

New reticulosan sponges from the middle Cambrian of Sonora, Mexico
MATILDE SYLVIA BERESI, JOSEPH P. BOTTING, JUAN J. PALAFOX, and BLANCA E. BUITRÓN SÁNCHEZ
Beresi, M.S., Botting, J.P., Palafox, J.J., and Buitrón Sánchez, B.E. 2017. New reticulosan sponges from the middle Cambrian of Sonora, Mexico. Acta Palaeontologica Polonica 62 (4): 691–703.
A small assemblage of extremely well-preserved fragments of new sponges has been discovered in calcipelites of the middle Cambrian El Mogallón Formation in the Cerro El Mogallón section, near Arivechi in eastern Sonora, Mexico. The assemblage includes two new reticulosan species referred to Ratcliffespongia arivechensis sp. nov. and Valospongia sonorensis sp. nov., combined with disarticulated remains assigned to Kiwetinokia and additional, currently unidentifiable taxa. The new species represent the first records of these Cambrian genera from Mexico, although they are widely distributed at low latitudes, being previously best known from Utah but extending through Laurentia and South China. This middle Cambrian fauna indicates that there was considerable continuity of the deeper-water hexactinellid sponges between the warm peri-platform of Laurentia and the peri-continental Cambrian platform of Sonora. The new material supports the impression of extremely wide distribution of Cambrian sponge genera, with local diversification at species level within regions, in contrast to much greater generic-level endemism during the Ordovician Period.
Key words: Porifera, Hexactinellida, Reticulosa, Burgess Shale-type fauna, biogeography, Cambrian, Mexico.
Matilde Sylvia Beresi [mberesi@mendoza-conicet.gob.ar], Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA) Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Av. A. Ruiz Leal s/n, Parque General San Martín, 5500, Mendoza, Argentina.
Joseph P. Botting [acutipuerilis@yahoo.co.uk], Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, China, and Department of Natural Sciences, Amgueddfa Cymru – National Museum Wales, Cathays Park, Cardiff CF10 3EP, UK.
Juan J. Palafox [palafox@correo.geologia.uson.mx], Departamento de Geología. Universidad de Sonora, Blvd. Luis Encinas y Rosales, CP 83000, Hermosillo, Sonora, México.
Blanca E. Buitrón Sánchez [blancab@unam.mx], Universidad Nacional Autónoma de México, Instituto de Geología. Departamento de Paleontología, Ciudad Universitaria, CP. 04510, México D.F., México.
Received 28 April 2017, accepted 21 August 2017, available online 22 November 2017.
Copyright © 2017 M.S. Beresi et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Sponges are one of the key components of the Cambrian Burgess Shale-type faunas, with diverse assemblages now described from the Burgess Shale (Rigby and Collins 2004), Chengjiang Biota (Rigby and Hou 1995; Wu et al. 2014), Kaili Biota (Zhao et al. 2011), the Wheeler and Marjum formations of Utah (Rigby et al. 2010), Sinsk Biota (Ivantsov et al. 2005) and the Sirius Passet Biota (Botting and Peel 2016). They are also present in other similar deposits such as the Emu Bay Shale (Paterson et al. 2016), but have not yet been described. However, due to their high diversity (including some degree of endemicity), lack of study, and differences in ages between sites, understanding of the global palaeobiogeography of Cambrian offshore sponges is still limited (Muir et al. 2013). Some indications suggest that sponge assemblages showed high temporal overturn, but with very wide generic distribution at any time (Botting and Peel 2016), nevertheless more information is necessary to assess how and when the major lineages spread through the Cambrian oceans. New faunas from otherwise unexplored regions therefore provide important information for understanding the evolutionary end ecological development of the Cambrian sponges.
The new material described here was collected from the middle Cambrian El Mogallón Formation, at Cerro El Mogallón quarry, near the town Arivechi, in Sonora State, Mexico (Fig. 1). Sponges from the Cambrian of Sonora have not previously been reported in detail, although isolated hexactine-based spicule assemblages have been recorded (Almazán-Vázquez 1989; McMenamin 2008; Cuen et al. 2013; Beresi et al. 2013).
Fig. 1. Location map showing position of the Arivechi locality in central Sonora, Mexico.
The quarry section at Cerro El Mogallón is the type section of the El Mogallón Formation. The formation is composed of thin-bedded red shales interbedded with dark gray limestone, the predominant lithology being shale with a bed thickness of less than 30 mm. The limestone beds have a thickness of around 100–200 mm, and comprise a subordinate component that yields a rich assemblage of shelly taxa. The exposed thickness at the type section is 23 m.
The formation conformably overlies the La Sata Formation and is overlain by the La Huerta Formation from the upper middle Cambrian (Fig. 2).
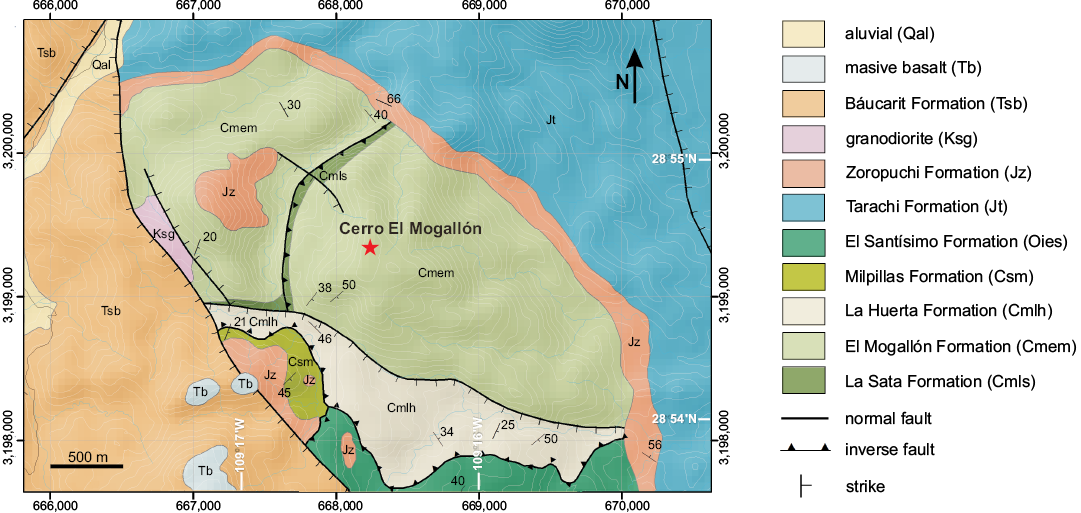
Fig. 2. Geological map showing position of the Cerro El Mogallón, and the Cambrian formations exposed in the Arivechi area, Sonora, Mexico.
The thickness of the middle Cambrian sequence in total reaches 700 m. In the El Mogallón Formation four lithostratgraphic units (in ascending order, members A–D; Fig. 3) were recognized by Almazán-Vázquez (1989), who mentioned hexactinellid spicules in members A and D. Member D of the formation is composed of thinly bedded gray mudstone, 100–200 mm thick, interbedded with red shale. Member D reaches up to 100 m thick, and is well exposed and free from obvious unconformities or hiatuses. In the basal part of this unit are found the brachiopods Lingulella sp., Linnarssonia ophirensis, Linnarssonia sp., and Prototreta sp. (Buitrón 1992; Almazán-Vázquez 1989). Above this, in the upper shale beds, the fauna becomes very abundant, including the trilobites Ptychagnostus atavus, Ptychagnostus sp., Onymagnostus hybridus?, Bathyuriscus sp., and Pagetia resseri (Almazán-Vázquez 1989).
