
First records of diapsid Palacrodon from the Norian, Late Triassic Chinle Formation of Arizona, and their biogeographic implications
BEN T. KLIGMAN, ADAM D. MARSH, and WILLIAM G. PARKER
Kligman, B.T., Marsh, A.D., and Parker, W.G. 2018. First records of diapsid Palacrodon from the Norian, Late Triassic Chinle Formation of Arizona, and their biogeographic implications. Acta Palaeontologica Polonica 63 (1): 117–127.
Vertebrates from the Triassic Period have broadly disparate tooth shapes and dentition patterns, the result of intense morphospace experimentation following the Permo-Triassic extinction. Here, we describe fragmentary tooth-bearing jaw elements of the rare Gondwanan Middle Triassic taxon Palacrodon that represent first occurrences from the Upper Triassic Chinle Formation in northeastern Arizona. These specimens come from two notably diverse macrovertebrate and microvertebrate assemblages, the “coprolite layer” horizon in Petrified Forest National Park, and the Placerias Quarry near St. Johns, Arizona. Palacrodon is diagnosed by the presence of acrodont maxillary and dentary dentition of labiolingually-widened, bulbous teeth, which are fused to one another mesiodistally, and are pyramidal in lateral view. We agree with previous workers and refer Palacrodon to Diapsida based on the presence of a posterior jugal spur and a quadrate that has a quadratojugal foramen and a concave posterior surface. Formerly known only from the Cynognathus Assemblage Zone in southern Africa and Antarctica, the presence of Palacrodon in the Norian of North America represents a temporal range extension of at least 15 million years and represents the first tetrapod genus known to be present in the Karoo Basin, Fremouw Formation, and the Upper Triassic of the southwestern United States. Range extensions such as this suggest that some vertebrate biostratigraphic hypotheses for the Triassic may be less robust than previously believed and subject to significant sampling biases.
Key words: Diapsida, Palacrodon, microvertebrate, biostratigraphy, Triassic, Chinle Formation, North America.
Ben T. Kligman [benkligman@gmail.com], Adam D. Marsh [adam_marsh@nps.gov], and William G. Parker [william_ parker@nps.gov], Division of Science and Resource Management, 1 Park Road, #2217, Petrified Forest National Park, Arizona 86028 USA.
Received 18 September 2017, accepted 2 January 2018, available online 8 February 2018.
Copyright © 2018 B.T. Kligman et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Palacrodon browni is a Triassic diapsid previously known from only three localities in South Africa and Antarctica (Broom 1906; Gow 1992, 1999). Palacrodon browni was erected from a specimen (SAM-PK-6215) collected in the Cynognathus Assemblage Zone (CAZ) of South Africa, based on a small isolated lower jaw with bulbous acrodont teeth, that are fused to one another mesiodistally (Broom 1906). Broom (1906: 380) classified the jaw as that of the “earliest known true rhynchocephalian” based on the acrodont implantation of the teeth and its stratigraphic position in what had been thought to be the Upper Triassic. The rhynchocephalian affinities of Palacrodon were supported by Hoffstetter (1955) and Huene (1956) who both classified Palacrodon as the earliest sphenodontian. Malan (1963: 215) disputed the rhynchocephalian affinities of Palacrodon, stating “[Palacrodon] could be an aberrant procolophonid or lizard just as easily as an aberrant rhynchocephalian.”
James Kitching collected the partial skull of a small diapsid (BP/1/5296) with labiolingually broadened acrodont dentition in 1970 from the lower Fremouw Formation (Sidor et al. 2008) in Antarctica, which was later described and named Fremouwsaurus geludens (Gow 1992). After noting the strong resemblance of the unique dentition of Fremouwsaurus to the dentition of Palacrodon, Gow (1999) synonymized Fremouwsaurus geludens with Palacrodon browni. He also described additional material of Palacrodon consisting of a partial right maxilla and dentary (BP/1/5672) collected in 1996 from CAZ rocks in South Africa (Gow 1999). Based on BP/1/5296 Palacrodon was diagnosed as a diapsid based on the presence of a posterior jugal spur, a quadrate with a quadratojugal foramen, and a curved lateral profile (Gow 1992, 1999). Previous classifications of Palacrodon as the earliest rhynchocephalian (Broom 1906; Hoffstetter 1955; Huene 1956) were rejected by Gow (1992, 1999) owing to the absence of a quadratojugal and the presence of a lacrimal in BP/1/5296.
We present here two new occurrences of the genus Palacrodon from the Upper Triassic Chinle Formation of northeastern Arizona. The oldest occurrence (PEFO 37247) was collected from the Blue Mesa Member of the Chinle Formation (middle Norian, ~221 Ma; Nordt et al. 2015) in Petrified Forest National Park, Arizona. The next occurrence is from lower part of the Chinle Formation (middle Norian, ~219 Ma; Ramezani et al. 2014) at the Placerias Quarry near St. Johns, Arizona. The Palacrodon specimens from the Placerias Quarry (MNA V3684, V11247) were collected during micro-screening efforts led by the Museum of Northern Arizona in 1978 and reported in Jacobs and Murry (1980), where they identified MNA V3684 as a member of Procolophonidae. This same specimen was briefly described and figured in Kaye and Padian (1994: fig. 109A, B), where it was identified as belonging to a reptile of uncertain taxonomic assignment. This specimen’s similarity to “Clevosaurus” latidens (Fraser, 1993) has also been noted. We recognized the similarity of this specimen to Palacrodon browni and subsequently identified additional Palacrodon material in the Placerias Quarry microvertebrate collection at the Museum of Northern Arizona. These occurrences represent the first reports of Palacrodon from the Norian and from Laurasia.
Institutional abbreviations.—BP, Bernard Price Institute for Paleontological Research, University of the Witwatersrand, Johannesburg, South Africa; MNA, Museum of Northern Arizona, Flagstaff, USA; PEFO, Petrified Forest National Park, Arizona, USA; SAM-PK, South African Museum, Iziko Museums of South Africa, Cape Town, South Africa.
Geological setting
The Upper Triassic Chinle Formation is exposed extensively in northeastern Arizona and is divided into five members, the Mesa Redondo Member, Blue Mesa Member, Sonsela Member, Petrified Forest Member, and Owl Rock Member (Martz and Parker 2010). The Blue Mesa Member is characterized by fine-grained siltstone, mudstone, and sandstone deposited by fluvial, lacustrine, and palustrine systems (Woody 2006). The Blue Mesa Member interfingers with the overlying Sonsela Member, that is characterized by predominantly coarser-grained sediments including sandstone and pebble conglomerate deposited in a fluvial system (Martz and Parker 2010). The paleolatitude of the Chinle Formation during the Late Triassic was within 10° north of the equator (e.g., Kent and Tauxe 2005; Zeigler and Geissman 2011), and the paleoclimate during deposition of these sediments was humid and warm with seasonal monsoonal precipitation (Dubiel et al. 1991; Dubiel 1994; Nordt et al. 2015).
PEFO 37247 was collected from a lens in the upper part of the Blue Mesa Member informally referred to as the “coprolite layer” (PFV 396; Fig. 1). The “coprolite layer” is a 5–12 cm thick grey-green siltstone with abundant coprolites, osteichthyan scales, vertebrate bones, and charcoal. A diverse assemblage has been collected from this locality, and taxa have been diagnosed using apomorphy-based identifications (sensu Norell 1989; Bell et al. 2004, 2010; Nesbitt and Stocker 2008). The vertebrate assemblage includes chondrichthyans, actinopterygians, sarcopterygians, temnospondyl amphibians, cynodonts, drepanosauromorphs, lepidosauromorphs, and a diverse collection of archosauromorphs (Kligman 2015; Kligman et al. 2017). The base of the fossiliferous unit is a dark purple mudstone with abundant pyrite that suggests the deposit was formed in a shallow pond or small lake with anoxic bottom conditions. High-precision U-Pb dates obtained from detrital zircons collected above and below the “coprolite layer” horizon at PFV 396 bracket the maximum age of deposition to between 223.036 ± 0.059 and 220.123 ±0.068 Ma (Fig. 1A; Ramezani et al. 2011; Atchley et al. 2013), and the estimated age of a “lacustrine mudstone” located stratigraphically close to the location of PEFO 37247 is 221.946 Ma (Nordt et al. 2015: 1804).
