

Expanding the geographic and geochronologic range of early pinnipeds: New specimens of Enaliarctos from Northern California and Oregon
ASHLEY W. POUST and ROBERT W. BOESSENECKER
Poust, A.W. and Boessenecker, R.W. 2018. Expanding the geographic and geochronologic range of early pinnipeds: New specimens of Enaliarctos from Northern California and Oregon. Acta Palaeontologica Polonica 63 (1): 25–40.
The early pinnipedimorph Enaliarctos was a marine-adapted carnivore with dental and locomotor features intermediate between terrestrial arctoids and living pinnipeds. New specimens of Enaliarctos are described from Oligocene and Miocene deposits on the Pacific coast of North America, and include the oldest enaliarctine mandible (Yaquina Formation, 30.6–27.4 Ma), the first enaliarctine from Northern California (Skooner Gulch Formation, 23.8–22 Ma), and the stratigraphically youngest fossil of the genus (Astoria Formation, 17.3–16.6 Ma). The wide biogeographic and temporal range of Enaliarctos provided the potential for interaction or competition with plotopterid birds, odontocete whales, and crown pinnipeds such as early odobenids, early otariids, and desmatophocids. The expansion of the known ranges of Enaliarctos species and the description of additional morphology, particularly of the mandible and lower dentition, provides insight into the origins of pinniped diversity and their possible interactions with other early Neogene coastal marine organisms.
Key words: Mammalia, Arctoidea, Pinnipedia, Enaliarctos, Miocene, Pacific.
Ashley W. Poust [ashley.poust@berkeley.edu], Department of Integrative Biology, Berkeley, California, 94720 USA; University of California Museum of Paleontology, Berkeley, California, 94720 USA.
Robert W. Boessenecker [boesseneckerrw@cofc.edu], University of California Museum of Paleontology, Berkeley, California, 94720 USA; Department of Geology, College of Charleston, South Carolina, 29424 USA.
Received 11 June 2017, accepted 4 September 2017, available online 22 December 2017.
Copyright © 2018 A.W. Poust et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pinnipeds are one of three diverse, extant clades of marine mammals and differ principally from the obligately aquatic cetaceans and sirenians in retaining the capability for terrestrial locomotion and returning to land to rest and breed. All pinnipeds evolved from a terrestrial common ancestor sometime in the Oligocene shortly after the Eocene diversification of Arctoidea (Berta et al. 1989). Enaliarctines are the geochronologically earliest pinnipedimorphs, and species of the Oligo-Miocene Enaliarctos already possessed a suite of locomotor adaptations for swimming including shortened humeri and femora and flipper-like manus and pes (Berta et al. 1989; Berta and Ray 1990) yet retained shearing carnassial teeth typical of terrestrial arctoids (Mitchell and Tedford 1973; Adam and Berta 2002; Churchill and Clementz 2016). Enaliarctines have been reported from the late Oligocene and early Miocene of the eastern North Pacific, and include Enaliarctos spp. (Chattian–Aquitanian, California and Oregon; Mitchell and Tedford 1973; Barnes 1979; Berta 1991), Pteronarctos goedertae (Barnes 1989, 1990; Berta 1994b), Pteronarctos piersoni and Pacificotaria hadromma (Barnes 1990, 1992; possible synonyms of P. goedertae according to Berta 1994b), and Pinnarctidion spp. (considered a basal phocoid according to Berta 1994a). Other aquatic arctoids, including the ursid-like molluskivore Kolponomos and the otter-like Puijila darwini, have been linked to Pinnipedimorpha (Tedford et al. 1994; Rybczynski et al. 2009).
This study reports several new specimens of Enaliarctos from California and Oregon, USA, including the oldest described mandible and lower dentition of Enaliarctos (Yaquina Formation, Oregon), the northernmost record of Enaliarctos mealsi (Skooner Gulch Formation, California), and the geochronologically youngest specimen of Enaliarctos (Burdigalian Astoria Formation, Oregon).
Institutional abbreviations.—UCMP, University of California Museum of Paleontology, Berkeley, USA; USNM, Smithsonian, Washington D.C., USA; UWBM, Burke Museum of Natural History and Culture, Seattle, USA.
Other abbreviations.—We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively. CV, cervical vertebra; TV, thoracic vertebra.
Geological setting
Skooner Gulch Formation.—The Skooner Gulch Formation crops out at a single locality along 500 meters of coastal cliffs near Gallaway, Mendocino County, California (Figs. 1C, 2B). This formation consists of 15–90 meters of massively bedded sandstone that become increasingly glauconitic towards the top of the unit. It nonconformably overlies the upper Oligocene Iversen Basalt and is conformably overlain by the lower Miocene Gallaway Formation (Phillips et al. 1976). The base of the Skooner Gulch Formation has yielded a molluskan faunule indicating correlation with the “Vaqueros” provincial megainvertebrate age and deposition in shallow (neritic) marine settings (Addicott 1967; Phillips et al. 1976). Abundant “turbidites” capped by intensely bioturbated glauconitic sandstone with phosphatic nodules indicate long periods of nondeposition between turbidity flows (Phillips et al. 1976), suggestive of submarine fan deposition (Loomis and Ingle 1995). Foraminifera from the Skooner Gulch Formation indicate a much deeper, middle bathyal setting (Phillips et al. 1976) and are indicative of Zemorrian–Saucesian benthic foraminiferal ages; Phillips et al. (1976) tentatively identified the Zemorrian–Saucesian boundary within the lower part of the overlying Gallaway Formation. According to Prothero (2001) the Zemorrian benthic foraminiferal stage is as young as 22 Ma in places (Aquitanian, early Miocene). A maximum age of 23.8 Ma is provided by a K/Ar date from the underlying Iversen Basalt (Turner 1970; Loomis and Ingle 1995). Miller (1981) notes that diagnostic Oligocene microfossils are absent in the Skooner Gulch Formation; owing to this, Barboza et al. (2017) indicate an entirely Miocene age. Therefore, the age of the Skooner Gulch Formation can be summarized as 23.03–22 Ma (earliest Miocene, Aquitanian correlative). Fossil vertebrates previously reported from the Skooner Gulch Formation include elasmobranchs (Carcharocles auriculatus, Chlamydoselachus sp., cf. C. tobleri, Centrophorus sp., Echinorhinus sp., cf. E. caspius, Galeocerdo sp., cf. G. acutus, Heterodontus sp., Hexanchus sp., cf. H. gigas, Isurus sp. A, Isurus sp. B, Megachasma applegatei, Mitsukurina sp., cf. M. maslinensis, Odontaspis sp., cf. O. ferox, Pristiophorus sp., ?Scymnodon sp.), desmostylians (Desmostylus sp., Archaeoparadoxia weltoni), cetaceans (cf. Argyrocetus, cf. Waipatiidae indet.), and the Enaliarctos mandible and metacarpal described herein (Addicott 1967; Phillips et al. 1976; Barnes 1979; Clark 1991; Shimada et al. 2014). The newly reported specimens were collected from UCMP locality V-75135, a glauconitic horizon in the upper part of the Skooner Gulch Formation, for which an age control of 23.03–22 Ma can be assigned (see above).
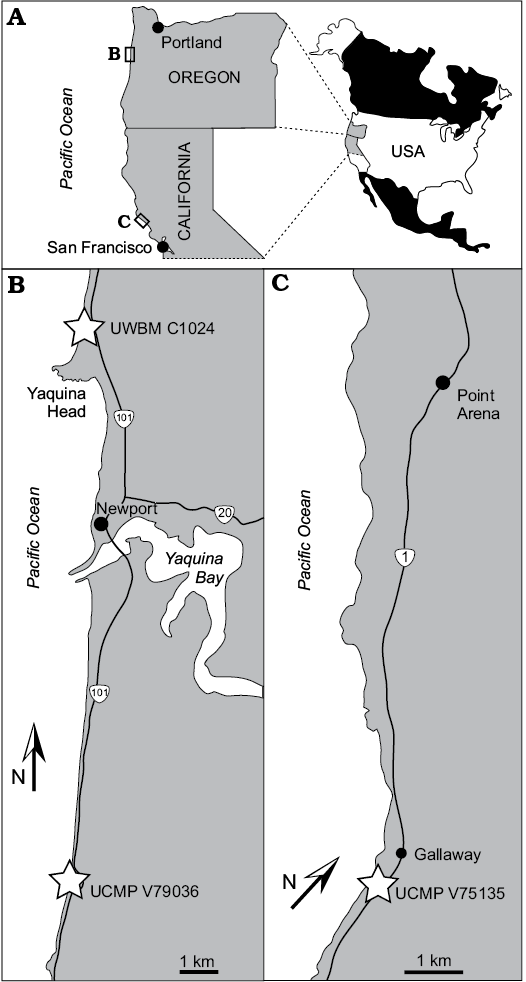
Fig. 1. Locality map for fossils of enaliarctines. Map of Oregon and California (A), the coastline near Newport, Oregon (B), the coastline near Point Arena, California (C).
Yaquina Formation.—The Yaquina Formation (Figs. 1B, 2A) crops out along the eastern shore of Yaquina Bay near Newport, Oregon, and along the coast between Seal Rock and Beaver Creek approximately 20 km south of Newport, Oregon. This formation consists of up to 700 m of fine-coarse cross-bedded sandstone with abundant carbonized wood and lenses of pebbly sandstone, tuffaceous siltstone, and coal (Snavely et al. 1976; Prothero et al. 2001b). It conformably overlies the lower Oligocene Alsea Formation and is in turn conformably overlain by the lower Miocene Nye Mudstone. Sedimentology and benthic foraminiferal assemblages indicate shallow marine shelf deposition, possibly deltaic in origin (Prothero et al. 2001b). Benthic foraminifera reported by Rau (1981) indicate a Zemorrian age (33–22 Ma; Prothero et al. 2001b) whereas mollusk assemblages indicate correlation with the Juanian provincial megainvertebrate stage (28.5–24 Ma; Prothero et al. 2001b) of the Pacific Northwest and the Vaqueros stage (30–19 Ma; Prothero et al. 2001b) of California (Addicott 1967). Fossil vertebrates reported from the Yaquina Formation include a megamouth shark (Megachasma applegatei), the enaliarctine pinniped Enaliarctos tedfordi, toothed mysticetes (Aetiocetus cotylalveus, Aetiocetus weltoni), and several desmostylians (Behemotops proteus, Cornwallius sookensis, Seuku emlongi; Emlong 1966; Domning et al. 1986; Berta 1991; Barnes et al. 1995; Deméré and Berta 2008; Shimada et al. 2014). The newly reported specimen (UCMP 253400) was collected as float within a boulder of sandstone matching the lithology of the Yaquina Formation. Though assignment to a specific stratigraphic interval within the Yaquina Formation is not possible, a recent paleomagnetic study by Prothero et al. (2001b) indicates that the type section of the Yaquina Formation to the Northeast at Yaquina Bay is older than considered by Armentrout (1981) and Deméré and Berta (2008), and ranging in age from 30.6–27.4 Ma (late Rupelian to early Chattian; chrons C12n–C9r; dates from Ogg 2012). An age older than Chattian is unlikely for the reported specimen given that only the uppermost few meters of the Yaquina Formation is exposed at these localities (Deméré and Berta 2008), but we retain the full range reported by Prothero et al. (2001b) given the lack of stratigraphic control and occurrence of UCMP 253400 within a loose block.
