
Palaeobiology of orthothecide hyoliths from the Cambrian Manto Formation of Hebei Province, North China
HAIJING SUN, JOHN M. MALINKY, MAOYAN ZHU, and DIYING HUANG
Sun, H., Malinky, J.M., Zhu, M., and Huang, D. 2018. Palaeobiology of orthothecide hyoliths from the Cambrian Manto Formation of Hebei Province, North China. Acta Palaeontologica Polonica 63 (1): 87–101.
Newly discovered specimens of the orthothecide hyoliths in Hebei Province of North China reveal new data on diversity, skeletal microstructure and palaeoecology of the order Orthothecida. Decoritheca cyrene possesses a well-defined skeletal structure consisting of rows of tubercles in the shell wall, which correspond to dimples on the internal surface of the conch. We describe a new species Cupitheca convexa sp. nov. characterised by a planar and interior convex operculum with a pair of bilobate cardinal processes, which differs from the more widespread C. holocyclata in opercular morphology and overall ornamentation of the conch, highting the significance of operculum in the classification of hyoliths. First discovery of the presumed unusual protoconch of C. convexa sp. nov. implies a possible lecithotrophic development to adapt to nutrient-fluctuant environments. Pitting structure on the operculum and shell as well as lamellar- fibrillar structure on the internal mould of C. convexa sp. nov. are observed herein.
Key words: Hyolitha, Orthothecida, ontogeny, larval conch, skeletal microstructure, Cambrian, China.
Haijing Sun [hjsun@nigpas.ac.cn] (corresponding author) and Diying Huang [dyhuang@nigpas.ac.cn] (corresponding author), State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China.
John M. Malinky [jmalinky@sbcglobal.net], Department of Physical Science, San Diego City College, 1313 Park Boulevard, San Diego, California, 92101, USA.
Maoyan Zhu [myzhu@nigpas.ac.cn], State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China, and College of Earth Sciences, University of Chinese Academy of Sciences, Beijing 100049, China.
Received 28 July 2017, accepted 20 October 2017, available online 29 January 2018.
Copyright © 2018 H. Sun et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Orthothecide hyoliths are one of the two main groups within the Hyolitha. They are most abundant and diverse worldwide in the Cambrian, with a progressive decline until their extinction in the Middle Devonian (Malinky 2009). They are characterized by a calcareous conical shell with a generally planar aperture and a retractable planar or slightly concave operculum. The boundary between the dorsal and ventral portions of the operculum is indistinct, although near the dorsal margin the operculum possesses an elevated knob-like structure referred to as the summit. The interior is ordinarily smooth except for a pair of cardinal processes that radiate from below the summit. The morphology of the gut tract as well as its sediment infilling suggests a benthic, deposit feeding life habit of some orthothecides (Marek 1967; Runnegar et al. 1975; Devaere et al. 2014), rather than suspension feeding as suggested for the hyolithides (Marek et al. 1997; Galle and Parsley 2005; Moysiuk et al. 2017).
Orthothecide hyoliths have been known from China for more than a century (Walcott 1905, 1911, 1913) with additional reports by Resser and Endo (1937) and Endo (1937). Interest in the group was revived by Qian (1977, 1978, 1983) and Qian et al. (1979) with more taxa being described (Xiao and Zhou 1984; Malinky 1987; Qian 1989; Jiang 1992; Xiao and Duan 1992; Qian et al. 2003; Steiner et al. 2007; Parkhaev and Demidenko 2010; Pan et al. 2015). Here we report a new occurrence of hyoliths from the Cambrian of North China. The specimens described here derive from the Cambrian Manto Formation (around the boundary of Cambrian Stage 4/ Stage 5) in the Zuojiawu section of Tangshan City in Hebei Province, North China (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app63-Sun_etal_SOM.pdf). Two species are recognised at the locality: Cupitheca convexa sp. nov. and Decoritheca cyrene (Walcott, 1905). Cupitheca Duan in Xing et al., 1984 is characterised by a generally tubular conch with a bluntly rounded apical termination and circumferential furrow. It ranges from the Cambrian Fortunian to Cambrian Stage 5 (Xing et al. 1984; Duan 1984; Bengtson et al. 1990; Pan et al. 2015) (see SOM 2). Owing to its unusual decollating ontogeny and lack of a convincing operculum, Cupitheca has long been excluded from the hyoliths and treated as a problematica form of uncertain affinity (Bengtson et al. 1990; Demidenko 2001; Wrona 2003; Malinky and Skovsted 2004; Pan et al. 2015), whereas others suggest a hyolith affinity and more close to Orthothecida (Kruse et al. 2004; Skovsted et al. 2016). The present material tends to support orthothecide affinity of Cupitheca.
Decoritheca is a globally distributed orthothecide ranging from the Cambrian Stage 3 to Stage 10 (Qian 1989; Qian and Xiao 1995; Malinky 2014; see also SOM 2), rendering it one of the longest ranging orthothecide genera. Here we report an unusual skeletal microstructure seen in D. cyrene that consists of tubercles covered the surface of internal mould of the shell.
Institutional abbreviations.—NIGPAS, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, China.
Geological setting
The hyolith specimens described in this paper were collected from the purplish-red shale of the Manto Formation at the Zuojiawu section, Fengrun District of Tangshan City, Hebei Province, located in the northeastern region of the North China Platform (see SOM 1). The Manto Formation is a heterogeneous succession characterised by purplish-red shale intercalated with limestone, marlstone, dolostone, and sandstone, which was generally considered to have been deposited in tidal flat or lagoon environments under arid conditions (Xiang et al. 1999). It crops out extensively in the northern and eastern parts of the North China Platform. The concept of this formation was revised and restricted to the lower part of the originally described unit (Lu and Dong 1953; Lu 1962). Subsequently, Xiang et al. (1999) reverted to the initial definition of the Manto Formation, which is followed herein. It includes nine trilobite zones, in ascending order, they are Redlichia nobilis, Qiaotouaspis, Yaojiayuella, Shantungaspis, Hsuchuangia–Ruichengella, Ruichengaspis, Sunaspis, Poriagraulos, and Bailiella–Lioparia zones (Wang et al. 2014), ranging from Cambrian Series 2 Stage 4 to Series 3 Stage 5 (Lungwangmiaoan to Hsuchuangian in age) based on Peng (2009) and Peng et al. (2012). The hyolith-yielding interval contains abundant trilobites including Luaspides lingyuanensis Duan, 1966 indicative of the Shantungaspis Zone (Duan et al. 2005), which was recently correlated with the Cambrian Stage 5 (Taijiangian) (Peng 2009; Wang et al. 2014). However, the precise age of the Shantungaspis Zone is uncertain, it could be late Stage 4 or early Stage 5 (Shanchi Peng, personal communication 2017). The hyolith-bearing interval also yields fragments of anomalocaridids and brachiopods (Huang et al. 2012) as well as other problematic fossils of uncertain affinity.
Material and methods
The specimens analysed herein were collected from the Manto Formation exposures of Tangshan, and the fossils are usually preserved as internal moulds with limited skeletal material. All specimens described here are deposited in NIGPAS under repository numbers 166329–166377.
Two types of preservation are recognised among the skeletal fossils from the Manto Formation. One style is represented by dense concentrations of shells preserved on the bedding planes (Fig. 1). These fossils consist of opercula, discarded segments and conchs of C. convexa sp. nov. in association with a few shells of D. cyrene (Fig. 1B, D), and numerous trilobite fragments and bioclasts of unidentified fossils. This assemblage is preserved as a non-size-sorted association that has a chaotic orientation without individuals being fully intact, suggesting rapid deposition from storm-induced density currents (O’Brien et al. 1980; Brett and Baird 1986; Simões and Aceñolaza 2009; Cherns et al. 2011). Mineral structures (Fig. 2A1–A3) and elemental analyses (Fig. 2A4) indicate that the pigment mineral hematite (diameter of hematite rosette 1–4 μm) accounts for the reddish colour of the shale and fossils (Ziegler and McKerrow 1975; Turner 1979), specifically the coating of the hyolith conchs (Fig. 2B). The presence of halite pseudomorphs (Fig. 2C1) and calcite impressions (Fig. 2C2) support the inference of an arid inner shelf/ lagoonal-mud flat depositional setting for the reddish shale (Xiang et al. 1999).

Fig. 1. Aggregations of orthothecide hyoliths preserved in purplish-red shales from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A. NIGPAS 166336. B. NIGPAS 166345. C. NIGPAS 166334. D. NIGPAS 166341b. E. NIGPAS 166346. Scale bars 5 mm, except E, 2 mm.
