
A new Mississippian hexactinellid sponge from the western Gondwana: Taxonomic and paleobiogeographic implications
MARCELO G. CARRERA, JUAN JOSE RUSTÁN, N. EMILIO VACCARI, and MIGUEL EZPELETA
Carrera, M.G., Rustán, J.J., Vaccari N.E., and Ezpeleta, M. 2018. A new Mississippian hexactinellid sponge from the western Gondwana: Taxonomic and paleobiogeographic implications. Acta Palaeontologica Polonica 63 (1): 63–70.
A Mississippian hexactinellid sponge from the western Argentina improves the extremely poor late Paleozoic sponge records from Gondwana. The sponge is included in the subfamily Thysanodictyinae of family Dictyospongiidae. The new genus and species Minitaspongia parvis is erected, and its well-preserved spicular structure is described in detail representing the first approximation of the spicule assemblage in Thysanodictyinae. The skeleton is clathrate, three-dimensional with at least two ranks of rectangular openings. This first report of this subfamily outside North America represents the best-known hexactinellid and the first dictyosponge record from the Carboniferous of Gondwana. Unlike the occurrences of Thysanodictyinae in North America, with thick skeletons linked to high-energy shallow water settings, Minitaspongia occurs in low-energy water siliciclastic settings related to a cold climate and glacimarine deposits. Accordingly, the complex wall structure of this sponge should not be invoked as a necessary adaptation to high energy and shallow water settings.
Key words: Porifera, Hexactinellida, Reticulosa, Carboniferous, Mississippian, Argentina.
Marcelo G. Carrera [mcarrera@unc.edu.ar], CICTERRA-CONICET Universidad Nacional de Córdoba, Facultad de Ciencias Exactas Físicas y Naturales, Av. Vélez Sarfield 1611, Ciudad Universitaria, 5016, Córdoba, Argentina.
Juan José Rustan [juanjorustan@gmail.com], N. Emilio Vaccari [evaccari@unc.edu.ar], and Miguel Ezpeleta [miguelezpeleta@gmail.com], CICTERRA-CONICET Universidad Nacional de Córdoba and Universidad Nacional de La Rioja, Av. Luis M. de la Fuente s/n, Ciudad Universitaria, 5300, La Rioja, Argentina.
Received 23 June 2017, accepted 18 September 2017, available online 6 February 2018.
Copyright © 2018 M.G. Carrera et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The worldwide records of Carboniferous sponges show a particular abundance of sphinctozoans, the lithistid rhizomorines and the hexactinellid dictyosponges, mainly concentrated in North America, China, United Kingdom, and Spain (García-Bellido and Rodríguez 2005 and references therein). Such a distribution, almost excluding Gondwana, suggests a sampling bias rather than a true paleobiogeographic pattern. As a consequence any new record outside these areas significantly contributes to the global diversity and biogeographic knowledge.
This contribution reports the find of a new genus of hexactinellid dictyosponge in the very limited record of late Paleozoic sponges from Gondwana. The sponge was found in Carboniferous marine periglacial deposits of western Argentina and represents one of a few examples of high-latitude occurrences, linked to glacial deposits. The finding also documents the first well-preserved spicule mesh composition, which allows describing in detail the elusive, and poorly known, spicule pattern of dictyosponges.
Some environmental insights are provided about the new genus Minitaspongia, in order to better interpret the recognized paleobiogeographic patterns.
Institutional abbreviations.—PULR-I, Museo de Ciencias Antropológicas y Naturales at Universidad Nacional de La Rioja, La Rioja, Argentina.
Geographic and geological setting
The stratigraphic record surveyed corresponds to a series of late Paleozoic basins developed in southwestern Gondwana linked to an active tectonic margin (Limarino and Spalletti 2006). In the context of the Central Andean foreland basin (Caminos et al. 1993), we focused on the Mississippian filling of the Río Blanco basin, a high paleolatitude southern basin (> 60° according to Isaacson et al. 2008). Strata surveyed crop out at the Sierra de las Minitas to the northwest of the small town of Jagüé, in central western Argentina (northern Argentine Precordillera). Fossils come from the southwestern region of the Sierra de Las Minitas (Fig. 1), to the west of de Agua Quemada syncline (see Fauqué and Limarino 1991 and Prestianni et al. 2015).
The studied material was recovered from two stratigraphically equivalent fossil localities, separated by a fault (tectonic block 1 and tectonic block 2), and located on the western flank of the Agua Quemada syncline (Fig. 1). The productive levels are correlated with the Agua de Lucho Formation, the main stratigraphic unit recording low (Tournaisian) Mississippian records in the area (González and Bossi 1987). In the studied sections, the Agua de Lucho Formation is approximately 800 m thick, without visible base, and it is composed of dominantly pelitic facies with thick interbedded diamictitic bodies yielding dropstones, and the succession is characteristic of a deltaic front and a prodelta, related to storm-dominated glacio-marine environments. A similar environmental interpretation was proposed by González and Bossi (1987) in the northern region of the basin.
Specifically, the specimens described herein come from a greenish-black pelitic interval of about 100 m thick, located immediately below a diamictite bed, laterally equivalent to the one reported in the base of the column of the Agua Quemada Syncline by Prestianni et al. (2015). In these dark, fine-grained fossiliferous interval, dropstones are recognizable in the uppermost part while a significant number of siliceous nodules occur in the sampled middle-lower portion. Fossil remains are diverse and well-preserved (usually articulated) either in the shales or in the nodules. In addition to the sponges studied here, brachiopods, bryozoans, hyolitids, conularids, bivalves, gastropods, nautiloids, crinoids, corals, fishes, and plant remains are also present. These sedimentological and taphonomic features largely suggest that suspension processes dominated over tractive ones, so that this succession is interpreted as a marine shelf environment with glacial influence.
