
Placoderms from the Lower Devonian “placoderm sandstone” of the Holy Cross Mountains, Poland with biostratigraphical and palaeobiogeographical implications
PIOTR SZREK and VINCENT DUPRET
Szrek, P. and Dupret, V. 2017. Placoderms from the Lower Devonian “placoderm sandstone” of the Holy Cross Mountains, Poland with biostratigraphical and palaeobiogeographical implications. Acta Palaeontologica Polonica 62 (4): 789–800.
The siliciclastic sequence of the Lower Devonian of the southern part of the Holy Cross Mountains in Poland is renown for abundant vertebrate fossils, including ostracoderm, sarcopterygian, acanthodian, chondrichthyan, and placoderm remains. Study of the vertebrate assemblage from the “placoderm sandstone” from Podłazie Hill in the Holy Cross Mountains reveals that the remains belong to Kujdanowiaspis sp. among other unspecified actinolepids and brachythoracids. The Polish actinolepid material is characterised by sizes bigger than those of the Podolian specimens; this may be related to geographic variation. Owing to the proximity between Podolia and Holy Cross Mountains we suggest that Kujdanowiaspis sp. from the Holy Cross Mountains may be a refugee of some species of Kujdanowiaspis from the Lochkovian–Pragian of Podolia and/or from Spain. Some anterolateral plates provisionally assigned to Arthrodira indet. probably represent a new genus due to the high overlapping surface for median dorsal and anteroventrolateral plates.
Key words: Placodermi, Arthrodira, Actinolepidoidei, palaeobiogeography, Devonian, Poland, Holy Cross Mountains.
Piotr Szrek [piotr.szrek@pgi.gov.pl], Polish Geological Institute-National Research Institute, ul. Rakowiecka 4, 00-975 Warszawa, Poland.
Vincent Dupret [vincent.dupret@anu.edu.au], Department of Applied Mathematics, Research School of Physics and Engineering, The Australian National University, Acton 2601 (Canberra) ACT, Australia.
Received 1 June 2017, accepted 8 August 2017, available online 30 October 2017.
Copyright © 2017 P. Szrek and V. Dupret. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The “placoderm sandstone” is a term introduced by Gürich (1896) for a mass-occurrence of the vertebrate remains considered by him to be placoderms. This term was later used by many geologists (e.g., Czarnocki 1919, 1936, 1950; Filonowicz 1968; Kowalczewski 1971; Łobanowski 1971; Tarnowska 1974, 1976; Szulczewski 1982, 1995). The occurrence of vertebrate remains was thought to be so characteristic that these rocks were colloquially named the “Placoderm Sandstone Formation”, and that this term was used to name the entire Lower Devonian strata especially in the Kielce region of the Holy Cross Mountains. This term should now be considered as an informal name for those portions of the section which are characterised by mass-accumulations of broken bones of storm origin (Szrek et al. 2014).
Placoderms have long been known to occur in the Holy Cross Mountains. Paradoxically, no placoderm from the “placoderm sandstone” has ever been formally described until now (but see Szrek 2003 and the Discussion herein). The presence of remains belonging to Arthrodira Actinolepidoidei and Brachythoraci is presented herein for the first time as a palaeontological study leading to biostratigraphical and palaeobiogeographical implications.
The occurrence of placoderms was ascertained by Czarnocki (1919, 1936) and mentioned, although not documented, by Tarnowska (e.g., 1981) and Szrek (2003). In contrast, an important literature focuses on agnathan remains owing to Tarlo (1957, 1961a, b, 1962, 1964, 1965) and Blieck (1980), while a single description of a sarcopterygian fish (Porolepis ex g. P. posnaniensis) was published by Kulczycki (1960). Tarlo (1957) noted that there were two bone-bearing localities in the Holy Cross Mountains: a vast amount of well-preserved vertebrate remains at Daleszyce (Podłazie Hill; Fig. 1), and at Dębska Wola (Czarnocki 1936: 181). Dębska Wola is not accessible for investigations at the moment because the locality is too vegetalised and requires large-scale excavations. Podłazie Hill near Daleszyce is now the only place in Poland where bone-bearing breccia is still available for investigations and is thus described below. Vertebrate remains were also recorded in boreholes (sections) by Tarnowska (1976). The most recent description (Szrek et al. 2015b) refers to a big homosteid arthrodire from Zagórze Formation of the Łysogóry region.
Institutional abbreviations.—Muz PGI, Geological Museum of the Polish Geological Institute-National Research Institute, Warsaw, Poland; MZ, Museum of Earth of the Polish Academy of Sciences, Warsaw, Poland.
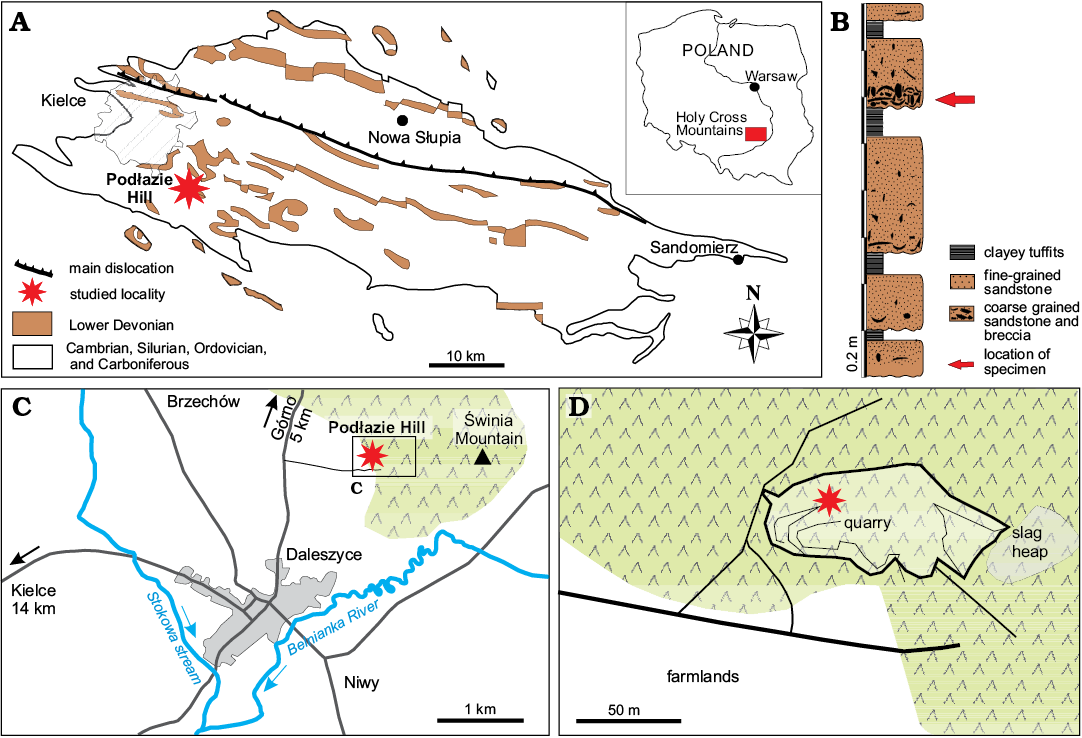
Fig. 1. A. Map of the Holy Cross Mountains showing distribution of Lower Dewonian deposits (modified from Kowalczewski 1971). B. Stratigraphic column at Podłazie Hill locality. C, D. Location and occurence of the Podłazie Hill outcrops.
Material and methods
All material is preserved as impressions (natural moulds). All elements consist of detached and disarticulated dermal bones of various sizes. Described elements are thus difficult to sort at the species level because it was not possible to compare their size and proportions, so they may represent one or several taxa.
Few specimens in the collections had rests of original bone preserved, which was systematically removed mechanically. All described and illustrated specimens were casted with silicone, whitened with Magnesium oxide and photographed. Selected specimens were drawn with a camera lucida. The studied material was collected mainly by Julian Kulczycki in the 1950s, and by Marek Dec, Grzegorz Niedźwiedzki, and PS during palaeontological fieldwork in the Holy Cross Mountains between 2004 and 2013. All specimens presented herein were found in the Podłazie Hill locality characterised in details by Szrek et al. (2014). The analysed material consists in 21 described specimens representing isolated parts of the disarticulated head- and trunk-shields. The fragmentary condition of the material precluded the authors to perform a cladistics analysis beyond the proposed systematic attribution.