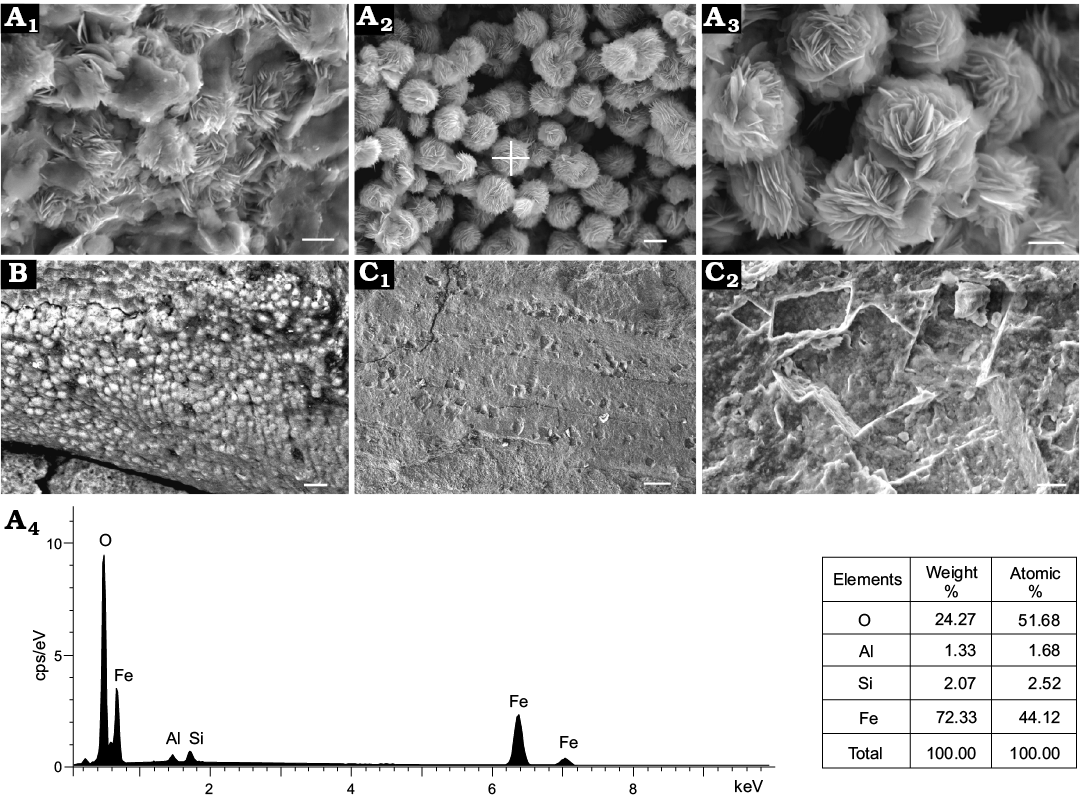
Fig. 2. Scanning electron micrographs of shell and shale fabrics as well as mineral elemental analyses of matrix from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A. NIGPAS 166334, fine grained clay minerals and hematite crystals in the matrix (A1). Tabular crystals of hematite forming rosette-shaped aggregations in the matrix (A2, A3); the cross in A2 shows the location of elemental analysis (A4). B. NIGPAS 166339a, skeleton coated with hematite. C. NIGPAS 166337, halite pseudomorphs distributed on the surface of undetermined fossil (C1), calcite imprints in the matrix (C2). Scale bars: A1, A2, C2, 2 μm; A3, 1 μm; B, 10 μm; C1, 50 μm.
A second type of preservation is characterized by individual conchs of orthothecides without opercula attached (Fig. 3B3, F, H), but associated with both fragmented and rare articulated trilobites, appendages and mouthparts of anomalocaridids, disarticulated brachiopods with convex-up oriented single valves, sponges and coprolitic material in purplish-red shales. No preferred orientation can be observed in this case. These fossil associations represent time-averaged assemblages (Zhao et al. 2009, 2014) and a few shells covered with algae (Fig. 3I) suggest a period of exposure on the seafloor prior to burial.
Specimens were examined using SU3500 Scanning Electron Microscopy (SEM) with Electron Dispersive X-ray (EDX) under UVD and BSE-COMP modes to observe the mineral and skeletal morphology in the Public Technology Service Center of NIGPAS. Illustrated fossils were imaged by Canon EOS 5D MarkIII with a Canon MP-E 65mm f/2.8 Macro lens and some of the specimens were coated with magnesium oxide to reduce reflection. Digital macrophotographs were edited and rendered using Adobe Photoshop CS6 and Coreldraw X4 softwares. Fossils were measured by using Digimizer software.
Systematic palaeontology
Phylum uncertain
Remarks.—Affinity of hyoliths has long been controversial. Previous works on the earliest developmental pattern of hyoliths and molluscs demonstrated a similarity between these groups to each other and to the annelids (e.g., Martí Mus et al. 2014). However, other studies of internal anatomy tend to favour a close relationship between hyoliths and sipunculans (Pojeta 1987; Sun et al. 2016). Recently, a novel idea that hyoliths are lophophorates close to the brachiopod-phoronid clade has been proposed by Moysiuk et al. (2017). However, the phylogenetic position of the hyoliths within the Lophotrochozoa, and whether these organisms constitute a monophyletic group remain open questions.
Class Hyolitha Marek, 1963
Order Orthothecida Marek, 1966
Family Novitatidae Sysoyev, 1968 (emended Malinky 1987)
Genus Decoritheca Sysoyev, 1972
Type species: Hyolithus (Orthotheca) excavatus Holm, 1893; middle Cambrian, Sweden.
Diagnosis.—See Berg-Madsen and Malinky (1999).
Remarks.—Decoritheca is morphologically similar to Contitheca Sysoyev, 1958, but they are separated by the prominent ridge present on the dorsum of the latter, giving rise to a heart-shaped cross section. The former possesses a rounded dorsum and lacks the middle dorsal ridge, creating a reniform cross section. Specimens described herein match well the traits of Decoritheca and are therefore assigned to that genus. Decoritheca also resembles Nephrotheca Marek, 1966 in cross section, but differs from the latter in solely having transverse lines (Malinky 1987), although, to some degree, ornamentation may not be the best criterion to differentiate taxon ranking above species. So far no operculum for Decoritheca has been documented. Decoritheca cyrene was described from the “middle Cambrian” (Cambrian Series 3) Changhsia Formation and the “upper Cambrian” (Cambrian Furongian) Chaomitien Formation (Walcott 1905; Malinky 1987); the long span of the species may also reflect a taxonomic problem. Until better fossil material is available to address the relationship between Nephrotheca, Decoritheca, and some other similar taxa, specimens described below are provisionally placed under Decoritheca.
Stratigraphic and geographic range.—Cambrian Series 2 Stage 3 to Furongian Stage 10. Distributed in China, Greenland, Sweden, British Columbia, Texas, Montana, possibly South Dakota, and New York State (Malinky 1987; 1990; 2014; see also SOM 2).
Decoritheca cyrene (Walcott, 1905)
Fig. 3.
1905 Orthotheca cyrene sp. nov.; Walcott 1905: 19.
1905 Orthotheca cyrene dryas sp. nov.; Walcott 1905: 19.
1913 Orthotheca cyrene Walcott, 1905; Walcott 1913: 94, pl. 5: 21, 21a.
1913 Orthotheca cyrene dryas Walcott, 1905; Walcott 1913: 93, pl. 5: 22, 22a–c.
1946 Hyolithes (Orthotheca) cyrene (Walcott), 1905; Sinclair 1946: 75.
1946 Hyolithes (Orthotheca) dryas (Walcott), 1905; Sinclair 1946: 75.
1987 Decoritheca cyrene (Walcott, 1905); Malinky 1987: 948–951, figs. 2.4, 6.1–6.6, 6.14.
Material.—Thirty-seven specimens (NIGPAS 166329–35, 166337, 166338, 166341, 166344, 166349–74) preserved as internal moulds of conchs.
Diagnosis.—See Malinky (1987: 948).
Description.—Conch straight with a planar (orthogonal) aperture, length of conchs ranges from 7.1–20.1 mm, and apertural width ranges from 2.9–5.5 mm. The dorsum is broadly rounded and covered with rows of tubercles (Fig. 3C), with diameters of 6–10 μm. Venter developed with a prominent longitudinal median concavity, becoming shallower from aperture to apex. Sub-rounded ridges adjacent to the concavity grade into narrowly rounded lateral margins (Fig. 3B1, B3, D, F, H) and lateral margins are straight in dorsal view. Ventral external surface is covered with closely spaced transverse lines (Fig. 3B2). The morphology of dorsum and venter gives rise to a reniform cross section, but only one specimen displays the full image of transverse section (Fig. 3E), the apertural height is 0.7 mm, apertural width is 1.4 mm, and apertural width/height ratio is 2. Conchs terminate in pointed apex, and angle of divergence at apex ranges from 15–18°. Operculum and helens unknown.