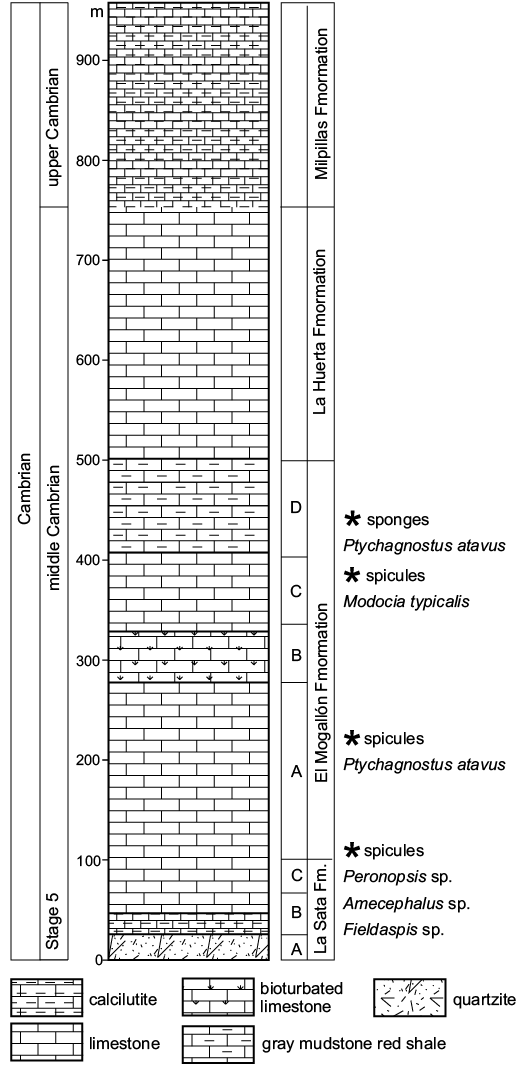
Fig. 3. Generalized stratigraphic column of the middle Cambrian El Mogallón Formation with members (A–D) with locations of sponges and spicules indicated (after Almazán-Vázquez 1989).
The age of the sponge-bearing layers has been determined, on the basis of the agnostid trilobite Ptychagnostus atavus, to correspond to the base of the P. atavus Zone, characteristic of Cambrian Series 3 of the global chronostratigraphic system (Babcock et al. 2007). The Ptychagnostus atavus Zone has been previously recognized in the El Mogallón Formation (Almazán-Vázquez 1989), and confirmed in the present study. This corresponds to the Drumian Stage of Cambrian Series 3 (Robison and Babcock 2011), and is equivalent to much of the Wheeler Formation of Utah (Rigby 1978; Rigby et al. 2010) and the lower part of the overlying Marjum Formation (Rigby 1969, 1983). The sediments of the El Mogallón Formation are, however, interpreted as representing an open-shelf environment (Almazán-Vázquez 1989) in comparison with the probably deeper-water environments represented in western Utah, particularly for the Wheeler Shale (Stewart et al. 2002).
The sponge remains described herein were collected in the lower and middle part of Member D. Sponge spicules are abundant (in contrast to the rare, very well preserved fragmentary body fossils), and are randomly distributed in patches or masses on bedding planes of red silty mudstone. Patches of irregularly distributed hexactine-based spicules and rarer triactine spicules, together with fragments of agnostid trilobites and phosphatic brachiopods, are also found on the bedding planes of calcipelite and grey-thin limestone, implying a life distribution through several sub-facies represented in the sequence.
The mixed carbonate-clastic platform, assigned to the Drumian Ptychagnostus atavus Zone (Almazán-Vázquez 1989), is broadly equivalent to the deeper-water communities from the middle Cambrian Wheeler Formation (Rigby 1978; Rigby et al. 2010) and Marjum Formation (Rigby 1969, 1983), which have yielded large numbers of reticulosan and protomonaxonid sponges. In this study we describe new species of the genera Ratcliffespongia and Valospongia, including information on their morphology, and representing the first records of these genera from Mexico and the region around it. For fossil sponge morphological terminology, refer to Finks et al. (2004).
Institutional abbreviations.—IANIGLA-PI, Colección de Paleontología del IANIGLA (Instituto Argentino de Nivologia, Glaciologia y Ciencias Ambientales) CONICET, Mendoza, Argentina; IGM, Instituto de Geología, Colección Nacional de Paleontología de la Universidad Nacional Autónoma de México, México; USDG, Departamento de Geología, Colección de Paleontología de la Universidad de Sonora, México.
Material and methods
The study area (Fig. 1) is 237.5 km east of Hermosillo, Sonora, and 10.5 km west of the town of Arivechi. The quarry is located on the hill Cerro El Mogallón (Fig. 2) which contains extensive exposures of the El Mogallón Formation, in parts affected by faulting and local deformation. The coordinates of the hill Cerro El Mogallón are: 28°54’30” N and 109°16’30” W.
Sponge specimens typically show almost no contrast when dry, and thus are very difficult to see in the field unless surfaces of split slabs are wetted. Whole-specimen photographs were taken with the specimens under water, using a Nikon D80 Digital SLR camera on a standard lens, and combination of external flash with low angle fibre-optic illumination. Detailed photographs were taken with a Canon EOS60D attached to a Leica EZ4 HD stereomicroscope.
Systematic palaeontology
Phylum Porifera Grant, 1836
“Reticulosa” sensu Botting and Muir, 2013
Remarks.—Reticulosans are thin-walled, hexactine-bearing sponges that have traditionally been regarded as hexactinellids (e.g., Finks and Rigby 2004), and the Reticulosa was erected by Reid (1958) as an extinct, third subclass of Hexactinellida. However, these sponges are now recognized as at least the stem group of Hexactinellida, Silicea and the early stem group of Demospongiae (Botting and Muir 2013, and references therein). Accordingly, the term is used informally in this context, and does not indicate a position within the crown or total group of Hexactinellida. Reticulosa is now recognised as a paraphyletic grouping, rather than a valid phylogenetic group and is used in this sense in this paper.
Family uncertain
Genus Ratcliffespongia Rigby, 1969
Type species: Ratcliffespongia perforata Rigby, 1969; lower part of the middle Cambrian Marjum Formation; Utah, USA.
Species included: The type species, Ratcliffespongia arivechensis sp. nov. and Ratcliffespongia multiforamina Wu, Yang, Jannussen, Steiner, Yang, and Zhu, 2005 from early Cambrian Huangbailing Formation of southern Anhui, South China (Wu et al. 2005) and R. freuchenensis Botting and Peel, 2016, from the early Cambrian of northern Freuchen Land, Greenland.
Remarks.—Ratcliffespongia has a complex taxonomic history, due to confusion by Rigby and Church (1990) over the nature of the type material of Rigby (1969), which they considered a decorticated outer layer of the bi-layered Hintzespongia Rigby and Gutschick, 1976. This view was further promulgated by Finks and Rigby (2004). The issue was discussed by Botting and Muir (2014) and Botting and Peel (2016), with the result that R. wheeleri Rigby and Church, 1990 was reassigned to Hintzespongia, and the genus Ratcliffespongia retained for the type species and others. Differences between the relevant genera and species are explained in the species discussion below.
Other parietal gap-bearing taxa from the Burgess Shale-type faunas include Stephenospongia Rigby, 1986, assigned to the Hintzespongiidae (Finks and Rigby 2004) and known from fragments only. It is a thin-walled sponge that can be distinguished by its variably sized (often bisected) and orthogonally arranged parietal gaps, by the large size of the spicules relative to the parietal gaps and the development of vertical and horizontal skeletal bundles between the gaps. Stephenospongia has also been reassigned to the family Hintzespongiidae (Finks and Rigby 2004; Rigby and Collins 2004), but this appears to be unwarranted given the absence of any evidence for a secondary skeletal wall, and differences from Hintzespongia in the preserved skeletal organisation.
The new material described here shows a simple, thin skeletal layer, with stauractines surrounding prominent parietal gaps, and without any dermal layer covering the openings. These features are shared with described species of Ratcliffespongia. There are small/slight differences between the species of Ratcliffespongia in the size and orientation of skeletal gaps, and differences in spiculation (Botting and Muir 2014), but the structure is fundamentally different to the double layer of Hintzespongia and allies. For these reasons, we prefer to leave the family-level assignment of Ratcliffespongia open.
Stratigraphic and geographic range.—Recognised from the early Cambrian of China and Greenland, and the middle Cambrian of the Marjum and Weeler formations of Utah, USA, and Sonora, Mexico (this paper).
Ratcliffespongia arivechensis sp. nov.
Fig. 4.
Etymology: From Arivechi, town in the state of Sonora, México, where the specimen was collected.
Holotype: UNAM 953, a very well-preserved fragment of body wall showing both skeletal details and parietal gaps outlined by ranked stauractine spicules.
Type locality: Cerro El Mogallón quarry, near Arivechi, 237.5 km East of Hermosillo city, Sonora, México.
