
Arthropod trace fossils from Eocene cold climate continental strata of King George Island, West Antarctica
ALFRED UCHMAN, ANDRZEJ GAŹDZICKI, and BŁAŻEJ BŁAŻEJOWSKI
Uchman, A., Gaździcki, A., and Błażejowski, B. 2018. Arthropod trace fossils from Eocene cold climate continental strata of King George Island, West Antarctica. Acta Palaeontologica Polonica 63 (2): 383–396.
Siltstone and sandstone beds of the Mount Wawel Formation (Eocene) contain trace fossils interpreted as insect resting traces and arthropod trackways, the latest determined as Glaciichnium australis isp. nov. and cf. Pterichnus isp. Glaciichnium is included in a new ichnofamily Protichnidae, which embraces invertebrate trackways composed of straight central trail(s) and lateral tracks. The same deposits contain fragments of plant stems in growth position, delicate fern-like plant twigs and leaves of Nothofagus. Their deposition took place in very shallow but flowing water, probably in a marginal part of a lake, perhaps in a delta. The presence of mudcracks proves incidental exposure of the sediment. The trace fossils were produced by arthropods, especially insects and/or isopods, between episodes of deposition and were influenced by the water flow and subtle changes in substrate consistency. This resulted in several morphological variants of the traces. Glaciichnium australis is similar to those produced by some caddisflies (Trichoptera) in shallow puddles in the Tatra Mountains of Poland. The arthropod-dominated trace fossil assemblage is similar to the Glaciichnium ichnocoenosis, which is known from some Pleistocene lacustrine varve sediments of Europe. This fits well with the beginning of climatic cooling in Antarctica during the late Eocene. This also shows the recurrence of some ichnological features on both ends of the globe in similar palaeoenvironmental conditions and supports basics of the ichnofacies concept.
Key words: Arthropoda, trace fossils, taphonomy, Antarctica, King George Island, Eocene.
Alfred Uchman [alfred.uchman@uj.edu.pl], Institute of Geological Sciences, Jagiellonian University, Gronostajowa 3a, 30-387 Kraków, Poland.
Andrzej Gaździcki [gazdzick@twarda.pan.pl] and Błażej Błażejowski [bblazej@twarda.pan.pl], Institute of Paleobiology, Polish Academy of Sciences, Twarda 51/55, 00-818 Warszawa, Poland.
Received 9 February 2018, accepted 19 April 2018, available online 22 May 2018.
Copyright © 2018 A. Uchman et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Southern Hemisphere Cenozoic glaciation began in the Antarctic at the end of the Eocene and led to the first appearance of ice sheets at the Eocene–Oligocene boundary, ca. 34 Ma (Birkenmajer et al. 2005; Davies et al. 2012). This climatic change significantly influenced continental biota. One of the most important records of the changes occurs in continental clastic-volcanoclastic sediments of the Eocene Mount Wawel Formation (Point Hennequin Group) on King George Island, South Shetlands (Fig. 1), in which leaves of the Nothofagus–Podocarpaceae assemblage have been found (Birkenmajer 1981, 2003; Birkenmajer and Zastawniak 1989; Dutra 2001, 2004; Hunt 2001; Cantrill and Poole 2012).
A contribution to the known invertebrate life in sedimentary environments of the Mount Wawel Formation comes from arthropod trace fossils preserved in several morphological variants, which have been found in the “Dragon Glacier plant fossil beds”. They are similar to some trace fossils from Pleistocene varved clay beds in the Northern Hemisphere. The aim of this paper is to describe and interpret these trace fossils, one of which appears to be a new ichnospecies. Its interpretation was aided by observations of Recent traces in the Tatra Mountains (Central Western Carpathians, Poland).
Institutional abbreviations.—GSE, British Geological Survey, Edinburgh, UK; ZPAL, Institute of Paleobiology, Polish Academy of Sciences, Warszawa, Poland.
