
Late Jurassic–Early Cretaceous oysters from Siberia: A systematic review
IGOR N. KOSENKO
Kosenko, I.N. 2017. Late Jurassic–Early Cretaceous oysters from Siberia: A systematic review. Acta Palaeontologica Polonica 62 (4): 759–778.
The present study reviews the taxonomy of Late Jurassic–Early Cretaceous oysters from the Northern and the Subpolar Urals (Western Siberia) and northern East Siberia. Previous studies have documented 10 species from the genus Liostrea (L. delta, L. cucurbita, L. praeanabarensis, L. anabarensis, L. plastica, L. gibberosa, L. planoconvexa, L. siberica, L. uralensis, L. lyapinensis), and 3 species from the genus Gryphaea (G. borealis and 2 species in open nomenclature). Liostrea gibberosa, L. planoconvexa, L. uralensis, and L. cucurbita are transferred in this study to the genus Pernostrea. Furthermore, two new species of Pernostrea are described: P. mesezhnikovi sp. nov. and P.? robusta sp. nov. Liostrea siberica is transferred to the genus Praeexogyra. Liostrea praeanabarensis and L. anabarensis are attributed to the subgenus Boreiodeltoideum (genus Deltoideum) as well as L. delta sensu Zakharov (1966) which is described here as new species Deltoideum (Boreiodeltoideum) borealis sp. nov. The similar shell morphology of the genera Deltoideum and Pernostrea provides a basis to establish the new tribe Pernostreini trib. nov. in the subfamily Gryphaeinae. Three species are recorded for the first time from Siberia: Nanogyra? cf. thurmanni, “Ostrea” cf. moreana and Gryphaea (Gryphaea) curva.
Key words: Bivalvia, Ostreoidea, Gryphaeidae, Jurassic, Cretaceous, Siberia, Russia.
Igor N. Kosenko [kosenkoin@ipgg.sbras.ru], Trofimuk Institute of Petroleum Geology and Geophysics SB RAS, prospect Koptyuga, 3, Novosibirsk, 630090 Russia (correspondence address); and Novosibirsk State University, Pirogova 2, Novosibirsk, 630090 Russia.
Received 19 May 2017, accepted 7 September 2017, available online 15 November 2017.
Copyright © 2017 I.N. Kosenko. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
It has been almost 40 years since Zakharov (1963, 1966, Zakharov in Saks et al. 1972; Zakharov in Zakharov and Mesezhnikov 1974; Zakharov 1978) have studied the rich oyster fauna of the Siberian Upper Jurassic and Lower Cretaceous. Since that time oyster classification has undergone several changes based on paleontological (Malchus 1990; Koppka 2015), anatomical (e.g., Harry 1985) and increasingly also on molecular phylogenetic analyses (e.g., Ó Foighil and Taylor 2000; Tëmkin 2010; Salvi et al. 2014; Raith et al. 2016). The objectives of the paper are to: (i) review of taxonomy of Late Jurassic–Early Cretaceous Siberian oysters based on recent data; (ii) describe new species; (iii) describe new tribe and clarify diagnoses of the genera Pernostrea and Deltoideum; (iv) summarize stratigraphic distribution of oysters in Siberian Kimmeridgian–Valanginian sections.
In this changing context, the present study sets out for a taxonomic reappraisal of the Siberian oysters using mainly the original collections of Victor A. Zakharov and other material available (Kosenko 2014, 2016a; Kosenko and Seltser 2016). All these data are included into a more complete picture of the evolutionary history of the Ostreoidea.
Institutional abbreviations.—TsSGM, Central Siberian Geological Museum, Novosibirsk, Russia; NHM, Lille Natural History Museum, France.
Other abbreviations.—D1, distance between posterior adductor muscle scar and anterior valve margin; D2, distance between posterior adductor muscle scar and posterior valve margin; D3, distance between posterior adductor muscle scar and ventral margin; D4, distance between posterior adductor muscle scar and dorsal margin; H, shell height; HCF, herringbone cross-foliated structure; Hla, ligament area height; Hpam, posterior adductor muscle scar height; L, shell length; LA, ligament area; LV, left valve; Lab, anterior bourrelet length; Lla, ligament area length; Lpam, posterior adductor muscle scar length; Lpb, posterior bourrelet length; Lr, resilifer length; PAM, posterior adductor muscle scar; RF, regularly foliated structure; RV, right valve.
Nomenclatural acts.—The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be re-solved and the associated information viewed through any standard web browser by appending the LSID to the pre-fix “http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:26FBF7FF-F554-48DC-B02D-069507B4202B. The electronic edition of this work was published in a journal with an eISSN 1732-2421, and has been archived and is available from the following digital repository: http://www.app.pan.pl/article/item/app003872017.html
Material and methods
The material was collected during the 1960s by Victor A. Zakharov in the Upper Jurassic–Lower Cretaceous successions of the eastern slopes of the Northern and Subpolar Ural mountains (along Tolya, Maurynya, and Yatriya rivers), northwestern Western Siberia, as well as in northern Eastern Siberia (Bolshaya Romanihka, Boyarka, and Dyabaka-Tari rivers) (Fig. 1). Additional material from the Jurassic/Cretaсeous boundary strata of the Maurynya River was provided by Olga S. Urman (Trofimuk Institute of Petroleum Geology and Geophysics, Novosibirsk, Russia) along with their stratigraphical position in the studied sections (Figs. 2–4). The material (including shells collected by Olga S. Urman) consists of 465 shells (exclusive of the shells already stocked in TsSGM) of Siberian oysters.
Further material used for comparison comes from the Upper Jurassic of England (collected by Victor A. Zakharov) and northern France (collected by Clémentine Colpaert, Boris L. Nikitenko, and INK; including the observation of Late Jurassic oysters in the collections of the Lille Natural History Museum). The oysters from the Upper Cretaceous and Paleogene (about 2000 shells) were collected by the present author and Vladimir V. Arkadyev in 2014 at the Maastrichtian sections “Baklinskaya kuesta” and “Korabelnaya kuesta”, the Paleogene sections “Balta-Chokrak” (Thanetian stage) and Thanetian–Lutetian section in a limestone quarry near the village Skalistoe (Mountain Crimea). Recent specimens of Crassostrea gigas (more than 100 shells) were collected by the author in 2014 at the coast of the Japan Sea (Russky Island, near the Far Eastern Federal University).
Along with a classic morphofunctional analysis, the oyster shell microstructure has been studied using light and scanning electron microscopy. Methods of preparing thin sections, as well as microstructure descriptions of oysters of Cheltsova (1969) were followed except for not using acid etching during the processing of the material. In order to study these thin sections using SEM, oyster shells were broken down into small pieces, and then were coated with sputtered gold. Main types of oyster shells microstructure were usually recognized. In Recent oysters, individual prisms forming the prismatic layer have been identified (Kosenko 2014: fig. 6). Metrical parameters of oyster shells have been measured with a standard caliper.
The morphological and microstructure terminology is adopted from Stenzel (1971), Malchus (1990), Carter (1990), and Carter et al. (2012). The main morphological characters of oyster shells from different subfamilies are shown in Fig. 3 and the measured parameters in Fig. 4.

Fig. 1. Geographical position of the studied oysters locations: 1, Lopsiya River; 2, Tolya River; 3, Maurynya River; 4, Yatriya River; 5, Boyarka River; 6, Bol’shaya Romanikha River; 7, Dyabaka-Tari River (after Zakharov 1966; Zakharov and Mesezhnikov 1974).

Fig. 2. Bio- and lithostratigraphy of Upper Jurassic–Lower Cretaceous sections from the Northern and the Subpolar Urals with the distribution of oysters in the Lopsiya River section (A), the Yatriya River section (B), and the Maurynya River section (C).
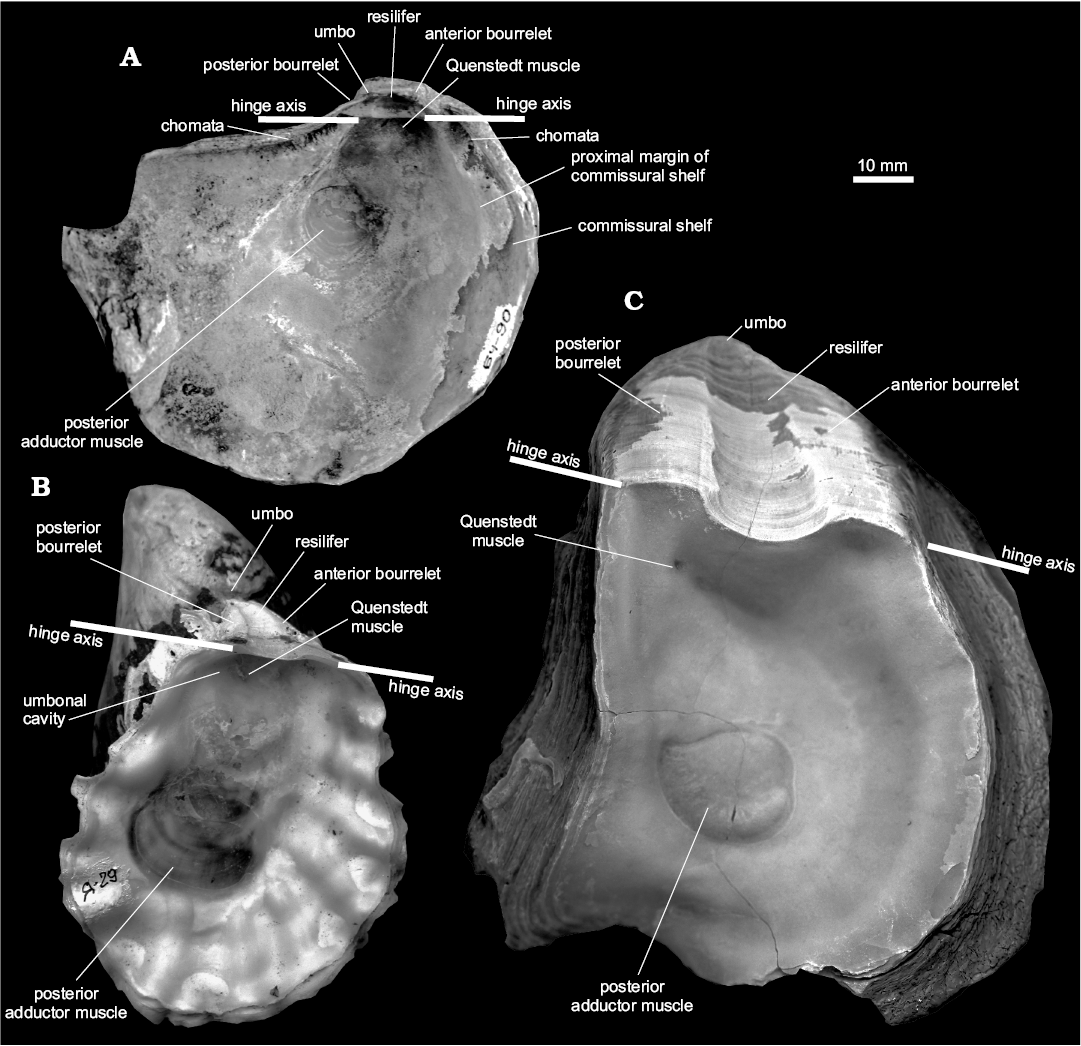
Fig. 3. Main morphological characters of oysters. A. Left valve of Phygraea (Gryphaeidae, Pycnodonteinae), TsSGM 2068/88, inner view. B. Left valve of Crassostrea (Flemingostreidae, Crassostreinae), TsSGM 2068/13, inner view. C. Left valve of Pernostrea (Gryphaeidae, Gryphaeinae), TsSGM 2068/2, inner view.