Astoria Formation.—The Astoria Formation crops out along the banks of the Columbia River near Astoria, Oregon, and along coastal exposures in Lincoln County near Newport, Oregon (Figs. 1B, 2B). Within the Newport embayment in coastal Oregon, the Astoria Formation consists of 150 m of massively bedded sandstone and mudrocks disconformably overlying the lower Miocene Nye Mudstone and is unconformably overlain by Pleistocene terrace deposits (Snavely et al. 1976). Mollusk assemblages suggest shelf deposition below storm weather wave base and as deep as 100 m (Moore 1963; Colbath 1985). A diverse marine vertebrate assemblage has been documented from the Astoria Formation near Newport including sharks (Hexanchus, Squalus, Pristiophorus, Squatina, Odontaspis, Carcharodon hastalis, Isurus planus, Cetorhinus piersoni, Carcharocles megalodon, Myliobatis; Welton 1972, 2015), birds (Pelagornis sp., Olson 1985), pinnipeds (Pteronarctos goedertae, Pteronarctos “piersoni”, Pacificotaria hadromma, Proneotherium repenningi, Desmatophoca oregonensis, “Allodesminae” indet.; Condon 1906; Barnes 1989, 1990, 1992; Kohno et al. 1995; Deméré and Berta 2001), cetaceans (Squalodontidae indet., Cophocetus oregonensis; Dooley 1994; Packard and Kellogg 1936), desmostylians (Desmostylus hesperus; Hannibal 1922), and even land mammals (Aphelops sp., Tylocephalonyx sp.; Mitchell and Repenning 1963; Munthe and Coombs 1979).
UWBM 89114 was collected as an isolated concretion from Moloch Beach and derived from the Iron Mountain Bed, a concretionary horizon within the upper part of the Astoria Formation which yielded the holotype and paratype specimens of the pinnipedimorphs Pteronarctos “piersoni”, Pacificotaria hadromma, and Proneotherium repenningi (Barnes 1990, 1992; Deméré and Berta 2001; Prothero et al. 2001a). Based on a combination of foraminiferal, molluscan, and land mammal biostratigraphy, Barnes (1990) estimated the Iron Mountain Bed to be approximately 16 Ma in age. Deméré and Berta (2001) adopted a more skeptical approach and considered the Astoria Formation (and the Iron Mountain Bed) to be 20–15 Ma. Paleomagnetic studies by Prothero et al. (2001a) vindicated both age assessments, providing a range of 20.7–15.1 Ma for the Astoria formation within the Newport embayment and a finer range of 17.3–16.6 Ma (Burdigalian, late early Miocene) for the Iron Mountain Bed.
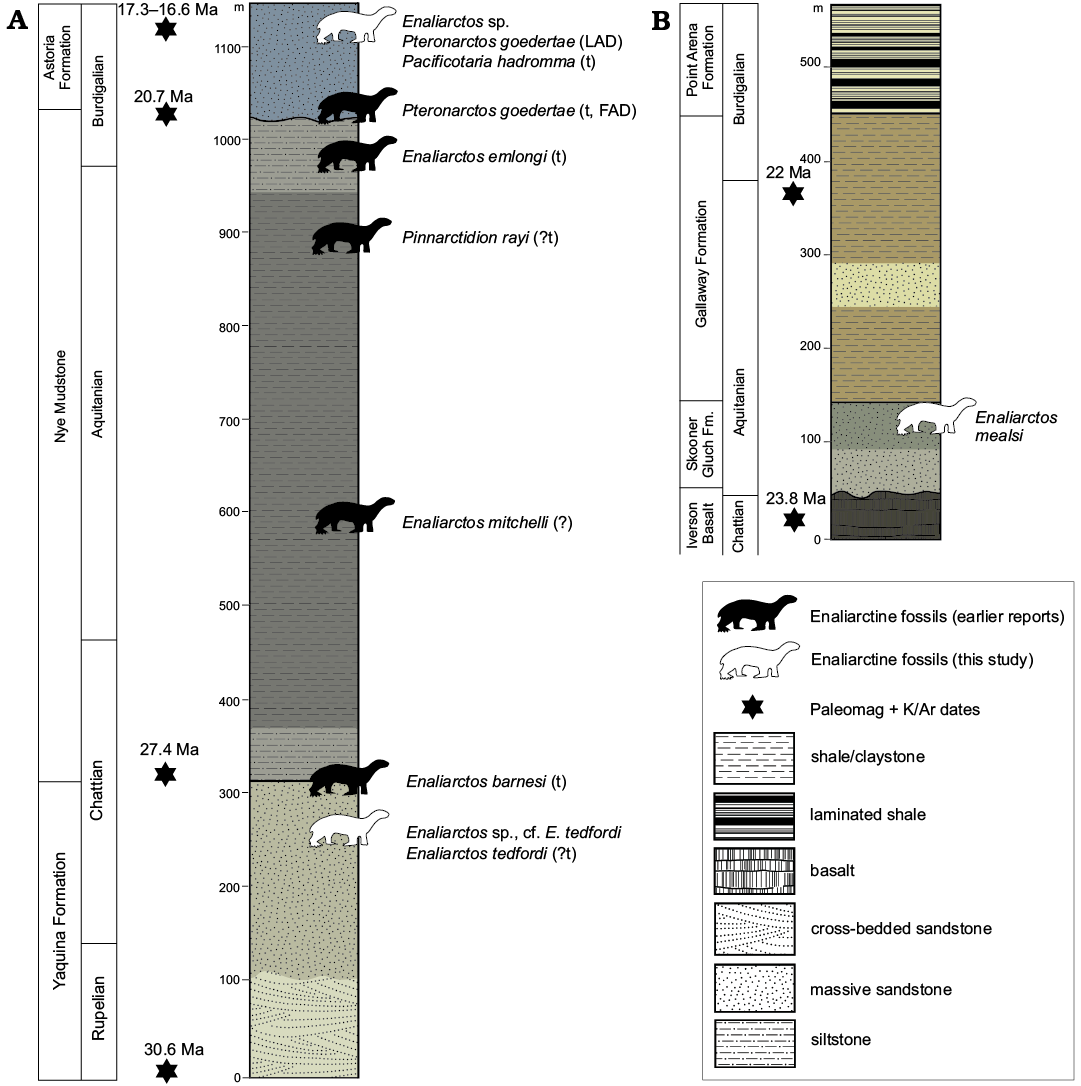
Fig. 2. Stratigraphic columns of Oregon (A) and California (B) localities. Columns based on Snavely et al. (1964) and Phillips et al. (1976); paleomagnetic and K/Ar dates from Prothero et al. (2001a, b) and Phillips et al. (1976). In the interest of clarity non-enaliarctine marine carnivores are omitted. Modified from Moore (1963) and Phillips et al. (1976). Abbreviations: FAD, first appearance datum; LAD, last appearance datum; t, type locality/horizon.
Systematic palaeontology
Mammalia Linnaeus, 1758
Carnivora Bowditch, 1821
Pinnipedimorpha Berta, Ray, and Wyss 1989
Genus Enaliarctos Mitchell and Tedford, 1973
Type species: Enaliarctos mealsi Mitchell and Tedford, 1973; Pyramid Hill, California, latest Oligocene to earliest Miocene.
Diagnosis.—A small pinnipedimorph different from all others in possessing the following combination of plesiomorphic characters: deep embrasure pit present between P4 and M1; P4–M2 with multiple roots; P4 with protocone shelf; C1 with a posterolingual bulge; m1 with well-developed hypoconid and trigonid cusps around talonid; six lumbar vertebrae; ulna lacking posteriorly extended olecranon process; radius lacking a strongly flattened and expanded distal end; fifth intermediate manual phalanx unreduced; teres femoris ligament pit present on femur; metapodials with cylindrical diaphysis, keeled heads, and strongly trochleated phalangeal articulations (modified from Berta 1991).
Enaliarctos mealsi Mitchell and Tedford, 1973
Figs. 3, 4; Tables 1, 2.
Material.—UCMP 114474, isolated left mandible (collected by Bruce J. Welton, October 6, 1975); UCMP 276804, partial metacarpal (collected by RWB and Richard Hilton, July 21, 2015); from Skooner Gulch I, Mendocino County, California (UCMP locality V75135); Skooner Gulch Formation, early Miocene (23.03–22 Ma).
Description.—UCMP 114474 includes the anterior left mandible, missing the posterior end behind the m2 alveolus (Fig. 3). The preserved portion of the horizontal ramus is rectangular, deepening only at the root of the canine located ventral of the p2. The anteroventral margin is broken and the genial tuberosity is missing. Two large (4.0 mm and 2.0 mm) mental foramina are positioned on the middle of the mandible below p2 and p3, respectively. The dorsal margin is flat except where the alveolar rims rise above this surface.
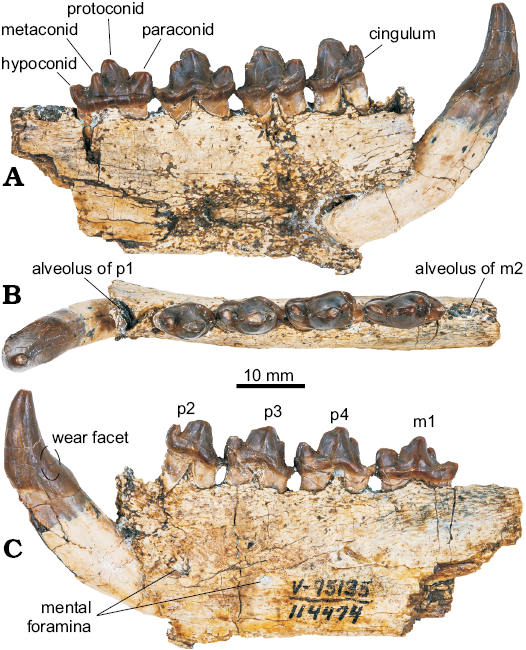
Fig. 3. Mandible of pinniped mammal Enaliarctos mealsi (UCMP 114474) from Schooner Gulch, Mendocino County, California, earliest Miocene; in lingual (A), occlusal (B), and labial (C) views.