Geological setting
The Eocene sedimentary facies occur in the dominantly volcanogenic succession of King George Island. They have been recognized in the Mount Wawel Formation (Point Hennequin Group) of Admiralty Bay (Figs. 1, 2), which is composed of volcaniclastic yellow, green, and grey tuffaceous sandstone and tuff beds, which show mud cracks and ripple marks (Fig. 3). A middle Eocene to late Oligocene age for the Point Hennequin Group has been proposed by Smellie et al. (1984), Birkenmajer (1989, 2001, 2003), Hunt (2001) and Hunt and Poole (2003). New data on the Mount Wawel Formation points to its latest middle–late Eocene age (Francis et al. 2008, 2009; Nawrocki et al. 2011; Mozer 2013; Pańczyk 2013).
The trace fossils discovered in loose, ex-situ slabs of fine-grained sandstone of the Dragon Glacier plant beds (informal unit) are associated with distinct facies on the 300-m high exposure, greatly affected by faulting ice covering and reworking (Hunt 2001). According to Zastawniak et al. (1985), the origin of the ex-situ “Dragon Glacier plant fossil beds” was the same as the in situ occurrences of the “Mount Wawel plant fossil beds” from the top of the Mount Wawel Formation. The sandstone beds contain sedimentary structures with wave-formed ripple marks (Fontes and Lindner Dutra 2010; Fig. 3A), elongate plant remains associated with mud cracks (Fig. 3B), delicate fern-like plant twigs (Fig. 3C), and well-preserved Nothofagus leaves (Fig. 3D), and oblique, longitudinally striated open tubes, which are probably lower plant (reed-like?) remains in growth position (Fig. 4). They occur irregularly or in patches. Extrapolated density fluctuates around 500 tubes/m2. In some bedding planes, the tops of tubes are surrounded by crescentic marks indicating a consistent direction of water flow.
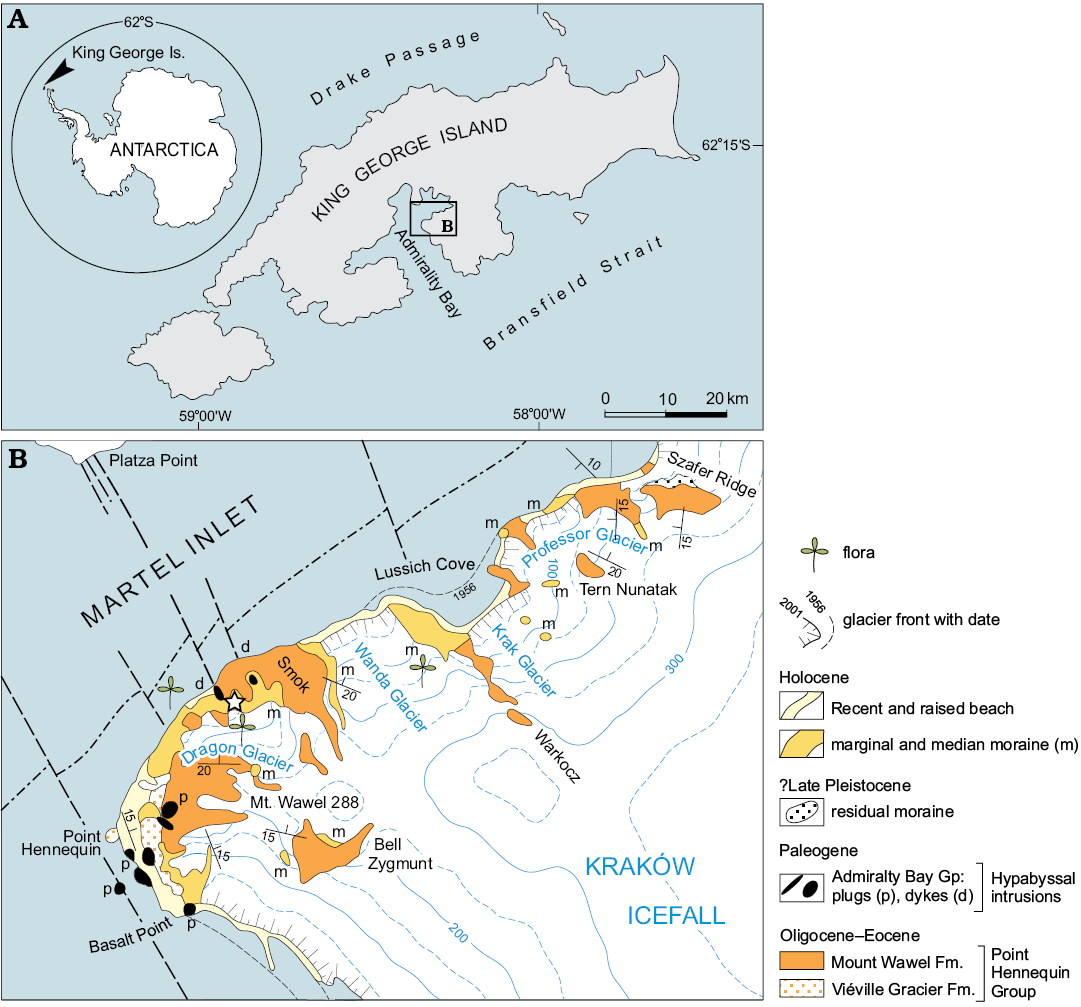
Fig. 1. Map of King George Island (A) and location of the study region (B), after Birkenmajer (2002) with locality indicated (star).