Since the Gondwanan palynomorph Waltzispora lanzonii Daemon, 1974, indicative of a Tournaisian age, has been recorded nearly 50 m above the overlying diamictites (see Prestianni et al. 2015), the sponge bearing levels are considered Tournaisian.
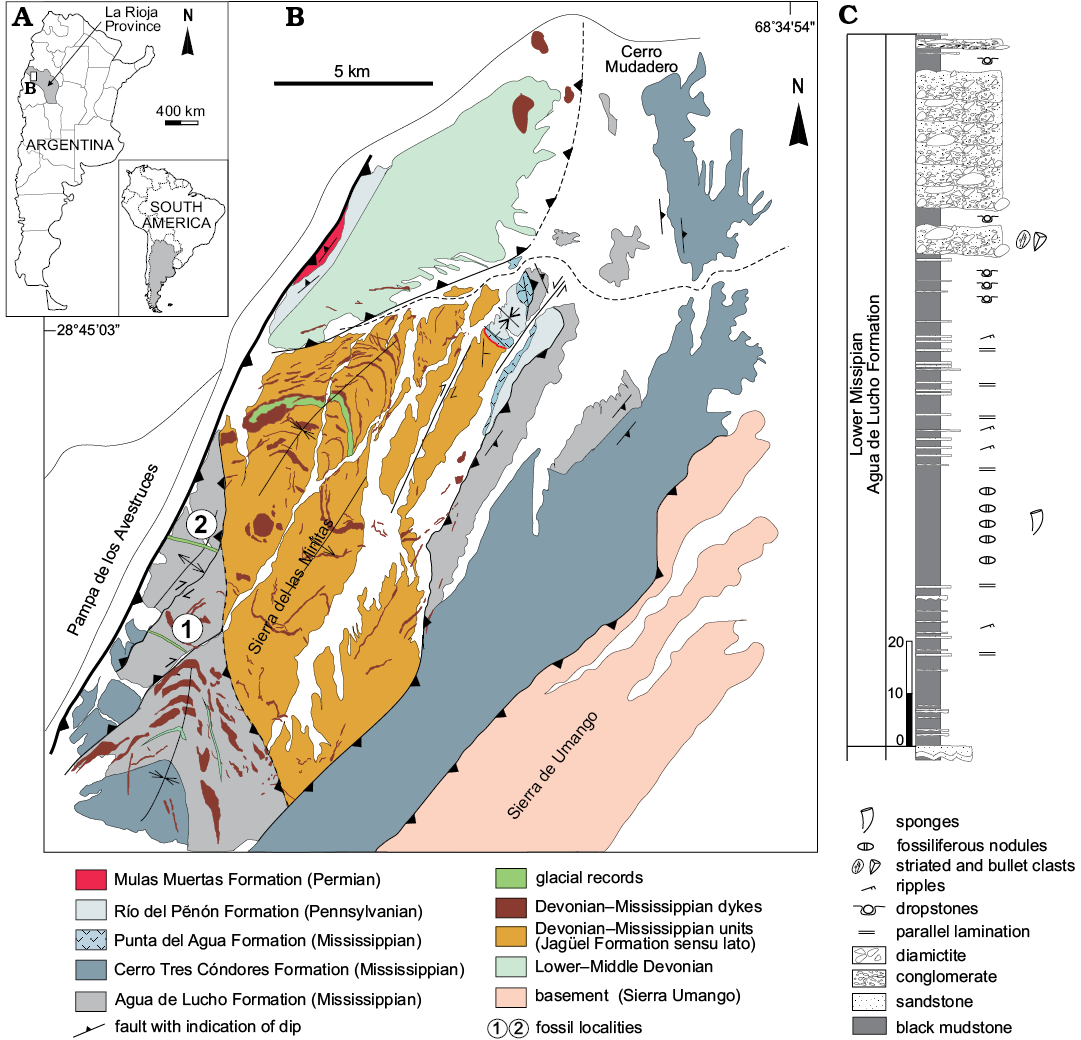
Fig. 1. A. Geographic location of the studied area. B. Geological map and location of the studied section. The west flank of the Agua Quemada Syncline is divided in some tectonic blocks (1 and 2) by inverse and dextral strip faults. Sponges come from nodules of equivalent fine-grained dark stratigraphical intervals of the Agua de Lucho Formation (Tournaisian), cropping out under glacigenic diamictitic in both tectonic blocks. C. Stratigraphic column of the Agua de Lucho Formation and the location of the sponge-bearing levels.
Systematic palaeontology
Class Hexactinellida Sollas, 1887
Order “Reticulosa” Reid, 1958
Informal group (see Botting and Muir 2013)
Family Dictyospongiidae Hall, 1882
Remarks.—The dictyospongid is a complex and varied group of hexactinellid sponges that certainly is in need of a major revision. The existing definitions of the superfamily Dictyospongioidea, family Dictyospongiidae, and the subfamilies are complex and unclear. Many taxa are currently included within groups that do not share very many of the same characteristics. Main definition of the superfamily Dictyospongioidea Hall, 1882 (according to Finks and Rigby 2004) includes “reticulosan” hexactinellids in which one or more layers of vertical and horizontal bundles of spicules developed beneath a dermal layer. However, a differentiated dermal layer is not clearly represented in all the included genera.
The definition of the family Dictiospongiidae referred to a fine autodermal and hypodermal structure (quadrate mesh) and the subdivision into six subfamilies (Finks and Rigby 2004: 372) are mainly based on the overall morphology of the sponge body: “basically tubular with prism facets”, “well developed prism facets and no protuberances”, “Conicocylidrical with annular expansions an quite large protuberances”.