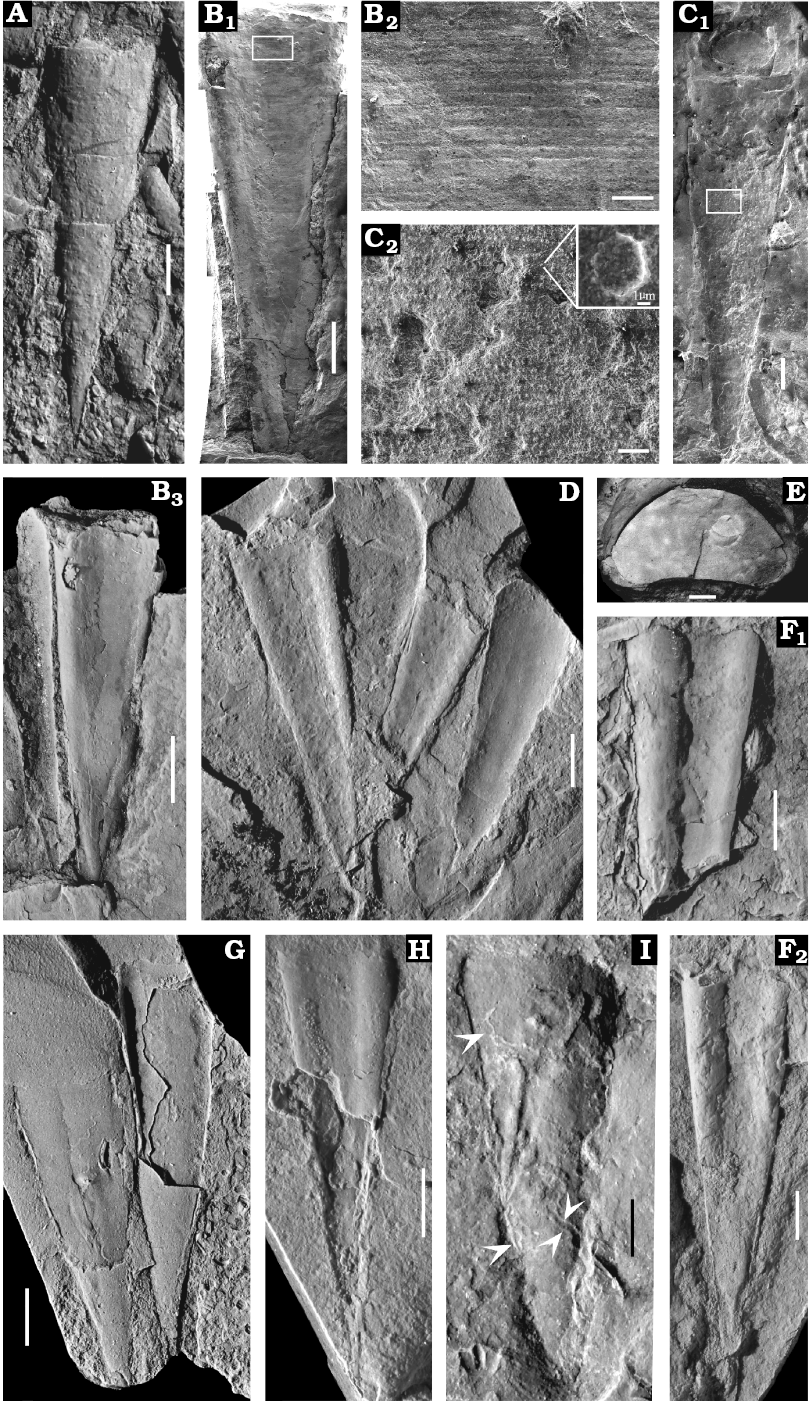
Fig. 3. Novitatid hyolith Decoritheca cyrene (Walcott, 1905) from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A. NIGPAS 166338 in dorsal view. B. NIGPAS 166335 in ventral (B1) and ventro-lateral (B3) views; detail showing transverse lines on the external surface of venter (B2). C. NIGPAS 166337 in dorsal view, conch of Decoritheca cyrene with operculum of Cupitheca convexa sp. nov. in apertural region (C1); rows of tubercles (C2), inset shows enlarged individual tubercle with nearly flat top. D. NIGPAS 166329 in ventral view. E. NIGPAS 166341b, aperture with reniform cross section. F. NIGPAS 166331 in ventral view; F1, incomplete shell showing well developed median concavity and adjacent ridges; F2, nearly complete shell with broken aperture and adapically shallowing median concavity. G. NIGPAS 166330, both shells display partial venter and partial dorsum from dorsal view. H. NIGPAS 166333 in ventral view. I. NIGPAS 166332 in dorsal view, white arrows indicate algae. Scale bars: A, D, B3, F–I, 2 mm; B1, C1, 1 mm; B2, 100 μm; C2, 20 μm; E, 200 μm.
Remarks.—This species bears resemblance to D. lineatula (Holm, 1893) from Sweden, but the presence of only longitudinal sculpture on the latter serves to distinguish it from the former. D. cyrene also bears limited resemblance to the Swedish D. excavata (Holm, 1893), but the two species can be differentiated by the absence of rounded ridges at the ventral lateral margins in the latter.
One specimen of this species (Fig. 3C1) appears to preserve a conch with an unattached operculum near the aperture. Given that the operculum is nearly circular but the cross section of D. cyrene is reniform, it is unlikely that the conch and operculum represent the same taxon. It is more likely that these specimens are coincidentally located next to each other as result of movement by currents or perhaps bioturbation. Circular opercula such as these are usually found in clusters of C. convexa sp. nov. shells, and therefore we suggest that these circular or oval opercula more reasonably belong to C. convexa sp. nov.
Stratigraphic and geographic range.—Cambrian Series 3 Stage 5 to Furongian Stage 10. The Manto Formation of Tangshan, Hebei Province (herein), the Chaomitien and Changhsia formations of Yantai, Shandong Province (Walcott 1905), and the Changhsia Formation in Anhui Province, China (Qian and Xiao 1995); Snowy Range Formation of Gallatin, Montana; Wilberns Formation of Burnet County, Texas (Malinky 1987; see also SOM 2).
Family Cupithecidae Duan, 1984
Genus Cupitheca Duan in Xing et al., 1984
Type species: Cupitheca brevituba Duan in Xing et al., 1984, lower Cambrian, South China.
Diagnosis.—See Bengtson et al. (1990: 203).
Remarks.—For detailed discussion and taxonomic history see Malinky and Skovsted (2004: 568–569), Pan et al. (2015: 388) and Skovsted et al. (2016: 125).
Stratigraphic and geographic range.—Cambrian; Terreneuvian Fortunian to Series 3 Stage 5; since the biostratigraphic correlation of “Lower Limestone” (= Maseong Formation) is not clear (Lee 2008), the age of C. cf. mira reported by Lee (2008) should be questioned. Distributed in China (Pan et al. 2015), Korea (Lee 2008), Australia (Kruse et al. 2004; Skovsted et al. 2016), Antarctica (Wrona 2003), Greenland (Malinky and Skovsted 2004), Newfoundland (Skovsted and Peel 2007), Spain (Jensen et al. 2010), India (Gilbert et al. 2016; Hughes 2016), possibly Kazakhstan and England (Malinky and Skovsted 2004; see also SOM 2).
Cupitheca convexa sp. nov.
Figs. 4–8.
Etymology: From Latin convexus, in reference to the convex nature of the interior of the operculum at the cardinal process area.
Type material: Holotype NIGPAS 166339a (Fig. 6D); Paratypes NIGPAS 166334, 166336–43 (Figs. 4, 6A–C) from the type locality.
Type locality: Zuojiawu section, Fengrun District of Tangshan City, Hebei Province, North China.
Type horizon: The purplish-red shales of the Manto Formation, Cambrian Stage 4/5.
Material.—Thirteen specimens in total, preserved as clusters of moulds of conchs and opercula (NIGPAS 166336–39a, b, 166340, 166341a, b, 166342–47), and 3 specimens (NIGPAS 166375–77) preserved as inarticulate shells. All are decalcified.
Diagnosis.—Cupitheca with interior surface of operculum convex near cardinal processes; cardinal processes are moderately well-developed and bilobate; conch cyrtoconic; ornamentation on conch is merely transverse lines, and both interior and exterior surfaces of operculum covered by concentric ribs.
Description.—Conchs cyrtoconic with pronounced curvature, degree of curvature slightly varies in different specimens (Fig. 4A, C–E); transverse lines on exterior surface (Figs. 4A3, 5A7) with intervening spaces 2–15 μm wide; cross section circular to oval (Fig. 4B); apical angle of divergence small (14–16°); sculpture at apical termination varies (Fig. 5A4, A5, B), with some having radial lines that probably represent the original microstructure of the shell (Fig. 5A4, A5), while others possess a smooth terminus with no lines or other features (Fig. 5A1–A3, A6, A7, C3); and still others have irregularly radiating, branching ornamentation (Fig. 5B), which may be an artefact of preservation. A circumferential furrow separates the apical region from the remainder of the shell.
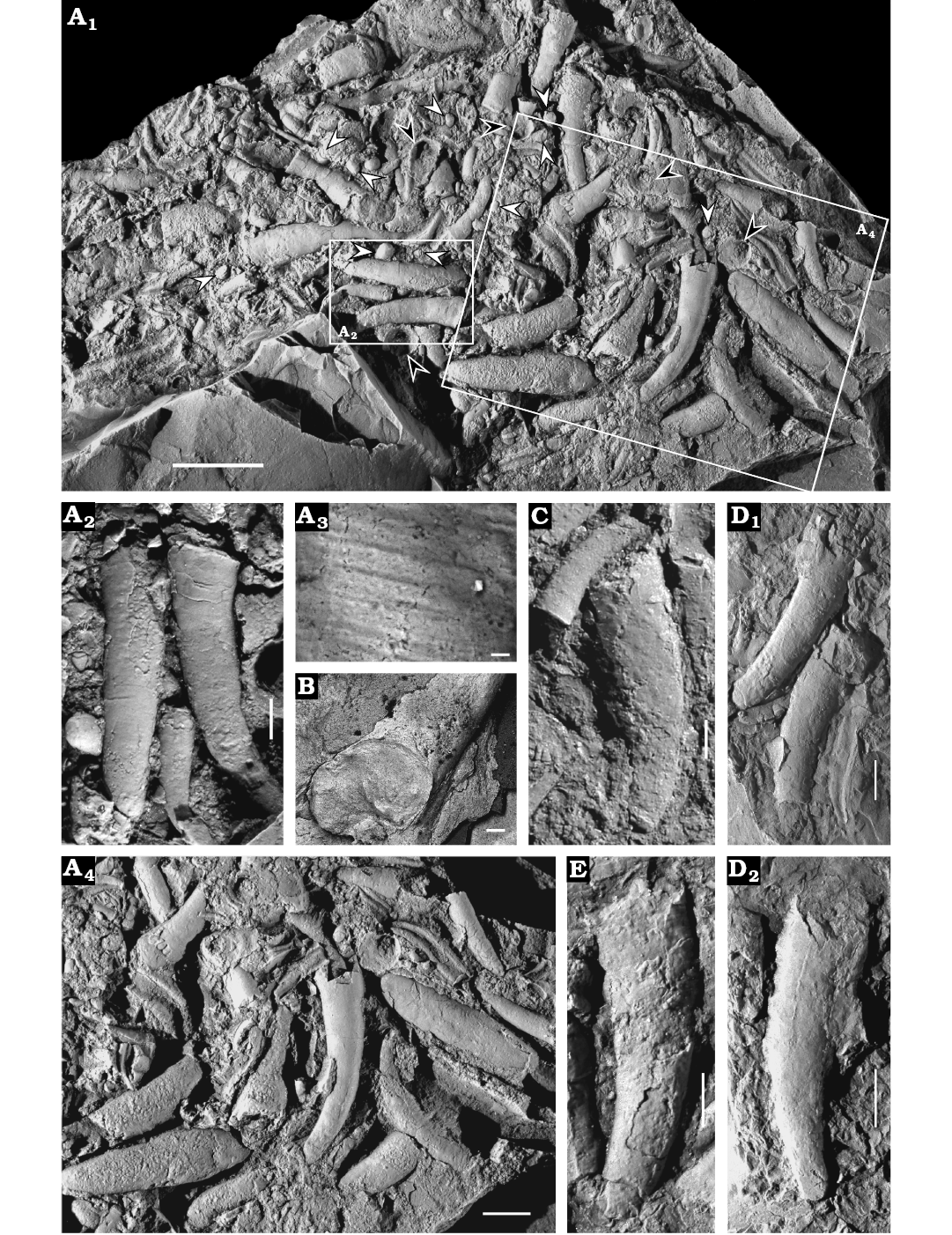
Fig. 4. Cupithecid hyolith Cupitheca convexa sp. nov. from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A. NIGPAS 166334, accumulation of conchs and opercula (A1), white arrows suggest discarded cylindrical segments, black arrows indicate opercula; three almost complete shells with apical parts buried in the matrix (A2); transverse lines on the dorsal surface of conch (A3); aggregation of skeletal parts with chaotic orientation (A4). B. NIGPAS 166336, incomplete conch shows full aperture. C–E. More or less complete conchs. C. NIGPAS 166338. D. NIGPAS 166340, two adjacent shells, the left one displays full apex (D1); single conch with partial broken aperture (D2). E. NIGPAS 166341b. Scale bars: A1, 5 mm; A2, C, 1 mm; A3, 5 μm; B, 100 μm; A4, D, E, 2 mm.
