
Sauropod dinosaur remains from a new Early Jurassic locality in the Central High Atlas of Morocco
CECILY S.C. NICHOLL, PHILIP D. MANNION, and PAUL M. BARRETT
Nicholl, C.S.C., Mannion, P.D., and Barrett, P.M. 2018. Sauropod dinosaur remains from a new Early Jurassic locality in the Central High Atlas of Morocco. Acta Palaeontologica Polonica 63 (1): 147–157.
Despite being globally widespread and abundant throughout much of the Mesozoic, the early record of sauropod dinosaur evolution is extremely poor. As such, any new remains can provide significant additions to our understanding of this important radiation. Here, we describe two sauropod middle cervical vertebrae from a new Early Jurassic locality in the Haute Moulouya Basin, Central High Atlas of Morocco. The possession of opisthocoelous centra, a well-developed system of centrodiapophyseal laminae, and the higher elevation of the postzygapophyses relative to the prezygapophyses, all provide strong support for a placement within Sauropoda. Absence of pneumaticity indicates non-neosauropod affinities, and several other features, including a tubercle on the dorsal margin of the prezygapophyses and an anteriorly slanting neural spine, suggest close relationships with various basal eusauropods, such as the Middle Jurassic taxa Jobaria tiguidensis and Patagosaurus fariasi. Phylogenetic analyses also support a position close to the base of Eusauropoda. The vertebrae differ from the only other Early Jurassic African sauropod dinosaurs preserving overlapping remains (the Moroccan Tazoudasaurus naimi and South African Pulanesaura eocollum), as well as stratigraphically younger taxa, although we refrain from erecting a new taxon due to the limited nature of the material. These new specimens represent one of the earliest eusauropod taxa and are an important additional data point for elucidating the early evolution of the clade.
Key words: Dinosauria, Sauropoda, Mesozoic, Gondwana, Morocco, Haute Moulouya Basin.
Cecily S.C. Nicholl [cecily.nicholl14@imperial.ac.uk] and Philip D. Mannion [philipdmannion@gmail.com], Department of Earth Science and Engineering, Imperial College London, South Kensington Campus, London, SW7 2AZ, UK.
Paul M. Barrett [p.barrett@nhm.ac.uk], Department of Earth Sciences, The Natural History Museum, Cromwell Road, London, SW7 5BD, UK.
Received 18 September 2017, accepted 30 October 2017, available online 29 January 2018.
Copyright © 2018 C.S.C. Nicholl et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Sauropods were a diverse clade of gigantic herbivorous dinosaurs that dominated many Mesozoic terrestrial ecosystems (Wilson 2002; Upchurch et al. 2004). However, our understanding of the early evolution and diversification of Sauropoda is hindered by a poor fossil record prior to the Middle Jurassic (Upchurch et al. 2004; Allain and Aquesbi 2008; McPhee et al. 2015). Revised phylogenies and new stratigraphic information mean that there are currently no unambiguous occurrences of sauropods in the Late Triassic (McPhee et al. 2014, 2015, 2017). However, definite sauropods are known from the Early Jurassic of China (e.g., McPhee et al. 2016), Europe (e.g., Wild 1978; Stumpf et al. 2015), India (e.g., Kutty et al. 2007; Bandyopadhyay et al. 2010), South America (e.g., Rauhut 2003; Cúneo et al. 2013) and Africa (e.g., Cooper 1984; Allain et al. 2004), although many of these occurrences are fragmentary, and their stratigraphic ages are often poorly constrained (McPhee et al. 2015). Consequently, any new sauropod material from the Early Jurassic provides an important data point that can augment our knowledge of early sauropod evolutionary history.
Although patchily distributed spatiotemporally, sauropod remains have been discovered throughout the Jurassic–Cretaceous of Africa (Mannion 2009; Mannion and Barrett 2013). Early Jurassic remains comprise Vulcanodon karibaensis from Zimbabwe (Raath 1972; Cooper 1984; Viglietti et al. 2018), Pulanesaura eocollum from South Africa (McPhee et al. 2015: although note that this taxon is not a sauropod under some definitions of the clade), and Tazoudasaurus naimi from the Central High Atlas of Morocco (Allain et al. 2004; Allain and Aquesbi 2008). Additional indeterminate remains and trackways have also been reported from Morocco (Termier 1942; Jenny et al. 1980; Hadri et al. 2007).
Here, we describe two sauropod cervical vertebrae from a new Early Jurassic locality in the Central High Atlas of Morocco, northwestern Africa. We place these new remains in context via comparisons with other early sauropods, incorporate them into phylogenetic data matrices, and review the Early–Middle Jurassic fossil record of African sauropods.
Institutional abbreviations.—BP, Evolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa; MNN, Musée National du Niger, Niamey, Republic of Niger; NHMUK, Natural History Museum, London, UK; PVL, Fundacion Miguel Lillo, Universidad Nacional de Tucumán, San Miguel de Tucuman, Argentina.
Other abbreviations.—ACDL, anterior centrodiapophyseal lamina; aEI, average Elongation Index; CPOL, centropostzygapophyseal lamina; CV, cervical vertebra; DP, diapophysis; PCDL, posterior centrodiapophyseal lamina; PCPL, posterior centroparapophyseal lamina; PODL, postzygodiapophyseal lamina; POSL, postspinal lamina; POZ, postzygapophysis; PP, parapophysis; PRDL, prezygodiapophyseal lamina; PRSL, prespinal lamina; PRZ, prezygapophysis; SPOL, spinopostzygapophyseal lamina; SPRL, spinoprezygapophyseal lamina; TPOL, interpostzygapophyseal lamina; TPRL, interprezygapophyseal lamina; V1, V2, vertebra 1, vertebra 2.
Geological and geographical setting
The vertebrae (NHMUK PV R36834) were found in the lower to middle Liassic sediments of the Haute Moulouya Basin, central Morocco (Fig. 1). The site lies near to an escarpment close to the River Moulouya in the Alto Muluya region, between the Middle and the Upper Central Atlas, 3 km southeast of the mineral locality of Aouli, Khénifra Province (32°49’5.03” N, 4°33’21.36” W). Sediments represent syntectonic continental basin infill formed during rifting of the Atlas region (Saâdi et al. 2012).
The specimens were obtained commercially and, although the position of the locality was recorded with precision, some uncertainty remains regarding the source horizon for these vertebrae. According to Bouabdellah and Sangster (2016), the coordinates place the locality within a lower to middle Liassic marine unit comprising conglomerate, sandstone, marl, clay, and massive dolostone and limestone rocks. If these marine units were the source of this material, then this would imply that some transportation of the vertebrae would have occurred; however, because they are well preserved and articulated (Fig. 2), with little evidence of abrasion, we propose that the vertebrae were not likely to have been transported far, and suggest that the marine units are unlikely to be the provenance of NHMUK PV R36834.
Saâdi et al. (2012) described several Lower Jurassic (Hettangian–Sinemurian) units within this region, some of which are terrestrial in origin, consisting predominantly of floodplain deposits cross cut by fluvial channels (termed the “Terrestrial Lateral Series”) (Fig. 1). Formed from three formations (Tizi n’Rechou, Tizi n’Toumelba, and Aghbalou Oumlil, listed from oldest to youngest), these Lower Jurassic terrestrial beds are composed primarily of shale and rusty clay, incised by fluvial conglomerate and sand. The matrix surrounding the vertebrae is formed of homogenous grey silt to mud grade sediment; however, this does not help to elucidate the likely formation of origin for the material, as shale is found within all three horizons, and we cannot be sure that this does not represent a lens within a larger scale rock unit. Consequently, the precise provenance of the vertebrae remains uncertain.