MNA V3684 and MNA V11247 were collected from the Placerias Quarry, located approximately 24 miles southeast of Petrified Forest National Park, south of the town of St. Johns, Arizona (MNA 207-2; Fig. 1). This quarry was discovered and first studied by Charles L. Camp in 1927 and consists of two distinct fossiliferous layers separated by about 60 cm of unfossiliferous sediment, with the lower layer containing the bones of mostly terrestrial vertebrates including the dicynodont Placerias, and the upper layer containing mostly bones of aquatic vertebrates (Camp and Welles 1956; Jacobs and Murry 1980; Long et al. 1989; Kaye and Padian 1994; Long and Murry 1995; Fiorillo et al. 2000). The lithology of the lower level is a clay-rich mudstone matrix with abundant macrovertebrate and microvertebrate bones and carbonaceous nodules (Fiorillo and Padian 1993; Fiorillo et al. 2000). The Downs’ Quarry (MNA 207-10), located 72 m away from the Placerias Quarry contains a similar lithology and vertebrate assemblage to the Placerias Quarry (Kaye and Padian 1994; Lucas et al. 1997). Specimens of Palacrodon were collected from the lower level of the Placerias Quarry (MNA 207-2), associating them with the predominantly terrestrial assemblage. A detailed taphonomic study by Fiorillo et al. (2000) interpreted the paleoenvironment of the lower level as an ephemeral watering hole subject to seasonal drought. The combined vertebrate assemblage of both the upper and lower levels of the Placerias Quarry and Downs’ Quarry is one of the most diverse Triassic vertebrate assemblages known, including chondrichthyans, actinopterygians, sarcopterygians, temnospondyl amphibians, synapsids, drepanosauromorphs, lepidosauromorphs, and diverse archosauromorphs (Camp and Welles 1956; Jacobs and Murry 1980; Murry 1987; Kaye and Padian 1994; Long and Murry 1995; Sues 1996; Lucas et al. 1997; Hunt et al. 1998; Heckert and Lucas 2002; Nesbitt et al. 2009a; Renesto et al. 2009; Ezcurra and Brusatte 2011; Mitchell et al. 2010). Kaye and Padian (1994) presented an exploratory study of the microvertebrate assemblage found at the Placerias Quarry.
High-precision U-Pb dates obtained from detrital zircons collected from the fossiliferous horizon at the Placerias Quarry date it to 219.39 ± 0.16 Ma (Ramezani et al. 2014), which is equivalent in age to the upper part of the Lot’s Wife beds of the Sonsela Member at PEFO (Fig. 1A). This is in contrast to previous interpretations of the stratigraphic position and age of the Placerias Quarry, which placed it near the base of the Chinle Formation (e.g., Lucas et al. 1997; Renesto et al. 2009).
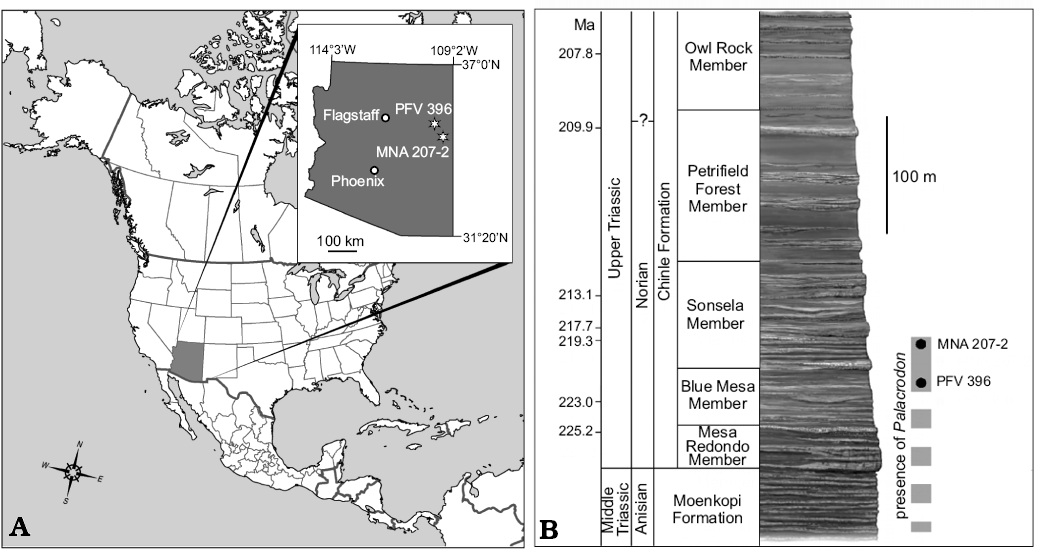
Fig. 1. Geographic and stratigraphic context of Palacrodon occurrences in Arizona. A. Map of Arizona showing the locations of PFV 396 and MNA 207-2. Map of North America modified from www.lakodosajta.info. B. Stratigraphic chart of the Chinle Formation at PEFO and the Placerias Quarry, with detrital zircon dates from Ramezani et al. (2011).
Material and methods
PEFO 37247 was collected by screenwashing and sorting matrix from the “coprolite layer” in the summer of 2015. Screenwashing was accomplished by rinsing water over small blocks of matrix through a series of five screens with a frame diameter of 8 inches, and screen diameter sizes of 6.33 mm, 4 mm, 2 mm, 1 mm, and 180 µm. All screens are made by USA Standard Test Sieve, (ASTM E-11 specification). Screened concentrate was examined under a Leica-Wild M7A microscope with a magnification 6–31 times.
In 1978 field crews from the Museum of Northern Arizona excavated 5.8 metric tons of sediment from the upper and lower fossiliferous horizons of the Placerias Quarry (MNA 207-2). This sediment was screenwashed and sorted under the leadership of Will Downs at the Museum of Northern Arizona (Jacobs and Murry 1980; Kaye and Padian 1994; Fiorillo et al. 2000). MNA V3684 and MNA V11247 were collected by this collection effort.
Photographs were taken with a Leica MZApo stereomicroscope and a Sony Nex5T digital camera, using image-stacking software HeliconFocus (v. 6.2.2 Lite, Method B depth map, full resolution, radius 50, smoothing 1). No adhesive or consolidant was used on PEFO 37247, and an unknown adhesive was used on MNA V3684.
We use the term “acrodonty” to describe the implantation of teeth on the apex of the jawbone, following the definition of Haridy et al. (2017). This term is used to describe the position of teeth in the jaw, and does not imply any mode of tooth replacement (no evidence of tooth replacement has been noted in any known Palacrodon material).
The Palacrodon specimens from the Chinle Formation are compared to Palacrodon specimens from South Africa and Antarctica in the discussion herein. Comparisons with BP/1/5672, and BP/1/5296 are made using photographs of the specimens, and comparisons with SAM-PK-6215 are made with a cast of the specimen.
Systematic palaeontology
Amniota Haeckel, 1866
Diapsida Osborn, 1903
Genus Palacrodon Broom, 1906
Type species: Palacrodon browni Broom, 1906, Aliwal North, South Africa, Middle Triassic.
Palacrodon browni Broom, 1906
Fig. 2.
