
A new brontothere from the Eocene of South China
ALEXANDER AVERIANOV, IGOR DANILOV, WEN CHEN, and JIANHUA JIN
Averianov, A., Danilov, I., Chen, W., and Jin, J. 2018. A new brontothere from the Eocene of South China. Acta Palaeontologica Polonica 63 (1): 189–196.
The upper Eocene Youganwo Formation in Maoming Province, Guangdong Province, China, produced abundant remains of turtles and crocodiles, but mammalian remains are rare. The previously found mammals from Youganwo Formation include a nimravid carnivore and an amynodontid perissodactyl. Here we report on a new brontotheriid perissodactyl from the Youganwo Formation. Maobrontops paganus gen. et sp. nov. is described based on a maxillary fragment with P4 and M1–2 (SYSU-M-4). Maobrontops paganus gen. et sp. nov. is characterized by the combination of a simple P4 without a hypocone with derived molars having large molar fossae and large anterolingual cingular cusp. A parsimony implied weighting character analysis places Maobrontops gen. nov. in the clade Embolotheriita as a sister taxon to the terminal subclade containing Nasamplus, Protembolotherium, and Embolotherium. Maobrontops paganus gen. et sp. nov. is one of the largest Asian brontotheres. The brontotheriid fauna of South China is endemic and includes at least three valid taxa: Dianotitan from Brontotheriita and Pygmaetitan and Maobrontops gen. nov. from Embolotheriita.
Key words: Mammalia, Perissodactyla, Brontotheriidae, Eocene, China.
Alexander Averianov [dzharakuduk@mail.ru], State Key Laboratory of Biocontrol and Guangdong Provincial Key Laboratory of Plant Resources, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China; Zoological Institute of the Russian Academy of Sciences, Universitetskaya Nab. 1, St Petersburg 199034, Russia; and Institute of Geology and Petroleum Technology, Kazan Federal University, Kremlevskaya Str. 4/5, 420008, Kazan, Russia.
Igor Danilov [igordanilov72@gmil.com], State Key Laboratory of Biocontrol and Guangdong Provincial Key Laboratory of Plant Resources, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China; and Zoological Institute of the Russian Academy of Sciences, Universitetskaya Nab. 1, St Petersburg 199034, Russia.
Wen Chen, Shengli State Farm of Guangdong Province, No.3, Xiupeichang of Shengli State Farm, Nantang Town of Gaozhou, Guangdong 525256, China.
Jianhua Jin [lssjjh@mail.sysu.edu.cn], State Key Laboratory of Biocontrol and Guangdong Provincial Key Laboratory of Plant Resources, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China.
Received 16 October 2017, accepted 16 November 2017, available online 11 January 2018.
Copyright © 2018 A. Averianov et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Brontotheres, or titanotheres, are extinct perissodactyls famous for their peculiar bunolophodont morphology and, in late representatives, large body size and bizarre nasofrontal horns. The phylogenetic position of brontotheres within the Perissodactyla has been debated; a recent analysis places them close to tapirs and chalicotheres (Rose et al. 2014). Brontotheres are common in the Eocene of North America and Eastern Asia (Osborn 1929b; Granger and Gregory 1943; Mihlbachler 2008). In China, most records of brontotheres come from Northern China, mainly from the Nei Mongol Province (Granger and Gregory 1943; Russell and Zhai 1987; Wang et al. 1999; Mihlbachler 2008). In South China, brontothere remains have been reported from the five intermountain basins: Lunan, Lijiang, and Yuezhow basins in Yunnan Province, Shinao Basin in Guizhou Province, and Bose Basin in Guangxi Zhuang Autonomous Region. The brontotheriid taxa identified previously from these basins by Chinese scientists include Dianotitan lunanensis (Chow and Hu, 1959), Protitan major Huang and Zheng, 2004, Protitan cf. robustus Granger and Gregory, 1943, Rhinotitan quadridens Xu and Chiu, 1962, and Brontotheriidae indet. from the middle Eocene Lumeiyi Formation of Lunan Basin (Chow and Hu 1959; Xu and Chiu 1962; Chow et al. 1974; Zheng et al. 1978; Huang and Zheng 2004); Brontotheriidae indet. from the middle Eocene Xiangshan Formation of Lijiang Basin (Zhang et al. 1978); Brontotheriidae indet. from the upper Eocene Caijiachong Formation of Yuezhow Basin (Zhang et al. 1978); Pygmaetitan panxianensis Miao, 1982 from the upper Eocene Shinao Formation of Shinao Basin (Miao 1982); cf. Protitan sp. and Metatelmatherium sp. from the middle Eocene Dongjun Formation of Bose Basin (Ding et al. 1977). According to the recent revision of the brontotheriid species-level taxonomy by Mihlbachler (2008), Dianotitan lunanensis and Pygmaetitan panxiensis are valid taxa, Protitan major and Rhinotitan quadridens are nomina dubia, and the material assigned to Protitan cf. robustus from the Lumeiyi Formation lacks sufficient diagnostic characters for species or genus identification. Identification of brontotheres from the Dongjun Formation is also based on fragmentary and not diagnostic specimens.
Here we report on a brontothere record in another intermountain basin in South China, represented by a maxillary fragment with P4 and M1-2 (SYSU-M-4). It comes from the dark brown oil shales in the upper part of the Youganwo Formation (Fig. 1) of the Maoming Basin in Guangdong Province, which were deposited under lacustrine conditions. This is the southernmost record of the Brontotheriidae in China. The Youganwo Formation has been dated based on magnetostratigraphy and palynomorph assemblages as upper Eocene (Wang et al. 1994; Jin 2008; Li et al. 2016), or middle-upper Eocene (Aleksandrova et al. 2015). The previously known vertebrate assemblage of the Youganwo Formation includes cyprinid fishes, carettochelyid, adocid, trionychid, and geoemydid turtles, tomistomine and alligatorid crocodiles, nimravid carnivore, and amynodontid perissodactyls (Chow and Liu 1955; Liu 1957; Ye 1958, 1963, 1994; Chow and Ye 1962; Wang et al. 2007; Tong et al. 2010; Claude et al. 2012; Danilov et al. 2013, 2017; Skutschas et al. 2014; Chen et al. 2015; Averianov et al. 2016, 2017).