Geological and stratigraphic setting
The Holy Cross Mountains is a geological region of central Poland divided into two units: a northern (Łysogóry) and a southern one (Kielce). Each region had separate geological histories and palaeogeographical positions during the Cambrian, Ordovician, Silurian, and part of the Devonian (Domeier and Torsvik 2014). The differences during the Devonian are illustrated by marine dominated conditions in the Łysogóry region and more continental conditions and only in some places marine in the Kielce region. Moreover the Devonian section in the Łysogóry region is almost complete whereas in the Kielce region it is characterised by the occurrence of many unconformities and gaps (see also Narkiewicz and Ziegler 2006: fig. 1 and Fig. 2 herein). The differences in the palaeontological record in the Lower Devonian deposits are also significant. In the Łysogóry region there is a rich assemblage of invertebrate fossils and ichnofossils (see Szulczewski 1995; Szulczewski and Porębski 2008) as well as vertebrate remains (Szrek et al. 2015b). In Kielce region there are no invertebrate remains apart from ichnofossils (Szrek et al. 2014, 2016), and the vertebrate remains are numerous and diversified (Tarlo 1957, 1961a, b, 1962, 1964, 1965; Kulczycki 1960; Blieck 1980).
Based on the vertebrate fauna, Czarnocki (1919, 1936) reported the same taxa for the Łysogóry and Kielce, notably Machaeracanthus polonicus. Later, Pawłowska (1954, 1961) discussed the stratigraphic sequences in both regions, accepting Czarnocki’s assessment of the different origins for northern and southern facies in the Devonian of the Holy Cross Mountains. Tarlo (1957) stated that Czarnocki (1936) disregarded vertebrates as potential useful age indicators. However, despite acknowledging that he had himself collected the same fauna in the northern and southern regions, Tarlo (1957) still considered that the correlation between the two regions was a matter of debate.
The Lower Devonian of the Kielce region represents a continental, marginal-marine and partly lacustrine sequence characterised by a progressing, stepwise and oscillatory transgression well recorded in the section (Kowalczewski 1971; Tarnowska 1976; Szrek et al. 2015a, b). The succession in the Kielce region consists of sandstone and mudstone beds, subordinately intercalated with siltstone, argilite, and conglomerate layers, and spans from the earliest Pragian to the Emsian based on tephrocorelation in tuffites and miospore analyses (Fijałkowska-Mader and Malec 2011; Fig. 2). The succession in the Kielce region was informally divided into the Haliszka and Winna formations, further subdivided into the lower sandstone, mudstone, and upper sandstone members (Tarnowska 1976, 1981). Tarlo (1957, 1964, 1965) described an essentially endemic psammosteid fauna (marine Lochkovian–Emsian Drepanaspis fauna) presumably older to the Middle Devonian continental psammosteid fauna assemblage of the Baltic area (see Mark-Kurik 2000). On stratigraphical and faunal grounds Tarlo (1957) regarded the Daleszyce fauna as entirely Emsian, possibly Lower to Middle.
The Lower Devonian of the Kielce region is now subdivided into four miospore zones (Verrucosisporites polygonalis–Dibolisporites wetteldorfensis, Emphanisporites annulatus–Brochotriletes bellatulus, Emphanisporites foveolatus–Verruciretusispora dubia, and Acinosporites apiculatus–Grandispora protea; Fijałkowska-Mader and Malec 2011) suggesting that the Haliszka and Winna formations range between the uppermost Pragian and the lowermost Eifelian. The age of analysed rocks cannot be precisely determined because of lack of precise stratigraphic markers. Based on correlations (Tarnowska 1976) and palynological analyses of boreholes it is most likely that the Lower Devonian of the Podłazie Hill represents the Winna Formation and is late Emsian in age (Fijałkowska-Mader and Malec 2011; Fig. 2).
The bone-bearing breccia of the Podłazie Hill mainly contains non-placoderm elements belonging to heterostracans, acanthodians, chondrichthyans, and osteichthyans which will be described in subsequent articles. Recent analyses reveal that the “placoderm sandstone” is actually tempestites containing less than 20% of placoderms remains out of the total vertebrate assemblage (Szrek et al. 2014: fig. 8). The mixed nature of this assemblage, together with abundance of sedimentological indicators such as currents and wave actions pledge for a storm synchrone of deposit (Szrek et al. 2014: figs. 4A, 7A). The marine influence is also consistent with the presence of ichnofossils (Szrek et al. 2014: 800–802). This diverse assemblage is composed of remains which were transported and deposited in shallow water conditions: during storms the skeletons were broken, blended together, size sorted, transported, and thrown in the direction of shallow water where they mixed with reworked sediment (Szrek et al. 2014).
Porolepis is very common in the “placoderm sandstone” (Kulczycki 1960; Ahlberg 1991; Clement 2001; Dec 2010; Szrek et al. 2014). It is an Early to early Middle Devonian genus mainly found in limestone rocks in the Arctic Canada (supposedly Lochkovian), Spitsbergen, East European Platform (Baltic area), Rhineland, Ural, Siberia, and eastern Australia. Such a vast distribution in shallow marine deposits is another evidence pledging for a shallow marine environment for the “placoderm sandstone” from Podłazie Hill.
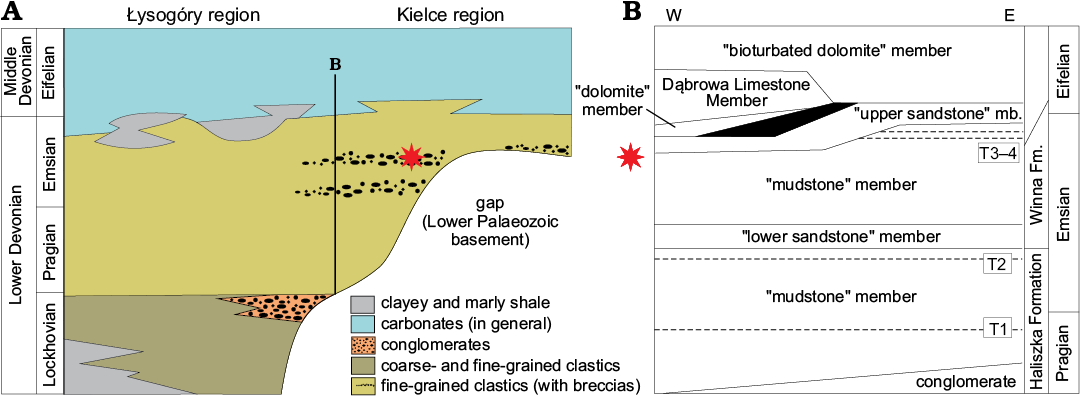
Fig. 2. A. Diagrammatic map the Holy Cross Mountains with indicated crossection from the base to the top of the Lower Devonian (modified after Szulczewski 1995). B. Lithostratigraphy of the upper Pragian to Eifelian in the Kielce Region (modified from Fijałkowska-Mader and Malec 2011). Star indicates Podłazie Hill locality; T1–4, pyroclastic horizons; black area in B is a stratigraphical gap.
Systematic palaeontology
Class Placodermi McCoy, 1848
Order Arthrodira Woodward, 1891
Suborder “Actinolepidoidei” Miles, 1973
Family Kujdanowiaspididae Berg, 1955
Genus Kujdanowiaspis Stensiö, 1942
Type species: Phlyctaenaspis buczacziensis Brotzen, 1934, Lower Devonian of Podolia (Ukraine).
Kujdanowiaspis sp.
Fig. 3A, C, E, F.
Material.—MZ-VIII/Vp-340 (Fig. 3A), Muz PGI 1733.II.37 (Fig. 3C), partially preserved head-shields; MZ-VIII/Vp-515 (Fig. 3E), Muz PGI 1733.II.390 (Fig. 3F), postorbital plates; Podłazie hill, Lower Devonian, Emsian.
