
New serpulid polychaetes from the Permian of western Sicily
ROSSANA SANFILIPPO, ANTONIETTA ROSSO, AGATINO REITANO, ALFIO VIOLA, and GIANNI INSACCO
Sanfilippo, R., Rosso, A., Reitano, A., Viola, A., and Insacco, G. 2018. New serpulid polychaetes from the Permian of western Sicily. Acta Palaeontologica Polonica 63 (3): 579–584.
Two new tubeworms, “Serpula” calannai sp. nov. and “Serpula” prisca sp. nov. are described from the Permian limestone of the Sosio Valley, western Sicily. Both species possess large tubes with long free anterior portions circular in cross-section. All morphological characters, such as tube shape and ornamentation, as well as inner structure of the wall, even if barely visible due to diagenesis, are still preserved. These two new species increase the previously known diversity of the Permian serpulid community, which flourished at the shelf edge of the western sector of the Palaeotethys and disappeared thereafter. The likely Wordian age of these serpulids, and their presumed absence in younger rocks up to the Middle–Late Triassic, suggests reduction, or even extinction, of these reef-related serpulid taxa during the end-Guadalupian biotic crisis, before the end-Permian extinction.
Key words: Polychaeta, Serpulidae, tube morphology, reef community, extinction events, Palaeozoic, Permian, Italy.
Rossana Sanfilippo [sanfiros@unict.it], Antonietta Rosso [rosso@unict.it], and Alfio Viola [a.viola@unict.it], Department of Biological, Geological and Environmental Sciences, University of Catania, Corso Italia 57, 95129 Catania, Italy.
Agatino Reitano [tinohawk@yahoo.it] and Gianni Insacco [g.insacco@comune.comiso.rg.it], Museo di Storia Naturale di Comiso, Via degli Studi 9, 97013 Comiso, Italy.
Received 2 December 2017, accepted 16 May 2018, available online 8 August 2018.
Copyright © 2018 R. Sanfilippo et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
True serpulids are commonly found in the fossil record and until recently they were considered widely distributed throughout the Mesozoic and Cenozoic, with about 46 genera and 350 species (ten Hove and Kupriyanova 2009; Kočí and Jäger 2015, among others). Small and simple forms dating back to the Middle Triassic, ca. 244 Ma, are the first records of unequivocal serpulids, although a number of problematic tubeworms of presumed serpulid affinity were reported from the Permian on (see Ippolitov et al. 2014). However, unexpected large-sized species with advanced characters, such as complex shape and ornamentation, were recently discovered in a limestone bed dated back to the middle-to-upper Permian, ca. 268–251 Ma (Sanfilippo et al. 2016, 2017b). These findings antedated serpulids’ first appearance even several millions years earlier and indicated that a surprisingly diversified serpulid community existed before the end-Permian mass extinction.
Two further new species unequivocally belonging to the serpulids, namely “Serpula” calannai sp. nov. and “Serpula” prisca sp. nov., were found in materials deposited in paleontological museums in Sicily. They originate from the same Permian rocks, specifically the “Pietra di Salomone” Limestone from the Sosio Valley, western Sicily. These new specimens consist of the long unattached anterior parts of large tubes, circular in cross section.
The aims of this study are to: (i) describe these new species; (ii) determine affinities and differences between the new species and the serpulid taxa already known from the same area and time span; (iii) discuss the role played by the serpulid community of the Sosio Limestone within reef palaeoenvironments of the Permian Mediterranean Palaeotethys.
Institutional abbreviations.—PMC, Museo di Paleontologia, University of Catania, Italy; MCSNC, Museo Civico di Storia Naturale di Comiso, Ragusa, Italy.
Material and methods
The two specimens examined in the present paper originate from the “Pietra di Salomone” Limestone (Fig. 1), the largest fossiliferous megablock (200 m long, 100 m wide and 30 m high) among those cropping out near Palazzo Adriano in the Sicani Mountains, western Sicily (for geological details see Sanfilippo et al. 2017b, and references therein). It consists of a coarse calcareous breccia of which the precise age is still debated, being referred to the Wordian by Flügel et al. (1991), but considered, at least partly, Capitanian, Wuchiapingian or even Changhsingian by Jenny-Deshusses et al. (2000).