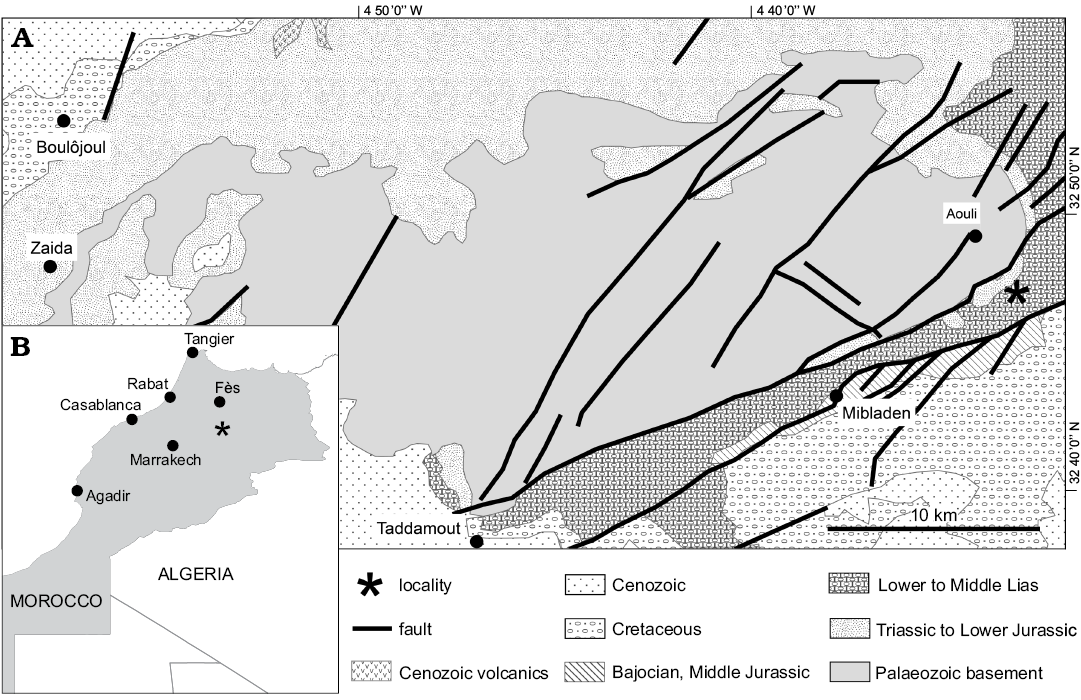
Fig. 1. A. Geological map of the Haute Moulouya Basin, Central High Atlas of Morocco, showing the location of the NHMUK PV R36834 site (based on Saâdi et al. 2012). B. Geographic location of the studied area (asterisk).
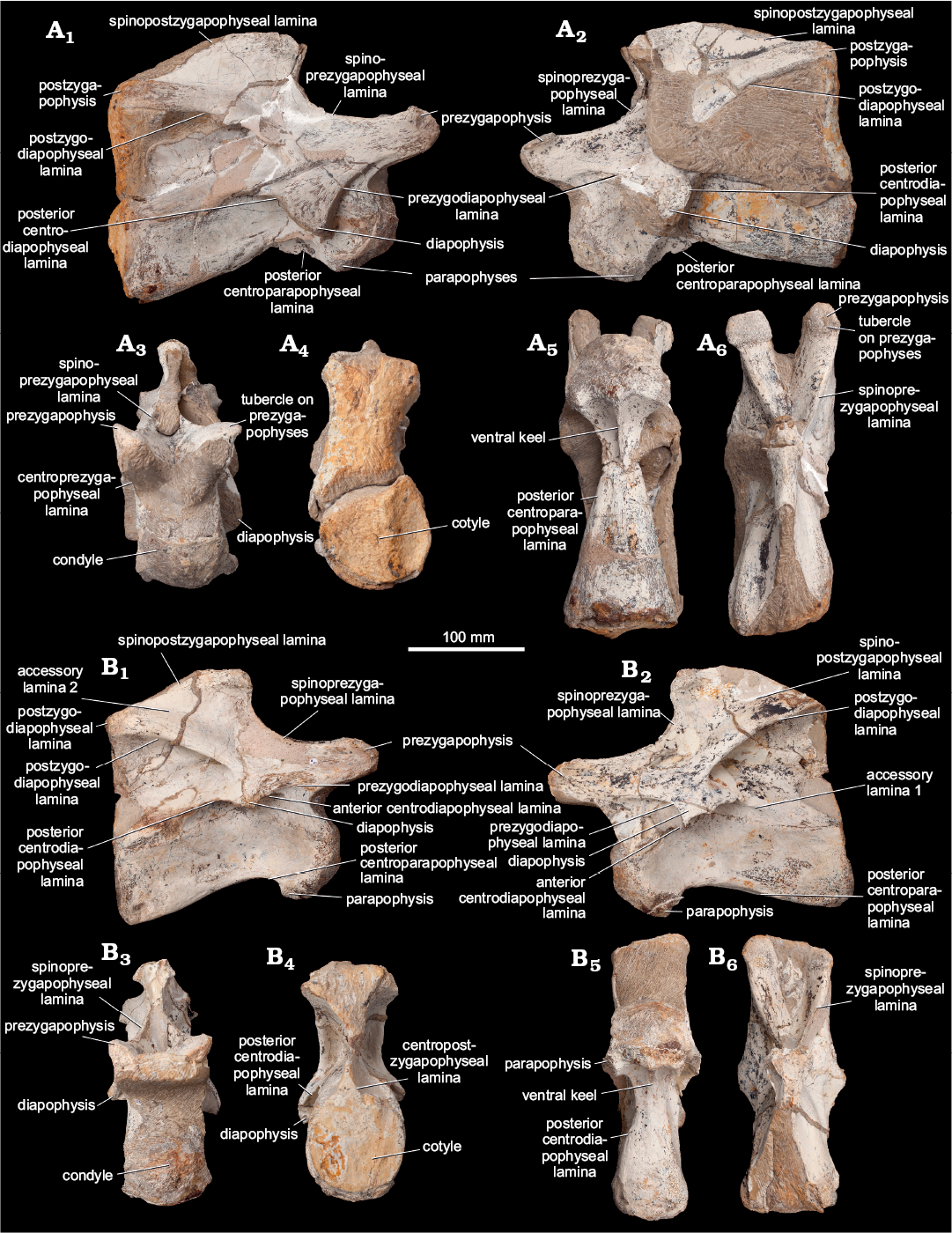
Fig. 2. Photographs of cervical vertebrae of Eusauropoda indet. (NHMUK PV R36834) from the Early Jurassic of the Haute Moulouya Basin, Morocco. Vertebra 1 (A) and vertebra 2 (B); in right lateral (A1, B1), left lateral (A2, B2), anterior (A3, B3), posterior (A4, B4), ventral (A5, B5), and dorsal (A6, B6) views.
Systematic palaeontology
Dinosauria Owen, 1842
Saurischia Seeley, 1887
Sauropodomorpha Huene, 1932
Sauropoda Marsh, 1878
Eusauropoda Upchurch, 1995
Eusauropoda indet.
Material.—NHMUK PV R36834, two complete cervical vertebrae from unknown formation within “lower Lias” unit, Early Jurassic; Haute Moulouya Basin, Central High Atlas, Khénifra Province, central Morocco (coordinates: 32°49’5.03” N, 4°33’21.36” W).
Description.—NHMUK PV R36834 comprises two cervical vertebrae that were found in articulation (Figs. 2, 3). Both are largely complete, consisting of a centrum fused to the neural arch and neural spine, although no ribs are present on either vertebra. Preservation of the vertebrae is generally good, but some areas, especially the anterior and posterior surfaces, are partly obscured by matrix and have been lightly reconstructed. Throughout, the specimens are referred to as vertebra 1 (V1) and vertebra 2 (V2). V2, the more posterior of the two vertebrae, is the most completely prepared. Vertebral laminae are described following the nomenclature of Wilson (1999). Measurements are provided in Table 1.