Dentition: The anterior portion of UCMP 114474 is missing, including the incisors. The canine is in place despite lacking much of the bone surrounding the root. It is relatively small and dorsoventrally short with mild posterior curvature. A shallow buccal wear facet is seen on the posterior bulge of the crown as in some other carnivores. The canine bears two cristae just lingual of its posterior edge. These converge slightly towards the tip, but do not meet.
All preserved postcanine teeth are double-rooted, but alveoli for p1 and m2 suggest these teeth were single-rooted. The p2 has a high but anteroposteriorly narrow crown with a large, distinct, nearly recurved, conical paraconid, no clear metaconid, and a low, crest-like hypoconid encircling a shallow posteromedially placed talonid basin. The p3 crown bears a larger, robust and conical paraconid, a high protoconid, a small metaconid, and a small hypoconid positioned posteriorly on a crest defining a somewhat larger talonid basin. The p4 continues the trend of enlarging the paraconid, possesses a high, pointed protoconid, and a large metaconid; the hypoconid resembles that of p3 but encircles a more deeply excavated talonid basin. This p4 talonid basin is separated into posterior and posteromedial portions by the base of the metaconid, which drops precipitously into the basin. In dorsal view the protoconid of p4 and m1 have two distinct wear surfaces separated by a narrow ridge. The metaconids of p3–m1 are high and positioned apically near the protoconid. All premolars exhibit moderately developed labial cingula and smaller, but distinct lingual cingula.
The m1 has a dorsoventrally lower crown with a lower, sharper protoconid than in the premolars. The metaconid is conical but apically rounded and offset lingually from the protoconid. The hypoconid is large, labiolingually broad and more apically prominent than in the premolars. No diastemata are present in the postcanines.
Metacarpal: UCMP 276804 is a partial distal metacarpal of uncertain position (Fig. 4). Proximally the cross-section is nearly circular. The distal end is somewhat hemispherical and bears a slight keel; it is rounded in dorsal view, differing from the flattened joint in extant pinnipeds. The keel protrudes strongly on the palmar side, suggesting a wide range of motion in the foreflipper of Enaliarctos, perhaps relating to prey acquisition and processing (e.g., Hocking et al. 2016).
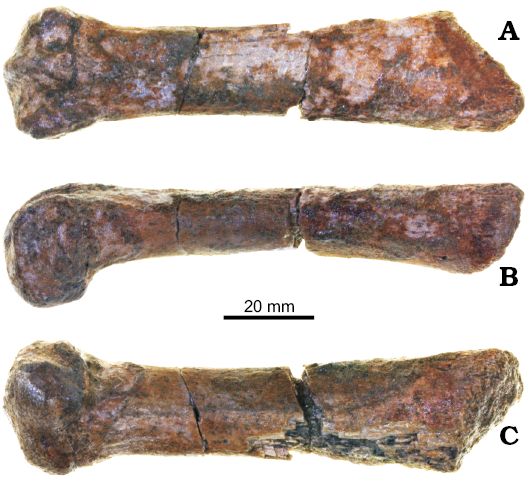
Fig. 4. Metacarpal of pinniped mammal Enaliarctos mealsi Mitchell and Tedford, 1973 (UCMP 276804) from Schooner Gulch, Mendocino County, California, earliest Miocene; in dorsal (A), lateral (B), and ventral/palmar (C) views.
Remarks.—The mandible, UCMP 114474, is identifiable as Enaliarctos in exhibiting a lingual bulge on the base of the canine crown, postcanine teeth with apically positioned, strongly developed metaconid cusps and trenchant paraconid cusps. Advanced wear of the cusps suggests this specimen represents an adult, and a relatively narrow canine in comparison with other enaliarctines suggests that UCMP 114474 is a female (Cullen et al. 2014). UCMP 114474 differs from Enaliarctos emlongi in exhibiting well-developed labial cingulum and cuspate posterolingual cingulum, and shares these features with E. mealsi and E. barnesi. UCMP 114474 differs from E. barnesi in exhibiting a deeper notch between the paraconid and protoconid on the m1, a lower m1 protoconid, and more apically prominent metaconids on p3–p4. These differences are not noted in comparison with isolated lower teeth referred to E. mealsi (Mitchell and Tedford 1973; Barnes 1979), the presumed p3–p4 and m1 of which are indistinguishable from UCMP 114474. Owing to these shared features (prominent metaconids, extensive labial and lingual cingulum, trenchant paraconids, low protoconid on m1 with deeper notch anteriorly) we refer UCMP 114474 to Enaliarctos mealsi. We note that E. barnesi and E. mealsi may be synonymous (see below) and another possibility may be that these dental features are attributable to sexual dimorphism, as the E. barnesi holotype is a presumed male (Cullen et al. 2014) and UCMP 114474 is a presumed female (see above). The metacarpal, UCMP 276804, is tentatively referred as Enaliarctos sp., cf. E. mealsi owing to its cylindrical diaphysis and strongly keeled distal trochlea (Berta 1991); metapodials are unknown for other enaliarctines (Pacificotaria, Pinnarctidion, Pteronarctos) or the enliarctine-like basal odobenid Proneotherium, rendering this referral provisional.
Stratigraphic and geographic range.—Skooner Gulch Formation, Schooner Gulch, Mendocino County, California; earliest Miocene.
Table 1. Measurements (in mm) of Enaliarctos sp. mandibles.
|
Measurement |
UWBM |
UCMP 253400 |
UCMP 114474 |
|
|
left |
right |
right |
left |
|
|
Total length
|
100.7 |
85.98 |
58.44 |
72.15 |
|
Length postcanine tooth row |
46.84 |
46.87 |
>45.19 |
>56.70 |
|
Anterior tip to base of coronoid |
58 |
– |
– |
– |
|
Greatest length |
– |
25.13 |
23.29 |
– |
|
Mandibular depth at: |
||||
|
c1 |
14.14 |
14.06 |
20.41 |
23.97 |
|
p1 |
14.64 |
17.38 |
22.20 |
24.06 |
|
p2 |
17.74 |
18.76 |
23.54 |
22.35 |
|
p3 |
16.06 |
– |
22.25 |
20.36 |
|
p4 |
16.44 |
– |
– |
21.32 |
|
m1 |
16.95 |
– |
– |
22.03 |
|
m2 |
16.71 |
– |
– |
– |
|
Length of toothrow c1–m1 |
50.53 |
53.4 |
57.22 |
51.70 |
|
Length m1 to base of coronoid |
14 |
– |
– |
– |
|
Anteroposterior length/dorsoventral depth: |
||||
|
i2 |
–/– |
3.81/2.54 |
–/– |
–/– |
|
i3 |
–/3.51 |
4.26/2.61 |
–/– |
–/– |
|
c1 |
7.29/11.55 |
–/– |
9.28/13.10 |
10.57/14.99 |
|
p1 |
6.54/4.91 |
5.71/– |
–/– |
–/– |
|
p2 |
7.9/4.39 |
–/– |
9.53/5.88 |
9.23/5.68 |
|
p3 |
7.91/6.36 |
–/– |
10.93/8.75 |
10.41/8.23 |
|
p4 |
8.14/5.35 |
–/– |
10.83/7.94 |
11.52/7.44 |
|
m1 |
9.16/4.25 |
–/– |
–/– |
–/– |
Table 2. Measurements (in mm) of Enaliarctos sp. postcrania. CV, cervical vertebra.
|
Measurement |
UWBM 89114 |
UCMP 253400 |
UCMP 276804 |
||||
|
atlas |
axis |
CV6 |
CV7 |
posterior thoracic |
CV6 or CV7 |
metacarpal |
|
|
Transverse width anterior facets |
45.28 |
– |
– |
– |
– |
– |
– |
|
Total height |
34.74 |
49.65 |
– |
60.26 |
59.01 |
– |
– |
|
Height neural foramen |
18.75 |
– |
10.2 |
9.0 |
8.32 |
13.79 |
– |
|
Transverse width neural foramen |
17.6 |
18.96 |
15.4 |
17.47 |
15.69 |
22.74 |
– |
|
Anteroposterior length neural spine |
– |
52.27 |
– |
– |
– |
– |
– |
|
Greatest width across transverse processes |
72.4 |
34.86 |
57.2 |
62.48 |
25.94 |
80.3 |
– |
|
Height of anterior articular surface |
17.46 |
14.9 |
16.06 |
– |
17.81 |
– |
– |
|
Transverse width anterior articular surface |
– |
31.2 |
18.09 |
23.5 |
19.19 |
– |
– |
|
Height of posterior articular surface |
– |
15 |
14.27 |
16 |
– |
17.5 |
– |
|
Transverse width posterior articular surface |
– |
18.5 |
19.86 |
22.3 |
– |
16.7 |
– |
|
Centrum length |
– |
43.13 |
25 |
32.33 |
29.58 |
36.04 |
– |
|
Midshaft dorsoventral height |
– |
– |
– |
– |
– |
– |
8.04 |
|
Midshaft transverse width |
– |
– |
– |
– |
– |
– |
10.63 |
|
Distal (head) dorsoventral height |
– |
– |
– |
– |
– |
– |
15.21 |
|
Distal (head) transverse width |
– |
– |
– |
– |
– |
– |
13.90 |
Enaliarctos sp., cf. E. tedfordi Berta, 1991
Fig. 5; Tables 1, 2.
Material.—UCMP 253400, associated right mandible, thoracic vertebra, and two ribs (collected by David G. Taylor, October 6, 1963); from Ona Beach, Lincoln County, Oregon (UCMP locality V79036); collected from a loose concretion of the Yaquina Formation, late Oligocene (30.6–27.4 Ma, late Rupelian–early Chattian equivalent; Prothero et al. 2001b). Detailed locality information available on request to qualified researchers from UCMP or the authors. The block containing UCMP 253400 remained unprepared and undescribed for over fifty years (Fig. 5A). The bones are preserved out of articulation but in contact with one another along a single plane in the block. The matrix is a well-sorted, moderately fine-grained sandstone. Immediately surrounding the bones the sandstone is iron-stained and more friable than the remainder of the matrix. In addition to the mammalian bones, the sandstone block includes fish vertebrae, scales, and bone fragments and some lignitized plant material. In places, the sandstone is composed of masses of small peloids with a radius of about a millimetre.