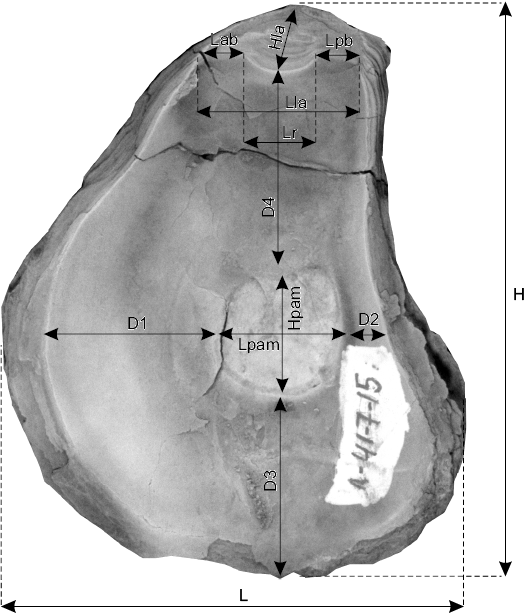
Fig. 4. Measurements of morphometric parameters of oyster shells. D1, distance between posterior adductor muscle scar and anterior valve margin; D2, distance between posterior adductor muscle scar and posterior valve margin; D3, distance between posterior adductor muscle scar and ventral margin; D4, distance between posterior adductor muscle scar and dorsal margin; H, shell height; HCF, herringbone cross-foliated structure; Hla, ligament area height; Hpam, posterior adductor muscle scar height; L, shell length; LA, ligament area; LV, left valve; Lab, anterior bourrelet length; Lla, ligament area length; Lpam, posterior adductor muscle scar length; Lpb, posterior bourrelet length; Lr, resilifer length; PAM, posterior adductor muscle scar; RF, regularly foliated structure; RV, right valve.
Geological setting
A detailed bed-by-bed description of sections at the eastern slopes of the Northern Urals and the Subpolar Urals (Lopsiya, Tolya, and Yatriya rivers) is given by Zakharov and Mesezhnikov (1974), while the section of the Jurassic–Cretaceous boundary strata located along the Maurynya River was studied by Saks et al. (1972), Alifirov et al. (2008), and Dzyuba et al. (2013). The Jurassic–Cretaceous boundary in Siberian sections is located within Craspedites taimyrensis Zone of the Upper Volgian (the necessity of usage of the Volgian stage in Siberia has been shown by Zakharov 2003 and Shurygin and Dzyuba 2015) based on complex of biostratigraphic, palaeomagnetic, and isotope data (Dzyuba et al. 2013; Shurygin and Dzyuba 2015). The stratigraphy of the Upper Jurassic and the Lower Cretaceous sections of the northern part of Eastern Siberia is summarized in Zakharov (1970).
In the Lopsiya River section oysters have been found in the Lower Kimmeridgian clayey silt and clay and the Upper Kimmeridgian–lowermost Volgian silty clay (Fig. 2). The Lower Kimmeridgian associations are allochthonous and the Upper Kimmeridgian associations are autochthonous. The base of the Volgian stage is marked by a large shell concentration of the oyster Pernostrea mesezhnikovi sp. nov. forming a shell bed. Palaeotemperatures calculated from the δ18 O values of belemnite rostra vary from +17°С to +24°С for the Kimmeridgian–Volgian interval of the Lopsiya River (Zakharov et al. 2005). The highest paleotemperature (+24°С) has been calculated for the lower part of Eurasenia pseudouralensis Zone (near the base of the Lower Kimeridgian). The other palaeotemperature peak (around +22°С) has been calculated for the Eosphinctoceras magnum Zone (near the Kimmeridgian/Volgian boundary) near to the abovementioned oyster shell bed. According to Zakharov et al. (1983), the Northern Urals during the Kimmeridgian was covered by a shallow-marine sea called the Lyapin Bay. A permanent presence of cephalopods such as ammonites and belemnites in the Lopsiya River section suggests a normal salinity of the Lyapin Bay (Zakharov et al. 2005).
During the Volgian age, the Subpolar and the Northern Urals were covered by a shallow sea with normal salinity (Zakharov et al. 1983). The Volgian deposits, exposed in the Yatriya River section, consist of silty calcareous sediments with glauconite, and yield numerous ammonites and bivalves. According to Zakharov (1981), the average annual palaeotemperatures based on isotope thermometry decreased in the Middle Volgian to +15.5°С and +12.7°С in the Late Volgian. In the Yatriya River section, oysters have been found in the Lower Volgian silt, the Middle Volgian calcareous silt and the Upper Volgian silt (Fig. 2). The most diversified oyster associations occur in the Middle Volgian silt.
In the Maurynya River section, oysters have been found in sand and calcareous sand of the Upper Volgian–Ryazanian (Fig. 2) and usually occur in shell accumulations (biostromes). Paleotemperatures based on δ18O values of oyster shells vary from 15°С to 21°C (Kosenko et al. 2013). Oysters found in the Tolya River section occur in clay of the Lower Volgian and silt of the Middle Volgian age (Zakharov and Mesezhnikov 1974).
In the sections located in northern Eastern Siberia, oysters have been found in fine-grained sand of the Kimmeridgian, in the Lower Volgian silt and silty fine-grained sand, and in fine-grained sand of the Ryazanian–Lower Hauterivian (Zakharov 1966). During the Kimmeridgian–Hauterivian, the north of Eastern Siberia was covered by a sea with normal salinity (Zakharov 1966; Zakharov et al. 1983).
According to the palaeogeographical map of Siberia during the Bazhenov time (the Volgian and beginning of the Ryazanian) (Devyatov et al. 2011), the studied oysters inhabited the upper sublittoral zone (0–20 m depth) which is in agreement with the favorite habitat of most of the Recent oysters (Scarlato 1960; Stenzel 1971).
Systematic palaeontology
Class Bivalvia Linnaeus, 1758
Subclass Autobranchia Grobben, 1894
Cohort Ostreomorphi, Férussac, 1822
Subcohort Ostreoini Férussac, 1822
Megaorder Ostreata Férussac, 1822
Superorder Ostreiformii Férussac, 1822 (= Eupteriomorphia Boss, 1982)
Order Ostreida Férussac, 1822
Suborder Ostreidina Férussac, 1822
Superfamily Ostreoidea Rafinesque, 1815
Family Gryphaeidae Vialov, 1936
Subfamily Gryphaeinae Vialov, 1936
Tribe Pernostreini nov.
Type genus: Pernostrea Munier-Chalmas, 1864.
Genera included: Pernostrea Munier-Chalmas, 1864, Deltoideum Rollier, 1917.
Diagnosis.—PAM orbicular (gryphaeoid type), centrally located or shifted to dorsal or posterior side. Ligament area ostreoid (occasionally multivincular), disposed with small angle to commissural plane. Umbo small, not protruding above the commissural plane. Inner surface of valves smooth. No chomata. Outer surface covered with growth squamae. Some species have radial striae.
Remarks.—The name of this new tribe was first mentioned in a conference abstract (Kosenko 2016b). However, according to ICZN Article 9.10 a conference abstract does not constitute a published work.
Stratigraphic and geographic range.—Middle Jurassic (Aalenian)–Lower Cretaceous (Hauterivian). Bajocian–Bathonian of Greenland. Bathonian–Tithonian of Europe (Alps, Poland, Romania, France, Germany, England, Switzerland, European part of Russia). Upper Jurassic of Saudi Arabia. Berriassian of Asia (Central Asia, Siberia). Kimmeridgian–Valanginian of Western Siberia, Kimmeridgian–Hauterivian of Eastern Siberia. The last representatives are known from the Hauterivian of northeastern Siberia.
Genus Deltoideum Rollier, 1917
Type species: Ostrea sowerbyana Bronn, 1836 (= Ostrea delta Smith, 1817), Kimmeridgian of England and France.
Diagnosis (modified from Stenzel 1971: N1102).—Shells medium to large, equivalve, rare slightly inequivalve. Outlines deltoid, crescentic or spatulate. Some species have well developed marginal growth foliations. Ligament area ostreoid, high, narrow, triangular to trapezoidal. Resilifer shallow, well marked, usually triangular. Anterior bourrelet usually longer than posterior one. Attachment area small to very large (almost all of LV). Angle between resilifer and commissural plane usually around 10°. PAM medium to small, circular, clearly marked with a posterocentral to posteroventral position. No umbonal cavity. No chomata. Surface of valves covered with commarginal growth squamae. RV of some species covered by radial striae. The crescentic depression for the gills and the brachytellum is well developed. Microstructure consists of regularly foliated structure (Fig. 5A).
Stratigraphic and geographic range (modified from Stenzel 1971: N1103).—Middle Jurassic (Bathonian)–Upper Jurassic (Tithonian) of Europe (England, France, Germany, Poland, European part of Russia) and Saudi Arabia; Berriassian of Mangyshlak (Kazakhstan); Upper Jurassic (Kimmeridgian)–Lower Cretaceous (Hauterivian) of Siberia.
Subgenus Boreiodeltoideum Kosenko, 2016a
Type species: Ostrea (Liostrea) anabarensis Bodylevsky, 1949; Upper Valanginian of the Anabar River (north of Eastern Siberia).
Diagnosis.—Shells medium to large, inequivalve. LV slightly convex, RV flat. Outlines deltoid to crescent. Umbo opisthogyrate. Ligament area ostreoid, small to medium, triangular to trapezoidal. Angle between LA and commissural plane very small (less than 10°). Resilifer triangular, wide, deeper on LV. Height of posterior bourrelet usually much smaller than height of anterior bourrelet. Attachment area small to large. Umbonal cavity absent, some species have a poorly developed depression located under the posterior bourrelet. PAM medium-sized, circular to length-oval in a posterocentral or posterodorsal position. No chomata. Outer surface of valves covered by concentric growth striae.
Stratigraphic and geographic range.—Upper Jurassic (Kimmeridgian)–Lower Cretaceous (Hauterivian) of Siberia.
Deltoideum (Boreiodeltoideum) borealis sp. nov.
Fig. 5A, E.
1963 Liostrea ex gr. delta (Smith, 1817); Zakharov 1963: 44, fig. 1a–c.
1966 Liostrea delta (Smith, 1817); Zakharov 1966: 101, pl. 37: 2a,b, pl. 38: 1.
ZooBank LSID: urn:lsid:zoobank.org:pub:26FBF7FF-F554-48DC-B02D-069507B4202B
Etymology: From Greek boreas, north; in reference to type area.
Holotype: TsSGM 150/3887 (Fig. 5E) left and right valves from the same shell with exposed ligament area.
Type locality: Outcrop along the Levaya Boyarka River (N70°33̍, E97°20̍), north of Eastern Siberia, Russia.
Type horizon: Kimmeridgian sand of the Sigovskaya Formation (the Sigovskaya Formation consists mainly of sandstone; for the detail information see Shurygin et al. 2000), the Levaya Boyarka River, north of Eastern Siberia.
Material.—Holotype and five left and right valves from the Lower Kimmerigian deposits exposed at the Lopsiya River (TsSGM, collection 2068, holotype TsSGM 150/3887, paratypes TsSGM 2068/31, 2068/83, 2068/84, 2068/85, 2068/86).
Diagnosis.—Shells medium to large, inequivalve, LV convex, RV flat or slightly concave, umbo opisthogyrate, ligament area triangular, with triangular resilifer, height of posterior bourrelet smaller than height of anterior bourrelet, PAM circular andin a posterodorsal position.
Description.—Shell inequivalve, outlines deltoid, subtriangular with height greater than length. LV convex, RV flat. Margins of valves taper dorsally. Attachment area small to relatively large, restricted to umbonal region. Maximum convexity close to anteroventral side. Sculpture of LV with growth lamellae, covered by weak radial stria. Growth lamellae of RV less thick than of LV. Ligament area ostreoid, small, triangular. Resilifer triangular, not deep, poorly marked, posteriorly directed. Height of anterior bourrelet greater than height of posterior bourrelet. Posterior adductor scar circular; position posterodorsal. Quenstedt muscle scar not observed. Chomata absent. Microstructure unknown.

Fig. 5. Microstructure of studied gryphaeid oysters. A. Deltoideum delta (Smith, 1817), TsSGM 2068/87; Kimmeridgian, Dorset, England. B. Pernostrea uralensis (Zakharov, 1972), TsSGM 2068/45; Upper Volgian, Maurynya River, eastern slopes of the Northern Urals. Microstructure (A1, B1), cross section (A2, B2). RF, regularly foliated structure; HCF, ?herringbone cross-foliated structure.
Remarks.—At the type locality, shells of D. (B.) borealis sp. nov. have been found in sand and glauconitic sand of Kimmeridgian age, as separated valves randomly distributed in the layers or rarely in lens-shaped shell accumulations (Zakharov 1966). According to Zakharov (1966) this species inhabited shallow, turbulent waters. However, in the Lopsiya River section, this species was also found in Lower Kimmeridgian clay. In this place the oysters inhabited a low-energy soft bottom environment. LV’s of oysters from the clay are more convex compared to specimens found in sand.