Type horizon: Ptychagnostus atavus Biozone, Series 3, middle Cambrian.
Diagnosis.—Ratcliffespongia with irregular array of subcircular parietal gaps, and smaller gaps in largest spaces between major gaps; skeleton of stauractines (perhaps also hexactines/pentactines) with some rays aligned to produce arcuate bundles; rays straight or very slightly curved; smaller stauractine spicules sometimes with one ray deflected.
Description.—Only a single fragment available, 18 × 19 mm (Fig. 4A) without base or osculum. Thin-walled, with a single skeletal layer pierced by prominent, large, circular to elliptical parietal gaps, partly outlined by irregularly- oriented, ranked stauractines of four size orders in non-parallel arrangement. The parietal gaps are separated by skeletal regions with width slightly less than their diameter, on average, and are arranged without any regular pattern. Largest circular gaps are 2.0–3.4 mm wide, with distance between them typically around 1–2 mm (see Table 1). Additional small circular openings, normally 0.4–1.0 mm across, present between right-angled rays of the largest stauractines in areas between the large gaps.
Skeleton (Fig. 4A, B) composed possibly of simple stauractines rather than hexactines, as there is no trace of distal or gastral rays in any spicules. At least four ranks of spicules evident (see Table 1). Largest stauractines may be locally aligned, but alignment is not constant across the fragment. Smaller stauractines in non-parallel arrangement occupy areas between parietal gaps. Individual spicules with rays straight and simple, not obviously tapering visibly. First-order spicules with rays 2.4–2.9 mm long and with basal ray diameter of 0.20–0.34 mm. Some larger-order spicules have closely-spaced, aligned rays, combining to form continuous, straight or sinuous strands or bundles across the preserved fragment, extending at least 20 mm. Strands are not parallel to each other, and show no clear geometric relationship.
Table 1. Measurements (in mm) of spicules of Ratcliffespongia arivechensis sp. nov. from middle Cambrian El Mogallón Formation, Sonora, Mexico.
| |
Skeletal net: stauractines |
Parietal gaps |
|
|
spicule length |
basal diameter |
||
|
First-order spicules |
2.4–3.6 |
0.20–0.34 |
3.8–4.0 × 2.6–2.8 |
|
3.4–3.6 × 2.4–2.9 |
|||
|
3.0 × 2.0–2.6 |
|||
|
Second-order spicules |
1.6–2.0 |
0.08–0.16 |
2.7–2.8 × 2.4–2.5 |
|
2.5 × 1.8–1.9 |
|||
|
Third-order spicules |
1.2–1.4 |
0.06–0.07 |
1.3–1.7 × 1.2 |
|
1.2 × 1.1 |
|||
|
1.0 × 0.8–1.2 |
|||
|
Fourth-order spicules |
0.6–1.0 |
0.03–0.05 |
0.6–0.7 × 0.5 |
|
Zero-order spicules |
7–10 |
0.35 |
|
|
Curved spicule rays |
16–22 |
0.35 |
|
Second-order, intermediate-sized spicules have rays 1.6–1.8 mm long and are 0.08–0.12 mm in basal diameter. Third-order spicules have rays 1.2–1.3 mm long and 0.06–0.07 mm wide. Fourth-order (smallest) spicules with rays 0.60–1.0 mm long and 0.03–0.05 mm in basal diameter.
Commonly three of the rays of smaller spicules are nearly at right angles, but the fourth is often at an odd angle, apparently to accommodate other spicules and the margins of small parietal gaps. Spicules replaced by pyrite, secondarily oxidised to ferroan oxides (goethite/limonite), with the body wall tinted by iron oxides that mark the extent of soft tissues and outline parietal gaps; the entire sponge fragment is surrounded by a pale oxidation halo.
No dermal or gastral layer preserved. No evidence for basalia or marginalia is seen in the specimen.
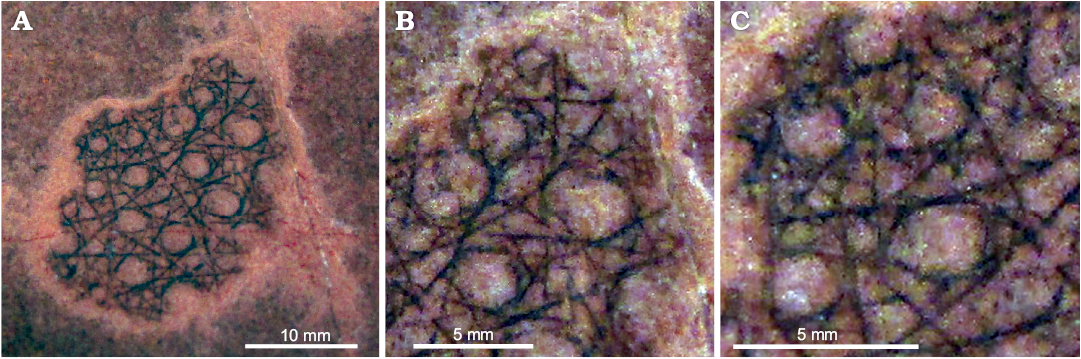
Fig. 4. Holotype UNAM 953 of the reticulosan sponge Ratcliffespongia arivechensis sp. nov. from the middle Cambrian El Mogallón Formation at Cerro El Mogallón, Sonora, Mexico. A. Skeletal layer with larger circular to elliptical parietal gaps. B. Detail showing reticulation of single skeletal layer. C. Enlargement showing that larger-order spicules are locally oriented with closely-spaced, aligned rays, combining to form straight or sinuous bundles.
Remarks.—The typical structure of the family Hintzespongiidae is confusing, as it has been described by different authors (Rigby and Church 1990: 331; Finks and Rigby 2004; Wu et al. 2005) as thin-walled, conico-cylindrical with one- or two-layered wall; if a double-layered wall, then one is a thin inner layer with prominent oval to circular parietal gaps surrounded by irregularly oriented stauractines, and the other an outer, regularly reticulate layer similar to Protospongia. and with possible rhabdodiactine prostalia. As discussed at the genus level, the second wall layer of Ratcliffespongia is an error introduced by misinterpretation of one of the species assigned to it, and the type species defines the single-layered skeletal architecture that characterises the genus.
The well-preserved fragment of R. arivechensis sp. nov. most closely resembles the type species of Ratcliffespongia, rather than Hintzespongia (formerly Ratcliffespongia) wheeleri, and shows very similar skeletal architecture. The most significant difference is the presence of arcuate strands composed of adjacent spicule rays, in a similar manner to that seen (and much more strongly developed) in Valospongia (see below). Ratcliffespongia multiforamina Wu, Yang, Jannussen, Steiner,Yang, and Zhu, 2005, from the Huangboling Formation of China, also conforms well with the type species of Ratcliffespongia, but shows few, if any, deflected rays, and includes pentactine or hexactines as well as stauracts. It is the only species of Ratcliffespongia known from virtually complete fossils, and was a broadly conical sponge; this may represent the body shape of the genus as a whole, but that cannot as yet be assessed. The early Cambrian R. freuchenensis Botting and Peel, 2016, from the Sirius Passet Biota, north Greenland, is known from fragments of a stauract-based skeleton that has smaller parietal gaps than R. arivechensis; most spicules are significantly smaller than in the current material, and more densely arranged, and the parietal gaps are separated by skeletal regions roughly as wide as their diameter. Many of these differences could be ontogenetic, but R. freuchenensis also shows strongly deflected rays in most of the larger spicules, rather than such deflections in some of the smaller spicules only. The two species are clearly closely related, but are certainly distinct. Liu et al. (2012: fig. 3) described as Protospongia? sp. a sponge from the top of the middle Cambrian Mantou Formation, Henan Province, China, which shows parietal gaps surrounded by an irregular array of hexactine-based spicules, some with strongly deflected rays. Although proximal and/or distal rays are present in some spicules, this specimen is probably assignable to Ratcliffespongia or a related lineage, and is very similar to R. freuchenenis.
Other parietal gap-bearing taxa from the Burgess Shale-type faunas include Stephenospongia Rigby, 1986, assigned to the Hintzespongiidae (Finks and Rigby 2004), which is also known from fragments only. This is a thin-walled sponge that can be distinguished by its elongate parietal gaps, the presence of numerous hexactines, arrangement of spicules to form dense tracts between the closely-spaced gaps, and by the large size of the spicules relative to the parietal gaps. The only non-Laurentian Cambrian sponge recorded as Hintzespongia (Yang et al. 2010) appears to show no evidence of a secondary spicule layer, and is likely to represent instead Ratcliffespongia or a related genus.
