

Highly derived eutherian mammals from the earliest Cretaceous of southern Britain
STEVEN C. SWEETMAN, GRANT SMITH, and DAVID M. MARTILL
Sweetman, S.C., Smith, G., and Martill, D.M. 2017. Highly derived eutherian mammals from the earliest Cretaceous of southern Britain. Acta Palaeontologica Polonica 62 (4): 657–665.
Eutherian mammals (Placentalia and all mammals phylogenetically closer to placentals than to marsupials) comprise the vast majority of extant Mammalia. Among these there is a phenomenal range of forms and sizes, but the origins of crown group placentals are obscure. They lie within the generally tiny mammals of the Mesozoic, represented for the most part by isolated teeth and jaws, and there is strongly conflicting evidence from phenomic and molecular data as to the date of origin of both Eutheria and Placentalia. The oldest purported eutherians are Juramaia from the Upper Jurassic of China, and Eomaia and Acristatherium from the Lower Cretaceous, also of China. Based on dental characters and analyses of other morphological and molecular data, doubt has recently been cast on the eutherian affinities of the Chinese taxa and consequently on the date of emergence of Eutheria. Until now, the only tribosphenic mammal recorded from the earliest Cretaceous (Berriasian) Purbeck Group of Britain was the stem tribosphenidan Tribactonodon. Here we document two new tribosphenic mammals from the Purbeck Group, Durlstotherium gen. nov. and Durlstodon gen. nov., showing highly derived eutherian molar characters that support the early emergence of this clade, prior to the Cretaceous.
Key words: Mammalia, Eutheria, dentition, Early Cretaceous, Purbeck Group, Britain, UK.
Steven C. Sweetman [steven.sweetman@port.ac.uk], Grant Smith [grant.smith1@myport.ac.uk], and David M. Martill [david.martill@port.ac.uk], University of Portsmouth, School of Earth and Environmental Sciences, Burnaby Building, Burnaby Road, Portsmouth, PO1 3QL, UK.
Received 4 July 2017, accepted 5 October 2017, available online 7 November 2017.
Copyright © 2017 S.C. Sweetman et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Purbeck Group “Mammal Bed”, bed number DB 83 (Clements 1993), exposed in Durlston Bay on the east coast of Dorset, southern United Kingdom (Figs. 1, 2) was, until the latter part of the 20th century, one of very few horizons world-wide to yield Mesozoic mammals. It was extensively sampled by Samuel Beckles in the mid-19th century at which time many iconic specimens were recovered (Owen 1854; Kielan-Jaworowska et al. 2004). These represented new taxa and also provided invaluable morphological information concerning the jaws and, in one case, the skull of Early Cretaceous mammals (Owen 1854, 1871; Kielan-Jaworowska et al. 2004). Following cessation of Beckles’ quarrying activities very few mammal specimens were reported from this horizon and the quarry is now obscured and inaccessible. Material collected in the 19th Century all represented non-therian clades, e.g., eutriconodontans, multituberculates, spalacotheroids, docodontans, and dryolestidans (Owen 1854; Kielan-Jaworowska et al. 2004). It was not until the latter part of the 20th century that a tooth crown and crown fragments representing a tribosphenic mammal, the stem tribosphenidan aegialodontid Tribactonodon bonfieldi Sigogneau-Russell, Hooker, and Ensom, 2001, were recovered from a horizon about 15 m higher in the succession. Recently, residue obtained from screen washing small samples taken from the “Mammal Bed” exposed at beach level in Durlston Bay have yielded isolated mammal teeth including two highly derived tribosphenic upper molars reported here. Dental morphology demonstrates that these both represent eutherian mammals.
Institutional abbreviations.—NHMUK PV M, The Natural History Museum, London, UK.
Nomenclatural acts.—The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be re-solved and the associated information viewed through any standard web browser by appending the LSID to the pre-fix “http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:12D080E5-88B1-4898-8D1B-96B44D247E77. The electronic edition of this work was published in a journal with an eISSN 1732-2421, and has been archived and is available from the following digital repositories: Internet Archive (https://archive.org/) and http://www.app.pan.pl/article/item/app004082017.html.