Fig. 2. Location of plant and trace fossil collection area as seen from Martel Inlet, Admiralty Bay, King George Island, Antarctica. Photograph by AG, January 2007.

Fig. 3. Some sedimentary and palaeobotanical features of the middle–late Eocene, Mount Wawel Formation, Martel Inlet Admiralty Bay, King George Island, Antarctica. A. Symmetric ripple marks on surface of very fine-grained sandstone. B. Fossil plant remains of unknown affinity in cracked mudstone. C. Delicate ferns on a parting surfaces. D. Leaves of Nothofagus sp.

Fig. 4. Lower plant (reed?) stems in siltstone-sandstone slabs from of the middle–late Eocene, Mount Wawel Formation, Martel Inlet Admiralty Bay, King George Island, Antarctica, slab ZPAL Tf.8/2007.20. A. Lower bedding-plane view with cross section of the stems (arrows) and needle-like plant detritus shallowly buried in the bed. B. Cross section of the stem in thin section showing sand-filled interior and carbonized, ribbed wall. C. Cross section of the bed with oblique stem, whose surface is carbonized. D. Cross section of a bed showing a fragment of stem with longitudinal ribbing.
Systematic palaeoichnology
Ichnofamily uncertain
Resting trace
Fig. 5A, D–E.
Material.—ZPAL Tf.8/2007.8, a slab of very fine-grained silty sandstone, middle–late Eocene, King George Island, Antarctica.
Description.—Hypichnial structure composed of an axial turbinate mound that is 21.3 mm long and associated with lateral, linear ridges. The turbinate mound is tripartite, with segments 3.5 m, 4.7 and 14 mm long. The longest segment is the widest reaching 3.5 mm in width. Three lateral, linear ridges, about 10 mm long, extended from one side of the wider part of the longest segment. The ridges are slightly curved, about 1 mm wide. Two of them are parallel, 3 mm apart. The third one, diverges at about 30° from the shorter segments. One ridge extends from the other side of the longest segment. It is 5 mm long, terminating in a hook, and inclined at about 25° to the axis of the turbinate mound. Additionally, two indistinct ridges are present on both sides of the longest segment. One of them is slightly winding, inverted over the ridges and passing on the other side, while the other runs straight outwards at an angle of 30° in respect to the axis of the segment.
Remarks.—The morphology suggests a resting trace of an insect or an imprint of a dead insect. In the second case, this is not a trace fossil, but it is impossible to judge this problem. However, its preservation in the same ways with other trace fossils in the slabs examined and general similarity to other resting traces of insects (e.g., Getty et al. 2013) is an argument for its ichnological origin. Segmentation of the axial turbinate mound corresponds to the head, thorax and abdomen of an insect. The main lateral ridges are imprints of legs. The leg imprints are complete on one side and incomplete on the other. The indistinct ridges could be cerci, however this is not clear, because they do not start from the termination of the interpreted abdomen. There are traces of wings or antennae. The systematic affiliation of this insect structure is undetermined because of the poor state of preservation; however, a beetle larva is possible.