Description.—Skull roof: The largest specimen (PGI 1733.II.37, Fig. 3C) shows very few details because of its high degree of weathering. It corresponds to the right part of a partial head shield. The surface is flat and shows no signs of secondary deformation. It is posteriorly concave just posteriorly to the central sensory line groove. Suture lines are not visible apart from the boundary between postorbital and paranuchal plates where it is emphasised by the linear pattern of ornamentation on a short distance. A faint supraorbital sensory groove can be distinguished. The central sensory line groove is well marked; it is 22 mm long and meets the infraorbital sensory groove at an angle of 63°. MZ-VIII/Vp-340 (Fig. 3A) corresponds to the left central and postorbital plates, belonging to a much larger individual: the central sensory line is 50 mm long, and meets infraorbital sensory groove at an angle of 61°. The surface is slightly depressed just behind the central sensory line groove. Ornamentation consists in randomly distributed tubercles whose diametre does not exceed 1.5 mm.
Postorbital plates: The postorbital plates are as wide as long and their outline is polygonal. The smallest specimen (Muz PGI 1733.II.390, Fig. 3F), which represents a right postorbital plate, is well preserved and ornamented with tubercles up to 1 mm in diametre arranged in circles. This allows to locate the radiation center, which is situated at the level of the trifurcation, between the central and the two rami of the infraorbital sensory line grooves. MZ-VIII/Vp-515 (Fig. 3E) corresponds to a left postorbital plate, and tubercles appear more worn than on the previous one. It also has a bigger angle between the infraorbital and central sensory grooves which equals 58°. The plate seems to be anteroposterioly elongated but no natural margin is preserved.
On the postorbital plates the common pattern of sensory grooves does not allow a precise identification of these specimens. They differ from the Kujdanowiaspis sp. described by Dupret (2010) by a different angle between the infraorbital and central sensory grooves which varies from 58° to 60° on Polish specimens and is 48–53° on Podolian specimens. All plates have broken margins and it is therefore impossible to determine relationships with other plates. Based on Muz PGI 1733.II.390 (Fig. 3F), which shows tubercules arranged in circular rows with the radiation centre more or less in the geometric centre of the specimen, it seems that the plate has an isometric shape.
Remarks.—The features visible on both head-shields do not allow a precise taxonomical identification because of the paucity of diagnostic sutures lines. However, the pattern of both central and partial infraorbital sensory grooves, together with the ornamentation are reminiscent of the Kujdanowiaspis and especially Kujdanowiaspis podolica (Brotzen, 1934) illustrated by Dupret (2010: fig. 13C). Polish specimens are however bigger than those illustrated by Dupret (2010: figs. 12, 13): the central sensory canal of K. podolica in Dupret (2010: fig. 13D) is almost 20 mm long whereas in MZ-VIII/Vp-340 (Fig. 3A) it is 50 mm long. In size only, described specimens would correspond to the Pragian actinolepid Eukaia elongata Mark-Kurik, 2013 described by Mark-Kurik (2013), to Erikaspis zychi (Stensiö, 1945) (see Dupret et al. 2007) or the Pragian Kujdanowiaspis buczacziensis (Brotzen, 1934) (see Dupret 2010), although the latter bears very minute and dense tubercles.
The occurence of such radiating tubercles from the geometric/growth centre of the plate fits the general shape of the postorbital plate in Kujdanowiaspis (Dupret 2010: fig. 12). Additionally, tubercles are more scattered than on Podolian specimens but are the same size as in K. podolica. Although precise measurement of the complete plates was not achievable, the preserved parts are sufficient to state that the studied material corresponds to bigger representatives than K. podolica. We thus attribute them to Kujdanowiaspis sp.
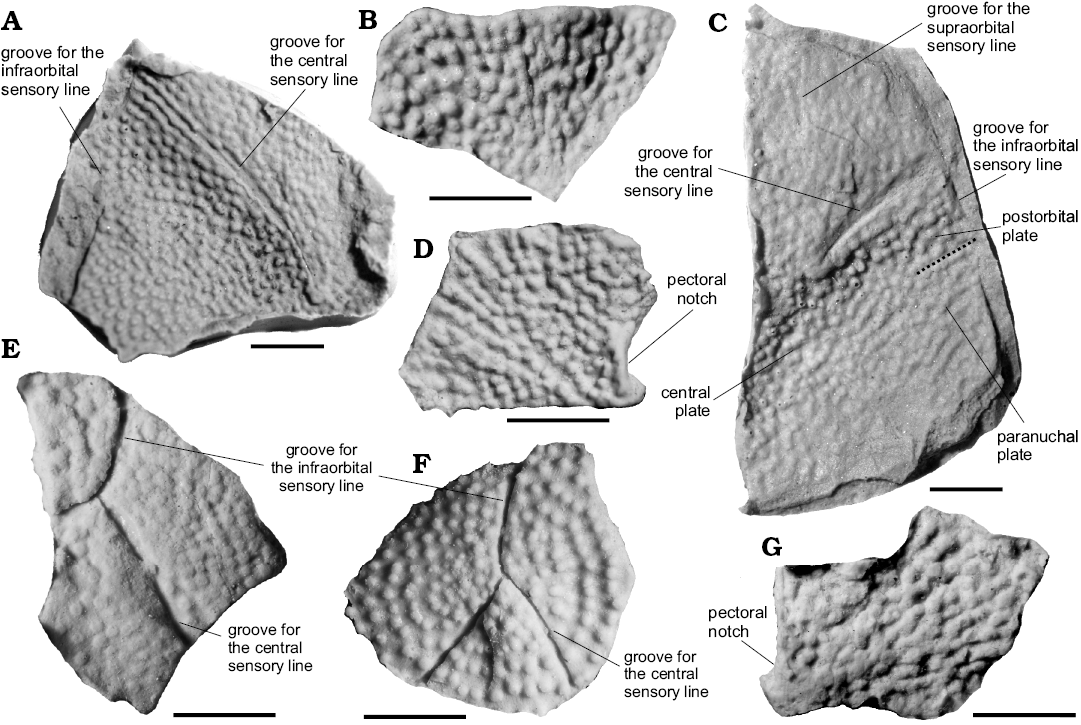
Fig. 3. Cranial and thoracic material of arthrodiran placoderm Kujdanowiaspis sp. (A, C, E, F) and Erikaspis sp. (B, D, F) from Lower Devonian (Emsian) of Podłazie Hill, Holy Cross Mountains, Poland. A. MZ-VIII/Vp-340, left fragmentary skull roof. B. MZ-VIII/Vp-480, posterodorsal fragment of a right anterior lateral plate. C. Muz PGI 1733.II.37, right fragmentary skull roof. D. Muz PGI 1733.II.183, left anterior lateral plate. E. MZ-VIII/Vp-515, left postorbital plate. F. Muz PGI 1733.II.390, right postorbital plate. G. Muz PGI 1733.II.383, subcomplete right anterior lateral plate. Scale bars 10 mm.
Genus Erikaspis Dupret, Goujet, and Mark-Kurik, 2007
Type species: Kujdanowiaspis zychi Stensiö, 1945 reasigned to Erikaspis zychi (Stensiö, 1945) since Dupret, Goujet, and Mark-Kurik, 2007, Podolia (Ukraine), Lower Devonian (Lochkovian).
Erikaspis sp.
Fig. 3B, D, F.
Material.—Muz PGI 1733.II.183 (Fig. 3D), Muz PGI 1733.II.383 (Fig. 3G), MZ-VIII/Vp-480 (Fig. 3B), anterior lateral plates; Podłazie hill, Lower Devonian, Emsian.
Description.—Anterior lateral plates: MZ-VIII/Vp-480 corresponds to the upper part of right plate (Fig. 3G), Muz PGI 1733.II.183, the completely preserved left plate (Fig. 3D), and Muz PGI 1733.II.383, the partially preserved right plate with its upper margin broken (Fig. 3G). The shape of the former two allows to determine the typical trapezoidal outline with straight slightly rounded and undulating boundary contacting the posterolateral and posterodorsolateral plates. The anteroventral margin is straight and narrow. All specimens appear to be similar in size. Tubercles (up to 1.5 mm in diametre) form oblique rows that radiate from the growth center to the angles of the plate which happens to be located close to the posteroventral corner of the plate (Fig. 3D, G). In the posterior part a pectoral notch is clearly visible as a shallow embayment.