Geological and palaeonvironmental setting
The Purbeck Group comprises a distinctive and heterolithic sequence of evaporites, thinly bedded sandstone and shelly limestone, interbedded with calcareous marl and shale. Palaeontological and sedimentological evidence shows that these were deposited in fluctuating freshwater, brackish, hypersaline and quasi-marine environments (Batten 2002). The fauna and flora is also indicative of terrestrial, lacustrine and variably saline, lagoonal associations (Batten 2002). The boundary with the underlying, massively bedded marine limestone of the Upper Jurassic Portland Group is easily recognised in exposures in the southern part of Durlston Bay. However, at Peveril Point at the northern end of the bay (Fig. 1) the junction between the Purbeck Group and the overlying clastic succession comprising the Wealden Group is obscured. The age of the Purbeck Group has been the subject of much debate and until the latter part of the twentieth century beds below the Cinder Member (commencing with DB 110, Fig. 2) were thought to be uppermost Jurassic. More recently the entire Purbeck Group has been considered to be Berriasian. However, the exact position of the Jurassic–Cretaceous boundary remains uncertain (Allen and Wimbledon 1991; Ogg et al. 1994; Hunt 2004). Evidence suggests that the climate during deposition of the Purbeck Group was Mediterranean-like. However, facies changes and palaeontological data indicate that it became wetter towards the end of the Berriasian (Batten 2002; Sladden and Batten 1984; Allen 1998; Radley 2002). The Marly Freshwater Member, in which the “Mammal Bed” is situated, primarily comprises calcareous mudstone with no evidence of evaporites. In contrast, the Soft Cockle Member below contains micrites with halite pseudomorphs and thicker evaporite deposits. The former is considered to have been deposited in a shallow, freshwater lake. The overlying Cherty Freshwater Member is dominated by limestone with subordinate calcareous mudstone and shale. Palaeontological data suggests deposition in environments with fluctuating salinity (freshwater to slightly saline) with higher salinity perhaps reflecting periods of drought (Allen 1998), and mudcracks and dinosaur footprints periods of complete desiccation.
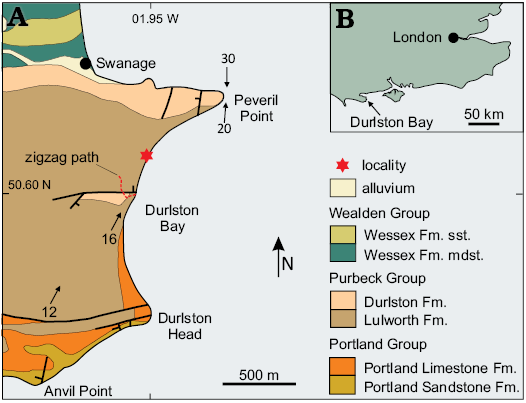
Fig. 1. A. Outline geological map of Durlston Bay and the southern part of Swanage Bay, east Dorset, United Kingdom showing the locality. Arrows indicate dip. B. Location map. Abbreviations: Fm., Formation; mdst., mudstone; sst., sandstone.

Fig. 2. Stratigraphic log of part of the Purbeck Group exposed in the northern part of Durlston Bay showing the horizons from which mammal remains have been recovered, and the horizon from which the new specimens described here were obtained. Redrawn from Clements (1993) using the author’s “erosional profile” but with additional bed names. Detailed comments on lithologies and palaeontology are contained in the above publication in which all bed numbers bear the prefix DB.
Material and methods
During the summer of 2015 bulk samples totalling approximately 55 kg were taken from a beach-level exposure of the “Mammal Bed” at British National Grid Reference SZ 03673 78377. At this location members of the public have free access to the foreshore and permission to collect fossils is not required under UK law. The samples were thoroughly air dried in the laboratory before being processed in small batches. Processing involved disaggregation in fresh water and hydrogen peroxide. Samples were then hand sieved in water using a sieve with a 0.25 mm mesh. Dried residues were divided into size fractions using a nest of sieves and those with a particle size 0.25–2 mm were picked under a binocular microscope using a gridded sorting tray. Mammal teeth were recovered from the 0.5–2 mm fraction, that with a particle size 0.25–0.5 mm being largely devoid of determinate vertebrate remains. Two determinable mammal teeth, as described below, were recovered from one of the samples and a very small incisor, a small double-rooted canine and the principal cusp of a substantial premolariform crown were recovered from others. Images of specimens, which were gold-palladium coated to avoid charging, were obtained using a Zeiss EVO MA10 electron microscope with an acceleration voltage of 10 kV. Normal light photographs were also obtained using a Leica EZ4W AiarLab microscope, which incorporates a 5 megapixel CMOS digital camera, and employing CombineZP image stacking software. Measurements were taken digitally using KLONK Image Measurement, software version 15.1.2.1 (available from http://klonk-image-measurement.en.softonic.com/). In descriptions below tooth terminology follows Kielan-Jaworowska et al. (2004) and wear facet numbers are based on Crompton (1971), but the homology of some facets is uncertain. Figured specimens are accessioned in the collections of the Natural History Museum, London under accession numbers NHMUK PV M 99991 and NHMUK PV M 99992.
In view of the limited number of character states that can be scored for these posterior-most teeth no attempt at phylogenetic analysis has been made. Furthermore, there are no coeval eutherians in which M3 is preserved. M3 is known for Juramaia from the Oxfordian of China but its tooth structure is plesiomorphic having a low protocone and lingually placed conules. M3 is also known for Acristatherium and has been reported for Eomaia from the Barremian of the same region. However, in the case of Eomaia, M3 is not preserved (see discussion in Averianov et al. 2010). The next temporarily closest taxa are from the Aptian–Albian of Asia and North America and are therefore at least 20 Ma younger than the Purbeck specimens. Tooth morphology of these taxa is, however, plesiomorphic compared to the specimens documented here. This together with their temporal and geographical separation renders any close phylogenetic relationship between them and the Purbeck taxa unlikely. Tooth morphologies of the new Purbeck taxa are more similar to those of Late Cretaceous taxa than any described Early Cretaceous taxon and this suggests both an early origin and divergence of Eutheria and the early appearance of derived eutherian clades.
