. During our visit to the property a small exposure of mudrock and sandstone, roughly 200 m from the farmhouse, revealed some scattered fragmentary fossil material. No other natural outcrop, except for practically inaccessible cliffs of the UEF and Clarens Formation (see eastern part of Fig. 2B), is observable anywhere on the farm. Based on its stratigraphic proximity to the mapped contact of the Molteno and Elliot formations (western part of Fig. 2B), as well as the high matrix content of its sandstone (wacke), we identified the exposure near the farm house as part of the LEF. This is consistent with the stratigraphic position of the recently described second species of Eucnemesaurus (E. entaxonis, McPhee et al. 2015b), which is located on farm Cannon Rock (Figs. 2C, 3; Table 1) at 1500 m elevation, some 50 m above the basal contact of the formation (contra McPhee et al. 2015b, who placed it somewhat closer to the contact of the Molteno–Elliot formations), as well as the referred material of E. fortis (Yates 2007a), which was collected from within the LEF donga system on Heelbo (Figs. 2I, 3; Table 1). Unfortunately, more precise stratigraphic correlation between these specimens is not possible at this time given the marked difference in LEF thickness between the southern and northern parts of the basin.
Blikanasaurus: The probable type locality for Blikanasaurus (Figs. 2G, 3; Table 1) was retraced with respect to the locality information given in Galton and Van Heerden (1985: 510): “1.6 km northeast of Blikana Trading Store, Herschel district, Transkei, South Africa, at an elevation of 1370 m, from the lowermost part of the lower Elliot”. These coordinates are broadly congruent with an isolated hillock exposing the Elliot Formation at the very base of the LEF, although the elevation of the Blikana fossil site is at 1550 m (the nearest elevation of 1370 m is >40 km west from Blikana in the Orange River valley at coordinates 30°30’29.62”S 27°12’16.76”E). Although we observed some isolated sauropodomorph vertebrae and fragmentary hindlimb material weathering out, this material is unlikely to relate to that recovered in the original excavation in 1962. Nonetheless, given the local sedimentological properties (identical to those mentioned at the Melanorosaurus type locality at Thaba ‘Nyama) and proximity of the fossil site to the nearest Molteno Formation exposures (<3.5 km NW from Blikana, at 30°32’16.00”S 27°35’59.35”E), the stratigraphic provenance for the Blikanasaurus holotype is considered here as very low down in the LEF (as per Charig et al. 1965).
Antetonitrus: The stratigraphic provenance of Antetonitrus has recently come into question (McPhee et al. 2015a; McPhee and Choiniere 2016; Bordy et al. 2017). In introducing the taxon, Yates and Kitching (2003) followed the biostratigraphic framework of Kitching and Raath (1984) whereby all large-bodied sauropodomorphs were essentially consigned to the LEF (“Euskelosaurus Range Zone”). A later stratigraphic assessment of the Edelweiss locality (~16 km NW of Ladybrand, eastern Free State Province) restated the position of Antetonitrus within the uppermost LEF, even though a layer of diagnostic massive, clast-supported carbonate nodule conglomerate—a potential index rock of the UEF (Bordy et al. 2004a)—was documented below the stratigraphic level from which Antetonitrus is believed to have been excavated (Abdala et al. 2007: fig. 1). Reiterating from Bordy et al. (2017), the following features are highly characteristic of the UEF in the immediate vicinity (including 20 m below) of the Antetonitrus type locality: (i) diagnostic carbonate nodule conglomerates; (ii) upward-fining, channel fill sandstone without lateral accretion surfaces; (iii) deep red, maroon to deep pink massive mudstone with well-developed pedogenic alteration features (e.g., great abundance of in situ carbonate nodules, calcareous rhizocretions, deeply penetrating desiccation cracks); (iv) deep red to maroon, clast-rich very fine grained sandstone with poorly sorted, 1–4 cm angular, rip-up mudstone clasts.
Antetonitrus is therefore an UEF taxon, albeit one from relatively low in the stratum.
Results of numeric analyses.—Our tests of stratigraphic congruence (i.e., MSM, GER, MIG, SCI) show that our current reconstruction of the stratigraphic relationships of Late Triassic and Early Jurassic Sauropodmorpha represents a significantly better explanation than if this data were generated at random (Figs. 6, 7). However, comparison between our previous hypotheses of Elliot Formation sauropodomorph biostratigraphy (when Antetonitrus and NMQR 3314, “Melanorosaurus” were assigned to the latest Triassic) suggests that the revised arrangement is actually slightly less congruent with phylogeny than the former (see SOM: table S2 vs table S3). This reduction in stratigraphic fit is likely primarily due to the increased temporal separation between Antetonitrus and Lessemsaurus, with these sister-taxa now spanning the mid-Norian to Sinemurian (see below).

Fig. 6. The hypothesis of sauropodomorph biostratigraphic relationships presented within, reconstructed with “equal” branch lengths (Bell and Lloyd 2015). This tree represents the “best-fit” from amongst the 48 original MPTs as per the modified Gap Excess Ratio (GER). Note that although the genus Eucnemesaurus is paraphyletic in the current topology, the relationship (E. entaxonis (Riojasaurus + E. fortis)) is not supported by synapomorphies present in Riojasaurus and E. fortis and absent in E. entaxonis, and instead appears to be a reflection of the higher number of localised autapomorphies present in E. entaxonis compared to E. fortis. The highly incomplete nature of the specimens grouped under Eucnemesaurus, along with the possibly chimerical status of Riojasaurus (see McPhee et al. 2015b), presents issues that exceed the scope of the current study to adequately address. Specimens from Elliot Formation shown in bold.
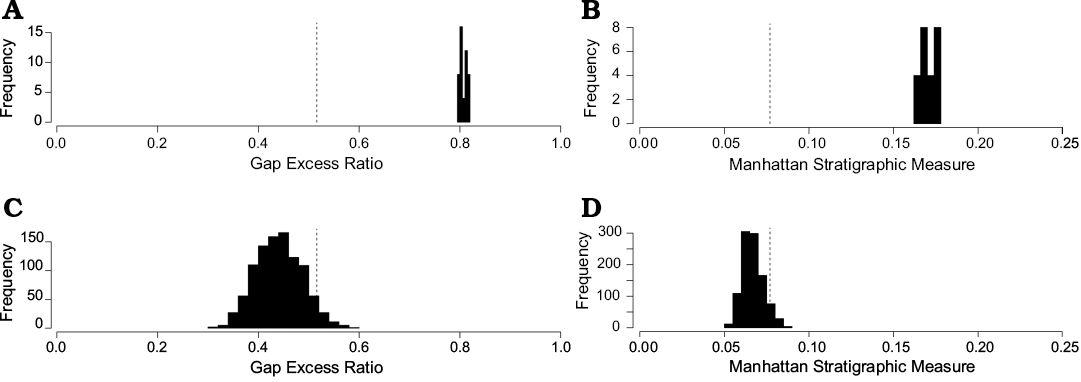
Fig. 7. Histograms indicating the level of stratigraphic fit of the 48 input trees (A, B) versus the randomly generated topologies (C, D). Only the results for the Gap Excess Ratio (C) and the modified Manhattan Stratigraphic Measure (D) are shown here. The vertical dashed line represents the critical value at which 95% of trees fail the randomization test. Thus, all 48 MPTs trees show a significantly better fit to stratigraphy than expected at random.
Our disparity analyses demonstrate that sauropodomorph morphological diversity within the upper Elliot Formation was greater than that of the lower Elliot Formation with respect to every available metric, including both total range and the degree of variance within that range (Fig 8; Table 2). Although these results are not markedly dissimilar, and unlikely to be distinct at the level of statistical significance, the Functional Richness metric (Table 2) nonetheless indicates that the morphospace volume occupied by upper Elliot Formation taxa is approximately twice that of lower Elliot Formation taxa. This trend is also observable across all analysed taxa of the Late Triassic (Norian–Rhaetian) and Early Jurassic (Hettangian–Pliensbachian), although it requires mentioning that both Functional Richness and the sum of disparity ranges are highly sensitive to sample size (Butler et al. 2012; Villéger et al. 2008; Table 2). Disparity between the two time-bins is especially marked if Vulcanodon and Gongxianosaurus (both taxa of highly uncertain age that are possibly older than the Toarcian age often assigned them; Yates et al. 2004; Chen et al. 2006) are included within the latter assemblage; it is further heightened if Thecodontosaurus and Pantydraco are removed from the former. (Ages for the British “Rhaetian” fissure fills are very poorly constrained, and could potentially date as early as the Carnian; Storrs 1994; Benton et al. 2000; although see Foffa et al. 2014; a possibility given tentative support by their very basal phylogenetic relationships). It is clear that this signal is primarily influenced by the basal divergence of Sauropoda within the earliest Jurassic. The relevance of changing environmental/ecological conditions across the TJB, as well as events associated with the boundary itself, to sauropodomorph diversity is explored in depth below.