The fossiliferous content of these Permian megablocks is famous world-wide for its rich invertebrate fauna, first discovered and described by Gemmellaro (1887–1889). In total, more than 300 species are known from the “Pietra di Salomone” alone, belonging to several taxonomic groups such as sponges, corals, bryozoans, brachiopods, as well as to the recently discovered serpulids (see Flügel et al. 1991; Sanfilippo et al. 2016, 2017b, and references therein). Like previously described serpulids, the two new species were included in the white crinoid-rich limestone from the upper part of the “Pietra di Salomone”. Serpulid tubes were mainly found attached to sponges, while the species described herein are broken off parts of unattached anterior ends found embedded in the carbonate rock.
The fragments belonging to “Serpula” calannai sp. nov. were separated mechanically from the slightly cemented calcilutite, and glued together to reconstruct a longer, although broken, tube part. The tube was accurately cleaned to examine the whole external surface and broken longitudinally at one end to investigate its microstructure. The tube belonging to “S.” prisca sp. nov. was cleaned, but not extracted from the enclosing rock, to avoid the separation of tube portions glued together in an old restoration.
Low magnification photos were acquired with a Zeiss Discovery V8A stereomicroscope equipped with an Axiovision acquisition system in order to document general tube morphology. Both specimens and the detached fragment were also examined uncoated under a LMU Tescan Vega Scanning Electron Microscope in Low Vacuum modality to investigate their micromorphology and microstructure.
“Serpula” calannai sp. nov. is stored in the Collection Mario Calanna of the MCSNC, while “S.” prisca sp. nov. is stored in the collections of the PMC.
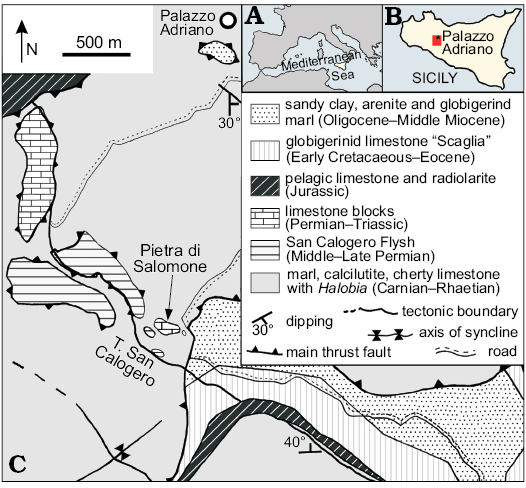
Fig. 1. A, B. Location of the study area in the Mediterranean and Sicily. C. Geological map of the Palazzo Adriano area (Sicani Mountains) showing the location of the “Pietra di Salomone” megablock (modified from Di Stefano and Gullo 1997).
Systematic palaeontology
Class Polychaeta Grube, 1850
Family Serpulidae Rafinesque, 1815
Genus Serpula Linnaeus, 1758
Type species: Serpula vermicularis Linnaeus, 1767; Recent of England.
“Serpula” calannai sp. nov.
Figs. 2, 3A–F.
Etymology: In honor of the paleontology amateur collector the late Mario Calanna (1945–2014), for his contribution to natural history collections, especially in malacology.
Holotype: MSNC 4549-33, one large tube portion (44 mm long and 7.8 mm wide), representing part of an unattached anterior erect portion, broken at both ends and partially filled by lithified sediment.
Type locality: Sosio Valley, western Sicily, Italy.
Type horizon: “Pietra di Salomone” Limestone, Wordian to upper Permian (see Material and methods).
Material.—Type material only.
Diagnosis.—Tube quite large, with a well-developed raised anterior end, circular in cross-section, with outer surface feebly rough. Ornamentations solely consisting of eristomes well developed and slightly flaring. Tube wall thick and layered.