Remarks.—Within anterior lateral plates the specimen MZ-VIII/Vp-480 (Fig. 3B) reflects some characteristics similar to those observed in Kujdanowiaspis podolica. These are: dorsal quadrant and postero-dorsal angle forming a rounded lobe. It resembles the right anterolateral plate of K. podolica illustrated by Dupret (2010: fig. 20B) and that of Aleosteus eganensis (Johnson et al. 2000: fig. 2J). Muz PGI 1733.II.183 and Muz PGI 1733.II.383 (Fig. 3D, G) are characterised by straighter margins, but they fit roughly to the morphology of the K. podolica plates (Dupret 2010: fig. 20A, B). The difference lays in the absence of crests and quadrant in specimen Muz PGI 1733.II.183 (Fig. 3D). Moreover, the ornamentation is extremelly similar to that observed in Erikaspis zychi, although the shape of the plate in this latter form is higher than long and resembles more that observed in Actinolepis magna (Dupret et al. 2007). It would be possible to consider that the square shape of the anterior lateral plate in Erikaspis is a juvenile character, and that the plate becomes higher as the animal get older. For this taxonomic determination, we put more weight on the ornamentation than on the shape of the plate.
Kujdanowiaspididae indet.
Fig. 4A–I.
Material.—MZ-VIII/Vp-485, 487, Muz PGI 1733.II.285, 340, 365, spinal plates (Fig. 4A–E); Muz PGI 1733.II.81, 384, 442, median dorsal plates (Fig. 4F–H); MZ-VIII/Vp-520, post median dorsal scute (Fig. 4I); Podłazie hill, Lower Devonian, Emsian.
Description.—Spinal plates: All spinal plates are incomplete. MZ-VIII/Vp-485 (Fig. 4B, D) and Muz PGI 1755.II.340 (Fig. 4E) are preserved as a cast of the dorsal side, as indicated by the groove for the insertion of the anterior lateral plate; Muz PGI 1733.II.340 is preserved as cast of the ventral side. Muz PGI 1733.II.365 and Muz PGI 1733.II.285 are preserved as three-dimensional casts of the distal part with broken tips. The largest plate (MZ-VIII/Vp-487, Fig. 4A) is 93 mm long and 25 mm wide in its proximal part. The plate is gently curved posteromesially. Its surface (dorsal and ventral) is ornamented with small densely distributed tubercles. The size of tubercles is uniform and does not exceed 1 mm in diametre. The mesial margin of the biggest specimens (MZ-VIII/Vp-485, 487) is ornamented with one row of anteriorly directed thorn-like spinelets which are bigger than the tubercles, and whose size increases posteriorly. Their amount varies from 6 (MZ-VIII/Vp-487, Fig. 4A) to 7 (Muz PGI 1733.II.485, Fig. 4B) or are absent (Muz PGI 1733.II.285, Fig. 4C; Muz PGI 1733.II.340, Fig. 4D), although this absence is most likely a preservational artefact. The numbers of those spinelets does not constitute a maximum because of the fragmentary state of preservation and the broken top of spinal plates.
Median dorsal plates: The median dorsal plate is almost as long as wide and has a rounded?-pentagonal outline. Specimens are similar in size: about 40 mm broad and 45 mm long. Width/length index is 88 (based on Muz PGI 1733.II.384 which is the most complete specimen). The latter possesses a low crest which is almost flattened anteriorly but more arched posteriorly. The entire surface is ornamented with coarse, generally irregularly arranged tubercles of a maximum of 1 mm in diametre. On the Muz PGI 1733.II.384 (Fig. 4G), tubercles in the posterior part are arranged in rows radiating from the center located approximately at the geometric center of the plate.
Posterior median dorsal scute: The plate is partially preserved and represents probably the first posterior median dorsal scute. Its anterior part is broken and missing. It is 24 mm wide, and was at least 20 mm long. In posterior view the triangular overlap area is visible (Fig. 4I2). The anterior area is characterised by pronounced raised area which makes a short ridge falling towards the front. The ornamentation consists of tubercles which vary from small (0.7–0.8 mm), densely arranged and coalesced in ridges in the center to bigger (1 mm) and more widely spaced on distal areas.
Remarks.—The spinal plates vary in sizes but all of them are bigger than the plates described and illustrated by Dupret (2010: 21–22, fig. 16A, B). The distance between the pectoral notch and the tip of the plate in Kujdanowiaspis podolica (Dupret 2010: fig. 16A, B) is 36 mm, whereas the length of the incomplete MZ-VIII/Vp-487 (Fig. 4A) reaches 68 mm. The described spinal plates differ from their homologues in K. podolica or Kujdanowiaspis buczaczensis by bigger sizes, more comparable with “Acanthaspis” armatus (Heintz 1929: fig. 4B). Thorn-like spinelets are prominent and similar to those described and illustrated by Dupret (2010: fig. 2F) and Dupret et al. (2011: fig. 3I). Features described above allow to identify MZ-VIII/Vp-485, MZ-VIII/Vp-487, and Muz PGI 1733.II.285, Muz PGI 1733.II.340 (Fig. 4A–E) as belonging to Kujdanowiaspis sp. All morphological features fit within the Kujdanowiaspis characters observed within the K. podolica, despite the much bigger sizes compared to that of the Podolian material. The absence of other articulated remains induces us to leave this identification in open nomenclature. Reasons of size differences may be related to regional, stratigraphic, ontogenetic and of course taxonomical/phylogenetic variation. Similar large actinolepids are, however, different in morphology and were described from the Pragian of the northwestern Siberian Platform (Eukaia elongata Mark-Kurik, 2013) and from the Givetian of the East European Platform (Actinolepis magna Mark-Kurik, 1973).
The overall shape of the median dorsal plate reminds the plate of K. podolica (Dupret 2010: fig. 17A, B) being as wide as long and more or less pentagonal, but the low posterior part and the coarse and dense tuberculation is more reminiscnent of Erikaspis zychi (Dupret et al. 2007). The plate is also low, reminding the Erikaspis disposition rather than the roof-shaped one in K. podolica or K. buczacziensis. The blunt posterior tip and the crest as long as the posterior half of the plate is also encoutered in Erikaspis zychi and Actinolepis magna. Moreover, median dorsal plate in K. podolica in the posterior part makes a prominent lobe. This character is absent in the studied Polish specimens and resembles the outline of median dorsal plate in Aleosteus eganensis (Johnson et al. 2000: fig. 2C) and Erikaspis zychi (Dupret et al. 2007). An other difference resides in the Polish specimens being relatively low in comparison with the roof-shaped median dorsal plate in K. podolica (Dupret 2010: fig. 17C) and Aleosteus eganensis (Johnson et al. 2000: fig. 2D). In this matter, the studied Polish specimens are very similar to Erikaspis zychi (Dupret et al. 2007: fig. 9A–C), which possesses the same ornamentation pattern, is low and possesses a low crest on the posterior half of its dorsal side. The only difference is that the specimen Muz. PGI 1733.II.384 (Fig. 3G) is anteriorly elongated when compared with Erikaspis zychi (Dupret et al. 2007: fig. 9A).
The posterior median dorsal scute plate does not show any exceptionally diagnostic features. Size, shape and ornamentation pattern are similar to those illustrated by Johnson et al. (2000: fig. 2A, B), Dupret (2010: fig. 24C–I), and Dupret et al. (2007: fig. 11 E–G). We think that the plate must belong with certainty to an actinolepidoideid placoderm, probably close or belonging to the Kujdanowiaspididae.