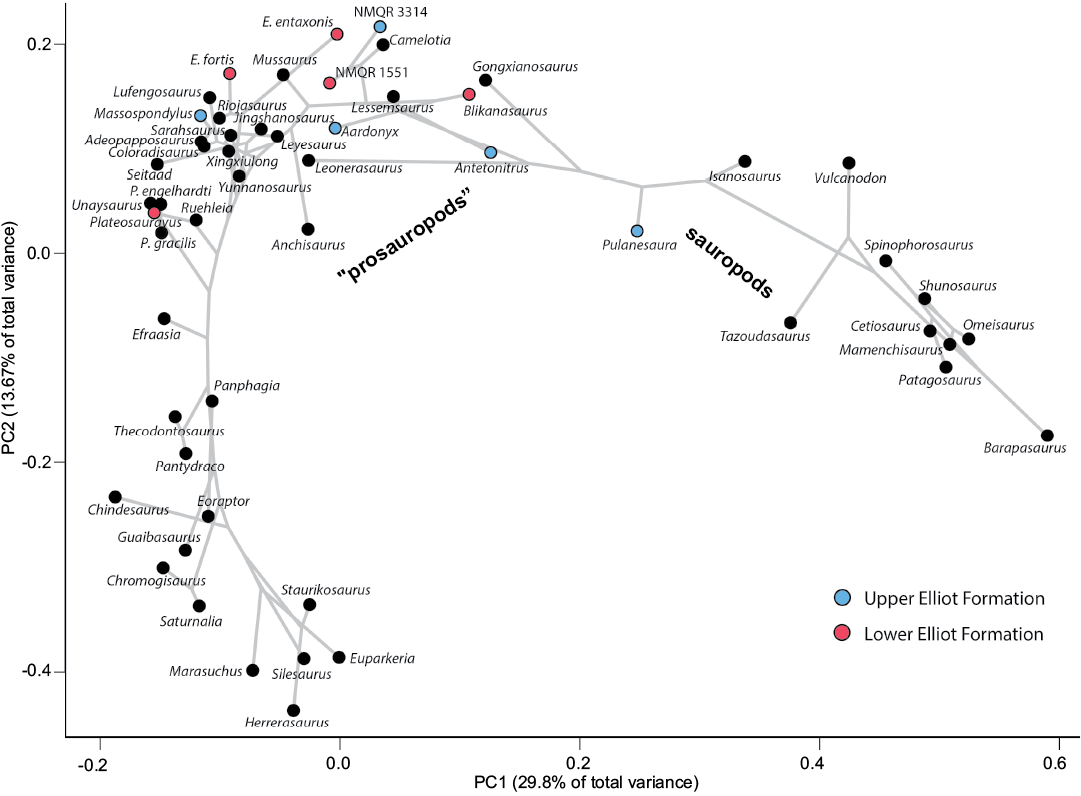
Fig. 8. Morphospace plot of the first two PCO axes generated in the R statistical environment (Claddis package). Branches are superimposed from a single representative topology selected from amongst the 48 MPTs.
Table 2. Results of the Principal Coordinate and Functional Richness analyses, showing the disparity metrics for the taxon-bins of interest. Asterisk denotes removal of Thecodontosaurus and Pantrydraco from the Late Triassic, and the addition of Gongxianosaurus and Vulcanodon to the Early Jurassic.
| |
Lower Elliot Formation |
Upper Elliot Formation |
Late Triassic |
Early Jurassic |
Late Triassic* |
Early Jurassic* |
|
Mean pairwise distance (pre-ordination) |
0.4113 |
0.4857 |
0.4443 |
0.4941 |
0.434 |
0.5193 |
|
Sum of ranges |
4.9669 |
5.0947 |
7.5229 |
8.1789 |
7.2197 |
8.8007 |
|
Sum of variance |
0.1401 |
0.1457 |
0.1334 |
0.1631 |
0.1285 |
0.1786 |
|
Functional richness |
1.0897 |
2.3212 |
3.31E-08 |
2.40E-11 |
2.26E-08 |
5.75E-05 |
|
Functional evenness |
0.875 |
0.9699 |
0.9117 |
0.8688 |
0.9082 |
0.8691 |
Discussion
Despite the increasingly widespread use of radiometric methods for age assessment, biostratigraphy remains one of the most useful methods for relative correlation of strata over both long and short distances. Moreover, it is our primary source of data for understanding faunal change over time. The Elliot Formation of South Africa and Lesotho is a world standard for understanding the early evolution of Sauropodomorpha, and for global biostratigraphic correlation of Upper Triassic to Lower Jurassic strata. However, the sauropodomorph biostratigraphy of the Elliot Formation has not been revised in more than 20 years, which have been some of the most active periods of work on southern African dinosaurs. Moreover, this unit potentially contains the TJB, and a finer-scale understanding of faunal distributions within the Elliot is necessary to understand the terrestrial effects of this extinction event.
Although never an explicit desideratum of Kitching and Raath (1984), the twofold partitioning of the Elliot Formation into “Euskelosaurus” and Massospondylus range zones resulted in the tendency for workers to orient themselves stratigraphically with reference to the size of the dinosaur bones they were finding. Hence, anything that was arbitrarily deemed to be “large” suggested the LEF, whereas a preponderance of smaller-bodied “prosauropods” indicated a higher position in the formation. Our investigations, along with taxonomic revision over the last decade and a half, have shown that this biostratigraphic scheme is no longer an accurate reflection of the stratigraphic distribution of Elliot Formation Sauropodomorpha. Below we summarise our revised understanding of the taxonomic content of both the LEF and UEF.
Lower Elliot Formation.—Building on the taxonomic work of Adam Yates (see references above), we have shown that the LEF contains at least four valid genera: Blikanasaurus, Plateosauravus, Eucnemesaurus spp., and Melanorosaurus (although at least one referred specimen of the latter is located within the UEF, see below). Nonetheless, the LEF so far lacks any taxa of comparable gracility to Massospondylus, suggesting a genuine ecological bias towards sauropodomorphs of larger body mass (see discussion in McPhee et al. 2015b). This, coupled with the relative morphological homogeneity of the LEF sauropodomorph assemblage (see below), undoubtedly contributed to past assertions of a single taxonomic occurrence (“Euskelosaurus”; Van Heerden 1979; Cooper 1980; Kitching and Raath 1984), although occasionally a specimen would be assigned to the vaguely defined “Melanorosauridae” on the grounds of either size, “robustness”, or a relatively derived character set (Charig et al. 1965; Raath 1972; Galton 1985a; see also “Blikanasauridae”, Galton and Van Heerden 1985, 1998).
In addition to the stratigraphic repositioning of Antetonitrus (see below), a major result of the present study is the demonstration that one of the key specimens referred to Melanorosaurus, NMQR 3314, is also from the UEF. Primarily of interest due to its preservation of a complete skull (Yates 2007b), the taxonomic affinity of this specimen has already come into question due to noted anatomical inconsistencies with NMQR 1551 (McPhee et al. 2015b). That this specimen is also stratigraphically inconsistent with both the Melanorosaurus syntype and NMQR 1551 provides robust grounds for the removal of NMQR 3314 from the Melanorosaurus hypodigm, and its revised taxonomic relationships should be treated independently in a future study. The issues surrounding NMQR 3314 are part of a broader series of concerns pertaining to the validity of Melanorosaurus. This uncertainty is due primarily to a highly problematic syntype series (SAM-PK 3449, 3450) which presents potentially unresolvable issues relating to inter-association and a lack of explicitly diagnosable characters (see Galton et al. 2005; Yates 2007b; McPhee et al. 2015b). The future erection of a neotype (e.g., NMQR 1551) may therefore be necessary to safeguard the validity of Melanorosaurus.
Our stratigraphic repositioning of NMQR 3314 has three major corollaries: (i) there is now a complete absence of sauropodomorph cranial material from the LEF; (ii) the basal ornithischian Eocursor (Butler et al. 2007), which was collected from within a few meters of NMQR 3314, is also an UEF taxon, removing any record of Ornithischia from the Late Triassic of South Africa (see also Olsen et al. 2011); and (iii) Yates’ (2008) documentation of a specimen of Blikanasaurus (BP/1/5271a) at Damplaats potentially draws this taxon across the LEF–UEF boundary (given the predominance of UEF outcropping on the farm). Although the second point is of clear interest, removing as it does one of the few Triassic occurrences for Ornithischia, any discussion of ornithischian biostratigraphy is beyond the scope of the current contribution, and will not be addressed further. Given that Blikanasaurus is one of the more distinctive LEF taxa, its potential presence at Damplaats warrants further discussion. BP/1/5271a is part of an assemblage of surface float probably collected from throughout erosional gully at Damplaats (Yates 2008). The LEF at Damplaats is restricted to the lowermost several meters of the exposure (see above). If this specimen came from a position higher in the section, then the presence of a sauropodomorph of similarly robust proportions to Blikanasaurus within the UEF is a possibility. Although the absence of precise provenance information makes any discussion of the stratigraphic relationships of BP1/5271a conjectural, it is worth noting that its manner of preservation is markedly dissimilar to that of NMQR 3314, being a deep purplish-grey (as opposed to red) and lacking any evidence of extensive subaerial exposure. It also lacks the strong hematite coating of other known elements collected from within the UEF at Damplaats (e.g., NMQR 3314, BP/1/6965; see below). This preservational signal is therefore more consistent with LEF fossil material, supporting a collection point from the very base of the Damplaats exposure—and hence from the uppermost sections of the LEF. Our interpretation thus provides tentative support for Yates’ (2008) assertion (based on the observed stratigraphic range for Blikanasaurus) of a relatively homogenous depositional and biostratigraphic scenario for the LEF.
The removal of what was previously the only (mostly) complete, articulated skeleton (NMQR 3314) from the LEF further confounds our understanding of LEF sauropodomorph diversity, with all genera confidently assignable to the subunit problematic with respect to either missing anatomy (Blikanasaurus, Eucnemesaurus, Plateosauravus), questionable inter-association of the type assemblage (Plateosauravus and Melanorosaurus), and diagnosability of the hypodigm (Plateosauravus and Melanorosaurus) (Table 3). This uncertainty makes it difficult to assess the evolutionary significance of the mosaic morphologies exhibited between specific LEF taxa noted by McPhee et al. (2015b); i.e., whether this is explicable via homoplastic convergence, simple intra-generic variation, or incomplete lineage sorting between certain LEF sauropodomorph populations. These caveats aside, at least three different morphotypes can be recognised within the LEF: a distinctive, highly robust form represented by Blikanasaurus (and additional, non-overlapping robust material recently described by McPhee and Choiniere 2016); a generic “prosauropod” grade typified by Eucnemesaurus and Plateosauravus (and possibly Sefapanosaurus); and a relatively derived (i.e., more “sauropod-like”) form represented by the most informative of the referred LEF Melanorosaurus assemblages: NMQR 1551 (Fig. 9). The latter is primarily differentiated from the “prosauropod” grade with respect to its 4-vertebra sacrum with an additional dorsosacral, anteroposteriorly compressed anterior caudal centra, and derived femoral morphology (see Galton et al. 2005). An additional morphotype, roughly intermediate between the first two mentioned above, is potentially represented by the relatively robust (although not to the extreme degree seen in Blikanasaurus) articulated hindlimb and tail collected from Tele River on the same British-South African expedition that recovered Blikanasaurus (SAM-PK 382, “Euskelosaurus”; Charig et al. 1965). This specimen is potentially of a similar taxonomic affinity to the wealth of material excavated from the LEF of Maphutseng, Lesotho between 1956 and 1971 (Ellenberger and Ellenberger 1956; Ellenberger and Ginsburg 1966; Gauffre 1993a; Bordy et al. 2015; Fig. 9), however, this suggestion requires substantiation via indepth re-examination of both assemblages.