Fig. 5. Arthropod trace fossil Glaciichnium australis isp. nov., resting trace, and lower plant stems on lower bedding surface, middle–late Eocene, Mount Wawel Formation, King George Island, Antarctica, slab ZPAL Tf.8/2007.8. A. General view, the long G. australis running from the base to the top shows different preservational variants. The knobs are plant stems. The resting trace (rt) in the lower part. B. Fragment of the long G. australis with thin blankets of underlying laminae covering the trace fossil. C. Fragment of the long G. australis crossed by another preserved mostly as the median trail, several cross sections of the lower plant stem. D. The resting trace (rt), several cross sections of the lower plant stem, and G. australis preserved mostly as the median trail. E. Resting trace, drawing (E1) and photograph of close view (E2).
Ichnofamily Protichnidae nov.
Type ichnogenus: Protichnites Owen, 1852; typified by Protichnites septemnotatus Owen, 1852; middle–late Cambrian, eastern Canada.
Included ichnogenera: Protichnites Owen, 1852, Stiallia Smith, 1909, Stiaria Smith, 1909, Siskemia Smith, 1909, Keircalia Smith, 1909, Mitchellichnus Smith, 1909, Paleohelcura Gilmore, 1926, Glaciichnium Walter, 1985.
Diagnosis.—Relatively small trackways composed of straight central trail(s) and lateral tracks.
Remarks.—Distinction between some members of the ichnofamily can be difficult, because their preservational variants can be very similar, as for instance between Stiaria, Siskemia, and Mitchellichnus (Genise 2017) or between these ichnogenera and Paleohelcura (Getty et al. 2017). This is mostly conditioned by subtle changes of substrate properties, which control morphology of trackway variants (Uchman and Pervesler 2006; Davies et al. 2007; Minter et al. 2007; Getty et al. 2017). Therefore, the new ichnofamily can be useful, especially when separation of the ichnogenera is difficult. The phrases “relatively small” and “straight” in the diagnosis are introduced in order to separate vertebrate trackways, as some of them also display a medial trail and lateral tracks but they are usually larger and commonly display sinuous median trail (e.g., Farlow et al. 2018). The use of ichnofamilies is in accordance to taxonomic rules (Rindsberg 2012) and becomes a more frequent practice in the latest decades (e.g., Genise 2000, 2017; Knaust 2015; Wisshak 2017).
Ichnogenus Glaciichnium Walter, 1985
Type ichnospecies: Glaciichnium liebegastensis Walter, 1985, Liebegast in Germany, Pleistocene.
Emended diagnosis.—Horizontal trace fossil composed of median trail and lateral, long, straight or slightly curved tracks. The tracks can be composed of up to four segments.
Remarks.—The original diagnosis by Walter (1985: 104) reads: “Relativ geringe Fährtenbreiten bis zu 5 mm. An die in Bewegungsrichtung liegenden C-Eindrücke schließen sich jeweils vorn B und A, die schräg zur Bewegunsrichtung liegen, in nach hinten gestaffelter Anordnung an”. This can be translated as: Relatively small trackways up to 5 mm wide. In the direction movement, C-footprints always terminate before footprints B and A, and adjoin toward the direction of movement, with diverse arrangement from the rear side”.
The original diagnosis is too interpretative and with use of the nomenclature (footprints A, B, C) used by Hanken and Størmer (1975) for eurypterid trackways, and which is generally not followed in research of arthropod traces. Therefore, its emendation is necessary.
Previously, Glaciichnium liebegastensis Walter, 1985 was the only ichnospecies of this ichnogenus, known only from Pleistocene varved clay in Europe (Uchman et al. 2009). It is produced by the isopod Asellus aquaticus (Linnaeus, 1758), as proven by Schwarzbach (1938) and Gibbard and Stuart (1974). This isopod lives in cold waters of the temperate zone in the Northern Hemisphere; it is an omnivorous animal, but feeds mostly on algae and plant detritus. It is able to survive temporary freezing (Gibbard and Dreimanis 1978). Glaciichnium isp. was described from the Carboniferous of southern Brazil (Lima et al. 2017). It was erroneously determined in an earlier paper as G. liebegastensis (see Lima et al. 2015).
Stratigraphic and geographic range.—Pleistocene in Germany, Poland, Lithuania, England, Sweden (Uchman et al. 2009), middle–late Eocene in King George Island, Antarctica (this study), Carboniferous in Brazil (Lima et al. 2017).
Glaciichnium australis isp. nov.
Figs. 5A–D, 6A–D, 7A–D, 8A.