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Subclass Tribosphenida McKenna, 1975
Infraclass Eutheria Gill, 1872
Order and Family incertae sedis
Genus Durlstotherium nov.
ZooBank LSID: 12D080E5-88B1-4898-8D1B-96B44D247E77.
Type species: Durlstotherium newmani sp. nov., monotypic, see below.
Etymology: In reference to Durlston Bay, Dorset, UK, the locality from where the holotype and only specimen was obtained; and from Greek, therion, beast; a commonly used suffix in the names of mammals.
Diagnosis.—As for type species.
Stratigraphic and geographic range.—As for the type species.
Durlstotherium newmani sp. nov.
Figs. 3–6.
Etymology: In honour of Charlie Newman, recognising his intimate local knowledge, willingly shared, and his help in the field.
Holotype: NHMUK PV M 99991, a well preserved upper right distal-most (M3) tooth lacking only the labial roots, Figs. 3, 4A, 5A, 6A.
Type locality: Durlston Bay, Dorset, southern England, at British National Grid Reference SZ 03673 78377 (Fig. 1).
Type horizon: A beach-level exposure of bed DB 83 (Clements 1993) of the Lower Cretaceous, Berriasian (Ogg et al. 1994; Hunt 2004), Purbeck Group. (Fig. 2).
Diagnosis.—Monotypic tribosphenic mammal with a unique combination of derived dental characters. Differs from stem therians including Juramaia and Acristatherium, from the Oxfordian and Barremian of China respectively, but resembles Late Cretaceous eutherians, particularly asioryctitheres, cimolestids, and gypsonictopsids in: height and expansion of the protocone relative to labial cusps; possession of conules placed labially immediately adjacent to the paracone and metacone with sharp internal cristae and distance between conules as a percentage of total tooth length >31% <50% (44%) (Luo et al. 2003, 2011). Differs from Prokennalestes from the Aptian–Albian of Mongolia in the presence of a pre- and postcingulum. Resembles Gypsonictops from the Late Cretaceous of North America in the presence of strong cingula but differs from it in lack of a hypocone. Also differs from it in having a more connate paracone. Resembles members of Asioryctidae in the presence of a transversely elongate crown but differs from them in presence of strongly developed cingula. Also differs from Asioryctes from the Late Cretaceous of Mongolia in presence of a metaconule. Resembles members of Kennalestidae in the presence of cingula but differs from them in the presence of a preparastyle. Resembles members of Cimolestidae, which possess anteroposteriorly short and transversely broad upper molars with an anteroposteriorly compresed protocone and a paracone that is much taller than the metacone, but differs in the presence of a pre- and postcingulum which are either weak or lacking in members of Cimolestidae.

Fig. 3. Interpretive line drawings of teeth of the eutherian mammals Durlstotherium newmani gen. et sp. nov., NHMUK PV M 99991 (A) and Durlstodon ensomi gen. et sp. nov., NHMUK PV M 99992 (B) from the Purbeck Group exposed in Durlston Bay, Dorset, UK, in occlusal view, showing dental terminology (A, B) and dental measurements (C, D).
Description.—NHMUK PV M 99991 is an upper molar lacking only the labial roots. It is otherwise well preserved although substantially worn as a result of occlusion with the lower dentition and dietary attrition. It is a distal-most upper right molar (M3), identified as such on the basis of its general outline in occlusal view, the lack of a metastylar lobe and the expanded parastylar wing as defined by Kielan-Jaworowska et al. (2004; see also Figs. 3, 4A, 5A, 6A). Measurements are shown in Fig. 3.
The connate and slightly labiolingualy compressed paracone dominates the labial side of the crown and descends vertically. It is approximately three times the height of the metacone with a basal diameter a little over one and a half times that of the latter. The paracone is truncated by a substantial wear facet (facet 3, Fig. 6A) resulting from dietary attrition, that extends along the distal margin and across the narrow centrocrista to the base of the metacone. A labiolingually narrow wear facet, an extension of facet 1 (Fig. 6A), extends from the apex to the base of the paracone on its mesial surface. The metacone is closely appressed to the paracone and is placed distal and lingual to it. It is more heavily worn than the paracone with no trace of the original apex of the cusp remaining (facet 4, Fig. 6A). The flat, apical surface ascends towards the base of the paracone at an angle of about 45°. There is no metastylar lobe and consequently no ectoflexus. A low and apicobasally narrow ectocingulum, entirely devoid of cusps, extends from the linguobasal extremity of the metacone to the distal extremity of the parastylar wing. The latter is a prominent feature of the crown extending mesiolabially from the base of the paracone. It is strongly mesiolabially expanded with no evidence of molar interlock and bears a preparacrista that extends from the base of the paracone to the parastyle, the apex of which appears to have been substantially blunted by wear. There appears to be no stylocone but there is a minute cuspule on the anterior margin of the parastylar wing a short distance lingual to the parastyle that might be a remnant of it. A more substantial, but worn cusp taken to be the preparastyle is located lingual to this. A small labiolingually elongate and apicobasally narrow wear facet extends from the preparastyle to the lingual margin of the cuspule labial to it on the mesial margin of the parastylar wing. The substantial, labially placed paraconule is considerably worn mesially, facet 1 (Fig. 6A) and somewhat less so distolingually, facet 3 (Figs. 4A, 6A). It is pyramidal with a basal profile approximating that of an acute isosceles triangle, the long axis of which is mesiolabially inclined with the apex of the triangle at the labial end. Its labial surface ascends steeply to the base of the paracone forming with it a steep sided valley. The metaconule is also a substantial, labially placed cusp. Its labial surface ascends more steeply than that of the paraconule forming with the lingual surface of the metacone a sharply V-shaped valley. It is considerably worn mesiolingually, facet 4, but is unworn labially and distally except along the very short region originally occupied by the postmetaconule crista but now comprising the sharp edge of facet 2 (Fig. 6A). It is also pyramidal but unlike the paraconule its basal profile is that of an equilateral triangle the apex of which points lingually. The trigon basin occupying the area between the cusps described above has a smooth occlusal surface. It is deep and substantially worn (primarily by facet 4, Figs. 4A, 6A).