Institutional abbreviations.—IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; SYSU-M, Mammal fossil collection from the Maoming Basin in the School of Life Sciences, Sun Yat-sen University, Guangzhou, China.
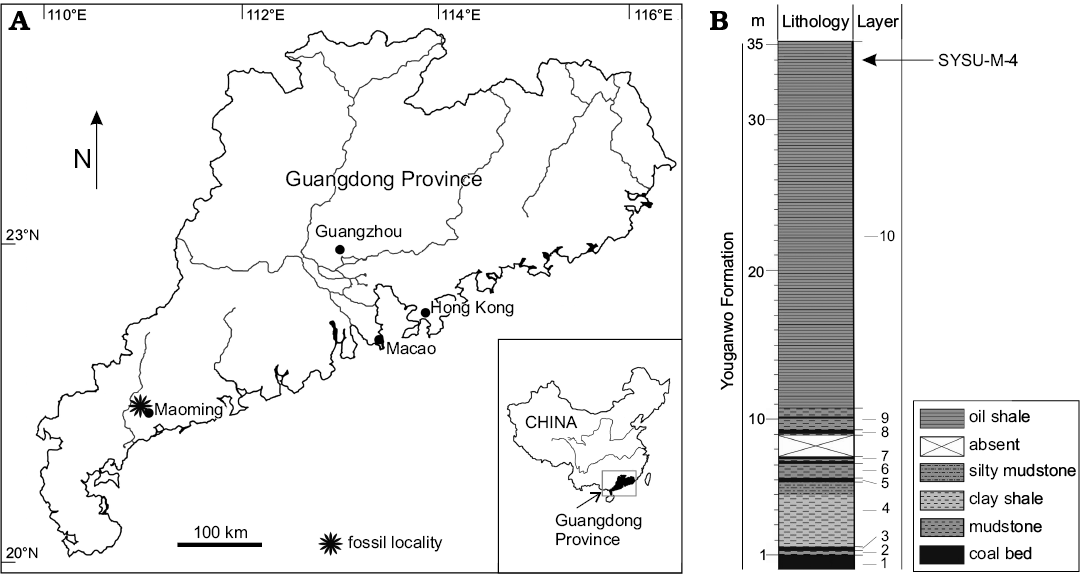
Fig. 1. A. Geographical map of the Maoming Basin, Guangdong Province, China; fossil site where SYSU-M-4 was found is indicated by a star. B. Stratigraphical column of the Youganwo Formation (position of SYSU-M-4 is indicated by an arrow).
Systematic palaeontology
Order Perissodactyla Owen, 1848
Family Brontotheriidae Marsh, 1873
Subfamily Brontotheriinae Marsh, 1873
Tribe Brontotheriini Marsh, 1873
Subtribe Brontotheriina Marsh, 1873
Infratribe Embolotheriita Osborn, 1929a
Genus Maobrontops nov.
Etymology: From Maoming City and Brontops, a currently invalid genus of North American brontotheres, used as a suffix for some Asian brontotheres.
Type species: Maobrontops paganus sp. nov., by monotypy, see below.
Diagnosis.―Same as for the type species.
Maobrontops paganus sp. nov.
Fig. 2.
Etymology: From Latin paganus, rustic; an allusion to the collection of many Maoming fossils by locals.
Type material: Holotype: SYSU-M-4, a left maxillary fragment with P4 and M1–2.
Type locality: The oil shale quarry (21°420 N, 110°530 E) located near Maoming City, Maoming Basin, Guangdong Province, China.
Type horizon: Youganwo Formation, upper Eocene (Li et al. 2016).
Diagnosis.―Referred to the Brontotheriidae based on W-shaped ectoloph and isolated lingual cusps of upper molars. Referred to the Brontotheriinae because of tall ectoloph with functional emphasis placed on shearing on the labial enamel of the ectoloph (i.e., the labial ectoloph enamel thicker than the lingual ectoloph enamel). Differs from all Brontotheriinae except some members of Embolotheriita (Nasamplus progressus, Protembolotherium efremovi, Embolotherium andrewsi, and Embolotherium grangeri) by upper molar anterolingual cingular cusp large. Differs from the above mentioned taxa of Embolotheriita by P4 paracone rib narrow and P4 hypocone lacking.
Description.—The maxilla is fragmentary, poorly preserved, and offers no morphological details concerning the cranial anatomy.
P3: The tooth is mostly destroyed. It is evident that this tooth was slightly narrower transversely than P4.
P4: The tooth is of rectangular outline in occlusal view, transversely wider than long. The ectoloph is almost straight, without a mesostyle and with a rather strong labial ridge at the paracone, weaker ridge at the parastyle, and a slight labial convexity at the metacone. The wear facet of the ectoloph is widest at the paracone (13.4 mm) and the metacone (9.3 mm), and narrowest between the paracone and the metacone and labial to the central fossa (7.6 mm). The exposed enamel is thicker along the labial side (~3.8 mm). On the lingual side, the exposed enamel is thicker at the paracone and metacone apices (maximum 2.5 mm) and decreases in thickness outwards. There are strong vertical ridges on the labial side of the paracone and metacone that bracket anteriorly and posteriorly a very deep central fossa.
The lingual heel is slightly shorter anteroposteriorly than the ectoloph. It is dominated by the cone-like protocone, which is as high as the labial cusps and lacks the preprotocrista and postprotocrisa. The protocone apex is slightly worn, with a small circle of exposed dentine that is flush with the surrounding exposed enamel. The protocone is surrounded by the anterior, lingual, and posterior cingula, from which it is separated by a distinct rounded groove. The anterior and posterior cingula are straight. The continuous lingual cingulum is rounded, convex lingually. There are cusp-like eminences at the anterior and posterior cingula.