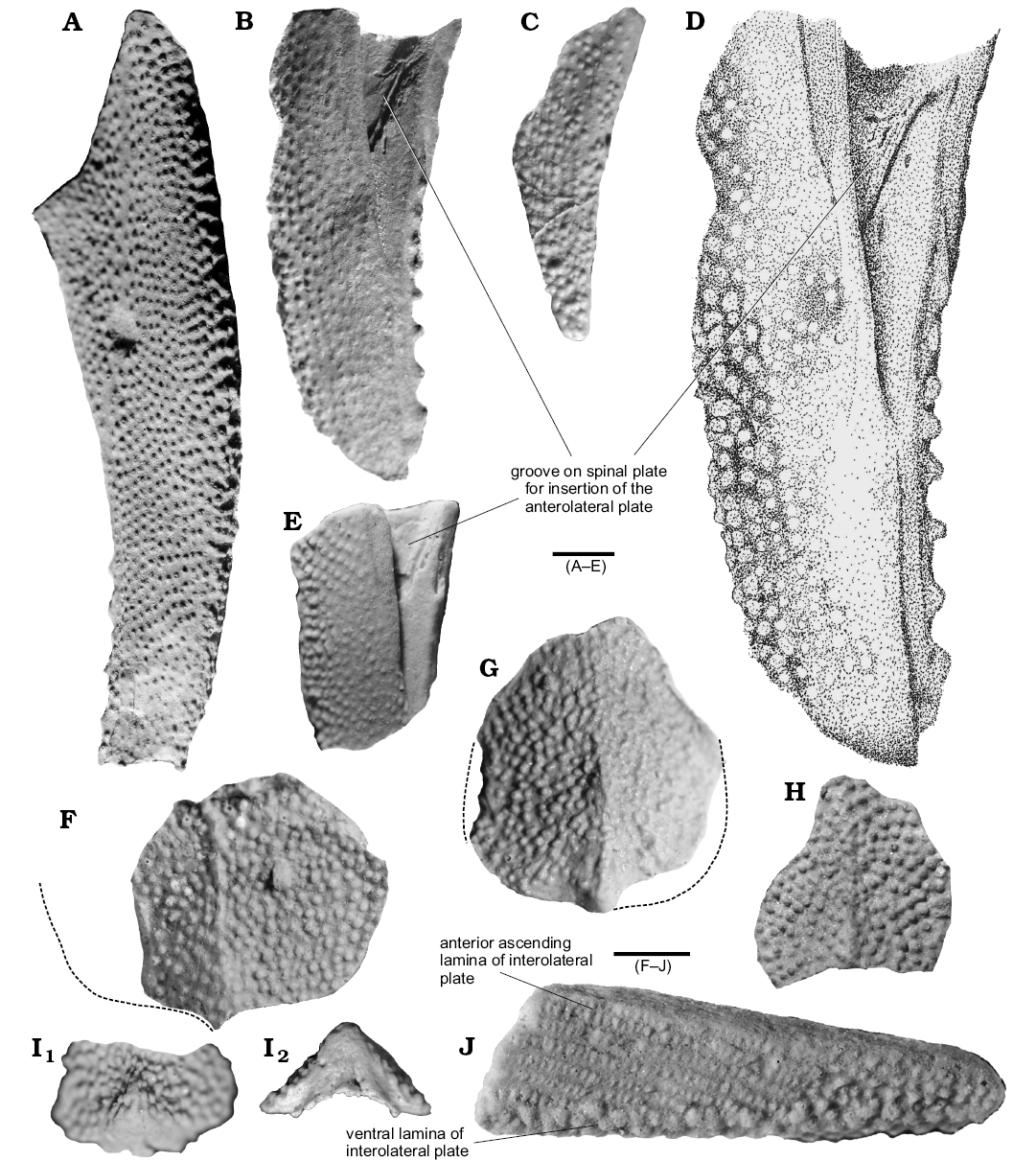
Fig. 4. Thoracic elements of arthrodiran placoderm Kujdanowiaspididae indet. (A–I) and Actinolepidoidei indet. (J) from Lower Devonian (Emsian) of Podłazie Hill, Holy Cross Mountains, Poland. A. MZ-VIII/Vp-487, left spinal plate. B. MZ-VIII/Vp-485, left spinal plate. C. Muz PGI 1733.II.285, right spinal plate. D. MZ-VIII/Vp-485, spinal plate (drawing) E. Muz PGI 1733.II.340, right spinal plate. F. Muz PGI 1733.II.442, posterior half of an incomplete median dorsal plate. G. Muz PGI 1733.II.384, subcomplete median dorsal plate. H. Muz PGI 1733.II.81, incomplete median dorsal plate. I. MZ-VIII/Vp-520, postmedian dorsal scute (tail cover) in dorsal (I1) and posterior (I2) views. J. Muz PGI 1733.II.174, right interolateral plate. Scale bars 10 mm.
Actinolepidoidei indet.
Fig. 4J.
Material.—Muz PGI 1733.II.174 interolateral plate (Fig. 4J); Podłazie hill, Lower Devonian, Emsian.
Description.—The sole specimen of the interolateral plate is incompletely preserved. It represents a right interolateral plate, of 34 mm long and 11 mm wide. On the specimen a special ornament typical of the ascending (internal) postbranchial lamina is well visible. The ornament is composed with seven rows of fine tubercules; the biggest is located on the anterior side. The ascending lamina widens laterally and reaches 7 mm in its highest preserved part. The ventral (external) lamina is ornamented with tubercles whose diametre can reach 1 mm, irregularly distributed in a high density.
Remarks.—The preserved part of the interolateral plate is too damaged to be diagnostic. Its elongation and similarity with the homologous plate in Dupret (2010: 38) and Dupret et al. (2011: 276) allows us to attribute this specimen to “actinolepids” (i.e., non Phyllolepida, non Wuttagoonaspididae, and non Antarctaspididae Actinolepidoidei).
Brachythoraci indet.
Fig. 5.
Material.—Muz PGI 1733.II.107 (Fig. 5B1), Muz PGI 1733.II.114 (Fig. 5A1), right and left, respectively, anterior dorsolateral plates; Muz PGI 1733.II.225 median dorsal plate (Fig. 5C); Podłazie hill, Lower Devonian, Emsian.
Description.—Anterior dorsolateral plates: The material is well preserved except for Muz PGI 1733.II.114 in which the anterior margin is only partially preserved (Fig. 5A1). The largest part of the plates corresponds to two overlapped areas (dorsally for the median dorsal plate and ventrally for the anterior lateral plate. Both specimens show a clearly reduced ornamented surface anteriorly. It is narrowest in the middle part of the plate, twice higher at the anterior margin and ten times higher at the posterior margin. The main lateral line groove is visible on the exposed area; it is straight on Muz PGI 1733.II.107 (Fig. 5B), and shows a distinct bifurcation on Muz PGI 1733.II.114 (Fig. 5A1). This bifurcation is low-angled (about 24°) and both lines beyond the bifurcation seem to have the same length so it is not obvious to determine which part is accessory. The preservation of both specimens as casts does not indicate the presence of either a glenoid condyle or a flattened sliding neck joint articulation. Ornamentation consists in scattered and irregularly arranged small tubercules measuring up to 0.7 mm in diametre.
Only a fragment of the posterior part of the median dorsal plate is preserved in visceral view. The natural margin is observable on the right side of the specimen only. It is slightly rounded and shows that the plate has an elongated outline, with a protruding posterior lobe. A keel is well visible in its middle part. It is 49 mm long in its maximum distance; the distance between the left margin and the keel is 20 mm so the calculated entire width must have reached 40 mm. The keel is broken at its base in the posterior part, where it is largest. The preserved part of the keel is 44 mm long.
