
A new burmagomphid dragonfly from the Eocene of Patagonia, Argentina
JULIÁN F. PETRULEVIČIUS
A new burmagomphid anisopteran, Satelitala soberana gen. et sp. nov. is described from the lower Eocene of Laguna del Hunco, Patagonia, Argentina. The new genus is characterised by hindwing characters such as the subdiscoidal triangle not elongated; anal loop divided longitudinally; paranal cell divided longitudinally; five terminal cells between RP and MA; five terminal cells between MP and CuA; and obtuse angle between PsA and CuP+AA. Burmagomphid dragonflies were represented so far only by one specimen from the middle Cretaceous of Southeast Asia. This new record extends the distribution to Patagonia, to the Cenozoic, and also to paleolake deposits.
Introduction
The family Burmagomphidae Zheng, Nel, and Wang, 2017 was recently described from the Cretaceous Burmese amber from Myanmar (Zheng et al. 2017). Burmagomphidae was included into the clade Gomphides Bechly, 2007 that includes 6 extant families and 2 extinct families from the Jurassic of Solnhofen (Germany), and Cretaceous of Santana (Brazil) (Bechly 2007).
The specimen studied here represents the second dragonfly from Laguna del Hunco and the first one belonging to Exophytica Bechly, 1996. The first dragonfly, Huncoaeshna corrugata Petrulevičius, Nel, and Voisin, 2010, was assigned to Aeshnidae (Petrulevičius et al. 2010). Other fossil adult representatives of Gomphides recorded in South America are all from the Santana Formation: the extinct Araripegomphidae Bechly, 1996 and Proterogomphidae Bechly, Nel, Martínez-Delclòs, and Fleck, 1998, and the extant family Lindeniidae Jacobson and Bianchi, 1905 (e.g., Araripegomphus cretacicus Nel and Paicheler, 1994, Cordulagomphus primaerensis Petrulevičius and Martins-Neto, 2007, Cratolindenia knuepfae Bechly, 2000, respectively). A gomphid-like nymph is also present in the close by locality of Confluencia (Ypresian?), together with other nymphs resembling families such as Synlestidae and Libellulidae (Petrulevičius 2012, 2013). Other specimens of Odonata (not anisopterans) described from Laguna del Hunco are representatives of Frenguelliidae Petrulevičius and Nel, 2003; and Austroperilestidae (Petrulevičius and Nel 2005, 2007).
Herein I record the second species of Burmagomphidae represented by a specimen coming from the Ypresian locality of Laguna del Hunco (52.2 Ma; paleolatitude c. 47°S), Chubut province, Patagonia, Argentina. Laguna del Hunco is a well sampled locality from which diverse insects, mainly Odonata, were recovered (Fidalgo and Smith 1987; Petrulevičius 2005, 2009, 2013, 2016; Petrulevičius and Nel 2003, 2005, 2007, 2013; Petrulevičius et al. 2010). Laguna del Hunco is also a hot spot of plant diversity of the Eocene (Wilf 2012; Wilf et al. 2003, 2005).
Institutional abbreviations.—MPEF-PI, Museo Paleontológico Egidio Feruglio, Trelew, Chubut, Argentina.
Other abbreviations.—AA, anterior anal; AL, anal loop; Ax0, Ax1, Ax2, primary antenodal crossveins; CuA, anterior cubitus; IR, intercalary radial veins; MA, anterior median; MP, posterior median; Mspl, median supplement; “O”, oblique vein; PC, paranal cell; PsA, pseudo-anal vein; R, radius; RA, anterior radius; RP, posterior radius; Rspl, radial supplement; ScP, posterior subcosta.
Material and methods
The material was collected from the volcanic caldera lake bed locality Laguna del Hunco, province of Chubut, Patagonia, Argentina (Wilf et al. 2003). It is represented by a single specimen, holotype MPEF-PI 1001, collected from pyroclastic debris of the locality LH-25, 42°30’S, 70°02’W (Wilf 2012; Wilf et al. 2003, 2005). The locality was dated using 40Ar/39Ar by Wilf et al. (2005) and recalculated by Wilf (2012), providing an age of 52.22 ± 0.22 (analytical 2 σ), ± 0.29 (full 2 σ) Ma.