M1: It has a quadrangular outline in occlusal view, distinctly longer and wider than P4. The W-shaped ectoloph is heavily worn, with the wear facet widest at the paracone (24 mm) and metacone (18 mm). There are strong labial ridges at the parastyle and mesostyle. The labial sides of the paracone and metacone between the labial ridges are slightly convex. The exposed enamel is thick on the labial side (~3.3 mm), and about twice as thin lingually around the paracone and metacone (~1.5 mm). There is a large central fossa of crescentic shape, which is bordered lingually by a prominent and significantly worn ridge. This ridge extends between the anterolingual cingular cusp and vertical ridge on the lingual side of the metacone.
The lingual heel is only a little shorter anteroposteriorly than the ectoloph. The protocone and hypocone are distinctly lower than the labial cusps. Both cusps have circular wear facets at the apices with exposed dentine and enamel. On the protocone wear facet the enamel rim is raised above the adjacent dentine surface, while on the hypocone wear facet the small exposed area of dentine is flush with the enamel. This suggests that the wear on the protocone started earlier than on the hypocone. The protocone is cone like, but with a flattened to slightly concave lingual side. The hypocone is more ridge-like and resembles the enlarged posterior cingulum of P4. The anterior cingulum is similar to that of P4 but larger. It has a large cusp-like eminence that is mostly worn down (anterolingual cingular cusp). There is no lingual cingulum.
M2: It is distinctly larger than M1 and is of trapezium outline in occlusal view. The anterior side is about 1.5 times wider than the posterior side, which results in an oblique position of the labial side relative to the lingual side. The W-shaped ectoloph is about twice less worn than that of the M1. The exposed enamel is ~3.0 mm thick along the labial side and ~2.5 mm thick around the paracone and metacone apices on the lingual side. There are strong labial ridges at the parastyle and mesostyle and a much weaker ridge at the metastyle. The labial side of the paracone and metacone is slightly convex between the ridges. There is a large central fossa of triangular shape that is bordered posteriorly by a sharp vertical ridge along the metacone lingual side and lingually by a distinct ridge.
The lingual cusps are much lower compared to the labial cusps and correspond in shape to those of the M1. With a flattened lingual side, the cone-like protocone is relatively larger than that of P4. Because of this, the size difference between the protocone and hypocone is more pronounced on M2 compared to M1. The ridge-like hypocone is obliquely extended along the postero-lingual corner of the tooth. There is only slight wear on the protocone and hypocone apices. There is a large cusp-like eminence along the anterior cingulum (anterolingual cingular cusp). This eminence is connected with the base of the hypocone by a ridge extending between the central fossa and the vertical ridge on the lingual side of the metacone. This ridge is unworn and bears several prominent irregular denticles. There is no lingual cingulum.
The enamel is mostly smooth, but there are areas of pitted or wrinkled enamel on the labial side of the P4 and molars, and on the lingual side of P4.
For dental measurements see Table 1.
Stratigraphic and geographic range.—Eocene of South China.
Table 1. Measurements (in mm) of upper dentition of SYSU-M-4, the holotype of Maobrontops paganus gen. et sp. nov.
|
P4 |
M1 |
M2 |
|||
|
Length |
Width |
Length |
Width |
Length |
Width |
|
42.1 |
49.0 |
56.1 |
57.4 |
73.3 |
69.8 |
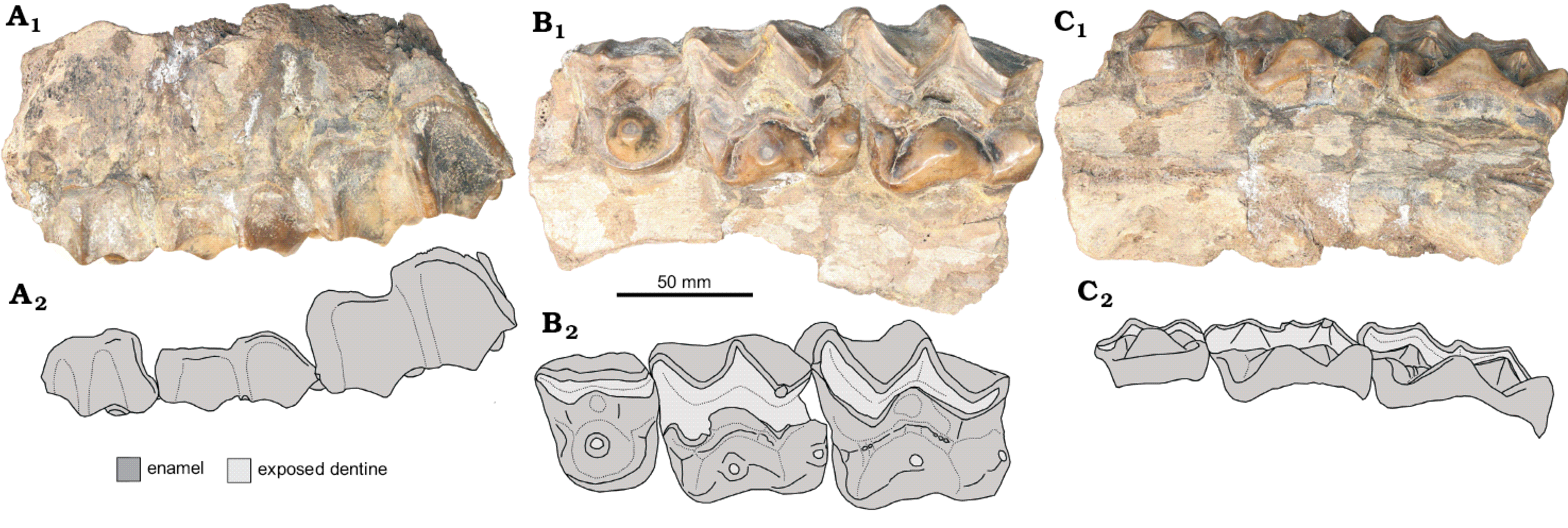
Fig. 2. Embolotheriit brontothere Maobrontops paganus gen. et sp. nov., SYSU-M-4 (holotype), from Maoming Basin, Guangdong Province, China; Youganwo Formation, upper Eocene. Left maxillary fragment with P4 and M1–2, in labial (A), occlusal (B), and lingual (C) views. Photographs (A1–C1) and interpretive drawings of tooth crowns (A2–C2).