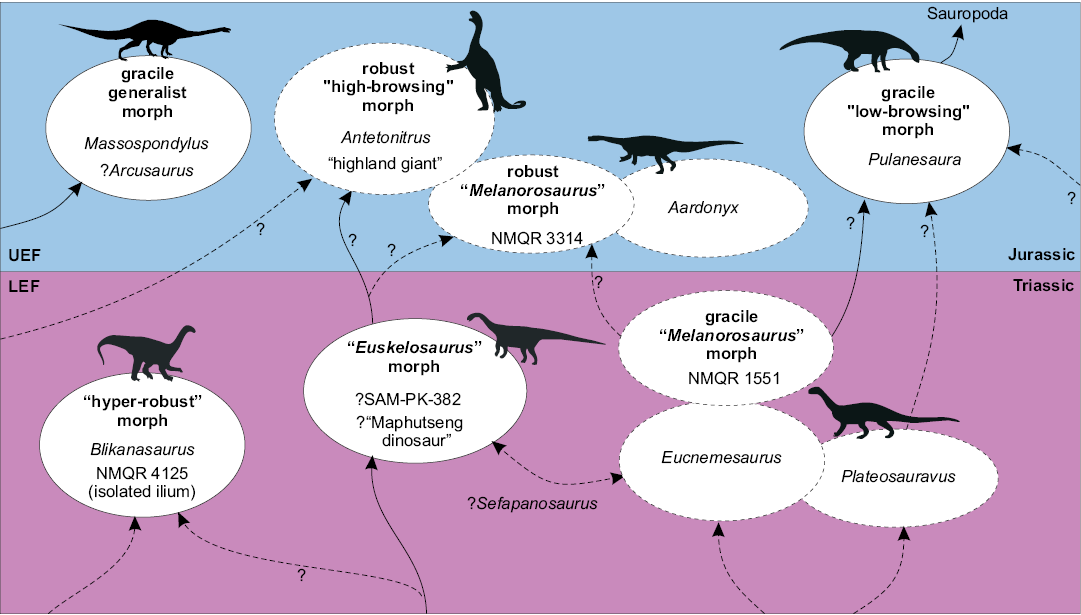
Fig. 9. Schematic representation of hypothetical ecomorphotype groupings and population dynamics of the sauropodomorph fauna of the Elliot Formation.
Table 3. Distinguishing characteristics of lower Elliot Formation sauropodomorph taxa (note that these are not intended as formal diagnoses). Asterisk denotes holotype/syntype.
|
Taxon |
Key specimens |
References |
Notable features |
|
Plateosauravus cullingworthi |
SAM-PK 3341–3356, 3602–3603*; SAM-PK 3607–3609 |
Haughton 1924; Van Heerden 1979; Yates 2003a, 2007a; BWM personal observation |
Lateral swellings at dorsal apex of pectoral vertebrae neural spines. Height of dorsal neural spines over 1.5 times the anteroposterior length of their bases. Humerus:femur ratio = ~0.77. Deltopectoral crest strongly sinuous and located entirely within proximal half of the humerus. Distinct “heel” on the posteroventral corner of the ischial peduncle. |
|
Eucnemesaurus spp. |
TM 119*; BP/1/6107, 6110–6115, 6220; BP/1/6234* |
Van Hoepen 1920; Yates 2007a; McPhee et al. 2015b; BWM personal observation |
Brevis fossa on iliac postacetabular process. Lesser trochanter taller than high with abrupt proximal termination. Fourth trochanter obliquely oriented. Posterior descending process of distal tibia does not extend as far laterally as anterior ascending process (observed only in E. entaxonis) |
|
Melanorosaurus readi |
SAM-PK 3449, 3450*; NMQR 1551 |
Haughton 1924; Yates 2003a; Galton et al. 2005; McPhee et al. 2015b; BWM personal observation |
Hyposphenes on dorsal vertebrae dorsoventrally deep and mediolaterally thin. Four-vertebra sacrum with additional dorsosacral. Centra of anterior caudal vertebrae strongly anteroposteriorly compressed. Shallow radial fossa on proximal ulna. Lesser trochanter located on lateral margin of femoral shaft. Femur markedly straight in anterior/posterior view. Pes relatively elongate (transverse width of proximal end of metatarsal I 0.57 times its total length). |
|
Blikanasaurus cromptoni |
SAM-PK 403*; BP/1/5271a |
Galton and Van Heerden 1985, 1998; Yates 2008; BWM personal observation |
Length of proximal surface of tibia 0.48 times its total length. Posterior descending process of distal tibia does not extend as far laterally as anterior ascending process. Smoothly rounded anteromedial corner of the astragalus. Pes extremely stout (transverse width of proximal end of metatarsal I 0.9 times its total length, metatarsal III 0.36 times the length of the tibia). Mediodorsal corner of proximal surface of metatarsal II strongly distally deflected. Pedal ungual I longer than metatarsal I. |
|
“Euskelosaurus” sp. |
SAM-PK 382 |
Charig et al. 1965; BWM personal observation |
Lesser trochanter located on lateral margin of femoral shaft. Pes stout (transverse width of proximal end of metatarsal I 0.79 times its proximodistal length). Pedal ungual I equal in length to metatarsal I. |
To summarize, the taxonomy of the LEF remains clouded by a lack of articulated specimens, large amounts of missing anatomical data for known taxa, and a lack of detailed reporting on important historically collected specimens (e.g., the Maphutseng assemblage, SAM-PK 382). Nonetheless, the partitioning of LEF sauropodomorph diversity into the three (or potentially four) crude morphotypes outlined above provides a starting point for future investigations into the population dynamics of LEF Sauropodomorpha. Finally, although there is no clear evidence of sauropod-grade taxa within the LEF, the retention of NMQR 1551 (?Melanorosaurus) within the LEF nonetheless indicates the incipient presence of a more derived sauropodomorph bauplan, providing a suggestive transitional link between at least one grade of LEF sauropodomorph and the suite of relatively derived forms that are now present in the UEF. Unfortunately, no taxon exhibits either the clearly delimited stratigraphic range or preservational completeness to provide a replacement eponym for “Euskelosaurus”, and we prefer to leave the LEF biozone unnamed for the time being.
Relevance of the LEF to Late Triassic global sauropodomorph biostratigraphy.—The palaeobiogeographical relationships of the LEF were recently discussed in McPhee et al. (2015b). This study reiterated previously noted similarities between the LEF sauropodomorph assemblage and that of the Argentinian Los Colorados Formation (Pol and Powell 2007; Apaldetti et al. 2012; Ezcurra and Apaldetti 2012; Kent et al. 2014). These observations have generally focused on the close phylogenetic relationships hypothesised between Eucnemesaurus + Riojasaurus and Antetonitrus + Lessemsaurus, with both taxonomic pairings recovered in most recent analyses (e.g., Yates 2007b; McPhee et al. 2015b; Otero et al. 2015). The Los Colorados fauna is restricted to the La Esquina Member (upper third of the formation), with an upper age bound of ~213 Ma (Martinez et al. 2011; Kent et al. 2014). This is consistent with the late Norian/Rhaetian age hypothesised for the LEF. However, the Early Jurassic age now inferred for Antetonitrus (see below) means that it is temporally separated from Lessemsaurus by a stratigraphic interval of upwards of ~15 my. This discrepancy potentially explains the decrease in stratigraphic congruence between our old hypothesis of sauropodomorph biostratigraphy and the one presented here (see above). Although this relationship is supported primarily by apomorphies of their atypically enlarged scapular blades (see McPhee et al. 2014), and it remains possible that this feature is convergent for both taxa, other features, such as the extreme proximodistal shortening of the manus, continue to indicate a close phylogenetic affinity. Additional fossil sampling from the Late Triassic of Argentina and South Africa may eventually resolve this stratigraphic incongruence via the substantiation of a larger “Lessemsauridae” clade extending from the Norian to the Early Jurassic.
Both Plateosauravus and Melanorosaurus bear some similarities with taxa from the late Norian/Rhaetian of Europe. The former is regularly resolved in a similar phylogenetic position (i.e., just outside of, or at the very base of, Plateosauria sensu Yates 2007b) to taxa from the German Keuper (i.e., Plateosaurus and Ruehleia; Yates 2003b; Moser 2003; McPhee et al. 2014, 2015b; Fig. 6), whereas the latter has been compared with the poorly known Camelotia from the English Westbury Formation (Galton 1985a, 1998). This lends further support to a similar age for the LEF. Blikanasaurus is often resolved in a relatively derived position close to Early Jurassic sauropododiform taxa such as Aardonyx and Anetonitrus. However, the persistent incompleteness of this taxon severely limits its contribution to our understanding of Late Triassic sauropodomorph biostratigraphy.
Upper Elliot Formation.—A relatively diverse assemblage of large-bodied, non-massospondylid taxa are now known from the UEF. This increase in taxonomic diversity has been due primarily to successive investigations undertaken on Heelbo (Spion Kop) farm (Yates et al. 2004, 2010, 2011; McPhee et al. 2015a). It was due to the relative abundance of large-bodied taxa discovered on Heelbo, in conjunction with the apparent absence of material referable to Massospondylus, that led Yates et al. (2011) to hypothesise the presence of a unique micro-climate/ecosystem upon the upper Elliot floodplain. Although the faunal assemblage present on the farm appears relatively atypical when compared to other known UEF localities (e.g., Bramley’s Hoek, Nova Barletta), subsequent investigation conducted by the current authors suggests that large-bodied sauropodomorph taxa had a much greater distribution throughout the UEF. Evidence in support of this position includes: A large, articulated hindlimb and posterior vertebral series from an indeterminate sauropodomorph excavated from near the UEF–Clarens Formation contact of the Barkley East district of the Eastern Cape (AM 6147); a yet-to-be-named massive sauropodomorph (the “Highland Giant”, BP/1/7120) from the UEF of the South Africa-Lesotho border near Clarens, Free State (own unpublished data); and the recent discovery of an anterior caudal vertebra (BP/1/6965) referable to Pulanesaura within the uppermost part of the Damplaats section.