Stratigraphic and geographic range.—Recognised from the middle Cambrian at the Cerro El Mogallón, Arivechi, central Sonora.
Family uncertain
Genus Valospongia Rigby, 1983
Type species: Valospongia gigantis Rigby, 1983; Series 3, middle Cambrian; Utah, USA.
Species included: The type species, Valospongia sonorensis sp. nov. and Valospongia bufo Botting and Muir, 2014, from the late Tremadocian Afon Gam Biota of North Wales, UK.
Remarks.—The genus is distinctive in the possession of dermal mounds covered by a finely reticulate skeleton. There are significant differences between the skeletal structure of the type species, and that seen in Valospongia bufo Botting and Muir, 2014, and these may necessitate subdivision of the genus in future, pending more information on related forms. The current material, however, shows a close similarity to the type species, but is unquestionably a Valospongia in the strict sense.
The higher-level classification of Valospongia was discussed by Botting and Muir (2014), where assignment to any existing family was rejected. There are some structural similarities to Lantianospongia palifera Xiao, Hu, Yuan, Parseley, and Cao, 2005, and it may be possible to unite the genera into a new family in future.
Valospongia sonorensis sp. nov.
Fig. 5A.
Etymology: After the state of Sonora, México, were the specimens were collected.
Type material: Holotype: IANIGLA-PI 3094a/b, a partial specimen showing well-preserved outer skeletal layer with mounds, and spiculation of inner layer. Paratype: IANIGLA-PI 3095, fragment with part of the spicular net, from the type locality; found in the central part of the section, approximately at 40 m stratigraphically above the base of the formation.
Type locality: Cerro El Mogallón quarry, near Arivechi, 237.5 km East of Hermosillo, Sonora, México.
Type horizon: Ptychagnostus atavus Biozone, Series 3, middle Cambrian.
Material.—Type material and IANIGLA-PI 3119, fragment with part of skeleton from the type locality.
Diagnosis.—Medium to large Valospongia with prominent subcircular mounds spaced slightly less than their diameter apart, and numerous small, rounded parietal gaps in the intervening body wall; primary skeleton composed of irregular network of spicular bundles formed by parallel, adjacent rays of large stauractines (and possibly diactines); irregular meshwork of smaller stauractines forms secondary skeleton, outlining the parietal gaps; mounds covered by additional fine skeletal reticulation of indistinct spicules, dense at edge of mounds and more open at summit.
Description.—Material is preserved as oxidised pyrite replacement of spicules, and also mineralized traces of soft tissue over substantial parts of the specimens. No complete material available, but fragments imply a large, broad sponge body (Fig. 5A); holotype 61 mm maximum wide and over 86.5 mm tall; counterpart 56 mm at widest and over 84 mm tall. Oscular margin probably not preserved, although one edge is concavely curved, and may represent oscular rim; no distinct marginalia or prostalia visible.
Specimen shows all the skeletal layers compressed onto each other. Outer skeletal wall with low, subcircular mounds, mostly 3–4 mm in diameter (total range 1–6.5 mm), evenly spaced but irregularly distributed over the entire preserved surface. The mounds are perforated at their crests and appear to represent parietal gaps through the thin sponge wall, possibly representing lateral oscula by analogy with extant euplectellid (and other) hexactinellids. These mounds are now filled by fine argillaceous matrix.
Some mounds are covered with a thin, irregular mesh (Fig. 5A8) of tiny stauractines up to 0.25 mm long, irregularly organised but with some indication of a consistent, orthogonal arrangement. The fine skeletal net extends across the mounds on both counterparts. The outer layer of the skeleton between the mounds is preserved as a thin sheet of oxidised iron minerals showing the original extent of soft tissues, containing tiny spicules, which outlines numerous small parietal gaps. Spicules in this layer could correspond to autodermalia, covering the exterior of the sponge including the lateral surface of the mounds. The best-preserved regions in the holotype show an apparently regular grid with minute spicules of 0.2 mm long, oriented diagonally relative to the larger spicules, and spaces consistently around 0.1 mm across, between spicules that appear to be simple stauractines in semi-regular arrangement surrounding and overlying the mounds. Also, a dense felt of small, fine spicules (mainly irregularly oriented stauractines with ray length up to 1 mm) outlines close-packed, circular to oval spaces that form a dense, disordered array. The spicules are arranged to form an irregular polygonal meshwork, with gaps up to maximum of 0.5–1 mm across. The skeletal areas between gaps are normally 0.5 mm wide at narrowest.
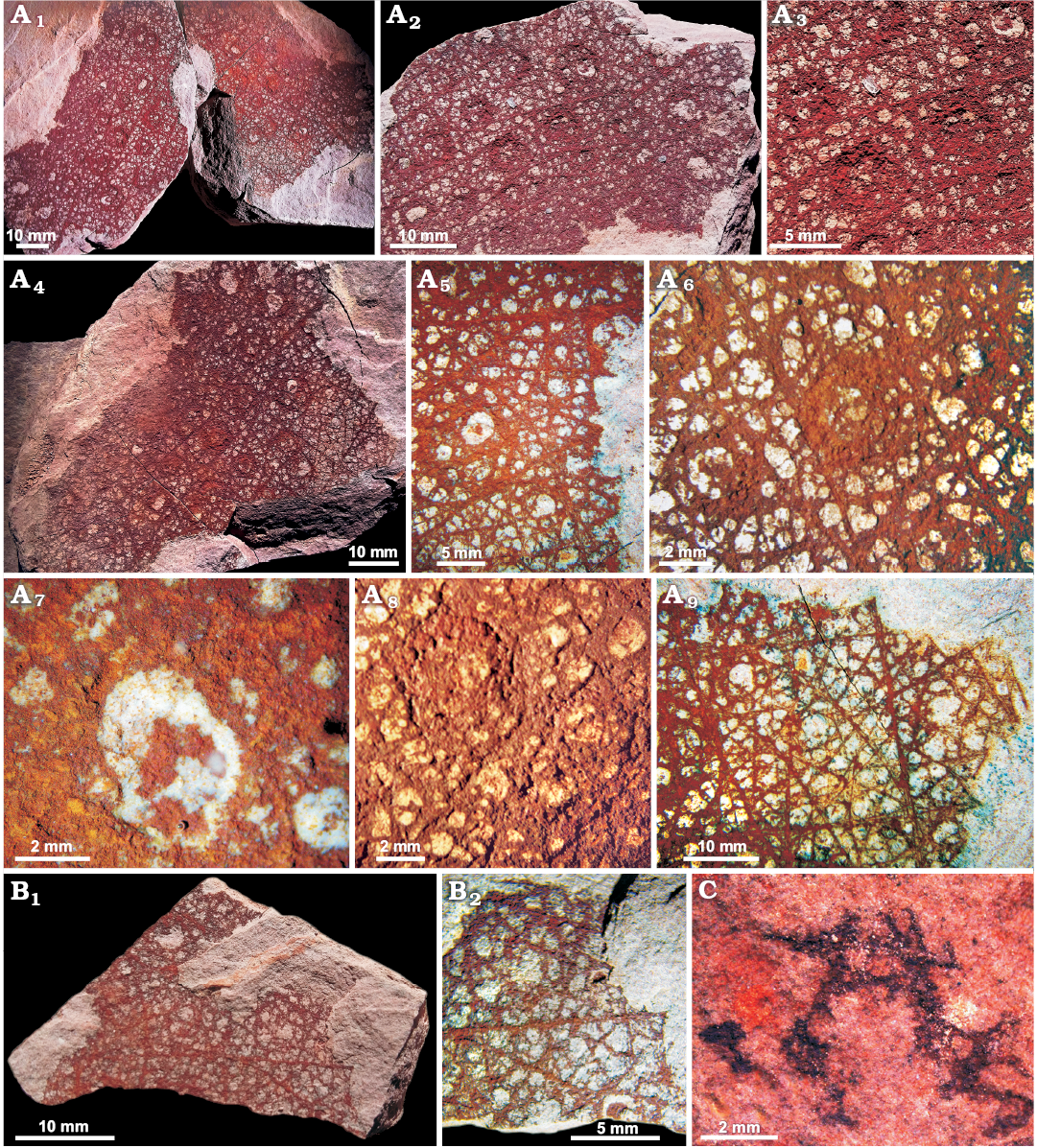
Fig. 5. Reticulosan sponge Valospongia sonorensis sp. nov. from the middle Cambrian El Mogallón Formation at Cerro El Mogallón, Sonora, Mexico. A. Holotype IANIGLA-PI 3094a/b, overall view of part and counterpart of specimen. A1, view of inner layer of the holotype; A2, detail showing base of mounds and largest spicules in semi-quadruled array; A3, view of outer layer; A4, part of the skeleton, showing semi-regular outer spicules, mounds and parietal gaps of inner layer; A5, detail of mound covered in parts by reticulate mesh, visible in places; A6, close-up of round recrystallised mound; A7, detail of a mound base with dermalia visible in places, and parietal gaps of inner layer; A8, view of part of parenchymal/choanosomal skeleton with largest spicules and smaller spicules distributed at parietal gap margins; A9, bundles composed of the largest distinct spicules, with coarse, straight and simple rays. B. Paratype IANIGLA-PI 3095; B1, overall view of the fragment; B2, detail showing largest spicules of primary reticulation and smaller spicules at parietal gap margins. C. IANIGLA-PI 3119, fragment of V. sonorensis with spiculation of the outer wall obscured by pyritisation at the margin of subcircular parietal gaps.