Etymology: From Latin australis, southern; in reference to Southern Hemisphere.
Type material: Holotype: ZPAL Tf.8/2007.1, a slab of very fine-grained silty sandstone (Figs. 6A, 8A); paratype: ZPAL Tf.8/2007.2.
Type horizon: Mount Wawel Formation, Eocene.
Type locality: N of the Dragon Glacier, Admiralty Bay area, King George Island, Antarctica.
Diagnosis.—Glaciichnium having unilobated or bilobated median trail and lateral tracks inclined at an angle of 65–85° in respect to the median trail.
Description.—The trace fossil was observed on parting, upper or lower surfaces that resulted from cracking the rock along laminae. It is preserved as trackways and their undertracks. The trace fossil is formed by the median trail and lateral tracks (in fact preserved as undertracks). The whole structure is 10 mm wide. It displays different morphologies with transitions between them. The median trail is a smooth, semicircular furrow when epichnial, and a ridge when hypichnial, 2–3 mm wide, with a transition to a bilobate furrow (Figs. 6D, 7A–C) or ridge (Figs. 5A, C, D, 7D), with a low median ridge or furrow (Fig. 6A, C). The furrow can be bounded by low levees (Fig. 7A). The course of the median trail is irregularly winding, or with longer, gently bent segments separated by more abrupt turns of 15–25° every 10–15 cm. Very rarely single loops are present (Fig. 7D). The lateral tracks are straight or slightly curved linear furrows or ridges, 7–9 mm long, less than 1 mm wide, except for the inner terminal part, which can show a triangular widening up to 1.5 mm at the base and is unifid (terminology after Trewin 1994) at the tip. Some of the tracks are segmented into two or three depressions or ridges (Fig. 6A). They start from the edge of the median trail or at a distance of 2 mm away. They are perpendicular to the median trail or slightly oblique under the angle of 65–85°. The lateral tracks are 2–4 mm apart. They are distributed symmetrically on both sides of the median trail or only on one side. In some specimens the lateral tracks are represented only by single depressions, which disappear along the course of the trace fossil. In this case, only the central furrow or ridge may be present (Figs. 6D, 7C). It can be bilobate with a transition to a unilobate, less distinct furrow (Fig. 7B) or a ridge without lateral tracks. Locally, the trace fossil is covered by thin, flattish blanket of sediment (Fig. 5B), up to 10 mm long and elevated over the lower bedding surface, fitting the width of the entire structure.

Fig. 6. Arthropod trace fossil Glaciichnium australis isp. nov., middle–late Eocene, Mount Wawel Formation, King George Island, Antarctica, slab ZPAL Tf.8/2007.1. A. The holotype (redrawn in Fig. 8A), epichnion. The narrow diagonal furrows are damage scratches. B. Very irregular form running between the lower plant stems. C. Hypichnial forms preserved mostly as the double central trail resembling the trace fossil Diplopodichnus. D. Epichnial, unilobated median trails resembling the trace fossil Helminthoidichnites. Also delicate median bilobated trails are present.