Deltoideum (Boreiodeltoideum) praeanabarensis (Zakharov, 1963) (Zakharov 1963: 50, pl. 7: 1–4; 1966: 103, pl. 38: 3–6, pl. 39: 1–2, pl. 40: 1–2; Kosenko 2016a: pl. 3: 12, 13, 15; Fig. 6B–D, F) is morphologically close to D. (B.) borealis sp. nov. It differs by a more elongated shell with the length greater than height, and the presence of an alate expansion posterior of the umbo.
D. (B.) anabarensis (Bodylevsky, 1949) (Zakharov 1966: 108, pl. 39: 3, pl. 40: 4, 5, pl. 42: 4–5; Kosenko 2016a: pl. 3: 9, 14, 16; SOM: fig. 3, Supplementary Online Material available at http://app.pan.pl/SOM/app62-Kosenko_SOM.pdf) differs by an extremely elongated and crescentic shaped shell.
“Ostrea” eduliformis (Zieten, 1832) (Zieten 1832: 60, pl. 45: 1; Madsen 1909: 200, pl. 7: 1–5) (this species belongs most likely to Pernostrea and it was usually assigned to Liostrea) from the Bajocian of Germany and Greenland differs by a less convex LV, a larger ligament area and the position of the PAM is less posteriorly shifted.
Stratigraphic and geographic range.—Lower Kimmeridgian of the Northern Urals, north of Eastern Siberia.

Fig. 6. Gryphaeid oysters from the subgenus Boreiodeltoideum. A, E. Deltoideum (Boreiodeltoideum) borealis sp. nov. A. TsSGM 2068/31, paratype, Lower Kimmeridgian, Lopsiya River, Northern Urals; exterior (A1) and interior (A2) of left valve. E. TsSGM 150/3887, holotype, Lower Kimmeridgian, Khatanga depression, north of Eastern Siberia; interior of RV (E1) and exterior of LV (E2). B–D, F. Deltoideum (Boreiodeltoideum) praeanabarensis (Zakharov, 1966), Lower Volgian, Dyabaka-Tari River, north of Eastern Siberia. B. TsSGM 2068/10, view on left (B1) and right (B2) valves. C. TsSGM 2068/57, interior of right valve. D. TsSGM 2068/8, exterior (D1) and interior (D2) of left valve. F. TsSGM 150/2031, interior of left valve.
Genus Pernostrea Munier-Chalmas, 1864
Type species: Perna bachelieri Orbigny, 1850 (= Ostrea luciensis Orbigny, 1850), Bathonian of France.
Emended diagnosis (modified from Stenzel 1971: N1104).—Shells medium to large, equivalve or inequivalve. Outlines rectangular, deltoid or crescentic. Some species have well developed marginal growth foliations. Ligament area ostreoid, occasionally multivincular, large, very broad, rectangular to trapezoidal. Attachment area small to very large (almost all of LV). Angle between resilifer and commissural plane usually around 10°–30°. PAM medium to large, circular to oval, clearly marked with a central to posterocentral position. No umbonal cavity. No chomata. Surface of valves covered with commarginal growth squamae. RV of some species covered by radial striae. Some species bear concentric plications and radial plicae (SOM: fig. 1A2). The crescentic depression for the gills and the brachytellum is well developed. Microstructure consists mainly of regularly foliated structure, but some species have interlayers with ?herringbone cross-foliated structure.
Remarks.—The name of the genus is based on the presence of a multivincular ligament, but actually only a few specimens of Pernostrea show this feature. This feature might be restricted to the type species.
Stratigraphic and geographic range.—Middle–Upper Jurassic of Europe (Alps, Poland, Romania, France, Germany, England, Switzerland) and Greenland, Lower Cretaceous of the Urals and the north of Eastern Siberia.
Pernostrea mesezhnikovi sp. nov.
Figs. 7–9, 10B, D.
2014 Deltoideum delta (Smith, 1817); Kosenko 2014: pl. 6: 4, 5, 7.
2014 Deltoideum ex gr. delta (Smith, 1817); Kosenko 2014: text-fig. 4, pl. 6: 6, 8.
2016 Deltoideum (Deltoideum) delta (Smith, 1817); Kosenko 2016a: 342, pl. 3: 4a–c, 5, 7, 8, 11.
ZooBank LSID: urn:lsid:zoobank.org:pub:26FBF7FF-F554-48DC-B02D-069507B4202B
Etymology: After Mikhail S. Mesezhnikov (1931–1989), specialist on the boreal Jurassic stratigraphy.
Type material: Holotype, a complete adult specimen, both valves with exposed ligament areas, TsSGM 2048/17 (Fig. 7A). Paratypes: TsSGM 2048/17-1, 2048/17-2, 2048/23, 2048/24, 2068/28, 2068/29, 2068/30, 2068/32, 2068/33, 2068/34, 2068/84-1, 2068/84-2, 2068/85, 2068/86, 2068/87); from the type locality.
Type locality: Lopsiya River, outcrop 41, the Northern Urals (North-West of Western Siberia), Russia.
Type horizon: Eosphinctoceras magnum Zone, base of the Lower Volgian, horizon with large “Ostrea” (sensu Mesezhnikov 1959).
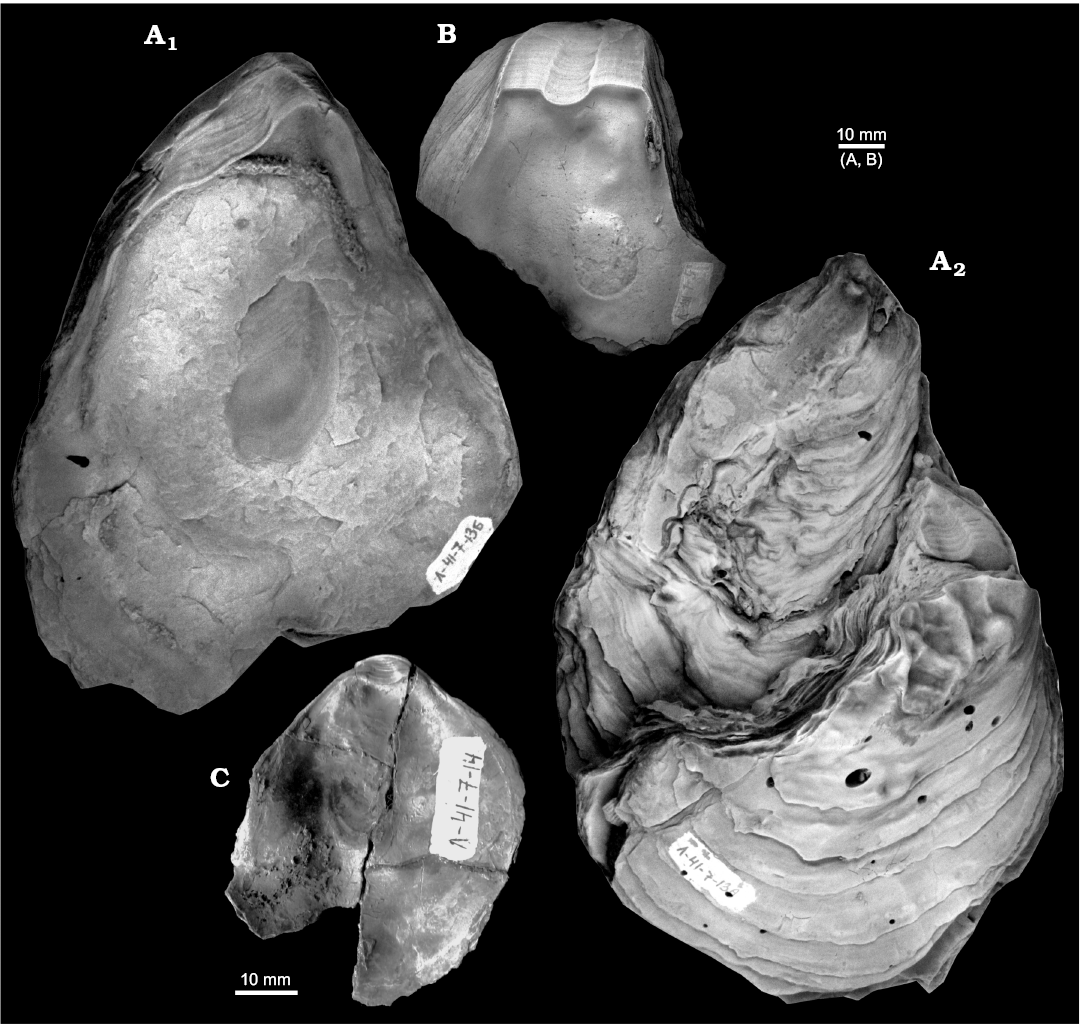
Fig. 7. Gryphaeid oyster Pernostrea mesezhnikovi sp. nov., Lower Volgian, Lopsiya River, Northern Urals. A. TsSGM 2048/17, holotype, interior of right valve (A1), exterior of left valve with attached oyster (A2). B. TsSGM 2048/24, paratype, interior of right valve. C. TsSGM 2068/33, paratype, interior of left valve.
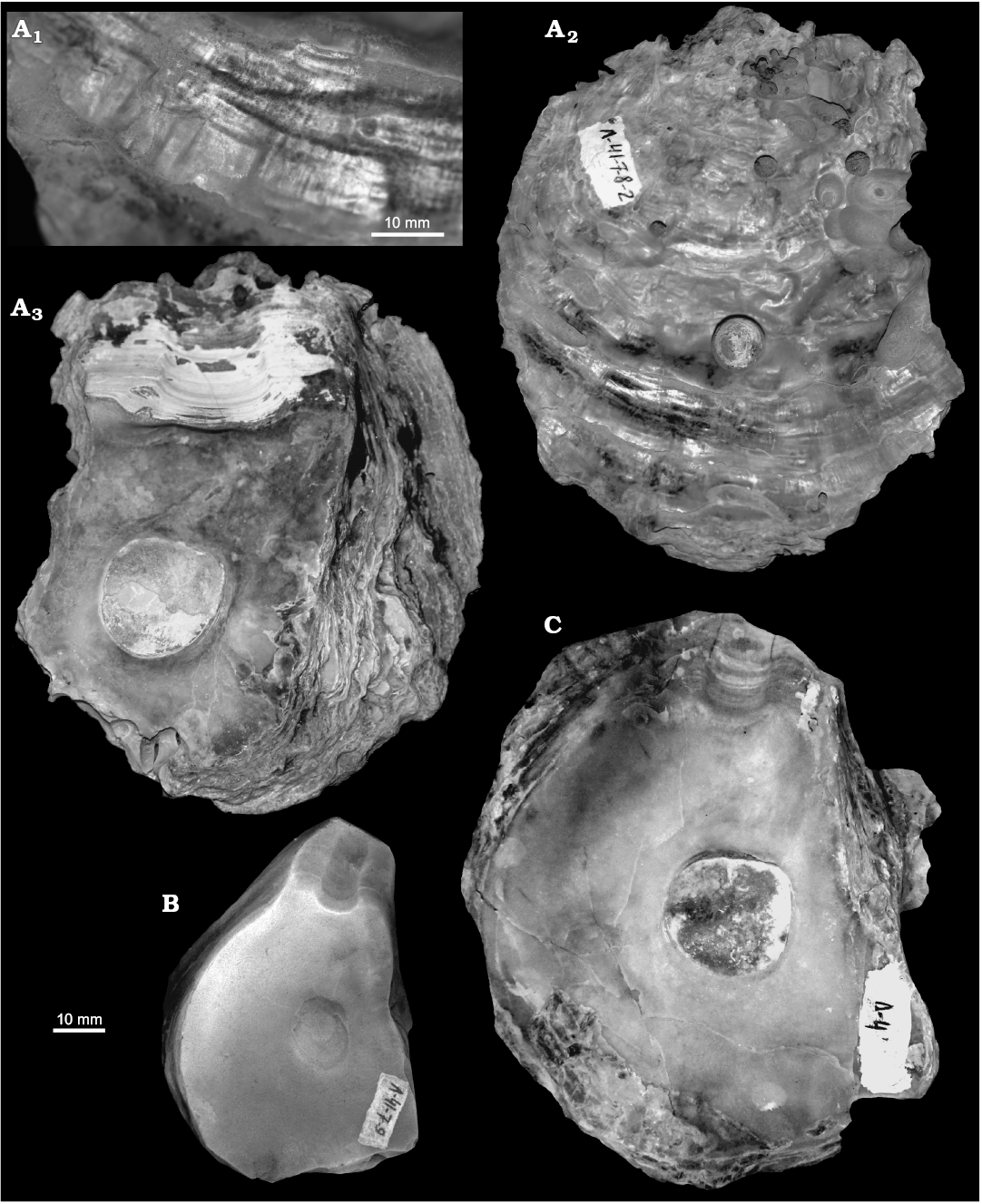
Fig. 8. Gryphaeid oyster Pernostrea mesezhnikovi sp. nov., Lower Volgian, Lopsiya River, Northern Urals. A. TsSGM 2068/28, paratype, details of right valve sculpture: fine radial striae (A1), exterior with Gastrochaenolites (A2), interior (A3). B. TsSGM 2048/21, interior of right valve. C. TsSGM 2068/29, interior of right valve.