The subfamily Thysanodictyinae (in which we include the studied material) is one of the few that includes the structure of the wall as an important characteristic in its definition. However, as we pointed earlier, the dictyopongids need a major revision which is beyond the scope of this work.
Subfamily Thysanodictyinae Hall, 1882
Genus Minitaspongia nov.
Type species: Minitaspongia parvis gen et. sp. nov.; monotypic; see below.
Etymology: From the Sierra de las Minitas locality.
Diagnosis.—As for type species by monotypy.
Minitaspongia parvis sp. nov.
Figs. 2, 3.
Etymology: From Latin parvis, small, young.
Type material: Holotype: PULR-I 4 nearly complete sponge specimen. Paratype: PULR-I 5, fragmentary specimen.
Type locality: Sierra de las Minitas section, La Rioja Province, Argentina. Holotype comes from tectonic block 1 at GPS: 28° 47 43 S/ 68° 45 22 W . Paratype comes from tectonic block 2 at GPS 28° 46 28 S/ 68° 44 40 W (Fig. 1).
Type horizon: Agua de Lucho Formation, Tournaisian. The sponge comes from nodules within a thick black fossiliferous mudstone interval, approximately 100–150 m underlying a diamictitic, glacigenic bed, and approximately 200–250 m below the record of the Tournaisian guide spore Waltzispora lanzonii Daemon, 1974 (see Prestianni et al. 2015).
Material.—Type material only.
Diagnosis.—Obconical to palmate thysanodictyine sponge with subcylindrical base expanding upward to define a broad spongocoel, with a wall consisting of a few layers of spicules. Skeleton clathrate, three-dimensional with at least two ranks of rectangular openings. First order quadrules 2 × 1.5–1.8 mm and the second order approximately 0.5 × 0.5 mm across. Vertical tracts supported by two or three aligned long-rayed hexactins, 3–4 mm in maximum length and a shorter ray 1.0–1.2 mm.
Description.—The specimens are small, obconical to palmate sponges. The holotype, a nearly complete specimen, shows an upward-expanding obconical shape, approximately 3–4 cm high. The specimen begins with a small cylindrical base 3.5 mm in diameter, apparently broken, from which the body expands abruptly in a fan or obconical shape reaching 20 mm wide. The walls are 1–2 mm thick and outline a wide and deep spongocoel that reaches almost to base.
The specimens lack nodes, wrinkles or other sculpture that typically characterizes other genera of the family. Externally, the mesh is marked by rectangular skeletal openings (defined mainly by impressions and limonite stains in silty matrix). The skeleton shows a tridimensional clathrate wall of regular vertical and horizontal blade-like skeletal straps.
The wall consists of box-like meshes outlined by spicular bundles of hexactins and hexactine-based spicules. The skeletal grid has typical rectangular reticulation of dictyosponges; with primary quadrules nearly 2 mm high and 1.5–1.8 mm wide, and spicule tracts of 0.1–0.3 mm wide (Figs. 2, 3).