Description.—Mandible: UCMP 253400 includes the anterior right mandible, missing the posterior end behind the p4 (Fig. 5). The preserved portion of the horizontal ramus is subrectangular and relatively dorsoventrally shallow. A strongly expressed genial tuberosity occurs below p1–p2 and marks the deepest point on the mandible. Two large mental foramina are positioned below p2 (3.8 mm) and p3 (3.1 mm), similar to those of the other enaliarctine mandibles described. The symphyseal surface is poorly preserved but appears to have been broadly lanceolate.
The alveolar margin is linear, though the p2, p3, and p4 are consistently higher with more root exposed; it is unclear if these teeth were slightly displaced out of their alveoli during decomposition and prior to burial. Posterior to the p4 the mandible is not preserved, but the impression of the m1 (see Fig. 5) shows that this trend was not continued in the molars, suggesting postmortem displacement of the anterior teeth.
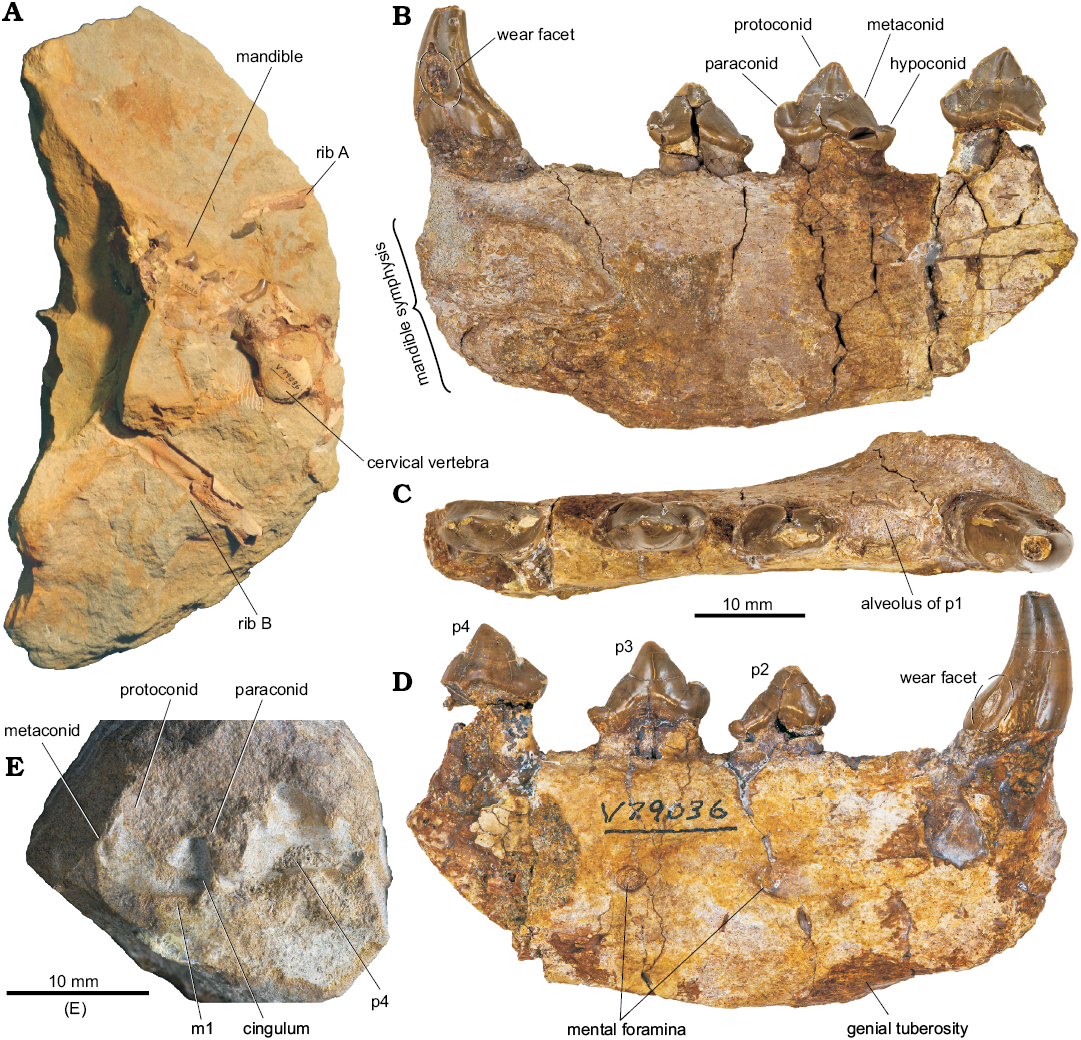
Fig. 5. Elements of pinniped mammal Enaliarctos sp., cf. E. tedfordi Berta, 1991 (UCMP 253400) from Ona Beach, Lincoln County, Oregon, Yaquina Formation, early late Oligocene; overview of original block (A), lingual (B), occlusal (C), and labial (D) views; matrix preserving impression of lingual side of p4 and m1 (E).
Dentition: No incisors are preserved in UCMP 253400. The canine is relatively small, dorsoventrally short, and curved posteriorly. The canine exhibits a posterior bulge on the base of the crown, the buccal side of which has a deep, oval wear facet from the upper canine that has penetrated the enamel. Above this bulge on the lingual side is an upward-tapering wear facet, formed from occlusion with I3. The c1 bears a double posterior crista. Towards the root, the ancillary ridge deflects medially, dividing the postero-medial half of the tooth crown into three triangular faces.
The alveolus of the p1 indicates that the tooth was single-rooted. The remaining premolars are double-rooted. The p2 has a large crown with a small conical paraconid, a very small metaconid on the mid-crown, and a low but well-defined hypoconid. The p3 crown bears a larger, more rounded paraconid, a high protoconid, and a metaconid elevated to mid-crown height; the hypoconid is a low cusp situated at the posterior terminus of the lingual cingulum which encircles a shallow talonid basin. The p4 has a larger conical paraconid, a high, sharp protoconid, and a well-developed metaconid, more distinct from the principal cusp than in p3; the hypoconid is broken, but the impression shows it was a very low, conical cusp. All premolars are labially smooth and lack labial cingulum, but possess well developed smooth lingual cingulum.
The m1 is lost, but an impression of the lingual surface remains in the sandstone block (Fig. 5E). The mold shows an anteroposteriorly elongate crown with a low protoconid cusp and a high trenchant paraconid. The protoconid has a similar height to p4, with a distinct, rounded metaconid; an impression of the hypoconid is not preserved. The diastema between p3 and p4 is particularly large at 5.5 mm, whereas no diastema in UCMP 114474 or UWBM 89114 exceeds 2 mm.
Posterior cervical vertebra: The convex, oval anterior articular surface of the centrum sits below a large dorsoventerally flattened neural canal (Fig. 5A). The laminae are flat, broad, and inclined slightly anteriorly. Gracile transverse processes extend from the anterior surface of the pedicles, tilting anterolaterally at approximately a 45° from the dorsal plane. The blades of the transverse processes bifurcate into a “T”-shape with a flat lateral edge. Prezygapophyses are not preserved; the postzygapophyses are broad, arranged at an angle greater than 45° from the sagittal plane, and have facets facing ventrolateral, more or less in line with the wings of the transverse processes. The neural spine is broken, but appears to have been low and posteriorly inclined. The posterior articular surface of the centrum is raised slightly dorsally relative to the anterior face to allow increased axial flexion.
Ribs: The two partial ribs of UCMP 253400 are short segments of the distal and mid-shafts respectively; positions are unclear. Rib A was preserved beneath the mandible (see Fig. 5A) and extends from a somewhat flattened mid-shaft to a distal end more circular in cross-section with rough, porous cortical bone where the element met costal cartilage. Rib B is very incomplete, and is represented in part by a natural mold of the internal surface of the mid-shaft showing a very distinct, anterior costal groove.
Comparisons.—UCMP 253400 shares with Enaliarctos emlongi smooth labial crown surfaces further lacking labial cingula, relatively small paraconid cusps, and low metaconid cusps. UCMP 253400 is from the Oligocene Yaquina Formation (28.1–27.4 Ma) and therefore much older than Enaliarctos emlongi from uppermost Nye Mudstone or lowermost Astoria Formation (20.2–19.1 Ma); owing to this age difference, UCMP 253400 is probably not referable to E. emlongi. However, E. tedfordi, which is not known from a mandible or lower dentition, shares loss of the labial cingulum and reduction of the lingual cingulum of the upper dentition with E. emlongi. Enaliarctos tedfordi originates from the Yaquina Formation, and assuming that the lower dentition parallels the upper dentition as in E. emlongi, UCMP 253400 is tentatively referable to E. tedfordi.
Enaliarctos sp.
Figs. 6, 7; Tables 1, 2.
Material.—UWBM 89114, a partial skeleton exposed in relief in a small concretion including left and right mandibles with i2–i3, c1, p1–p4, m1, atlas, axis, CV6, CV7, TV1, several other thoracic vertebrae, one rib, and parts of three sternebrae (Collected by Jason Love in March 2004, from UWBM locality C1024); from Moloch Beach, Lincoln County, Oregon, USA, Iron Mountain Bed, Astoria Formation, late early Miocene (17.3–16.6 Ma; Burdigalian equivalent). Detailed locality information available on request to qualified researchers from UWBM or the authors.
Description.—Mandible: UWBM 89114 includes both mandibles, with the right mandible missing the posterior end and worn down below the alveolar margin posterior to the p2 (Figs. 6A, 7). Minimal wear of the teeth argues against advanced age, but the postcanine teeth are not crowded as in juveniles, suggesting subadult or possibly adult status; the canine is relatively large and similar in proportion to the presumed male E. emlongi holotype, suggesting that UWBM 89114 is a male. The left mandible is nearly complete and is missing only the coronoid process, mandibular condyle, and angular process. The medial surface of the right mandible and lateral surface of the left mandible are exposed. The horizontal ramus of the mandible is subrectangular, relatively shallow dorsoventrally, and is deepest posteriorly at the level of the p3. Three mental foramina are positioned on the middle of the mandible below p2–p3. The symphyseal surface is rugose and is lenticular in shape; no genial tuberosity is developed, but a delicate ventral crest is present posteroventral to the symphysis.