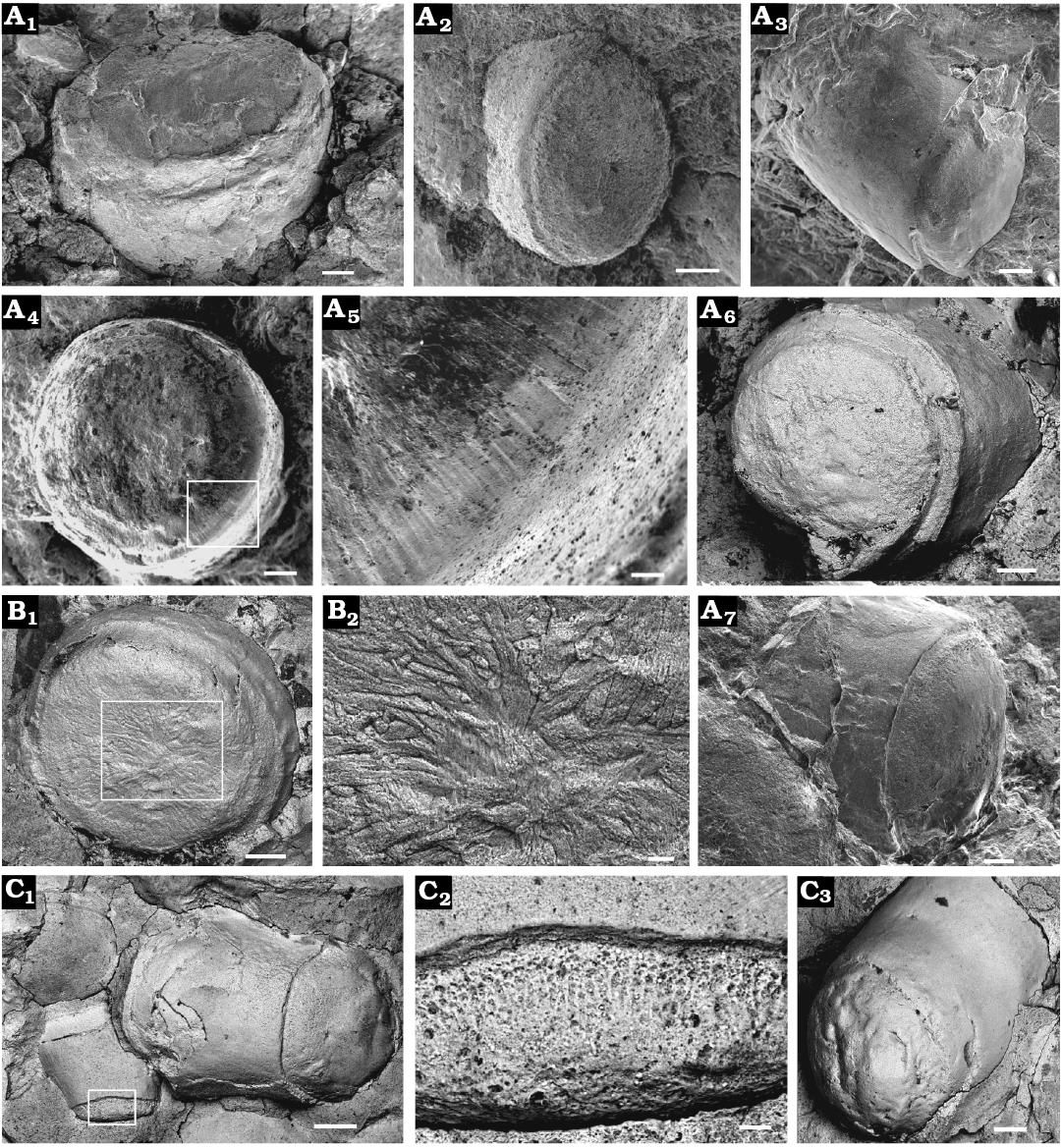
Fig. 5. Internal moulds of decollated segments of cupithecid hyolith Cupitheca convexa sp. nov. from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A. NIGPAS 166334, discarded apical segments with smooth surface (A1–A3, A6); bottom of the apex (A4); apical bottom covered with radial sculptures (A5); discarded segment preserves faint transverse lines on the surface of conch (A7). B. NIGPAS 166343, decollated segment displaying apical bottom, probable diagenetic artefacts on the bottom of apex (B1); branching ornamentation on the bottom of apex (B2). C. NIGPAS 166339a, decollated segments two segments preserved together the left one exhibits pits and longitudinal groove structures on the surface of terminal part (C1); detail (C2); discarded apical segments with smooth surface (C3). Scale bars: A1, A3, A6, A7, B1, C3, 100 μm; A2, A4, 50 μm; C1, 200 μm; A5, B2, C2, 20 μm.
Operculum circular to sub-circular or slightly oval (opercular width and length are subequal, Fig. 7A); protooperculum nearly flat (Fig. 6A2, A3) and circular (Fig. 6A, B1, B3), 250–530 μm in diameter (Fig. 7A), with a location toward the dorsal margin. Bilobate cardinal process can be observed near the dorsal margin of the operculum on the interior surface, forming a convex area and grading downward into a flattened marginal area (Fig. 6B4, C, D1, D3), impressions of cardinal processes can also be observed on the exterior of the operculum (Fig. 6A, B1–B3), which may be the result of high compression; these structures are generally rectangular in shape, and terminate at the distal end in a bluntly rounded surface. Angles of divergence of cardinal processes are 99–113° and 100° on average. Cardinal processes are covered from proximal to distal terminations with scattered sub-circular pits (shown as tubercles on the counterparts; e.g., Fig. 6D), 4–9 μm in diameter. Exterior of operculum is covered by a series of concentric lamellae or ribs (Fig. 6A1, A3, B1–B3). Interior also covered with concentric lines or ribs (Fig. 6B4, D1, D3) that correspond to those on the exterior.