Additional evidence for the presence of relatively derived, large-bodied taxa within the UEF also extends from our revised stratigraphic position of the Antetonitrus type locality. This suggests that several relatively derived sauropodomorph characters, previously thought to originate within the Late Triassic, do not appear prior to the earliest Jurassic (e.g., dorsal neural spines well over 1.5 times as high as anteroposteriorly long (cf. Plateosauravus); development of sheet-like spinopostzygapophyseal laminae in posterior dorsal vertebrae; transverse compression of the proximal tibia). Until recently, this also would have had major implications for the timing of the origins of Sauropoda (see below), leaving only Isanosaurus (Buffetaut et al. 2000; see also Lessemsaurus, Pol and Powell 2007) from Thailand as the remaining putative sauropod from the Late Triassic. However, more recent research has questioned both the sauropodan affinity of Antetonitrus (McPhee et al. 2014, 2015a), as well as the Late Triassic age of Isanosaurus (Racey and Goodall 2009; McPhee et al. 2015a).
The UEF (and, by extension, the earliest Jurassic) therefore presents a much more dynamic palaeoecological scenario than inferred by previous assessments of UEF sauropodomorph diversity, which tended to argue that the progressive aridification of the Stormberg sequence favoured generalist taxa such as Massospondylus (e.g., Barrett 2000). In contrast, the decline in large-bodied, non-massospondylid taxa was hypothesised as either being due to a preference for highly restricted, mesic subenvironments (Barrett 2009; Yates et al. 2011), or as a form of regional abandonment in favour of more temperate, high latitude climes (McPhee et al. 2014). However, although from a purely numerical perspective it is clear that certain environmental trends within the earliest Jurassic favoured the radiation of small-bodied massospondylids, the equally broad distribution of large-bodied taxa across the upper Elliot floodplain suggests that a drier climate, instead of negatively impacting sauropodiform diversity, may have encouraged the generation of biomechanical and palaeoecological novelty (see below). This possibility was discussed recently by McPhee et al. (2015a), who suggested that the earliest radiation of Sauropoda was coincident with a novel locomotory strategy centred on obligate quadrupedality and a unique form of energetically conservative low-to-mid browsing. The relative abundance of potentially higher-browsing taxa sympatrically (or parapatrically) associated with Pulanesaura (e.g., Antetonitrus, Aardonyx, the “Highland Giant”) suggests that this diversity of forms is a product of the fine-scale partitioning of niche space in a deteriorating (fluvially speaking) environment—a possibility that warrants future investigation (see also below).
The above review of UEF sauropodomorph diversity suggests occupation by three different ecomorphotypes: an abundant, gracile (and potentially omnivorous; Barrett 2000) form represented primarily by Massospondylus; a physically robust, facultatively bipedal grade of basal sauropodiforms represented by Aardonyx and Antetonitrus (and possibly the undescribed “Highland Giant” and NMQR 3314); and a comparatively gracile, obligate quadruped represented by Pulanesaura (Fig. 9). The latter taxon may also be the earliest sauropod currently known, although anatomical incompleteness at this part of the tree makes substantiation of this suggestion difficult (McPhee et al. 2015a). Yates et al. (2004) also raised the possibility of Vulcanodon-grade sauropods within the UEF based on a single anterior caudal vertebra found during the initial exploration of Heelbo. However, of the two proposed apomorphies reported for this element one (ventral sulcus of the centrum) is now known from a wide range of basal sauropodomorphs (e.g., Lufengosaurus, Melanorosaurus, Leyesaurus) and is potentially variable within genera (and even within the caudal series of a single individual, cf. Camelotia; BWM personal observations 2014), whereas the other (presence of a hyposphene) has been documented in specimens of Melanorosaurus (Yates et al. 2004) as well as being present in Pulanesaura. We therefore tentatively suggest that BP/1/6105 (Yates et al. 2004) is another specimen of Pulanesaura, or a closely related taxon. Although our knowledge of the range and relative abundance of larger-bodied animals within the UEF is clearly increasing, Massospondylus (or a putative clade of massospodylids) nonetheless remains the most abundant taxon throughout the subunit (and possibly within the Clarens Formation [= Forest Sandstone of Zimbabwe], which is still to produce any large-bodied sauropodiform taxa). We therefore retain the original biozonation of the upper Elliot and Clarens formations proposed by Kitching and Raath (1984): the Massospondylus Range Zone.
Relevance of the UEF to Early Jurassic global sauropodomorph biostratigraphy.—The UEF shows marked similarities with the Lufeng Formation of China. The latter contains a diverse assemblage of sauropodomorphs ranging from possible massospondylids (Lufengosaurus), sauropodiforms (e.g., Yunnanosaurus, Jingshanosaurus, Xingxiulong; although the latter may represent a more basal taxon, see Fig. 6), and indeterminate taxa potentially representing basal members of Sauropoda (e.g., Kunmingosaurus) (Young 1941, 1942; Barrett 1999; McPhee et al. 2016: fig. 1; Wang et al. 2017). A similar, if less well-sampled, fauna appears to have also been present within the the Upper Dharmaram Formation of India (Kutty et al. 2007; Novas et al. 2011). These deposits, along with the UEF, highlight the rapid diversification and respective specialization of various sauropodomorph lineages occurring in the earliest Jurassic—a phenomenon that appears to have been global in character. A similarly global radiation of the possibly monophyletic Massospondylidae sensu Yates (2007b) also appears to have taken within the earliest Jurassic, with members of this putative clade found in Antarctica (Smith and Pol 2007), South America (Martinez 2009; Apaldetti et al. 2011), China (Young 1941), India (Novas et al. 2011), and South Africa. The UEF sauropodomorph assemblage is therefore emblematic of broader evolutionary trends occurring at the outset of the Jurassic.
Disparity and diversity: the effects of the TJB on Sauropodomorpha.—Our revision of the stratigraphic relationships of the Sauropodomorphs of the Elliot Formation has implications for the broader understanding of sauropodomorph evolution across the TJB. Generally considered one of the “big five” crashes in global biodiversity, the effects of the events associated with this boundary on sauropodomorph diversity remain relatively poorly understood. Although the abundance of material known from Early Jurassic deposits in China, South America, and South Africa (e.g., Martinez 2009; Apaldetti et al. 2011; McPhee et al. 2016; Wang et al. 2017) suggests that this transition did little to impede the rapid radiation of Sauropodomorpha that began in the Late Triassic, taxic diversity plots produced by Irmis (2011) and, to a lesser extent, Mannion et al. (2011) suggest some decline for the group in the earliest Jurassic (or possibly the Rhaetian; Barrett and Upchurch 2005). However, as also noted by these authors, this signal is highly sensitive to a combination of sampling bias and an unevenly distributed rock record (to which we might add the potentially erroneous binning of taxa, e.g., Antetonitrus).
Given the problems associated with inferring accurate taxonomic diversity (abundance) measures from a highly incomplete fossil record (see further discussion in e.g., Mannion et al. 2011; Butler et al. 2011; Upchurch et al. 2011), other means of investigating macroevolutionary trends have become increasingly popular in recent years. Of these, morphological diversity (disparity) and evolutionary rates analyses have received perhaps the greatest attention (e.g., Brusatte et al. 2008, 2014; Butler et al. 2012; Benson et al. 2014; Lloyd 2016). The former quantifies the degree of bauplan similarity between organisms, whereas the (much more methodologically complex) latter isolates significant deviations from the average tempo of trait acquisition (of e.g., discrete characters or size estimates) across the nodes and branches of a time-calibrated cladogram. Our analysis of sauropodomorph disparity across the TJB is in accordance with our more general taxonomic observations: First, our results clearly demonstrate that UEF sauropodomorph taxa are somewhat more morphologically disparate than those of the LEF. Second, sauropodomorph morphological disparity occupies a greater total range of morphospace within the first ~15 million years of the Jurassic than in the final 15 million years of the Triassic (Fig. 8; Table 2). Given that these two crude time bins almost certainly represent a simplification of the true temporal ranges of the taxa in question, it is possible that improved precision in age constraints may increase this gap.
The increase in sauropodomorph disparity (and potentially generic abundance) is intriguing, given the dramatic turnover in global faunas generally inferred at the TJB. Previous hypotheses related the marked increase in dinosaurian diversity (as well as body size threshold) to ideas of “ecological release” following the extinction of incumbent large-bodied (non-crocodylomorph) crurotarsans and (non-cynodont) synapsids at the end of the Triassic (e.g., Olsen et al. 2002; see also Brusatte et al. 2008). Our results can therefore be interpreted as lending some support to macroevolutionary theories positing the expedited filling of available morpho/niche-space following events associated with the TJB. However, the degree to which the ultimate success of Sauropodomorpha was dependent on this “release” remains to be shown. Although several lines of research have demonstrated the tendency for surviving faunas to dramatically (re)expand within morphospace directly following a biodiversity crisis (see Erwin 2008 for review; Halliday and Goswami 2016), other research into the effects of mass-extinction on terrestrial faunas (e.g., anomodonts) suggests that survivorship is not necessarily followed by a concomitant increase in phenotypic novelty (Ruta et al. 2013). Furthermore, the temporal proximity of the UEF to the TJB (with the latter potentially located at the base of the former, see above) raises questions regarding the time-dependent sensitivity of these metrics to adequately express post-crisis morphospace expansion so soon after the event—although the slightly higher values within the UEF suggests that the ETE did little to negatively impact sauropodomorph populations.
Other lines of evidence indicate that the adaptive radiation of Sauropodomorpha took place against the staggered decline/extinction of specific herbivore communities close to the Carnian–Norian boundary (i.e., rhynchosaurs and dicynodonts), followed by non-crocodylomorph crurotarsans towards (or at) the end of the Triassic (Benton 1983, 1986; Brusatte et al. 2010; Langer et al. 2010). This suggests a more complex scenario of iterative competition and contingency whereby the vacation (or extirpation) of existing communities from their respective browsing niches encouraged the progressive development of catholic macroherbivory in sauropodomorphs, in turn limiting feeding opportunities for other contemporaneous herbivores. This is consistent with current evidence which strongly suggests that sauropodomorphs already dominated the large-bodied herbivore guild prior to the end of the Triassic. In contrast, non-dinosaurian herbivorous archosaurs i.e., aetosaurs, are either relatively rare (if not entirely unknown) in the Late Triassic deposits in which sauropodomorphs are common (e.g., the Elliot Formation), or clearly occupied a different dietary niche (e.g., Neoaetosauroides, Los Colorados Formation; Desojo and Vizcaíno 2009) (see Barrett et al. 2011 for further discussion). Although the precise dynamics at play in the ascendency of Sauropodomorpha are likely to remain largely speculative, it is nonetheless clear that the major suite of sauropodomorph morphological and palaeoecological novelties were already well established prior to the end of the Triassic (Barrett and Upchurch 2007; Langer et al. 2010; Brusatte et al. 2010).