Vertical tracts are coarse (tridimensional) and mainly supported by two or three aligned long-rayed hexactins (probably stauracts). These primary hexactines have a long vertical tangential ray, 3–4 mm in maximum length and a short tangential horizontal ray 1.0–1.2 mm, disposed perpendicularly to the vertical tracts (Fig. 2B4–B7). The third axis, perpendicular to the sponge surface, appeared as a knob-like distal ray, the other end is uncertain, it probably continues to the interior of the wall, in which case the spicule is a pentactin not a stauract (Fig. 2B7).
Second order quadrules, are approximately 0.5 mm high and 0.5 mm wide. In well preserved patches small hexactins, with equal ray length (0.8–1.0 mm), have been observed as main part of the quadrules. In the external surface this second order quadrules occur as a cross-like center dividing the first order quadrules (Fig. 2A). In the periphery of the sponge wall a marginal concentration of second order quadrules, implies a more complex structure including fine autodermal and hypodermal quadrate meshes (Fig. 2B1–B3).
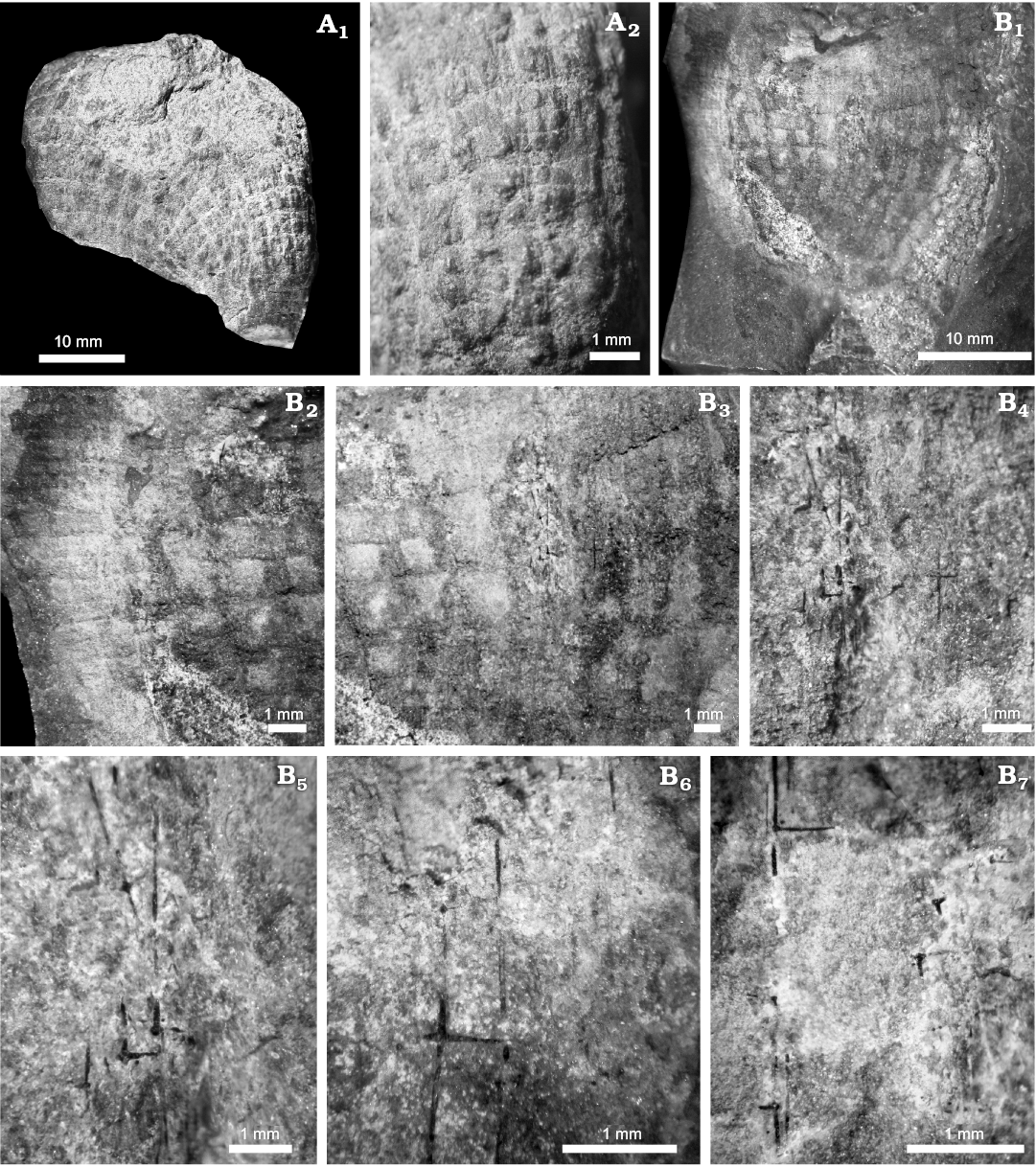
Fig. 2. Hexactinellid sponge Minitaspongia parvis gen. et sp. nov. from the Tournaisian Carboniferous, Sierra de las Minitas section, La Rioja Province, Argentina. A. Holotype PULR-I 4, complete obconical specimen, showing the cylindrical acute base. External surface of the holotype showing deep impressions of vertical and horizontal tracts and cross-shaped spicule molds as second order elements in the middle of first order quadrules (A2). B. Paratype PULR-I 5, external impression with original spicules forming the reticulate skeleton (B1); detail showing a well preserved external area of the second order quadrules (B2); detail of the first order quadrules showing preserved hexactine-based spicules aligned (B3); detail of the tract with vertically oriented, main long axis hexactine- based spicules, bundles of two or three long spicule axis occur in the tract (B4); enlarged section of B4 showing nodes of the perpendicular axis of the hexactins (B5); details of the spicules in the tracts, long rayed hexactins, and small second order hexactins with equal-ray dimensions (B6, B7).
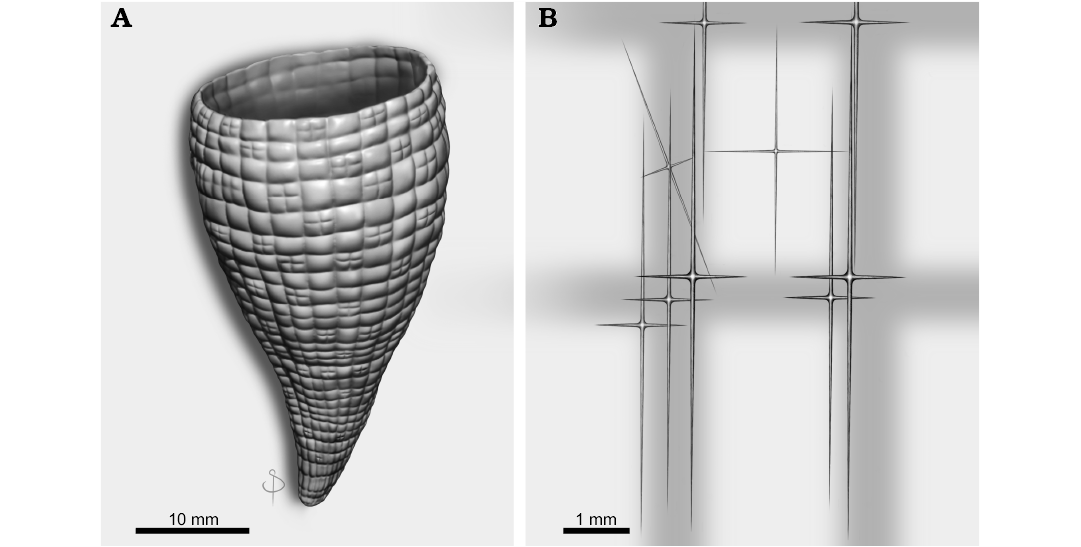
Fig. 3. A. Schematic reconstruction of the complete sponge Minitaspongia parvis gen et. sp. nov. B. The spicule framework of the wall as main parts of the vertical and horizontal bands.
Remarks.—The specimens are included in the subfamily Thysanodictyinae Hall, 1882 (family Dictyospongiidae Hall, 1882) and are related to Clathrospongia Hall, 1882 or Thysanodictya Hall and Clarke, 1899. Both genera exhibit a typical coarse ridge-like, quadrate mesh of spicule bundles based on pentactins and hexactins. This skeletal structure is uncommon among dictyosponges, which in general have very thin skeletons made of thin strap-like tracts.
Thysanodictya and Clathrospongia both have a clathrate or grid-like skeleton, which is three-dimensional with box- like mesh openings of at least two ranks in a very symmetrical pattern.