Description.—The single available specimen is a large tube portion of the anterior end, 44 mm long and 7.8 mm wide. It is slightly bent and circular in cross-section, not increasing in diameter. Outer surface is feebly lumpy to smooth, with only irregular and inconspicuous growth lines, which are about 10 µm thick. Peristomes are well developed, differently pronounced and slightly flaring, with smooth, circular edges, locally crenulated. Peristomes are protruding up to 300 µm and irregularly spaced, at intervals of 2–6 mm. Tube wall is about 600 µm thick, with a seemingly layered internal structure, as barely visible along some sections, which cross areas that are not completely altered by recrystallization. Posterior (attached) tube portion is lacking.
Remarks.—The combination of tube characters of “Serpula” calannai sp. nov. are considered sufficient to describe it as a new species. Particularly, this species clearly differs from the other five serpulids recently mentioned from the same area (Sanfilippo et al. 2016, 2017b) by the relevant size of the tube, which is significantly larger and thicker than those of these other species, and by the presence of evident peristomes. Obvious differences also exist with species that have erect anterior ends, but show different co-occurring characters: Palaeotubus sosiensis Sanfilippo, Reitano, Insacco, and Rosso, 2016, has a tube ornamented by a number of longitudinal keels; Serpula distefanoi Sanfilippo, Rosso, Reitano, and Insacco, 2017b, has a tube end with evident growth lines and a rougher outer surface; Serpulidae gen. et. sp. indet. sensu Sanfilippo et al. 2017b, has a very smooth tube, with regular growth lines and only weak annulations but no peristomes. “S.” calannai sp. nov. only roughly recalls Propomatoceros permianus Sanfilippo, Rosso, Reitano, and Insacco, 2017b, of which exclusively its posterior part is known. However, it is not unlikely that, even if this latter species developed an anterior adult part circular in cross section and ornamented with peristomes (as it is in some other serpulid species), it would have retained typical transverse ribs that are lacking in “S.” calannai sp. nov.
Contrary to the specific attribution, the generic allocation is difficult, owing to the absence of preserved characters of taxonomical importance (e.g., Weedon 1994, Vinn and Mutvei 2009, Vinn et al. 2008) in the available material, because neither the morphology of the attached tube portion nor the pristine structure of the tube wall are fully known. Like some species of the genus Vermiliopsis Saint-Joseph, 1894, “Serpula” calannai sp. nov. possesses distinct peristomes, but its tube does not increase in diameter and lacks the longitudinal keels typical of that genus (Jäger 2005, 2011; ten Hove and Kupriyanova 2009). In contrast, particularly large-sized tubes with circular anterior ends and evident peristomes can be present in Serpula (Sanfilippo and Mòllica 2000; ten Hove and Kuprianova 2009; Sanfilippo et al. 2013). Consequently, we provisionally assign the new species to “Serpula”, a genus name often misused to allocate species of the family Serpulidae whose characters are insufficiently known and/or do not allow an unequivocal generic attribution (see Jäger 1993). If further specimens will be found, including attached tube parts with less altered wall structure, it would be possible to support the present attribution more consistently or disregard it.
“Serpula” calannai sp. nov. resembles the present-day S. cavernicola Fassari and Mòllica, 1991, more than the other species in the genus, for its very long erect part with a relevant diameter and comparable, and even major, wall thickness, as well as for the occurrence of peristomes. However, S. cavernicola has longitudinal keels. This species also shows a special micromorphology of the outer surface (Sanfilippo and Mòllica 2000: figs. 1, 2), which seems to be different from that of S. calannai. In the new species growth lines are obvious and perfectly preserved (Fig. 3E) evidencing that the outer surface is not affected by any process of destructive taphonomic processes (e.g., abrasion, dissolution).