Table 1. Measurements (in mm) of NHMUK PVR36834.
|
Measurement |
Vertebra 1 |
Vertebra 2 |
|
|
Height |
240 |
241 |
|
|
Length |
313 |
310 |
|
|
Centrum |
height (measured at posterior articular surface) |
88 |
89 |
|
width (measured at posterior articular surface) |
95 |
83 |
|
|
length (excluding condyle) |
211 |
191 |
|
|
length (including condyle) |
247 |
225 |
|
|
Neural arch height |
80 |
57 |
|
|
Neural spine |
height |
72 |
95 |
|
length |
205 |
197 |
|
|
width (widest point) |
89 |
88 |
|
|
width (narrowest point) |
15 |
23 |
|
|
Width across |
diapophyses |
105 |
101 |
|
prezygapophyses |
103 |
89 |
|
|
postzygapophyses |
86 |
88 |
|
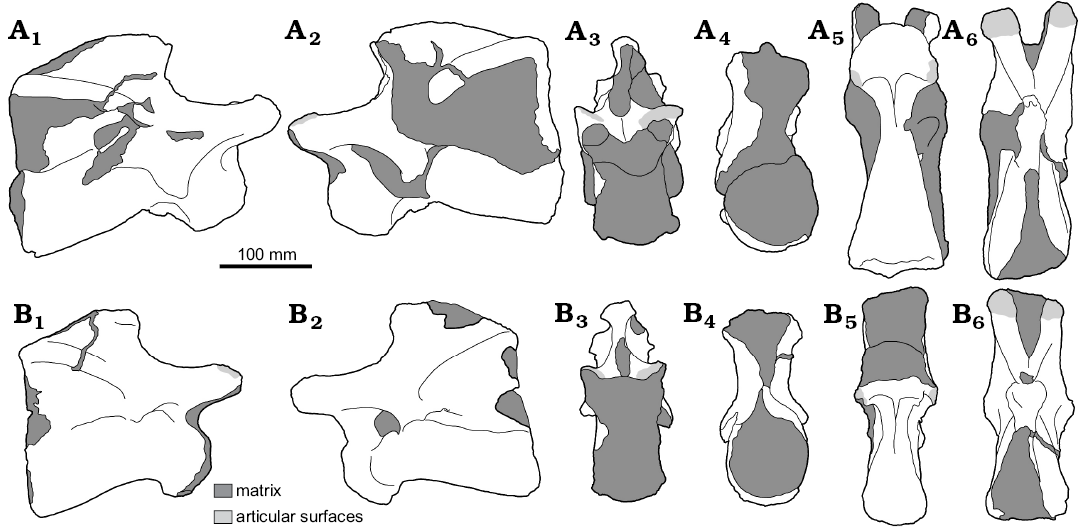
Fig. 3. Line drawings of cervical vertebrae of Eusauropoda indet. (NHMUK PV R36834) from the Early Jurassic of the Haute Moulouya Basin, Morocco. Vertebra 1 (A) and vertebra 2 (B), in right lateral (A1, B1) and left lateral (A2, B2), anterior (A3, B3), posterior (A4, B4), ventral (A5, B5), and dorsal (A6, B6) views.
Based on comparisons with the near-complete cervical vertebral series (MNN TIG F39-49; PDM personal observation 2013) of the non-neosauropod eusauropod Jobaria tiguidensis (Sereno et al. 1999; Fig. 4), we suggest that NHMUK PV R36834 are from the middle region of the neck, probably falling between the fifth and tenth position in the cervical series. This individual was likely to have been a juvenile or sub-adult at the time of death, as indicated by the partial fusion between the centrum and neural arch, and the presence of visible neurocentral sutures, alluding to incomplete growth.
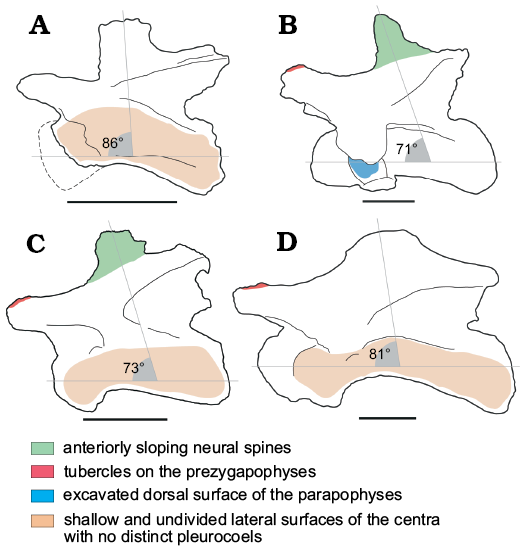
Fig. 4. Morphological comparisons between Lower and Middle Jurassic African sauropod middle cervical vertebrae. A. Pulanesaura (BP/1/6199; McPhee et al. 2015: fig. 4). B. Jobaria tiguendis (MNN TIG F40-49; PDM personal observation 2013). C. Eusauropoda indet. (NHMUK PV R36834). D. Spinophorosaurus nigerensis (Remes et al. 2009: fig. 3A). Scale bars 100 mm.
The centra are opisthocoelous, with an anterior convex condyle that protrudes 25 mm from the main body of the centrum, and a concave posterior cotyle, which has an approximate maximum depth of 24 mm (measurements based on V2). The condyle is orientated anteroventrally, with the apex pointing around 30° from the horizontal, whereas the posterior margin of the centrum is closer to vertical. The cotyle in V1 is shorter dorsoventrally than mediolaterally (ratio of 0.93); however, this ratio is 1.07 in V2. The average Elongation Index (aEI = anteroposterior length of the centra divided by the average of height and width of the posterior articular surface) is 2.26.
In lateral view, the ventral surface is strongly concave, especially anteriorly. Transversely, the anterior one-third of the ventral surface is slightly concave, but is essentially flat along the posterior two-thirds. A midline ventral keel is present along the anterior two-thirds of the centrum; it is prominent in its anterior half, fading out posteriorly. The keel is more strongly developed in V1 than in V2.
Parapophyses project ventrolaterally and slightly posteriorly, although they do not extend far beneath the ventral margin of the centrum. Their dorsal surfaces are unexcavated. A posterior centroparapophyseal lamina (PCPL) runs along the ventrolateral margin of the centrum. The lateral surface of the centrum is gently excavated, but does not form a sharp-lipped opening, and no dividing accessory laminae are present. CT scans reveal that the internal tissue structure of the vertebrae is largely solid (acamerate).
The neural arch (measured up to the base of the postzygapophyses) is dorsoventrally short relative to centrum height (0.91 and 0.64 in V1 and V2, respectively). Only visible in anterior view, the anterior centrodiapophyseal lamina (ACDL) forms a small subvertical ridge between the ventrolateral surface of the diapophysis and the suture line between the centrum and neural arch. A weakly developed ridge (“accessory lamina A”) runs parallel and dorsal to the ACDL. This lamina can only be seen in left lateral view due to damage on the right lateral surface of V2. The presence of accessory lamina A in V1 is questionable as the vertebrae are quite heavily damaged in this section: a ridge is present, but it appears to be along the suture line, and is less ventrally inclined than in V2. A posterior centrodiapophyseal lamina (PCDL) forms a thin ridge that projects ventrolaterally between the posteroventral region of the diapophysis and the lateral surface of the centrum. It is orientated subhorizontally, but dips mildly posteroventrally before fading out in the posterior region of the centrum. Each diapophysis projects ventrolaterally, and lacks any posterior process. The diapophyses shift posteriorly from V1 to V2.
Centroprezygapophyseal laminae (CPRLs) are generally well-preserved as simple, unbifurcated laminae; however, their medial surfaces are largely obscured by matrix. Their anterior margins are strongly concave in lateral view. Although partially obscured by matrix in both vertebrae, the anterior surface of V2 is partly exposed and shows no evidence for a distinct interprezygapophyseal lamina (TPRL). The prezygapophyses are prominent and extend well beyond the anterior-most point of the condyle. Their articular surfaces are flat to very mildly convex, and face slightly dorsomedially. Although no pre-epipophyses are present, a small tubercle is situated on the dorsomedial margin of each prezygapophysis, close to its anterior tip. The prezygodiapophyseal lamina (PRDL) is subhorizontal along its anterior portion, increasing in prominence and becoming ventrolaterally deflected posteriorly.