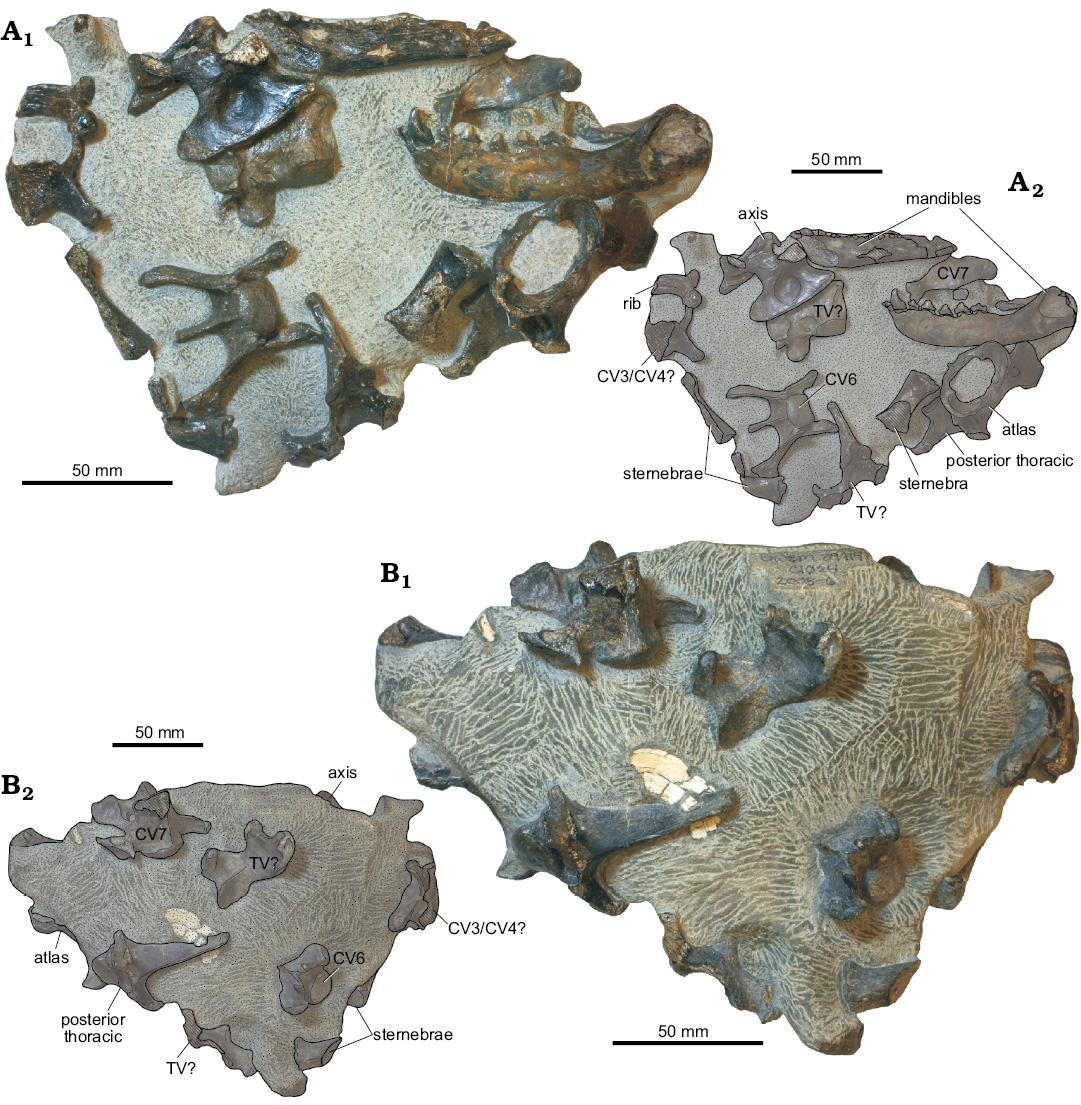
Fig. 6. Mandibles and vertebrae of pinniped mammal Enaliarctos sp. (UWBM 89114) from Moolack Beach, Lincoln County, Oregon, Astoria Formation, late early Miocene. Two sides of the block (A, B). Photographs (A1, B1), explanatory drawings (A2, B2). Abbreviations: CV, cervical vertebra; TV, thoracic vertebra.
The alveolar margin is undulatory and descends ventrally at the level of the p3, giving the toothrow a somewhat dorsally concave profile in lateral view. the alveolar margin rises toward the m1 and descends again to give the margin anterior to the coronoid process a dorsally concave profile. The masseteric fossa is shallow and does not extend ventral to the toothrow.
Dentition: Both i2 and i3 are preserved and exhibit transversely narrow roots with slightly wider crowns bearing apical transverse grooves (Fig. 7A2, C). The canine is relatively small, transversely wide but dorsoventrally short and posteriorly curving. It exhibits a posterior bulge on the base of the crown, and bears a posterior crista, mesial crista, and lingual cingulum. A small wear facet from the upper canine is present labially on the base of the crown as in Enaliarctos sp., cf. E. tedfordi (UCMP 253400, see above).

Fig. 7. Mandibles and vertebrae of pinniped mammal Enaliarctos sp. (UWBM 89114) from Moolack Beach, Lincoln County, Oregon, Astoria Formation, late early Miocene. A. Left mandible in labial view; photograph (A1), explanatory drawing (A2). B. Left mandible in occlusal view. C. Right mandible in occlusal view.
The p1 is single-rooted and bears a small crown with an indistinct paraconid (= anterior accessory cusp), a low protoconid (= principal cusp), and a low hypoconid (= posterior accessory cusp); the metaconid is absent. As in other enaliarctines the p2–p4 are double-rooted. The p2 bears a somewhat larger crown with an indistinct paraconid, a small metaconid, and a small hypoconid. The p3 crown is lower than the p2 owing to the “dip” in the alveolar margin of the mandible and bears a small conical paraconid, a high protoconid, and a metaconid elevated to mid-crown height; the hypoconid is obscured by matrix. The p4 has a larger conical paraconid, a high but apically worn protoconid, and a well-developed metaconid like p3; the hypoconid is low and conical. All premolar crowns are labially smooth and lack a labial cingulum.
The m1 is double-rooted and bears a low, anteroposteriorly elongate crown with a low, trenchant paraconid, a low protoconid, an indistinct metaconid, and a conical and low hypoconid. The m2 is absent but a small partially prepared alveolus indicates a single circular root. All lower postcanines are tightly spaced and no diastemata are apparent.
Atlas: The anterior condylar facets are concave, transversely narrow ventrally and broad and widely separated dorsally (Fig. 6A). A low neural spine is present. The transverse process is subrectangular and posteroventrally oriented in lateral view, approximately 15–20° from vertical. The transverse process is pierced by a lateral vertebral canal positioned medially within a fossa on the anterior surface.
Axis: The axis bears a well developed conical odontoid process with a convex and transversely narrow atlantal facet. The centrum bears a longitudinal ridge dorsally within the neural foramen and a deep ridge ventrally, flanked by deep ventrolateral fossae. A secondary fossa is present on the posterolateral surface below the pedicle of the neural arch, separated by the ventrolateral fossa by a thin ridge. The neural spine is large, falcate, and extends anteriorly to the level of the odontoid apex in lateral view; the neural spine rises posteriorly towards its dorsal apex. A small tongue-shaped postzygapophysis is present. A small, posterolaterally projecting transverse process is developed.
Third? or fourth? cervical vertebra: One incompletely preserved vertebra exhibits a circular lateral vertebral canal larger than in CV6; it bears a low subrectangular neural spine with an apical tubercle and a small fan-shaped transverse process that is less elongate than in CV6. Owing to the short transverse process this vertebra represents an anterior cervical.
Sixth cervical vertebra: A small, rectangular anterior articular surface of the centrum is flanked by small oval lateral vertebral canals (Fig. 6B). The transverse process is elongate, ventrolaterally projecting and bears a sharp anterior crest. Small triangular prongs are present anteriorly at the level of the ventral margin of the centrum; the ventrolateral apex of the transverse process is bifurcated. A low neural spine is present; spoon-like prezygapophyses are present and have dorsomedially facing articular surfaces approximately 45° from the sagittal plane, whereas the postzygapophyses have subhorizontal facets with small medial tubercles.
Seventh cervical vertebra: The CV7 differs from the CV6 in having a taller neural spine and a short, transversely oriented transverse process with a laterally deepening, fan-like shape. The posterior articular surface of the centrum is oval-shaped. The neural spine bears an apical tubercle for the nuchal ligament.
Thoracic vertebrae: The TV1 is similar to the CV7 but has a further dorsally positioned transverse process with a concave facet for the first rib and a dorsal spur, and a slightly taller neural spine. Two other thoracic vertebrae of uncertain position are preserved. One exhibits a more cylindrical centrum than the other vertebrae, while the second consists only of a neural arch and spine. Both show a posteriorly inclined neural spine lacking an apical tubercle, and horizontal postzygapophyses on the posterior part of the neural arch.
Ribs: The proximal end of one rib is preserved and includes a bulbous, spherical capitulum, a short neck, and a flattened tubercle. A low lateral ridge is present anteriorly.
Sternebrae: Parts of three sternebrae are present (Fig. 6A, B). One is complete and subcylindrical in overall form and exhibits a rugose and quadrate articular surface for the next sternebra; it is transversely and dorsoventrally narrow or “pinched” in the middle.
Remarks.—UWBM 89114 differs from all other enaliarctines for which a mandible is known (Enaliarctos barnesi, Enaliarctos emlongi, Enaliarctos mealsi, Pteronarctos goedertae) in its smaller absolute size, mandible with sinuous alveolar margin, and a masseteric fossa not extending ventral to the toothrow, and further differs from E. barnesi and E. mealsi in lacking labial cingulum (which it shares with E. emlongi). UWBM 89114 is identifiable as Enaliarctos based on the presence of a lingual bulge on the base of the canine crown (shared with Potamotherium, Puijila, and other arctoid “fissipeds”), differing from all pinnipediformes; it shares this feature with all Enaliarctos spp. for which a lower canine is known. UWBM 89114 further differs from E. emlongi in possessing somewhat more inflated protoconid cusps on p2–p4, more basally positioned metaconids on p2–m1, and a lower protoconid on m1. UWBM 89114 shares similar small size with E. mitchelli, and has a postcanine toothrow length of 45 mm, which is somewhat smaller than the upper postcanine toothrow length of E. mitchelli (56–59 mm); in E. emlongi, it is approximately 70 mm, and in E. barnesi it measures 59 mm (measurements approximated from Berta 1991: tables 2 and 3). Within Enaliarctos, loss of upper postcanine labial cingulum is shared by E. emlongi, E. mitchelli, and E. tedfordi, with E. emlongi showing loss of the labial cingulum in both the upper and lower dentition (Berta 1991). With the exception of Enaliarctos mealsi (Berta and Ray 1990), postcrania are sporadically reported amongst enaliarctine pinnipeds. The transverse process of the atlas is subrectangular, unlike the rounded margin in Pteronarctos (Berta 1994b). The axis shares an anteroposteriorly elongate, fan-shaped neural spine with E. mealsi. Thoracic vertebrae, ribs, and sternebrae do not differ from E. mealsi except in size. Body length estimation using mandibular equations from Churchill et al. (2015) consistently overestimated the body size (2–3 m) of Enaliarctos sp. (UWBM 89114). Mandibular measurements are approximately 67% the size of E. emlongi, which Churchill et al. (2015) estimated at 1.7 m in length; simple scaling suggests that Enaliarctos sp. was 1.1 m in length, similar in length to the extant Baikal seal (Pusa sibirica) and rivalling Eotaria spp. as the smallest pinnipedimorph of all time. Though UWBM 89114 is much younger chronologically than all reported species of Enaliarctos, its small size may suggest affinities with E. mitchelli, the youngest named species of Enaliarctos, recorded from the approximate level of the Nye Mudstone/Astoria Formation contact (Berta 1994b), and dated to 24–19.1 Ma (Prothero et al. 2001a). Owing to the stratigraphic separation between UWBM 89114 and the referred cranium of E. mitchelli from the Nye/Astoria contact, we conservatively identify this specimen as Enaliarctos sp.