On the lingual part of the crown the worn preprotocrista extends from the apex of the protocone to overlie the paracone mesially. The postprotocrista extends labially to the lingual extremity of the metaconule. Wear also extends across the preparaconule crista, across the apex of the paraconule and along the postparaconule crista. On the distal side it extends from the apex of the sharply pointed metaconule, down the short premetaconule and postmetaconule cristae (facet 2, Fig. 6A). All cristae in this part of the crown form sharp shearing surfaces. The protocone is a robust cusp only slightly less elevated than the paracone. It is placed directly lingual to the paracone and mesiolingual to the metacone. The apex is mesiodistally compressed but expands basally and labially. Its apicobasal axis is tilted apicomesially. Its dorsoventral axis is mesially inclined at ca. 70°. There is a well-developed precingulum which has a wear facet on its mesial margin (facet 5, Fig. 6A). In apparent continuation of this there is also a small facet at the apex of the protocone on its mesial side. The precingulum overlies the mid part of the preprotocrista for about two thirds of its length. A substantial postcingulum, lacking a hypocone, extends from the lingual margin of the postmetaconule crista to a point opposite the lingual extremity of the precingulum and is of approximately the same length. It is somewhat apicobasally broader than the precingulum and it bears a correspondingly broader wear facet (facet 6, Fig. 6A). In contrast, however, there is no expression of this facet on the apex of the protocone on its distal margin.
The root overlying the protocone is very robust, apicobasally short and labiolingually broad. It is mesiodistally compressed with a mesiolingually projecting and expanded flange at its apex. Roots overlying the parastylar wing and the area between the paracone and metacone are broken away close to their bases. That part of the root overlying the parastylar wing is laterally compressed following the axis of the wing and is large compared to that overlying the junction between the paracone and metacone. The former is about two thirds the size in basal area of that overlying the protocone. The root occupying the area between the paracone and metacone is approximately circular in outline and is the smallest of the roots. Its diameter is approximately half the labiolingual width of its labiomesial neighbour.
Stratigraphic and geographic range.—Currently known only from the type horizon and locality.

Fig. 4. Stereo scanning electron micrographs of studied eutherian mammal specimens from the Berriasian Purbeck Group of Dorset, southern England; in occlusal view. A. Durlstotherium newmani gen. et sp. nov., NHMUK PV M 99991. B. Durlstodon ensomi gen. et sp. nov., NHMUK PV M 99992.
Genus Durlstodon nov.
ZooBank LSID: 12D080E5-88B1-4898-8D1B-96B44D247E77.
Type species: Durlstodon ensomi sp. nov., monotypic, see below.
Etymology: In reference to Durlston Bay, Dorset, UK, the locality from where the holotype and only specimen was obtained; and from Greek, odontos, tooth.
Diagnosis.—As for type species.
Stratigraphic and geographic range.—As for the type species.
Durlstodon ensomi sp. nov.
Figs. 3–6.
Etymology: In honour of Paul Ensom, recognising his major contribution to Purbeck palaeontology.
Holotype: NHMUK PV M 99992, a well preserved upper right distal-most (M3) tooth crown lacking the parastylar wing (Figs. 3, 4B, 5B, 6B.
Type locality: Durlston Bay, Dorset, southern England, at British National Grid Reference SZ 03673 78377 (Fig. 1).
Type horizon: A beach-level exposure of bed DB 83 (Clements 1993) of the Lower Cretaceous, Berriasian (Ogg et al. 1994; Hunt 2004), Purbeck Group (Fig. 2).