Material.—Type material only.
Diagnosis.—Ligament area broad with resilifer subparallel to anterior and posterior bourrelets, PAM circular in a posterocentral position. Outer surface of valves covered by concentric growth squamae. RV with radial striae.
Description.—Large (maximum H = 170 mm, maximum L = 116 mm), usually equivalve (Table 1). Outlines variable, including oval, deltoid, trapezoidal and subtriangular shapes, usually slightly elongated in height. Shell shape sometimes irregular in clusters. Attachment area commonly very large, sometimes cemented with entire LV. Rarely attachment areas are very small and in such a case the shells have convex LVs and flat RVs (see Fig. 9). Shell height to length ratio (H/L) varies 0.95–1.46 with average of 1.23 (based on measurements of 15 shells from one layer). LV and RV covered by concentric growth squamae and by weak radial striae. Ligament area ostreoid, well developed, very wide, trapezoidal, rare rectangular, with a straight or slightly undulating resilifer. LV resilifer deep, flanked by equally broad, elevated bourrelets, angle between resilifer and commissural plane around 10°–30°. RV resilifer less deep than LV, cavity under RV resilifer absent. Ligament area length to shell length ratio (Lla/L) varies from 0.26 to 0.55 with average of 0.41 (based on measurements of 11 shells). PAM orbicular to oval; position usually subcentral or posterocentraland in LV usually buttressed. Quenstedt muscle scar displaced to posterior side of valve, its position corresponds to projection of posterior side of resilifer. No chomata. No chambers in shell microstructure. LV and RV with simply foliated structure.
Table 1. Results of measurments of morphometric parameters of gryphaeid oysters Pernostre mesezhnikovi sp. nov. Abbreviations: D1, distance between posterior adductor muscle scar and anterior valve margin; D2, distance between posterior adductor muscle scar and posterior valve margin; D3, distance between posterior adductor muscle scar and ventral margin; D4, distance between posterior adductor muscle scar and dorsal margin; H, shell height; Hla, ligament area height; Hpam, posterior adductor muscle scar height; L, shell length; LA, ligament area; LV, left valve; Lab, anterior bourrelet length; Lla, ligament area length; Lpam, posterior adductor muscle scar length; Lpb, posterior bourrelet length; Lr, resilifer length. adductor musc structure; RV, right valve.
|
Specimen |
H |
L |
Hla |
Lla |
Lab |
Lr |
Lpb |
Hpam |
Lpam |
D1 |
D2 |
D3 |
D4 |
H/L (%) |
Hla/H (%) |
Lla/L (%) |
D1/D2 |
|
TsSGM 2048/23 |
78 |
61 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
127.7 |
– |
– |
– |
|
TsSGM 2048/21 |
84 |
62 |
20 |
23 |
9 |
9 |
8 |
16 |
14 |
26 |
16 |
21 |
24 |
135.5 |
23.8 |
37.1 |
1.6 |
|
TsSGM 2048/22 |
69 |
56 |
8 |
19 |
5 |
8 |
5 |
16 |
15 |
25 |
13 |
24 |
22 |
123.6 |
11.6 |
33.9 |
1.9 |
|
TsSGM 2048/24 |
79 |
71 |
21 |
33 |
11 |
11 |
12 |
19 |
15 |
37 |
21 |
25 |
14 |
111.7 |
26.6 |
46.5 |
1.8 |
|
TsSGM 2048/17-1 |
170 |
115 |
18 |
49 |
14 |
24 |
12 |
– |
– |
– |
– |
– |
– |
147.8 |
10.6 |
42.6 |
– |
|
TsSGM 2048/17-2 |
120 |
109 |
19 |
24 |
7 |
11 |
7 |
– |
– |
– |
– |
– |
– |
109.3 |
15.8 |
22.0 |
– |
|
TsSGM 2068/84-1 |
86 |
93 |
14 |
42 |
13 |
13 |
16 |
20 |
22 |
39 |
28 |
26 |
26 |
93.1 |
16.3 |
45.2 |
1.4 |
|
TsSGM 2068/84-2 |
100 |
93 |
17 |
45 |
16 |
17 |
13 |
27 |
30 |
37 |
29 |
38 |
20 |
107.9 |
17.0 |
48.4 |
1.3 |
|
TsSGM 2068/28 |
108 |
90 |
31 |
53 |
14 |
27 |
13 |
25 |
24 |
18 |
12 |
24 |
29 |
119.3 |
28.7 |
58.9 |
1.5 |
|
TsSGM 2068/85 |
138 |
109 |
19 |
42 |
12 |
21 |
10 |
– |
– |
– |
– |
– |
– |
126.9 |
13.8 |
38.5 |
– |
|
TsSGM 2068/86 |
105 |
80 |
21 |
28 |
8 |
14 |
7 |
21 |
21 |
39 |
13 |
41 |
19 |
130.9 |
20.0 |
35.0 |
3.0 |
|
TsSGM 2068/30 |
111 |
116 |
20 |
56 |
16 |
26 |
14 |
24 |
25 |
53 |
32 |
24 |
41 |
95.5 |
18.0 |
48.3 |
1.7 |
|
TsSGM 2068/32 |
– |
77 |
30 |
36 |
12 |
11 |
12 |
– |
– |
– |
– |
– |
– |
– |
– |
46.8 |
– |
|
TsSGM 2068/33 |
59 |
47 |
5 |
13 |
5 |
6 |
4 |
10 |
9 |
22 |
12 |
16 |
24 |
125.3 |
8.5 |
27.7 |
1.8 |
|
TsSGM 2068/87 |
– |
– |
5 |
13 |
4 |
6 |
3 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
TsSGM 2068/29 |
116 |
104 |
20 |
36 |
11 |
14 |
10 |
27 |
28 |
39 |
19 |
26 |
37 |
111.6 |
17.2 |
34.6 |
2.1 |
|
TsSGM 2068/34 |
123 |
80 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
155.0 |
– |
– |
– |

Fig. 9. Gryphaeid oyster Pernostrea mesezhnikovi sp. nov., Lower Volgian, Lopsiya River, Northern Urals. A. TsSGM 2048/23, left (A1) and right (A2) valve views, view from posterior side (A3). B. TsSGM 2068/34, left (B1) and right (B2) valve views. C. TsSGM 2068/32, exterior (C1) and interior (C2) of left valve, view from anterior side (C3).
Remarks.—The species has been found in the uppermost Kimmeridgian–lowermost Volgian clay of the Lopsiya River section in association with ammonites, belemnites and bivalves. According to Zakharov in Zakharov and Mesezhnikov (1974), this assemblage is autochtonous or paraautochtonous. The species is found usually in aggregates consisting of two shells. Individually grown specimens are characterized by inequivalve shells with a convex LV and a concave RV (morphotype “Gryphaea-like recliner”, see Kosenko 2014: fig. 5).
Some species of Pernostrea and morphologically similar forms are discussed below in alphabetic order. Pernostrea luciensis (Orbigny, 1850) (Stenzel 1971: N1104, fig. J78: 1–3) is type species of the genus Pernostrea distributed in Bathonian–Callovian of France and England. It can be distinguished from P. mesezhnikovi sp. nov. by rectangular and circular outlines and broader ligament area (occasionally multivincular). Deltoideum delta (Smith, 1817) (Smith 1817: 18, pl. 5: 6; Arkell 1932: 149–155, text-figs. 22–26, pl. 16: 2–4, pl. 15: 6, pl. 14: 6; Fig. 10A, C) is widely distributed in Kimmeridgian–Tithonian species. Furthermore, Bogdanova (1988: 147, pl. 23: 2, pl. 24: 1, pl. 25: 4, pl. 26: 12) described D. delta from the Berriassian of Mangyshlak (Central Asia). This species first time has been described from the Upper Oxfordian–Lower Kimmeridgian of England. It can be distinguished from P. mesezhnikovi sp. nov. by extremely flat both LV and RV, narrowing of shell near ligament area and narrow and usually strongly dorsally elongated shape of ligament area. P. cucurbita (Zakharov, 1966) (Zakharov 1966: 111–112, pl. 41: 1, 2, pl. 42: 1; SOM: fig. 4) from the Valanginian of north of Eastern Siberia (Khatanga depression). This species differs by a dorsoposterior position of PAM, a less wide ligament area and the absence of radial striae on RV. “Ostrea” expansa Sowerby, 1819 (Sowerby 1819: 65, pl. 238: 1) was first described from the Tithonian of England. This species is also common in the Tithonian of northern France and was recently assigned to Helvetostrea by Koppka (2015). The species differentiates from P. mesezhnikovi sp. nov. by having a reniform shapeed PAM and a less developed ligament area. P. gibberosa (Zakharov, 1974) (Zakharov 1974: 147–148, pl. 31: 1, pl. 32: 1; SOM: fig. 2B, C) from the Lower–Middle Volgian of the Northern and the Subpolar Urals. This species can be distinguished by having a well-developed radial fold on the LV and a corresponding radial depression on its RV. Pernostrea? planoconvexa (Zakharov, 1974) (Zakharov 1974: 148, pl. 33: 1) from the Middle Volgian of the Subpolar Ural differs by an inequivalve shaped shell with a convex LV and a concave or flat RV. Pernostrea? robusta sp. nov. (Fig. 10) differs by a broader ligament area with a less deep resilifer and the strongly depressed, oval elongated, reniform PAM. P. uralensis (Zakharov, 1972) (Zakharov in Saks et al. 1972: 225–226, pl. 41: 1, pl. 42: 9, pl. 43: 5; SOM: fig. 1A) from the Upper Volgian–Ryazanian stage of the Northern and the Subpolar Urals. This species differs from P. mesezhnikovi sp. nov. by its isometric orbicular shell outlines, a central position of PAM and a low and broad ligament area. Some inequivalved specimens of P. mesezhnikovi sp. nov. with a prolonged height are similar to Helvetostrea and Crassostrea but differ from these genera by the absence of an umbonal cavity and structural chambers.
Stratigraphic and geographic range.—Upper Kimmeridgian–Lower Volgian of the Northern Urals (Lopsiya and Tolya rivers), the northwest of Western Siberia, Russia.