Phylogenetic analysis
Maobrontops paganus gen. et sp. nov. can be coded using 19 of the 91 characters (20.9%) appearing in the data matrix of Mihlbachler and Samuels (2016). The character codings are as follows:
Character 50 (2): Labial upper premolar ribs: paracone rib narrow, metacone rib very weak. This character state is found in brontotheres which are more terminal than Palaeosyops. Microtitan, Brontotheriita. The terminal Embolotheriita (Nasamplus, Protembolotherium, and Embolotherium) have a more derived condition, all labial ribs very weak on upper premolars.
Character 53 (1): Upper premolar lingual morphology: lingual side of crown is somewhat wider, protocone often associated with lingual crest. In Maobrontops gen. nov., there is no lingual crest on P4. Actually, the morphology of P4 lingual side is intermediate between the states 0 and 1 of this character. The protocone is cone-like, but the lingual side is not narrow, although it is somewhat narrower than in most other derived brontotheres. Most of the derived brontotheres have a variously developed hypocone on P4. Aktautitan hippopotamopus Mihlbachler, Lucas, Emry, and Bayshashov, 2004 from the middle Eocene of Kazakhstan is the phylogenetically closest taxon to Maobrontops gen. nov. iaxon of brontotheres which lack hypocone on P4 (Mihlbachler et al. 2004). Although the hypocone is lacking on P4 of Aktautitan, the lingual side of P4 is almost as wide as the labial side in that taxon. In Maobrontops gen. nov., the lingual side of P4 is distinctly narrower than the labial side.
Character 54 (1): Upper molar parastyle high. Found in all brontotheres and outgroup taxa.
Character 55 (1): Upper molar parastyle mainly labial to paracone. Distribution as in the previous character.
Character 56 (1): Upper molar anterior cingulum does not form parastyle shelf. Found in brontotheres more terminal than Palaeosyops.
Character 57 (1): Upper molar metastyle angled labially. Found in all brontotheres and outgroup taxa.
Character 58 (1): Upper molar ectoloph hypsodont. Found in brontotheres more terminal than Bunobrontops.
Character 59 (2): Labial side of upper molar paracone and metacone lingually angled and convex. Found in brontotheres more terminal than Palaeosyops.
Character 60 (1): Molar ribs weak. Found in brontotheres more terminal than Bunobrontops.
Character 61 (1): Upper molar mesostyle present. Found in all brontotheres and outgroup taxa.
Character 62 (1): Lingual margins of molar paracone and metacone wedged. Found in brontotheres more terminal than Bunobrontops.
Character 63 (3): Thickness of the lingual enamel of the upper molar ectoloph completely thinner than labial ectoloph enamel. Found in brontotheres more terminal than Mesatirhinus.
Character 64 (2): Labiolingual width of upper molar ectoloph: wide. Found in brontotheres more terminal than Eotitanops.
Character 65 (1): Central molar fossa present. Found in brontotheres more terminal than Mesatirhinus, but reversed in Microtitan, Metatelmatherium, Wickia, Telmatherium, and Qufutitan.
Character 66 (4): Upper molar protoloph and paraconules absent. Found in brontotheres more terminal than Dolichorhinus, but reversed in Wickia, Telmatherium, and Nanotitanops.
Character 67 (3): M1–2 metaloph absent. Found in brontotheres more terminal than Mesatirhinus. The second state included in this character by Mihlbachler (2008) (vestigial metaloph-like ridge) refers to the ridge extended between the anterolingual cingular cusp and the hypocone. This ridge borders the molar fossa lingually. This ridge is not connected with the hypocone apex and more likely is a neomorph character rather than the remnant of the metaloph. Mihlbachler (2008) noted the presence of this ridge in Megacerops kuwagatarhinus Mader and Alexander, 1995 (Mader and Alexander 1995: fig. 5) and Pygmaetitan, and considered this a reversal from the metaloph absence (character state 3). Based on the illustrations in Mihlbachler (2008), this ridge has a wider distribution, being present in Dolichorhinus hyognathus (Osborn, 1889), Fossendorhinus diploconus (Osborn, 1895), Metarhinus fluviatilis Osborn, 1908, Metarhinus abbotti (Riggs, 1912), Aktautitan hippopotamopus, Metatitan primus Granger and Gregory, 1943, Metatitan relictus Granger and Gregory, 1943, Protembolotherium efremovi Yanovskaya, 1954, Embolotherium andrewsi Osborn, 1929a, Embolotherium grangeri Osborn, 1929a, and Dianotitan lunanensis. Distribution of this character has no clear phylogenetic pattern, but it is notably present in a clade including Maobrontops gen. nov., Protembolotherium, and Embolotherium.
Character 68 (2): Upper molar anterolingual cingular cusp large. Present in Maobrontops gen. nov., Protembolotherium, and Embolotherium.
Character 91 (1): M1–2 parastyle and mesostyle extend an equal distance labially.
Character 92 (1): Upper molar protocone distinctly larger than the hypocone, at least 50% on M2.