Thysanodictya has a discoidal base (wide basal disc), and a cylindrical morphology with big and rectangular, vertically oriented quadrules.
Clathrospongia is a conico-cylindrical sponge with coarse ridges and more quadrate openings. Hall and Clarke (1899) referred four species of Devonian and Carboniferous dictyosponges to the genus Clathrospongia, in addition other four species were referred to the genus with question. Variation in grid sizes is significant and it was used for species differentiation: the type species Clathrospongia abacus Hall, 1882, has a primary grid of 9 × 10 mm. C. fenestrata (Hall 1863), C. caprodonta Hall and Clarke, 1899, and C. vascellum (Hall, 1890), all have a coarser grid, ranging from 0.9 × 10 mm, to a maximum of 26 × 30 mm in C. vascellum (the original spelling is C. vascellum and not C. bascellum, as cited by Rigby and Keyes 1990).
Rigby and Keyes (1990) described Clathrospongia bangorensis from the upper Mississippian of Alabama which is similar in its external morphology to the Argentinean species. However, the grid is bigger, 4 × 6 mm for the primary grid with second order openings 3 × 3 mm across.
These grid sizes are considerably larger than the primary grid of our specimens (2.0 × 1.5 mm). However, the size of the grid alone should not be taken as a reliable character. The reticulosans grew by expansion (Botting 2003) and skeletal dimensions will be dependent on growth stage. Only when the proportion of the quadrule sides is maintained in relation with other taxa known from larger specimens, a more confident comparison could be considered.
Other related genus is Thamnodictya Hall, 1884 which has a very acute base that expands very rapidly given the sponge a fan-like appearance. This morphology is clearly different from the features observed in the Argentinean material.
The Argentinean sponge is a small and probably young specimen. The clathrate skeleton, three-dimensional with at least two ranks of rectangular openings is shared by Thysanodictya and Clathrospongia. However, the overall structure and arrangement of the quadrules in the wall, and their relative side dimensions are different from those seen in these genera. The vertical tracts supported by two or three aligned long-rayed hexactins could be considered as a distinct characteristic of the new genus. Unfortunately, there are no descriptions of the spicule details and size in the related genera of the subfamily.
There are very few records of preserved spicule net among the family. The Devonian Mattaspongia apache Rigby, 1970 shows spicule tracts composed of subparallel hexactines and smaller rhabdodiactines, that also occur as prostalia. This spicule net composition clearly differs from the structure observed in the Argentinean sponge.
The genus Corticospongia Caster, 1939, belonging to the subfamily Calathospongiinae Hall and Clarke, 1899 (Finks and Rigby 2004), shows preserved spicules forming the reticulate skeleton with main hexactine spicules similar to those of Minitaspongia parvis gen. et sp. nov., a knoblike distal ray and a long main tangential ray but disposed horizontally. The general structure and the overall morphology of this form are clearly different from the Argentinean species.
The marginal distribution of the second order quadrules in the sponge wall of Minitaspongia gen. nov. (Fig. 2B2), implies a more complex structure, fine autodermal and hypodermal meshes, which is comparable, to the restoration of Hall and Clarke (1899) or Finks and Rigby (2004) for Clathrospongia or Thysanodictya.
Hall and Clarke (1899) interpreted the quadrate mesh as representing radially, solid erected lamellae, as a common pattern seen in the subfamily. The coarse ridge mesh in our material is three-dimensional forming an apparent vertical wall expanded towards the interior of the sponge body. The vertical ridges are internally irregular composed of vertically and horizontally arranged bundles of subparallel hexactine-based spicules. This structure is different to the interpretation of quadrules with laminar solid walls envisaged by Hall and Clarke (1899), a view also questioned by Rigby and Keyes (1990) and Finks and Rigby (2004).
A remaining unclear feature is the whole composition of the vertical and horizontal parietal tracts. It is clear that the main scaffold structure of the spicular skeleton is supported by the long vertical rayed hexactines, but unknown smaller elongated spicules (monaxon types?) may also participate in the spicular bundle construction.
Stratigraphic and geographic range.—Tournaisian, Carboniferous; Agua de Lucho Formation, Las Minitas section, La Rioja Province, northern Argentine Precordillera.
The paleobiogeographic significance of the Argentinean sponge
The Carboniferous shows an abundance of calcified sponges, the type of sponge organization known as “sphinctozoans”, characterized by hypercalcified chambered skeletons. This group occurs mainly in areas of carbonate sedimentation (King 1938; Van de Graff 1969; Rigby and Mundy 2000; Rigby and Mapes 2000; García-Bellido et al. 2004). Their diversity and abundance in equatorial areas and reef settings increased during the Permian (Finks 1960; Finks and Rigby 2004).
Another well-represented group in the Carboniferous are the “lithistid” demosponge rhizomorines. Their abundance is in fact due to particular expansion of some genera such as Haplistion Young and Young, 1877 and Chaunactis Finks, 1960 recorded mainly in North America (Finks 1960; Rigby et al. 1970; Rigby and Manger 1994; Rigby and Mapes 2000; Dilliard and Rigby 2001); Northern England (Rigby and Mundy 2000) and Spain (García-Bellido and Rigby 2004). These forms occur in areas with carbonate sedimentation but they are also found in mixed carbonate-siliciclastic settings.
Another group commonly recorded in these settings are the calcareous heteractinid sponges whose records include specimens of the wewokellid family, mainly the type genus Wewokella Girty, 1911 (King 1938; Rigby and Moyle 1959; Croneis and Toomey 1965; Rigby and Church 1993; Rigby and Mapes 2000).