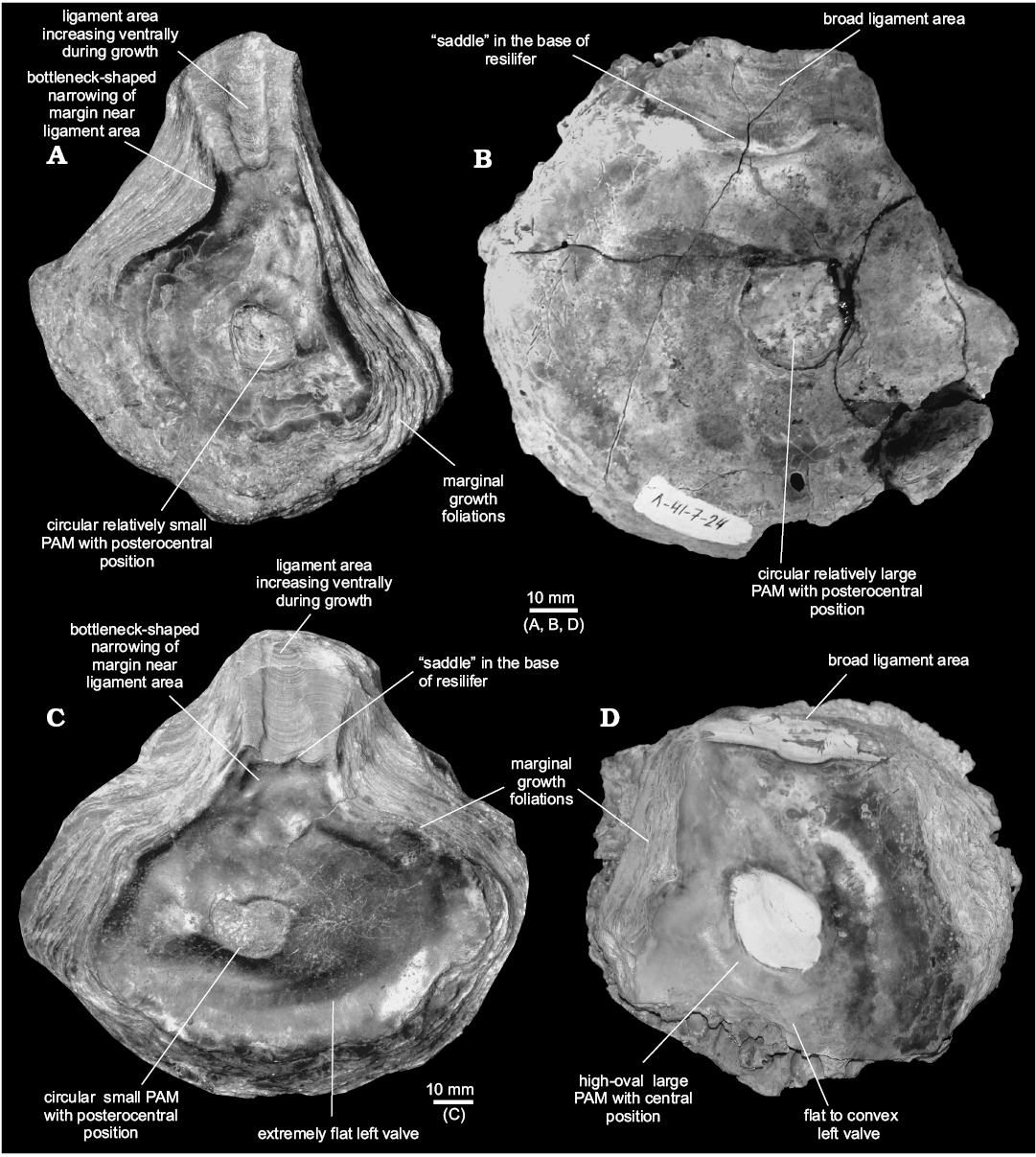
Fig. 10. Comparison of morphology Deltoideum vs. Pernostrea. A, C. Deltoideum delta (Smith, 1817) (labelled as “Ostrea deltoidea”), Kimmeridgian, La Hève, France. A. NHM 204314, interior of right valve. C. NHM 204314, interior of left valve. B, D. Pernostrea mesezhnikovi sp. nov., Lower Volgian, Lopsiya River, Northern Urals. B. TsSGM 2068/30, interior of right valve. D. TsSGM 2068/84, interior of left valve.
Pernostrea? robusta sp. nov.
Fig. 11.
2016 Deltoideum (Deltoideum) delta (Smith, 1817); Kosenko 2016a: pl. 3: 3.
ZooBank LSID: urn:lsid:zoobank.org:pub:26FBF7FF-F554-48DC-B02D-069507B4202B
Etymology: From Latin robustus, powerful, massive; in reference to gross shell morphology.
Type material: Holotype, adult right valve with exposed ligament area: TsSGM 2068/35 (Fig. 11C). Paratypes: TsSGM 2068/36, TsSGM 2068/38 from the type locality (Fig. 11B, D) and TsSGM 2068/37 from the Yatriya River (Fig. 11A).
Type locality: Tolya River, the Northern Urals (north-west of Western Siberia), Russia.
Type horizon: Middle Volgian silt (Pavlovia iatriensis Zone, Strajevskya strajevskyi Subzone) of the Tolya River.
Material.—Type material only.
Diagnosis.—Inequivalve; LV convex with deep internal cavity, RV flat; ligament area extremely large; PAM length-oval, strongly depressed.
Description.—Shell inequivalve, outlines subrectangular or trapezoidal. LV convex, very thick (maximum thickness 10 mm), relatively capacious (deepest part of LV located near anteroventral margin with maximum depth of 15 mm). RV slightly convex, relatively capacious, maximum depth near anteroventral margin as in LV. Attachment area large, almost entire LV. Observed maximum size (measurements were made on complete shells, hypothetic size of specimen 2068/37 is bigger than the measured specimens), H=110 mm, L = 10 mm. Shell height to length ratio (H/L) varies 0.94–1.21 (average 1.11, based on measurements of 5 shells). LV and RV covered with thick growth lamellae. Radial stria questionable because preservation of shells does not allow to observe them. Ligament area ostreoid, trapezoidal or rectangular, extremely elongated in length. Resilifer very shallow and extremely wide. Ligament area length to shell length ratio (Lla/L) varies 0.47–0.79 with average of 0.65 (based on the results of measuring 5 shells). Resilifer length to ligament area length ratio (Lr/Lla) varies 0.42–0.54 with average of 0.49 (based on the results of measuring 5 shells). PAM length-oval, very depressed, posterocentral. Quenstedt muscle located under posterior border of resilifer. Chomata absent. Microstructure not observed.
Remarks.—This species is characterized by length-oval shape of PAM whereas other oysters from the genus Pernostrea have circular shape of PAM. Due to these differences attribution of this species to the genus Pernostrea is questionable. This species has been found in the Middle Volgian silt of Tolya River and the Middle Volgian silt of Yatriya River. Only LV presented in studied collection is attached to valve of other oyster.
Helvetostrea expansa (J. Sowerby, 1819) is the most morphologically similar species to Pernostrea? robusta sp. nov. Helvetostrea expansa differs by having a much less broad ligament area. Koppka (2015) noted, that shells of H. expansa have a small umbonal cavity and large chambers below the ligament area, while such structural chambers are absent in P.? robusta sp. nov. Pernostrea gibberosa Zakharov, 1974 (Zakharov and Mesezhnikov 1974: 147, pl. 31: 1, pl. 32: 1; SOM: fig. 2B, C) differentiates by the presence of an U-shaped fold on the LV and a corresponding depression on the RV. Pernostrea? planoconvexa Zakharov, 1974 (Zakharov and Mesezhnikov 1974: 148. pl. 33: 1) differs by a less broad triangular ligament area, more convex elongated in length LV and the orbicular shape of PAM which is situated in a subcentral position. Pernostrea uralensis Zakharov, 1972 (SOM: fig. 1A) differs by having an orbicular and centrally positioned PAM. Other species of Pernostrea can be distinguished by the presence of an orbicular PAM, while the PAM of P.? robusta sp. nov. is length-oval and strongly depressed.
Stratigraphic and geographic range.—Middle Volgian of the Northern and the Subpolar Urals (Yatriya and Tolya rivers), the northwest of Western Siberia, Russia.
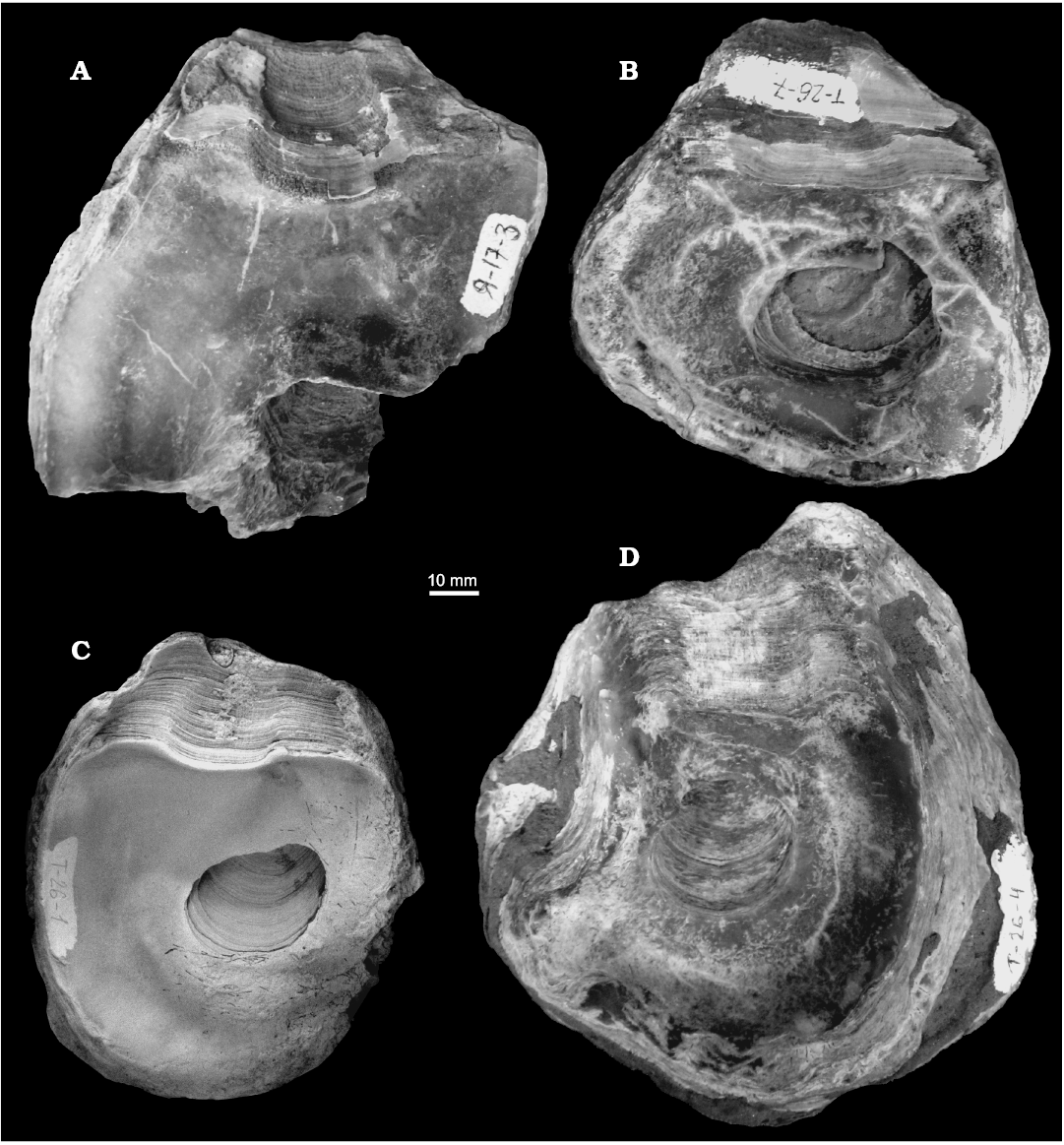
Fig. 11. Gryphaeid oyster Pernostrea? robusta sp. nov., Middle Volgian, Yatriya River, Subpolar Urals, . A. TsSGM 2068/37, interior of right valve. B. TsSGM 2068/36, interior of right valve. C. TsSGM 2068/35, interior of right valve. D. TsSGM 2068/38, interior of left valve attached to other oyster shell.
Systematic review of Late Jurassic–Early Cretaceous Ostreoidea from Siberia
Family Flemingostreidae Stenzel, 1971
Subfamily Crassostreinae Scarlato and Starobogatov, 1979
Genus Helvetostrea Koppka, 2015
Helvetostrea exotica (Kosenko, 2014); Upper Kimmeridgian (Fig. 12L, M)
Subfamily Liostreinae Vialov, 1983
Genus Praeexogyra Charles and Maubeuge, 1953
Praeexogyra siberica (Zakharov, 1974); Middle Volgian–Ryazanian (Fig. 12A, B, E, F, G)
Family Gryphaeidae Vialov, 1936
Subfamily Gryphaeinae Vialov, 1936
Tribe Pernostreini nov.