The character-taxon matrix with scores for Maobrontops paganus gen. et sp. nov. (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app63-Averianov_etal_SOM.pdf) was analyzed using the traditional search option of TNT version 1.1 (Goloboff et al. 2008) with 1000 replications and 10 trees saved per replication, followed by the tree bisection and reconnection (TBR) swapping algorithm of the trees in memory; 10 000 trees were set up in memory. The tree statistics and distribution of characters were obtained using WinClada 1.00.08 (Nixon 2002).
Separate analyses were conducted under equal weights and implied weighting (Table 2). The phylogenetic position of Maobrontops paganus gen. et sp. nov. was unresolved in the equal weights and the implied weighting analyses with the constant K ranging from 1 to 9. Only the implied weighting analysis with the constant K = 10 produced a strict consensus tree with the resolved position of Maobrontops paganus gen. et sp. nov. A fragment of this tree is shown in Fig. 3. For the branch support we used the Group present/Contradict (GC) frequency values calculated on symmetric resampling with 1000 replicates using TNT software (P = 33).
Table 2. Statistics for equal weight and implied weighting analyses. MPT, most parsimonius tree; * overflow.
|
Analysis |
Number of MPT |
Tree length |
|
|
Equal weight |
10000* |
350 |
|
|
Implied weighting |
K = 1 |
189 |
53.81 |
|
K = 2 |
378 |
41.34 |
|
|
K = 3 |
1890 |
33.85 |
|
|
K = 4 |
1890 |
28.79 |
|
|
K = 5 |
1485 |
26.06 |
|
|
K = 6 |
1485 |
22.20 |
|
|
K = 7 |
1485 |
19.94 |
|
|
K = 8 |
1485 |
18.11 |
|
|
K = 9 |
1485 |
16.59 |
|
|
K = 10 |
99 |
15.31 |
|
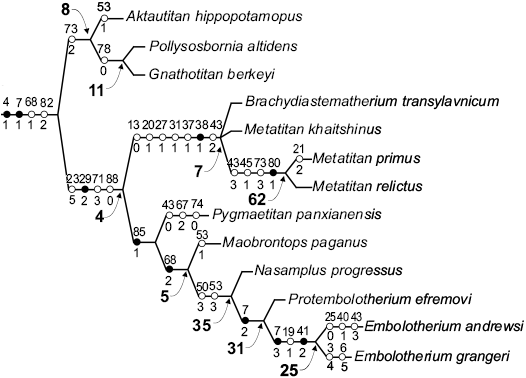
Fig. 3. Part of the strict consensus tree of 99 most parsimonious trees produced by TNT implied weighting character analysis (K = 10), showing the interrelationships within the infratribe Embolotheriita Osborn, 1929a sensu Mihlbachler (2008). Only unambiguous characters are shown (black circles are nonhomoplasies and white circles are homoplasies). The numbers next to the circles are characters (above) and states (below). The numbers at nodes in bold face are GC values for branch support.
Principal component analysis
We performed a principal component analysis (PCA) to analyze variation in size dimensions of P4 and M1–2 in brontotheres based on measurements provided in Mihlbachler (2008). These teeth were chosen because only P4 and M1–2 are represented in the specimen SYSU-M-4, described in the present paper. The teeth in the middle part of the tooth row are usually less variable in size compared with the mesial and distal cheek teeth: anterior premolars or M3. Only specimens with known values for the length (L) and width (W) of all three teeth (P4, M1, and M2) were used in this study. Our dataset includes 30 specimens belonging to ten species of the Embolotheriita. Pygmaetitan panxianensis was excluded from this sample because it is far removed from the morphospace of other Embolotheriita owing to its very small size. The principal component analysis was performed using the software PAST version 3.01 (Hammer et al. 2001). Principal component 1 (PC 1) represents 73% of the variance in the dataset (Table 3). The length and width of M2 have the greater loadings on PC 1 (Table 3).
On the plot PC 1 versus PC 2 (Fig. 4) the holotype of Maobrontops paganus (SYSU-M-4) is within the morphospace of Metatitan relictus. Positionally, the holotype is located most closely to two specimens belonging to that species as well as to the holotype of “Protitan” major (IVPP V13802). The morphospaces of three species of Metatitan and of Rhinotitan andrewsi Osborn, 1925 are strongly overlapped.
Table 3. Principal component loadings of teeth variables in analyzed sample of brontotheres. PC, Principal Component.
|
Variable |
PC 1 |
PC 2 |
|
|
P4 |
length |
0.1274 |
0.04419 |
|
width |
0.026053 |
0.2259 |
|
|
M1 |
length |
0.20221 |
-0.41584 |
|
width |
0.41112 |
0.50834 |
|
|
M2 |
length |
0.61 |
-0.63253 |
|
width |
0.57783 |
0.33999 |
|
|
Eigenvalue |
165.009 |
32.4286 |
|
|
Variance |
72.96% |
14.34% |
|
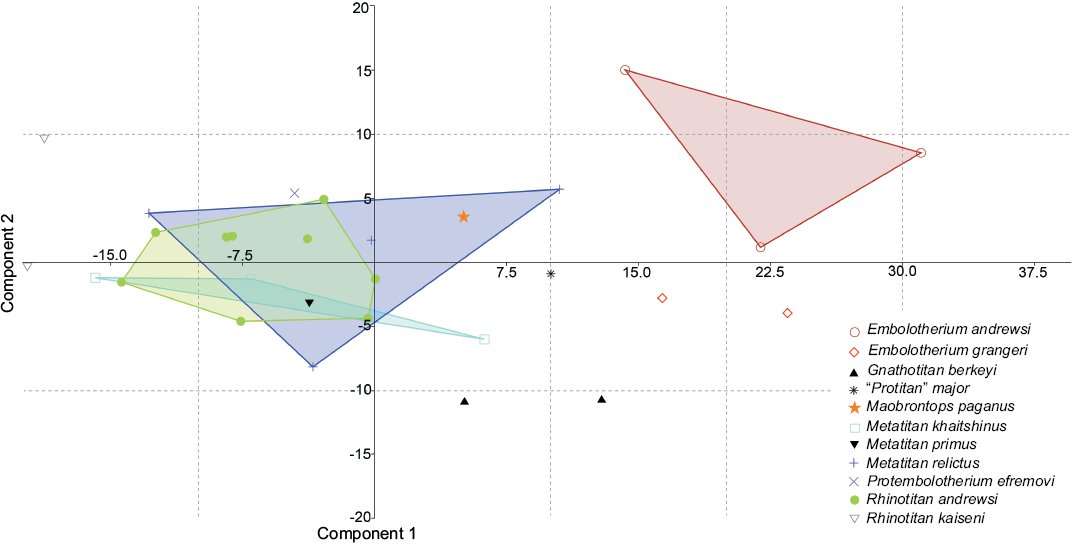
Fig. 4. Principal component analysis of 32 specimens of ten species of Embolotheriita based on measurements of P4 and M1–2 (see SOM 2).