Although present, there are comparatively fewer occurrences of hexactinellid sponges in calcareous rocks. Some of these examples are the Bangor limestone of Alabama (Rigby and Keyes 1990) where some species of dictyosponges occur, Nevada (Rigby and Washburn 1972), Alaska (Rigby et al. 1979), and China (Rigby et al. 1999), where the genus Stereodictyum, Finks 1960, has been cited. Other records of Carboniferous hexactinellids occur in calcareous deposits of Poland (Hurcewicz and Czarniecki 1985) and in microbial buildups of Morocco (Aretz and Herbig 2008).
In contrast, Carboniferous records from exclusively siliciclastic settings show a remarkable diversity of hexactinellid dictyosponges. These sponges were mainly recorded by Hall and Clarke (1899) from Late Devonian and early Carboniferous units of New York, Ohio, Wyoming, Indiana, and Pennsylvania in the United States, and Ontario in Canada. Most of these associations occur in the Waverly Sandstone Group, notably in shallow water siliciclastics settings. Other Carboniferous dictyosponges were reported by Rigby and Ausich (1981), also in shallow water siliciclastic facies of the Edwardsville Formation of Southern Indiana and one Carboniferous record of hexactinellids, was reported in deep black shale facies of the Carbondale Formation of northwestern Illinois, Rigby and Von Bitter (2005).
The related genus Clathrospongia in particular, was reported by Hall and Clarke (1899) from the Upper Devonian and Carboniferous Chemung Group of New York, as well as from the Waverley Group of Indiana. The genus was also recorded in the Carboniferous Bangor limestone of Alabama (Rigby and Keyes 1990).
Hence, Carboniferous hexactinellid sponges exhibit a noticeable bias to Laurussian records, Gondwana being known to a very much lesser extent.
In the context of Carboniferous Gondwanan seas, reports on sponges come from Uruguay (Kling and Reif 1969), corresponding to a poorly known hemidiscosan hexactinellid spicules and calcareous and siliceous spicules recorded in the Carboniferous of Chile (Ling et al. 1985); Morocco and Spain, being northern Gondwana (García-Bellido and Rodríguez 2005), comparatively better known.
Such evidence suggests that this group of sponges is well represented in Gondwana but has been overlooked in paleontological reports, putatively indicating an artifact in the pattern distribution (defined by a sampling or study bias). In fact, this Carboniferous scarcity contrasts with the Devonian sponge records from Gondwana which are somewhat more diverse, showing important associations mainly in Australia (Pickett 1969; Rigby 1979, 1986; Pickett and Rigby 1983). Although very limited, some Lower Devonian sponge records are also known in southwestern Gondwana (the Malvinokaffric Realm), including the hexactinellid dictyospongid Prismodyctia Hall and Clarke, 1899 from Ghana and Brasil (Saul et al. 1963), and the genus Talacastospongia Carrera and Rustán, 2016, from Argentina.
In this paleobiogeographic scenario, Minitaspongia is the first well-documented hexactinellid sponge report from the Carboniferous of Gondwana.
Hexactinellid sponges are commonly considered to be associated with deep-water settings. This view is probably related to their recent distribution and the abundant occurrences of early-middle Paleozoic species in fine-grained siliciclastic environments, although sporadic occurrences in shallow water and even reefs are known (Finks 1970; Rigby and Webby 1988; Rigby and Keyes 1990; Carrera and Rigby 2004; Carrera and Botting 2008). In contrast, late Paleozoic hexactinellids, in particular the diverse dictyosponges, show more records in shallow, sandy facies. For example, as pointed out, best records of Late Devonian and Carboniferous hexactinellids come from coarse siliciclastic sediments of North America (Hall and Clarke 1899; Rigby and Ausich 1981).
The Devonian and Carboniferous hexactinellids, especially the dictyosponges, acquired thicker and more complex skeletons, apparently to reach best adaptations to these more rough environmental settings. The spicule bundles typical of the family may have served to strengthen the skeleton (Finks 1983, 2004). However, Minitaspongia gen. nov. challenges these assumptions. Unlike the occurrences of the structurally comparable genera (i.e., Thysanodictya, Clathrospongia) in North America, Minitaspongia occurs in low-energy water siliciclastic settings, related to a cold climate, glacimarine deposits. In spite of the small size, being a young specimen, its particularly complex skeletal structure should not be invoked as a direct consequence of an adaptation to high energy, shallow water settings.
Acknowledgements
Appreciation is extended to the reviewers of the manuscript Joseph Botting (Nanjing Institute of Geology and Palaeontology, Nanjing, China) and John Pickett (Geological Survey of New South Wales, New South Wales, Australia) for theirs invaluable comments and suggestions. Diego Balseiro (CICTERRA, Universidad Nacional de Córdoba, Córdoba, Argentina) and Cyrille Prestianni (Royal Belgian Institute of Natural Sciences, Brussels, Belgium) collaborated invaluably in the field works. The company Benito Roggio S.A. and Vialmani S.A. provided lodging facilities during the field trips. We thanked Santiago Druetta (CICTERRA) for the schematic, artistic sponge reconstruction. This is a contribution to the grant PUE 2016-CONICET-CICTERRA. We acknowledge Research Council of Argentina (CONICET) for financial support to MGC and Universidad Nacional de La Rioja to JJR.
References
Aretz, M. and Herbig, H. 2008. Microbial-sponge and microbial-metazoan buildups in the Late Viséan basin-fill sequence of the Jerada Massif (Carboniferous, NE Morocco). Geological Journal 43: 307–336. Crossref
Botting, J.P. 2003. Growth patterns of Early Palaeozoic sponges. Lethaia 36: 41–52. Crossref
Botting, J.P. and Muir, L.A. 2013. Spicule structure and affinities of the Late Ordovician hexactinellid-like sponge Cyathophycus loydelli from the Llanfawr Mudstones Lagerstӓtte, Wales. Lethaia 46: 454–469.