Genus Pernostrea Munier-Chalmas, 1864
Pernostrea cucurbita (Zakharov, 1966); Lower Valanginian (SOM: fig. 4)
Pernostrea gibberosa (Zakharov, 1974); Lower Volgian (SOM: fig. 2B, C)
Pernostrea mesezhnikovi sp. nov.; Upper Kimmeridgian–Lower Volgian (Figs. 7, 9, 10B, D)
Pernostrea? robusta sp. nov.; Middle–Upper Volgian (Fig. 11)
Pernostrea cf. quadrangularis (Arkell, 1927); Lower Kimmeridgian (SOM: fig. 5)
Pernostrea uralensis (Zakharov, 1972); Upper Volgian–Ryazanian (SOM: fig. 1A)
Genus Deltoideum Rollier, 1917
Subgenus Boreiodeltoideum Kosenko, 2016a
Deltoideum (Boreiodeltoideum) borealis sp. nov.; Lower Kimmeridgian (Fig. 5A, E)
Deltoideum (Boreiodeltoideum) praeanabarensis (Zakharov, 1963); Lower Volgian (Fig. 5B–D, F)
Deltoideum (Boreiodeltoideum) anabarensis (Bodylevsky, 1949); Ryazanian–Lower Hauterivian (SOM: fig. 3)
Tribe Gryphaeini Vialov, 1936
Genus Gryphaea Lamarck, 1801
Subgenus Gryphaea (Gryphaea) Lamarck, 1801
Gryphaea (Gryphaea) curva (Gerasimov, 1955); Lower Kimmeridgian (Fig. 12C, D)
Gryphaea? borealis Zakharov, 1966; Lower Valanginian (Fig. 12J)
Gryphaea sp.; Lower Volgian (Fig. 12I)
Subfamily Exogyrinae Vialov, 1936
Tribe Amphidontini Vialov, 1983
Genus Nanogyra Beurlen, 1958
Nanogyra? cf. thurmanni (Étallon, 1862); Middle Volgian (Fig. 12O)
Nanogyra? sp.; Lower Kimmeridgian (Fig. 12H)
Subfamily ?Pycnodonteinae Stenzel, 1959
Genus Argutostrea Kosenko in Kosenko and Seltser, 2016
Argutostrea roemeri (Quenstedt, 1843); Kimmeridgian–Middle Volgian (Fig. 12K, N)
Subfamily indet.
Genus indet.
“Ostrea” cf. moreana (Buvignier, 1852); Middle Volgian (SOM: fig. 2A)
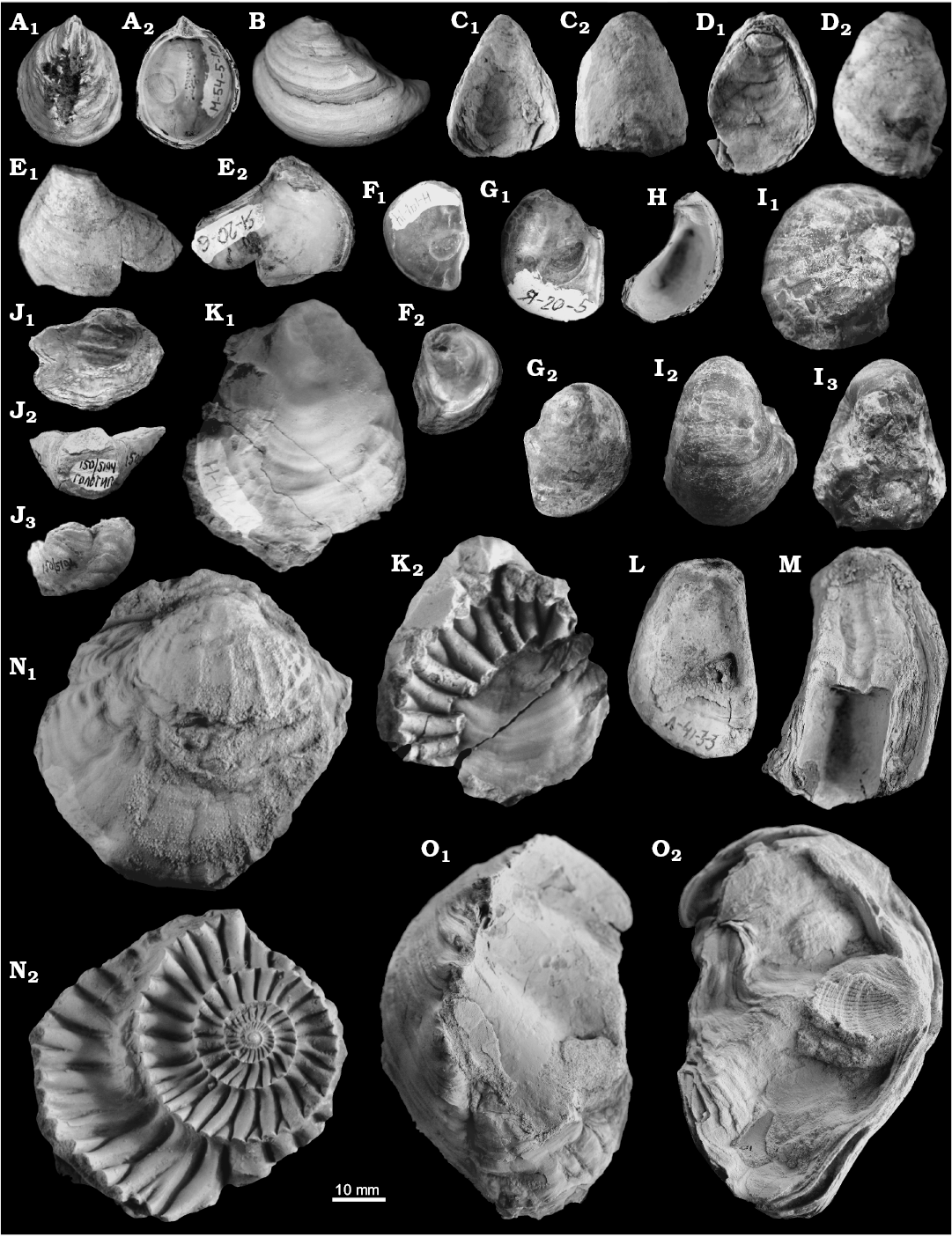
Fig. 12. Late Jurassic–Early Cretaceous oysters from Siberia. A, B, E. Praeexogyra siberica Zakharov, 1974, Upper Volgian, Maurynya River, the Northern Urals. A. TsSGM 2068/75, exterior (A1) and interior (A2) of left valve. B. TsSGM 2068/74, exterior of left valve. E. TsSGM 2068/76, exterior (E1) and interior (E2) of left valve. C, D. Gryphaea (Gryphaea) curva (Gerasimov, 1955), Upper Kimmeridgian, Lopsiya River, the Northern Urals. C. TsSGM 2068/71, interior (C1) and exterior (C2) of left valve. D. TsSGM 2068/72, right (D1) and left (D2) valve views. F, G. Praeexogyra siberica Zakharov, 1974, Middle Volgian, Yatriya River, the Subpolar Urals. F. TsSGM 2068/41-1, interior (F1) and exterior (F2) of right valve. G. TsSGM 2068/41-2, interior (G1) and exterior (G2) of right valve. H. Nanogyra? sp., TsSGM 2068/42, Kimmeridgian, Lopsiya River, the Northern Urals; interior of left valve. I. Gryphaea (Gryphaea) sp., TsSGM 2068/43 (collection 150), Lower Volgian, Khatanga depression, north of Eastern Siberia; posterior (I1), left (I2) and right (I3) valves views. J. Gryphaea? borealis (Zakharov, 1966), TsSGM 150/5194, Lower Valanginian, Khatanga depression, north of Eastern Siberia; right valve (J1), umbo (J2), and posterior (J3) views. K, N. Argutostrea roemeri (Quenstedt, 1843), Middle Volgian, Yatriya River, the Subpolar Urals. K. TsSGM 2068/77, exterior of right valve (K1), exterior of left valve with impression of ammonite shell (K2). N. TsSGM 2068/79, exterior of right (N1) and left (N2) valves with xenomorphic sculpture. L, M. Helvetostrea exotica (Kosenko, 2014), Upper Kimmeridgian, Lopsiya River, the Northern Urals. L. TsSGM 2048/6, paratype, interior of right valve. M. TsSGM 2048/2, paratype, interior of left valve. O. Nanogyra? cf. thurmanni (Étallon, 1862), TsSGM 2068/73, Middle Volgian, Yatriya River, the Subpolar Urals; exterior of left (O1) and right (O2) valves.
Discussion
Variability and the problem of species identification Deltoideum vs. Pernostrea.—Zakharov (1963, 1966; in Saks et al. 1972; in Zakharov and Mesezhnikov 1974) regarded all oysters included here in the genus Pernostrea as belonging to Liostrea. The Siberian oysters can be distinguished from that genus by the absence of an umbonal cavity in the LV. However, Zakharov (1978) assigned them later to the genus Deltoideum, a decision which was followed by Kosenko (2014, 2016a). However, when the shell morphology of Siberian oysters has been compared to characteristics of the genera Pernostrea and Deltoideum (Table 2) it turned out that the Siberian oysters (classified here as Pernostrea) share some morphological characters typical for Pernostrea as well as for Deltoideum. For example, Siberian oysters usually have extremely broad ligament areas with a broad resilifer subparallel to the anterior and posterior bourrelets what is typical for Pernostrea, and radial striae on the outer surface of RV, what is characteristic for Deltoideum. There are also other morphological characters in common for Siberian Pernostrea and Deltoideum (e.g., flatness of both valves and sometimes shell outlines) and altogether it suggests that Pernostrea and Deltoideum are morphologically very close to each other. In addition, the principal construction of the ligament area of Deltoideum and Pernostrea is basically the same, except for the occasional presence of multivincular ligament areas which are restricted solely to Pernostrea. Both genera have normally an ostreoid type of ligament area and the difference lies mainly in its relative length which is short in Pernostrea and long and curved in Deltoideum. Another morphological character, which distinguishes Pernostrea from Deltoideum, is the typical bottleneck-shaped narrowing of valve margins ventral of the ligament area in Deltoideum and absent in Pernostrea. The construction of the ligament area, suggests the large Siberian oysters “Liostrea” gibberrosa, “L.” uralensis, “L.” cucurbita, and morphologically similar species described in present paper for the first time need to be included in Pernostrea. The position of the PAM cannot be regarded as an important character to distinguish these two genera because of its wide range of variability. For example, Deltoideum usually has its PAM close to the posterior margin, but more symmetrical shells of Deltoideum have a centered PAM (Fig. 10C).
In the present paper, the Siberian oysters are included in the genus Pernostrea due to their extremely broad ligament areas and the lack of transversally constricting anterior and posterior margins ventral to the ligament area as it is present in Deltoideum (Stenzel 1971: N1103, fig. J76: 4, 5; Fig. 10A, C). Moreover, observations of Siberian oysters show, that the variability of their shells is very high and this makes it sometimes difficult to identify the generic affiliation of untypical (xenomorphic/aberrant) specimens. The shape of the shell strongly depends on the type of sediment and the size of the attachment area (e.g., morphotypes of P. mesezhnikovi sp. nov.).
Table 2. Comparison of morphological characters of Deltoideum, Pernostrea and Siberian oysters. “Saddle” in the base of resilifer is a deformation of the ventral margin of the resilifer dividing this margin into two parts (see also Fig. 10B, C).
|
Morphological character |
Pernostrea |
Deltoideum |
Siberian oysters |
|
Posterior adductor muscle (shape and position) |
orbicular, posterocentral |
orbicular, posterodorsal, posterocentral or central |
orbicular, posterocentral or central |
|
Ligament area |
broad ligament area with broad rectangular resilifer and equally developed subparallel bourrelets; some shells with multiple resilifer |
anterior bourrelet usually is approximately 1.5 times longer than a posterior one; ventrally prolonged ligament area |
broad ligament area with equally developed subparallel bourrelets and a wide straight resilifer; some shells have “saddle” in the base of resilifer (characters of Pernostrea) |
|
Shell form |
equivalve or sligthly inequivalve |
flat equivalve shells |
equivalve or inequivalve, sometimes flat (characters of Pernostrea and Deltoideum) |
|
Shell outlines |
oval, orbicular, rectangular |
triangular, spatulate, pear-or crescent-shaped, prominent branchitellum, strong constrictions near ligament area |
oval, orbicular, rectangular, deltoid (characters of Pernostrea) |
|
Radial sculpture |
without radial sculpture |
weakly marked radial striae |
weakly marked radial striae (characters of Deltoideum) |
|
Marginal growth foliations |
no marginal growth foliations |
marginal growth foliations |
marginal growth foliations sometimes well developed |
|
Shell microstructure |
unknown |
regularly foliated |
regularly foliated with interlayers of ?herringbone cross foliated structure |
|
Geographic distribution |
Europe, Greenland |
Europe, Saudi Arabia, Central Asia |
Siberia |
Taxonomic position of Pernostrea and Deltoideum in the superfamily Ostreoidea.—The position of Pernostrea and Deltoideum within the Ostreoidea is controversial. Stenzel (1971) included these genera in the subfamily Gryphaeinae due to the presence of a circular PAM in a central or centrodorsal position. Todd (1993) shared the same opinion and Malchus (1990) considered these genera to be a part of the tribe Liostreini (he also included to this tribe the genera Liostrea, Catinula, Praeexogyra, and Bosostrea) incorporated into the subfamily Liostreinae Vialov, 1983. Kosenko (2016a) followed Malchus (1990) and included the genus Deltoideum in the subfamily Liostreinae. Later, Nicolaus Malchus (personal communication 2015), suggested returning to Stenzel’s (1971) point of view that these genera belong to the subfamily Gryphaeinae (because of the shape of PAM); however, he believed that there is not enough data for the final solution at the moment.