Discussion and conclusions
The phylogenetic relationships within the Embolotheriita in our resulting cladogram (Fig. 3) are very similar to those found in the strict consensus of heuristic search performed with ordered multistate characters by Mihlbachler (2008: fig. 194). Except for the inclusion of the new taxon, Maobrontops paganus gen. et sp. nov., the only difference between the two cladograms is the position of Pygmaetitan panxianensis. In the strict consensus provided by Mihlbachler (2008: fig. 194) Pygmaetitan forms a polytomy with the clades Brachiadiastematherium + Metatitan and Nasamplus + (Protembolotherium + Embolotherium). In our consensus tree (Fig. 3) the position of Pygmaetitan is fully resolved: it is the sister taxon to the clade Maobrontops gen. nov. + (Nasamplus + (Protembolotherium + Embolotherium)). However, the resampling analysis places Pygmaetitan outside Embolotheriita as the sister taxon to Sthenodectes with a GC branch support value of 12. In our analysis, Pygmaetitan is united with the derived Embolotheriita by one unambiguous synapomorphy, lower molar occlusal basins deep (character 85[1]). This unique character is present also in Embolotherium, but unknown for Nasamplus, Protembolotherium, and Maobrontops gen. nov. Pygmaetitan comes from the Shinao Basin of Guizhou Province, which is relatively close to the Maoming site. In spite of its close geographic and phylogenetic position relative to Maobrontops gen. nov., Pygmaetitan is markedly different from the latter having a much smaller size (Fig. 5), P4 hypocone, and relatively short M1 compared to other molars. Another brontothere from South China, Dianotitan, is nested within the sister clade to the Embolotheriita, the Brontotheriita, in our analysis and in Mihlbachler (2008: fig. 194). It is also different from Maobrontops gen. nov. by having a smaller size (Fig. 5) and P4 hypocone.
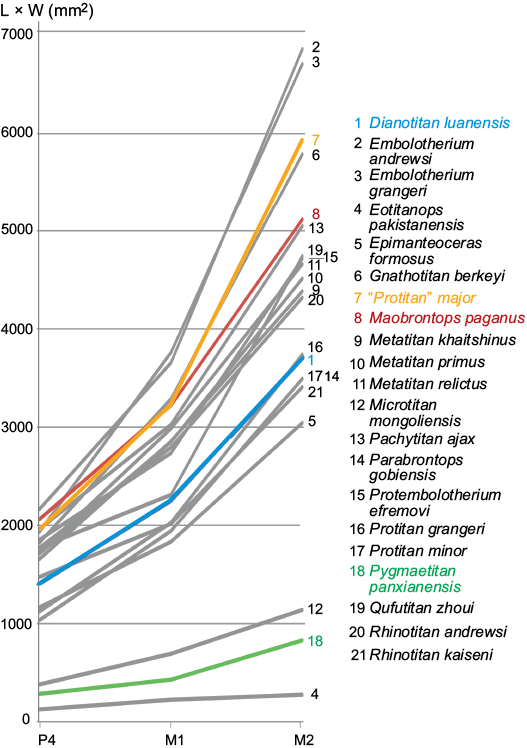
Fig. 5. The size of middle cheek teeth, P4 and M1–2 (L × W), in Asian brontotheres (based on data present in SOM 2). Taxa found in South China are Dianotitan lunanensis (1), “Protitan” major (7), Maobrontops paganus gen. et sp. nov. (8), and Pygmaetitan panxianensis (18).
There is a single unambiguous synapomorphy that unites Maobrontops gen. nov. with derived Embolotheriita: the upper molar anterolingual cingular cusp is large (character 68[2]; Fig. 3). This character is present also in Protembolotherium and Embolotherium.
Maobrontops paganus gen. et sp. nov. is similar to “Protitan” major from the middle Eocene Lumeiyi Formation of Lunan Basin in Southern China (Huang and Zheng 2004; Mihlbachler 2008) in its large size and lack of hypocone on P4. However, Maobrontops paganus gen. et sp. nov. is clearly different from that taxon in having a much more pronounced antero-lingual cusp and deep furrows between the lingual cusps of the upper molars. The latter character is found also in Embolotherium and may be structurally related to the deep lower molar basins found in that taxon and related taxa. This character was not included in the data matrix provided by Mihlbachler (2008) or Mihlbachler and Samuels (2016). However, its presence in Maobrontops gen. nov. and derived Embolotheriita supports the inclusion of the new taxon in that clade.
Maobrontops paganus gen. et sp. nov. is one of the largest known brontotheres from Asia, being inferior in size only to Embolotherium (Fig. 5). Janis (1990) published equations relating body mass to M2 area (length × width) in hyracoids and perissodactyls. Based on those formulas, the body mass of the holotype of Maobrontops paganus gen. et sp. nov. was approximately 792 kg. However, this increase in size was likely evolved in southern Chinese brontotheres independently of those living in northern China, as Protembolotherium is smaller than Maobrontops gen. nov.