Centropostzygapophyseal laminae (CPOLs) are preserved in both specimens, although their true shape is obscured by matrix. The dorsolaterally orientated postzygapophyses are generally not well preserved, and are heavily covered in matrix. They are elevated relative to the level of the prezygapophyses and do not extend beyond the posterior margin of the centrum. Their articular surfaces face ventrolaterally. There is no evidence for either an interpostzygapophyseal (TPOL) lamina or for epipophyses, although both absences might be preservational artefacts. The postzygodiapophyseal lamina (PODL) slopes anteroventrally at an angle close to 45° from the horizontal. This lamina is the most prominent of those connecting to the diapophyses, and extends for approximately half of the length of the vertebra. A second accessory lamina (“B”) runs parallel and dorsal to the PODL, at least in V2. In V1, the presence of accessory lamina B is disputable, as it appears to combine with the PODL to form a thicker ridge, and there is no medial depression between the two, perhaps indicating that this is just an extension of the same bony sheet.
The neural spine extends for around two-thirds of the length of the centrum, but is difficult to fully study in both vertebrae because of poor exposure. It is simple, showing no bifurcation, and forms a transversely thin bony sheet that thickens posteriorly, reaching its widest point between the postzygapophyses. In lateral view, the anterior margin of the neural spine is concave, with the apex of the spine deflected slightly anteriorly, whereas the posterior margin slopes to face posterodorsally. The neural spine is essentially formed from the spinoprezygapophyseal (SPRLs) and spinopostzygapophyseal laminae (SPOLs). The SPRLs and SPOLs define the prespinal and postspinal fossae, respectively. No prespinal or postspinal laminae are present. Neural spine height increases from V1 to V2 (arch to spine height ratio of 0.90 and 1.67 in V1 and V2, respectively).
Comparisons with other early sauropods.—Although the vertebrae of NHMUK PV R36834 are well preserved, ontogenetic changes and variation along the cervical series complicate comparisons with other sauropods. We focus our comparisons on Early to Middle Jurassic sauropods that preserve anterior to middle cervical vertebrae, taking into account such possible variation.
The apneumatic internal bone structure of NHMUK PV R36834 is consistent with that seen in the presacral vertebrae of most non-neosauropods (Wilson and Sereno 1998; Wedel 2003; Carballido et al. 2011). However, as NHMUK PV R36834 represents an immature individual, it is important to note that the extent of internal pneumaticity has an ontogenetic, as well as phylogenetic signal (Wedel 2003).
The cervical centra are opisthocoelous, which characterises all sauropods more derived than Gongxianosaurus shibeiensis (Upchurch 1995; Upchurch et al. 2007a; Yates 2007). However, it should be noted that the cervical series of most taxa around the sauropodomorph-sauropod transition are very poorly known, especially with regards to whether or not they are opisthocoelous (e.g., Vulcanodon). In lateral view, the ventral surfaces are arched dorsally. Although this is the case in most sauropodomorphs, the arching in NHMUK PV R36834 is anteriorly restricted, similar to the condition in basal eusauropods such as Jobaria (MNN TIG F41-46; PDM personal observation 2013) and Patagosaurus fariasi (Bonaparte 1986b), whereas the arching extends for most of the centrum length in more basal taxa (e.g., Massospondylus carinatus [BP/1/4934], PMB personal observation 2015; Pulanesaura, McPhee et al. 2015: fig. 4), as well as many neosauropods (e.g., Apatosaurus; Gilmore 1936). The presence of a midline ventral keel in cervical vertebrae is the plesiomorphic sauropodomorph condition that is retained in many basal sauropods (Upchurch 1995), including Amygdalodon patagonicus (Rauhut 2003), Tazoudasaurus (Allain and Aquesbi 2008), Lapparentosaurus madagascariensis (Upchurch 1998), Spinophorosaurus nigerensis (Remes et al. 2009) and Shunosaurus lii (Zhang 1988). A keel is variably present in the cervical centra of Jobaria (MNN TIG F40-49, present in CV3 and in CV9 onwards; PDM personal observation 2013), but is absent in most derived sauropods (Upchurch 1998), including the non-neosauropod eusauropod Cetiosaurus oxoniensis (Upchurch and Martin 2002).
The absence of sharp-lipped excavations on the lateral surfaces of the centra means that NHMUK PV R36834 retains the plesiomorphic sauropodomorph condition (Upchurch 1998; Yates et al. 2012), as is also the case in Shunosaurus (Zhang 1988), Amygdalodon (Rauhut 2003) and Tazoudasaurus (Allain and Aquesbi 2008), contrasting with the development of distinct “pleurocoels” in most eusauropods (Upchurch 1995, 1998). Similarly, the absence of dorsally excavated parapophyses in NHMUK PV R36834 is consistent with the phylogenetic distribution of pleurocoels, although this feature is secondarily lost in many taxa (Upchurch 1998).
The ratio of posterior centrum height to width varies between taxa and along the cervical series (Upchurch 1998; Wilson 2002; Carballido et al. 2012; Mannion et al. 2013). With an average ratio of 1.0, the vertebrae of NHMUK PV R36834 have a similar value to many of the taxa close to the sauropodomorph–sauropod transition (Carballido et al. 2012), including basal sauropods such as Tazoudasaurus (Allain and Aquesbi 2008: fig. 9J) and Jobaria (MNN TIG F41-46; PDM personal observation 2013). An aEI of 2.26 is similar to that of the anterior–middle cervical centra of many basal sauropod taxa (Carballido et al. 2012), including Amgydalodon (Rauhut 2003) and Tazoudasaurus (Allain and Aquesbi 2008), although this is higher than in Jobaria (Sereno et al. 1999) or Patagosaurus (PVL 4170; PDM personal observation 2013), both of which have ratios < 2.0.
Diapophyseal laminae are well developed in NHMUK PV R36834. This is the case in most sauropods, including Tazoudasaurus (Allain and Aquesbi 2008) and Spinophorosaurus (Remes et al. 2009), whereas diapophyseal laminae are rudimentary or absent in most non-sauropodan sauropodomorphs (Wilson 2002; Yates 2007). The CPOLs are similarly prominent in NHMUK PV R36834, contrasting with Tazoudasaurus, in which these laminae are absent (Allain and Aquesbi 2008). Tazoudasaurus also differs from NHMUK PV R36834 in that the former possesses distinct TPRLs (Allain and Aquesbi 2008).
The presence of tubercles on the dorsal surfaces of the anterior ends of the prezygapophyses in NHMUK PV R36834 appears to be uncommon in sauropodomorph cervical vertebrae, and we have only been able to recognize this feature confidently in Jobaria (MNN TIG F39-49; PDM personal observation 2013) and possibly Spinophorosaurus (Remes et al. 2009: fig. 3A; Fig. 4). However, this is a subtle feature that is difficult to detect without firsthand study, and so might be more widespread. The higher elevation of the postzygapophyses relative to the prezygapophyses is a feature that is restricted to sauropods (McPhee et al. 2015), e.g., Patagosaurus (Bonaparte et al. 1986b), Tazoudasaurus (Allain and Aquesbi 2008) and Pulanesaura (if the latter is considered a “true” sauropod; McPhee et al. 2015).
The anterior slanting of the anterior–middle cervical neural spines of NHMUK PV R36834 is shared with Jobaria (MNN TIG F39-49; PDM personal observation 2013; Fig. 4) and Patagosaurus (Bonaparte 1986b), as well as more posterior cervical vertebrae of Cetiosaurus (Upchurch and Martin 2002). A more extreme anterior deflection of the cervical neural spines is also present in most dicraeosaurids (Rauhut et al. 2005). Other sauropods preserving cervical vertebrae have subvertical or posteriorly slanting anterior margins. This appears to be the case in the only near-complete cervical vertebra of Tazoudasaurus, but the anterior margin of its neural spine is not well-preserved (Allain and Aquesbi 2008: fig. 9).