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 2. New serpulid polychaetes from the Wordian to upper Permian, “Pietra di Salomone” Limestone, Sicily, Italy. A. “Serpula” calannai sp. nov. (holotype MSNC 4549-33); general views (A1–A3), cross section of tube, white limestone partly filling lumen (A4). B. “Serpula” prisca sp. nov. (holotype PMC.S5.15.09.2017); general view of anterior end, partially embedded in rock. Arrow indicates growth directions for all tubes.

Fig. 3. Serpulid polychaete “Serpula” calannai sp. nov. (holotype MSNC 4549-33) from the Wordian to upper Permian, “Pietra di Salomone” Limestone, Sicily, Italy. A. Tube fragment before assembling. Growth direction, inferred from the slightly flaring peristomes, marked by an arrow. B. Detail of a peristome and growth lines. C. Longitudinal fracture of tube displaying wall section and inner surface covered by secondary crystals. D. Oblique view of tube showing prominence of ornamentations. E. Detail of C showing layered structure of tube wall, only locally preserved. F. Circular cross section of tube, with relatively thick wall locally showing its likely original layering. All SEM images.
“Serpula” prisca sp. nov.
Figs. 2B, 4A–D.
Etymology: From Latin priscus, ancient; referring to both the age of the outcrop where the species was found, and to the primordial position of the species in the serpulid family.
Holotype: PMC.S5.15.09.2017, a fragment, broken at both ends, of a slightly curved free anterior tube portion. The fragment is 43 mm long and 3.9 mm wide, and appears recrystallized.
Type locality: Sosio Valley, western Sicily, Italy.
Type horizon: “Pietra di Salomone” Limestone, Wordian to upper Permian (see Material and methods).
Material.—Type material only.
Diagnosis.—Tube relatively large, with a well-developed raised anterior end. Tube circular in cross-section, outer surface smooth with very thin growth lines and nearly imperceptible longitudinal striations.
Description.—The single specimen consists of a slightly curved tube fragment corresponding to a long anterior tube portion. It is circular in cross-section with a constant diameter of 3.9 mm. Lumen 3.3 mm wide. Tube wall 0.3 mm thick. Outer surface smooth, with very thin and smooth growth lines spaced at intervals of usually 100 to 300 µm, but locally more closely spaced and evident. Even thinner and nearly imperceptible longitudinal striations, locally intersect the growth lines. Posterior encrusting tube portion unknown. Original microstructure of the tube wall not preserved.
Remarks.—Even incomplete, the measurements of the fragment most probably indicate that it belonged to an adult specimen. The available tube portion provided distinctive characters justifying the erection of a new species. “Serpula” prisca sp. nov. has a unique, extremely smooth tube, that differs from all other serpulids from the Sosio Limestone, only superficially resembling Serpulidae sp. nov. indet. Sanfilippo et al. 2017b. Nevertheless, the latter species is smaller-sized, shows more evident growth lines than S. prisca sp. nov. and lacks longitudinal striations.
Like for the previous species, generic allocation is difficult, mostly because of the absence of the posterior attached part. The tube roughly resembles that of species belonging to Protula Risso, 1826, which may have comparable long raised anterior tube portions, not (noticeably) increasing in diameter, and lacking keels and peristomes. The delicate micromorphology with imperceptible longitudinal striations is somewhat similar to that of some scaphopod gadilinidae shells (Scarabino 1995), but differs for disposition of growth lines. Also, the smooth outer morphology of the new species is comparable to that observed in Protula but in this species the outer surface is rougher (RS, unpublished data). In absence of a more fitting genus, we provisionally assign this species to “Serpula”, pending further support from more complete tubes with attached portions. Large secondary calcite crystals occur in the tube wall, whose original structure has been obliterated by diagenesis. The tube of S. prisca sp. nov. does not show any evidence of a worst preservation in respect to that of the other species (S. calannai sp. nov.) described herein: both species come from the same outcrop, thus presumably underwent similar process of fossilization which led to similar degree of diagenetic preservation (heavy recrystallization, with large secondary calcite crystals). In spite of this, tubes of the two species possess different micromorphology. The tube of S. calannai sp. nov. displays a lumpy surface, with irregular growth lines, about 10 micron thick (Fig. 3E). In addition, several well-developed peristomes occur in the tube of S. calannai sp. nov. They are irregularly spaced and surprisingly very frequent along the tube (at intervals of 2–6 mm). Conversely, tube of S. prisca sp. nov. is perfectly smooth and lacks even slightly developed peristome. Also, both tubes represent anterior erect parts that largely differ in size: S. calannai sp. nov. is twice the size of S. prisca sp. nov. (outer diameter 7.8 mm and 3.9 mm) and has a thick wall, twice that of the other.