In spite of a far less abundant record of brontotheres in South China compared to North China and Mongolia, the brontothere fauna of that geographic region is quite diverse. It includes at least three taxa (Dianotitan, Pygmaetitan, and Maobrontops gen. nov.), and possible one more taxon (“Rhinotitan quadridens”). Dianotitan (Brontotheriita) was likely derived from a North American immigrant (Mihlbachler 2008), while Pygmaetitan and Maobrontops gen. nov. were parts of the endemic radiation (Embolotheriita). Embolotheriita is mostly an Asian clade with a single North American representative (Pollyosbornia). Brachydiastematherium comes from eastern Europe, which was biogeographically part of Asia in the Eocene (Mihlbachler 2008). The Embolotheriita might have originated in Southern China, and from there, more derived members of the group dispersed to the northern parts of eastern Asia.
Acknowledgements
We thank Matthew Mihlbachler (New York Institute of Technology, Old Westbury, USA) and Bryn Mader (Queensborough Community College, Bayside, USA) for reading the manuscript and providing useful comments. This study was supported by the National Natural Science Foundation of China (Grant No. 41210001), the Key Project of Sun Yat-sen University for inviting foreign teachers. AA was supported by Zoological Institute, Russian Academy of Sciences (project АААА-А17-117022810195-3) and Russian Foundation for Basic Research (project 16-04-00294). The work is performed according to the Russian Government Program of Competitive Growth of Kazan Federal University.
References
Aleksandrova, G.N., Kodrul, T.T., and Jin, J. 2015. Palynological and paleobotanical investigations of Paleogene sections in the Maoming basin, South China. Stratigraphy and Geological Correlation 23: 300–325. Crossref
Averianov, A.O., Danilov, I.G., Jin, J., and Wang, Y. 2017. A new amynodontid from the Eocene of South China and phylogeny of Amynodontidae (Perissodactyla: Rhinocerotoidea). Journal of Systematic Palaeontology 15: 927–945. Crossref
Averianov, A.O., Obraztsova, E.M., Danilov, I.G., Skutschas, P.P., and Jin, J. 2016. First nimravid skull from Asia. Scientific Reports 6: 25812. Crossref
Chen, G.J., Chang, M.-M., and Liu, H.Z. 2015. Revision of Cyprinus maomingensis Liu, 1957 and the first discovery of Procypris-like cyprinid (Teleostei, Pisces) from the Late Eocene of South China. Science China Earth Sciences 58: 1123–1132. Crossref
Chow, M.-C. and Hu, C.C. 1959. A new species of Parabrontops from the Oligocene of Lunan, Yunnan. Acta Palaeontologica Sinica 7: 85–88.
Chow, M.-C. and Liu, C.-L. 1955. A new anosterine turtle from Maoming, Kwangtung. Acta Palaeontologica Sinica 3: 275–282.
Chow, M.-C. and Ye, H.-K. 1962. A new emydid from Eocene of Maoming, Kwangtung. Vertebrata PalAsiatica 6: 225–229.
Chow, M.-C., Chang, Y., and Ting, S. 1974. Some Early Tertiary Perrisodactyla from Lunan Basin, E. Yunnan. Vertebrata PalAsiatica 12: 262–273.
Claude, J., Zhang, J.-Y., Li, J.-J., Mo, J.-Y., Kuang, X.-W., and Tong, H. 2012. Geoemydid turtles from the late Eocene Maoming Basin, Southern China. Bulletin de la Société géologique de France 183: 641–651. Crossref
Danilov, I.G., Obraztsova, E.M., Chen, W., and Jin, J. 2017. The cranial morphology of Anosteira maomingensis (Testudines, Pan-Carettochelys) and the evolution of pan-carettochelyid turtles. Journal of Vertebrate Paleontology: e1335735. Crossref
Danilov, I.G., Syromyatnikova, E.V., Skutschas, P.P., Kodrul, T.T., and Jin, J. 2013. The first ‘true’ Adocus (Testudines, Adocidae) from the Paleogene of Asia. Journal of Vertebrate Paleontology 33: 1071–1080. Crossref
Ding, S., Zheng, Y., Zhang, Y., and Tong, Y. 1977. The age and characteristic of the Liuniu and the Dongjun faunas, Bose Basin of Guangxi. Vertebrata PalAsiatica 15: 35–45.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT (Tree Analysis Using New Technology) (BETA). Published by the authors, Tucamán.
Granger, W. and Gregory, W.K. 1943. A revision of the Mongolian titanotheres. Bulletin of the American Museum of Natural History 80: 349–389.
Hammer, Ø., Harper, D.A.T., and Ryan, P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9.
Huang, X. and Zheng, J. 2004. Brontotheres (Perissodactyla, Mammalia) from the middle Eocene of Lunan, Yunnan. Vertebrata PalAsiatica 42: 334–339.
Janis, C.M. 1990. Correlation of cranial and dental variables with body size in ungualtes and macropodids. In: J. Damuth and B.J. MacFadden (eds.), Body Size in Mammalian Paleobiology: Estimation and Biological Implications, 255–300. Cambridge University Press, Cambridge.
Jin, J. 2008. On the age of the Youganwo Formation in the Maoming Basin, Guangdong Province. Journal of Stratigraphy 32: 47–50.
Li, Y.X., Jiao, W.J., Liu, Z.H., Jin, J., Wang, D.H., He, Y.X., and Quan, C. 2016. Terrestrial responses of low-latitude Asia to the Eocene–Oligocene climate transition revealed by integrated chronostratigraphy. Climate of the Past 12: 255-272. Crossref
Liu, H.-T. 1957. A new fossil cyprinid fish from Maoming, Kwangtung. Vertebrata PalAsiatica 1: 151–153.
Mader, B.J. and Alexander, J.P. 1995. Megacerops kuwagatarhinus n. sp., an unusual brontothere (Mammalia, Perissodactyla) with distally forked horns. Journal of Paleontology 69: 581–587. Crossref
Marsh, O.C. 1873. Notice of new Tertiary mammals. American Journal of Science, Series 3 5: 485–488. Crossref
Miao, D. 1982. Early Tertiary fossil mammals from the Shinao basin, Panxian County, Guizhou Province. Acta Palaeontologica Sinica 21: 527–536.