Diagnosis.—Monotypic tribosphenic mammal with a unique combination of derived characters. Differs from stem therians including Juramaia and Acristotherium, from the Oxfordian and Barremian of China respectively, but resembles Late Cretaceous eutherians, particularly members of Zhelestidae (sensu Archibald and Averianov 2012) in: height and expansion of the protocone relative to labial cusps; possession of conules placed labially immediately adjacent to the paracone and metacone with sharp internal cristae; protocone anteroposteriorly expanded and similar in height to the metacone; conular region width more than 51% of the total molar width (c. 64%); lower molars with a substantial entoconid as evidenced by a wear facet on the distal side of the protocone. Differs from members of Zhelestidae in possession of a paracone that is substantially taller than the protocone and metacone. Differs from Durlstotherium gen. nov. in: the presence of a rectangular trigon basin with internal conular cristae rather than a triangular trigon basin lacking these; in the more expanded protocone lacking any apical constriction; in presence of an angled postprotocrista (exaggerated by wear); in possession of an apparently more robust paraconule (possibly exaggerated by wear); in greater separation and relative sizes of the paracone and metacone; possession of a cuspidate ectocinglum; absence of pre- and postcingulum; pattern of wear (see below); and greater size. Other characters cannot be determined due to loss of the mesiolabial corner of the tooth.
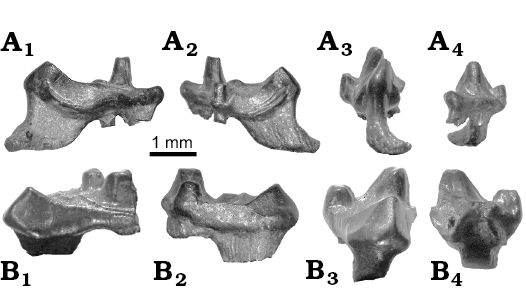
Fig. 5. Normal light photographs of studied specimens of eutherian mammal teeth from the Berriasian Purbeck Group of Dorset, southern England; in mesial (A1, B1), distal (A2, B2), lingual (A3, B3), and labial (A4, B4) views. A. Durlstotherium newmani gen. et sp. nov., NHMUK PV M 99991. B. Durlstodon ensomi gen. et sp. nov., NHMUK PV M 99992.
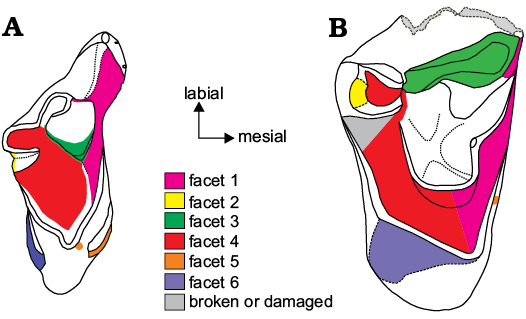
Fig. 6. Explanatory drawing of studied specimens of eutherian mammal teeth from the Berriasian Purbeck Group of Dorset, southern England; in occlusal view showing wear facets described in the text (from Crompton 1971). A. Durlstotherium newmani gen. et sp. nov., NHMUK PV M 99991. B. Durlstodon ensomi gen. et sp. nov., NHMUK PV M 99992.
Description.—NHMUK PV M 99992 is an upper molar lacking the labial roots and the parastylar wing. It is otherwise well preserved but bears a number of cracks in the region of the paracone. As with NHMUK PV M 99991 described above, it is substantially worn due to occlusion with the lower dentition and dietary attrition. It is also a distal-most upper right molar (M3) identified on the basis of its general outline in occlusal view and the lack of a metastylar lobe (Figs. 3, 4B, 5B, 6B). Measurements are shown in Fig. 3.
The connate paracone is somewhat labiolingually compressed apically and is the more robust of the labial cusps and descends vertically. It is a little less than twice the height of the metacone but the relative height of the cusps prior to substantial wear cannot be determined. The basal diameter of the paracone is twice that of the metacone. The paracone is truncated by a substantial wear facet (facet 3, Fig. 6B). This extends down the distal margin and across the centrocrista which is substantially broader than that of NHMUK PV M 99991. A wear facet (facet 1, Fig. 6B), occupies the apex of the paracone on its mesial surface and extends mesially along the preparacrista. The metacone is separated from the paracone by a U-shaped valley and is placed distal and slightly lingual to it. A metastylar lobe is absent but a low ectocingulum extends from the linguobasal extremity of the metacone to the distolabial extremity of the broken surface. It bears minute cuspules, the largest of which is at the base of the metacone at its distolabial corner. There is a substantial but heavily worn paraconule. It is separated from the paracone by a narrow valley the mesial margin of which comprises the preparaconule crista. This extends from the apex of the cusp to the broken surface at the labiomesial corner of the preserved part of the crown. The paraconule is considerably worn mesially, facet 1 (Figs. 4B, 6B). The metaconule is missing. However, examination of the lingual margin of the metacone and the area immediately lingual to it suggests that breakage has occurred here and that a substantial metaconule with a triangular basal profile was present and located close to the metacone.
The central part of the crown lingual to the labial cusps comprises a valley surrounded by a substantial ridge. This extends from the mesiolingual corner of the base of the metacone following the postprotocrista to a point opposite a sharp change in angle as the latter approaches the protocone. Here the ridge bends at right-angles mesially to approach the preprotocrista but before reaching this it bends again at right-angles to reach the lingual margin of the paraconule. Except as described above in connection with wear on the distolingual surface of the paraconule the valley appears to be unworn, other than over a narrow area adjacent to the mesiolingual corner of the metacone. In contrast, the remainder of the occlusal surface external to the valley is heavily worn (Figs. 4B, 6B).