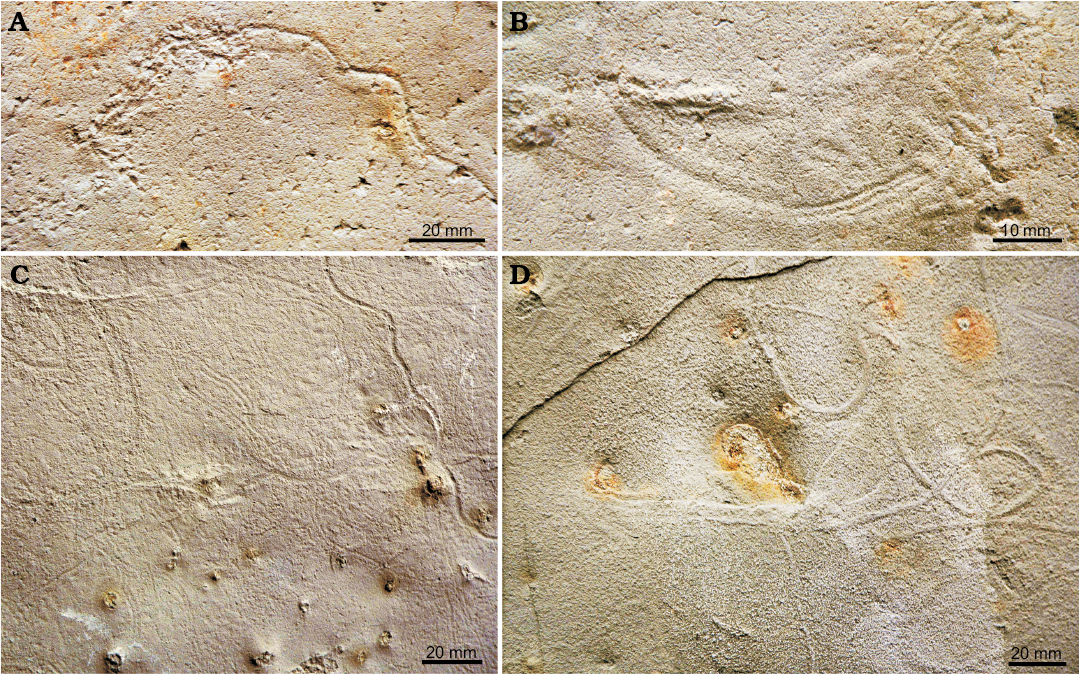
Fig. 7. Arthropod trace fossil Glaciichnium australis isp. nov. preserved mostly as unilobated or bilobated median trails, middle–late Eocene, Mount Wawel Formation, King George Island, Antarctica, slab ZPAL Tf.8/2007.25. A. Unilobate median trail passes into irregular trackway, produced probably in low cohesive substrate. B. A transition between unilobate and bilobate median trail. C. Unilobate and bilobate median trails as epichnial furrows. D. Hypichnial ridges, which are unilobated median trails resembling the trace fossil Helminthoidichnites.
Remarks.—In general, the emended ichnogeneric diagnosis matches partly the diagnosis of Protichnites Owen, 1852 by Keighley and Pickerill (1998: 95), which reads: “Trackways that, in epirelief, consist of any number of simple, or short striae imprints in similar series on either side of a central, intermittent or permanent, groove or grooves.” The main difference is that the imprints in Glaciichnium are long when completely preserved. The same concerns the diagnosis of Stiaria Smith, 1909 typified by S. quadripedia Smith, 1909 (Fig. 8F, G), which reads: “A trackway consisting of a single groove between two rows of imprints which occur in groups of two to four. These groups are set transversely or obliquely to the midline of the trackway” (Walker 1985: 290). However, the imprints in S. quadripedia are short. Also S. intermedia Smith, 1909 (Figs. 8H, 9) shows similar features. Moreover, its tracks are locally asymmetric in respect to the midline. Siskemia Smith, 1909, typified by S. elegans, 1909 (Fig. 8E) displays bilobate or trilobate central ridge, short and curved imprints (Walker 1985). However, the median trail of Glaciichnium australis is also locally bilobate. It is an open problem if these ichnogenera should be synonymised on the basis of the common two elements: central groove(s) and lateral tracks. Genise (2017: 498) suggested that Stiaria, Siskemia, and Mitchellichnus should be synonymised. The ichnotaxonomic relation of Glaciichnium to these ichnogenera is a separate problem. However, in this stage of knowledge, when the intraspecific variability of these ichnogenera is still poorly known, the possible synonymization would be premature. Instead, they can be grouped in the new ichnofamily Protichnidae.
The median trail and oblique, long or slightly curved tracks are the feature of Glaciichnium, which till now was a monospecific ichnogenus represented only by Glaciichnium liebegastensis Walter, 1985. However, the latter is narrower by half and its lateral tracks are inclined at a smaller angle with respect to the median trail (30–40° vs. 65–85° in G. australis; Fig. 8B, C). The bilobate median trail of G. australis, without lateral tracks (Figs. 6C, 7C), resembles the trace fossil Diplopodichnus Brady, 1947 (Uchman et al. 2011), but this morphotype of G. australis is only an incomplete morphological variant of this ichnotaxon, probably preserved as an undertrack. Unilobate central trails, preserved as epichnial furrows or hypichnial ridges (Figs. 6D, 7C, D), probably undertracks, are very similar to the trace fossil Helminthoidichnites Fitch, 1850 (e.g., Uchman et al. 2009). Nevertheless, the transitions from the unilobate to the bilobate trail and to the bilobate trail with lateral tracks show that they are only preservational variants of G. australis.