It is worth emphasising, at this point, the singular nature of the sauropodomorph bauplan, which has no obvious analogue, either before or since. Although dinosaurs are generally thought to have been relatively minor components of many early Late Triassic ecosystems (Brusatte et al. 2008, 2010; Langer et al. 2010; Martinez et al. 2011), from its origins ~225 Ma the sauropodomorph bauplan rapidly established itself as a highly successful evolutionary innovation. This is evidenced not only by the numerical dominance of sauropodomorphs in the majority of terrestrial deposits from the mid-Norian onwards, but by the subsequent 150 million years in which this bauplan provided the physical blueprint for the evolution of the Earth’s largest terrestrial animals. In this respect, the unabated radiation of Sauropodomorpha across the TJB, as well as its continued diversification in the Early Jurassic, was potentially promoted by the acquisition of one or more “key innovations” (Hunter 1998; Galis 2001). While the acquisition of a herbivorous diet has already been proposed as a possible key innovation contributing to the relative numerical dominance of Sauropodomorpha in the Late Triassic (Barrett et al. 2011), a host of other (morphological) features are also likely to have played a role (e.g., small head to body ratio, lack of mastication, long neck, extensive post-cranial pneumaticity, habitual/facultative quadrupedality; see Barrett and Upchurch 2007; Sander et al. 2011; McPhee et al. 2015a). In this scenario, an integrated suite of dietary and biomechanical novelties potentially buffered the group from the effects of the end-Triassic extinction (possibly applicable to Dinosauria as a whole), while also providing ripe selective terrain for continued niche space exploration in response to the changing climatic conditions of the earliest Jurassic (see above).
The above hypothesis is undoubtedly sensitive to continued taxon-sampling and further refinement of taxon age-ranges. Of primary interest are the effects of the extinction of the “prosauropod” grade at the end of the Pliensbachian on general sauropodomorph disparity, with the basal divergence of the derived sauropod bauplan certainly contributing to the increased sum of sauropodomorph disparity ranges in the earliest Jurassic. Unfortunately (at least compared to the latest Triassic and earliest Jurassic), the transition from the Early to the Middle Jurassic is poorly represented in the fossil record, with our current assemblage of (unequivocal) basal sauropods probably representing a fraction of the true diversity. The progressive development of statistical methods designed to adjust for missing data (i.e., when correlated with the available rock record, see Mannion et al. 2011), as well as continued fossil sampling, will hopefully ameliorate this situation in the near future. Nonetheless, the current study provides a relatively robust insight into sauropodomorph evolutionary dynamics proximate to the TJB. With reference to the above, we emphasize a scenario whereby ecological drivers associated with environmental changes in the earliest Jurassic encouraged the continued diversification of an already highly successful clade, with the end-Triassic extinction event playing an important, if not decisive, role in that success (see further discussion in Langer et al. 2010: 78–82).
Conclusions
Our review of the basal sauropodomorph record of South Africa’s Elliot Formation overturns recent consensus that the LEF contains the greater taxonomic wealth, contrasting with a relatively “depauperate” UEF typified primarily by Massospondylus remains. Instead, the LEF contains a handful of poorly known forms (due primarily to their fragmentary and/or disassociated condition) that are not easily distinguishable with respect to either character diagnoses or morphospace occupation. This situation is compounded by the stratigraphic repositioning of the important transitional sauropodiform Antetonitrus. However, the observation of at least three coarse morphotypes suggests that our understanding of the taxonomic and morphological content of the LEF will improve with only a modest amount of additional in situ fossil sampling. Moreover, the presence of the enigmatic Blikanasaurus within the LEF represents an intriguing deviation from the relatively homogenous LEF sauropodomorph assemblage.
In comparison, the UEF is now known to contain a much greater diversity of sauropodomorphs than previously appreciated, both in terms of taxic abundance and morphological diversity. This observation, primarily the fruit of work conducted within the past decade, is underscored by the three discrete ecomorphotypes recognised here, one of which potentially represents the basal-most divergence of Sauropoda. This taxonomic range suggests that progressive aridification of the Elliot Formation, instead of restricting sauropodomorph diversification, may have encouraged the exploration of novel niche-space. The increase in taxonomic contribution to the UEF is also reflected in a concomitant increase in sauropodomorph morphological disparity in the earliest Jurassic.
These results thus allow for a more nuanced appreciation of sauropodomorph evolutionary dynamics across the TJB, with current evidence suggesting that the end-Triassic extinction event contributed, in part, to the continued diversification of Sauropodomorpha in the earliest Jurassic. However, it also closed a period (beginning in the Norian) in which the sauropodomorph bauplan evolved many of its characteristic features, ultimately becoming the dominant phenotype within the large-bodied herbivore guild of many Late Triassic ecosystems. Thus, the increased morphological diversity observable within the UEF is interpreted as a combination of the opening-up of niche space following the events of the TJB, as well as a reflection of changing climatic and ecological drivers within the Early Jurassic. Nonetheless, increased taxon sampling and improved chronostratigraphic control is required to further corroborate the hypotheses presented here, and it remains plausible that the oft-discussed “extinction” was more staggered than generally appreciated. In this respect, the Elliot Formation remains a key index formation for correlating international faunas proximate to the TJB—both sauropodomorphan and otherwise.
Acknowledgments
We would like to extend our sincere gratitude to all the farmers and landowners of the Karoo who warmly granted us entry onto their property throughout the course of this fieldwork. For assistance in both lab and field, BWM would like to thank Kimi Chapalle, Kathleen Dollman, Pia Viglietti, Katherine Clayton, Cory Dinter, Casey Staunton (all BP), and Natasha Phillips (Centre for Archaeological Science, University of Wollongong, Australia). Reviews by Paul Barrett (Natural History Museum, London, UK) and Fabian Knoll (Fundación Conjunto Paleontológico de Teruel-Dinópolis, Teruel, Spain) contributed meaningfully to the overall quality of this study. The support of the DST-NRF Centre of Excellence in Palaeosciences (CoE in Palaeosciences) towards this research is hereby acknowledged. Funding from the Palaeontological Scientific Trust (PAST) (to BWM) and the Royal Society (to JNC) made possible the fieldwork conducted during the course of this research. Support for BWM in RSA was provided by an African Origins (NRF) post-doctoral bursary to JNC, and in Brazil by a FAPESP post-doctoral research grant. JNC acknowledges the support of the Friedel Sellschop Award. Ongoing support from both the University of the Witwatersrand and the University of Cape Town is gratefully acknowledged.
References
Abdala, F., Damiani, R., Yates, A., and Neveling, J. 2007. A non-mammaliaform cynodont from the Upper Triassic of South Africa: a therapsid Lazarus taxon? Palaeontologia Africana 42: 17–23.
Anderson, J.M., Anderson, H.M., and Cruickshank, A.R. 1998. Late Triassic ecosystems of the Molteno/Lower Elliot biome of southern Africa. Palaeontology 41: 387–412.
Apaldetti, C., Martinez, R.N., Alcober, O.A., and Pol, D. 2011. A New Basal Sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina. PLoS ONE 6 (11): e26964. Crossref
Apaldetti, C., Martinez, R.N., Pol, D., and Souter, T. 2014. Redescription of the skull of Coloradisaurus brevis (Dinosauria, Sauropodomorpha) from the Late Triassic Los Colorados Formation of the Ischigualasto-Villa Union Basin, northwestern Argentina. Journal of Vertebrate Paleontology 34: 1113–1132. Crossref
Apaldetti, C., Pol, D., and Yates, A. 2012. The postcranial anatomy of Coloradisaurus brevis (Dinosauria: Sauropodomorpha) from the late Triassic of Argentina and its phylogenetic implications. Palaeontology 2012: 1–25.
Barrett, P.M. 1999. A sauropod dinosaur from the lower Lufeng Formation (Lower Jurassic) of Yunnan province, People’s Republic of China. Journal of Vertebrate Paleontology 19: 785–787. Crossref
Barrett, P.M. 2000. Prosauropod dinosaurs and iguanas: speculations on the diets of extinct reptiles. In: H.-D. Sues (ed.), Evolution of herbivory in terrestrial vertebrates: perspectives from the fossil record, 42–78. Cambridge University Press, Cambridge. Crossref
Barrett, P.M. 2004. Sauropodomorph dinosaur diversity in the upper Elliot Formation (Massospondylus range zone: Lower Jurassic) of South Africa: research letter. South African Journal of Science 100: 501–503.
Barrett, P.M. 2009. A new basal sauropodomorph dinosaur from the upper Elliot Formation (Lower Jurassic) of South Africa. Journal of Vertebrate Paleontology 29: 1032–1045. Crossref
Barrett, P.M. and Upchurch, P. 2005. Sauropodomorph Diversity through Time: paleoecological and macroevolutionary implications. In: K.A. Curry Rogers and J.A. Wilson (eds.), The Sauropods: Evolution and Paleobiology, 125–126. University of California Press, Berkeley.
Barrett, P.M. and Upchurch, P. 2007. The evolution of feeding mechanisms in early sauropodomorph dinosaurs. Special Papers in Palaeontology 77: 91–112.
Barrett, P.M., Butler, R.J., and Nesbitt, S.J. 2011. The roles of herbivory and omnivory in early dinosaur evolution. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 383–396. Crossref
Bell, M.A. and Lloyd, G.T. 2015. Strap: an R package for plotting phylogenies against stratigraphy and assessing their stratigraphic congruence. Palaeontology 58: 379–389. Crossref
Benson, R.B.J., Campione, N.E., Carrano, M.T., Mannion, P.D., Sullivan, C., Upchurch, P., and Evans, D.C. 2014. Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage. PLOS Biology 12 (5): e1001853. Crossref
Benton, M.J. 1983. Dinosaur success in the Triassic: a noncompetitive ecological model. The Quarterly Review of Biology 58: 29–55. Crossref
Benton, M.J. 1986. The Late Triassic tetrapod extinction events. In: K. Padian (ed.), The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic-Jurassic boundary, 303–320. Cambridge University Press, Cambridge.