Material.—PEFO 37247, fragment of a right maxilla with two complete teeth and two partial teeth. Obtained from PFV 396, the “coprolite layer” near Billing’s Gap in Petrified Forest National Park, Arizona, USA. A fossiliferous light green matrix-supported siltstone in the upper Blue Mesa Member of the Chinle Formation, between 223.036 ± 0.059 and 220.123 ± 0.068 Ma (Ramezani et al. 2011). These dates place the occurrence within the Adamanian Land Vertebrate Estimated Holochron (Martz and Parker 2017; previously the Adamanian Land Vertebrate Faunachron of Lucas and Hunt 1993). Detailed locality information is on file at Petrified Forest National Park and available to qualified researchers.
Diagnosis.—Diapsid reptile with bulbous, labio-lingually broadened teeth that are ankylosed directly onto the apex of the jaw with acrodont implantation. Teeth are fused to each other mesiodistally along their entire labiolingual width. The margin of each tooth is flanged on its mesial and distal edges. The teeth have a pyramidal lateral profile with concave mesial and distal crown surfaces. Wear facets are present on the apex of each tooth. Tooth margins form a rectangular profile in occlusal view, and have a trapezoidal profile in anteroposterior view. This combination of characters is unique to Palacrodon browni.
Description.—PEFO 37247 is a partial right maxilla with four preserved teeth, two of which are complete, and two are largely fragmented. PEFO 37247 has an anteroposterior length of 7.2 mm, a greatest dorsoventral depth of 3.8 mm, and a mediolateral width of 1.1 mm. The maxilla is incomplete along its dorsal margin and both anteroposterior ends because of damage during processing. Two nutrient foramina with a width of 0.5 mm are located on the labial side of the maxilla and open into a vacuity in the interior of the element (Fig. 2B1); these foramina are similar to those found on BP/1/5296. A slot opens posteromedially at the posterodorsal end of the bone, likely where the jugal articulated onto the maxilla.
Four teeth are present, with the two middle ones being complete. One of the two complete teeth is larger than the other (Table 1). This size difference indicates that the larger tooth was further back in the jaw than the smaller one (Gow 1992, 1999). The teeth are ankylosed directly onto the apical surface of the dentary in an acrodont fashion. There is a row of small pits each with a diameter of 0.01 mm where the labial and lingual margins of the base of each tooth meets the maxillary bone (Fig. 2C). The tooth bases bulge out 0.3 mm from the labial and lingual margins of the maxilla, making the teeth wider labiolingually than the maxilla itself. PEFO 37247 is identical to SAM-PK-6215 in that the teeth are pyramidal in lateral view and each complete tooth has a flat lemniscate wear facet on its apical surface (Fig. 2A1), cutting through the thin layer of enamel that covers the tooth surface and exposing the dentine center of the tooth. The wear facets on PEFO 37247 are nearly identical in shape to those found in the partial dentary of BP/1/5672. Concave shelves are present on the mesial and distal sides of the wear facet on each tooth, extending two-thirds of the way down the tooth (Fig. 2B2). A flange extends below the mesial and distal shelves on each tooth to meet the flange of the adjacent tooth. The teeth contact each other mesiodistally and the respective flanges are firmly fused together along this line of contact (Fig. 2A2); three such tooth-to-tooth contacts are present in this specimen.
Stratigraphic and geographic range.—Currently known from Cynognathus Assemblage Zone rocks (South Africa), the lower Fremouw Formation (Antarctica), and the Chinle Formation (Arizona, USA).
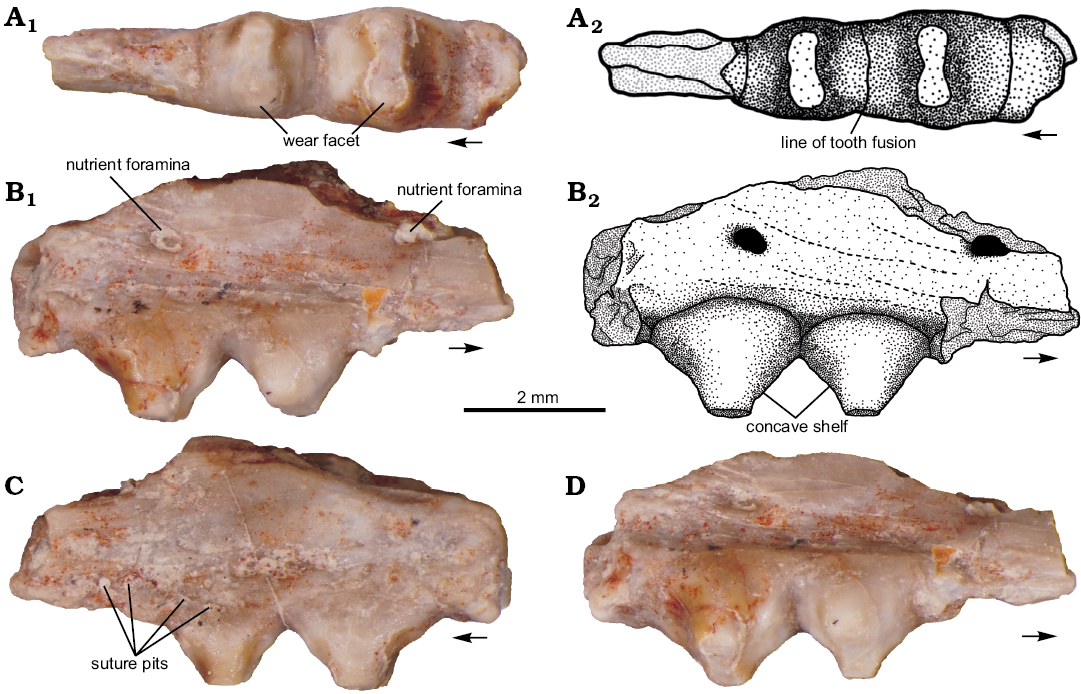
Fig. 2. Diapsid Palacrodon browni Broom, 1906 (PEFO 37247) from Norian, Petrified Forest National Park, Arizona, USA; in occlusal (A), labial (B), lingual (C), and oblique (D) views. Arrows point in anterior direction. Photographs (A1, B1, C, D), explanatory drawings (A2, B2).
Palacrodon sp.
Fig. 3.
1980 Procolophonidae; Jacobs and Murry 1980: 63.
1983 Reptilia; Tannenbaum 1983: 63, fig. 109.
1993 Clevosaurus latidens; Fraser 1993: 138.
1994 Reptilia; Kaye and Padian 1994: 189, fig. 9.109.
2000 Procolophonidae; Edwards 2000: 320–321, fig. 4.6.3A.
Material.—MNA V3684, fragment of marginal dentition with two complete teeth; MNA V11247, isolated tooth. Obtained from MNA 207-2, the Placerias Quarry, St. John’s, USA. A fossiliferous horizon with abundant macrovertebrate and microvertebrate bones and carbonaceous nodules in a clay rich mudstone matrix (Fiorillo et al. 2000) from the Chinle Formation, dated to 219.39 ± 0.16 Ma based on U-Pb detrital zircon dates (Ramezani et al. 2014). These dates place the occurrence solidly in the Adamanian Land Vertebrate Estimated Holochron (Martz and Parker 2017). Detailed locality information is on file at Museum of Northern Arizona and is available to qualified researchers.
Diagnosis.—Differs from Palacrodon browni in having teeth that have a bicuspid apex and a mesially recurved labial profile. The base of the distal shelf is concave in labial view, halfway towards the apex becoming convex to the apex of the tooth. The mesial tooth shelf is concave in labial view, and the apical edge of the tooth reaches an angle just greater than 90°. The labiolingually-broadened apex of the tooth possesses two cusps, one on the labial side, and one on the lingual side. These cusps are joined by a narrow ridge that forms a saddle between the cusps in mesiodistal view.
Description.—MNA V3684 is a tooth-bearing fragment of a maxilla or dentary (Fig. 3A). The assignment to either element is difficult because the bone is broken along all margins except for the tooth-bearing surface, and will be referred to as a dentary in this description. Two complete teeth are preserved. The jaw fragment has a mesiodistal length of 7.5 mm, and a labiolingual width of 3.6 mm, and is incomplete on along its labial, lingual, ventral, posterior, and anterior surfaces. Measurements of the teeth are found in Table 1.