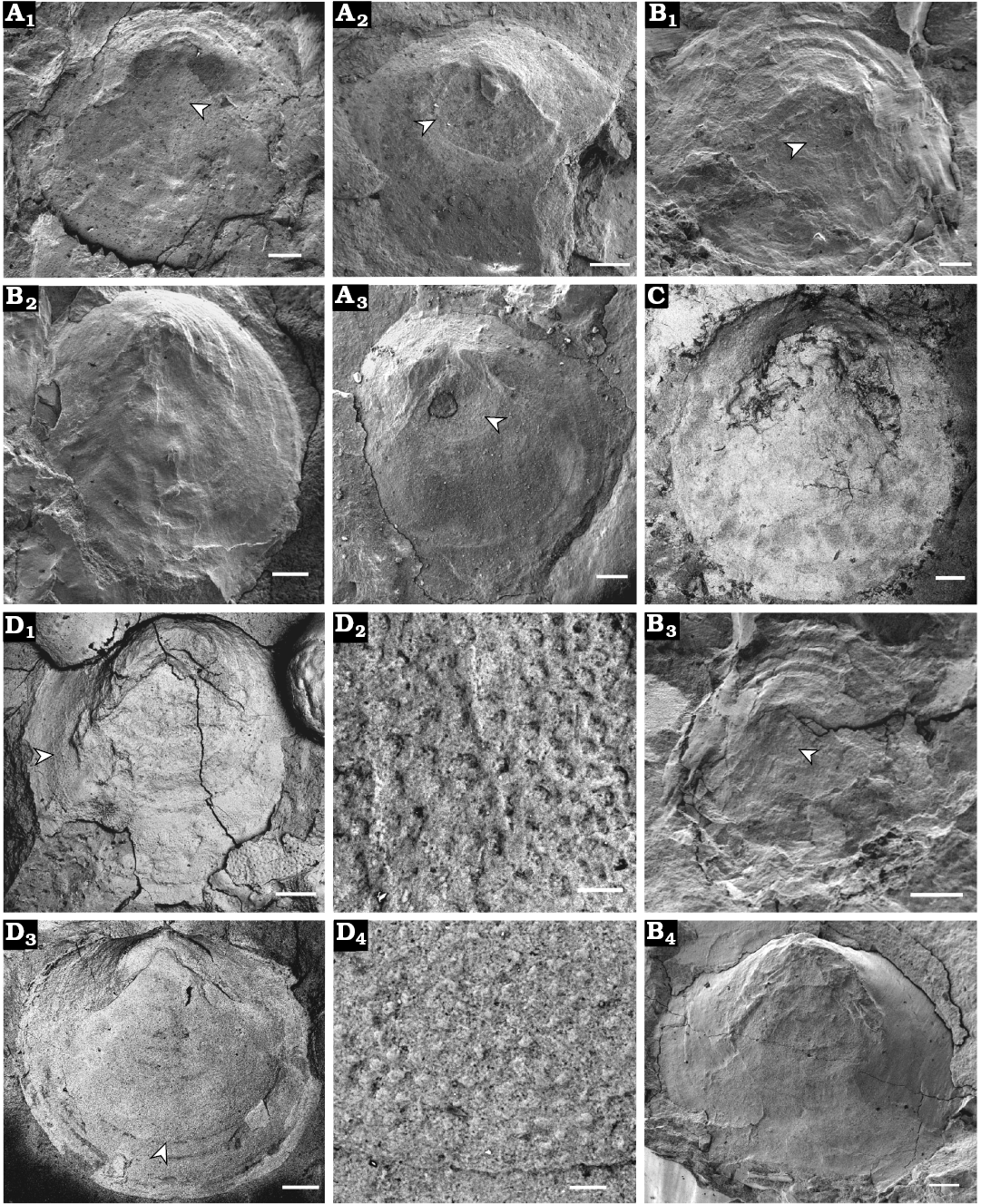
Fig. 6. Opercula of cupithecid hyolith Cupitheca convexa sp. nov. preserved as internal or external moulds, which are prepared from natural casts, collected from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A, B. Internal moulds of opercula, NIGPAS 166337 (A) and NIGPAS 166342 (B); showing exteriors covered with concentric ribs, protoopercula (indicated by white arrows) and imprints of bilobate cardinal processes (A1–A3, B1–B3); interior preserved concentric ribs and cardinal processes (B4). C, D. Counterparts of internal moulds of opercula displaying interior with imprints of bilobate cardinal processes, NIGPAS 166343 (C) and NIGPAS 166339a (D, holotype). Internal mould exhibiting interior surface of the operculum with concentric ribs and cardinal processes preserved (D1), arrow indicates the location of D2; pitting structure on the surface near opercular margin (D2); specimen showing opercular interior covered by concentric ribs and tubercles on the surface near margin (D3), arrow indicates position of D4; rounded tubercles (D4), which correspond to pits on the surface of interior of operculum, as they are shown in (D2). Scale bars: A, B2, B4, C, D1, D3, 200 μm; B1, B3, 100 μm; D2, D4, 20 μm.
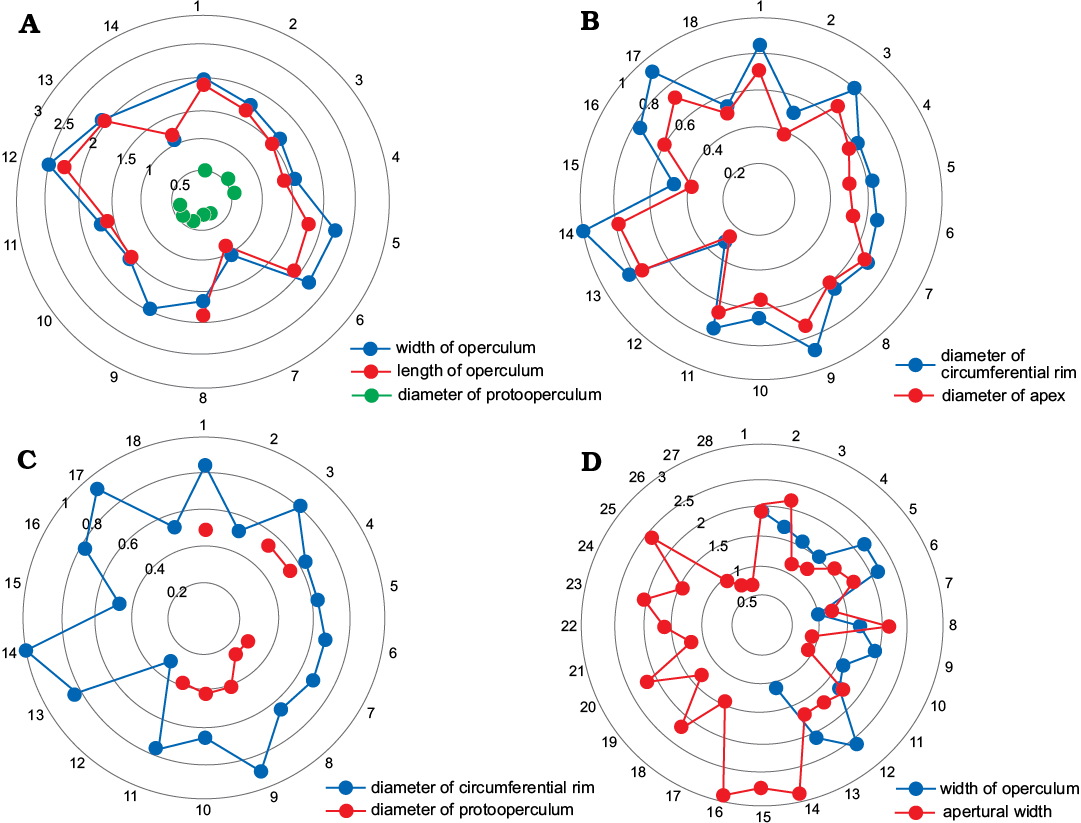
Fig. 7. Statistical distribution of different skeletal parts, junctions of the outermost ring represent the number of data, and concentric circles represent length (in mm).
Remarks.—Specimens of Cupitheca recovered from the Manto Formation on the North China Platform are distinctive from the more widely distributed and therefore better known C. holocyclata Bengtson in Bengtson et al., 1990 (Bengtson et al. 1990; Skovsted et al. 2016) in terms of ornamentation of the conch and overall shape of the operculum. C. holocyclata from Australia (Bengtson et al. 1990) and several individuals reported from North China from Cambrian Stage 4 (Skovsted et al. 2016) have a dominant transverse ornament, with minimal to subordinate longitudinal overprint. In addition, the interior of the operculum of C. holocyclata is concave, while that of C. convexa sp. nov. is convex, and cardinal processes seen in C. holocyclata appear to be more robust than those of C. convexa sp. nov.. The protooperculum of the former is a circular elevated platform with a concave base and marginal rim (Skovsted et al. 2016), which is different from the flattened and circular protooperculum in C. convexa sp. nov.. The new species is distinguished from C. costellata Xiao and Zhou, 1984 by possession of distinct longitudinal ornament in the latter. Our material also resembles C. manicae Duan, 1984 from the Lower Cambrian of Hubei Province, South China (Duan 1984) in general morphology and cross section of the shell, but the latter possesses no ornament on the conch (possibly a taphonomic bias) and no known operculum, which makes detailed comparison difficult. This is also the case with the similar taxa C. brevituba Duan in Xing et al., 1984, C. intermedia (Duan, 1984), and C. mira (He in Qian, 1977) from the lower Cambrian of South China (Xiao and Zhou 1984; Duan 1984). These previous identifications as Cupitheca were based mainly on the ornament on the conchs, here we highlight the importance of the operculum in the classification of this group. The occurrence of C. holocyclata without opercula preserved should be questioned (see SOM 2).
The conch of specimen NIGPAS 166334 (Fig. 8A) possesses two distinct zones that are separated by a transverse discontinuity. The adapertural portion is characterised by transverse lines (Fig. 8D), whereas the adapical tapering zone is covered by longitudinal to sub-radial ornament (Fig. 8C). Although the apical termination cannot be well separated from the matrix, based on the distinct ornamentation covering of the two zones, the adapertural region is regarded as early juvenile portion of the conch, whereas the adapical portion is the presumed protoconch. Therefore the transverse discontinuity or furrow is possible the site where the first septum-like structure developed (Bengtson et al. 1990). Furthermore, the diameter of the shell at the furrow is 450 μm, which overlaps with the ranges of diameters of the protoopercula in this collection (Fig. 7C). No weakening points or septum-like structures have been observed on the presumed protoconch. The ridge-like structure (Fig. 8C) seems to be deceptive, appearing as a biological structure, but in fact is most likely the result of mechanical damage because the outmost layer is squeezed to form an uneven ridge (Fig. 8C) rather than a dissolved groove (Fig. 8B). Although the apex is partially broken and peeled away, part of the lateral sub-radial lines could still be observed on the mould (Fig. 8E).
Stratigraphic and geographic range.—The Manto Formation, late Cambrian Series 2 Stage 4 or early Series 3 Stage 5, Hebei Province, North China (see SOM 2).

Fig. 8. Internal mould of the conch of cupithecid hyolith Cupitheca convexa sp. nov. (NIGPAS 166334) from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section of Tangshan, China. A. General view. B. Boundary between presumed protoconch and the rest of the shell. C. Transverse lines preserved on the surface of the conch. D. Conch covered with longitudinal to sub-radial ornament and mechanical compression forming a transverse ridge-like structure. E. Apical portion. F. Lamellar-fibrillar structure (indicated by white arrow in A). Scale bars: A, E, 200 μm; B, 20 μm; C, 2 μm; D, 50 μm; F, 5 μm.