If these tubes belonged to the same species, the dimensional range (variability of the diameter of 4 mm) would be too large, and the hypothesis of an intraspecific variability would be little supportable. Likewise, the difference in thickness of the tube wall (0.3 mm) is likely too much to fit into a variability. It should also be considered that usually the raised parts of the tubes maintain substantially a constant diameter and a constant wall thickness. It is likely that tubes with such obvious differences belong to two different species rather than represent variants of the same species.
Stratigraphic and geographic range.—Type locality and horizon only.

Fig. 4. Serpulid polychaete “Serpula” prisca sp. nov. (holotype PMC.S5.15.09.2017) from the Wordian to upper Permian, “Pietra di Salomone” Limestone, Sicily, Italy. A. Central portion of the tube glued together, with only feeble growth lines on its outer surface. B. Tube wall with large secondary calcite crystals pointing to complete recrystallization. C. Close-up of outer surface showing very thin and smooth growth lines. D. Magnification of C, with closely spaced, relatively prominent growth lines, crossed by nearly imperceptible longitudinal striations. All SEM images.
Conclusions
Since their origin till nowadays serpulids represent a relevant component of coral/algal/metazoan bioconstructions in shallow-shelf habitats (among others Nishi 1996; Di Geronimo et al. 2002; Mallela 2007), as well as in bathyal (Sanfilippo 2009a; Rosso et al. 2010; Kupriyanova et al. 2011; D’Onghia et al. 2015) and cave environments (Rosso et al. 2013, Sanfilippo et al. 2013, 2015, 2017a; Guido et al. 2017). Increase in abundance and diversity of serpulids throughout geological time appears to be related to the presence of reef ecosystems, where skeletons of frame-builders constituted heterogeneous substrata available for colonization. This is the case with some Jurassic sponge build-ups (among others Pisera 1991) and Jurassic–Cretaceous coral reefs, comprising abundant and diversified serpulid associations (among others Parsch 1956; Jäger 1993, 2011), and recently acquired data (Sanfilippo et al. 2016, 2017b, present paper) suggest that this scenario may also explain high diversity of serpulids in the Permian. At that time eight serpulid species belonging to five nominal genera, Palaeotubus, Filograna, “Serpula”, Propomatoceros, and Pyrgopolon, thrived in the Palaeotethys metazoan bioherms, already exhibiting different shapes and ornamentations.
As a whole, the stout and well-calcified serpulids from the Sosio Valley, flourishing in metazoan reef-like buildups dominated by sponges, crinoids and brachiopods, may represent an adaptive response to an exposed shallow-shelf environment. Furthermore, the frame of reef-builders probably offered differentiated microenvironments including sheltered parts, where minute and brittle forms like Filograna, or species like “S.” calannai sp. nov. and “S.” prisca sp. nov., having tubes with long anterior ends raising from the substrate, were adapted to live. Analogously to living species of Protula (see Sanfilippo 2009b: 2021–2022), in the new species the presence of an anterior part raising from the substrate for a considerable length also suggests a life habit in relatively sheltered environments of the shallow-water reef, less exposed to hydrodynamic action.