Two teeth are present. One of the two complete teeth is larger than the other (Table 1). Palacrodon teeth increase in size distally, indicating that the larger tooth was further distal in the jaw than the smaller one (Gow 1992, 1999). The teeth are ankylosed directly onto the apical surface of the dentary in an acrodont fashion, and are bulbous, bulging out slightly from the margins of the jaw. No tooth root is apparent in the jaw where it is fractured less than 1 mm below the tooth bases and it can be assumed that if rooting is present, it is very shallow. There is a row of small pits on the lingual side of the distal tooth, each with a diameter of 0.01 mm where the lingual margin of the tooth base meets bone, similar to the pitting seen in PEFO 37247 (Fig. 2C).
The teeth are rectangular in occlusal view with most of their surface taken up by mesial and distal shelves that meet at the apex of the tooth. The base of the mesial shelf starts as a flange that meets and fuses to the tooth distal to it, and is concave in labial view. Halfway towards the apex the shelf becomes convex. The distal tooth shelf is concave in labial view and reaches an angle just greater than 90° with respect to the jaw, creating a recurved profile in labial view. The labiolingually-broadened apex of the teeth possesses two cusps, one on the labial side, and one on the lingual side (Fig. 3A2). These cusps are joined by a narrow ridge that forms a concave saddle between the cusps in mesial view (Fig. 3B1). The mesial and distal shelves are concave basally, with maximum concavity at the labiolingual midline of the teeth forming a slight basin. The presence of these basins on the mesial and distal sides of the apex creates a constriction of the apical ridge at the labiolingual midline of the teeth, and the ridge expands to its widest at the cusps on either side of the apex. The apex of the posterior tooth exhibits a lemniscate wear facet spreading across both cusps and the saddle between them. The apex of the anterior tooth has slight wear facets on the tips of both cusps, but no wear on the saddle connecting them. These wear facets cut through a thin layer of enamel exposing dentine below.
The tooth-bearing fragment described herein is likely from the posterior or middle of the maxilla or dentary, because as seen in BP/1/5296 and SAM-PK-6215, the teeth in Palacrodon maxillae and dentaries become increasingly cylindrical and peg-like towards the anterior end of the jaw and lose their mesiodistal contact.
MNA V11247 (Fig. 3B) is an isolated tooth attached to a fragment of jaw. The tooth is 2.7 mm in mesiodistal length and 3.2 mm in labiolingual width, and 2.1 mm at its greatest apicobasal height. Remnant fragments of adjacent teeth can be seen fused to both the mesial and distal margins of this tooth and a small fragment of bone is attached to the base of the tooth on its labial or lingual side. Like the anteriormost tooth in MNA V3684, this tooth is worn largely on the apex of both cusps exposing dentine, and the wear facets extend onto the saddle connecting the cusps creating a lemniscate shape.

Fig. 3. Diapsid Palacrodon sp. from Norian, Placerias Quarry, St. John’s, USA. A. MNA 3684, fragment of marginal dentition with two complete teeth in occlusal (A1, A2), labial (A3, A4), lingual (A5), and oblique (A6) views. B. MNA V11247, isolated tooth in occlusal (B1) and distal (B2) views. Arrows point in anterior direction. Photographs (A1, A3, A5, A6, B), explanatory drawings (A2, A4).
Discussion
Determining the evolutionary relationships of Palacrodon is difficult because of the fragmentary nature of all known specimens, which include one partial skull and multiple fragmentary maxillae and dentaries (Gow 1992, 1999). The partial skull of Palacrodon (BP/1/5296) has several features diagnostic to Diapsida including the presence of a posterior jugal spur and a bowed quadrate with a quadratojugal foramen and a concave posterior surface (Gow 1992, 1999). We agree with this assignment, and while Gow (1992, 1999) did not provide a phylogenetic context for his diagnosis, these are characters that are commonly used to diagnose Diapsida (Benton 1985; Gauthier et al. 1988). It is also worth noting that BP/1/5296 lacks a quadratojugal and accordingly lacks an infratemporal fenestra and lower temporal bar. The lower temporal bar is lost and reacquired multiple times within diapsid lineages (Gauthier et al. 1988; Müller 2004), so until additional Palacrodon material is found this feature is not currently informative for determining its relationships.
The dentition of Palacrodon bears a unique suite of dental characters that is unlike any other Triassic diapsid, with posterior maxillary and dentary teeth displaying pyramidal labiolingual profiles, labiolingual-widening, flat apical wear facets, and mesiodistal fusion of adjacent teeth at their bases (Broom 1906; Malan 1963; Gow 1992, 1999). Other Triassic diapsids have labiolingually widened teeth, including the allokotosaurids Teraterpeton hrynewichorum Sues, 2003 and the three species of Trilophosaurus (Mueller and Parker 2006; Spielmann et al. 2008; Nesbitt et al. 2015). Unlike Palacrodon, the teeth of Trilophosaurus are characterized by three raised cusps, thecodont implantation, and a lack of mesiodistal tooth to tooth fusion (Gregory 1945). The bulbous teeth of Teraterpeton superficially resemble those of Palacrodon, but their crown structure, bulbous columnar shape, and implantation are significantly different (Sues 2003).
Because Palacrodon is likely a diapsid reptile, we refrain from extensive comparison of its dentition to those of procolophonids, but note that no known procolophonid displays mesiodistal fusion of adjacent teeth (Cisneros 2008). While many rhynchocephalians have teeth that technically contact each other mesiodistally, Palacrodon is thought not to belong to that group owing to the temporal bar and quadratojugal, the presence of lacrimal, and the fact that the quadratojugal foramen passes through the quadrate (Gow 1992). Several Late Triassic sphenodontians share some characteristics with Palacrodon, including acrodont dentition with transversely widened teeth that touch each other mesiodistally, such as “Clevosaurus” latidens Fraser, 1993, Pelecymala robustus Fraser, 1986, and Sphenotitan leyesi Martinez, Apaldetti, Colombi, Praderio, Fernandez, Santi Malnis, Correa, Abelin, and Alcobar, 2013 (Fraser 1986, 1993; Martinez et al. 2013). However, unlike Palacrodon, sphenodontian teeth are not bulbous and lack flat apical wear facets. Moreover sphenodontian teeth display vertical wear facets on the lingual side of the maxillary teeth and on the labial side of the dentary teeth, thought to be caused by shearing or propalinal action on a vertical plane between the teeth of the upper and lower jaws (Fraser 1986, 1993; Martinez et al. 2013). This is indicative of a different mode of jaw movement during mastication than what would be present in Palacrodon, which has flat wear facets on the apical surfaces of its teeth.
PEFO 37247 is referred here to Palacrodon browni, the species that has been used to refer all previously known Palacrodon material, which share the same general dental structure despite some slight variation in the dentition of different specimens (Broom 1906; Gow 1999). Palacrodon material described by Gow (1999) includes BP/1/5672, BP/1/5296, SAM-PK-6215, and SAM-PK-5871. A range of morphological variation can be seen in the posterior maxillary and dentary teeth of these specimens as well as the new Palacrodon material described above, specifically by the apicobasal height to transverse width quotient, the mesiodistal length to transverse width quotient, and the apicobasal height to mesiodistal length quotient (Table 1). Additionally, these specimens display a range of the degree of wear on the apical surface of the teeth. The morphology of PEFO 37247 places this specimen well within the range of morphological variation seen in other Palacrodon browni specimens, and allows for its assignment to this taxon. Besides the morphological variation in dentition noted above, the labial side of the dentary in BP/1/5296 is covered in pits and grooves (Gow 1992), whereas on the holotype dentary SAM-PK-5871 the labial side of the dentary is smooth. The presence of more than one species may account for this variation, but more material would need to be described to make this conclusion.