In this work, I follow the wing venation nomenclature of Kukalová-Peck (1983), amended by Kukalová-Peck (1991, 2008), also contributions by Riek and Kukalová-Peck (1984), Nel et al. (1993), Bechly (1996), and Petrulevičius and Gutiérrez (2016). The higher classification of fossil and extant Odonata is based on the phylogenetic system of Bechly (1996, 2007).
The new species was drawn and photographed in detail using a camera lucida and a Leica digital camera (DMC2900), attached to a Leica M205C stereomicroscope. Habitus photographs of the specimen were taken using a camera Nikon Coolpix 7100.
Systematic palaeontology
Odonata Fabricius, 1793
Gomphides Bechly, 1996
Burmagomphidae Zheng, Nel, and Wang, 2017
Genus Satelitala nov.
Etymology: From Castilian satélite, satellite and Greek ala, wing.
Type species: Satelitala soberana sp. nov. (by monotypy); see below.
Diagnosis.—As for the type species by monotypy.
Satelitala soberana sp. nov.
Figs. 1, 2.
Etymology: From Castilian soberana, sovereign.
Holotype: MPEF-PI 1001, counterpart of an incoplete hind wing.
Type locality: Laguna del Hunco, Chubut, Patagonia, Argentina.
Type horizon: Ypresian (52 Ma), lower Eocene.
Diagnosis.—The genus is characterized by the following hindwing characters: (1) wing medium sized (c. 29 mm long); (2) narrow wing base, gradually widening from base towards nodus; (3) arculus close to Ax1; (4) bases of RP and MA close at arculus; (5) discoidal triangle, hypertriangle and subdiscoidal triangle free; (6) discoidal triangle elongated; (7) trigonal planate well developed; (8) CuA1+2 distinctly shortened with reduced pectinate branching; (9) midfork one cell distal to discoidal triangle; (10) RP branches symmetrical at midfork; (11) one antefurcal crossvein; (12) no Mspl nor Rspl; (13) PsA aligned with anterior side of triangle; (14) extremely acute (c. 10º) angle between CuA+MP and PsA; (15) ScP basally close to RA+RP; (16) subdiscoidal triangle not elongated; (17) anal loop closed (?) and longitudinally two (?) celled; (18) paranal cell rounded divided longitudinally; (19) “cordulegastrid gap”; (20) obtuse (c. 140°) angle between PsA and CuP+AA; (21) five terminal cells between RP and MA; (22) five terminal cells between MP and CuA.
Characters 1–15 are diagnostic characters of the family according to Zheng et al. (2017). Characters 13–15 are newly proposed for Burmagomphidae. Characters 16–22 are unique from the new genus. Two characters proposed by Zheng et al. (2017) have to be removed from the diagnosis of the family and placed in the genus Burmagomphides Zheng, Nel, and Wang, 2017 because of their absence in Satelitala gen. nov., i.e. subdiscoidal triangle elongated; no “cordulegastrid gap”.