The morphology of the genera Pernostrea and Deltoideum has a number of characters that are not typical for the majority of the Gryphaeinae. These characters include a flattened shell shape (typical Gryphaea has a strongly convex LV and a flat or even concave RV), absence of “beaked” umbo (Gryphaea has a strongly “beaked” orthogyrate umbo on LV), and the ligament area meeting the commissural plane in a small angle (for Gryphaea, the ligament area is usually subperpendicular to commissural plane), as well as a greater degree of shell shape variation. Characters of Pernostrea and Deltoideum in common with a typical Gryphaea are a circular shape of PAM and its position (subcentral or centrodorsal), as well as the absence of a marked umbonal cavity and chambers in microstructure of the shell (Fig. 6). However, the microstructure of examined Pernostrea-shells consists mainly of simply foliated structure (P. uralensis has interlayers of structure similar to herringbone cross-foliated structure) while the microstructure of a typical Gryphaea consists of simply cross-foliated, belt-foliated, and irregular complex cross-foliated structures (Malchus 1990). Main difference from the typical Liostreinae is the lack of a marked umbonal cavity (Table 3). In addition, representatives of the subfamily Liostreinae usually have their PAM shifted to the ventral side or a comma-shaped PAM in a posterodorsal position, and generally a small triangular ligament area. For basal Liostreinae (such as Liostrea), as well as for Gryphaeinae, typical is the absence of structural chambers in their shells (Malchus 1998). Furthermore, species of Pernostrea from Siberia share some common characters with Crassostreinae such as the circular PAM with subcentral position, spatulate outlines and a similar mode of life. However, they differ from Crassostreinae by absence of umbonal cavity and absence of chambers in microstructure of shell. This short review shows that the genera Deltoideum and Pernostrea differ both from the typical Gryphaeinae as well as from Liostreinae and at the same time these genera share several morphological characters with both families.
According to Stenzel (1971), the shape and position of the PAM is a taxonomical character of family rank. The family Gryphaeidae is characterized by an orbicular PAM with a centerodorsal to posterodorsal position. Based on this, genera as Pernostrea and Deltoideum need to be included to the Gryphaeidae. The absence of an umbonal cavity in Pernostrea and Deltoideum is an additional argument for including them in the Gryphaeidae and not to the Liostreinae; the absence of chomata and a vesicular microstructure and the ostreoid ligament area suggests that these genera belong to the subfamily Gryphaeinae and not to the Pycnodontinae.
The close morphological similarity of Pernostrea and Deltoideum and their differences from a typical Gryphaea provide the basis to distinguish the new tribe Pernostreini trib. nov. of the subfamily Gryphaeinae. Only Gryphaea, including the subgenera Africogryphaea, Bilobissa, and Gryphaea is here regarded as a member of the tribe Gryphaeini Vialov, 1936.
Table 3. Comparison of morphological characters of subfamilies Liostreinae, Gryphaeinae, Crassostreinae, and tribe Pernostreini trib. nov.
|
|
Liostreinae |
Gryphaeinae |
Crassostreinae |
Pernostreini |
|
Ligament area |
ostreoid, peaked triangular |
gryphaeoid |
ostreoid or gryphaeoid |
ostreoid, triangular or rectangular |
|
Posterior adductor muscle scar (shape and position) |
high-oval posteroventral, biconcave posteroventral, comma-shaped posterodorsal, crescent/reniform posteroventral |
circular posterodorsal, reniform centerodorsal |
circular centered, high-oval posteroventral, comma-shaped posteroventral, reniform posteroventral |
circular posterodorsal, posterocenter, centerodorsal or central, reniform centered |
|
Microstructure |
regularly foliated |
simply cross-foliated, belt- foliated, irregular complex cross-foliated |
irregular cross-foliated, regularly foliated |
regularly foliated, ?herringbone cross-foliated |
|
Chambers |
sometimes present |
absent |
present |
absent |
|
Chomata |
absent or present |
absent |
absent or present |
absent |
|
Umbonal cavity |
present or absent |
absent |
present |
absent |
Conclusions
The taxonomy of Siberian Late Jurassic–Early Cretaceous oysters has been revised. Oysters with thick shells were firstly described by Zakharov (1963, 1966; in Saks et al. 1972; in Zakharov and Mesezhnikov 1974) and attributed to the genus Liostrea (L. gibberosa, L. uralensis, L. cucurbita) and are reassigned herein to the genus Pernostrea. A comparison of the shell morphology of the genera Deltoideum and Pernostrea shows that they are morphologically close to each other but differ from Gryphaea in many ways, which allow to introduce a new tribe Pernostreini trib. nov. as part of the subfamily Gryphaeinae. Thus, the subfamily Gryphaeinae is divided into the tribes Pernostreini trib. nov. and Gryphaeini Vialov, 1936. Three new species of the new tribe are described: Deltoideum (Boreiodeltoideum) borealis sp. nov., Pernostrea mesezhnikovi sp. nov. and Pernostrea? robusta sp. nov. The species Nanogyra? cf. thurmanni and “Ostrea” cf. moreana are recorded for the first time from the Middle Volgian of Siberia.
Acknowledgements
I am grateful to Krzysztof Hryniewicz (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland), Jens Koppka (University of Greifswald, Germany) and Clémentine Colpaert (Novosibirsk State University, Russia) for the assistance in correction of English language, Nikolaus Malchus (ICP Catalan Institute of Palaeontology Miquel Crusafont, Barcelona, Spain), Jens Koppka, Krzysztof Hryniewicz, and Dario Lazo (Universidad de Buenos Aires, Argentina) for their review of the article, useful comments and advices during the review process and taxonomical discussions. Thanks also to Boris Shurygin, Oksana Dzyuba, Aleksandr Igol’nikov (all Institute of Petroleum Geology and Geophysics, Siberian Branch, Russian Academy of Science, Novosibirsk, Russia) for their useful advice and valuable remarks during the preparation of the present paper. Furthermore, I am indepted to Viktor Zakharov, Mikhail Rogov (both Geological Institute, Russian Academy of Sciences, Moscow, Russia), Tamara Bogdanova (VSEGEI, All Russia Research Geological Institute, St. Petersburg, Russia), Vladimir Arkadiev (St. Petersburg State University, Russia), Vasiliy Mitta (Paleontological Institute, Russian Academy of Sciences, Moscow, Russia), and to Jonathan Todd (Natural History Museum, London, UK) for useful advices on oyster taxonomy. I am thankful to Thierry Oudoire (Lille Natural History Museum, France) and Jessie Cuvelier (University Lille, Sciences and Technologies, UMR 8198 Evo-Eco-Paleo, France) for the opportunity to work with the collections of oysters in the Lille Natural History Museum. The work was performed with support of Russian Foundation of Basic Research (project 16-35-00003). This paper is a contribution to the UNESCO-IUGS IGCP Project 632.
References
Alifirov, A.S., Igol’nikov, A.E., and Dzyuba, O.S. [Dzûba, O.S.] 2008. Ammonites and the structure of the section of Volgian–Berriasian deposits on the Mauryn’ya River (Subpolar Urals): New data [in Russian]. In: O.S. Dzûba, V.A. Zaharov, and B.N. Šurygin (eds.), Melovaâ sistema Rossii i bližnego zarubež’â: problemy stratigrafii i paleogeografii, 20–23. IPGG SB RAS, Novosibirsk.
Arkell, W.J. 1927. The Corallian rocks of Oxford, Berks and North Wilts. Philosophical Transactions of the Royal Society of London, Series B 216: 67–181.
Arkell, W.J. 1932. A monograph of British Corallian Lamellibranchia. Monograph of Palaeontographical Society 4: 133–180.
Beurlen, K. 1958. Die Exogyren. Ein Beitrag zur phylogenetischen Morphogenese der Austern. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1958 (5): 197–217.
Bodylevsky, V.I. [Bodylevskij, V.I.], Mordvilko, T.A., and Luppov, N.P. 1949. Class Lamellibranchiata [in Russian]. In: N.P. Luppov (ed.), Аtlas rukovodâŝih form iskopaemyh faun SSSR, tom 10, nižnij mel, 120–159. Gosudarstvennoe izdatel’stvo geologičeskoj literatury, Moskva.
Bronn, H.G. 1834–1838. Lethaea geognostica. 1–48 (1834), 49–224 (1835) + Atlas 1–8 (1836), 225–480 (1837), i–vi + 481–768 (1838), 769–1350 + Atlas 9–16 (1838). Schweizerbart, Stuttgart,
Bogdanova, T.N. 1988. Type Mollusca. Class Bivalvia [in Russian]. In: G.Y. Krymgolts (ed.), Berrias Mangyšlaka, 134–156. Nauka, Leningrad.
Boss, K.J. 1982. Mollusca. In: S.P. Parker (ed.), Synopsis and Classification of Living Organisms. Vol. 1, 945–1166. McGraw-Hill, New York.
Buvignier, A. 1852. Statistique géologique, minéralogique, minérallurgique et paléontologique du département de la Meuse, ouvrage accompagné d’un Atlas de 32 planches. 694 pp. + Atlas 52 pp. Baillière, Paris.
Carter, J.G., 1990. Shell microstructural data for the Bivalvia. Part IV. Order Ostreoida. In: J.G. Carter (ed.), Skeletal Biomineralization: Patterns, Processes and Evolutionary Trends. Vol. 1, 347–362. Van Nostrand Reinhold, New York.
Carter, J.G., Harries, P.J., Malchus, N., Sartori, A.F., Anderson, L.C., Bieler, R., Bogan, A.E., Coan, E.V., Cope, J.C.W., Cragg, S.M., García-March, J.R., Hylleberg, J., Kelley, P., Kleemann, K., Kříž, J., McRoberts, C., Mikkelsen, P.M., Pojeta, J. jr., Tëmkin, I., Yancey, T., and Zieritz, A. 2012. Illustrated Glossary of the Bivalvia, Part N, Revised. Treatise Online 48: 1–209.
Charles, R.P. and Maubeuge, P.-L. 1953. Les huîtres plissées jurassiques de l’est du Bassin Parisien (pt. 2). Bulletin du Musée d’Histoire Naturelle de Marseille 12: 113–123.
Cheltsova, N.A. [Čeltsova, N.A.] 1969. Značenie mikrostruktury rakoviny melovyh ustric dlâ ih sistematiki. 83 pp. Nauka, Moskva.
Devyatov, V.P. [Devâtov, V.P.], Nikitenko, B.L., and Shurygin, B.N. [Šurygin, B.N.] 2011. Jurassic paleobiogeography of Siberia during major changeovers [in Russian]. Novosti paleontologii i stratigrafii 16–17: 87–101.
Dzyuba, O.S., Izokh, O.P., and Shurygin, B.N. 2013. Carbon isotope excursions in Boreal Jurassic–Cretaceous boundary sections and their correlation potential. Palaeogeography, Palaeoclimatology, Palaeoecology 381–382: 33–46.
Étallon, A. 1962. Etudes paléontologiques sur le Haut-Jura – Vertébrés, articulés, mollusques. Mémoires de la Société d’Émulation du Département du Doubs 3 (6): 53–260.