Table 1. Comparisons of tooth measurements and quotients from Palacrodon specimens. Abbreviations: ant., anterior teeth; post., posterior teeth; NA, not available.
|
Measurement |
Maxilla |
Dentary |
Unknown |
||||||
|
PEFO 37247 |
BP/1/5672 |
BP/1/5296 |
BP/1/5296 |
BP/1/5672 |
SAM-PK-6215 |
MNA V3684 |
|||
|
post. |
ant. |
post. |
post. |
post. |
post. |
post. |
post. |
ant. |
|
|
Maximal labial apicobasal height (mm) |
1.5 |
1.1 |
1.8 |
1.9 |
1.9 |
4.1 |
1.9 |
2.5 |
2.4 |
|
Maximal transverse width (mm) |
2.2 |
1.7 |
4.9 |
not |
not |
7.9 |
1.9 |
3.7 |
3.5 |
|
Maximal mesiodistal length (mm) |
2.1 |
1.8 |
2.3 |
3.0 |
3.1 |
5.6 |
2.4 |
3.2 |
3.1 |
|
Apicobasal height to transverse width |
0.68 |
0.65 |
0.36 |
NA |
NA |
0.52 |
0.79 |
0.68 |
0.69 |
|
Mesiodistal length to transverse width |
0.96 |
1.05 |
0.46 |
NA |
NA |
0.71 |
1.3 |
0.86 |
0.89 |
|
Apicobasal height to mesiodistal length |
0.71 |
0.61 |
0.78 |
0.63 |
0.61 |
0.73 |
0.79 |
0.78 |
0.77 |
MNA V3684 and MNA V11247 exhibit several features that differentiate them from other known Palacrodon material. The teeth of these specimens lack the mesiodistal symmetry seen in the pyramidal teeth of P. browni, the MNA specimens are recurved in labial view and lack a prominent mesial flange. The tooth apices also differ. In MNA V3684 and MNA V11247 the apices of the teeth are bicuspid with minimal wear on the cusps, whereas in P. browni a flat wear facet occupies the entire apical surface of the tooth. This difference in wear patterns may be explained by ontogenetic change, where younger individuals begin with bicuspid teeth, and as wear accumulates through life the teeth are ground down to the degree seen in Palacrodon browni, where there is a continuous flat facet on the apex of the tooth. The occlusal pattern apparent in Palacrodon browni and Palacrodon sp. (MNA V3684, V11247) appears to show the same jaw closing mechanism, where tooth wear only occurs on the apical surface of the crown, lending support for an ontogenetic explanation for the differences in tooth wear. Additionally, the recurved nature of the teeth in MNA V3684 and MNA V11247 would be masked as wear on the teeth accumulated, making the teeth pyramidal in labiolingual view with lemniscate wear facets, much like Palacrodon browni. A growth series of Palacrodon dentition is required to support any argument regarding tooth ontogenetic variation in this taxon, and because there are no known Palacrodon growth series, we refrain from making further ontogenetic conclusions about MNA V3684 and MNA V11247, and refer them to Palacrodon sp.
The presence of Palacrodon in the middle Norian (Adamanian) of the southwest United States significantly expands the stratigraphic range and biogeographic distribution of this genus (Fig. 4), as it was previously only known from the Middle Triassic of South Africa and Antarctica (Gow 1999; Sidor et al. 2008). Although the exact provenance of the holotype material of Palacrodon browni is unknown, Gow (1999) believes that it came from the Cynognathus Assemblage Zone (CAZ). The base and subzones A and B of the CAZ are thought to be Early Triassic (Olenekian) in age, but the uppermost subzone C may be Middle Triassic (Anisian) in age based on the presence of Paracyclotosaurus (Hancox 2000; Damiani and Hancox 2003; Abdala et al. 2005; Ogg 2012). The occurrence of Palacrodon from the Driefontein in the Paul Roux District locality in South Africa is also from the CAZ (Gow 1999; Sidor et al. 2008). The Antarctica occurrence of Palacrodon came from the lower Fremouw Formation which is of uncertain Middle Triassic age, and may be older than the CAZ based on the biostratigraphic correlation of the upper Fremouw Formation to the CAZ in South Africa (Kitching 1995; Sidor et al. 2007, 2008). The exact age of the CAZ in South Africa is not known, and can only be inferred through biostratigraphic correlation, though recent detrital zircon U-Pb dating of Cynognathus-bearing rocks from the Puesto Viejo Group in Argentina indicates that either all CAZ rocks are of Carnian age (235.8 ± 2.0 Ma; Ottone et al. 2014), or that the inferred Middle Triassic age of the CAZ in Argentina cannot be correlated to other CAZ strata. Therefore, the relative ages of the South Africa and Antarctica Palacrodon occurrences are difficult to determine, and may be Anisian to Carnian. In either case, the presence of Palacrodon from middle Norian rocks in the Chinle Formation extends the stratigraphic range of Palacrodon by at least 15 million years. It also makes Palacrodon the only tetrapod genus known to be present in both the Karoo Basin and the Upper Triassic of the southwestern United States.
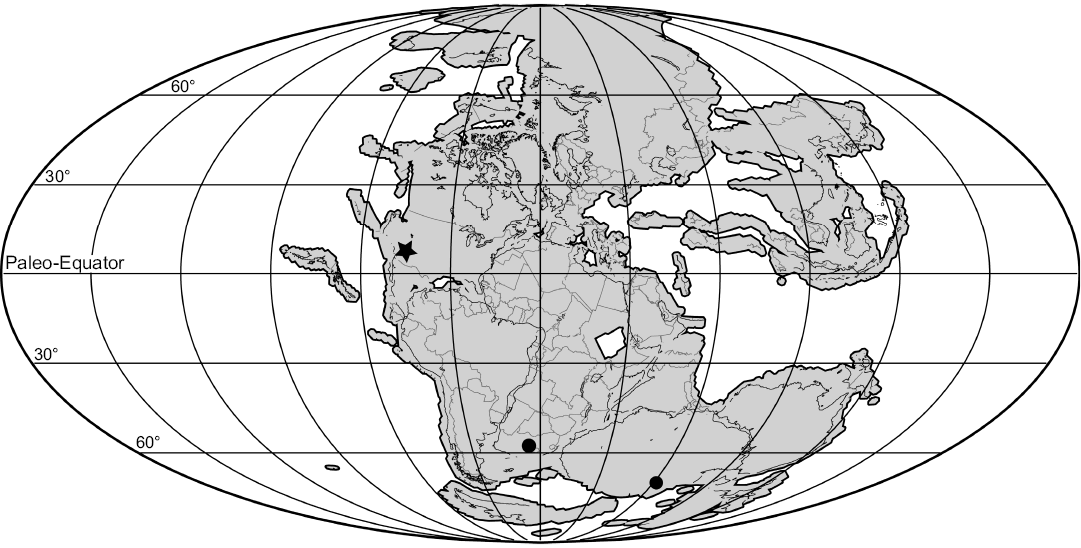
Fig. 4. Paleoreconstruction map of Pangaea in the Triassic with Gondwanan occurrences of Palacrodon marked with circles and Laurasia occurrences of Palacrodon marked with a star (note the two Arizona occurrences overlap in this scale). Paleogeography modified from www.pubs.usgs.gov/gip/dynamic/historical.html.
The actinopterygian Saurichthys is a taxon whose only known Late Triassic freshwater occurrence is at the same “coprolite layer” locality as Palacrodon (PFV 396; Kligman et al. 2017). Freshwater members of this genus have been found at multiple locations in Gondwana in the Early and Middle Triassic (Romano et al. 2012). The discovery of Palacrodon and freshwater Saurichthys at PFV 396, and of Palacrodon at the Placerias Quarry, taxa previously known only from Gondwana, is evidence of greater homogeneity between high and low latitude vertebrate communities in the Triassic.