This surprisingly diversified Permian serpulid community was associated with heavily mineralized demosponges and calcisponges able to produce carbonate frameworks that reached their maximum extent and high diversity during the middle Permian, in the Guadalupian (Sremac et al. 2016). However, these habitats were strongly affected by the end-Guadalupian (pre-Lopingian) biotic crisis, presumed to be linked to global warming, ocean acidification and anoxia events. This crisis caused a steady decrease in biodiversity, especially for some groups (brachiopods, molluscs) and the demise of sponge-metazoan reefs (Clapham et al. 2009; Bond et al. 2010; Sremac et al. 2016). A concomitant decrease in diversity and a virtual disappearance paralleling the fate of other taxa could be hypothesized also for serpulids, which formed a part of the diversified reef-dwelling community. Indeed, serpulids so far have not been recorded from younger Permian and Lower Triassic rocks, up to the simple-shaped Filograna specimens, found in sheltered shallow shelf settings of the very late Middle Triassic (Flügel et al. 1984; Cirilli et al. 1999; Stiller 2000; among others).
If the Wordian or Capitanian age for the Sosio megablocks is confirmed (see above), it seems that the serpulid palaeocommunity might have indeed suffered a sharp reduction to almost extinction during the end-Guadalupian biotic crisis, before the end-Permian extinction. However, data on late Permian–Early Triassic serpulids are still fragmentary and further research is needed to fill this gap.
Acknowledgements
The authors thank the late Emiliano Calanna (Catania, Italy), who donated his uncle’s paleontological collections to the Museo di Storia Naturale di Comiso. The reviewers Harry ten Hove (Naturalis Biodiversity Center, The Nederlands), Manfred Jäger (Rosenfeld, Germany) and Alexei Ippolitov (Geological Institute of Russian Academy of Sciences, Moscow, Russia) greatly improved the paper with their invaluable comments and suggestions. This paper was financially supported by University of Catania grants to RS (Catania Palaeontological Research Group: contribution no. 437).
References
Bond, D.P.G., Hilton, J., Wignall, P.B., Ali J.R., Stevens, L.G., Sun Y., and Lai, X. 2010. The Middle Permian (Capitanian) mass extinction on land and in the oceans. Earth-Science Reviews 102: 100–116. Crossref
Cirilli, S., Iannace, A., Jadoul, F., and Zamparelli, V. 1999. Microbial-serpulid build-ups in the Norian–Rhaetian of the Western Mediterranean area: ecological response of shelf margin communities to stressed environments. Terra Nova 11: 195–202. Crossref
Clapham, M.E., Shen, S., and Bottjer D.J. 2009. The double mass extinction revisited: reassessing the severity, selectivity, and causes of the end-Guadalupian biotic crisis (Late Permian). Paleobiology 35: 32–50. Crossref
Di Geronimo, I., Di Geronimo, R., Rosso, A., and Sanfilippo, R. 2002. Structural and taphonomic analysis of a columnar corallinae build-up from a SE Sicily shelf bottom. Géobios, Mémoire Spécial 24: 86–95. Crossref
Di Stefano, P. and Gullo, M. 1997. Permian deposits of Sicily. A review. Geodiversitas 19: 193–202.
D’Onghia, G., Capezzuto, F., Cardone, F., Carlucci, R., Carluccio, A., Chimienti, G., Corriero, G., Longo, C., Maiorano, P., Mastrototaro, F., Panetta, P., Rosso, A., Sanfilippo, R., Sion, L., and Tursi, A. 2015. Macro- and megafauna recorded in the submarine Bari Canyon (southern Adriatic, Mediterranean Sea) using different tools. Mediterranean Marine Science 16: 180–196. Crossref
Flügel, E., Di Stefano, P., and Senowbari-Daryan, B. 1991. Microfacies and depositional structure of allochthonous carbonate base-of-slope deposits: The Late Permian Pietra di Salomone megablock, Sosio Valley (western Sicily). Facies 25: 147–186. Crossref
Flügel, E., Flügel-Kahler, E., Martin, J.M., and Martin-Algarra, A. 1984. Middle Triassic Reefs from Southern Spain. Facies 11: 173–218. Crossref
Gemmellaro, G.G. 1887–1889. La fauna dei calcari con Fusulina della valle del fiume Sosio nella Provincia di Palermo. Cephalopoda, Ammonoidea, Nautiloidea, Gastropoda. Giornale di Scienze Naturali ed Economiche di Palermo 19: 1–106; 20: 1–26, 97–182.