Phylogenetic analysis
A cladistic analysis of the phylogenetic relationships of NHMUK PV R36834 was performed using adapted versions of the data matrices of Carballido et al. (2015) and McPhee et al. (2015). The former includes an extensive sample of taxa from throughout the sauropodomorph tree, whereas the latter is focused on basal sauropodomorph interrelationships. Both updated datasets are provided as TNT files in the SOM (Supplementary Online Material available at http://app.pan.pl/SOM/app63-Nicholl_etal_SOM.pdf). Ordering of multistate characters followed that of Carballido et al. (2015) and McPhee et al. (2015). Both matrices were analysed using the “Stabilize Consensus” option in the “New Technology Search” in TNT vs. 1.1 (Goloboff et al. 2008). The searches were executed using sectorial searches, drift, and tree fusing, and the consensus was stabilized five times, prior to using the resultant trees as the starting trees for a “Traditional Search” using Tree Bisection-Reconstruction.
Coding of 14 out of a possible 370 characters in the Carballido et al. (2015) matrix produced 50 most parsimonious trees (MPTs) with tree lengths of 1079 steps. The strict consensus tree of these MPTs places NHMUK PV R36834 in a polytomy with the non-neosauropods Patagosaurus (Bonaparte 1986b) and Cetiosaurus (Upchurch and Martin 2002), within Eusauropoda, in a position more derived than Shunosaurus and Barapasaurus tagorai (Fig. 5A). Coding of 17 out of 365 characters in the McPhee et al. (2015) matrix resulted in 180 MPTs with tree lengths of 1266 steps. The strict consensus tree of these MPTs places NHMUK PV R36834 in a polytomy with Omeisaurus tianfuensis, Mamenchisaurus youngi, and Barapasaurus, outside of Neosauropoda (Fig. 5B). Again, NHMUK PV R36834 is recovered as a eusauropod more derived than Shunosaurus. Bremer supports for both trees are generally weak, with a value of 1 for the base of Eusauropoda.
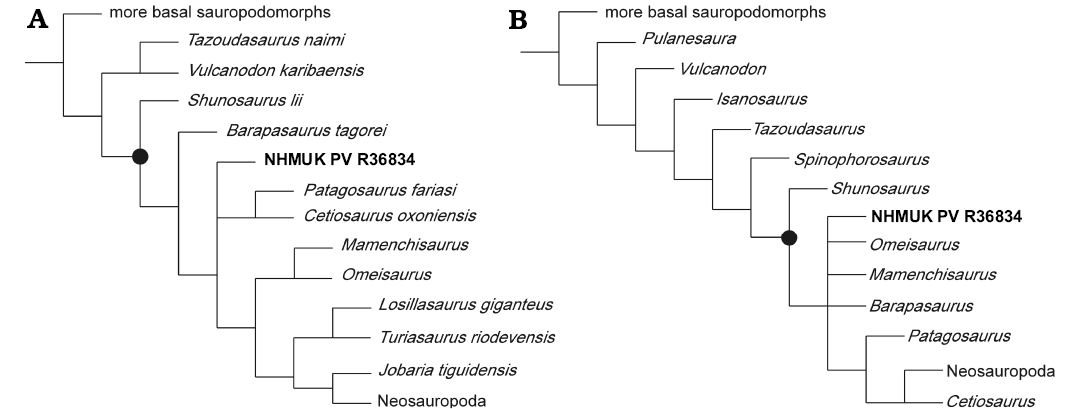
Fig. 5. Strict consensus trees obtained from addition of NHMUK PV R36834 to the data matrices of Carballido et al. 2015 (A) and McPhee et al. 2015 (B). Black circle indicates node for Eusauropoda. Taxa basal to Sauropoda collapsed for simplicity.
Discussion
Phylogenetic affinities.—Our comparisons and phylogenetic analyses provide strong support for referral of NHMUK PV R36834 to Sauropoda. This is based on the possession of opisthocoelous centra (the condition in all sauropods more derived than Gongxianosaurus), the presence of a well-developed centrodiapophyseal laminae system (which is absent or poorly developed in most non-sauropodan sauropodomorphs), and the higher elevation of the postzygapophyses relative to the prezygapophyses (Upchurch 1995; Wilson 2002; Upchurch et al. 2007a; Yates 2007; McPhee et al. 2015). We also propose that the vertebrae likely represent a non-neosauropod, based primarily on the absence of internal or external evidence for pneumaticity, as well as the retention of a midline keel (Upchurch 1995, 1998).
Whether NHMUK PV R36834 belongs within or outside of Eusauropoda is slightly more difficult to ascertain, although both matrices place it within the clade. Several features of NHMUK PV R36834 (the anteriorly-biased arching of the ventral margin in lateral view; the tubercle on the prezygapophyses; and the anteriorly slanting neural spine) are suggestive of placement within Eusauropoda. The main exception is the absence of sharp-lipped excavations on the lateral surfaces of the cervical centra, which might suggest that NHMUK PV R36834 lies outside of Eusauropoda (Upchurch 1998; Carballido et al. 2012). However, the basal eusauropod Shunosaurus also lacks such excavations (Zhang 1988) and thus we tentatively conclude that NHMUK PV R36834 is a non-neosauropod eusauropod. This makes NHMUK PV R36834 the second recognized eusauropod from the Early Jurassic, following an undescribed partial skeleton from the Toarcian of Argentina (Cúneo et al. 2013). Although not explicitly referred to Eusauropoda, McPhee et al. (2016) noted a number of features of the late Early Jurassic Chinese sauropod Sanpasaurus yaoi that might indicate a third Early Jurassic eusauropod.
Finally, although NHMUK PV R36834 lacks autapomorphies, the vertebrae appear to be distinct from those of other contemporaneous sauropod taxa. However, because of the limited material available, and the lack of anatomical overlap with other taxa, we refrain from naming a new taxon based on a unique combination of features.
Review of the Early–Middle Jurassic African sauropod record.—Sauropod remains have been recovered from several Early–Middle Jurassic African localities. Here, we briefly review this fossil record in an attempt to elucidate the early evolution of sauropods on this continent (see Table 2 for a list of named taxa; Fig. 6).
Table 2. Early–Middle Jurassic named sauropod dinosaur taxa from Africa.
|
Taxon |
Country |
Stratigraphic unit |
Age |
References |
|
Pulanesaura eocollum |
South Africa |
Upper Elliot Formation |
Hettangian–Sinemurian |
|
|
Vulcanodon karibaensis |
Zimbabwe |
Forest Sandstone |
Sinemurian–Pliensbachian |
|
|
Tazoudasaurus naimi |
Morocco |
Toundoute Group |
Toarcian |
|
|
Spinophorosaurus nigerensis |
Niger |
Irhazer II Formation |
Bajocian–Bathonian |
|
|
Atlasaurus imelakai |
Morocco |
Guettioua Formation |
Bathonian |
|
|
“Cetiosaurus” mogrebiensis |
Morocco |
El Mers Formation |
Bathonian |
|
|
“Bothriospondylus madagascariensis” |
Madagascar |
Isalo III Formation |
Bathonian |
|
|
Lapparentosaurus madagascariensis |
Madagascar |
Isalo III Formation |
Bathonian |
|
|
Archaeodontosaurus descouensi |
Madagascar |
Isalo III Formation |
Bathonian |
|
|
Jobaria tiguidensis |
Niger |
Tiourarén Formation and Tegama Group |
Bathonian–Callovian |
|
|
Chebsaurus algeriensis |
Algeria |
Aïssa Formation |
Callovian |
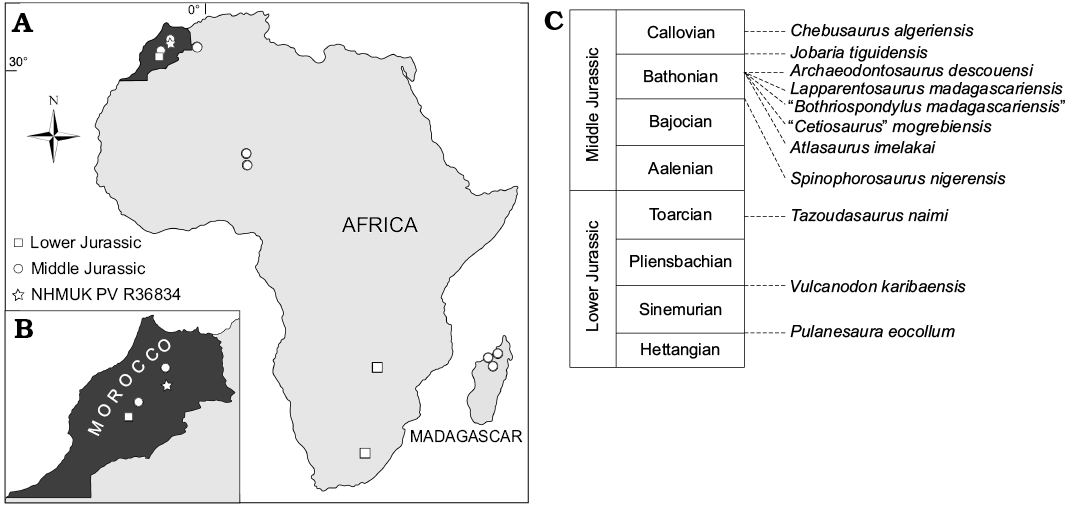
Fig. 6. Map of Africa (A) and enlarged map of Morocco (B), showing Lower and Middle Jurassic sauropod localities. C. Stratigraphic ages of named taxa.