Description.—Specimen consists of 3/4 part of a hyaline hindwing (Fig. 1); preserved length 22 mm, 7.5 mm wide; reconstructed length of wing c. 29 mm; distance between base and arculus, 4.3 mm, between arculus and nodus, 8 mm; ScP almost touching R at wing base and ending at midpoint between anterior wing margin and RA at nodus; primary antenodals present; Ax0 (?) near wing base, 1.7 mm from Ax1; Ax1 0.8 mm basal to arculus; no secondary antenodals between Ax0 and Ax1; Ax2 (?) at level of distal end of discoidal triangle; two (?) secondary antenodals between Ax1 and Ax2; two (?) secondary antenodal and three (?) antesubnodal crossveins distal Ax2, 4 preserved postnodal and two (?) postsubnodal crossveins preserved before pterostigma; postnodal and postsubnodal crossveins not aligned; RP and MA separate at arculus; median and submedian spaces free of crossveins; hypertriangle, discoidal triangle and subtriangle free; anterior side of hypertriangle well curved, 3 mm long and 0.56 mm wide; triangle 0.7 mm distal to arculus, anterior side 2.2 mm long, basal side 1 mm long and distal side 2.4 mm long; distal side of triangle somewhat curved at anterior part, subdiscoidal triangle not elongated, with anterior side (PsA) strong and oblique aligned with the anterior side of discoidal triangle (Figs. 1B, 2A), PsA 1.1 mm long, posterior side 1.6 mm long and distal side 0.9 mm long; MP+CuA and PsA at an extremely acute angle (c. 10°); obtuse angle (c. 140°) between PsA and CuP+AA; postdiscoidal area with one (?) row of cells basally to c. 18 rows of cells at wing margin; no Rspl nor Mspl; area between RA and RP basal to RP3+4 with one crossvein and one just at RP3+4 base; “cordulegastrid gap” present; one antefurcal crossvein basal to midfork; midfork one cell distal to end of triangle and five (?) cells to nodus; RP2 arise at subnodus; nodus vertical, subnodus oblique (Figs. 1B, 2B); area between RP2 and IR2 with one row of cells before “O”, expanded distally; RP3/4 and MA running almost parallel, with one row of cells before “O” and five rows along wing margin; area between CuA and MP with one row of cells basally, and five cells along wing margin; anal area broad with 4–5 cells; anal loop closed (?) with a longitudinal division forming at least two cells; paranal cell rounded divided longitudinally into two cells; cubito anal area broad with five rows of cells between CuA1+2 and wing margin; CuA1+2 with two (?) posterior branches; CuA3+4 strongly curved reaching obliquely the posterior margin; two recognisable AA veins.
Remarks.—The wing is quite wrinkled; the posterior wing margin at the level of CuA1+2 and MP is not in its original position, and is supposed to be actually closer to the wing (the interpreted position is indicated by the dotted line in Fig. 1B). According to this, the width of the wing is gradually increasing from the wing base towards the nodus. A part of the anterior wing margin is inversed in position (see curved arrow in Fig. 1B).
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 1. Burmagomphid dragonfly Satelitala soberana gen. et sp. nov., holotype, MPEF-PI 1001, from Laguna del Hunco (Chubut, Patagonia, Argentina); Ypresian, lower Eocene. Photograph (A), explanatory drawing (B).
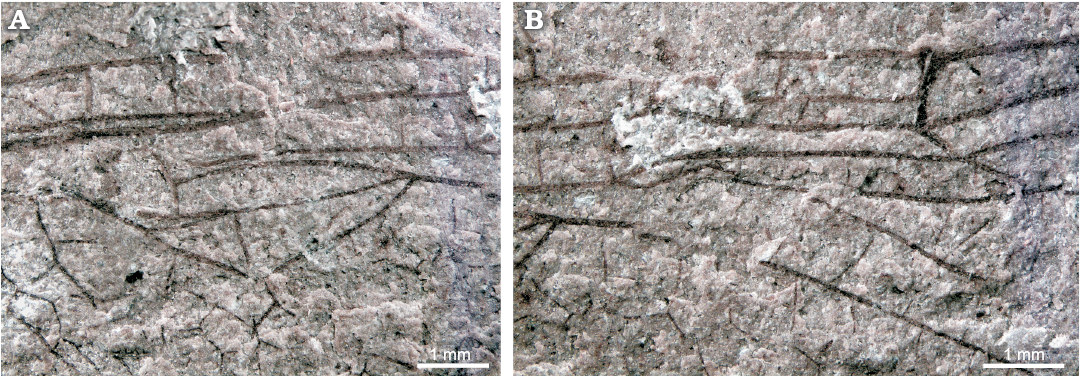
Fig. 2. Details of wing of burmagomphid dragonfly Satelitala soberana gen. et sp. nov., holotype MPEF-PI 1001 from Laguna del Hunco (Chubut, Patagonia, Argentina); Ypresian, lower Eocene. Basal part with triangles and arculus (A), median part with nodus (B).