Primary, presumed choanosomal, skeletal layer (or possibly layers according to Rigby 1983; Finks and Rigby 2004; Botting and Muir 2014) composed of continuous strands formed by overlapping spicule rays, and forming an irregular, pseudo-orthogonal and pseudo-diagonal reticulation. Spicules may be entirely stauractines, but diactines are also probably present in the largest strands; hexactines cannot be excluded, but no evidence for distal or gastral rays observed. Primary strands or fibres typically separated by 5–8 mm. The largest distinct spicules forming the bundles have coarse, straight and simple rays up tp 30–40 mm long and with basal ray diameter of approximately 0.3–0.6 mm. Some rays appear curved and sinuous, including strand-like structures up to 37 mm long and 0.3–0.4 mm in diameter, but it is unclear whether these represent single spicule rays or bundles composed of a few straight rays that overlap in an arc. Spaces between primary strands are subdivided by similar strands composed of smaller spicules, ultimately resulting in square to irregular spaces of typically 7–13 mm.
A range of measurements for spicules of Valospongia sonorensis sp. nov. are provided in Table 2.
Table 2. Measurements (in mm) of spicules of Valospongia sonorensis sp. nov., at Cerro El Mogallón, Sonora, Mexico.
|
|
Holotype
part A |
Holotype
counterpart B |
Paratype |
|
|
Tall × wide |
|
84 × 56 |
40 × 23 |
|
|
Mounds diameter |
first-order |
5.0–6.2 |
5.8–6.0 |
|
|
second-order |
2.0–3.0 |
3.0 |
2.0–2.5 |
|
|
Parietal gaps |
first-order |
1.0–3.0 |
2.0 |
1.2–1.4 |
|
second-order |
0.5–0.7 |
0.5–1.0 |
0.5–0.75 |
|
|
Outer layer spicules |
length |
1.0–1.2 |
1.0–1.3 |
1.0 |
|
diameter |
0.5–0.6 |
|
0.25 |
|
|
Inner layer spicules |
length |
25–60 |
35 –55 |
25 |
|
diameter |
0.25 |
0.25–0.35 |
0.20 |
|
|
Quadrules |
8.5 × 7.0; 8.0 × 6.5; 8.0 × 6.0 |
7.0 × 4.0; 6.5 × 4.5; 5.0 × 4.0 |
4.5 × 3.0 |
|
|
Curved ray |
35 |
25–60 |
17 |
|
|
Dermalia |
rays length |
0.24–0.25 |
0.24 |
0.24 |
|
quadrules |
0.25 × 0.20 |
0.25 × 0.25 |
|
|
Remarks.—The structure of Valospongia is complex and disputed, with implications for systematic placement and relationships with other early reticulosans. The main components of its skeletal architecture are summarised here:
(i) Dermalia: a dense surface layer with small (1 mm) circular spaces over the entire surface, perhaps also with distinct, semi-regular lattice of tiny stauractines as autodermalia; these also cover the summits of low mounds.
(ii) Parenchymal/choanosomal skeleton: consisting predominantly of continuous inner layer or layers of semi-quadruled, semi-regular stauractines (and/or diactines and hexactines) combining to form irregularly orthogonal and diagonal elements. The long rays of stauractines form the primary skeletal support, with greatly elongated overlapping rays outlining roughly rectangular or triangular regions, subdivided by finer skeletal strands. The largest strands, probably composed at least partly of long diactines, are irregularly orientated as part of the primary skeletal structure.
Only two species of Valospongia have been previously described: the type species V. gigantis Rigby, 1983 from the middle Cambrian of Utah, and V. bufo Botting and Muir, 2014 from the late Tremadocian of North Wales. Valospongia gigantis Rigby, 1983 was assigned to Hydnodictydae Rigby, 1971 and was characterized as a thin-walled sponge sculptured with low mounds and fine-scale dermal spiculation, both over and outside the mound surfaces, together with prominent skeletal strands that were also emphasized by Finks and Rigby (2004). These features are also observed in the new material from Sonora, leaving no doubt as to the generic assignment.
The main difference between the new species and V. gigantis Rigby, 1983 is in the presence of numerous small rounded gaps between the spicule strands. As the type material of both preserves evidence of soft tissue, the difference is obvious, but in purely skeletal remains the distinction may be obscure. In that case, skeletal proportions and the wider separation of the mounds are perhaps the best discriminating characters. The mounds and the inter-mound areas of the new species appear to be covered by a reticulate meshwork of tiny stauractines, suggesting distinct autodermalia; this has not been preserved in the type species, however, so its taxonomic significance remains unclear.
The other known species, V. bufo Botting and Muir, 2014 from the late Tremadocian of Wales, is known from numerous specimens that are all far smaller than the type species or V. sonorensis sp. nov. It also has smaller and more close-packed mounds than in the Mexican species, and its inner layer is composed of dense felt of small, fine hexactines or derivatives in parallel, perhaps partly quadruled array, which is very different to the well developed skeletal bundles in the other species.
A possible Valospongia was described by Wu et al. (2014) as V. cf. gigantis, but the features of this partial sponge are very unclear. There appear to be three-dimensional mounds projecting from the body wall, but the spicules are weakly preserved. As this is the only record of the genus from China, however, it should be treated with some caution.
Stratigraphic and geographic range.—The range is restricted to the middle Cambrian El Mogallón Formation, Arivechi, central Sonora, Mexico.
Family Protospongiidae? Hinde, 1887
Remarks.—The Protospongiidae (Finks and Rigby 2004) includes many different thin-walled, reticulate sponges, some of which have since been placed in separate families on the basis of specific derived features (e.g., Asthenospongiidae Botting, 2004). The group is clearly paraphyletic and doubtless includes a range of groups that could reasonably be separated as distinct monophyletic families. We follow here the restricted familial definition of Botting and Muir (2011), which means that many of these isolated spicules cannot be assigned confidently to the Protospongiidae, and may be better regarded as indeterminate reticulosans (discussed separately below). Other slabs in the collection show dissociated stauractine and triactine spicules, which are characteristic of the Kiwetinokia Walcott, 1920.
Genus Kiwetinokia Walcott, 1920
Type species: K. utahensis Walcott, 1920; Series 3, middle Cambrian, Utah, USA.
Kiwetinokia sp.?
Fig. 6M.
Material.—USDG 001–5, several isolated triradiate spicules from the middle Cambrian at the Cerro El Mogallón, Arivechi, Sonora.
Description.—Occasional triradiate or Y-shaped spicules occur in the assemblage, associated with dissociated stauractines of diverse sizes. Triactine-like prodiaene spicules have presumed proximal rays 5.5 mm long, and two somewhat longer, presumed distal rays, with maximum ray lengths of 10.3 mm (mean = 5.5 mm). Basal ray diameter in largest spicules is 0.40–0.50 mm.