Africa’s stratigraphically oldest known sauropod, Pulanesaura eocollum, comes from the Early Jurassic (Hettangian–Sinemurian) upper Elliot Formation of South Africa, and consists of teeth and partial postcranial remains representing at least two sub-adult to adult individuals (McPhee et al. 2015). It either represents a very primitive sauropod or a close relative that lies just outside the clade, depending upon the preferred definition of Sauropoda adopted (McPhee et al. 2015). Vulcanodon karibaensis is known from the postcranial remains of a single individual (Raath 1972) from the Sinemurian to Pliensbachian-aged Forest Sandstone of Zimbabwe (Viglietti et al. 2018). Originally described as a “prosauropod” (Raath 1972), it was subsequently recognized as a basal sauropod (Cruickshank 1975; Cooper 1984), a placement later supported through phylogenetic analysis (e.g., Upchurch 1995; Wilson and Sereno 1998). A caudal vertebra from the upper Elliot Formation in South Africa was suggested to be closely related to Vulcanodon (Yates et al. 2004), although McPhee et al. (2017) argued that this is more likely referable to either Pulanesaura or a closely related taxon.
Tazoudasaurus naimi is the only named sauropod taxon from the Early Jurassic of North Africa, and was recovered in the same general region as NHMUK PVR36834, i.e., in the Central High Atlas of Morocco (Allain et al. 2004; Allain and Aquesbi 2008). Known from both adult and juvenile partial skeletons, including cranial material, Tazoudasaurus was found in the late Early Jurassic (Toarcian) Toundoute Group, and has been consistently recovered as a non-eusauropod sauropod (e.g., McPhee et al. 2015; Otero et al. 2015), which might form a clade with Vulcanodon (Allain and Aquesbi 2008). Fragmentary sauropod remains and tracks have also been documented from contemporaneous deposits in this region of Morocco (Termier 1942; Jenny et al. 1980; Hadri et al. 2007; Allain and Aquesbi 2008).
The Middle Jurassic African record is slightly richer, and three taxa have been described from northwestern Africa. An almost complete skeleton, including a skull, from the late Middle Jurassic (Bathonian) Guettioua Formation of the Central High Atlas of Morocco, was described by Monbaron et al. (1999) as Atlasaurus imelakai. Although initially considered to lie closer to Brachiosaurus than to other sauropods (Monbaron et al. 1999), Atlasaurus has since been demonstrated to represent a non-neosauropod eusauropod (e.g., Royo-Torres et al. 2006; D’Emic 2012; Mannion et al. 2013). The lateral equivalent of the Guettioua Formation in the Middle Atlas of Morocco, the El Mers Formation (Allain and Aquesbi 2008), has yielded partial postcranial remains of at least three sauropod individuals. A new species of Cetiosaurus, C. mogrebiensis, was erected for this material (Lapparent 1955), although subsequent authors have shown that this material is unlikely to be referable to this genus (e.g., Upchurch and Martin 2003). The phylogenetic analysis of Läng and Mahammed (2010) recovered “Cetiosaurus” mogrebiensis as a non-neosauropod eusauropod. Sauropod tracks have also been described from the El Mers Formation (Jenny et al. 1981). Chebsaurus algeriensis is a non-neosauropod eusauropod from the late Middle Jurassic (Callovian) Aïssa Formation of the Western Saharan Atlas, Algeria, and is represented by two partial juvenile skeletons, including cranial material (Mahammed et al. 2005; Läng and Mahammed 2010).
The Middle Jurassic Irhazer Group in Niger has yielded two sauropod genera. Spinophorosaurus nigerensis is known from two partial skeletons, preserving cranial remains and most of the postcranium (Remes et al. 2009). It was collected from the Irhazer II Formation, which is probably Bajocian–Bathonian in age, and was recovered as a non-eusauropod sauropod (Remes et al. 2009). Jobaria tiguidensis is a non-neosauropod eusauropod known from multiple near-complete skeletons from the overlying Tiourarén Formation (Sereno et al. 1999). Although initially identified as Early Cretaceous in age by Sereno et al. (1999), this stratigraphic unit has since been assigned to the Middle Jurassic, and is most likely Bathonian–Callovian (Rauhut and López-Arbarello 2009).
Multiple sauropod remains have been discovered in the Bathonian-aged Isalo III Formation of northwest Madagascar, which at the time would have been in contact with mainland Africa. Many of these remains have a complicated taxonomic history, although a recent review recognized three distinct sauropod taxa (Mannion 2010). Lapparentosaurus madagascariensis was named by Bonaparte (1986a) for subadult remains of at least five individuals, including fragmentary cranial elements. Although some authors have suggested titanosauriform affinities (e.g., Upchurch 1995), Lapparentosaurus is almost certainly a non-neosauropod eusauropod (Bonaparte 1986a; Läng and Mahammed 2010; Mannion et al. 2013). Archaeodontosaurus descouensi is known only from a dentary and is a non-neosauropod sauropod, possibly belonging to Eusauropoda (Buffetaut 2005; see also Upchurch et al. 2007b). The remaining taxon, “Bothriospondylus madagascariensis”, comprises fragmentary postcranial remains that show differences to Lapparentosaurus, indicating the presence of a third non-neosauropod eusauropod in the Bathonian of Madagascar (Mannion 2010). Other remains have uncertain affinities and might ultimately demonstrate an even higher diversity (e.g., Läng and Goussard 2007). Sauropod tracks are also known from the slightly older (Bajocian–Bathonian) Bemerara Formation in western Madagascar (Wagensommer et al. 2010). Finally, sauropod tracks have been documented from the Middle–Late Jurassic Dande Sandstone Formation of Zimbabwe (Ait-Kaci Ahmed et al. 2004).
Conclusions
The discovery of two well preserved cervical vertebrae from a new Early Jurassic locality in the Haute Moulouya Basin, Central High Atlas of Morocco, demonstrates the presence of a sauropod that appears to differ from contemporaneous taxa. We identify the cervical vertebrae as being from a sauropod based on their opisthocoelous centra, the presence of a well-developed centrodiapophyseal laminae system, and elevated postzygapophyses (relative to the prezygapophyses). The absence of external or internal evidence for pneumaticity indicates non-neosauropod affinities, whereas several other features, coupled with phylogenetic analyses, support a position within Eusauropoda. Despite the limited material, NHMUK PV R36834 provides an important data point as potentially one of the earliest eusauropods known, and improves the record of Early Jurassic sauropods in Africa.