The preprotocrista appears to extend from the apex of the protocone to overlie the paracone but substantial wear has separated it from the now apparently overlying preparaconule crista and exposed the intervening dentine. Mesially a residual enamel ridge extends as far as the point of breakage labial to the paracone. The ventral surface is worn over its entire length except where obscured by the postparaconule crista. There is also lateral wear extending from a point above the apex of the paraconule to the labial end of the crest as preserved. Confluent lateral wear is also present on the postparaconule crista along its length. The postprotocrista is also heavily worn. It extends labially to the point of breakage at the lingual extremity of the missing metaconule.
The protocone is a robust cusp slightly taller than the heavily worn metacone. It is placed lingual to the distal margin of the paracone and mesiolingual to the metacone. The apex defined by the cristae is broadly V-shaped and the postprotocrista is sharply inflected midway along its length, this inflection being exaggerated by a substantial wear facet (facet 6, Fig. 6B). The protocone expands basally and its apicobasal axis is tilted apicomesially at c. 75°, more so than in NHMUK PV M 99991. A precingulum is absent but there is a basal swelling on the mesial surface of the protocone and a small, vertically orientated facet is also present on its mesial surface (facet 5, Fig. 6B). It is situated below the preprotocrista midway between its apex and that of the paraconule. There is also a swelling towards the base of the protocone on the distal side. This appears to be more pronounced than that on the mesial side. However, there is a large wear facet (facet 6, Fig. 6B) which has partially obliterated it. This facet is at a shallower angle than the apicobasal axis of the cusp and must represent wear from a substantial entoconid on the occluding lower molar.
The root overlying the protocone is very robust, mesiodistally narrow and labiolingually broad. It is broken away a short distance from its base. Despite being robust it overlies a smaller area of the crown than the root in a similar position in NHMUK PV M 99991. The root overlying the paracone, and which would evidently also have extended to support the missing parastylar wing, is also laterally compressed as in NHMUK PV M 99991 and is large compared to that overlying the metacone. It appears to be about two thirds the size in basal area of that overlying the protocone but its labiobasal end is missing. The root above the metacone is approximately circular and the smallest of the roots. Its diameter is smaller than the remaining part of the root above the paracone. Proportionately, however, it is of greater diameter relative to the other roots than the comparable but somewhat more mesially placed root in NHMUK PV M 99991.
Stratigraphic and geographic range.—Currently known only from the type horizon and locality.
Concluding remarks
Juramaia from the Oxfordian (Late Jurassic) of China is presently the oldest tribosphenidan. If, as claimed (Luo et al. 2011), it is a eutherian, it predates by more than 30 My the previous earliest purported records for this clade, Eomaia (Ji et al. 2002) and Acristatherium (Hu et al. 2009) from the Barremian (Early Cretaceous) of China. It also exceeds one estimate (Bininda-Emonds et al. 2007) for the early divergence of Eutheria from a therian ancestor which placed this at between 148 and 130 Ma based on molecular data. However, all of these records are at odds with a more recent analysis which employed large datasets incorporating both phenomic and molecular characters (O’Leary et al. 2013), and controversially concluded that Eutheria is considerably younger than estimated from previous studies. The study also concluded that Eomaia falls outside of Eutheria as a stem taxon to Theria (Juramaia and Acristatherium were not included in this analysis). O’Leary et al. (2013) also conclude that the earliest date of emergence of Eutheria, constrained by taxa such as Maelestes (Wible et al. 2009) is 91 Ma and that of Theria is 127.5 Ma, strongly contrasting with one molecular-based estimate that suggests a date of emergence of 190 Ma for the latter (Meredith et al. 2011). A subsequent morphological study (Averianov 2015) also excludes Juramaia and Eomaia from Eutheria and has led to a reappraisal of other Early Cretaceous taxa previously assigned to Eutheria, relegating some to a position outside of this clade. In North America, Early Cretaceous tribosphenidans are known from the Aptian–Albian Antlers Formation of the Trinity Group of Oklahoma and Texas (Davis and Cifelli 2011; Averianov 2015) and the Albian part of the Cloverly Formation of Montana and Wyoming (Cifelli and Davis 2015) with some originally assigned to Eutheria. However, considerable debate still exists as to the phylogenetic placement of the so-called Trinity therians and in the most recent review (Averianov 2015) none are considered eutherians. Some dental characters of Juramaia, and most upper molar characters for Eomaia, are unknown (Ji et al. 2002); however, a low and unexpanded protocone and the lingual placement of conules lacking cristae suggest that Juramaia should also be excluded from Eutheria, as should Acristatherium for similar reasons. Based on dental characters accepted as diagnostic for Eutheria, the taxa described here, which date from approximately 145 Ma, display highly derived tooth morphologies that, despite being distal-most upper molars, support their assignment to Eutheria. These characters include reduction of the stylocone and the presence of a wide, flange-like parastylar lobe, clearly seen in Durlstotherium (Davis and Cifelli 2011), and are otherwise summarised above and by Averianov (2015). The presence of these highly derived eutherians in the earliest Cretaceous of southern Britain (Fig. 7) is remarkable and supports an early emergence of Eutheria from a therian ancestor. Furthermore, if, as seems likely, Juramaia is excluded from Eutheria (Averianov 2015; O’Leary et al. 2013), Durlstotherium gen. nov. and Durlstodon gen. nov. represent the earliest eutherians currently recorded. Similarities in tooth morphology to taxa occurring in the Late Cretaceous, asioryctitheres, cimolestids, and gypsonictopsids in the case of the former and zhelestids in the case of the latter, also suggest an early divergence of Eutheria and an early emergence of derived eutherian clades including these and others.