Fig. 8. Comparison of drawings of holotype Glaciichnium australis sp. nov. (A) to Glaciichnium liebegastensis (B holotype, C), trackway of a Recent caddisfly larvae Philopotamus montanus (D), lectotype of Siskemia elegans (E), Stiaria quadripedia (F neotype, G), and lectotype of Stiaria intermedia (H). B, C from Walter (1985: fig. 4A and B, respectively); E from Walker (1985: fig. 2a, part); F from Pollard and Walker (1984: pl. 2: 2); G from Walker (1985: fig. 5a); H from Walker (1985: fig. 5c).

Fig. 9. The lectotype of Stiaria intermedia Smith, 1909 (see also Fig. 8H), Epichnion, Old Red Sandstone, Lower Devonian, Dunure (GSE 14075), Scotland, UK. Published with permission of the British Geological Survey in Edinburgh.
Most specimens of Glaciichnium liebegastensis, including the holotype (Fig. 8B), display well expressed lateral tracks composed of two or three segments, and the median trail bounded by discontinuous, longitudinal, inwardly concave arcs. However, in some specimens, the median trail is continuous (Walter 1985: pl. 2: 5, 7; Uchman et al. 2009: fig. 5C). These specimens of G. liebegastensis are the most similar to G. australis, which shows the continuous median trail. The continuous median trail occurs in all preservational variants of G. australis, while some preservational variants of G. liebegastensis do not display the median trail. The differences somewhat challenge the ichnogeneric affinity of G. australis, but solution of the problem requires extensive studies of the ichnofamily Protichnidae, which are beyond the scope of this paper.
In general, Glaciichnium australis is very similar to the Recent trace of the caddisfly larva Philopotamus montanus (Donovan, 1813) and other caddisfly larvae of the family Leptoceridae, with agglutinated house tubes observed in shallow puddles on a dirt road along the stream in the Lejowa Valley, Tatra Mountains, southern Poland (Figs. 8D, 10). The larvae walked on the very soft muddy bottom of a puddle using three pairs of segmented legs. P. montanus lives in colder waters in submontane areas of this region. Its size is similar but the lateral tracks are inclined at an angle 30–40° with respect to the median trail. However, similar traces can be produced by apterygote insects (Getty et al. 2013).
The median trail is a trace of the body dragged on the muddy bottom, while the lateral tracks are footprints made during locomotion. The bilobate segments of the furrow indicate some double appendages along the body, such as the uropods of isopods. The levees resulted from accumulation of sediments pushed out from the furrow by ploughing the body. The local blanket of sediment covering the trace fossil consists of remnants of laminae on which the trace was imprinted. The local asymmetry of the trace fossils can be caused by currents tilting the tracemaker, which responded by placing greater pressure of appendages from one side than from the other side of its body. The variations in preservation can be caused also by subtle changes in sediment consistency. The segments showing transitions from bilobated to unilobated furrow without lateral tracks can be interpreted as a lateral transition from more to less cohesive substrate (compare Uchman and Pervesler 2006).
Glaciichnium australis can be interpreted as a locomotion and possibly a feeding trace (pascichnion). Possibly, the animal fed on microbe-rich detritus at the surface. The tracemaker of Glaciichnium australis is unknown, but isopods (by similarity to Asellus aquaticus, tracemaker of G. liebegastensis, and to Recent trace of Armadillidium vulgare; see Uchman et al. 2011) and trichopteran insects (by direct observations of traces of Philopotamus montanus and others) are so far the most reliable candidates.
Stratigraphic and geographic range.—Middle–late Eocene, King George Island, Antarctica.