Acknowledgements
We are grateful to all those who have helped with access to sauropod material in their care, especially Tyler Keillor, Bob Masek, and Paul Sereno (all University of Chicago, USA) for facilitating P.D.M.’s examination of Jobaria. Material was prepared by Mark Graham (NHMUK, Conservation Centre), CT-scanned by Amin Garbout (NHMUK, Imaging and Analysis Centre), and photographed by Harry Taylor (NHMUK, Image Resources). We thank Kendal Martyn and Brian Eberhardie (Moussa, UK) for providing detailed locality information with the material. Simon Wills (NHMUK) is thanked for drafting Fig. 1. Reviews by José Carballido (Museo Paleontológico Egidio Feruglio, Trelew, Argentina) and Blair McPhee (Universidade de São Paulo, Brazil) improved an earlier draft of the manuscript. Funds to purchase the specimens were obtained from the NHMUK Earth Sciences Departmental Investment Fund. CSCN was supported by an Undergraduate Research Bursary awarded by The Palaeontological Association. PDM’s contribution was supported by a Leverhulme Trust Early Career Fellowship (ECF-2014-662) and a Royal Society University Research Fellowship (UF160216).
References
Ait-Kaci Ahmed, A., Lingham-Soliar, T., and Broderick, T.J. 2004. Giant sauropod tracks from the Middle–Late Jurassic of Zimbabwe in close association with theropod tracks. Lethaia 37: 467–470. Crossref
Allain, R. and Aquesbi, N. 2008. Anatomy and phylogenetic relationships of Tazoudasaurus naimi (Dinosauria, Sauropoda) from the late Early Jurassic of Morocco. Geodiversitas 30: 345–424.
Allain, R., Aquesbi, N., Dejax, J., Meyer, C., Monbaron, M., Montenat, C., Richir, P., Rochdy, M., Russell, D., and Taquet P. 2004. A basal sauropod dinosaur from the Early Jurassic of Morocco. Comptes Rendus Palevol 3: 199–208. Crossref
Bandyopadhyay, S., Gillette, D.D., Ray, S., and Sengupta, D.P. 2010. Osteology of Barapasaurus tagorei (Dinosauria: Sauropoda) from the Early Jurassic of India. Palaeontology 53: 533–569. Crossref
Bonaparte, J.F. 1986a. Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétiosauridés) du Jurassique moyen de Cerro Cóndor (Chubut, Argentina). Annales de Paléontologie 72: 325–386.
Bonaparte, J.F. 1986b. The early radiation and phylogenetic relationships of the Jurassic sauropod dinosaurs, based on vertebral anatomy. In: K. Padian (ed.), The Beginning of the Age of Dinosaurs, 247–258. Cambridge University Press, Cambridge.
Bouabdellah, M. and Sangster, D.F. 2016. Geology, geochemistry, and current genetic models for major Mississippi Valley-type Pb-Zn deposits of Morocco. In: M. Bouabdellah and J. Slack (eds.), Mineral Deposits of North Africa, 1st Ed., 463–495. Springer International Publishing, Cham. Crossref
Buffetaut, E. 2005. A new sauropod dinosaur with prosauropod-like teeth from the Middle Jurassic of Madagascar. Bulletin de la Société Géologique de France 176: 483–489. Crossref
Carballido, J.L., Pol, D., Parra Ruge, M.L., Padilla Bernal, S., Páramo-Fonseca, M.E., and Etayo-Serna F. 2015. A new Early Cretaceous brachiosaurid (Dinosauria, Neosauropoda) from northwestern Gondwana (Villa de Leiva, Colombia). Journal of Vertebrate Paleontology 35: e980505. Crossref
Carballido, J.L., Rauhut, O.W.M., Pol, D., and Salgado, L. 2011. Osteology and phylogenetic relationships of Tehuelchesaurus benitezii (Dinosauria, Sauropoda) from the Upper Jurassic of Patagonia. Zoological Journal of the Linnean Society 163: 605–662. Crossref
Carballido, J.L., Salgado, L., Pol, D., Canudo, J.I., and Garrido, A. 2012. A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin; evolution and biogeography of the group. Historical Biology 24: 631–654. Crossref
Cooper, M.R. 1984. A reassessment of Vulcanodon karibaensis Raath (Dinosauria: Saurischia) and the origin of the Sauropoda. Palaeontologia Africana 25: 203–231.
Cruickshank, A.R.I. 1975. The origin of sauropod dinosaurs. South African Journal of Science 71: 89–90.
Cúneo, R., Ramezani, J., Scasso, R., Pol, D., Escapa, I., Zavattieri, A.M., and Bowring, S.A. 2013. High-precision U-Pb geochronology and a new chronostratigraphy for the Cañadón Asfalto Basin, Chubut, central Patagonia: Implications for terrestrial faunal and floral evolution in Jurassic. Gondwana Research 24: 1267–1275. Crossref
D’Emic, M.D. 2012. The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society 166: 624–671. Crossref
Gilmore, C.W. 1936. Osteology of Apatosaurus with special reference to specimens in the Carnegie Museum. Memoirs of the Carnegie Museum 11: 175–300.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 1–13. Crossref
Hadri, M., Boutakiout, M., and Pérez-Lorente, F. 2007. Nuevos yacimientos de icnitas de dinosaurios carixiensis. Sur del Alto Atlas Central (Goulmima. Marruecos). Geogaceta 41: 107–110.
Huene, F. von 1932. Die fossile Reptil-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monographien zur Geologie und Palaeontologie (series 1) 4: 1–361.
Jenny, J., Jenny-Deshusses, C., Le Marrec, A., and Taquet, P. 1980. Découverte d’ossements de Dinosauriens dans le Jurassique inférieur (Toarcien) du Haut Atlas central (Maroc). Comptes Rendus de l’Académie des Sciences à Paris, série D 290: 839–842.
Jenny, J., Le Marrec, A., and Monbaron, M. 1981. Les empreintes de pas de dinosauriens dans le Jurassique moyen du Haut Atlas central (Maroc): nouveaux gisements et precisions stratigraphiques. Géobios 14: 427–431. Crossref
Kutty, T.S., Chatterjee, S., Galton, P.M., and Upchurch, P. 2007. Basal sauropodomorphs (Dinosauria, Saurischia) from the Lower Jurassic of India: their anatomy and relationships. Journal of Paleontology 81: 1552–1574. Crossref
Läng, É. and Goussard, F. 2007. Redescription of the wrist and manus of ?Bothriospondylus madagascariensis: new data on carpus morphology in Sauropoda. Geodiversitas 29: 549–560. Crossref
Läng, E. and Mahammed, F. 2010. New anatomical data and phylogenetic relationships of Chebsaurus algeriensis (Dinosauria, Sauropoda) from the Middle Jurassic of Algeria. Historical Biology 22: 142–164.
Lapparent, A.F. 1955. Étude paléontologique des vertébrés du Jurassique d’El Mers (Moyen Atlas). Notes et Mémoires du Service Géologique du Maroc 124: 1–36.
Lydekker, R 1895. On bones of a sauropodous dinosaur from Madagascar. Quarterly Journal of the Geological Society of London 51: 329–336. Crossref
Mahammed, F., Läng, E., Mami, L., Mekahli, L., Benhamou, M., Bouterfa, B., Kacemi, A., Chérief, S., Chaouati, H., and Taquet, P. 2005. The “Giant of Ksour”, a Middle Jurassic sauropod dinosaur from Algeria. Comptes Rendus Palevol 4: 707–714. Crossref
Mannion, P.D. 2009. Review and analysis of African sauropodomorph dinosaur diversity. Palaeontologia africana 44: 108–111.