Concluding remarks
Geographic and stratigraphic distribution of Enaliarctos.—Enaliarctines (Enaliarctos, Pacificotaria, Pinnarctidion, Pteronarctos) have been named only from the late Oligocene and early Miocene of California and Oregon, though the published record is already quite densely sampled (Mitchell and Tedford 1973; Barnes 1979, 1989, 1990, 1992; Berta 1991, 1994a, b; Berta et al. 1989; Cullen et al. 2014; this study); unnamed enaliarctines are also known from the Pysht Formation of Washington (Hunt and Barnes 1994). Enaliarctos in particular is now reported from the Jewett Sand at Pyramid Hill, California, the Skooner Gulch Formation in Mendocino County, California, and the Yaquina Formation, Nye Mudstone, and Astoria Formation of the Newport Embayment in Lincoln County, Oregon (Table 3). This study critically reports Enaliarctos sp. from the “Iron Mountain Bed” of the Astoria Formation (17.4–16.6 Ma; Prothero et al. 2001a), extending the range of the genus into the late Burdigalian stage (Fig. 8). This new record clearly indicates that Enaliarctos geochronologically overlapped with not only other more derived enaliarctines (Pteronarctos goedertae, Pacificotaria hadromma, Pinnarctidion rayi; Barnes 1989, 1990, 1992; Berta 1994a, b) but also the earliest odobenids (Proneotherium repenningi, Pelagiarctos; Kohno et al. 1995; Deméré and Berta 2001; Boessenecker and Churchill 2013) and the earliest otariids (Eotaria; Boessenecker and Churchill 2015). Early otariids and small imagotariines were enaliarctine-like in craniodental morphology (Kohno et al. 1995; Boessenecker and Churchill 2015; Velez-Juarbe 2017) and no obvious differences in feeding ecology among these groups are apparent, suggesting unexplained diversity in the eastern North Pacific during the late early Miocene. The increased temporal overlap of these clades suggests that further analysis, such as functional diversity modelling (e.g., Novack-Gottshall 2007; Dineen et al. 2014) or various specimen-based approaches (e.g., Kloess and Parham 2017), may inform our understanding of their respective diversifications.
Table 3. Fossil occurrences and geochronologic age control for enaliarctine pinnipedimorphs.
|
Taxon |
Formation |
Age (Ma) |
References |
|
|
Age |
Record |
|||
|
Enaliarctos barnesi |
Uppermost Yaquina Formation or lower Nye Mudstone |
28.1–26.4 |
||
|
Enaliarctos emlongi |
Uppermost Nye Mudstone or lowermost Astoria Formation |
20.2–19.1 |
||
|
Enaliarctos mealsi |
Jewett Sand, Skooner Gulch Formation |
24–21.5 |
Scheirer and Magoon 2007; |
|
|
Enaliarctos mitchelli |
Nye Mudstone, Jewett Sand |
24–19.1 |
Prothero et al. 2001b; Scheirer and Magoon 2007; Barboza et al. 2017 |
|
|
Enaliarctos tedfordi |
Yaquina Formation |
28.1–27.4 |
||
|
Enaliarctos sp. |
Astoria Formation |
17.3–16.6 |
this study |
|
|
Pteronarctos goedertae |
Astoria Formation |
20.7–16.6 |
||
|
Pacificotaria hadromma |
Astoria Formation |
17.3–16.6 |
||
|
Pinnarctidion bishopi |
Jewett Sand |
24–21.5 |
Scheirer and Magoon 2007; |
|
|
Pinnarctidion rayi |
Nye Mudstone |
20.7–17.3 |
||
The geochronologically youngest enaliarctines are all reported from the „Iron Mountain Bed” of the upper Astoria Formation, which preserves a diverse pinniped fauna including three enaliarctines (Enaliarctos sp., Pacificotaria hadromma, Pteronarctos goedertae), the desmatophocid seal Desmatophoca oregonensis and a larger undescribed Allodesmus-like desmatophocid, and the early odobenid Proneotherium repenningi (Barnes 1987, 1989, 1990, 1992; Berta 1994b; Deméré and Berta 2001, 2002; this study). Geochronologically earlier (Chattian–Aquitanian) assemblages are lower in diversity and consist entirely of enaliarctines (e.g., Yaquina Formation, Nye Mudstone, Jewett Sand; Table 3). Later assemblages lack enaliarctines, but possess small-bodied early diverging odobenids and otariids as well as larger-bodied odobenids and species of Allodesmus (Miyazaki et al. 1995). Reasons for enaliarctine extinction are unclear, yet it seems likely that small imagotariines and early otariids may reflect surviving ecological analogs in the middle and late Miocene. Further studies of early Miocene enaliarctines, feeding morphology, and stable isotope analyses may highlight possible modes of competition and extinction.
Taxonomy and diversity of Enaliarctos.—The taxonomy of Enaliarctos has not been evaluated since publication of Berta (1991), who reported three new species (Enaliarctos barnesi, E. emlongi, and E. tedfordi) from the Yaquina Formation, Nye Mudstone, and/or Astoria Formation of Oregon. With five species reported, it may be that the genus Enaliarctos is oversplit. For example, Enaliarctos barnesi and Enaliarctos mealsi both share, to the exclusion of all other species, widely flaring nasals with a concave lateral margin and strongly developed labial and lingual cingula (Berta 1991); they differ only in the presence of a posterolingual shelf on p3 and a more spur-like palatine process of the maxilla in E. barnesi (Berta 1991). It is possible that these are either sister taxa or the same species, possibilities further supported by their similar geochronologic age (Table 3; Berta 1991; Prothero et al. 2001b; Barboza et al. 2017)—although Berta (1991: fig. 21) did not recover a sister taxon relationship between these species (Fig. 8). Possible synonymies within the remaining species are more difficult to assess owing to their stratigraphic separation. Enaliarctos mitchelli is distinctive in small adult size (Barnes 1979; Berta 1991) but Enaliarctos emlongi differs from Enaliarctos tedfordi only in exhibiting larger metacone crests on p2–p3 and p3 with a larger posterolingual shelf. Craniodental similarities and limited differences between these two species, in concert with stratigraphic separation, suggest they may represent chronospecies within a single lineage. Dental similarities (e.g., loss of cingula), and the cladogram of Berta (1991: fig. 21) further suggest that E. mitchelli could be a terminal species in this lineage (though E. emlongi and E. mitchelli may temporally overlap; Fig. 8). Description of many additional undescribed enaliarctine specimens within LACM and USNM collections will permit re-evaluation of enaliarctine variation, taxonomy, and phylogeny, and test hypotheses presented herein.
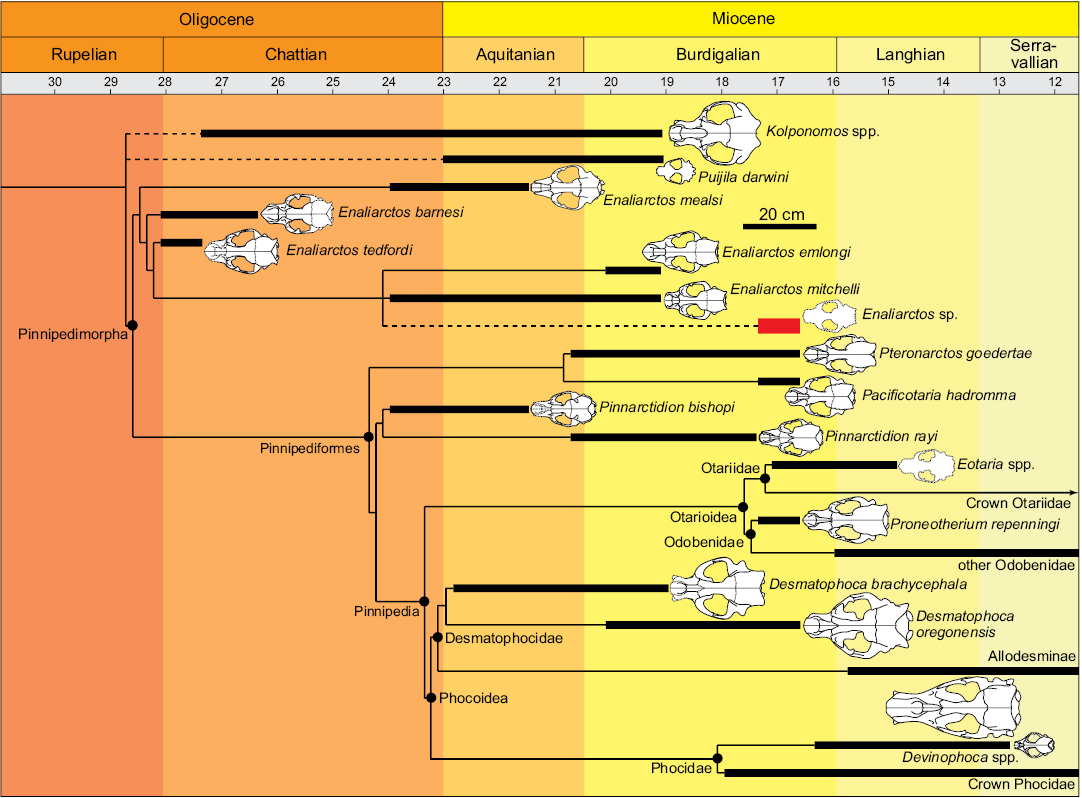
Fig. 8. Time calibrated composite phylogenetic hypothesis of early pinnipedimorphs and putative allies, with an emphasis on Oligo-Miocene pinnipedimorphs from the eastern North Pacific. Topology based on Boessenecker and Churchill (2015), except as follows: Enaliarctos from Berta (1991); Phocoidea after Boessenecker and Churchill (in press). Placement of Pacificotaria from Deméré and Berta (2001), conjectural placement (dashed lines) of Puijila and Kolponomos after Tedford et al. (1994) and Rybczynski et al. (2009), Enaliarctos sp. (UWBM 89114) based on shared dental synapomorphies with E. emlongi and E. mitchelli. Geochronologic ranges from Table 3 (this study), Boessenecker and Churchill (2015: supplementary information, in press: supplementary information), and Rybczynski et al. (2009).