Guido, A., Jimenez, C., Achilleos, K., Rosso, A., Sanfilippo, R., Hadjioannou, L., Petrou, A., Russo, F., and Mastandrea, A. 2017. Cryptic serpulid-microbialite bioconstructions in the Kakoskali submarine cave (Ciprus, Eastern Mediterranean). Facies 63 (3) [published online, 10.1007/s10347-017-0502-3]. Crossref
Hove, H.A. ten and Kupriyanova, E.K. 2009. Taxonomy of Serpulidae: the state of affairs. Zootaxa 2036: 1–126.
Ippolitov, A.P., Vinn, O., Kupriyanova, E.K., and Jäger, M. 2014. Written in stone: history of serpulid polychaetes through time. Memoirs of Museum Victoria 71: 123–159. Crossref
Jäger, M. 1993. Danian Serpulidae and Spirorbidae from NE Belgium and SE Netherlands: K/T boundary extinction, survival, and origination patterns. Contributions to Tertiary and Quaternary Geology 29: 73–137.
Jäger, M. 2005. Serpulidae und Spirorbidae (Polychaeta sedentaria) aus Campan und Maastricht von Norddeutschland, den Niederlanden, Belgien und angrenzenden Gebieten. Geologisches Jahrbuch A 157: 121–249.
Jäger, M. 2011. Sabellidae, Serpulidae and Spirorbinae (Polychaeta sedentaria) from the Barremian (Lower Cretaceous) of the Serre de Bleyton (Drôme, SE France). Annalen des Naturhistorischen Museums in Wien, Serie A 113: 675–733.
Jenny-Deshusses, C., Marrini, R., and Zaninerri, L. 2000. Découverte du foraminifère Colaniella Likharev dans le Permien supérieur de la vallée du Sosio (Sicile). Comtes Rendus de l’Académie des Sciences 330: 799–804.
Kočí, T. and Jäger, M. 2015. Sabellid and serpulid worms (Polychaeta, Canalipalpata, Sabellida, Sabellidae, Serpulidae) from the rocky coast facies (Late Cenomanian) at Předboj near Prague. Sborník Národního muzea v Praze, řada B, Přírodní vědy. Acta Musei Nationalis Pragae, Series B, Historia naturalis 71: 31–50.
Kupriyanova, E.K, Bailey-Brock, J.H., and Nishi, E. 2011. New records of Serpulidae (Annelida, Polychaeta) collected by R/V “Vityaz” from bathyal and abyssal depths of the Pacific Ocean. Zootaxa 2871: 43–60.
Mallela, J. 2007. Coral reef encruster communities and carbonate production in cryptic and exposed coral reef habitats along a gradient of terrestrial disturbance. Coral Reefs 26: 775–785. Crossref
Nishi, E. 1996. Serpulid Polychaetes Associated with Living and Dead Corals at Okinawa Island, Southwest Japan. Publications Seto Marine Biological Laboratory 37: 305–318. Crossref
Parsch, K.O.A. 1956. Die Serpuliden-Fauna des südwestdeutschen Jura. Palaeontographica A 107: 211–240.
Pisera, A. 1991. Upper Jurassic Sponge Megafacies in Spain: Preliminary Report. In: J. Reitner and H. Keupp (eds.), Fossil and Recent Sponges, 486–497. Springer, Berlin.
Rosso, A., Sanfilippo, R., Taddei Ruggieri, E., and Di Martino, E. 2013. Faunas and ecological groups of Serpuloidea, Bryozoa and Brachiopoda from submarine caves in Sicily (Mediterranean Sea). Bollettino della Società Paleontologica Italiana 52: 167–176.