Discussion
Mode of growth, early life habit, and affinity of Cupitheca.—Bengtson in Bengtson et al. (1990) presented evidence for an unusual manner of growth for Cupitheca in which the animal decollated portions of its conch at various times by the development of a septum-like transverse wall in the apical region, which may have been a strategy similar to coiling in order to avoid reaching an extreme length. Cupitheca convexa sp. nov. probably evolved a new life habit with a preference for shallow, nearshore environments where the availability of resources may have periodically fluctuated. The decollating life habit would have been especially well suited to this type of environment, in that by shortening itself, the animal would have required fewer resources in order to survive. The decollating mode of growth is similar to that of some extant terrestrial and littoral-fringe gastropods (Vermeij 1973; Gittenberger and Povel 1995; Páll-Gergely and Németh 2008) as well as gastropods from deep-water hot vents and cold seeps (Kaim et al. 2008). The functions of decollation for these animals include repair of an incidentally damaged apex or protection against desiccation by evaporation (Gittenberger and Povel 1995). Cupitheca convexa sp. nov. having lived in shallow water, may also have developed decollation for repair of injured apex. Regarding this mode of growth, it is questionable as to whether the presumed gastropod ancestor of the extant decollating forms possessed the ability to shed portions of its shell or whether decollation evolved within this group, convergent with Cupitheca.
The protoconch of Cupitheca remained unknown until now. Although orthothecide protoconchs have been frequently documented (Dzik 1980; Duan 1984; Qian and Zhang 1985; Qian 1989; Wrona 2003; Steiner et al. 2007; Topper et al. 2009), most were described for taxonomic purposes. The protoconchs observed on Bactrotheca, Conotheca, Neogloborilus and some other orthothecide taxa either have a smooth sub-spherical apex (Dzik 1978, 1980; Qian and Zhang 1985; Qian 1989; Wrona 2003; Steiner et al. 2007; Topper et al. 2009) or fusiform with a pointed apex (Dzik 1978, 1980; Duan 1984; Qian and Zhang 1985; Martí Mus et al. 2014), separated by a constriction of variable size from the remainder of the conch. In this case the constriction is well-defined on the internal mould and this type of feature ordinarily has been interpreted as having recorded hatching or the beginning of metamorphosis (Martí Mus et al. 2014). However, the presumed protoconch of Cupitheca described herein differs from the protoconchs mentioned above in terms of overall shape (fusiform versus rounded). The size of the C. convexa sp. nov. protoconch, however, surpasses the dimensions of all other members of hyoliths (Bengtson et al. 1990; Martí Mus et al. 2014), and hence must be considered unusual among Cambrian hyoliths, which implies a possible lecithotrophic development as suggested for other metazoan groups (Allen and Pernet 2007; Laibl et al. 2017). Lecithotrophy of this group may decrease reliance on exogenous nutrition associated with unpredictable food resources (Laibl et al. 2017) and thereby maximize chances of survival (Allen and Pernet 2007; Mercier et al. 2013). The protooperculum of C. convexa sp. nov. is flat rather than elevated as in C. holocyclata (Skovsted et al. 2016), indicating that an attachment or anchoring larval lifestyle is unlikely and that the larva was possibly free-lying on the sea floor, with limited locomotion or passively movable by currents.
The presumed protoconch of C. convexa sp. nov. does not show abrupt morphological change, and the larval period terminated with a juvenile body form that gradually acquired adult characteristics, as in previously documented groups (Martí Mus et al. 2014). With the growth of the conch, C. convexa sp. nov. graded into the decollating stage (see SOM 3). Observed diameters of protoopercula are usually smaller than those of the circumferential rims of the discarded segments (Fig. 7C), which suggests that decollation started following metamorphosis. Some discarded fragments that have a circumferential diameter similar to that of protoopercula (Fig. 7C), might be indicative of early decollating stages.
The hyolith affinity of Cupitheca is clearly established (Skovsted et al. 2016), and it has been proposed that Cupitheca is closer to the orthothecide hyoliths as has been suggested previously by Kruse et al. (2004). This suggestion is based mainly on the lack of helens and clavicles, although the presence of a cardinal shield in C. holocyclata is more reminiscent of hyolithides than the orthothecides (Skovsted et al. 2016). As is typical for orthothecides, the operculum of C. convexa sp. nov. possesses cardinal processes only and is planar without differentiation of cardinal or conical shields. One characteristic trait of orthothecides not yet seen in Cupitheca is an operculum that may be retracted a short distance into the conch (Marek 1963, 1966, 1967; Malinky 2009; Valent and Fatka 2013), which is thought to have been a strategy for minimizing sediment intake into the shell, given the absence of a ligula in that group. Although no specimens of Cupitheca possess an attached operculum, the width range of opercula matches that of the apertures (Fig. 7D), which in some degree may provide a proxy of the supposed retractable manner of C. convexa sp. nov. Consequently, an assignment of Cupitheca to Orthothecida is suggested.
Skeletal microstructures of Cupitheca and Decoritheca.—Most of the specimens studied herein were entirely decalcified and the original skeletal microstructures have been largely destroyed by diagenesis or erosion. However, some shells fortunately preserve replicas of the original microstructures.
One specimen (NIGPAS 166339a) of C. convexa sp. nov. has well preserved pits and longitudinal gutters connecting them on the outer surface of the cylindrical rim (Fig. 5C1, C2). The pits correspond to the radial tubules in the rim of the convex wall, which may have functioned as channels for releasing shell-dissolving fluid to the specific area of transverse wall formation (Bengtson et al. 1990). The gutters connected to the pits may be the result of erosion by groundwater and therefore are a taphonomic rather than biologic feature. Lamellar-fibrillar microstructure commonly seen in Cambrian molluscan forms (Feng and Sun 2003; Vendrasco et al. 2010) has been observed on one internal mould of C. convexa sp. nov. (Fig. 8F) with crystal bundles oriented at varying angles. Whether this structural resemblance is the result of homology or convergence remains an open question, as the lamellar-fibrillar structure has evolved convergently several times even within the molluscs (Vendrasco et al. 2010). Some opercula of C. convexa sp. nov. exhibit scattered sub-circular, shallow and nearly flat-bottomed pits (preserved as nodules on the counterparts; e.g., Fig. 6D) on the marginal area of the internal surface. The pits documented here may correspond to the terminations of rods in the opercula (Kouchinsky 2000b) whereas similar structures on the exterior of some brachiopod shells are interpreted as impressions of biomineralised tablets in the primary layer (Cusack et al. 1999).
The skeletal microstructure of Decoritheca has been reported only once before (Dzik 1980) as consisting of phosphatized rods in the shell body wall. However, the conch of D. excavata illustrated by Dzik (1980) possesses a sharply triangular cross section unlike the reniform cross section of typical Decoritheca, therefore the identification as Decoritheca by Dzik (1980) is doubtful. Despite generally good preservation of most specimens of D. cyrene the shell microstructure has been unknown until now. One internal mould from the Manto Formation exhibits rows of tubercles on the dorsal surface of D. cyrene (Fig. 3C). The tubercles are circular to sub-circular with a nearly flat top, which correspond to dimples on the internal surface of the conch. These have been interpreted as casts of tubules in the shell wall that provided a framework to reinforce the skeleton (Kouchinsky 2000b), and they are similar to those of some molluscs (Kouchinsky 2000a). Tubercles are also common structures in trilobites, and some that are smooth and circular may even be comparable to sensory organs of extant arthropods (Størmer 1980). Similar structures such as sensory protrusions (papillae) can also be observed beneath the cuticle of sipunculans (Cutler 1994). To function as sensors, the tubercles would need the reach the external surface of the shell, which is not the case for D. cyrene and the sensory function seems unlikely.
Conclusions
Newly discovered hyoliths from the Manto Formation (Cambrian Stage 4/5) in the Zuojiawu section in Hebei Province, North China are assigned to the species Cupitheca convexa sp. nov. and Decoritheca cyrene (Walcott, 1905). Cardinal processes on opercula of C. convexa sp. nov. support a hyolith affinity for this group (Skovsted et al. 2016). The absence of helens and the planar outline of the operculum without clavicles would argue for its placement within the order Orthothecida. C. convexa sp. nov. and C. holocyclata exhibit strong morphological differences in their opercula, whereas their conchs are far less dissimilar, highlighting the crucial role of the operculum in taxonomy. Discovery of the presumed protoconch of C. convexa sp. nov. demonstrates that this feature is distinct from those of other hyoliths which suggests a strong adaptability and a diversity of early life habit of Cambrian orthothecides. Replicates of the original skeletal microstructures (pits and lamellar-fibrillar structure) of C. convexa sp. nov. bear similarities with those of brachiopods and molluscs, however, whether this is the result of convergence or homology remains an open question. The function of decollation of the conch in C. convexa sp. nov. may have been to repair injuries to the animal, or as an alternative of coiling to avoid an extreme length to deal with the fluctuation of nutrient resources. Specimens of the orthothecide D. cyrene from the Manto Formation possess tubercle structures on the dorsum and these may represent casts of tubules in the shell wall, which possibly have provided a framework to reinforce the skeleton. Our results affirm the diversity of skeletal structure and ecology of Cambrian orthothecides.
Acknowledgements
We acknowledge Martin Valent (National Museum, Prague, Czech Republic) for help with photography and helpful suggestions, and Junming Zhang and Shanchi Peng (both NIGPAS) for beneficial discussions. Jian Gao (Tangshan, China) and Chenyang Cai (NIGPAS) are thanked for help with fieldwork. Peter D. Kruse (South Australian Museum, Normanville, Australia) and an anonymous reviewer provided constructive and helpful comments that greatly improved the manuscript. This research was supported partly by grants from the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB18000000) and the National Sciences Foundation of China (Nos. 41602002, 41222013), Key Laboratory of Economic Stratigraphy and Palaeogeography, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (No. 2016KF08).