Early pinnipedimorphs, cetaceans, and flightless seabirds.—Several studies have investigated possible evolutionary interactions between early pinnipedimorphs, cetaceans, and sea birds (Olson 1985; Olson and Hasegawa 1979; Ando and Fordyce 2014; Marx et al. 2015). Most Oligocene toothed baleen whales were quite small (e.g., 2–4 m body length, Fucaia buelli; Marx et al. 2015) and some at least were raptorial piscivores (Barnes et al. 1995) like early pinnipedimorphs (Churchill and Clementz 2015, 2016), raising the possibility that they competed for prey. Furthermore, the diversification of toothed mysticetes during the Oligocene coincides with the appearance of the earliest pinnipedimorphs (Marx and Fordyce 2015). We note that large-bodied (e.g., > 2 m body length) pinnipeds do not appear until the late early Miocene (Burdigalian) (Desmatophoca, Pelagiarctos; Churchill et al. 2015), well after the presumed Chattian–Aquitanian extinction of toothed mysticetes (but see Staley and Barnes 2008). Pinnipedimorph diversification has been implicated in the extinction of the giant flightless wing-propelled penguin-like plotopterids and giant Oligo-Miocene penguins (Olson 1985; Olson and Hasegawa 1979). However, analyses of diversity trends find little long term evidence for competitive displacement of penguins and plotopterids by pinnipeds, instead highlighting echolocating toothed whales (Odontoceti) as competitors (Ando and Fordyce 2014). Given various differences in feeding morphology between early pinnipedimorphs and toothed mysticetes (Churchill and Clementz 2015, 2016; Marx and Fordyce 2015; Marx et al. 2016), perhaps odontocetes occupied a more competitive role that could explain the extinction of toothed mysticetes. Lastly, we note that the diminutive enaliarctine Enaliarctos sp. now coincides in age with the tiny enaliarctine-like otariid Eotaria from California (Fig. 8; Boessenecker and Churchill 2015; Velez-Juarbe 2017). The overlap of these small-bodied taxa suggests the possibility of niche partitioning and further highlights the geochronologic antiquity of diverse pinniped communities in the North Pacific (e.g., Velez-Juarbe 2017).
Enaliarctines, Puijila, Kolponomos, and the origin of Pinnipedia.—Enaliarctines are regarded as the earliest diverging and geochronologically earliest pinnipedimorphs (Mitchell and Tedford 1973; Berta et al. 1989). Recently, a semiaquatic otter-like arctoid, Puijila darwini, was reported from lower Miocene nonmarine deposits of the Canadian arctic (Rybczynski et al. 2009). Despite lacking flipper-like manus and pes, shortened tail, and postdating the earliest Enaliarctos by nearly 10 my, this taxon exhibits several craniodental features unique to pinnipedimorphs including a posteriorly expanded hard palate, an enlarged infraorbital foramen, a shelf-like p4 protocone, and a reduced and lingually positioned m2; cladistic analysis supported Puijila and the anatomically similar Potamotherium valletoni (Oligo-Miocene, France and Germany) as sister to Enaliarctos (Rybczynski et al. 2009). If these relationships are borne out by future cladistic study, it is important to note that most of these features are also present in the enigmatic amphicynodontid Kolponomos (except shelf-like p4 protocone; Tedford et al. 1994), a bear-like molluskivore with enlarged sea otter-like cheek teeth, binocular vision, and adaptations for high bite force convergent with sabretooth cats (Tedford et al. 1994; Tseng et al. 2016). Like the nonmarine otter-like Potamotherium and Puijila, initial cladistic analysis identified Kolponomos as the possible sister taxon to Pinnipedimorpha (Tedford et al. 1994). Since these taxa are rare, continued description of fossils from the long record of enaliarctines such as we present here holds the promise of more confidently resolving these relationships through a better understanding of species-level taxonomy and an expanded matrix including more non-cranial characters. Further cladistic analysis is needed, but if indeed these taxa are all early pinniped relatives, the clade comprising them may represent a wider radiation of arctoid carnivores into various aquatic niches, including littoral molluskivores (Kolponomos), small otter-like freshwater taxa (Potamotherium, Puijila), and the highly successful marine pinnipedimorphs.
Acknowledgements
First and foremost, we thank Jason Love (Seattle, Washington, USA) for donating UWBM 89114 and for other donations to UWBM. UCMP provided laboratory facilities for the preparation of UCMP 253400. Thanks to Lawrence G. Barnes, Samuel A. McLeod, and Vanessa Rhue (all Natural History Museum of Los Angeles County, USA), Patricia A. Holroyd and Mark Goodwin (both UCMP), Ron Eng, Meredith Rivin, and Christian Sidor (all UWBM), and David J. Bohaska and Nicholas D. Pyenson (both USNM) for access to collections under their care. We thank David Strauss for photography in Figs. 3 and 5. This study benefited from discussions of early pinniped evolution with L.G. Barnes, Annalisa Berta (San Diego State University, USA), Thomas A. Deméré (San Diego Natural History Museum, USA), and Morgan Churchill (University of Wisconsin Oshkosh, USA). This study benefited greatly from constructive reviews by A. Berta, Naoki Kohno (National Museum of Nature and Science, Tokyo, Japan), and Olivier Lambert (Royal Belgian Institute of Natural Sciences, Brussels, Belgium). RWB was supported by a Vertebrate Paleontology Collection Study Grant from the University of Washington Burke Museum of Natural History and Culture. This is UCMP contribution number 2084.
References
Adam, P.J. and Berta, A. 2002. Evolution of prey capture strategies and diet in the Pinnipedimorpha (Mammalia, Carnivora). Oryctos 4: 83–107.
Addicott, W.O. 1967. Age of the Skooner Gulch Formation, Mendocino County, California. United States Geological Survey Bulletin Contributions to Stratigraphy 1254C: 1–11.
Ando, T. and Fordyce R.E. 2014. Evolutionary drivers for flightless, wing-propelled divers in the Northern and Southern hemispheres. Palaeogeography, Palaeoclimatology, Palaeoecology 400: 50–61. Crossref
Armentrout, J.M. 1981. Correlation and ages of Cenozoic chronostratigraphic units in Oregon and Washington. Geological Society of America Special Paper 184: 137–148. Crossref
Barboza, M.M., Parham, J.F., Santos, G.-P., Kussman, B.N., and Velez-Juarbe, J. 2017. The age of the Oso Member, Capistrano Formation, and a review of fossil crocodylians from California. PaleoBios 34: 1–16.
Barnes, L.G. 1979. Fossil enaliarctine pinnipeds (Mammalia: Otariidae) from Pyramid Hill, Kern County, California. Contributions in Science, Natural History Museum of Los Angeles County 318: 1–41.
Barnes, L.G. 1990. A new Miocene enaliarctine pinniped of the genus Pteronarctos (Mammalia: Otariidae) from the Astoria Formation, Oregon. Contributions in Science, Natural History Museum of Los Angeles County 422: 1–20.
Barnes, L.G. 1989. A new enaliarctine pinniped from the Astoria Formation, Oregon, and a classification of the Otariidae (Mammalia: Carnivora). Contributions in Science, Natural History Museum of Los Angeles County 403: 1–26.
Barnes, L.G. 1992. A new genus and species of middle Miocene enaliarctine pinniped (Mammalia, Carnivora, Otariidae) from the Astoria Formation in coastal Oregon. Contributions in Science, Natural History Museum of Los Angeles County 431: 1–27.
Barnes, L.G., Kimura, M., Furusawa, H., and Sawamura, H. 1995. Classification and distribution of Oligocene Aetiocetidae (Mammalia; Cetacea; Mysticeti) from western North America and Japan. The Island Arc 3: 392–431. Crossref
Berta, A. 1991. New Enaliarctos (Pinnipedimorpha) from the Oligocene and Miocene of Oregon and the role of “Enaliarctids” in pinniped phylogeny. Smithsonian Contributions to Paleobiology 69: 1–36. Crossref
Berta, A. 1994a. A new species of phocoid pinniped Pinnarctidion from the early Miocene of Oregon. Journal of Vertebrate Paleontology 14: 405–413. Crossref
Berta, A. 1994b. New specimens of the pinnipediform Pteronarctos from the Miocene of Oregon. Smithsonian Contributions to Paleobiology 78: 1–30. Crossref
Berta, A. and Ray, C.E. 1990. Skeletal morphology and locomotor capabilities of the archaic pinniped Enaliarctos mealsi. Journal of Vertebrate Paleontology 10: 141–157. Crossref
Berta, A., Ray, C.E., and Wyss, A.R. 1989. Skeleton of the Oldest Known Pinniped, Enaliarctos mealsi. Science 244: 60–62. Crossref
Boessenecker, R.W. and Churchill, M. 2013. A Reevaluation of the morphology, paleoecology, and phylogenetic relationships of the enigmatic walrus Pelagiarctos. PLoS ONE 8 (1): e54311. Crossref
Boessenecker, R.W. and Churchill, M. 2015. The oldest known fur seal. Biology Letters 11: 20140835. Crossref
Boessenecker, R.W. and Churchill, M. (in press). The last of the desmatophocid seals: a new species of Allodesmus from the upper Miocene of Washington, USA, and a revision of the taxonomy of Desmatophocidae. Zoological Journal of the Linnean Society. Crossref
Bowditch, T.E. 1821. An Analysis of the Natural Classifications of Mammalia for the Use of Students and Travelers. 151 pp. J. Smith, Paris.