Discussion
Wings of Gomphaeshnidae and Gomphides are similar at first glance but after a more detailed examination of characters both groups can be differentiated. These clades belong to Aeshnoptera and Exophytica respectively, sister groups of Euanisoptera. The wing studied here could be excluded from Aeshnoptera because of the absence of Rspl and Mspl, and a non-undulated RP3+4 and MA (Bechly 2007; Zheng et al. 2017).
The specimen could be attributed to Gomphides because the PsA and the subdiscoidal triangle are distinct; the discoidal triangle has at least a somewhat angulated distal side, caused by a more or less distinct supplementary sector (trigonal planate); the hypertriangle has a distinctly curved anterior margin; and there is a strong tendency towards development of the straight arculus (Bechly 2007; Zheng et al. 2017). Within Gomphides, attribution of the described specimen to Progomphidae could be excluded by their absence of an anal loop and a paranal cell. Likewise, assignation to the Araripegomphidae could be excluded because this family is characterized by having two or more antefurcal crossveins (contra one in the new species) and a wide hindwing base. Desmoproctida Bechly, 1996, the third clade within Gomphides, has no autapomorphies in wing venation (Bechly 2007) making attribution of the new species to this clade problematic. Notwithstanding, one included clade, the Lindeniidae, could be excluded by their numerous antefurcal crossveins (contra one in the new species), the higher number of terminal cells between RP and MA and between MP and CuA and a wide hindwing base. The other clade of Desmoproctida, the Oligophlebiata, includes three taxa, the Hagenioidea, Brevicubitalia and Burmagomphidae (Zheng et al. 2017). The new species shares with them the symmetrical RP branches at the midfork. It differs from Proterogomphidae (Hagenioidea) by having a narrower hind wing base and only one antefurcal crossvein, and from Hageniidae by having the MP and CuA1+2 not strongly divergent towards the wing margin (Bechly 2007). Bechly (2007) considers that Brevicubitalia has a “CuAa (CuA1+2 in present paper) distinctly shortened and with a distinct course (distally converging with the wing margin, instead of running parallel to it), and with a reduced pectinate branching. This character is present in the new species but the CuA1+2, as in Burmagomphidae, is even shorter. The Zonophoridae (Brevicubitalia) have the discoidal triangle somewhat elongated but differ from the new species by having a triangle divided, numerous antefurcal crossveins and no paranal cell developed. The remaining Brevicubitalia, the Gomphida, have a reduced anal loop and no paranal cell. It should be also noted that the Zonophoridae and Epigomphidae (Gomphida) have a tendency to a parallelization of the PsA with the anterior side of the triangle but this tendency does not reach the parallel position present in the new species and the Burmagomphidae. The new species also shares with the Burmagomphidae a narrow hind wing base, its width gradually increasing from the wing base towards the nodus (Zheng et al. 2017) and two other synapomorphies proposed in this paper: the extremely acute angle between CuA+MP and PsA, and a ScP basally close to RA+RP.
Concluding remarks
I report a burmagomphid compression fossil from the Eocene caldera lake insect beds of Laguna del Hunco in Patagonia, Argentina. So far burmagomphid dragonflies were only reported in an amber inclusion from the Cretaceous of Myanmar (Burma) (Zheng et al. 2017). The new species could be included into a new genus Satelitala because of some differences with the only described genus of Burmagomphidae, Burmagomphides. Satelitala gen. nov. reveals a less elongated subdiscoidal triangle, an anal loop divided longitudinally, a bigger paranal cell, five terminal cells between RP and MA, five terminal cells between MP and CuA, and an obtuse angle between PsA and CuP+AA.