References
Allen, J.D. and Pernet, B. 2007. Intermediate modes of larval development: bridging the gap between planktotrophy and lecithotrophy. Evolution & Development 9: 643–653. Crossref
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., and Runnegar, B.N. 1990. Early Cambrian fossils from South Australia. Memoirs of the Association of Australasian Palaeontologists 9: 1–364.
Berg-Madsen, V. and Malinky, J.M. 1999. A revision of Holm’s late Mid and Late Cambrian hyoliths of Sweden. Palaeontology 42: 841–885. Crossref
Brett, C.E. and Baird, G.C. 1986. Comparative taphonomy: a key to paleoenvironmental interpretation based on fossil preservation. Palaios 1: 207–227. Crossref
Cherns, L., Wheeley, J.R., and Wright, V.P. 2011. Taphonomic bias in shelly faunas through time: early aragonitic dissolution and its implications for the fossil record. In: P.A. Allison and D.J. Bottjer (eds.), Taphonomy: Process and Bias through Time, 79–105. Springer, New York.
Cusack, M., Williams, A., and Buckman, J.O. 1999. Chemico—structural evolution of linguloid brachiopod shells. Palaeontology 42: 799–840. Crossref
Cutler, E.B. 1994. The Sipuncula: Their Systematics, Biology, and Evolution. 453 pp. Cornell University Press, New York.
Demidenko, Y.E. 2001. Small shelly fossils. In: E.M. Alexander, J.B. Jago, A.Y. Rozanov, and A.Y. Zhuravlev (eds.), The Cambrian Biostratigraphy of the Stansbury Basin, South Australia, 85–117. Transactions of the Palaeontological Institute, Russian Academy of Sciences. IAPC Nauka/Interperiodica, Moscow.
Devaere, L., Clausen, S., Alvaro, J.J., Peel, J.S., and Vachard, D. 2014. Terreneuvian orthothecid (Hyolitha) digestive tracts from northern Montagne Noire, France; taphonomic, ontogenetic and phylogenetic implications. PLoS One 9 (2): e88583. Crossref
Duan, C.H. 1984. Small shelly fossils from the lower Cambrian Xihaoping Formation in the Shennongjia district, Hubei Province—Hyoliths and fossil skeletons of unknown affinities [in Chinese, with English abstract]. Bulletin, Tianjin Institute of Geology and Mineral Resources 7: 143–183.
Duan, J.Y. 1966. Cambrian stratigraphy and trilobites from northeastern Hebei and western Liaoning [in Chinese]. Symposium of Changchun Geological Institute 4: 137–152.
Duan, J.Y., An, S.L., Liu, P.J., Peng, X.D., and Zhang, L.Q. 2005. The Cambrian Stratigraphy, Fauna and Palaeogeography in Eastern Part of North China Plate [in Chinese, with English abstract]. 255 pp. Yayuan Publishing House, Hong Kong.
Dzik, J. 1978. Larval development of hyolithids. Lethaia 11: 293–299. Crossref
Dzik, J. 1980. Ontogeny of Bactrotheca and related hyoliths. Geologiska Föreningen i Stockholm Förhandlingar 102: 223–233. Crossref
Endo, R. 1937. Addenda to Parts I and II. In: R. Endo and C.E. Resser (eds.), The Sinian and Cambrian Formations and Fossils of Southern Manchoukuo. Manchurian Science Museum Bulletin 1: 302–366.
Feng, W. and Sun, W. 2003. Phosphate replicated and replaced microstructure of molluscan shells from the earliest Cambrian of China. Acta Palaeontologica Polonica 48: 21–30.
Galle, A. and Parsley, R.L. 2005. Epibiont relationships on hyolithids demonstrated by Ordovician trepostomes (Bryozoa) and Devonian tabulates (Anthozoa). Bulletin of Geosciences 80: 125–138.
Gilbert, I.R., Hughes, N.C., and Myrow, P.M. 2016. Cambrian microfossils from the Tethyan Himalaya. Journal of Paleontology 90: 10–30. Crossref
Gittenberger, E. and Povel, G.D.E. 1995. Shell growth and decollation in terrestrial gastropods. The Nautilus 109: 38–40.
Holm, G.J. 1893. Sveriges Kambrisk–Siluriska Hyolithidæ och Conulariidæ. Sveriges Geologiska Undersökning, Series C 112: 1–172.
Huang, D.Y., Wang, Y.Y., Gao, J., and Wang, Y.N. 2012. A new anomalocaridid frontal appendage from the Middle Cambrian Mantou Formation of the Tangshan area, Hebei [in Chinese, with English abstract]. Acta Palaeontologica Sinica 51: 411–415.
Hughes, N.C. 2016. The Cambrian palaeontological record of the Indian subcontinent. Earth-Science Reviews 159: 428–461. Crossref
Jensen, S., Palacios, T., and Martí Mus, M. 2010. Revised biochronology of the Lower Cambrian of the Central Iberian zone, southern Iberian massif, Spain. Geological Magazine 147: 690–703. Crossref
Jiang, Z.W. 1992. The Lower Cambrian fossil record of China. In: J.H. Lipps and P.W. Signor (eds.), Origin and Early Evolution of the Metazoa, 311–333. Plenum Press, New York.
Kaim, A., Jenkins, R.G., and Warén, A. 2008. Provannid and provannid-like gastropods from the Late Cretaceous cold seeps of Hokkaido (Japan) and the fossil record of the Provannidae (Gastropoda: Abyssochrysoidea). Zoological Journal of the Linnean Society 154: 421–436. Crossref
Kouchinsky, A.V. 2000a. Shell microstructures in Early Cambrian molluscs. Acta Palaeontologica Polonica 45: 119–150.
Kouchinsky, A.V. 2000b. Skeletal microstructures of hyoliths from the Early Cambrian of Siberia. Alcheringa 24: 65–81. Crossref
Kruse, P.D., Laurie, J.R., and Webby, B.D. 2004. Cambrian geology and palaeontology of the Ord Basin. Memoirs of the Association of Australasian Palaeontologists 30: 1–58.
Laibl, L., Esteve, J., and Fatka, O. 2017. Giant postembryonic stages of Hydrocephalus and Eccaparadoxides and the origin of lecithotrophy in Cambrian trilobites. Palaeogeography, Palaeoclimatology, Palaeoecology 470: 109–115. Crossref
Lee, B.S. 2008. Stratigraphic implications of skeletal microfossils from the Cambrian of Korea. Acta Geologica Sinica, English Edition 82: 1152–1159.
Lu, Y.H. 1962. Collection of Scientific Papers Submitted to the National Congress on Stratigraphy, Cambrian System of China [in Chinese]. 117 pp. Science Press, Beijing.
Lu, Y.H. and Dong, N.T. 1953. New survey of type sections of Cambrian from Shandong [in Chinese]. Acta Geologica Sinica 32: 164–201.
Malinky, J.M. 1987. Taxonomic revision of lower and middle Paleozoic Orthothecida (Hyolitha) from North America and China. Journal of Paleontology 61: 942–959. Crossref
Malinky, J.M. 1990. Cambrian Hyolitha from northeast Canada: Reappraisal of the hyolith orders Camerothecida and Diplothecida. Journal of Paleontology 64: 587–595. Crossref
Malinky, J.M. 2009. First occurrence of Orthotheca Novák, 1886 (Hyolitha, Early Devonian) in North America. Journal of Paleontology 83: 588–596. Crossref
Malinky, J.M. 2014. Cambrian Hyolitha and problematica from West Laurentian North America: taxonomy and palaeobiogeography. Alcheringa 38: 338–363. Crossref
Malinky, J.M. and Skovsted, C.B. 2004. Hyoliths and small shelly fossils from the Lower Cambrian of North-East Greenland. Acta Palaeontologica Polonica 49: 551–578.
Marek, L. 1963. New knowledge on the morphology of Hyolithes. Sborník geologickych věd, řada Paleontologie 1: 53–72.
Marek, L. 1966. New hyolithid genera from the Ordovician of Bohemia. Časopis národního muzea 135: 89–92.
Marek, L. 1967. The class Hyolitha in the Caradoc of Bohemia. Sborník geologickych věd, řada Paleontologie 9: 51–113.
Marek, L., Parsley, R.L., and Galle, A. 1997. Functional morphology of hyolithids based on flume studies. Věstník Českého geologického ústavu 72: 351–358.
Martí Mus, M., Jeppsson, L., and Malinky, J.M. 2014. A complete reconstruction of the hyolithid skeleton. Journal of Paleontology 88: 160–170. Crossref
Mercier, A., Doncaster, E.J., and Hamel, J.-F. 2013. Contrasting predation rates on planktotrophic and lecithotrophic propagules by marine benthic invertebrates. Journal of Experimental Marine Biology and Ecology 449: 100–110. Crossref
Moysiuk, J., Smith, M.R., and Caron, J.-B. 2017. Hyoliths are Palaeozoic lophophorates. Nature 541: 394–397. Crossref
O’Brien, N.R., Nakazawa, K., and Tokuhashi, S. 1980. Use of clay fabric to distinguish turbiditic and hemipelagic siltstones and silts. Deep-Water Turbidite Systems: 271–271.
Páll-Gergely, B. and Németh, L. 2008. Observations on the breeding habits, shell development, decollation, and reproductive anatomy of Pontophaedusa funiculum (Mousson 1856) (Gastropoda, Pulmonata, Clausiliidae, Phaedusinae). Malacologica Bohemoslovaca 7: 11–14.