Rosso, A., Vertino, A., Di Geronimo, I., Sanfilippo, R., Sciuto, F., Di Geronimo, R., Violanti, D., Corselli, C. Taviani, M., Mastrototaro, F., and Tursi, A. 2010. Hard and soft-bottom thanatofacies from the Santa Maria di Leuca deep-water coral province, Mediterranean. Deep Sea Research II. Topical Studies in Oceanography 57: 360–379. Crossref
Sanfilippo, R. 2009a. New species of Hyalopomatus Marenzeller, 1878 (Annelida, Polychaeta, Serpulidae) from Recent Mediterranean deep-water coral mounds and comments on some congeners. Zoosystema 31: 147–161. Crossref
Sanfilippo, R. 2009b. Systematics and life habit in Serpula israelitica Amoureux, 1976 (Polychaeta Serpulidae) from the Mediterranean with remarks on other soft-bottom serpulids. Journal of Natural History 43: 2009–2025. Crossref
Sanfilippo, R. and Mòllica, E. 2000. Serpula cavernicola Fassari & Mòllica, 1991 (Annelida Polychaeta): diagnostic features of the tubes and new Mediterranean records. Marine Life 10: 27–32.
Sanfilippo, R., Reitano, A., Insacco, G., and Rosso, A. 2016. A new tubeworm of possible serpulid affinity from the Permian of Sicily. Acta Paleontologica Polonica 61: 621–626.
Sanfilippo, R., Rosso, A., Guido, A., and Gerovasileiou, V. 2017a. Serpulid communities from two marine caves in the Aegean Sea, Eastern Mediterranean. Journal of the Marine Biological Association of the United Kingdom 97: 1059–1068. Crossref
Sanfilippo, R., Rosso, A., Guido, A., Mastandrea, A., Russo, F., Ryding, R., and Taddei Ruggero, E. 2015. Metazoan/microbial biostalactites from modern submarine caves in the Mediterranean Sea. Marine Ecology 36: 1277–1293. Crossref
Sanfilippo, R., Rosso, A., Reitano, A., and Insacco, G. 2017b. First record of sabellid and serpulid polychaetes from the Permian of Sicily. Acta Paleontologica Polonica 62: 25–38. Crossref
Sanfilippo, R., Vertino, A., Rosso, A., Beuck, L., Freiwald, A., and Taviani, M. 2013. Serpula aggregates and their role in deep-sea coral communities in the southern Adriatic Sea. Facies 59: 663–677. Crossref
Scarabino, V. 1995. Scaphopoda of the tropical Pacific and Indian Oceans, with descriptions of 3 new genera and 42 new species. In: P. Bouchet (ed.), Résultats des Campagnes MUSORSTOM, Vol. 14. Mémoires du Muséum national d’Histoire naturelle, 167: 189–379.
Sremac, J., Jurkovšek, B., Aljinović, D., and Kolar-Jurkovšek, T. 2016. Equatorial Palaeotethys as the last sanctuary for late Permian metazoan reef-builders: New evidence from the Bellerophon Formation of Slovenia. Palaeogeography, Palaeoclimatology, Palaeoecology 454: 91–100. Crossref
Stiller, F. 2000. Polychaeta (Annelida) from the Upper Anisian (Middle Triassic) of Qingyan, south-western China. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 217: 245–266. Crossref
Vinn, O. and Mutvei, H. 2009. Calcareous tubeworms of the Phanerozoic. Estonian Journal of Earth Sciences 58: 286–296. Crossref
Vinn, O., Hove, H.A. ten, and Mutvei, H. 2008. On the tube ultrastructure and origin of calcification in sabellids (Annelida Polychaeta). Palaeontology 51: 295–301. Crossref
Weedon, M.J. 1994. Tube microstructure of recent and Jurassic serpulid polychaetes and the question of the Palaeozoic “spirorbids”. Acta Palaeontologica Polonica 39: 1–15.
Acta Palaeontol. Pol. 63 (3): 579–584, 2018
https://doi.org/10.4202/app.00448.2017