Remarks.—The described specimens represent only a small portion of a trunk-shield and therefore only possess few distinctive features. The general morphology with extended overlap areas and narrow exposed area are not common characters in placoderms. Very high overlap area is observed in Taemasosteus novaustrocambricus White, 1952 (White 1978: fig. 102) but on our specimens it is even higher. The shape of overlapping surfaces visible on the Polish specimens (Fig. 5A, B) suggests that anterolateral plate had a blunt dorsal blade. The smooth posterior edge and the lack or proper overlapping area indicates the absence of a posterior dorsolateral plate. Bifurcation of the lateral line canal seems to be unique for this specimen, but this phenomenom was ascertained also in T. novaustrocambricus (White 1978: fig. 102). Besides, our sample is far from being statistically significant. White’s (1978: fig. 102) specimen differs from the Podłazie specimen by having an accessory twig obviously secondary than bifurcation of equal branches of sensory lines. Neither specimens belong to actinolepids nor phlyctaenidids because of the completely different shape of the anterior dorso-lateral plate and the very high overlap area for the median dorsal and anterolateral plates. This feature is reminiscent of what is observed in the family Homosteidae to which T. novaustrocambricus belongs (White 1978: 193), or to ptyctodontids. The exceptional characters of those plates suggest a close phylogenetic relationship with homosteids, but more specimens are required to ascertain this attribution. Homosteids were big placoderms with flattened bodies and armours, with probably benthic lifestyles, which occurred wide in Europe (Russian Arctic, Baltic area, Siberia, Spitsbergen, and Greenland) and Canada (Denison 1978). The stratigraphic range of this group covers almost the entire Early Devonian (Siegenian and Emsian) and early Middle Devonian (Givetian). The occurrence of the Emsian homosteids (Lelièvre 1988), in the Rhineland and Aragon as well as the material described in this paper and by Szrek et al. (2015), is in agreement with palaeogeographic reconstructions (Carls 2003) for the end of the Early Devonian. According to the position of the southern shelf of the Laurussian continent in the Early Devonian and localities of all species mentioned above, material from Poland represents a transitional position between Rhineland, Germany (Gross 1942), and the West Siberian Platform in Perevozinskoe, Russia (Krasnov and Kurik 1982; Szrek et al. 2015b; Fig. 6).
Based on the presence and shape of the keel and the general morphology of the available elements, the considered form belongs to arthrodires with a low crest and with a prominent ventral keel what is characteristic for many brachythoracids (Denison 1978).
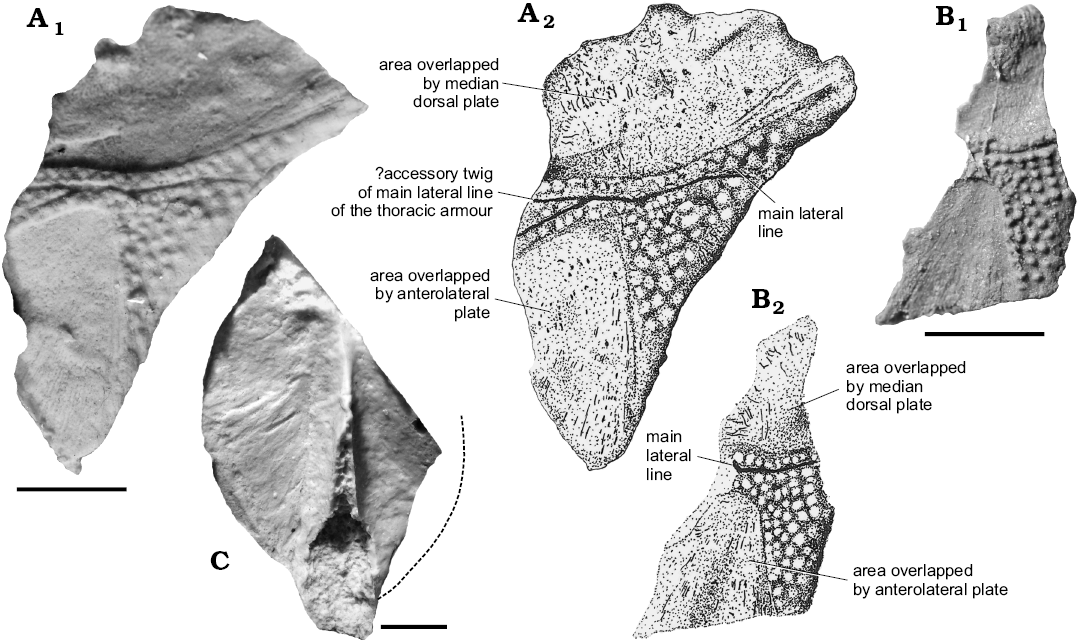
Fig. 5. Arthrodiran placoderm Brachythoraci indet. from Lower Devonian (Emsian) of Podłazie Hill, Holy Cross Mountains, Poland. A. Muz PGI 1733.II.114, print of subcomplete left anterior dorsolateral plate (A1, photograph; A2, “negative drawing”). B. Muz PGI 1733II.107, countreprint subcomplete right anterior dorsolateral plate (B1, photograph; B2, drawing). C. Muz PGI 1733.II.225, ventral side of an incomplete median dorsal plate. Scale bars 10 mm.
Discussion
All described elements are broken, disarticulated and mixed with other vertebrate remains. Due to their different sizes they certainly correspond to different individuals, and the possibility that they belong to different taxa must be taken into consideration. Disarticulation is the main reason why all elements were identified generally on an anatomical basis rather than a taxonomic one and in only two cases Kujdanowiaspis and Erikaspis, the authors agreed on a generic determination.
The material studied corresponds to the latest form of the Kujdanowiaspis and one of the biggest actinolepids. Those large forms are represented by two fragmentary head shields attributed to Actinolepidoidei and four spinal plates attributed to Kujdanowiaspis sp. It is possible that both head shields and spinal plates mentioned above represent the same form of Kujdanowiaspis sp., but it would require to be verified by more complete specimens in the future, which is very elusive considering the preservation nature of the specimens in tempestites. Despite its general bigger size, the Polish material resembles the Podolian material of Kujdanowiaspis. A reason for this difference could lay in geographic and/or stratigraphic variety, as well as biological matters such as different ontogenetic stages (from Podolian and Spanish forms), intraspecific variation or a different phylogenetic assignment within the Kujdanowiaspididae.
Kielce region in the Holy Cross Mountains was a part of the Małopolska block during Early Devonian. It belonged to the same southern margin of Baltica as Podolia (Domeier and Torsvik 2014). Close spatial relationships and connections between Podolia and the Holy Cross Mountains during the Lochkovian and early Pragian are evidenced by the co-occurrence of the “Dittonian” agnathan fauna (Lochkovian–early Pragian; Pawłowska 1954, 1961; Kulczycki 1960). This fauna occurs in classical Old Red Sandstone facies which is very common in Podolia but does not occur in the higher stages in the Kielce region (late Pragian and Emsian). Thus, the most important difference is the age of each assemblage (Podolia and Holy Cross Mountains), the Spanish one, although in a calcareous facies, being contemporaneous of the lower part of the Podolian one (Lochkovian; Dupret et al. 2011). In Podolia most of the specimens of Kujdanowiaspis described by Stensiö (1944), Dupret et al. (2007) and Dupret (2010) are late Lochkovian and early Pragian in age (Blieck and Cloutier 2000; Dupret and Blieck 2009). The studied Holy Cross Mountains series is dated as upper Emsian (Fijałkowska-Mader and Malec 2011; Szrek et al. 2014). The Polish assemblage is therefore the first occurrence of Kujdanowiaspis outside of Podolia but also 10–15 million years younger. According to Early Devonian palaeogeography (Domeier and Torsvik 2014), the occurrence of Kujdanowiaspis representatives in the Kielce region of the Holy Cross Mountains may suggest its southwards dispersal (Fig. 6). It also illustrates a wider geographical distribution and stratigraphic surviving span of the species until the end of the lower Devonian than previously thought. The Holy Cross Moutains may have acted as a refuge area for this Kujdanowiaspis fauna.
Apart from actinolepids, other placoderm remains are very interesting. The description of anterior dorsolateral plates (Fig. 5A, B) characterised by very large overlapped areas needs to be supplemented by further comparative material to potentially identify a new form of placoderm.
According to Szrek et al. (2014), the assemblages recorded at Podłazie Hill contain forms reworked from the open shelf (acanthodians, chondrichthyans) and mixed with those that lived in a marginal-marine environment (placoderms and sarcopterygians). Most if not all actinolepids were bottom-dwellers, according to their ventrally flattened body morphology, and their well-developed lateral spines (Denison 1978; Janvier 1996), added to a poor estimated swimmability. The mixed character of the bone assemblages leads to the conclusion that the life habitat of placoderms must have been located away from the place of final burial of their carcasses. This was resulted from a high-energy environment, such as a stormy episode, during which the redeposition and final burial of faunal remains of different ecological niche origins occured.