The present discovery of a new genus of Burmagomphidae in the Eocene of Patagonia is noteworthy for the reconstruction of the biogeographic and phylogenetic history of the Gomphides. Their oldest record in the Cenomanian of Asia proposes a distribution in Gondwana for the clade at least at the Early Cretaceous. The presence of extinct families in the Eocene of Patagonia adds complexity to future discussions and also questions the taxonomic attribution to extant families of the immature specimens from Confluencia (considered “gomphid-like” by Petrulevičius 2013), and of endophytic ovipositions from Laguna del Hunco (attributed to Coenagrionidae by Sarzetti et al. 2009). It cannot be excluded that these specimens should be assigned to burmagomphid nymphs and frenguelliid ovipositions, respectively.
Acknowledgements.—Thanks are due to Rubén Cúneo and Eduardo “Dudu” Ruigómez (both Museo Egidio Feruglio, Trelew, Argentina), and also to reviewers, Daran Zheng (Chinese Academy of Sciences, Nanjing, China) and Stefan Pinkert (Philipps Universität Marburg, Germany), for helpful comments. Funding support for the fieldtrip and labo studies came from grants: PIP 0834 from the National Research Council of Argentina (CONICET); PICT-2016-4297 from the National Agency of Scientific and Technological Promotion of Argentina (ANPCyT); and DEB-1556666/1556136 from the National Science Foundation of USA (NSF).
References
Bechly, G. 1996. Morphologische Untersuchungen am Flügelgeäder der rezenten Libellen und deren Stammgruppenvertreter (Insecta; Pterygota; Odonata), unter besonderer Berücksichtigung der Phylogenetischen Systematik und des Grundplanes der *Odonata. Petalura 2: 1–402.
Bechly, G. 2007. Phylogenetic Systematics of Odonata. https://bechly.lima-city.de/system.htm
Fidalgo, P. and Smith, D.R. 1987. A fossil Siricidae (Hymenoptera) from Argentina. Entomological News 98: 63–66.
Kukalová-Peck J. 1983. Origin of the insect wing and wing articulation from the arthropodan leg. Canadian Journal of Zoology 61: 1618–1669. Crossref
Kukalová-Peck, J. 1991. Fossil history and the evolution of hexapod structures. In: I.D. Naumann (ed.), The Insects of Australia, A Textbook for Students and Research Workers, 2nd Ed., Vol. 1, 141–179. Melbourne University Press, Melbourne.
Kukalová-Peck, J. 2008. Phylogeny of higher taxa in Insecta: finding synapomorphies in the extant fauna and separating them from homoplasies. Evolutionary Biology 35: 4–51. Crossref
Nel, A., Martínez-Delclòs, X., Paicheler, J.C., and Henrotay, M. 1993. Les “Anisozygoptera” fossiles. Phylogénie et classification. (Odonata). Martinia, Numéro Hors-Série 3: 1–311.
Petrulevičius, J.F. 2005. Avances en el conocimiento de la diversidad de insectos de Laguna del Hunco, Eoceno inferior de Chubut, Patagonia, Argentina. Ameghiniana 42 (Suplement 4): 40R.
Petrulevičius, J.F. 2009. A Panorpoidea (Insecta: Mecoptera) from the lower Eocene of Patagonia, Argentina. Journal of Paleontology 83: 994–997. Crossref
Petrulevičius, J.F. 2012. Libélulas (Insecta: Odonata) del Eoceno de Patagonia, Argentina y su aporte al paleoambiente de Laguna del Hunco y Confluencia. Ameghiniana 49: R154.