Remarks.—Walcott (1920) proposed the genus Kiwetinokia, designating the new species K. utahensis as the type, to include clusters of scattered spicules in the absence of any articulated material. Similar disarticulated remains from Little Métis, Quebec (Dawson and Hinde 1889) were reassigned to the genus from Hyalostelia. There is thus a severe risk that the spicule assemblage is itself not natural, although the name is effectively tied to the most distinctive spicules, consisting of triradiate spicules, Y-shaped spicules resembling demosponge prodiaenes, and spicules with helically twisted rays. Walcott (1920) considered the cruciform spicules associated with the more distinctive morphologies to be characteristic of species included in that genus. Finks and Rigby (2004) and other authors considered that the twisted spicular rods may be part of the basalia of the co-occurring (at Little Métis) Palaeosaccus Hinde, 1893. It is therefore possible that Kiwetinokia may be a junior synonym of Palaeosaccus, although the full range of distinctive spicules included in the genus is unlikely to be accommodated in that way. Until articulated remains are discovered, the genus remains recognisable as a spicule assemblage.
In the Arivechi material the spirally twisted triradiate spicules are not yet known. However, from the early Cambrian of the Caborca, northern Sonora, McMenamin (2008) described one spicule of this type, assigning it to Kiwetinokia.
Isolated hexactine-based spicules
Fig. 6A–H, J.
Additional dissociated spicules cannot be described taxonomically, but are typical of the Protospongiidae in general (Fig. 6A–H, J). Isolated hexactine-based spicules include spicules substantially larger than those described above, and are dominantly stauractines with four smooth coplanar rays (Fig. 6A–G), although some are possibly pentactines. Stauractine spicules show a wide range of sizes. Large stauractines have rays 7.5–11.7 mm long, with a basal ray diameter of 0.35–0.50 mm; the rays are smooth and taper to sharp point. Small stauractines have ray length of 1.4–1.7 mm and, maximum basal ray diameter of 0.06–0.10 mm (Fig. 6C).
In some spicules bearing four smooth rays there is evidence of a dermal boss that represents an aborted distal ray, and these could be interpreted as pentactines (Fig. 6A, D, F) no well-developed distal rays are seen, however. The majority of spicules exhibit 90° angles between rays, indicating a geometry of three orthogonal axes, although others have one curved or deflected ray. The preservation of rays varies within individual spicules, exhibiting a greater degree of deformation, such as foreshortening and irregularity in outline (Fig. 6); this may imply that some rays were deflected slightly from the horizontal plane.
The large size and simple morphology of the largest stauracts is typical of Protospongia hicksi Rigby, 1966 and related taxa, but many other reticulosans could also generate smaller spicules of this type. No generic or specific determination is possible with this material, although it indicates a greater diversity than is currently known from the locality.
Root tufts and basalia
Fig. 6H, J, L.
Long isolated monaxons occur in association with stauractines (Fig. 6H, L). The spicules are 6–15.5 mm long with ray diameters of approximately 0.6–0.8 mm. A single possible anchoring spicule is also preserved, with two strongly recurved rays of 0.55 mm and 0.90 mm length respectively (Fig. 6J). Unusually, it is preserved as silica. This spicule is not associated with any sponge body fragments, but as basalia are not known for either Valospongia or Ratcliffespongia, these basal structures most likely represent additional taxa.

Fig. 6. Isolated spicules and associated fossils from the middle Cambrian El Mogallón Formation at Cerro El Mogallón, Sonora, Mexico. A, B, D. Spicules exhibiting 90° angles between four coplanar rays, and a vestigial fifth perpendicular ray, preserved as a central boss (arrows), indicating a geometry of three orthogonal axes. C. Assemblage of diverse disarticulated spicules. E. Stauractine with three smooth equal long rays and the forth longer. F, G. Fragments of possibly semi-articulated skeleton composed of stauractines and monaxons, with traces of a sub-regular, perhaps quadruled array. H. Fine monaxial spicules. I. Unidentified agnostoid trilobite and a typical cruciform stauractine. J. Possible anchoring spicule with two strongly recurved rays. K. Ptychagnostus atavus (Tullberg, 1880). L. Isolated, partial, presumed root tuft with two long parallel monaxons. M. USDG 001, triactine (Kiwotinokia) and a long isolated monaxon; the arrow points to a possible ovoid sponge.
Discussion
In most of the early and middle Cambrian (Series 2 and 3) Burgess Shale-type faunas, protomonaxonids are the dominant group of sponges (Rigby and Collins 2004; Yang et al. 2005; Rigby et al. 2010; Wu et al. 2014; Botting and Peel 2016). However, deeper-water assemblages from the interval host a high proportion of reticulosans (Wu et al. 2005; Xiao et al. 2005), in a pattern that parallels that seen in Ordovician assemblages (Muir and Botting 2015). Many of these reticulosans possessed semi-regular skeletons with parietal gaps such as Ratcliffespongia or Stephenospongia, clearly bilaminar walls (Hintzespongia) or with both complex layered walls and mounds (Valospongia). Many of these reticulosan genera are distributed widely around Laurentian and South Chinese shelf seas, although there are also many taxa known only from one fauna (e.g., Xiao et al. 2005; Botting and Peel 2016).
During the earliest Cambrian, Sonora was at lower latitude, between 5° and 10° South (Fortey and Cocks 2003; Cocks and Torsvik 2011), having broken off from Laurentia around the end of the Cambrian. Additional Laurentian terrains included most of North America, the Chukotka Peninsula of Siberia, Greenland, Spitsbergen, and the NW British Isles, but from most of these there is no information regarding contemporaneous sponge faunas.
The taxa described here have been previously found in Laurentia and South China (Table 3). In Laurentia, the reticulosan genera have been found from the north of Greenland, and across the Great Basin of Utah, but the new material is in both genera most similar to the species previously described from the Wheeler and Marjum formations of Utah (e.g., Rigby 1983; Rigby et al. 2010). This additional record from the southern continental margin, in Sonora, is temporally equivalent to both formations of the Utah carbonate-dominated platform, but significantly younger than the Sirius Passet assemblage of Greenland (Botting and Peel 2016).
Table 3. Distribution of the sponges found in the Arivechi locality in comparison with other regions.
|
Species |
West Utah (USA) |
North Greenland |
Wales (UK) |
South China |
Arivechi, Sonora (México) |
Reference |
|
Ratcliffespongia |
||||||
|
R. perforata |
middle Cambrian |
|
|
|
|
|
|
R. freuchenensis |
|
Cambrian |
|
|
|
|
|
R. multiforamina |
|
|
|
Cambrian |
|
|
|
R. arivechensis |
|
|
|
|
middle Cambrian |
this paper |
|
Valospongia |
||||||
|
V. gigantis |
middle Cambrian |
|
|
|
|
|
|
V. buffo |
|
|
early Ordovician |
|
|
|
|
V. sonorensis |
|
|
|
|
middle Cambrian |
this paper |
The lithology and the occurrence of agnostoid trilobites in the El Mogallón Formation allow an inference as an open, peri-platform environment. Free-swimming agnostoid trilobites are found articulated (and therefore of local origin) in deposits of the outer belt in western Utah facing the open ocean to the west, in a similar manner to that interpreted for the El Mogallón Formation. The carbonates and siliciclastics were deposited on and around the southwestern continental margin of North America (Laurentia) during the Cambrian, which was localized near the paleoequator during Cambrian times (see Fig. 7; Page et al. 2003).
This report augments our knowledge of middle Cambrian reticulosan sponges. Some of these taxa also occur as rare specimens in the protomonaxonid-dominated Burgess Shale fauna (Rigby and Collins 2004), but are subordinate members of the assemblage. The Utah assemblages are more mixed, with the most abundant sponges being choiid protomonaxonids and the reticulosan Diagoniella. It is possible that the limited discoveries from Arivechi so far are not representative of the whole fauna, and more taxa should be anticipated in future.
The slight differences of the new sponges from the type species described from other areas are suggestive of only minor local evolution, and reinforce the view (Botting and Peel 2016) that Cambrian sponge genera were surprisingly widespread at any one time. Many of the same taxa are present in widely disparate regions, at least at low latitudes, but with many families and genera continuing into the early Ordovician at high latitudes (Botting 2016). These new finds confirm that the diversity of these faunas remains incompletely documented, even though many of the major genera are ubiquitous.