Benton, M.J. 1994. Palaeontological data and identifying mass extinctions. Trends in Ecology & Evolution 9: 181–185. Crossref
Benton, M.J. 1995. Diversification and extinction in the history of life. Science 268: 52. Crossref
Benton, M.J., Juul, L., Storrs, G.W., and Galton, P.M. 2000. Anatomy and systematics of the prosauropod dinosaur Thecodontosaurus antiquus from the upper Triassic of southwest England. Journal of Vertebrate Paleontology 20: 77–108. Crossref
Blackburn, T.J., Olsen, P.E., Bowring, S.A., McLean, N.M., Kent, D.V., Puffer, J., McHone, G., Rasbury, E.T., and Et-Touhami, M. 2013. Zircon U-Pb geochronology links the end-Triassic extinction with the Central Atlantic Magmatic Province. Science 340: 941–945. Crossref
Bonnan, M.F. and Yates, A.M. 2007. A new description of the forelimb of the basal sauropodomorph Melanorosaurus: implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs. Special Papers in Palaeontology 77: 157–168.
Bordy, E.M. and Eriksson, P. 2015. Lithostratigraphy of the Elliot Formation (Karoo Supergroup), South Africa. South African Journal of Geology 118: 311–316. Crossref
Bordy, E.M., Hancox, P.J., and Rubidge, B.S. 2004a. Basin development during the deposition of the Elliot Formation (Late Triassic–Early Jurassic), Karoo Supergroup, South Africa. South African Journal of Geology 107: 397–412. Crossref
Bordy, E.M., Hancox, P.J., and Rubidge, B.S. 2004b. Fluvial style variations in the Late Triassic–Early Jurassic Elliot Formation, main Karoo Basin, South Africa. Journal of African Earth Sciences 38: 383–400. Crossref
Bordy, E.M., Hancox, P.J., and Rubidge, B.S. 2005. The contact of the Molteno and Elliot formations through the main Karoo Basin, South Africa: a second-order sequence boundary. South African Journal of Geology 108: 351–364. Crossref
Bordy, E.M., Sciscio, L., Abdala, F., McPhee, B.W., and Choiniere, J.N. 2017. First Lower Jurassic vertebrate burrow from southern Africa (upper Elliot Formation, Karoo Basin, South Africa). Palaeogeography, Palaeoclimatology, Palaeoecology 468: 362–372. Crossref
Bordy, E.M., Van Gen, J., Tucker, R., and McPhee, B.W. 2015. Maphutseng fossil heritage: Stratigraphic context of the dinosaur trackways and bone bed in the Upper Triassic–Lower Jurassic Elliot Formation (Karoo Supergroup, Lesotho). In: H. Saber, A. Lagnaoui, and A. Belahmira (eds.), First International Congress on Continental Ichnology [ICCI-2015], Proceeding Abstracts, 67. Chouaïb Doukkali University, El Jadida
Broom, R. 1911. On the dinosaurs of the Stormberg, South Africa. Annals of the South African Museum 7: 291–308.
Brusatte, S.L., Benton, M.J., Ruta, M., and Lloyd, G.T. 2008. Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science 321 (5895): 1485–1488. Crossref
Brusatte, S.L., Lloyd, G.T., Wang, S.C., and Norell, M.A. 2014. Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition. Current Biology 24: 2386–2392. Crossref
Brusatte, S.L., Nesbitt, S.J., Irmis, R.B., Butler, R.J., Benton, M.J., and Norell, M.A. 2010. The origin and early radiation of dinosaurs. Earth-Science Reviews 101: 68–100. Crossref
Buffetaut, E., Suteethorn, V., Cuny, G., Tong, H., Le Loeuff, J., Khansubha, S., and Jongautchariyakul, S. 2000. The earliest known sauropod dinosaur. Nature 407: 72–74. Crossref
Butler, R.J. 2005. The “fabrosaurid” ornithischian dinosaurs of the upper Elliot Formation (Lower Jurassic) of South Africa and Lesotho. Zoological Journal of the Linnean Society 145: 175–218. Crossref
Butler, R.J., Benson, R.B., Carrano, M.T., Mannion, P.D., and Upchurch, P. 2011. Sea level, dinosaur diversity and sampling biases: investigating the “common cause” hypothesis in the terrestrial realm. Proceedings of the Royal Society of London B: Biological Sciences 278: 1165–1170. Crossref
Butler, R.J., Brusatte, S.L., Andres, B., and Benson, R.B. 2012. How do geological sampling biases affect studies of morphological evolution in deep time? A case study of pterosaur (Reptilia: Archosauria) disparity. Evolution 66: 147–162. Crossref
Butler, R.J., Smith, R.M.H., and Norman, D.B. 2007. A primitive ornithischian dinosaur from the Late Triassic of South Africa, and the early evolution and diversification of Ornithischia. Proceedings of the Royal Society B: Biological Sciences 274: 2041–2046. Crossref
Butler, R.J., Yates, A.M., Rauhut, O.W., and Foth, C. 2013. A pathological tail in a basal sauropodomorph dinosaur from South Africa: evidence of traumatic amputation? Journal of Vertebrate Paleontology 33: 224–228. Crossref
Catuneanu, O., Hancox, J.P., and Rubidge, B.S. 1998. Reciprocal flexural behaviour and contrasting stratigraphies: a new basin development model for the Karoo retroarc foreland system, South Africa. Basin Research 10: 417–439. Crossref
Charig, A.J., Attridge, J., and Crompton, A.W. 1965. On the origin of the sauropods and the classification of the Saurischia. Proceedings of the Linnean Society of London 176: 197–221. Crossref
Chen, P.-J., Li, J., Matsukawa, M., Zhang, H., Wang, Q., and Lockley, M.G. 2006. Geological ages of dinosaur-track-bearing formations in China. Cretaceous Research 27: 22–32. Crossref
Choiniere, J., Benson, R.B.J., Irmis, R.B., and Jinnah, Z. 2014. Volcanic tuffs from the Elliot Formation: Implications for the Triassic-Jurassic terrestrial boundary. Journal of Vertebrate Paleontology 34: 108A.
Cohen, K.M., Finney, S.C., Gibbard, P.L., Fan, J.-X. 2013. The ICS International Chronostratigraphic Chart. Episodes 36: 199–204. http://www.stratigraphy.org/ICSchart/ChronostratChart2016-12.pdf
Cole, D.I. 1992. Evolution and development of the Karoo Basin. In: M.J. de Witt and I.G.D. Ransome (eds.), Inversion Tectonics of the Cape Fold Belt, Karoo and Cretaceous Basins of Southern Africa, 87–99. A.A. Balkema, Rotterdam.
Cooper, M.R. 1980. The first record of the prosauropod dinosaur Euskelosaurus from Zimbabwe. Arnoldia Zimbabwe 9 (3): 1–17.
Cooper, M.R. 1981. The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance. Occasional Papers, National Museums and Monuments of Rhodesia, Series B 6: 689–840.
de Fabrègues, C.P. and Allain, R. 2016. New material and revision of Melanorosaurus thabanensis, a basal sauropodomorph from the Upper Triassic of Lesotho. PeerJ 4: e1639. Crossref
Desojo, J.B. and Vizcaíno, S.F. 2009. Jaw biomechanics in the South American aetosaur Neoaetosauroides engaeus. Paläontologische Zeitschrift 83: 499. Crossref
Ellenberger, F. and Ellenberger, P. 1956. Le gisement de dinosauriens de Maphutseng (Basutoland, Afrique du sud). Comptes Rendus Sommaires de la Societe Geologique de France 8: 99–101.
Ellenberger, F. and Ginsburg, L. 1966. Le gisement de dinosauriens de Maphutseng (Basutoland, Afrique du sud) et l’origine des sauropodes. Comptes Rendus de l’Académie des Sciences, Paris 262: 444–447.
Ellenberger, F., Ellenberger, P., and Ginsburg, L. 1969. The appearance and evolution of dinosaurs in the Trias and Lias: a comparison between South African Upper Karroo and Western Europe based on vertebrate footprints. In: A.J. Amos (ed.), Gondwana Stratigraphy: IUGS Symposium Paris, 333–354. UNESCO, Buenos Aires.
Ellenberger, P. 1970. Establissement de Zones Straigraphiques Détaillées dans le Stromberg du Lesotho (Afrique du Sud) (Trias Supérieur à Jurassique). In: S.H. Haughton (ed.), Proceedings and Papers of the Second Gondwanaland Symposium, 343–370. Council for Scientific and Industrial Research, Pretoria.
Erwin, D.H. 2008. Extinction as the loss of evolutionary history. Proceedings of the National Academy of Sciences 105 (Supplement 1): 11520–11527. Crossref
Ezcurra, M.D. and Apaldetti, C. 2012. A robust sauropodomorph specimen from the Upper Triassic of Argentina and insights on the diversity of the Los Colorados Formation. Proceedings of the Geologists’ Association 123 (1): 155–164. Crossref
Foffa, D., Whiteside, D.I., Viegas, P.A., and Benton, M.J. 2014. Vertebrates from the Late Triassic Thecodontosaurus-bearing rocks of Durdham Down, Clifton (Bristol, UK). Proceedings of the Geologists’ Association 125: 317–328.
Galis, F. 2001. Key innovations and radiations. In: G.P. Wagner (ed.), The Character Concept in Evolutionary Biology, 581–605. Academic Press, San Diego. Crossref
Galton, P.M. 1985a. Notes on the Melanorosauridae, a family of large Prosauropod Dinosaurs (Saurischia: Sauropodomorpha). Geobios 18: 671–676. Crossref
Galton, P.M. 1985b. The poposaurid thecodontian Teratosaurus suevicus v. Meyer, plus referred specimens mostly based on prosauropod dinosaurs, from the Middle Stubensandstein (Upper Triassic) of Nordwurttemberg. Stuttgart Beitrage zur Naturkunde (B) 116: 1–29.