The Late Triassic Land Vertebrate biozones first proposed by Gregory (1957) were based on hypothesized turnovers of phytosaurian archosauriforms and later supplemented by the ranges and purported turnovers of aetosaurs and other taxa (e.g., Lucas and Hunt 1993 ; Hunt et al. 1998; Heckert 2004; Heckert and Lucas 2006). Most significant in the Chinle Formation is the Adamanian–Revueltian turnover, which for years was hypothesized to approximate the Carnian–Norian boundary (e.g., Long and Padian 1986; Lucas and Hunt 1993; Lucas 1998, 2010), but is now recognized to represent primarily a shift in aetosaur, phytosaur, and palynomorph taxa (Parker and Martz 2011; Reichgelt et al. 2013; Baranyi et al. 2017). The continued discovery of vertebrate taxa such as Saurichthys (Kligman et al. 2017) and Palacrodon as well as long-lived species such as Vancleavea campi (Parker and Barton 2008; Nesbitt et al. 2009b) in the Adamanian of the Chinle Formation demonstrate that perceived stratigraphic ranges of vertebrate taxa in the Triassic may be significantly affected by sampling as well as the conservation of key phylogenetic characters.
The occurrence of Palacrodon browni (PEFO 37247) has two important and conflicting implications for Palacrodon browni as a species, bearing in mind that the specimen from the Chinle Formation is only known from a fragmentary jaw. Either Palacrodon browni is a long lived, broadly-distributed taxon, or its dentition is conserved and distinguishing some taxa in this group to the species level based solely on dentition is difficult. The only known specimen more complete than fragmentary jaw material is the partial skull of BP/1/5296, and the other occurrences are all from geographically disparate locations of sometimes great stratigraphic and chronologic separation. The possibility that the dentition of Palacrodon browni reflects a conserved morphology throughout much of the group cannot be evaluated further without more complete skeletons that might show morphological variation in bones other than jaws. This same concept should be applied to other species which are known and classified from one or a few bones bearing distinct apomorphies as is common with microvertebrates. Besides Palacrodon browni, other taxa in the Chinle Formation known from only a few diagnostic bones and known from multiple localities include Lissodus humblei, Xenacanthus moorei, Colognathus obscurus, Crosbysaurus harrisae, Acallosuchus rectori, Kraterokheirodon colberti, and Protecovasaurus lucasi (Long and Murry 1995; Heckert and Lucas 2006; Irmis and Parker 2005). Many more taxa fit this criterion globally in the fossil record. Thus, other than the well-developed biostratigraphic zonations of phytosaurs and aetosaurs in North America (and possibly Europe; Kammerer et al. 2016; Martz and Parker 2017), the use for Triassic biostratigraphy of taxa known only from specific skeletal elements, including microvertebrate taxa, should be utilized with caution. Their biostratigraphic utility, even across epoch boundaries, may be less robust than previously believed and appear highly susceptible to sampling biases.
Conclusions
The new occurrences of Palacrodon from the Chinle Formation of the southwestern United States provide further evidence of a little-known group of diapsids bearing highly apomorphic dentition, adding to this group’s diversity, and morphological variation. The range of morphological variation of Palacrodon material described herein opens the possibility that more than one species exists, and that Palacrodon may display ontogenetic variation in their tooth morphology. The presence of Palacrodon in the Norian marks a large temporal range expansion, indicating that this taxon is present in strata at least 15 million years younger than previously known. Additionally, the presence of Palacrodon in Laurasia is a significant geographic range expansion from its previously Gondwanan distribution, as it is now known from Antarctica, Africa, and North America. These temporal and geographic range extensions of Palacrodon demonstrate the uncertainty involved with using fragmentary fossils in biostratigraphic hypotheses.
Acknowledgements
We thank Adam Pritchard (Yale University, New Haven, USA) for providing photographs of BP/1/5672, BP/1/5296, and SAM-PK-5871. Christian Sidor (University of Washington, Seattle, USA) provided casts of SAM-PK-5781. David and Janet Gillette (both MNA) facilitated our use of the Placerias Quarry microvertebrate collection. Funding for BTK was provided by the Petrified Forest Museum Association. The efforts of a team of MNA researchers and students resulted in the collection of MNA V3684. Nicholas Fraser (National Museums Scotland, Edinburgh, UK) and Andrew Heckert (Appalachian State University, Boone, USA) provided detailed and helpful reviews of the manuscript. This is Petrified Forest National Park Paleontological Contribution No. 52. The conclusions presented here are those of the authors and do not represent the views of the United States Government.
References
Abdala, F., Hancox, P.J., and Neveling, J. 2005. Cynodonts from the uppermost Burgersdorp Formation, South Africa, and their bearing on the biostratigraphy and correlation of the Triassic Cynognathus assemblage zone. Journal of Vertebrate Paleontology 25: 192–199. Crossref
Atchley, S.C., Nordt, L.C., Dworkin, S.I., Ramezani, J., Parker, W.G., Ash, S.R., and Bowring, S.A. 2013. A linkage among Pangean tectonism, cyclic alluviation, climate change, and biologic turnover in the Late Triassic: the record from the Chinle Formation, Southwestern United States. Journal of Sedimentary Research 83: 1147–1161. Crossref
Baranyi, V., Reichgelt, T., Olsen, P.E., Parker, W.G., and Kürschner, W.M. 2017. Norian vegetation history and related environmental changes: New data from the Chinle Formation, Petri ed Forest National Park (Arizona, SW USA). GSA Bulletin [published online].
Bell, C.J., Gauthier, J.A., and Bever, G.S. 2010. Covert biases, circularity, and apomorphies: a critical look at the North American Quaternary Herpetofauna Stability Hypothesis. Quaternary International 217: 30–36. Crossref
Bell, C.J., Head, J.J., and Mead, J.I. 2004. Synopsis of the herpetofauna from Porcupine Cave. In: A.D. Barnosky (ed.), Biodiversity Response to Climate Change in the Middle Pleistocene: The Porcupine Cave Fauna from Colorado, 117–126. University of California Press, Berkeley. Crossref
Benton, M.J. 1985. Classification and phylogeny of the diapsid reptiles. Zoological Journal of the Linnean Society 84: 97–164. Crossref
Broom, R. 1906. On a new South African Triassic rhynchocephalian. Transactions of the South African Philosophical Society 16: 379–380. Crossref
Camp, C.L. and Welles, S.P. 1956. Triassic dicynodont reptiles. Part I, The North American genus Placerias. Memoirs of the University of California 13: 255–304.
Cisneros, J.C. 2008. Phylogenetic relationships of procolophonid parareptiles with remarks on their geological record. Journal of Systematic Palaeontology 6: 345–366. Crossref
Damiani, R.J. and Hancox, P.J. 2003. New mastodonsaurid temnospondyls from the Cynognathus Assemblage Zone (Upper Beaufort Group; Karoo Basin) of South Africa. Journal of Vertebrate Paleontology 23: 54–66. Crossref
Dubiel, R.F. 1994. Triassic deposystems, paleogeography, and paleoclimate of the Western Interior. In: M.V. Caputo, J.A. Peterson, and K.J. Francyzk (eds.), Mesozoic Systems of the Rocky Mountain Region, USA, 133–168. Rocky Mountain Section, Society for Sedimentary Geology (SEPM), Denver.
Dubiel, R.F., Parrish, J.T., Parrish, J.M., and Good, S.C. 1991. The Pangean Megamonsoon: evidence from the Upper Triassic Chinle Formation, Colorado Plateau. Palaios 6: 347–370. Crossref
Edwards, B.T. 2000. The Upper Triassic Microvertebrate Assemblage of Ruthin Quarry, South Wales. 393 pp. Ph.D. Dissertation, University College London, London.
Ezcurra, M.D. and S.L. Brusatte. 2011. Taxonomic and phylogenetic reassessment of the early neotheropod dinosaur Camposaurus arizonensis from the Late Triassic of North America. Palaeontology 54: 763–772. Crossref
Fiorillo, A.R. and Padian, K. 1993. Taphonomy of the Late Triassic Placerias Quarry (Petrified Forest Member, Chinle Formation) of eastern Arizona. New Mexico Museum of Natural History and Science Bulletin 3: 133–134.