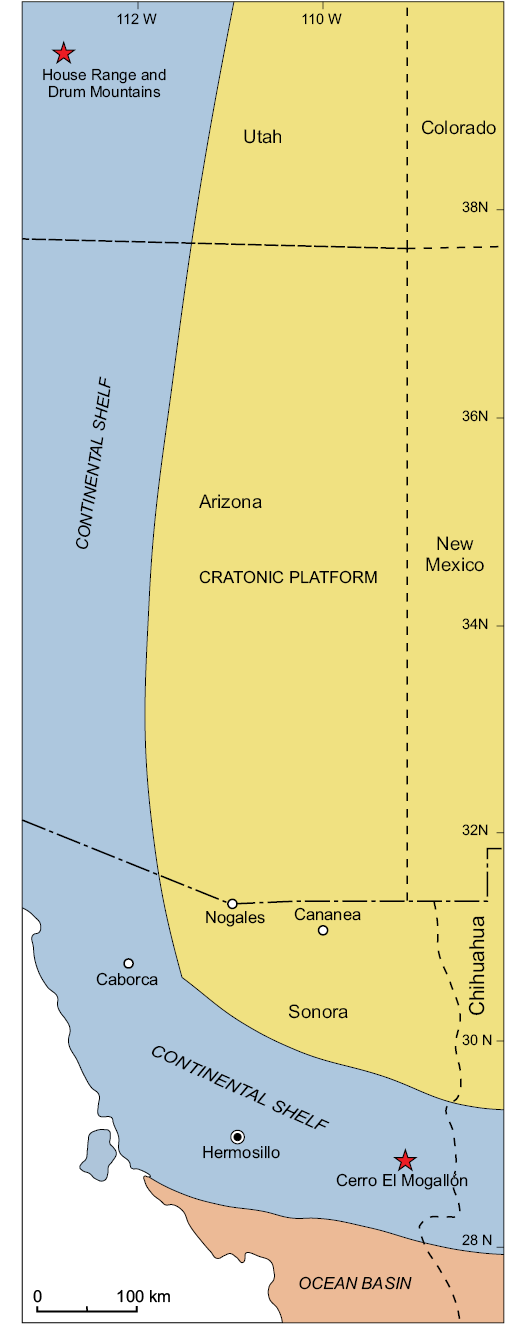
Fig. 7. Generalized map of southern part of Laurentia showing the distribution of Cambrian platform, shelf basin deposits, and the location of the Cerro El Mogallón, at Arivechi, Sonora (after Page et al. 2012).
Conclusions
A new fauna of very well preserved fragments of sponges has been recovered from the middle Cambrian of Arivechi, Mexico. Fragmentation of the remains occurred in an at least episodically turbulent environment with violent transport prior to decay, and the resulting torn pieces of sponge were preserved through pyritisation of both spicules and soft tissues. Ratcliffespongia arivechensis sp. nov. and Valospongia sonorensis sp. nov., together with disarticulated remains of Kiwetinokia, are the first records of sponges from this new locality, although by comparison with contemporaneous assemblages in Utah, Canada, and China, more species should be anticipated in future.
All the sponges described here belong to previously-recognized genera. The extension of the ranges of these genera into a new region supports previous observations of the widespread nature of many of the Cambrian sponge lineages. The subtle differences of the Sonoran species from the type species (described from Utah) indicates only a small degree of evolution within each lineage; that is, that dispersal of genera was the dominant constraint on middle Cambrian sponge palaeobiogeography, with only limited intra-regional diversification. The homogeneity of these faunas is remarkable in comparison with Ordovician sponge communities (cf. Muir and Botting 2015), supporting a very rapid and sudden dispersal of the genera following their origination in an unknown area. Intra-regional diversification, as seen in Ordovician sponge faunas, leads to numerous endemic genera and endemic, morphologically distinctive taxa at higher taxonomic levels, even on small spatial and temporal scales (Muir and Botting 2015; Botting et al. 2017). The homogeneity of middle Cambrian sponges, with only slight differences between related taxa in different regions, strongly suggests that dispersal of those genera was rapid (preventing substantial evolution during the process). The subtlety of the differences between species also suggests that there has been little diversification of the genera within each region, perhaps due to slow evolutionary rates or short timescales following their colonization.
Acknowledgements
Support for the fieldwork of BEB, JJP, and MB was provided by the Sonora geologists Francisco Cuen, Dulce Reyes, Amilcar Casarrubias, and Glenda Danzós (all Department of Geology, Sonora University, México) and the logistics by Rosendo Dueñas (Sonora, Mexico) are gratefully acknowledged. We thank Rafael Bottero (IANIGLA) for the making of the figures and Marcelo Bourget (IANIGLA) for the preparation of the samples, and Andrzej Pisera (Institute of Paleobiology PAS, Warsaw, Poland) and two anonymous referees for constructive comments. Authors thank the financial support from the Projects: M13-U01 ECOS-Francia, ANUIES (to JJP), and CONACY-Mexico 165826 (to BEB and to MSB) and 235970 (to JJP).
References
Almazán-Vázquez, E. 1989. El Cámbrico-Ordovícico de Arivechi, en la región centro oriental del estado de Sonora. Universidad Nacional Autónoma de México, Instituto de Geología, Revista 8 (1): 58–66.
Babcock, L.E., Robison, R.A., Rees, M.N., Peng, S., and Saltzman, M.R. 2007. The Global boundary Stratotype Section and Point (GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA. Episodes 30 (2): 85–95.
Beresi, M.S, Cuén, R., Montijo, G.A., Buitrón, B.E., Minjarez, J.L., De la O, M., Palafox, J. 2013. Chancelloriida Walcott, 1920 y Reticulosa Reid, 1958 (Protospóngidos) del Cámbrico de San José de gracia, Sonora. In: VIII y XIII Congreso Mexicano de Paleontología, Libro de Resúmenes, 38. Sociedad Mexicana de Paleontología A.C., Museo Dugès, Universidad de Guanajuato, México.
Botting, J.P. 2004. An exceptional Caradoc sponge fauna from the Llanfawr Quarries, central Wales, and phylogenetic implications. Journal of Systematic Palaeontology 2: 31–63. Crossref
Botting, J.P. 2016. Diversity and ecology of sponges in the Early Ordovician Fezouata Biota, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 75–86. Crossref
Botting, J.P. and Muir, L.A. 2011. A new Middle Ordovician (late Dapingian) hexactinellid sponge from Cumbria, UK. Geological Journal 46: 501–506. Crossref
Botting, J.P. and Muir, L.A. 2013. Spicule structure and affinities of the Late Ordovician hexactinellid-like sponge Cyathophycus loydelli (Llanfawr Mudstones Lagerstätte, Wales). Lethaia 46: 454–469.
Botting, J.P. and Muir, L.A. 2014. First post-Cambrian records of the reticulosan sponges Valospongia and Hintzespongia from the late Tremadocian of North Wales. Acta Palaeontologica Polonica 59: 241– 252.
Botting, J.P. and Peel, J.S. 2016. Early Cambrian Sponges of The Sirius Passet Biota, North Greenland. Papers in Palaeontology 2: 1–25. Crossref
Botting, J.P., Muir, L.A., Zhang, Y., Ma, X., Ma, J., Wang, L., Zhang, J., Song, Y., and Fang, X. 2017. Flourishing sponge-based ecosystems after the End-Ordovician mass extinction. Current Biology 27: 556–562. Crossref
Buitrón, B.E. 1992. Las rocas sedimentarias del Paleozoico inferior de México y su contenido biótico. In: J.G. Gutiérrez Marco, J. Saavedra, and I. Rábano (eds.), Paleozoico inferior de Ibero-América, 193–201. Universidad de Extremadura, Cáceres.