We also supplement and correct Szrek’s (2003) considerations, who identified presumable antiarch and arthrodire remains. The current study did not confirm those identifications. Three specimens illustrated in that paper (Szrek 2003: figs. 2B, C, 3B) were rather misinterpreted and should now be referred generally to indeterminable vertebrate (placoderm or sarcopterygian) remains.
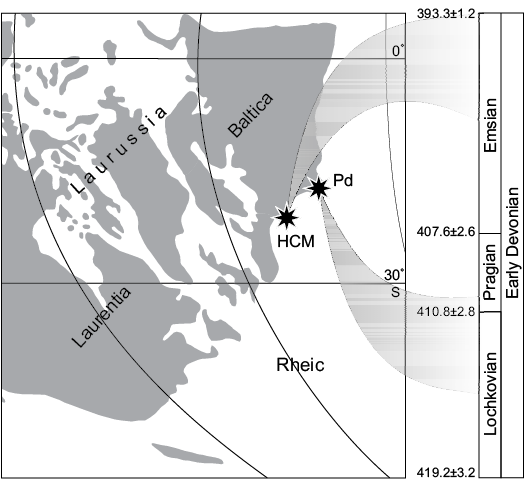
Fig. 6. Palaeogeographic reconstruction during the Early Devonian (after Domeier and Torsvik 2014) and stratigraphic occurences of the “Polish Placoderm Sandstone” and the Ukrainian “Kujdanowiaspis series” in Podolia. Abbreviations: HCM, Holy Cross Mountains; Pd, Podolia.
Conclusions
The material presented herein is the first placoderm assemblage from the Lower Devonian “placoderm sandstone” reported and described exhaustively. The assemblage comprises the youngest representatives of the Kujdanowiaspis fauna, as well as unspecified actinolepids and unidentified brachythoracid arthrodires. This study also provides a supplement to the “placoderm sandstone” assemblage composition: placoderms now implement already studied agnathans, acanthodians, and sarcopterygians.
Actinolepids remains differ from those described from the adjacent area of Podolia. This is probably due to their higher stratigraphic position, but other geological or biological reasons should not be omitted. The occurrence of the genus of Kujdanowiaspis points out close relations between Podolia and Holy Cross Mountains during the Early Devonian. Therefore, the Kujdanowiaspis sp. from the upper Emsian of the Holy Cross Mountains maybe considered as a refugee of some species of Kujdanowiaspis from the Lochkovian–Pragian of Podolia and perhaps of Spain. Although this is not the first occurrence of Kujdanowiaspis out of Podolia and the most western (according to modern geography) and southern (according to Early Devonian palaeogeography), it represents, however, the youngest occurence of the genus, and invites us to consider the Holy Cross Mountain as a potential refuge for this iconic Early Devonian Kujdanowiaspis assemblage. Current works on the remains from the “placoderm sandstone”, especially the unindentified brachythoracids, points out the necessity of further studies and research in other places in the Holy Cross Mountains. Our next spot of interest remains in a 2 m thick bone-bearing breccia was reported in the past (Czarnocki 1919, 1936), which still needs to be studied in details.
Acknowledgements
We are grateful to the former Mayor of Daleszyce, Wojciech Furmanek for his support during field-work. We thank also Marta Hodbod and Maria Andruszkiewicz-Gorzelak (both Polish Geological Institute-National Research Institute, Warsaw, Poland), Przemysław Gorzelak and Marek Dec (both Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland), Daniel Tyborowski (Museum and Institute of Zoology, Polish Academy of Sciences, Warsaw, Poland), and Aleksandra Gronkowska (Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland) who helped us during the fieldwork. The Podłazie locality was examined during the fieldwork in 2011–2012 financed by the Ministry of Science and Higher Education (grant IP2010 041470 to PS).
References
Ahlberg, P.E. 1991. A re-examination of sarcopterygian interrelationships, with special reference to the Porolepiformes. Zoological Journal of the Linnean Society 103: 241–287. Crossref
Berg, L.S. 1955. Sistemariboobraznik i rib, ninie jivooŝih i iskopaiemyh. 2nd Edition. 286 pp. Moskva, Leningrad.
Blieck, A. 1980. Le genre Rhinopteraspis Jaekel (Vertébrés, Hétérostracés) du Dévonien inférieur: systématique, morphologie, répartition. Bulletin du Muséum national d’Histoire naturelle, 4ème série, 2, section C 1: 25–47.
Blieck, A. and Cloutier, R. 2000. Biostratigraphical correlations of Early Devonian vertebrate assemblages of the Old Red Sandstone Continent. In: A. Blieck and S. Turner (eds.), Palaeozoic Vertebrate Biochronology and Global Marine/Non-Marine Correlation. Final Report of IGCP 328 (1991–1996). Courier Forschunginstitut Senckenberg 223: 223–269.
Carls, P. 2003. Tornquist’s Sea and the Rheic Ocean are Illusive. Courrier Forschunginstitut Senckenberg 242: 89–109.
Clément, G. 2001. Evidence for lack of choanae in the Porolepiformes. Journal of Vertebrate Paleontology 21: 795–802. Crossref
Czarnocki, J. 1919. Stratygrafia i tektonika Gór Świętokrzyskich. Prace Towarzystwa Naukowego Warszawskiego 28: 1–172.
Czarnocki, J. 1936. Przegląd stratygrafii i paleogeografii dewonu dolnego Gór Świętokrzyskich. Sprawozdania Państwowego Instytutu Geologicznego 7: 129–200.
Czarnocki J. 1950. Geology of the Łysa Góra region (Holy Cross Mountains) in the connection with the problem of iron ores at Rudki [in Polish with English summary]. Prace Państwowego Instytutu Geologicznego 18: 1–308.
Dec, M. 2010. Osteichthyans from the Lower Devonian placoderm sandstones of the Holy Cross Mountains. 49 pp. Unpublished M.Sc. Thesis, University of Warsaw, Warsaw.
Denison R.H. 1978. Placodermi. In: H.-P. Schultze (ed.), Handbook of Paleoichthyology 2. 128 pp. Gustav Fischer Verlag, Stuttgart.
Domeier, M. and Torsvik, T.H. 2014. Plate tectonic in the late Paleozoic. Geoscience Frontiers 5: 303–350. Crossref
Dupret, V. 2010. Revision of the genus Kujdanowiaspis Stensiö, 1942 (Placodermi, Arthrodira, “Actinolepida”) from the Lower Devonian of Podolia (Ukraine). Geodiversitas 32: 5–63. Crossref
Dupret, V. and Blieck, A. 2009. The Lochkovian–Pragian boundary in Podolia (Lower Devonian, Ukraine) based upon placoderm vertebrates. Comptes Rendus Geoscience (Paris) 341: 63–70. Crossref
Dupret, V., Goujet, D., and Mark-Kurik, E. 2007. A new genus of placoderm (Arthrodira: “Actinolepida”) from the Lower Devonian of Podolia (Ukraine). Journal of Vertebrate Paleontology 27: 266–284. Crossref
Dupret, V., Carls, P., Martínez-Pérez, C., and Botella, H. 2011. First Perigondwanan record of actinolepids (Vertebrata: Placodermi: Arthrodira) from the Lochkovian (Early Devonian) of Spain and its palaeobiogeographic significance. Palaeogeography, Palaeoclimatology, Palaeoecology 310: 273–282. Crossef
Fijałkowska-Mader, A. and Malec, J. 2011. Biostratigraphy of the Emsian to Eifelian in the Holy Cross Mountains (Poland). Geological Quarterly 55: 109–138.
Filonowicz, P. 1968. Objaśnienia do szczegółowej mapy geologicznej Polski. Arkusz Kielce. 100 pp. Wydawnictwa Geologiczne, Warszawa.
Gross, W. 1942. Die Fischfaunen des baltischen Devons und ihre biostratigraphische Bedeutung. Naturforscher-Vereins zu Riga, Korrespondenzblatt 64: 373–436.