Caminos, R., Fauqué, L., Cingolani, C., Varela, R., and Morel, E. 1993. Estratigrafía y estructura del Devónico-Carbonífero en el sector septentrional de la sierra de la Punilla, Precordillera de la Rioja y San Juan. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 2: 31–41.
Carrera, M.G. and Botting, J. 2008. Evolutionary history of Cambrian spiculate sponges: implication for the Cambrian evolutionary fauna. Palaios 23: 124–138. Crossref
Carrera, M.G. and Rigby, J.K. 2004. Sponges. In: B.D. Webby, M.L. Droser, F. Paris, and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, 102–111. Columbia University Press, New York.
Carrera, M.G. and Rustán, J.J. 2016. The new genus Talacastospongia: insights on the first record of a Devonian sponge from South America. Journal of Paleontology 89: 912–919. Crossref
Caster, K.E. 1939. Siliceous sponges from Mississippian and Devonian strata of the Penn-York Embayment. Journal of Paleontology 13: 1–20.
Croneis, C. and Toomey, D.F. 1965. Gunsight (Virgilian) wewokellid sponges and their depositional environment. Journal of Paleontology 39: 1–16.
Daemon, R.F. 1974. Palinomorfos-Guias do Devoniano Superior e Carbonifero Inferior das Bacias do Amazonas e Parnaiba. Anais da Academia Brasileira de Ciencias 46: 549–587.
Dilliard, K.A. and Rigby, J.K. 2001. The new demosponges, Chaunactis olsoni and Haplistion nacoense, and associated sponges from the Pennsylvanian Naco Formation, central Arizona. Brigham Young University Geology Studies 46: 1–11.
Fauqué, L.E. and Limarino, C.O. 1991. El Carbonífero de Agua de Carlos (Precordillera de La Rioja), su importancia tectónica y paleoambiental. Revista de la Asociación Geológica Argentina 46: 103–114.
Finks, R.M. 1960. Late Paleozoic sponge faunas of the Texas region: The siliceous sponges. American Museum of Natural History Bulletin 120: 1–160.
Finks, R.M. 1970. The evolution and ecologic history of sponges during Palaeozoic times. Symposium of the Zoological Society of London 25: 3–22.
Finks, R.M. 1983. Fossil Hexactinellida. In: T.W. Broadhead (ed.), Sponge and Spongiomorphs. Notes for a short course (organized by Rigby J.K. and C.W. Stearn). Studies in Geology 7: 101–115.
Finks, R.M. 2004. Ecology and Paleoecology of sponges In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology, Pt. E. Porifera (revised) 2, 244–260. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Finks, R.M. and Rigby, J.K. 2004. Paleozoic Hexactinellids. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology, Pt. E. Porifera (revised) 3. E319–E556. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
García-Bellido, D.C. and Rigby, J.K. 2004. Devonian and Carboniferous sponges from Spain. Journal of Paleontology 78: 431–455. Crossref
García-Bellido, D.C. and Rodriguez, S. 2005. Palaeobiogeographical relationships of poriferan and coral assemblages during the late Carboniferous and the closure of the western Palaeotethys Sea-Panthalassan Ocean connection. Palaeogeography, Palaeoclimatology, Palaeoecology 219: 321– 331. Crossref
García-Bellido, D.C., Senowbari-Daryan, B., and Rigby, J.K. 2004. Cystothalamia vandegraaffi new species and other sphinctozoan sponges from the Upper Carboniferous of Spain. Journal of Paleontology 78: 1037–1055. Crossref
Girty, G.H. 1911. On some new genera and species of Pennsylvanian fossils from the Wewoka Formation of Oklahoma. Annals of the New York Academy of Science 21: 119–156. Crossref
González, C.R. and Bossi, G. 1987. Descubrimiento del Carbónico inferior marino al oeste de Jagüel, La Rioja. 4º Congreso Latinoamericano de Paleontología (Santa Cruz de la Sierra). Actas 2: 713–729.
Hall, J. 1863 Observations upon the genera Uphantaenia and Dictyophyton with notice of some species from the Chemung Group of New York and the Waberly sandstones of Ohio. New York State Cabinet 16: 84–91.
Hall, J. 1882. Notes on the Family Disctyospongiidae. Issued in advanced of the 35th Annual Report of the New York State Museum of Natural History, 1884. Pl. 17–20 (with explanations).
Hall, J. 1884. Description of the species of fossil reticulate sponges, constituting the family Dictyospongidae. Thirty-fifth Annual Report on the New York State Museum of Natural History, 465–481. Wynkoop Hallenbeck Crawford Company v. Gaines, Albany.
Hall, J. 1890. Fossil Dictyospongidae of the Devonian and Carboniferous formations, new forms of Dictyospongidae from rocks of the Chemung Group. Ninth Annual Report New York State Geologist, 56–60. Wynkoop Hallenbeck Crawford Company v. Gaines, Albany.
Hall, J. and Clarke, 1899. A memoir on the Paleozoic reticulated sponges constituting the family Dictyospongidae. Memoir of the New York State Museum 2 (for 1898): 1–350.
Hurcewicz, H. and Czarniecki, S. 1985. Lyssakide sponges from the Carboniferous limestone and the Culm of southern Poland and their environmental differentiation. Annales Societatis Geologorum Poloniae 55: 333–354.
Isaacson, P.E., Díaz-Martínez, E., Grader, G.W., Kalvoda, J., Babek, O., and Devuyst, F.X. 2008. Late Devonian–earliest Mississippian glaciation in Gondwanaland and its biogeographic consequences. Palaeogeography, Palaeoclimatology, Palaeoecology 268: 126–142. Crossref
King, R.H. 1938. Pennsylvanian sponges of north-central Texas. Journal of Paleontology 12: 498–504.