Galton, P.M. 1998. Saurischian dinosaurs from the Upper Triassic of England: Camelotia (Prosauropoda, Melanorosauridae) and Avalonianus (Theropoda, ?Carnosauria). Palaeontographica Abteilung A 250: 155–172.
Galton, P.M. and Upchurch, P. 2004. Prosauropoda. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria: 2nd Edition, 232–258. The University of California Press, Berkeley. Crossref
Galton, P.M. and Van Heerden, J. 1985. Partial hindlimb of Blikanasaurus cromptoni n. gen. and n. sp., representing a new family of prosauropod dinosaurs from the upper triassic of South Africa. Geobios 18: 509–516. Crossref
Galton, P.M. and Van Heerden, J. 1998. Anatomy of the prosauropod dinosaur Blikanasaurus cromptoni (Upper Triassic, South Africa), with notes on the other tetrapods from the lower Elliot Formation. Paläontologische Zeitschrift 72: 163–177. Crossref
Galton, P.M., Van Heerden, J., and Yates, A.M. 2005. Postcranial anatomy of referred specimens of the sauropodomorph dinosaur Melanorosaurus from the Upper Triassic of South Africa. In: V. Tidwell and K. Carpenter (eds.), Thunder-Lizards: the Sauropodomorph Dinosaurs, 1–37. Indiana University Press, Bloomington.
Gauffre, F.-X. 1993a. Biochronostratigraphy of the Lower Elliot Formation (Southern Africa) and preliminary results on the Maphutseng dinosaur (Saurischia: Prosauropoda) from the same formation of Lesotho. New Mexico Museum of Natural History & Science Bulletin 3: 147–149.
Gauffre, F.-X. 1993b. The most recent Melanorosauridae (Saurischia, Prosauropoda), Lower Jurassic of Lesotho, with remarks on the prosauropod phylogeny. Neues Jahrbuch für Geologie und Paläontologie 11: 648–654.
Gow, C.E., Kitching, J.W., and Raath, M.A. 1990. Skulls of the prosauropod dinosaur Massospondylus carinatus Owen in the collections of the Bernard Price Institute for Palaeontological Research. Palaeontologia Africana 27: 45–58.
Hallam, A. and Wignall, P. 1999. Mass extinctions and sea-level changes. Earth-Science Reviews 48: 217–250. Crossref
Halliday, T.J.D. and Goswami, A. 2016. Eutherian morphological disparity across the end-Cretaceous mass extinction. Biological Journal of the Linnean Society 118: 152–168. Crossref
Haughton, S.H. 1924. The fauna and stratigraphy of the Stormberg Series. Annals of the South African Museum 12: 323–497.
Huene, F. von 1932. Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monographien zur Geologie und Paläontologie 4: 1–361.
Hunter, J.P. 1998. Key innovations and the ecology of macroevolution. Trends in Ecology & Evolution 13: 31–36. Crossref
Huxley, T.H. 1866. On the remains of large dinosaurian reptiles from the Stormberg Mountains, South Africa. Geological Magazine 3: 563.
Irmis, R.B. 2011. Evaluating hypotheses for the early diversification of dinosaurs. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 397–426. Crossref
Kent, D.V., Malnis, P.S., Colombi, C.E., Alcober, O.A., and Martínez, R.N. 2014. Age constraints on the dispersal of dinosaurs in the Late Triassic from magnetochronology of the Los Colorados Formation (Argentina). Proceedings of the National Academy of Sciences 111: 7958–7963. Crossref
Kitching, J.W. and Raath, M.A. 1984. Fossils from the Elliot and Clarens formations (Karoo sequence) of the northeastern Cape, Orange Free State and Lesotho, and a suggested biozonation based on tetrapods. Palaeontologia Africana 25: 111–125.
Knoll, F. 2004. Review of the tetrapod fauna of the “Lower Stormberg Group” of the main Karoo Basin (southern Africa) : implication for the age of the Lower Elliot Formation. Bulletin de la Societe Geologique de France 175: 73–83. Crossref
Knoll, F. 2005. The tetrapod fauna of the Upper Elliot and Clarens formations in the main Karoo Basin (South Africa and Lesotho). Bulletin de la Societe Geologique de France 176: 81–91. Crossref
Knoll, F. 2010. A primitive sauropodomorph from the upper Elliot Formation of Lesotho. Geological Magazine 147: 814–829. Crossref
Kutty, T., Chatterjee, S., Galton, P.M., and Upchurch, P. 2007. Basal sauropodomorphs (Dinosauria: Saurischia) from the Lower Jurassic of India: their anatomy and relationships. Journal of Paleontology 81: 1218–1240. Crossref
Laliberté, E. and Legendre, P. 2010. A distance-based framework for measuring functional diversity from multiple traits. Ecology 91: 299–305. Crossref
Laliberté, E., Legendre, P., Shipley, B., and Laliberté, M.E. 2014. Package “FD”: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version. 28 pp. https://cran.r-project.org/web/packages/FD/FD.pdf
Langer, M.C., Ezcurra, M.D., Bittencourt, J.S., and Novas, F.E. 2010. The origin and early evolution of dinosaurs. Biological Reviews 85: 55–110. Crossref
Lloyd, G.T. 2016. Estimating morphological diversity and tempo with discrete character-taxon matrices: implementation, challenges, progress, and future directions. Biological Journal of the Linnean Society 118: 131–151. Crossref
Lucas, S.G. and Hancox, J.P. 2001. Tetrapod-based correlation of the nonmarine Upper Triassic of southern Africa. Albertiana 25: 5–9.
Lucas, S.G., Tanner, L.H., and Heckert, A.B. 2005. Tetrapod biostratigraphy and biochronology across the Triassic–Jurassic boundary in northeastern Arizona. Vertebrate Paleontology in Arizona. New Mexico Museum of Natural History and Science Bulletin 29: 83–92.
Lucas, S.G., Tanner, L., Donohoo-Hurley, L., Geissman, J., Kozur, H., Heckert, A., and Weems, R. 2011. Position of the Triassic–Jurassic boundary and timing of the end-Triassic extinctions on land: Data from the Moenave Formation on the southern Colorado Plateau, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 302: 194–205. Crossref
Mannion, P.D., Upchurch, P., Carrano, M.T., and Barrett, P.M. 2011. Testing the effect of the rock record on diversity: a multidisciplinary approach to elucidating the generic richness of sauropodomorph dinosaurs through time. Biological Reviews 86: 157–181. Crossref
Martinez, R.N. 2009. Adeopapposaurus mognai, gen. et sp. nov. (Dinosauria: Sauropodomorpha), with comments on adaptations of basal Sauropodomorpha. Journal of Vertebrate Paleontology 29: 142–164. Crossref
Martinez, R.N., Apaldetti, C., Correa, G., Colombi, C.E., Fernández, E., Malnis, P.S., Praderio, A., Abelín, D., Benegas, L.G., and Aguilar-Cameo, A. 2015. A new Late Triassic vertebrate assemblage from northwestern Argentina. Ameghiniana 52: 379–390. Crossref
Martinez, R.N., Sereno, P.C., Alcober, O.A., Colombi, C.E., Renne, P.R., Montañez, I.P., and Currie, B.S. 2011. A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea. Science 331: 206–210. Crossref
McGhee, G.R., Clapham, M.E., Sheehan, P.M., Bottjer, D.J., and Droser, M.L. 2013. A new ecological-severity ranking of major Phanerozoic biodiversity crises. Palaeogeography, Palaeoclimatology, Palaeoecology 370: 260–270. Crossref
McPhee, B.W. and Choiniere, J.N. 2016. A hyper-robust sauropodomorph dinosaur ilium from the Upper Triassic–Lower Jurassic Elliot Formation of South Africa: Implications for the functional diversity of basal Sauropodomorpha. Journal of African Earth Sciences 123: 177–184. Crossref
McPhee, B.W., Bonnan, M.F., Yates, A.M., Neveling, J., and Choiniere, J.N. 2015a. A new basal sauropod from the pre-Toarcian Jurassic of South Africa: evidence of niche-partitioning at the sauropodomorph–sauropod boundary? Scientific Reports 5: 13224. Crossref
McPhee, B.W., Choiniere, J.N., Yates, A.M., and Viglietti, P.A. 2015b. A second species of Eucnemesaurus Van Hoepen, 1920 (Dinosauria, Sauropodomorpha): new information on the diversity and evolution of the sauropodomorph fauna of South Africa’s lower Elliot Formation (latest Triassic). Journal of Vertebrate Paleontology 35: e980504. Crossref
McPhee, B.W., Upchurch, P., Mannion, P.D., Sullivan, C., Butler, R.J., and Barrett, P.M. 2016. A revision of Sanpasaurus yaoi Young, 1944 from the Early Jurassic of China, and its relevance to the early evolution of Sauropoda (Dinosauria). PeerJ 4: e2578. Crossref
McPhee, B.W., Yates, A.M., Choiniere, J.N., and Abdala, F. 2014. The complete anatomy and phylogenetic relationships of Antetonitrus ingenipes (Sauropodiformes, Dinosauria): implications for the origins of Sauropoda. Zoological Journal of the Linnean Society 171: 151–205. Crossref
Moser, M. 2003. Plateosaurus engelhardti Meyer, 1837 (Dinosauria: Sauropodomorpha) aus dem Feuerletten (Mittelkeuper; Obertrias) von Bayern. Zitteliana 24: 3–186.
Novas, F.E., Ezcurra, M.D., Chatterjee, S., and Kutty, T.S. 2011. New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of Central India. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101 (Special Issue 3–4): 333–349.
Olsen, P.E. and Galton, P.M. 1984. A review of the reptile and amphibian assemblages from the Stormberg of southern Africa, with special emphasis on the footprints and the age of the Stormberg. Palaeontologia Africana 25: 87–110.