Fig. 7. Artist’s impression of the Purbeck lagoon at dusk with Durlstodon gen. nov. (left foreground), Durlstotherium gen. nov. (right and center foreground) and the theropod Nuthetes holding a captured Durlstotherium (centre middle distance). Artwork by Mark Witton.
Acknowledgements
We thank Charlie Newman (Worth Matravers, Dorset, UK) for his hospitality and help with fieldwork, and Paul Ensom (Falmouth, Cornwall, UK) for advice concerning stratigraphy and for general discussions concerning Purbeck Group mammals. Alexander Averianov (Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia), Richard Cifelli (School of Geology and Geophysics, University of Oklahoma, Norman, USA), Brian Davis (Department of Anatomical Sciences and Neurobiology, University of Louisville School of Medicine, USA), and Jerry Hooker (Natural History Museum, London, UK) are thanked for their generous assistance and comments while drafting the manuscript. We are very grateful to Alexander Averianov and Brian M. Davis for their thorough and constructive reviews of the submitted manuscript which resulted in significant improvements. Pip Brewer (Natural History Museum, London, UK) provided curatorial assistance for which we are grateful. Jo Dunlop (University of Portsmouth, UK) provided assistance while obtaining SEM images and Emily Roberts (University of Portsmouth, UK) while obtaining stacked digital images. We are grateful to both. Mark Witton (University of Portsmouth, UK) is also thanked for his excellent artwork. We have no competing interests and all funding for this project was provided by the University of Portsmouth.
References
Allen, P. 1998. Purbeck–Wealden (Early Cretaceous) climates. Proceedings of the Geologists’ Association 109: 197–236.
Allen, P. and Wimbledon, W.A. 1991. Correlation of NW European Purbeck−Wealden (non-marine Lower Cretaceous) as seen from the English type−areas. Cretaceous Research 12: 511–526. Crossref
Archibald, J.D. and Averianov, A.O. 2012. Phylogenetic analysis, taxonomic revision, and dental ontogeny of the Cretaceous Zhelestidae (Mammalia: Eutheria). Zoological Journal of the Linnean Society 164: 361– 426. Crossref
Averianov, A.O. 2015. Taxonomic revision of tribosphenic mammals from the Lower Cretaceous Antlers Formation of Texas and Oklahoma, USA. Proceedings of the Zoological Institute RAS 319: 141–181.
Averianov, A.O., Archibald, J.D., and Ekdale, E.G. 2010. New material of the Late Cretaceous deltatheroidan mammal Sulestes from Uzbekistan and phylogenetic reassessment of the metatherian-eutherian dichotomy. Journal of Systematic Palaeontology 8: 301–330. Crossref
Batten, D.J. 2002. Palaeoenvironmental setting of the Purbeck Limestone Group of Dorset, southern England. In: A.R. Milner and D.J. Batten (eds.), Life and Environments in Purbeck Times. Special Papers in Palaeontology 68: 13–20.
Bininda-Emonds, O.R.P., Cardillo, M., Jones, K.E., MacPhee, R.D.E., Beck, R.M.D., Grenyer, R., Price, S.A., Vos, R.A., Gittleman, J.L., and Purvis, A. 2007.The delayed rise of present-day mammals. Nature 449: 507–512. Crossref
Cifelli, R.L. and Davis, B.M. 2015. Tribosphenic mammals from the Lower Cretaceous Cloverly Formation of Montana and Wyoming. Journal of Vertebrate Paleontology 35 (3): e920848. Crossref
Clements, R.G. 1993. Type-section of the Purbeck Limestone Group, Durlston Bay, Swanage, Dorset. Proceedings of the Dorset Natural History and Archaeological Society 114: 181–206.
Crompton, A.W. 1971. The origin of the tribosphenic molar. In: D.M. Kermack and K.A. Kermack (eds.). Early Mammals. Zoological Journal of the Linnean Society 50 (Supplement 1): 65–87.
Davis, B.M. and Cifelli, R.L. 2011. Reappraisal of the tribosphenidan mammals from the Trinity Group (Aptian–Albian) of Texas and Oklahoma. Acta Palaeontologica Polonica 56: 441–462. Crossref
Gill, T.N. 1872. Arrangement of the families of mammals. With analytical tables. Smithsonian Miscellaneous Collections 11: 1–98.