Mannion, P.D. 2010. A revision of the sauropod dinosaur genus “Bothriospondylus” with a redescription of the type material of the Middle Jurassic form “B. madagascariensis”. Palaeontology 53: 277–296. Crossref
Mannion, P.D. and Barrett, P.M. 2013. Additions to the sauropod dinosaur fauna of the Cenomanian (early Late Cretaceous) Kem Kem beds of Morocco: palaeobiogeographical implications of the mid-Cretaceous African sauropod fossil record. Cretaceous Research 45: 49–59. Crossref
Mannion, P.D., Upchurch, P., Barnes, R.N., and Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society 168: 98–206. Crossref
Marsh, O.C. 1878. Principal characters of American Jurassic dinosaurs. Part I. American Journal of Science, Series 3 16: 411–416. Crossref
McPhee, B.W., Bonnan, M.F., Yates, A.M., Neveling, J., and Choiniere, J.N. 2015. A new basal sauropod from the pre-Toarcian Jurassic of South Africa: evidence of niche-partitioning at the sauropodomorph–sauropod boundary? Scientific Reports 5: 13224. Crossref
McPhee, B.W., Bordy, E.M., Sciscio, L., and Choiniere, J.N. 2017. The sauropodomorph biostratigraphy of the Elliot Formation of southern Africa: Tracking the evolution of Sauropodomorpha across the Triassic–Jurassic boundary. Acta Palaeontologica Polonica 62: 441–465. Crossref
McPhee, B.W., Upchurch, P., Mannion, P.D., Sullivan, C., Butler, R.J., and Barrett, P.M. 2016. A revision of Sanpasaurus yaoi Young, 1944 from the Early Jurassic of China, and its relevance to the early evolution of Sauropoda (Dinosauria). PeerJ 4: e2578. Crossref
McPhee, B.W., Yates, A.M., Choiniere, J.N., and Abdala, F. 2014. The complete anatomy and phylogenetic relationships of Antetonitrus ingenipes (Sauropodiformes, Dinosauria): implications for the origins of Sauropoda. Zoological Journal of the Linnean Society 171: 151–205. Crossref
Monbaron, M., Russell, D.A., and Taquet, P. 1999. Atlasaurus imelakei, n. g., n. sp., a brachiosaurid-like sauropod from the Middle Jurassic of Morocco. Comptes Rendus de l’Académie des Sciences, Paris 329: 519–526.
Otero, A., Krupandan, E., Pol, D., Chinsamy, A., and Choiniere, J. 2015. A new basal sauropodiform from South Africa and the phylogenetic relationships of basal sauropodomorphs. Zoological Journal of the Linnean Society 174: 589–634. Crossref
Owen, R. 1842. Report on British fossil reptiles, Part II. Reports of the British Association for the Advancement of Sciences 11: 60–204.
Raath, M.A. 1972. Fossil vertebrate studies in Rhodesia: a new dinosaur (Reptilia: Saurischia) from near the Trias–Jurassic boundary. Arnoldia 5: 1–37.
Rauhut, O.W.M. 2003. Revision of Amygdalodon patagonicus Cabrera, 1947 (Dinosauria, Sauropoda). Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe 6: 173–181.
Rauhut, O.W.M. and López-Arbarello, A. 2009. Considerations on the age of the Tiouaren Formation (Iullemmeden Basin, Niger, Africa): Implications for Gondwanan Mesozoic terrestrial vertebrate faunas. Palaeogeography, Palaeoclimatology, Palaeoecology 271: 259–267. Crossref
Rauhut, O.W.M., Remes, K., Fechner, R., Cladera, G., and Puerta, P. 2005. Discovery of a short-necked sauropod dinosaur from the Late Jurassic of Patagonia. Nature 435: 670–672. Crossref
Remes, K., Ortega, F., Fierro, I., Joger, U., Kosma, R., Marín, J.M., Project PALDES, Niger Project SNHM, Ide, O.A., and Maga, A. 2009. A new basal sauropod dinosaur from the Middle Jurassic of Niger and the early evolution of Sauropoda. PLoS ONE 4: e6924. Crossref
Royo-Torres, R., Cobos, A., and Alcalá, L. 2006. A giant European dinosaur and a new sauropod clade. Science 314: 1925–1927. Crossref
Saâdi, Z., Fedan, B., Laadila, M., Azzouz, O., and Sanz de Galdeano, C. 2012. Terrigenous successions of the Late Triassic-Early Jurassic in the High Moulouya (Morocco): stratigraphy and geodynamic context. Estudios Geológicos 68: 41–56.
Seeley, H.G. 1887. On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London 43: 165–171. Crossref
Sereno, P.C., Beck, A.L., Dutheil, D.B., Larsson, H.C.E., Lyon, G.H., Moussa, B., Sadleir, R.W., Sidor, C.A., Varricchio, D.J., Wilson, G.P., and Wilson, J.A. 1999. Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs. Science 286: 1342–1347. Crossref
Stumpf, S., Ansorge, J., and Krempien, W. 2015. Gravisaurian sauropod remains from the marine late Early Jurassic (lower Toarcian) of North Eastern Germany. Geobios 48: 271–279. Crossref
Termier, H. 1942. Données nouvelles sur le Jurassique rouge à Dinosauriens du Grand et Moyen-Atlas (Maroc). Bulletin de la Société géologique de France 12: 199–207.
Upchurch, P. 1995. Evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London, Series B 349: 365–390. Crossref
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124: 43–103. Crossref
Upchurch, P. and Martin, J. 2002. The Rutland Cetiosaurus: The anatomy and relationships of a Middle Jurassic British sauropod dinosaur. Palaeontology 45: 1049–1074. Crossref
Upchurch, P. and Martin, J. 2003. The anatomy and taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the Middle Jurassic of England. Journal of Vertebrate Paleontology 23: 208–231. Crossref
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, 2nd Ed., 259–322. University of California Press, Berkeley. Crossref
Upchurch, P., Barrett, P.M., and Galton, P.M. 2007a. The phylogenetic relationships of basal sauropodomorphs: implications for the origin of sauropods. Special Papers in Palaeontology 77: 57–90.
Upchurch, P., Barrett, P.M., Zhao, X.J., and Xu, X. 2007b. A re-evaluation of Chinshakiangosaurus chunghoensis Ye vide Dong 1992 (Dinosauria, Sauropodomorpha): implications for cranial evolution in basal sauropod dinosaurs. Geological Magazine 144: 247–262. Crossref
Viglietti, P.A., Barrett, P.M., Broderick, T.J., Munyikwa, D., MacNiven, R., Broderick, L., Chapelle, K., Glynn, D., Edwards, S., Zondo, M., Broderick, P., and Choiniere, J.N. 2018. Stratigraphy of the Vulcanodon type locality and its implications for regional correlations within the Karoo Supergroup. Journal of African Earth Science 137: 149–156. Crossref
Wagensommer, A., Latiano, M., and Nicosia, U. 2010. First report of dinosaur footprints from Madagascar: two tracksites from the Middle Jurassic Bemaraha Formation. Ichnos 17: 127–136. Crossref
Wedel, M.J. 2003. The evolution of vertebral pneumaticity in sauropod dinosaurs. Journal of Vertebrate Paleontology 23: 344–357. Crossref
Wild, R. 1978. Ein Sauropoden-Rest (Reptilia, Saurischia, Sauropodomorpha) aus dem Posidonienschiefer (Lias, Toarcium) von Holzmaden. Stuttgarter Beiträge zur Naturkunde, Serie B 41: 1–15.
Wilson, J.A. 1999. A nomenclature for vertebral laminae in sauropods and other saurischian dinosaurs. Journal of Vertebrate Paleontology 19: 639–653. Crossref
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society of London 136: 217–276. Crossref
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5: 1–68. Crossref
Yates, A.M. 2007. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria). Special Papers in Palaeontology 77: 9–55.
Yates, A.M., Hancox, J.P., and Rubidge, B.S. 2004. First record of a sauropod dinosaur from the Upper Elliot Formation (Early Jurassic) of South Africa. South African Journal of Science 100: 504–506.
Yates, A.M., Wedel, M.J., and Bonnan, M.F. 2012. The early evolution of postcranial skeletal pneumaticity in sauropodomorph dinosaurs. Acta Palaeontologica Polonica 57: 85–100. Crossref
Zhang, Y.H. 1988. The Middle Jurassic Dinosaur Fauna From Dashanpu, Zigong, Sichuan. III. Sauropod Dinosaurs. 1. Shunosaurus [in Chinese, with English abstract]. 106 pp. Sichuan Publishing House of Science and Technology, Chengdu.
Acta Palaeontol. Pol. 63 (1): 147–157, 2018
https://doi.org/10.4202/app.00425.2017