Olsen, P.E., Kent, D.V., Sues, H.-D., Koeberl, C., Huber, H., Montanari, A., Rainforth, E., Fowell, S., Szajna, M.J., and Hartline, B.W. 2002. Ascent of dinosaurs linked to an iridium anomaly at the Triassic-Jurassic boundary. Science 296: 1305–1307. Crossref
Olsen, P.E., Kent, D.V., and Whiteside, J.H. 2011. Implications of the Newark Supergroup-based astrochronology and geomagnetic polarity time scale (Newark-APTS) for the tempo and mode of the early diversification of the Dinosauria. Earth and Environmental Science Transactions-Royal Society of Edinburgh 101: 201. Crossref
Otero, A., Krupandan, E., Pol, D., Chinsamy, A., and Choiniere, J. 2015. A new basal sauropodiform from South Africa and the phylogenetic relationships of basal sauropodomorphs. Zoological Journal of the Linnean Society 174: 589–634. Crossref
Owen, R. 1854. Descriptive catalogue of the Fossil organic remains of Reptilia and Pisces contained in the Museum of the Royal College of Surgeons of England. 184 pp. Taylor and Francis, London.
Pálfy, J., Mortensen, J.K., Carter, E.S., Smith, P.L., Friedman, R.M., and Tipper, H.W. 2000. Timing the end-Triassic mass extinction: First on land, then in the sea? Geology 28: 39–42. Crossref
Pol, D. and Norell, M.A. 2001. Comments on the Manhattan Stratigraphic Measure. Cladistics 17: 285–289. Crossref
Pol, D. and Norell, M.A. 2006. Uncertainty in the age of fossils and the stratigraphic fit to phylogenies. Systematic Biology 55: 512–521. Crossref
Pol, D. and Powell, J.E. 2007. New information on Lessemsaurus sauropoides (Dinosauria: Sauropodomorpha) from the Upper Triassic of Argentina. Special Papers in Palaeontology 77: 223–243.
Pol, D., Garrido, A., and Cerda, I.A. 2011. A new sauropodomorph dinosaur from the Early Jurassic of Patagonia and the origin and evolution of the sauropod-type sacrum. PLoS ONE 6 (1): e14572. Crossref
Pol, D., Norell, M.A., Siddall, M.E. 2004. Measures of stratigraphic fit to phylogeny and their sensitivity to tree size, tree shape, and scale. Cladistics 20: 64–75. Crossref
R Core Team 2017. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/.
Raath, M.A. 1972. Fossil vertebrate studies in Rhodesia: a new dinosaur (Reptilia: Saurischia) from near the Trias-Jurassic boundary. Arnoldia 5 (30): 1–37.
Racey, A. and Goodall, J.G.S. 2009. Palynology and stratigraphy of the Mesozoic Khorat Group red bed sequences from Thailand. Geological Society of London Special Publication 315: 69–83. Crossref
Raup, D.M. and Sepkoski, J.J. 1982. Mass extinctions in the marine fossil record. Science 215: 1501–1503. Crossref
Reisz, R.R., Evans, D.C., Roberts, E.M., Sues, H.-D., and Yates, A.M. 2012. Oldest known dinosaurian nesting site and reproductive biology of the Early Jurassic sauropodomorph Massospondylus. Proceedings of the National Academy of Sciences 109: 2428–2433. Crossref
Rubidge, B.S., Day, M.O., Barbolini, N., Hancox, P.J., Choiniere, J.N., Bamford, M.K., Viglietti, P.A., McPhee, B.W., and Jirah, S. 2016. Advances in Nonmarine Karoo Biostratigraphy: Significance for Understanding Basin Development. In: B. Linol and M.J. de Wit (eds.), Origin and Evolution of the Cape Mountains and Karoo Basin, 141–149. Springer, Switzerland.
Ruta, M., Angielczyk, K.D., Fröbisch, J., and Benton, M.J. 2013. Decoupling of morphological disparity and taxic diversity during the adaptive radiation of anomodont therapsids. Proceedings of the Royal Society B 280: 20131071. Crossref
Sander, P.M., Christian, A., Clauss, M., Fechner, R., Gee, C.T., Griebeler, E.-M., Gunga, H.-C., Hummel, J., Mallison, H., Perry, S.F., Preuschoft, H., Rauhut, O.W.M., Remes, K., Tütken, T., Wings, O., and Witzel, U. 2011. Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews 86: 117–155. Crossref
Schaltegger, U., Guex, J., Bartolini, A., Schoene, B., and Ovtcharova, M. 2008. Precise U-Pb age constraints for end-Triassic mass extinction, its correlation to volcanism and Hettangian post-extinction recovery. Earth and Planetary Science Letters 267: 266–275. Crossref
Sciscio, L., de Kock, M., Bordy, E.M., and Knoll, F. 2017. Magnetostratigraphy across the Triassic–Jurassic boundary in the main Karoo Basin. Gondwana Research [published online]. Crossref
Sertich, J.J.W. and Loewen, M.A. 2010. A new basal sauropodomorph Dinosaur from the Lower Jurassic Navajo Sandstone of Southern Utah. PLoS ONE 5 (3): e9789. Crossref
Smith, N.D. and Pol, D. 2007. Anatomy of a basal sauropodomorph from the Early Jurassic Hanson Formation of Antarctica. Acta Palaeontologica Polonica 52: 657–674.
Smith, R. and Kitching, J. 1997. Sedimentology and vertebrate taphonomy of the Tritylodon acme zone: a reworked palaeosol in the Lower Jurassic Elliot Formation, Karoo Supergroup, South Africa. Palaeogeography, Palaeoclimatology, Palaeoecology 131: 29–50. Crossref
Storrs, G.W. 1994. Fossil vertebrate faunas of the British Rhaetian (latest Triassic). Zoological Journal of the Linnean Society 112: 217–259. Crossref
Sues, H.-D. and Olsen, P.E. 2015. Stratigraphic and temporal context and faunal diversity of Permian-Jurassic continental tetrapod assemblages from the Fundy rift basin, eastern Canada. Atlantic Geology 51: 139–205. Crossref
Upchurch, P., Mannion, P.D., Benson, R.B., Butler, R.J., and Carrano, M.T. 2011. Geological and anthropogenic controls on the sampling of the terrestrial fossil record: a case study from the Dinosauria. Geological Society of London Special Publications 358: 209–240. Crossref
Van Heerden, J. 1979. The morphology and taxonomy of Euskelosaurus (Reptilia: Saurischia: Late Triassic) from South Africa. Navorsinge Van die Nasionale Museum 4: 21–84.
Van Hoepen, E.C.N. 1920. Contributions to the knowledge of the reptiles of the Karoo Formation. 6. Further dinosaurian material in the Transvaal Museum Annals of the Transvaal Museum 7: 93–140.
Vasconcelos, C. and Yates, A. 2004. Sauropodomorph biodiversity of the upper Elliot Formation (Lower Jurassic) of southern Africa. Geoscience Africa 2004: 670.
Villéger, S., Mason, N.W., and Mouillot, D. 2008. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89: 2290–2301. Crossref
Wang, Y.-M., You, H.-L., and Wang, T. 2017. A new basal sauropodiform dinosaur from the Lower Jurassic of Yunnan Province, China. Scientific Reports 7: 41881. Crossref
Wills, M.A. 1999. Congruence between phylogeny and stratigraphy: randomization tests and the gap excess ratio. Systematic Biology 48: 559–580. Crossref
Wills, M.A., Barrett, P.M., and Heathcote, J.F. 2008. The modified gap excess ratio (GER*) and the stratigraphic congruence of dinosaur phylogenies. Systematic Biology 57: 891–904. Crossref
Yates, A.M. 2003a. A definite prosauropod dinosaur from the lower Elliot Formation (Norian: Upper Triassic) of South Africa. Palaeontologia Africana 39: 63–68.
Yates, A.M. 2003b. The species taxonomy of the sauropodomorph dinosaurs from the Löwenstein Formation (Norian, Late Triassic) of Germany. Palaeontology 46: 317–337. Crossref
Yates, A.M. 2004. The Death of a Dinosaur: dismembering Euskelosaurus. Geoscience Africa: 715.
Yates, A.M. 2005. A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods. Palaeontologia Africana 41: 105–122.
Yates, A.M. 2007a. Solving a dinosaurian puzzle: the identity of Aliwalia rex Galton. Historical Biology 19: 93–123. Crossref
Yates, A.M. 2007b. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria). Special Papers in Paleontology 77: 9–55.
Yates, A.M. 2008. A second specimen of Blikanasaurus (Dinosauria: Sauropoda) and the biostratigraphy of the lower Elliot Formation. Palaeontologia Africana 43: 39–43.
Yates, A.M. and Barrett, P.M. 2010. Massospondylus carinatus Owen 1854 (Dinosauria: Sauropodomorpha) from the Lower Jurassic of South Africa: Proposed conservation of the usage by designation of a neotype. Palaeontologia Africana 45: 7–10.
Yates, A.M. and Kitching, J.W. 2003. The earliest known sauropod dinosaur and the first steps towards sauropod locomotion. Proceedings of the Royal Society of London. Series B: Biological Sciences 270: 1753–1758. Crossref
Yates, A.M. and Vasconcelos, C.C. 2005. Furcula-like clavicles in the prosauropod dinosaur Massospondylus. Journal of Vertebrate Paleontology 25: 466–468. Crossref
Yates, A.M., Bonnan, M.F., and Neveling, J. 2011. A new basal sauropodomorph dinosaur from the Early Jurassic of South Africa. Journal of Vertebrate Paleontology 31: 610–625. Crossref
Yates, A.M., Bonnan, M.F., Neveling, J., Chinsamy, A., and Blackbeard, M.G. 2010. A new transitional sauropodomorph dinosaur from the Early Jurassic of South Africa and the evolution of sauropod feeding and quadrupedalism. Proceedings of the Royal Society B: Biological Sciences 277: 787–794. Crossref
Yates, A.M., Hancox, J.P., and Rubidge, B.S. 2004. First record of a sauropod dinosaur from the Upper Elliot Formation (Early Jurassic) of South Africa. South African Journal of Science 100: 504–506.
Young, C.-C. 1941. A complete osteology of Lufengosaurus huenei Young (gen. et sp. nov.) Palaeontologica Sinica, Series C 7: 1–53.
Young, C.-C. 1942. Yunnanosaurus huangi Young (gen. et sp. nov.), a new Prosauropoda from the Red Beds at Lufeng, Yunnan. Bulletin of the Geological Society of China 22: 63–104.
Acta Palaeontol. Pol. 62 (3): 441–465, 2017
https://doi.org/10.4202/app.00377.2017