Mihlbachler, M.C. 2008. Species taxonomy, phylogeny and biogeography of the Brontotheriidae (Mammalia, Perissodactyla). Bulletin of the American Museum of Natural History 311: 1–475. Crossref
Mihlbachler, M.C. and Samuels, J.X. 2016. A small-bodied species of Brontotheriidae from the middle Eocene Nut Beds of the Clarno Formation, John Day Basin, Oregon. Journal of Paleontology 90: 1233–1244. Crossref
Mihlbachler, M.C., Lucas, S.G., Emry, R.J., and Bayshashov, B.U. 2004. A new brontothere (Brontotheriidae, Perissodactyla, Mammalia) from the Eocene of the Ily Basin of Kazakstan and a phylogeny of Asian “horned” brontotheres. American Museum Novitates 3439: 1–43. Crossref
Nixon, K.C. 2002. WinClada version 1.00.08. Avaiable at http://www.cladistics.com.
Osborn, H.F. 1889. The Mammalia of the Uinta Formation. Part III. The Perissodactyla. Transactions of the American Philosophical Society 16: 505–530.
Osborn, H.F. 1895. Fossil mammals of the Uinta Basin. Expedition of 1894. Bulletin of the American Museum of Natural History 7: 71–105.
Osborn, H.F. 1908. New or little known titanotheres from the Eocene and Oligocene. Bulletin of the American Museum of Natural History 24: 599–617.
Osborn, H.F. 1925. Upper Eocene and lower Oligocene titanotheres of Mongolia. American Museum Novitates 202: 1–12.
Osborn, H.F. 1929a. Embolotherium, gen. nov., of the Ulan Gochu, Mongolia. American Museum Novitates 353: 1–20.
Osborn, H.F. 1929b. Titanotheres of ancient Wyoming, Dakota, and Nebraska. United States Geological Survey Monographs 55: 1–894.
Owen, R. 1848. Description of teeth and portions of jaws of two extinct anthracotherioid quadrupeds (Hyopotamus vectianus and Hyop. bovinus) discovered by the Marchioness of Hastings in the Eocene deposits on the NW coast of the Isle of Wight: with an attempt to develop Cuvier’s idea of the classification of pachyderms by the number of their toes. Quarterly Journal of the Geological Society of London 4: 103–141. Crossref
Riggs, E.S. 1912. New or little known titanotheres from the lower Uintah formations with notes on the stratigraphy and distribution of fossils. Field Museum of Natural History Publications Geological Series 4: 17–41.
Rose, K.D., Holbrook, L.T., Rana, R.S., Kumar, K., Jones, K.E., Ahrens, H.E., Missiaen, P., Sahni, A., and Smith, T. 2014. Early Eocene fossils suggest that the mammalian order Perissodactyla originated in India. Nature Communications 5: 5570. Crossref
Russell, D.E. and Zhai, R. 1987. The Paleogene of Asia: Mammals and stratigraphy. Memoires du Museum National d’Histoire Naturelle, Series C 52: 1–488.
Skutschas, P.P., Danilov, I.G., Kodrul, T.T., and Jin, J. 2014. The first discovery of an alligatorid (Crocodylia, Alligatoroidea, Alligatoridae) in the Eocene of China. Journal of Vertebrate Paleontology 32: 471–476. Crossref
Tong, H., Zhang, J.-Y., and Li, J.-J. 2010. Anosteira maomingensis (Testudines: Carettochelyidae) from the Late Eocene of Maoming, Guangdong, southern China: new material and re-description. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 256: 279–290. Crossref
Wang, J., Li, H., Zhu, Z., Seguin, M.K., Yang, J., and Guomei, Z. 1994. Magnetostratigraphy of Tertiary rocks from Maoming Basin, Guangdong Province, China. Chinese Journal of Geochemistry 13: 165–175. Crossref
Wang, Y., Guo, J., and Wang, J. 1999. A review of Chinese brontotheres. In: Y. Wang and T. Deng (eds.), Proceedings of the Seventh Annual Meeting of the Chinese Society of Vertebrate Paleontology, 139–147. China Ocean Press, Beijing.
Wang, Y.-Y., Zhang, Z.-H., and Jin, J. 2007. Discovery of Eocene fossil mammal from Maoming Basin, Guangdong. Acta Scientiarum Naturalium Universitatis Sunyatseni 46: 131–133.
Xu, Y.-X. and Chiu, C.-S. 1962. Early Tertiary mammalian fossils from Lunan, Yunnan. Vertebrata PalAsiatica 6: 313–332.
Yanovskaya, N.M. [Âanovskaâ, N.M.] 1954. A new genus of Embolotheriinae from the Paleogene in Mongolia [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 55: 5–43.
Ye, H.-K. 1958. A new crocodile from Maoming, Kwangtung. Vertebrata PalAsiatica 2: 237–242.
Ye, H.-K. 1963. Fossil turtles of China. Paleontologica Sinica, New Series C 18: 1–112.
Ye, H.-K. 1994. Fossil and Recent Turtles of China. 112 pp. Science Press, Beijing.
Zhang, Y.P., You, Y.-Z., Ji, H.X., and Ding, S. 1978. Cenozoic Stratigraphy of Yunnan. Professional Papers of Stratigraphy and Palaeontology 7: 1–21.
Zheng, J.J., Tang, Y.J., Zhai, R., Ding, S., and Huang, X.S. 1978. Early Tertiary strata of Lunan Basin, Yunnan. Professional Papers of Stratigraphy and Palaeontology 7: 22–29.
Acta Palaeontol. Pol. 63 (1): 189–196, 2018
https://doi.org/10.4202/app.00431.2017