Hu, Y., Meng, J., Li, C., and Wang, Y. 2009. New basal eutherian mammal from the Early Cretaceous Jehol biota, Liaoning, China. Proceedings of the Royal Society, B 277: 229–236. Crossref
Hu, Y., Meng, J., Wang, Y., and Li, C. 2005. Large Mesozoic mammals fed on young dinosaurs. Nature 433: 149–152. Crossref
Hunt, C.O. 2004. Palynostratigraphy of the classic Portland and Purbeck sequences of Dorset, southern England, and the correlation of Jurassic–Cretaceous boundary beds in the Tethyan and Boreal realms. Geological Society, London, Special Publications 230: 175–186. Crossref
Ji, Q., Luo, Z.-X., Yuan, C.-X., Wible, J.R., Zhang, J.-P., and Georg, J.A. 2002. The earliest known eutherian mammal. Nature 416: 816–822. Crossref
Kielan-Jaworowska, Z., Cifelli, R.C., and Luo, Z.-X. 2004. Mammals from the Age of Dinosaurs: Origins, Evolution and Structure. 630 pp. Columbia University Press, New York. Crossref
Linnaeus, C. 1758. Systema naturae perregna tria naturae, secundum classis, ordines, genera, species cum characteribus, differentiis, synonymis, locis. Vol. 1, Regnum animale. Editio decima, reformata. 824 pp. Laurentii Salvii, Stockholm.
Luo, Z.-X., Ji, Q., Wible, J.R, and Yuan, C.-X. 2003. An Early Cretaceous tribosphenic mammal and metatherian evolution. Science 302: 1934–1940. Crossref
Luo, Z.-X., Yuan, C.-X., Meng, Q.-J., and Ji, Q. 2011. A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature 476: 442–445. Crossref
McKenna, M.C. 1975. Toward a phylogenetic classification of the Mammalia. In: W.P. Luckett and F.S. Szalay (eds.), Phylogeny of the Primates, 21–46. Plenum Press, New York. Crossref
Meredith, R.W., Janečka, J.E., Gatesy, J., Ryder, O.A., Fisher, C.A., Teeling, E.C., Goodbla, A., Eizirik, E., Simão, T.L.L., Stadler, T., Rabosky, D.L., Honeycutt, R.L., Flynn, J.J., Ingram, C.M., Steiner, C., Williams, T.L., Robinson, T.J., Burk-Herrick, A., Westerman, M., Ayoub, N.A., Springer, M.S., and Murphy, W.J. 2011. Impacts of the Cretaceous Terrestrial Revolution and KPg extinction on mammal diversification. Science 334: 521. Crossref
Ogg, J.G., Hasenyager, R.W., and Wimbledon, W.A. 1994. Jurassic–Cretaceous boundry: Portland-Purbeck magnetostratigraphy and possible correlation to the Tethyan faunal realm. Geobios 17: 519–527. Crossref
O’Leary, M.A.O., Bloch, J.I., Flynn, J.J, Gaudin, T.J., Giallombardo, A., Giannini, N.P., Goldberg, S.L., Kraatz, B.P., Luo, Z.-X., Meng, J., Ni, X., Novacek, M.J., Perini, F.P., Randall, Z.S., Rougier, G.W., Sargis, E J., Silcox, M.T., Simmons, N.B., Spaulding, M., Velazco, P.M., Weksler, M., Wible, J.R., and Cirranello, A.L. 2013. The placental mammal ancestor and the post-K/Pg radiation of placentals. Science 339: 662–667. Crossref
Owen, R. 1854. On some fossil reptilian and mammalian remains from the Purbecks. Quarterly Journal of the Geological Society 10: 420–433. Crossref
Owen, R. 1871. Monograph of the fossil Mammalia of the Mesozoic formations. Monograph of the Palaeontological Society 33 (for 1870): 1–115.
Radley, J.D. 2002. Distribution and palaeoenvironmental significance of molluscs in the late Jurassic–early Cretaceous Purbeck Formation of Dorset, southern England: a review. In: A.R. Milner and D.J. Batten (eds.), Life and Environments in Purbeck Times. Special Papers in Palaeontology 68: 41–52.
Sigogneau-Russell, D., Hooker, J.J., and Ensom P. 2001. The oldest tribosphenic mammal from Laurasia (Purbeck Limestone Group, Berriasian, Cretaceous, UK) and its bearing on the “dual origin” of Tribosphenida Comptes Rendus de l’Académie des Sciciences, Paris 333: 141–147.
Sladen, C.P. and Batten, D.J. 1984. Source-area environments of Late Jurassicand Early Cretaceous sediments in southeast England. Proceedings of the Geologists’ Association 95: 149–163.
Wible, J.R., Rougier, G.W., Novacek, M.J., and Asher, R.J. 2009. The eutherian Maelestes gobiensis from the Late Cretaceous of Mongolia and the phylogeny of Cretaceous Eutheria. Bulletin of the American Museum of Natural History 327: 1–123. Crossref
Acta Palaeontol. Pol. 62 (4): 657–665, 2017
https://doi.org/10.4202/app.00408.2017