Pan, B., Miao, L.Y., Yang, H.N., and Li, G.X. 2015. Enigmatic tubular fossil Cupitheca from the Lower Cambrian Xinji Formation of Luonan, Shaanxi Province [in Chinese, with English abstract]. Acta Micropalaeontologica Sinica 32: 384–395.
Parkhaev, P.Y. and Demidenko, Y.E. 2010. Zooproblematica and Mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China. Paleontological Journal 44: 883–1161. Crossref
Peng, S.C. 2009. Review on the studies of Cambrian trilobite faunas from Jiangnan Slope Belt, South China, with notes on Cambrian correlation between South and North China. Acta Palaeontologica Sinica 48: 437–452.
Peng, S.C., Babcock, L.E., and Cooper, R.A. 2012. The Cambrian Period. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), The Geologic Time Scale 2012, 437–488. Elsevier BV, Amsterdam. Crossref
Pojeta, J. 1987. Phylum Hyolitha. In: R.S. Boardman, A.H. Cheetham, and A.J. Rowell (eds.), Fossil Invertebrates, 416–444. Blackwell, Palo Alto.
Qian, Y. 1977. Hyolitha and some problematica from the Lower Cambrian Meishucun Stage in central and S.W. China [in Chinese, with English abstract]. Acta Palaeontologica Sinica 16: 255–275.
Qian, Y. 1978. The Early Cambrian hyolithids in central and southwest China and their stratigraphical significance [in Chinese, with English abstract]. Memoirs of the Nanjing Institute of Geology and Palaeontology, Academia Sinica 11: 1–38.
Qian, Y. 1983. Hyolitha [in Chinese]. In: Nanjing Institute of Geology and Mineral Resources (ed.), Paleontological Atlas of East China; Volume of the Early Paleozoic, 286–289. Geological Publishing House, Beijing.
Qian, Y. 1989. Early Cambrian Small Shelly Fossils of China with Special Reference to the Precambrian–Cambrian Boundary (2). 341 pp. Nanjing University Publishing House, Nanjing.
Qian, Y. and Xiao, L.G. 1995. Hyolitha [in Chinese]. 262 pp. Science Press, Beijing.
Qian, Y. and Zhang, S. 1985. On the systematic position of small orthoconic fossils with bulbous initial part from the early Lower Cambrian [in Chinese, with English abstract]. Acta Micropalaeontologica Sinica 2: 1–13.
Qian, Y., Chen, M.E., and Qian, Y.Y. 1979. Hyolithids and other small shelly fossils from the Lower Cambrian Huangshandong Formation in the eastern part of the Yangtze Gorge [in Chinese, with English abstract]. Acta Palaeontologica Sinica 18: 207–230.
Qian, Y., Li, G.X., Zhu, M.Y., and Yin, G.Z. 2003. Hyoliths from the Lower Cambrian Xiaoerbulakr Formation of Wushi, Xinjiang [in Chinese, with English abstract]. Acta Micropalaeontologica Sinica 20: 342–349.
Resser, C.E. and Endo, R. 1937. Description of the fossils. In: R. Endo and C.E. Resser (eds.), The Sinian and Cambrian Formations and Fossils of Southern Manchoukuo. Manchurian Science Museum Bulletin 1: 103–301
Runnegar, B., Pojeta, J., Morris, N.J., Taylor, J.D., Taylor, M.E., and McClung, G. 1975. Biology of the Hyolitha. Lethaia 8: 181–191. Crossref
Simões, M.G. and Aceñolaza, G.F. 2009. Hyolith-dominated shell beds from the Lampazar Formation in NW Argentina: patterns and processes of origin in the Late Cambrian (Furongian) seas of western Gondwana. Memoirs of the Association of Australasian Palaeontologists 37: 453–462.
Sinclair, G.W. 1946. Notes on the nomenclature of Hyolithes. Journal of Paleontology 20: 72–85.
Skovsted, C. and Peel, J.S. 2007. Small shelly fossils from the argillaceous facies of the Lower Cambrian Forteau Formation of western Newfoundland. Acta Palaeontologica Polonica 52: 729–748.
Skovsted, C.B., Pan, B., Topper, T.P., Betts, M.J., Li, G., and Brock, G.A. 2016. The operculum and mode of life of the lower Cambrian hyolith Cupitheca from South Australia and North China. Palaeogeography, Palaeoclimatology, Palaeoecology 443: 123–130. Crossref
Størmer, L. 1980. Sculpture and microstructure of the exoskeleton in chasmopinid and phacopid trilobites. Palaeontology 23: 237–271.
Steiner, M., Li, G., Qian, Y., Zhu, M., and Erdtmann, B.-D. 2007. Neoproterozoic to Early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China). Palaeogeography, Palaeoclimatology, Palaeoecology 254: 67–99. Crossref
Sun, H., Babcock, L.E., Peng, J., and Zhao, Y. 2016. Three-dimensionally preserved digestive systems of two Cambrian hyolithides (Hyolitha). Bulletin of Geosciences 91: 51–56. Crossref
Sysoyev, V.A. [Sysoev, V.A.] 1958. The superorder Hyolithoidea [in Russian]. In: N.P. Luppov and V.V. Druŝic (eds.), Osnovy paleontologii, mollyuski i golovonogie, 184–190. Akademiâ Nauk SSSR, Moskva.
Sysoyev, V.A. [Sysoev, V.A.] 1968. Stratigrafiâ i hiolity drevnejših sloev nižnego kembriâ Sibirskoj Platformy. 67 pp. Âkutskij Gosudarstvennyj Universitet and Institut Geologii, Âkutskij Filial Sibirskogo Otdeleniâ, Akademiâ Nauk SSSR, Âkutsk.
Sysoyev, V.A. [Sysoev, V.A.] 1972. Biostratigrafiâ i hiolity ortotecimorfy nižnego Kembriâ Sibirskoj platformy. 152 pp. Nauka, Moskva.
Topper, T.P., Brock, G.A., Skovsted, C.B., and Paterson, J.R. 2009. Shelly fossils from the Lower Cambrian Pararaia bunyerooensis Zone, Flinders Ranges, South Australia. Memoirs of the Association of Australasian Palaeontologists 37: 199.
Turner, P. 1979. Diagenetic origin of Cambrian marine red beds: Caerfai Bay shales, Dyfed, Wales. Sedimentary Geology 24: 269–281. Crossref
Valent, M. and Fatka, O. 2013. Gracilitheca astronauta n. sp. and Nephrotheca sophia n. sp. (Hyolitha, Orthothecida) from the Cambrian of Czech Republic. Annales de Paléontologie 99: 207–216. Crossref
Vendrasco, M.J., Porter, S.M., Kouchinsky, A., Li, G., and Fernandez, C.Z. 2010. New data on molluscs and their shell microstructures from the Middle Cambrian Gowers Formation, Australia. Palaeontology 53: 97–135. Crossref
Vermeij, G.J. 1973. Molluscs in mangrove swamps: physiognomy, diversity, and regional differences. Systematic Zoology 22: 609–624. Crossref
Walcott, C.D. 1905. Cambrian faunas of China. Proceedings of the U.S. National Museum 29: 1–106. Crossref
Walcott, C.D. 1911. Cambrian faunas of China. In: C.D. Walcott (ed.), Cambrian Geology and Paleontology II. Smithsonian Miscellaneous Collections 57: 69–108.
Walcott, C.D. 1913. The Cambrian faunas of China. In: C.D. Walcott (ed.), Research in China, 1–276. Carnegie Institute of Washington, Washington D.C.
Wang, Z.J., Huang, Z.G., Yao, J.X., and Ma, X.L. 2014. Characteristics and main progress of The Stratigraphic Chart of China and directions [in Chinese, with English abstract]. Acta Geoscientica Sinica 35: 271–276.
Wrona, R. 2003. Early Cambrian molluscs from glacial erratics of King George Island, West Antarctica. Polish Polar Research 24: 181–216.
Xiang, L.W., Zhu, Z.L., Li, S.J., and Zhou, Z.Q. 1999. Chinese Stratigraphic Lexicon (The Cambrian) [in Chinese]. 95 pp. Geological Publishing House, Beijing.
Xiao, B. and Duan, C.H. 1992. Review of small shelly fauna of Yultus, Early Cambrian of Xinjiang [in Chinese, with English abstract]. Xinjiang Geology 10: 212–232.
Xiao, L.G. and Zhou, B.H. 1984. Early Cambrian Hyolitha from Huainan and Huoqiu County in Anhui Province [in Chinese, with English abstract]. Professional Papers of Stratigraphy and Palaeontology 13: 141–151.
Xing, Y.S., Ding, Q.X., Luo, H.L., He, T.G., and Wang, Y.G. 1984. The Sinian–Cambrian boundary of China [in Chinese]. Bulletin of the Institute of Geology, Chinese Academy of Geological Sciences 10: 1–262.
Zhao, F.C., Caron, J.-B., Hu, S.X., and Zhu, M.Y. 2009. Quantitative analysis of taphofacies and paleocommunities in the early Cambrian Chengjiang Lagerstätte. Palaios 24: 826–839. Crossref
Zhao, F.C., Caron, J.-B., Bottjer, D.J., Hu, S.X., Yin, Z.J., and Zhu, M.Y. 2014. Diversity and species abundance patterns of the early Cambrian (Series 2, Stage 3) Chengjiang Biota from China. Paleobiology 40: 50–69. Crossref
Ziegler, A. and McKerrow, W. 1975. Silurian marine red beds. American Journal of Science 275: 31–56. Crossref
Acta Palaeontol. Pol. 63 (1): 87–101, 2018
https://doi.org/10.4202/app.00413.2017