Kling, S.A. and Reif, W. 1969. The Paleozoic history of amphidisc and hemidisc sponges: new evidence from the Carboniferous of Uruguay. Journal of Paleontology 43: 1429–1434.
Limarino, C.O. and Spalletti, L.A. 2006. Paleogeography of the upper Paleozoic basins of southern South America: An overview. Journal of South American Earth Sciences 22: 134–155. Crossref
Ling, H.Y., Forsythe, R.D., and Douglass, R.C. 1985. Late Paleozoic microfaunas from southernmost Chile and their relation to Gondwanaland forearc development. Geology 13: 357–360. Crossref
Pickett, J. 1969. Middle and Upper Palaeozoic sponges from New South Wales. Geological Survey of New South Wales Palaeontological Memoir 16: 1–24.
Pickett, J. and Rigby, J.K. 1983. Sponges from the Early Devonian Garra Formation, New South Wales. Journal of Paleontology 57: 720–741.
Prestianni, C., Rustán, J.J., Balseiro, D., Vaccari, E., Sterren, A.F., Steemans, P., and Astini, R.A. 2015. Early seed plants from Western Gondwana: Paleobiogeographical and ecological implications based on Tournaisian (Lower Carboniferous) records from Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 417: 210–219. Crossref
Reid, R.E.H. 1958. A monograph of the Upper Cretaceous Hexactinellida of Great Britain and Northern Ireland. Part 1. Palaeontographical Society of London, Monographs 111: I–XIV.
Rigby, J.K. 1970. Two new Upper Devonian hexactinellid sponges from Alberta. Journal of Paleontology 44: 7–16.
Rigby, J.K. 1979. Patterns in Devonian sponge distribution. In: M.R. House, C.T. Scrutton, and M.G. Bassett (eds.), The Devonian System. Special Papers in Palaeontology 23: 225–228.
Rigby, J.K. 1986. Late Devonian sponges of Western Australia. Report of the Geological Survey of Western Australia 18: 1–59.
Rigby, J.K. and Ausich, W. 1981. Lower Mississippian sponges from the Edwarsdville Fomation, Soutern Indiana. Journal of Paleontology 55: 370–382.
Rigby, J.K. and Church, S.B., 1993. Wewokella and other sponges from the Pennsylvanian Minturn Formation of North-Central Colorado. Journal of Paleontology 67: 909–916. Crossref
Rigby, J.K. and Keyes, R., Jr. 1990. First Report of Hexactinellid Dictyosponges and other sponges from the Upper Mississippian Bangor Limestones, Northwestern Alabama. Journal of Paleontology 64: 886–897. Crossref
Rigby, J.K. and Manger, W.L. 1994. Morrowan lithistid demosponges and hexactinellids from the Ozark Mountains of northwestern Arkansas. Journal of Paleontology 68: 734–746. Crossref
Rigby, J.K. and Mapes, R.H. 2000. Some Pennsylvanian and Permian sponges from southwestern Oklahoma and north-central Texas. Brigham Young University Geology Studies 45: 25–67.
Rigby, J.K. and Mundy, D.J.C. 2000. Lower Carboniferous sponges from the Craven reef belt of North Yorkshire. Proceedings of the Yorkshire Geological Society 53: 119–128. Crossref
Rigby, J.K. and Moyle, R.W. 1959. Some Mississippian and Pennsylvanian sponges from Utah. Journal of Paleontology 33: 399–403.
Rigby, J.K. and Von Bitter, P. 2005. Sponges and associated fossils from the Pennsylvanian Carbondale Formation of Northwestern Illinois. Journal of Paleontology 79: 460–469. Crossref
Rigby, J.K. and Washburn, A.T. 1972. A new hexactinellid sponge from the Mississippian–Pennsylvanian Diamond Peak Formation in eastern Nevada. Journal of Paleontology 46: 266– 270.
Rigby J.K. and Webby, B.D. 1988. Late Ordovician sponges from the Malongulli Formation of central New South Wales. Palaeontographica Americana 56: 1–147.
Rigby, J.K., Chamberlain, C.K., and Black, B.A. 1970. Mississippian and Pennsylvanian sponges from the Ouachita Mountains of Oklahoma. Journal of Paleontology 44: 816– 832.
Rigby, J.K., Keyes, R., and Horowitz, A. 1979. Two new Mississippian sponges from Northwestern Alaska. Journal of Paleontology 53: 709–719.
Rigby, J.K., Waters, J.A., Maples, C.G., Lane, N.G., Liao, Z.-T., and Liu, L.-J. 1999. Hexactinellid and calcareous sponges from the Pennsylvanian (Moscovian) Dikanre Formation from Xinjiang-Uygur Autonomous Region, northwestern People’s Republic of China. Journal of Paleontology 73: 380–388. Crossref
Saul, J.M., Boucot, A.J., and Finks, R.M. 1963. Fauna of the Accraian Series (Devonian of Ghana) including a revision of the gastropod Plectonotus. Journal of Paleontology 37: 1042–1053.
Sollas, W.J. 1875. Sponges. In: T.S. Baynes (ed.), Encyclopedia Britannica, Ninth Edition, 427–446. Adam and Charles Black, Edinburgh.
Van de Graaff, W.J.E. 1969. Carboniferous Sphinctozoa from the Cantabrian mountains, Spain. Leidse Geologische Mededelingen 42: 239–257.
Young, J. and Young, J. 1877. On a Carboniferous Hyalonema and other sponges from Ayrshire. Annals and Magazine of Natural History, Series 4 (20): 425–432.
Acta Palaeontol. Pol. 63 (1): 63–70, 2018
https://doi.org/10.4202/app.00403.2017