Gürich, G. 1896. Das Paläozoicum im Polnische Mittelgebirge. Verhandlungen der Russichen-Kaiserlichen Mineralogischen Gesellscheft zu St-Petersburg 2 (32): 1–539.
Heintz, A. 1929. Die downtonischen und devonischen Vertebraten von Spitzbergen II. Acanthaspida. Skrifeter om Svalbard og Ishavet 22: 1–81.
Janvier, P. 1996. Early vertebrates. Oxford Monographs on Geology and Geophysics 33: 1–393.
Johnson, H.G., Elliot, D.K., and Wittke, J.H. 2000. A new actinolepid arthrodire (Class Placodermi) from the Lower Devonian Sevy Dolomite, East-Central Nevada. Zoological Journal of the Linnean Society 129: 241–266. Crossref
Kowalczewski, Z. 1971. Main geological problems of the Lower Devonian in the Świętokrzyskie Mts. Geological Quarterly 15: 263–283.
Krasnov, V.N. and Kurik, E. 1982. First find of fossil fish in limestones of the Tashtyp Formation of the South-Minusinsk Depression [in Russian]. In: O.V. Ûferev (ed.), Pervaâ nahodka iskopaemoj ryby v izvestnâkah taštypskoj svity Južno-Minusinskoj vpadiny. Stratigrafiâ i paleontologiâ devona i karbona. Transactions of the Institute of Geology and Geophysics, Academy of Sciences of the USSR, Siberian Branch 483: 47–52.
Kulczycki, J. 1960. Porolepis (Crossopterygii) from the Lower Devonian of the Holy Cross Mountains. Acta Palaeontologica Polonica 5: 65–103.
Lelièvre, H. 1988. New material of Antineosteus lehmani Leliévre, 1984 (Placodermi, Brachythoraci) and of acanthodians from the Lower Devonian (Emsian) of Algeria. Bulletin du Muséum national d’histoire naturelle, 4ème série, section C 10: 287–302.
Łobanowski, H. 1971. The Lower Devonian in the western part of the Klonów Belt (Holy Cross Mountains), Part I. Upper Emsian. Acta Geologica Polonica 21: 629–687.
Mark-Kurik, E. 1973. Actinolepis (Arthrodira) from the Middle Devonian of Estonia. Palaeontographica A 143: 89–108.
Mark-Kurik, E. 2000. The Middle Devonian fishes of the Baltic States (Estonia, Latvia) and Belarus. Courier Forschung-Institute Senckenberg 223: 309–324.
Mark-Kurik, E. 2013. A new Lower Devonian arthrodire (Placodermi) from the NW Siberian Platform. Estonian Journal of Earth Sciences 62: 131–138. Crossref
McCoy, F. 1848. On some new fossil fish of the Carboniferous period. Annals and Magazine of Natural History 2: 1–10. Crossref
Miles, R.S. 1973. An actinolepid arthrodire from the Lower Devonian Peel Sound Formation, Prince of Wales Island. Palaeontographica A 143: 109–118.
Narkiewicz, M. and Ziegler, P.A. 2006 Poland: junction of the main geological provinces of Europe. Geological Quarterly, Special Anniversary Volume 50: 1–210.
Pawłowska, K. 1954 Nowe dane o paleozoiku na północ od Sandomierza – komunikat wstępny. Przegląd Geologiczny 11: 461–462.
Pawłowska, K. 1961. W sprawie wieku warstw bostowskich w związku z problemem granicy pomiędzy sylurem i dewonem w Górach Świętokrzyskich. Geological Quarterly 5: 526–537.
Stensiö, E. 1942. On the snout of arthrodires. Kungliga Svenska Vetenskaps Akademiens Handlingar 20: 1–32.
Stensiö, E. 1944. Contributions to the knowledge of the vertebrate fauna of the Silurian and Devonian of Podolia. II. Note on two arthrodires from the Downtonian of Podolia. Arkiv for Zoologi 35: 1–83.
Szrek, P. 2003. Nowe dane na temat fauny kręgowców w „piaskowcach plakodermowych” z rejonu Daleszyc (Góry Świętokrzyskie). Przegląd Geologiczny 51: 409–411.
Szrek, P., Niedźwiedzki, G., and Dec, M. 2014. Storm origin of bone-bearing beds in the Lower Devonian placoderm sandstone from Podłazie Hill (Holy Cross Mountains, central Poland). Geological Quarterly 58: 795–806. Crossref
Szrek, P., Salwa, S., and Niedźwiedzki, G. 2015a. Plant-root system in the Lower Devonian of Poland. Estonian Journal of Earth Sciences 64: 95–98. Crossref
Szrek, P., Dec, M., and Niedźwiedzki, G. 2015b. The first placoderm fish from the Lower Devonian of Poland. Journal of Vertebrate Paleontology 35 (3): e930471.
Szrek, P., Salwa, S., Niedźwiedzki, G., Dec, M., Ahlberg, P.E., and Uchman, A. 2016. A glimpse of a fish face—an exceptional fish feeding trace fossil from the Lower Devonian of the Holy Cross Mountains, Poland. Palaeogeography, Palaeoclimatology, Palaeoecology 454: 113–124.
Szulczewski, M. 1982. Podstawowe problemy stratygrafii dewonu w Polsce. Biuletyn Geologiczny 25: 267–297.
Szulczewski, M. 1995. Depositional evolution of the Holy Cross Mountains in the Devonian and Carboniferous—a review. Geological Quarterly 39: 471–488.
Szulczewski, M. and Porębski, S. 2008. Stop 1—Bukowa Góra, Lower Devonian. In: G. Pieńkowski and A. Uchman (ed.), Ichnological Sites of Poland. The Holy Cross Mountains and the Carpathian Flysch. The Second International Congress on Ichnology. Cracow, Poland, August 29–September 8, 2008. The Pre-Congress and Post-Congress Field Trip Guidebook, 18–37. Polish Geological Institute, Warszawa.
Tarlo, L.B. 1957. A preliminary note on new ostracoderms from the Lower Devonian (Emsian) of central Poland. Acta Palaeontologica Polonica 2: 225–233.
Tarlo, L.B. 1961a. Psammosteids from the Middle and Upper Devonian of Scotland. The Quarterly Journal of the Geological Society of London 117: 367–402. Crossref
Tarlo, L.B. 1961b. Rhinopteraspis cornubica (McCoy), with notes on the clssification and evolution of the pteraspids. Acta Palaeontologica Polonica 6: 367–402.
Tarlo, L.B. 1962. The classification and evolution of the Heterostraci. Acta Palaeontologica Polonica 7: 249–286.
Tarlo, L.B. 1964. Psammosteiformes (Agnatha)—a review with descriptions of new material from the Lower Devonian of Poland. I—General part. Palaeontologia Polonica 13: 1–135.
Tarlo, L.B. 1965. Psammosteiformes (Agnatha)—a review with descriptions of new material from the Lower Devonian of Poland. II—Systematic part. Palaeontologia Polonica 15: 1–168.
Tarnowska, M. 1974. Poszukiwanie złóż piaskowców dolnodewońskich w rejonie Iwanisk. 64 pp. Archiwum PIG w Kielcach, Kielce.
Tarnowska, M. 1976. Korelacja litologiczna dewonu dolnego we wschodniej części Gór Świętokrzyskich. Biuletyn Instytutu Geologicznego 296: 75–117.
Tarnowska, M. 1981. Dewon dolny w centralnej części Gór Świętokrzyskich. In: H. Żakowa (ed.), Przewodnik 53 Zjazdu Polskiego Towarzystwa Geologicznego, Kielce, 57–67. Wydawnictwa Geologiczne, Warszawa.
White, E.I. 1952. Australian arthrodires. Bulletin of the British Museum (Natural History) (Geology) 1: 249–304.
White, E.I. 1978. The large arthrodiran fishes from the area of the Burrinjuck Dam, N.S.W. Transactions of the Zoological Society of London 34: 149–262. Crossref
Woodward, A.S. 1891. Catalogue of the Fossil Fishes in the British Museum (Natural History). Part II. 567 pp. British Museum (Natural History), London.
Acta Palaeontol. Pol. 62 (4): 789–800, 2017
https://doi.org/10.4202/app.00395.2017