Cocks, L.R. and Torsvik, T.H. 2011. The Palaeozoic geography of Laurentia and western Laurussia: A stable craton with mobile margins. Earth-Science Reviews 106: 1–51. Crossref
Cuen, R.F., Beresi, M.S., Montijo, A., Buitrón, B.E., Minjárez, I., De la O, M., and Palafox, J.J. 2013. Chancelloriia Walcott, 1920 y Reticulosa Reid, 1958 del Cámbrico medio de San José de Gracia, Sonora, México. Boletín de la Sociedad Geológica Mexicana 65: 581–590. Crossref
Dawson, J.W. and Hinde, G.J. 1889. On new species of fossil sponges from the Siluro-Cambrian at Little Métis on the Lower St. Lawrence; including notes on the specimen by Dr. G.J. Hinde, F.G.S. Transactions of the Royal Society of Canada (section IV), Geological and Biological Sciences 7: 31–55.
Finks, R.M. and Rigby, J.K. 2004. Paleozoic hexactinellid sponges. In: R.L. Kaessler (ed.), Treatise on Invertebrate Paleontology, Part E, Porifera (revised), vol. 3, 319–448. Geological Society of America, Boulder and University of Kansas, Lawrence.
Finks, R.M., Reid, R.E.H., and Rigby, J.K. 2004. Introduction to the Porifera. In: R.L. Kaessler (ed.), Treatise on Invertebrate Paleontology, Part E, Porifera (revised), Vol. 2. 345 pp. Geological Society of America, Boulder and University of Kansas, Lawrence.
Fortey, R.A. and Cocks, L.R.M. 2003. Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions. Earth-Science Reviews 61: 245–307. Crossref
Hinde, G.J. 1887. A monograph of the British fossil sponges, part 1. Palaeontographical Society Monograph, London 1: 1–92.
Hinde, G.J. 1893. On Palaeosaccus dawsoni Hinde, a new genus and species of hexactinellid sponge from the Quebec Group (Ordovician) at Little Métis, Quebec, Canada. Geological Magazine (new series, decade III) 10: 56–59.
Ivantsov, A.Y. [Ivancov, A.Û.], Zhuravlev, A.Y. [Žuravlev, A.Û.], Krassilov, V.A. [Krasilov, V.A.], Leguta, A.V., Melnikova, L.M. [Mel’nikova, L.M.], Urbanek, A., Ushatinskaya, G.T. [Ušatinskaâ, G.T.], and Malakhovskaya, Y.E. [Malahovskaâ, Â.E.] 2005. Unique Sinsk localities of Early Cambrian organisms (Siberian Platform) [in Russian]. Trudy Paleontologičeskogo instituta 284: 1–143.
Liu Q., Huang D., and Gong Y. 2012. Sponge fossil from the Cambrian Mantou Formation of Hebi, Henan Central China. Earth Science Journal of China University of Geoscience 37 (Supplement 2): 129–135.
McMenamin, A.S. 2008. Early Cambrian sponge spicules from the Cerro Clemente and Cerro Rajón, Sonora, Mexico. Geologica Acta 6: 363–367.
Muir, L.A. and Botting, J.P. 2015. An outline of the distribution and diversity of Porifera in the Ordovician Builth Inlier (Wales, UK). Palaeoworld 24: 176–190. Crossref
Muir, L.A., Botting, J.P., Carrera, M.G., and Beresi, M., 2013. Cambrian, Ordovician and Silurian non-stromatoporoid Porifera. Geological Society, London, Memoirs 38: 81–95.
Page W.R., Harris, A.G., Poole F.G., and Repetski J.E. 2003. Reinterpretation of the stratigraphy and structure of the Rancho Las Norias area, central Sonora, Mexico. Journal of South American Earth Sciences 16: 523–540. Crossref
Paterson, J.R., García-Bellido, D.C., Jago, J.B., Gehling, J.G., Lee, M.S., and Edgecombe, G.D. 2016. The Emu Bay Shale Konservat-Lagerstätte: a view of Cambrian life from East Gondwana. Journal of the Geological Society 173: 1–11. Crossref
Reid, R.E.H. 1958. A monograph of the Upper Cretaceous Hexactinellida of Great Britain and Northern Ireland, Part 1. Palaeontographical Society of London, Monographs 111: 1–45.
Rigby, J.K. 1966. Protospongia hicksi Hinde from the Middle Cambrian of western Utah. Journal of Paleontology 40: 549–554.
Rigby, J.K. 1969. A new Middle Cambrian hexactinellid sponge from western Utah. Journal of Paleontology 43: 125–128.
Rigby, J.K. 1971. Sponges of the Ordovician Cat Head Member, Lake Winnipeg, Manitoba. In: D.C. McGregor, F.H. Cramer, R.H. Flower, and J.K. Rigby. Contributions to Canadian Paleontology, Fossils of the Ordovician Red River Formation (Cat Head Member), Manitoba. Geological Survey of Canada Bulletin 202: 35–78.
Rigby, J.K. 1978. Porifera of the Middle Cambrian Wheeler Shale, from the Wheeler Amphipheater, House Range, in western Utah. Journal of Paleontology 52: 1325–1345.
Rigby, J.K. 1983. Sponges of the Middle Cambrian Marjum Limestone from the House Range and Drum Mountains of western Millard County, Utah. Journal of Paleontology 57: 240–270.
Rigby, J.K. 1986. Sponges of the Burgess Shale (Middle Cambrian) British Columbia. Palaeontographica Canadiana 2: 1–105.
Rigby, J.K. and Church, S.B. 1990. A new Middle Cambrian hexactinellid, Ratcliffespongia wheeleri, from western Utah and skeletal structure of Ratcliffespongia. Journal of Paleontology 64: 331–334. Crossref
Rigby, J.K. and Collins, D. 2004. Sponges of the Middle Cambrian Burgess and Stephen Shale Formations, British Columbia. Royal Ontario Museum Contributions in Science 1: 1–164.
Rigby, J.K. and Gutschick, R.C. 1976. Two new Lower Paleozoic hexactinellid sponges from Utah and Oklahoma. Journal of Paleontology 50: 79–85.
Rigby, J.K. and Hou, X.-G. 1995. Lower Cambrian demosponges and hexactinellid sponges from Yunnan, China. Journal of Paleontology 69: 1009–1019. Crossref
Rigby, J.K., Church, S.B., and Anderson, N.K. 2010. Middle Cambrian sponges from the Drum Mountains and House Range in western Utah. Journal of Paleontology 84: 66–78. Crossref
Robison, R.A. and Babcock, L.E. 2011. Systematics, paleobiology, and taphonomy of some exceptionally preserved trilobites from Cambrian Lagerstätten of Utah. Paleontological Contributions 5: 1–47.
Stewart, J.H., Amaya-Martínez, R., and Palmer, A.R. 2002. Neoproterozoic and Cambrian strata of Sonora, Mexico: Rodinian supercontinent to Laurentian Cordillera margin. In: A. Barth (ed.), Contributions to crustal evolution of the Southwestern United States. Geological Society of America, Special Paper 365: 5–47. Crossref
Walcott, C.D. 1920. Cambrian geology and paleontology IV, no. 6, Middle Cambrian Spongiae. Smithsonian Miscellaneous Collections 67: 261–364.
Wu, W., Yang, A., Janussen, D., Steiner, M., Yang, X., and Zhu, M. 2005. Hexactinellid sponges from the Early Cambrian Black Shale of Anhui, China. Journal of Paleontology 79: 1043–1051. Crossref
Wu, W., Zhu, M., and Steiner, M. 2014. Composition and tiering of the Cambrian sponge communities. Palaeogeography, Palaeoclimatology, Palaeoecology 398: 86–96. Crossref
Xiao, S., Hu, J., Yuan, X., Parsley, R.L., and Cao, R. 2005. Articulated sponges from the Lower Cambrian Hetang Formation in southern Anhui, South China: their age and implications for the early evolution of sponges. Palaeogeography, Palaeoclimatology, Palaeoecology 220: 89–117. Crossref
Yang, X., Zhao, Y., Zhu, M., Cui, T., and Yang, K. 2010. Sponges from the Early Cambrian Niutitang Formation at Danzhai, Guizhou and their environmental background. Acta Palaeontologica Sinica 49: 348–359.
Zhao, Y., Zhu, M., Babcock, L.E., and Peng, J. (eds.) 2011. The Kaili Biota: Marine Organisms from 508 Million Years Ago [in Chinese]. 251 pp. Guizhou Publishing Group, Guiyang.
Acta Palaeontol. Pol. 62 (4):
691–703, 2017
https://doi.org/10.4202/app.00378.2017