Fiorillo, A.R., Padian, K., and Musikasinthorn, C. 2000. Taphonomy and depositional setting of the Placerias Quarry (Chinle Formation: Late Triassic, Arizona). Palaios 15: 373–386. Crossref
Fraser, N.C. 1986. New Triassic sphenodontids from South West England and a review of their classification. Palaeontology 29: 165–186.
Fraser, N.C. 1993. A new sphenodontian from the early Mesozoic of England and North America: implications for correlating early Mesozoic continental deposits. New Mexico Museum of Natural History and Science Bulletin 3: 135–139.
Gauthier, J., Estes, R., and Queiroz, K.D.E. 1988. A phylogenetic analysis of Lepidosauromorpha. In: R. Estes and G. Pregill (eds.), Phylogenetic Relationships of the Lizard Families: Essays Commemorating Charles L. Camp, 15–98. Stanford University Press, Redwood City.
Gow, C.E. 1992. An enigmatic new reptile from the Lower Triassic Fremouw Formation of Antarctica. Palaeontologia Africana 29: 21–23.
Gow, C.E. 1999. The Triassic reptile Palacrodon browni Broom, synonymy and a new specimen. Palaeontologia Africana 35: 21–23.
Gregory, J.T. 1945. Osteology and relationships of Trilophosaurus. University of Texas Publication 4401: 273–359.
Gregory, J.T. 1957. Significance of fossil vertebrates for correlation of Late Triassic continental deposits of North America. Report of the 20th Session, Intemational Geological Congress Section II: 7–25.
Hancox, P.J. 2000. The continental Triassic of South Africa. Zentralblatt für Geologie und Paläeontologie Teil 1 1998: 1285–1324.
Haridy, Y., LeBlanc, A.R., and Reisz, R.R. 2017. The Permian reptile Opisthodontosaurus carrolli: a model for acrodont tooth replacement and dental ontogeny. Journal of Anatomy [published online].
Heckert, A.B. 2004. Late Triassic microvertebrates from the lower Chinle Group (Otischalkian–Adamanian: Carnian), southwestern USA. New Mexico Museum of Natural History and Science Bulletin 27: 1–170.
Heckert, A.B. and Lucas, S.G. 2002. Acaenasuchus geoffreyi (Archosauria: Aetosauria) from the upper Triassic Chinle Group: juvenile of Desmatosuchus haplocerus. New Mexico Museum of Natural History and Science Bulletin 21: 205–214.
Heckert, A.B. and Lucas, S.G. 2006. Micro-and small vertebrate biostratigraphy and biochronology of the Upper Triassic Chinle Group, southwestern USA. New Mexico Museum of Natural History and Science Bulletin 37: 94–104.
Hoffstetter, R. 1955. Rhynchocephalia. In: J. Piveteau (ed.), Traité de Paléontologie 5, Amphibiens, Reptiles, Oiseaux, 556–576. Masson et Cie, Paris.
Huene, F. von 1956. Paläontologie und Phylogenie der Niederen Tetrapoden. 716 pp. Gustav Fischer, Jena.
Hunt, A.P., Lucas, S.G., Heckert, A.B., Sullivan, R.M., and Lockley, M.G. 1998. Late Triassic dinosaurs from the western United States. Geobios 31: 511–531. Crossref
Irmis, R.B. and Parker, W.G. 2005. Unusual tetrapod teeth from the Upper Triassic Chinle Formation, Arizona, USA. Canadian Journal of Earth Sciences 42: 1339–1345. Crossref
Jacobs, L.L. and Murry, P.A. 1980. The vertebrate community of the Triassic Chinle Formation near St. Johns, Arizona. In: L.L. Jacobs (ed.), Aspects of Vertebrate History: Essays in Honor of Edwin Harris Colbert, 55–71. Museum of Northern Arizona Press, Flagstaff.
Kammerer, C.F., Butler, R.J., Bandyopadhyay, S., and Stocker, M.R. 2016. Relationships of the Indian phytosaur Parasuchus hislopi Lydekker, 1885. Papers in Palaeontology 2: 1–23. Crossref
Kaye, F.T. and Padian, K. 1994. Microvertebrates from the Placerias Quarry: a window on Late Triassic vertebrate diversity in the American Southwest. In: N.C. Fraser and H.D. Sues (eds.), In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods, 171–196. Cambridge University Press, New York.
Kent, D.V. and Tauxe, L. 2005. Corrected Late Triassic latitudes for continents adjacent to the North Atlantic. Science 307: 240–244.
Kligman, B.T. 2015. A new assemblage of small tetrapods from the Upper Triassic Chinle Formation, Arizona. In: A. MacKenzie, E. Maxwell, and J. Miller-Camp (eds.), Society of Vertebrate Paleontology 75th Annual Meeting, Program and Abstracts, 155. Society of Vertebrate Paleontology, Bethesda, MD.
Kligman, B.T., Parker, W.G., and Marsh, A.D. 2017. First record of Saurichthys (Actinopterygii) from the Late Triassic (Chinle Formation: Norian) of western North America. Journal of Vertebrate Paleontology 37: e1367304. Crossref
Kitching, J.W. 1995. Biostratigraphy of the Cynognathus Assemblage Zone. In: B.S. Rubridge (ed.), Biostratigraphy of the Beaufort Group (Karoo Supergroup), Biostratigraphic Series, 140–145. South African Committee for Stratigraphy, Pretoria.
Lucas, S.G. and Hunt, A.P. 1993. Tetrapod biochronology of the Chinle Group. New Mexico Museum of Natural History and Science Bulletin 3: 327–329.
Long, R.A. and Murry, P.A. 1995. Late Triassic (Carnian and Norian) Tetrapods from the Southwestern United States. New Mexico Museum of Natural History and Science Bulletin 4: 1–254.
Long, R.A. and Padian, K. 1986. Vertebrate biostratigraphy of the Late Triassic Chinle Formation, Petrified Forest National Park, Arizona: preliminary results. In: K. Padian, (ed.), The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic–Jurassic Boundary, 161–169. Cambridge University Press, Cambridge.
Long, R.A., Lucas, S.G., Hunt, A.P., and McCrea, R.T. 1989. Charles Camp: collecting Late Triassic vertebrates in the American Southwest during the 1920’s and 1930’s. In: S.G. Lucas and A.P. Hunt (eds.), Dawn of the Age of Dinosaurs in the American Southwest, 65–71. New Mexico Museum of Natural History, Albuquerque.
Lucas, S.G. 1998. Global Triassic tetrapod biostratigraphy and biochronology. Palaeogeography, Palaeoclimatology, Palaeoecology 143: 347–384. Crossref
Lucas, S.G. 2010. The Triassic timescale based on nonmarine tetrapod biostratigraphy and biochronology. In: S.G. Lucas (ed.), The Triassic Timescale, 447–500. The Geological Society of London, London. Crossref
Lucas, S.G., Heckert, A.B., and Hunt, A.P. 1997. Lithostratigraphy and biostratigraphic significance of the Placerias Quarry, east-central Arizona. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 203: 23–46.
Malan, M.E. 1963. The dentitions of the South African rhynchocephalia and their bearing on the origin of the rhynchosaurs. South African Journal of Science 59: 14–220.