Churchill, M. and Clementz, M.T. 2015. Functional implications of variation in tooth spacing and crown size in Pinnipedimorpha (Mammalia: Carnivora). The Anatomical Record 289: 878–902. Crossref
Churchill, M. and Clementz M.T. 2016. The evolution of aquatic feeding in seals: insights fron Enaliarctos (Carnivora: Pinnipedimorpha), the oldest known seal. Journal of Evolutionary Biology 29: 319–334. Crossref
Churchill, M., Clementz, M.T., and Kohno, N. 2015. Cope’s rule and the evolution of body size in Pinnipedimorpha (Mammalia: Carnivora). Evolution 69: 201–215. Crossref
Clark, J.M. 1991. A new early Miocene species of Paleoparadoxia (Mammalia: Desmostylia) from California. Journal of Vertebrate Paleontology 11: 490–508. Crossref
Colbath, S.L. 1985. Gastropod predation and depositional environments of two molluscan communities from the Miocene Astoria Formation at Beverly Beach State Park, Oregon. Journal of Paleontology 59: 849–869.
Cullen, T.M., Fraser, D., Rybczynski, N., and Schröder-Adams, C. 2014. Early evolution of sexual dimorphism and polygyny in pinnipedia. Evolution 68: 1469–1484. Crossref
Condon, T. 1906. A new fossil pinniped (Desmatophoca oregonensis) from the Miocene of the Oregon coast. University of Oregon Bulletin Supplement 3 (3): 1–14.
Deméré, T.A. and Berta, A. 2001. A reevaluation of Proneotherium repenningi from the Miocene Astoria Formation of Oregon and its position as a basal odobenid (Pinnipedia: Mammalia). Journal of Vertebrate Paleontology 21: 279–310. Crossref
Deméré, T.A. and Berta, A. 2002. The Miocene pinniped Desmatophoca oregonensis Condon, 1906 (Mammalia: Carnivora) from the Astoria Formation, Oregon. In: R.J. Emry (ed.), Later Cenozoic Mammals of Land and Sea: Tributes to the Career of Clayton E. Ray. Smithsonian Contributions to Paleobiology 93: 113–147.
Deméré, T.A. and Berta, A. 2008. Skull anatomy of the Oligocene toothed mysticete Aetioceus weltoni (Mammalia; Cetacea): implications for mysticete evolution and functional anatomy. Zoological Journal of the Linnean Society 154: 308–352. Crossref
Dineen, A.A., Fraiser, M.L., and Sheehan, P.M. 2014. Quantifying functional diversity in pre- and post-extinction paleocommunities: a test of ecological restructuring after the end-Permian mass extinction. Earth-Science Reviews 136: 339–349.Crossref
Domning, D.P., Ray, C.E., and McKenna, M.C. 1986. Two new Oligocene desmostylians and a discussion of tethytherian systematics. Smithsonian Contributions to Paleobiology 59: 1–56. Crossref
Dooley, A.C., Jr. 1994. The first well preserved squalodont (Cetacea) from the west coast of North America. Journal of Vertebrate Paleontology 14 (supplement 3): 23A.
Emlong, D.R. 1966. A new archaric cetacean from the Oligocene of Northwest Oregon. Bulletin of the Museum of Natural History of Oregon 3: 1–51.
Hannibal, H. 1922. Notes on Tertiary sirenians of the genus Desmostylus. Journal of Mammalogy 3: 238–240. Crossref
Hocking, D.P., Ladds, M.A., Slip, D.J., Fitzgerald, E.M.G., and Evans, A.R. 2016. Chew, shake, and tear: prey processing in Australian sea lions (Neophoca cinerea). Marine Mammal Science 33: 541–557. Crossref
Hunt, R.M. and Barnes, L.G. 1994. Basicranial evidence for ursid affinity of the oldest pinnipeds. Proceedings of the San Diego Society of Natural History 29: 57–67.
Kloess, P.A. and Parham, J.F. 2017. A Specimen-based approach to reconstructing the Neogene seabird communities of California. Palaeogeography, Palaeoclimatology, Palaeoecology 468: 473–484. Crossref
Kohno, N., Barnes, L.G., and Hirota, K. 1995. Miocene fossil pinnipeds of the genera Prototaria and Neotherium (Carnivora; Otariidae; Imagotariinae) in the north Pacific Ocean: evolution, relationships, and distribution. The Island Arc 3: 285–308. Crossref
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis,locis. Editio decima. 824 pp. Laurentius Salvius, Holmiae.
Loomis, K.B. and Ingle, J.C. 1995. Subsidence and uplift of the Late Cretaceous–Cenozoic margin of California: new evidence from the Gualala and Point Arena basins. Geological Society of America Bulletin 106: 915–931. Crossref
Marx, F.G. and Fordyce, R.E. 2015. Baleen boom and bust: a synthesis of mysticete phylogeny, diversity, and disparity. Royal Society Open Science 2: 140434. Crossref
Marx, F.G., Hocking, D.P., Park, T., Ziegler, T., Evans, A.R., and Fitzgerald, E.M.G. 2016. Suction feeding preceded filtering in baleen whale evolution. Memoirs of Museum Victoria 75: 71–82.Crossref
Marx, F.G., Tsai, C.-H., and Fordyce, R.E. 2015. A new Early Oligocene toothed “baleen” whale (Mysticeti: Aetiocetidae) from western North America: one of the oldest and the smallest. Royal Society Open Science 2: 150476. Crossref
Miller, P.L. 1981. Tertiary calcareous nannoplankton and benthic foraminifera biostratigraphy of the Point Arena area, California. Micropaleontology 27: 419–443. Crossref
Mitchell, E.D. and Repenning, C.A. 1963. The chronologic and geographic range of desmostylians. Los Angeles County Museum Contributions in Science 78: 3–20.
Mitchell, E.D. and Tedford, R.H. 1973. The Enaliarctinae A new group of extinct aquatic carnivora and a consideration of the origin of the otariidae. Bulletin of the American Museum of Natural History 151: 201–284.
Miyazaki, S., Horikawa, H., Kohno, N., Hirota, K., Kimura, M., Hasegawa, Y., Tomida, Y., Barnes, L.G., and Ray, C.E. 1995. Summary of the fossil record of pinnipeds of Japan, and comparisons with that from the eastern North Pacific. The Island Arc 3: 361–372. Crossref
Munthe, J. and Coombs, M.C. 1979. Miocene dome-skulled chalicotheres (Mammalia, Perissodactyla) from the western United States: a preliminary discussion of a bizarre structure. Journal of Paleontology 53: 77–91.
Moore, E.J. 1963. Miocene marine mollusks from the Astoria Formation in Oregon. U.S. Geological Survey Professional Paper 419: 1–109.
Novack-Gottshall, P.M. 2007. Using a theoretical ecospace to quanitfy the ecological diversity of Paleozoic and modern marine biotas. Paleobiology 33: 273–294. Crossref
Ogg, J.G. 2012. Geomagnetic polarity timescale. In: F.M. Gradstein, J.G. Ogg, M. Scmiyz, and G. Ogg (eds.), The Geologic Time Scale 2012, 85–113. Elsevier, Amsterdam.
Olson, S.L. 1985. The Fossil Record of Birds. In: D.S. Farner, J.R. King, and K.C. Parkes (eds.), Avian Biology, Volume 8, 79–238. Academic Press, New York. Crossref
Olson, S.L. and Hasegawa, Y. 1979. Fossil counterparts of giant penguins from the North Pacific. Science 206: 688–689. Crossref
Packard, E.L., and Kellogg, R. 1934. A new cetothere from the Miocene Astoria Formation of Newport, Oregon. Carnegie Institution of Washington Publications 447: 1–62.
Phillips, F.J., Welton, B.J., and Welton, J. 1976. Paleontologic studies of the middle Tertiary Skooner Gulch and Gallaway Formations. Proceedings of the SEPM, Pacific Section: 137–154.
Prothero, D.R. 2001. Chronostratigraphic calibration of the Pacific Coast Cenozoic: a summary. Pacific Section SEPM Special Publications 9: 377–394.
Prothero, D.R., Bitboul, C.Z., Moore, G.W., and Moore, E.J. 2001a. Magnetic stratigraphy of the lower and middle Miocene Astoria Formation, Lincoln County, Oregon. Pacific Section SEPM Special Publication 91: 272–283.
Prothero, D.R., Bitboul, C.Z., Moore, G.W., and Niem, A.R., 2001b. Magnetic stratigraphy and tectonic rotation of the Oligocene Alsea, Yaquina, and Nye formations, Lincoln County, Oregon. Pacific Section SEPM Special Publications 91: 184–194.
Rau, W.W. 1981. Pacific Northwest Tertiary benthic foraminiferal biostratigraphic framework—an overview. Geological Society of America Special Paper 184: 67–84. Crossref
Rybczynski, N., Dawson, M. R., and Tedford, R.H. 2009. A semi-aquatic Arctic mammalian carnivore from the Miocene epoch and the origin of Pinnipedia. Nature 458: 1021–1024. Crossref
Shimada, K., Welton, B.J., and Long, D.J. 2014. A new megamouth shark (Lamniformes, Megachasmidae) from the Oligocene–Miocene of the western United States. Journal of Vertebrate Paleontology 34: 281–290. Crossref
Snavely, P.D., MacLeod, N.S., Wagner, H.C., and Rau, W.W. 1976. Geologic map of the Yaquina and Toledo quadrangles, Lincoln County, Oregon. In: US Geological Survey Miscellaneous Investigations Series, Map I-867.
Staley, M. and Barnes, L.G. 2008. Early Miocene toothed cetaceans from the Vaqueros Formation, Orange County, California. Journal of Vertebrate Paleontology 28 (Supplement to No. 3): 147A.
Tedford, R.H., Barnes, L.G., and Ray, C.E. 1994. The early Miocene littoral ursoid carnivoran Kolponomos: systematics and mode of life. Proceedings of the San Diego Society of Natural History 29: 11–32.
Tseng, Z.J., Grohe, C., and Flynn, J.J. 2016. A unique feeding strategy of the extinct marine mammal Kolponomos: convergence on sabertooths and sea otters. Proceedings of the Royal Society B 283: 2160044. Crossref
Turner, D. 1970. Potassium-argon dating of Pacific Coast Miocene foraminiferal stages. In: O.L. Bandy (ed.), Radiometric Dating and Paleontological Zonation. Geological Society of America Special Paper 124: 91–129. Crossref
Velez-Juarbe, J. 2017. Eotaria citrica sp. nov., a new stem otariid from the “Topanga” formation of Southern California. PeerJ 5: e3022. Crossref
Welton, B.J. 1972. Fossil sharks of Oregon. The Ore Bin 34 (10): 161–172.
Welton, B.J. 2015. A new species of late early Miocene Cetorhinus (Lamniformes: Cetorhinidae) from the Astoria Formation of Oregon, and coeval Cetorhinus from Washington and California. Natural History Museum of Los Angeles County Contributions in Science 523: 67–89.
Acta Palaeontol. Pol. 63 (1): 25–40, 2018
https://doi.org/10.4202/app.00399.2017