Gerasimov, P.A. 1955. Rukovodâŝie iskopaemye mezozoâ Central’nyh oblastej evropejskoj časti SSSR. 379 pp. Gosgeoltehizdat, Moskva.
Férussac, A.E.J.A. de 1821–1822. Tableaux Systèmatiques des Animaux Mollusques suivis d’un Prodrome Général pour tous les Mollusques Terrestres ou Fluviatiles Vivants ou Fossiles. Première Partie, Tableaux Systématiques Généraux. 114 pp. Arthus-Bertrand, Paris.
Grobben, C. 1894. Zur Kenntniss der Morphologie, der Verwandtschaftsverhältnisse und des Systems der Mollusken. Kaiserliche Akademie der Wissenschaften (Mathematisch-Naturwissenschaftlichen Classe), Sitzungsberichte 103 (1): 61–86.
Harry, H.W. 1985. Synopsis of the supraspecific classification of living oysters (Bivalvia: Gryphaeidae and Ostreidae). The Veliger 28: 121–158.
Koppka, J. 2015. Revision of the Bivalvia from the Upper Jurassic Reuchenette Formation, Northwest Switzerland—Ostreoidea. Zootaxa 3927: 1–117.
Kosenko, I.N. 2014. On Late Jurassic and Early Cretaceous oysters (Bivalvia, Ostreidae) from northern Siberia. Paleontological Journal 48: 380–388.
Kosenko, I.N. 2016a. On Late Jurassic and Early Cretaceous oysters of the genus Deltoideum Rollier (Bivalvia, Ostreoidea) from Siberia. Paleontological Journal 50: 336–346.
Kosenko, I.N. 2016b. Upper Jurassic to Early Cretaceous Oysters (Bivalvia, Ostreoidea) from Siberia: Taxonomy and Variations of Carbon and Oxygen Isotopes. International Journal of Geological and Environmental Engineering 3 (11), waset.org/abstracts/52889.
Kosenko, I.N. and Seltser, V.B. 2016. Argutostrea gen. nov.: Questions of taxonomy and ethology of Middle–Late Jurassic epibiont oysters (Bivalvia, Ostreoidea). Paleontological Journal 50: 578–584.
Kosenko, I.N., Dzyuba, O.S. [Dzûba, O.S.], and Shurygin, B.N. [Šurygin, B.N.] 2013. Variations of carbon and oxygen isotopes around the Jurassic/Cretaceous boundary strata of the Northern Urals: comparative analyses of belemnites and oysters [in Russian]. In: V.A. Zaharov (ed.) Ûrskaâ sistema Rossii: problemy stratigrafii i paleobiogeografii. Pâtoe Vserossijskoe soveŝanie, 123–125. IzdatNaukaServis, Ekaterinburg.
Lamarck, J.B. de 1801. Système des animaux sans vertèbres, ou tableau général des classes, des ordres et des genres de ces animaux. viii + 432 pp. Chez Deterville, Paris.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Madsen, V. 1909. In Jurassic Fossils from East Greenland. Meddelelser om Grønland 29: 157–208.
Malchus, N. 1990. Revision der Kreide-Austern (Bivalvia, Pteriomorphia) Ägyptens (Biostratigraphie, Systematik). Berliner Geowissenschaftliche Abhandlungen, Reiche A 125: 1–231.
Malchus, N. 1998. Multiple parallel evolution and phylogenetic significance of shell chambers and chomata in the Ostreoidea (Bivalvia). In: P.A. Johnston and J.W. Haggart (eds.), Bivalves: An Eon of Evolution, 393–407. University of Calgary Press, Calgary.
Mesezhnikov, M.S. [Mesežnikov, M.S.] 1984. Kimeridžskij i volžskij ârusy severa SSSR. 224 pp. Nedra, Leningrad.
Munier-Chalmas, E.C.P.A. 1864. Description d’un nouveau genre monomyaire du terrain jurassique. Journal de Conchyliologie: comprenant l’étudie des mollusques vivants et fossils, Serie 3 4: 71–75.
Ó Foighil, D. and Taylor, D. 2000. Evolution of parental care and ovulation behavior in oysters. Molecular Phylogenetics and Evolution 15: 301–313.
Orbigny, A. de 1850. Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés. Tome 1 & 2. 394 pp. and 428 pp. Victor Masson, Paris.
Quenstedt, F.A. 1843. Das Flӧzgebirge Würtembergs. Mit besonderer Rücksicht auf den Jura. 558 pp. Laupp, Tübingen.
Raith, M., Zacherl, D.C., Pilgrim, E.M., and Eernisse, D.J. 2016. Phylogeny and species diversity of Gulf of California oysters (Ostreidae) inferred from mitochondrial DNA. American Malacological Bulletin 33: 263–283.
Rafinesque, C.S. 1815. Analyse de la nature ou tableau de l’Université et des corps organisés, etc. 223 pp. Jean Barravecchia, Palermo.
Rollier, L. 1917. Fossiles nouveaux ou peu connus des terrains secondaries (Mesozoïques) du Jura et des contrées environnantes. Mémories de la Société Paléontologique Suisse, 6–7 partie 42: 501–696.
Saks, V.N. (ed.), Basov, V.A., Burdykina, M.D., Gol’bert, A.V., Dagis, A.S., Zakharov, V.A. [Zaharov, V.A.], Ivanova, E.F., Klimova, I.G., Mesezhnikov, M.S. [Mesežnikov, M.S.], Nal’nyaeva, T.I. [Nal’nâeva, T.I.], Pokhilajnen, V.P. [Pohilajnen, V.P.], Romanova, E.E., Ronkina, Z.Z., Sazonova, I.G., Trushnikova, L.Ya. [Trušnikova L.Â.], Shul’gina, N.I. [Šul’gina, N.I.], Yudovnyj, E.G. [Ûdovnyj E.G.]. 1972. Granica ûry i mela i berriasskij ârus v Boreal’nom poâse. 371 pp. Nauka, Novosibirsk.
Salvi, D., Macali, A., and Mariottini, P. 2014. Molecular phylogenetics and systematics of the bivalve family Ostreidae based on rRNA sequence-structure models and multilocus species tree. PLoS ONE 9 (9): e108696.
Scarlato, O.A. 1960. Dvustvorčatye mollûski dal’nevostočnyh morej SSSR (otrâd Dysodonta). 151 pp. Akademiâ Nauk SSSR, Leningrad.
Scarlato, O.A. and Starobogatov, Y.I. [Starobogatov, Â.I.] 1979. The general features of the evolution and systematics of the Bivalvia [in Russian]. In: Â.I. Starobogatov (ed.), Morfologiâ, sistematika i filogeniâ mollûskov. Trudy Zoologičeskogo Instituta Аkademii Nauk SSSR 80: 5–38.
Shurygin, B.N. and Dzyuba, O.S. 2015. The Jurassic/Cretaceous boundary in northern Siberia and Boreal-Tethyan correlation of the boundary beds. Russian Geology and Geophysics 56: 652–662.
Shurygin, B.N. [Šurygin, B.N.], Nikitenko, B.L., Devyatov, V.P. [Devâtov, V.P.], Ilyina, V.I., Meledina, S.V., Gaideburova, S.V., Dzyuba, O.S. [Dzûba, O.S.], Kazakov, A.M., and Mogucheva, N.K. [Mogučeva, N.K.] 2000. Stratigrafiâ neftegazonosnyh bassejnov Sibiri. Ûrskaâ sistema. 480 pp. SB RAS, Department GEO, Novosibirsk.
Smith, W. 1817. Stratigraphical System of Organized Fossils, With Reference to the Specimens of the Original Geological Collection in the British Museum: Explaining Their State of Preservation and Their Use in Identifying the British Strata. 118 pp. E. Williams, London.
Sowerby, J. 1819. The Mineral Conchology of Great Britain. Vol. 3 (2), 41–98. Arding & Merett, London.
Stenzel, H.B. 1959. Cretaceous oysters of southwestern North America. International Geological Congress, XX Session, Mexico City, 1956, El Sistema Cretácico 1: 15–37.
Stenzel, H.B. 1971. Oysters. In: R.C. Moore (ed.), Treatise on Invertabrate Paleontology (N) Mollusca 6, Bivalvia 3, N953–N1224. University of Kansas, Lawrence.
Tёmkin, I. 2010. Molecular phylogeny of pearl oysters and their relatives (Mollusca, Bivalvia, Pterioidea). BMC Evolutionary Biology 10 (342): 2–28. Crossref
Todd, J.A. 1993. The bivalve shell as a preservation trap, as illustrated by the Late Jurassic gryphaeid Deltoideum delta (Smith). Scripta Geologica, Special Issue 2: 417–433.
Vialov, O.S. 1936. On the classification of the oysters [in Russian]. Doklady Akademii Nauk SSSR 4 (1): 2–5.
Vialov, O.S. 1983. General classification of oysters [in Ukrainian]. Doklady Аkademii Nauk Ukrainskoj SSR, Seriâ B 11: 6–8.
Zakharov, V.A. [Zaharov, V.A.] 1963. On Late Jurassic and Early Cretaceous oysters of the Arctic Seas of Siberia [in Russian]. Paleontologičeskij žurnal 1963 (4): 44–52.
Zakharov, V.A. [Zaharov, V.A.] 1966. Pozdneûrskie i rannemelovye dvustvorčatye mollûski severa Sibiri (otrâd Anisomyaria) i usloviâ ih suŝestvovaniâ. 189 pp. Nauka, Moskva.
Zakharov, V.A. [Zaharov, V.A.] 1970. Pozdneûrskie i rannemelovye dvustorčatye mollûski severa Sibiri i usloviâ ih suŝestvovaniâ. Čast’ II. Semejstvo Astartidae. 144 pp. Nauka, Moskva.
Zakharov, V.A. [Zaharov, V.A.] 1972. Berriasian marine fauna. Bivalvia [in Russian]. In: V.N. Saks (ed.), Granica ûry i mela i berriasskij ârus v Boreal’nom poâse, 215–229. Nauka, Novosibirsk.
Zakharov, V.A. [Zaharov, V.A.] 1978. Arctic Mesozoic oysters [in Russian]. In: L.A. Nevesskaâ, V.A. Sobeckij, and A.N. Ânakevič (eds.), Tezisy dokladov II Vsesoûznogo simpoziuma po morfologii, sisteme, filogenii i èkogenezu dvustvorčatyh mollûskov, 20–23. Gosudarstvennyj Pedagogičeskij Institut, Tiraspol’.
Zakharov, V.A. [Zaharov, V.A.] 1981. Buhiidy i biostratigrafiâ boreal’noj verhnej ûry i neokoma. 270 pp. Nauka, Moskva.
Zakharov, V.A. 2003. In defence of the Volgian Stage. Stratigraphy and Geological Correlation 11: 585–593.
Zakharov, V.A. [Zaharov, V.A.] and Mesezhnikov, M.S. [Mesežnikov, M.S.] 1974. Volžskij ârus Pripolârnogo Urala. 216 pp. Nauka, Novosibirsk.
Zakharov, V.A., Baudin, F., Dzyuba, O.S., Daux, V., Zverev, K.V., and Renard, M. 2005. Isotopic and faunal record of high paleotemperatures in the Kimmeridgian of Subpolar Urals. Russian Geology and Geophysics 46: 3–20.
Zakharov, V.A. [Zaharov, V.A.], Mesezhnikov, M.S. [Mesežnikov, M.S.], Ronkina, Z.Z., Repin, Y.S., Chirva, S.A. [Čirva, S.A.], Shurygin, B.N. [Šurygin, B.N.], Basov, V.A., Vereninova, T.A., Kalacheva, E.D. [Kalačeva, E.D.], Kravets, V.S. [Kravec, V.S.], Meledina, S.V., Nalnyaeva, T.I. [Nalnâeva, T.I.], Paraketsov, K.V. [Parakecov, K.V.], Polubotko, I.V., and Sei I.I. 1983. Paleogeografiâ SSSR v ûrskom periode. 192 pp. Nauka, Novosibirsk.
Zieten, C.H. 1832. Die Versteinerungen Württembergs. 102 pp. Expeditum des Werkes unserer Zeit, Stuttgart.
Acta Palaeontol. Pol. 62 (4): 759–778, 2017
https://doi.org/10.4202/app.00387.2017