Martinez, R.N., Apaldetti, C., Colombi, C.E., Praderio, A., Fernandez, A., Santi Malnis, P., Correa, G.A., Abelin, D., and Alcobar, O. 2013. A new sphenodontian (Lepidosauria: Rhynchocephalia) from the Late Triassic of Argentina and the early origin of herbivore opisthodontians. Proceedings of the Royal Society B: Biological Sciences 1772: 2013–2057. Crossref
Martz, J.W. and Parker, W.G. 2010. Revised lithostratigraphy of the Sonsela Member (Chinle Formation, Upper Triassic) in the southern part of Petrified Forest National Park, Arizona. PLoS One 5(2): e9329. Crossref
Martz, J.W. and Parker, W.G. 2017. Revised formulation of the Late Triassic land vertebrate “faunochrons” of Western North America: recommendations for codifying nascent systems of vertebrate biochronology. In: K.E. Zeigler and W.G. Parker (eds.), Terrestrial Depositional Systems: Deciphering Complexities through Multiple Stratigraphic Methods, 39–113. Elsevier Science, Amsterdam.
Mitchell, J.S., Heckert, A.B., and Sues, H.D. 2010. Grooves to tubes: Evolution of the venom delivery system in a Late Triassic “reptile”. Naturwissenschaften 97: 1117–1121. Crossref
Mueller, B.D. and Parker, W.G. 2006. A new species of Trilophosaurus (Diapsida: Archosauromorpha) from the Sonsela Member (Chinle Formation) of Petrified Forest National Park, Arizona. Museum of Northern Arizona, Bulletin 62: 119–125.
Müller, J. 2004. The relationships among diapsid reptiles and the influence of taxon selection. In: G. Arratia, M.V.H. Watson, and R. Cloutier (eds.), Recent Advances in the Origin and Early Radiation of Vertebrates, 379–408. Dr Friedrich Pfeil, Munich.
Murry, P.A. 1987. New reptiles from the Upper Triassic Chinle Formation of Arizona. Journal of Paleontology 61: 773–786. Crossref
Nesbitt, S.J. and Stocker, M.R. 2008. The vertebrate assemblage of the Late Triassic Canjilon Quarry (northern New Mexico, USA), and the importance of apomorphy-based assemblage comparisons. Journal of Vertebrate Paleontology 28: 1063–1072. Crossref
Nesbitt, S.J., Flynn, J.J., Pritchard, A.C., Parrish, M.J., Ranivoharimanana, L., and Wyss, A.R. 2015. Postcranial osteology of Azendohsaurus madagaskarensis (?Middle to Upper Triassic, Isalo Group, Madagascar) and its systematic position among stem archosaur reptiles. Bulletin of the American Museum of Natural History 398: 1–126. Crossref
Nesbitt, S.J., Irmis, R.B., Parker, W.G., Smith, N.D., Turner, A.H., and Rowe, T. 2009a. Hindlimb osteology and distribution of basal dinosauromorphs from the Late Triassic of North America. Journal of Vertebrate Paleontology 29: 498–916. Crossref
Nesbitt, S.J., Stocker, M.R., Small, B.J., and Downs, A. 2009b. The osteology and relationships of Vancleavea campi (Reptilia: Archosauriformes). Zoological Journal of the Linnean Society 157: 814–864. Crossref
Nordt, L.C., Atchley, S.C., and Dworkin, S.I. 2015. Collapse of the Late Triassic megamonsoon in western equatorial Pangaea, present-day American southwest. Geological Society of America Bulletin 127: 1798–1815. Crossref
Norell, M.A. 1989. Late Cenozoic lizards of the Anza Borrego Desert, California. Natural History Museum of Los Angeles County Contributions in Science 414: 1–31.
Ogg, J.G. 2012. Triassic. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), The Geologic Time Scale 2012, 681–730. Elsevier, Oxford.
Ottone, E.G., Monti, M., Marsicano, C.A., Marcelo, S., Naipauer, M., Armstrong, R., and Mancuso, A.C. 2014. A new Late Triassic age for the Puesto Viejo Group (San Rafael depocenter, Argentina): SHRIMP U-Pb zircon dating and biostratigraphic correlations across southern Gondwana. Journal of South American Earth Sciences 56: 186–199. Crossref
Parker, W.G. and Barton, B.J. 2008. New information on the Upper Triassic archosauriform Vancleavea campi based on new material from the Chinle Formation of Arizona. Palaeontologia Electronica 11: 1–20.
Parker, W.G. and Martz, J.W. 2011. The Late Triassic (Norian) Adamanian–Revueltian tetrapod faunal transition in the Chinle Formation of Petrified Forest National Park, Arizona. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101: 231–260. Crossref
Ramezani, R., Fastovsky, D.E., and Bowring, S.A. 2014. Revised chronostratigraphy of the lower Chinle Formation strata in Arizona and New Mexico (USA): high-precision U-Pb geochronological constraints on the Late Triassic evolution of dinosaurs. American Journal of Science 314: 981−1008. Crossref
Ramezani, R., Hoke, G.D., Fastovsky, D.E., Bowring, S.A., Therrien, F., Dworkin, S.I., Atchley, S.C., and Nordt, L.C. 2011. High-precision U-Pb zircon geochronology of the Late Triassic Chinle Formation, Petrified Forest National Park (Arizona, USA): temporal constraints on the early evolution of dinosaurs. Geological Society of America Bulletin 123: 2142–2159. Crossref
Reichgelt, T., Parker, W.G., Martz, J.W., Conran, J.G., Van Konijnenburg-Van Cittert, J.H., and Kürschner, W.M. 2013. The palynology of the Sonsela member (Late Triassic, Norian) at Petrified Forest National Park, Arizona, USA. Review of Palaeobotany and Palynology 189: 18–28. Crossref
Renesto, S.C., Spielmann, J.A., and Lucas, S.G. 2009. The oldest record of drepanosaurids (Reptilia: Diapsida) from the Late Triassic (Adamanian Placerias Quarry, Arizona, USA) and the stratigraphic range of the Drepanosauridae). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 252: 315–325. Crossref
Romano, C., Kogan, I., Jenks, J., Jerjen, I., and Brinkman, W. 2012. Saurichthys and other fossil fishes from the late Smithian (Early Triassic) of Bear Lake County (Idaho, USA), with a discussion of saurichthyid paleogeography and evolution. Bulletin of Geosciences 87: 543–570. Crossref
Sidor, C.A., Damiani, R.R., and Hammer, W.R. 2008. A new Triassic temnospondyl from Antarctica and a review of Fremouw Formation biostratigraphy. Journal of Vertebrate Paleontology 28: 656–663. Crossref
Sidor, C.A., Steyer, S.J., and Damiani, R.R. 2007. Parotosuchus (Temnospondyli: Mastodontosauridae) from the Triassic of Antarctica. Journal of Vertebrate Paleontology 27: 232–235. Crossref
Spielmann, J.A., Lucas, S.G., Rinehart, L.F., and Heckert, A.B. 2008. The Late Triassic archosauromorph Trilophosaurus. New Mexico Museum of Natural History and Science Bulletin 43: 1–177.
Sues, H.D. 1996. A reptilian tooth with apparent venom canals from the Chinle Group (Upper Triassic) of Arizona. Journal of Vertebrate Paleontology 16: 571–572. Crossref
Sues, H.D. 2003. An unusual new archosauriform from the Upper Triassic Wolfville Formation of Nova Scotia. Canadian Journal of Earth Sciences 40: 635–649. Crossref
Tannenbaum, F.A. 1983. The Microvertebrate Fauna of the Placerias and Downs’ Quarries, Chinle Formation (Upper Triassic) Near St. Johns, Arizona. 114 pp. Master’s Thesis, University of California Berkeley, Berkeley.
Woody, D.T. 2006. Revised stratigraphy of the lower Chinle Formation (Upper Triassic) of Petrified Forest National Park, Arizona. Museum of Northern Arizona Bulletin 62: 17–45.
Zeigler, K.E. and Geissman, J.W. 2011. Magnetostratigraphy of the Upper Triassic Chinle Group of New Mexico: Implications for regional and global correlations among Upper Triassic sequences. Geosphere 7: 802–829. Crossref
Acta Palaeontol. Pol. 63 (1): 117–127, 2018
https://doi.org/10.4202/app.00426.2017