Petrulevičius, J.F. 2013. Palaeoenvironmental and palaeoecological implications from body fossils and ovipositions of Odonata from the Eocene of Patagonia, Argentina. Terrestrial Arthropod Reviews 6: 53–60. Crossref
Petrulevičius, J.F. 2016. A new pentatomoid bug from the Ypresian of Patagonia, Argentina. Acta Palaeontologica Polonica 61: 863–868. Crossref
Petrulevičius, J.F. and Nel, A. 2003. Frenguelliidae, a new family of dragonflies from the earliest Eocene of Argentina (Insecta: Odonata). Phylogenetic relationships within Odonata. Journal of Natural History 37: 2909–2918. Crossref
Petrulevičius, J.F. and Nel, A. 2005. Austroperilestidae, a new family of damselflies from the earliest Eocene of Argentina (Insecta: Odonata). Phylogenetic relationships within odonata. Journal of Paleontology 79: 658–662. Crossref
Petrulevičius, J.F. and Nel, A. 2007. Enigmatic and little known Odonata (Insecta) from the Paleogene of Patagonia and northwest Argentina. Annales de la Societé Entomologique de France 43: 341–347. Crossref
Petrulevičius, J.F. and Nel, A. 2013. A new Frenguelliidae (Insecta: Odonata) from the early Eocene of Laguna del Hunco, Patagonia, Argentina. Zootaxa 3616: 597–600. Crossref
Petrulevičius, J.F. and Gutiérrez, P.R. 2016. New basal Odonatoptera (Insecta) from the lower Carboniferous (Serpukhovian) of Argentina. Arquivos Entomolóxicos 16: 341–358.
Petrulevičius, J.F., Nel, A., and Voisin, J.-F. 2010. Discovery of a new genus and species of darner dragonfly (Aeshnidae: Odonata) from the lower Eocene of Laguna del Hunco, Patagonia, Argentina. In: A. Nel, D. Azar, and J.F. Petrulevičius (eds.), Fossil Insects, Systematics, Phylogeny, and Palaeoecology. Annales de la Société Entomologique de France, Special Issue 46: 271–275.
Riek, E.F. and Kukalová-Peck, J. 1984. A new interpretation of dragonfly wing venation based upon Early Carboniferous fossils from Argentina (Insecta: Odonatoidea) and basic characters states in pterygote wings. Canadian Journal of Zoology 62: 1150–1166. Crossref
Sarzetti, L.C., Labandeira, C.C., Muzón, J., Wilf, P., Cúneo, N.R., Johnson, K.R., and Genise, J.F. 2009. Odonatan endophytic oviposition from the Eocene of Patagonia: the ichnogenus Paleoovoidus and implications for behavioural stasis. Journal of Paleontology 83: 431–447. Crossref
Wilf, P. 2012. Rainforest conifers of Eocene Patagonia: attached cones and foliage of the extant southeast-Asian and Australasian genus Dacrycarpus (Podocarpaceae). American Journal of Botany 99: 562–584. Crossref
Wilf, P., Cúneo, N.R., Johnson, K.R., Hicks, J.F., Wing, S.L., and Obradovich, J.D. 2003. High plant diversity in Eocene South America: Evidence from Patagonia. Science 300: 122–125. Crossref
Wilf, P., Johnson, K.R., Cúneo, N.R., Smith,
M.E., Singer, B.S., and Gandolfo, M.A. 2005. Eocene plant diversity at
Laguna del Hunco and Río Pichileufú, Patagonia, Argentina. The American Naturalist 165: 634–650. Crossref
Zheng, D., Nel, A., Chang, S.-C., Jarzembowski, E.A., Zhang, H., and Wang, B. 2017. A well-preserved true dragonfly (Anisoptera: Gomphides: Burmagomphidae fam. nov.) from Cretaceous Burmese amber. Journal of Systematic Palaeontology [published online]. Crossref
Julián F. Petrulevičius [levicius@fcnym.unlp.edu.ar], CONICET and División Paleozoología Invertebrados, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque s/n, La Plata (1900), Argentina.
Received 22 September 2017, accepted 16 November 2017, available online 21 November 2017.
Copyright © 2017 J.F. Petrulevičius. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 62 (4): 779–783, 2017
https://doi.org/10.4202/app.00427.2017