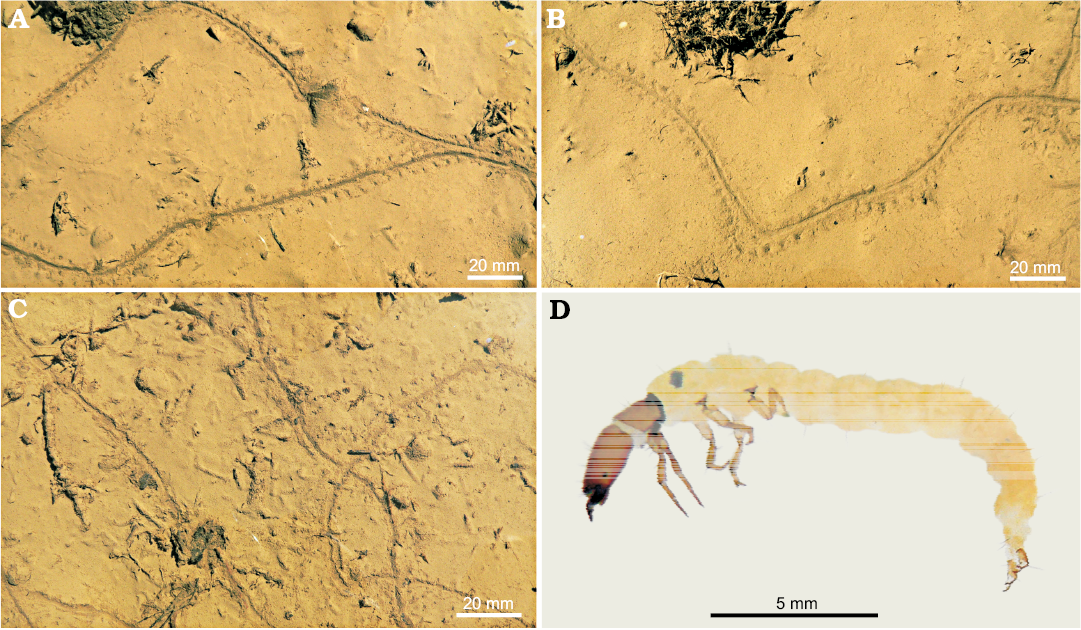
Fig. 10. Recent traces (A–C) of the caddisfly larvae Philopotamus montanus (Donovan, 1813) (D), Lejowa Valley, Tatra Mountains, southern Poland.
Ichnofamily uncertain
Ichnogenus Pterichnus Hitchcock, 1865
Type ichnospecies: Acanthichnus tardigradus Hitchcock, 1858; Triassic, north-eastern USA.
cf. Pterichnus isp.
Fig. 11A, B.
Material.—ZPAL Tf.8/2007.14, a slab of very fine-grained silty sandstone, middle–late Eocene, King George Island, Antarctica.
Description.—Hypichnial, winding trackway, 8–9 mm wide, composed of two rows of elongate ridges. The ridges are straight, curved or slightly sinuous, 1.5–3 mm long, about 1 mm wide. The ridges are inclined at an angle of 10–20° with respect to the course of the trackway. Locally, the ridges are slightly larger on one side than the other. At one place, three series of ridges are visible in the rows. Four ridges are clearly visible within each series (Fig. 11A). Ridges in the series are oriented in the same direction, which is inclined at an angle of 10–20° with respect to the course of the trackway. In another part of the same trackway, its elements are less ordered (Fig. 11B). Two rows of small ridges are accompanied by additional imprints on both sides.
Remarks.—The trackway was produced by a crawling animal that used at least four pairs of crawling appendages. Four imprints in a series are typical of Octopodichnus Gilmore, 1927, which is typified by O. didactylus Gilmore, 1927, and interpreted as a trackway of spider (Abel 1935; Braddy 2003), which suggests arid continental environments. This large trackway is characterized by two pairs of anterior bifid imprints. Bifid imprints are unclear in the described trace fossil and the rows with four imprints are visible only in a part of the trackway. The sinuous ridges (imprints) resemble the type 3 morphotype of Pterichnus isopodicus Gaillard, Hantzpergue, Vannier, Margerard, and Mazin, 2005, which is an isopod trackway from Upper Jurassic tidal flat sediments of France (Gaillard et al. 2005). The type 1 morphotype of this trace fossil also displays four imprints in per at least some series, Moreover, the inclination of imprints in respect to the course of the trackway (10–20°) is close to the value (15–20º) provided in the diagnosis of Pterichnus (Häntzschel 1975; Gaillard et al. 2005). It is not excluded that an isopod could be a candidate for the tracemaker of the trace fossil described.

Fig. 11. Hypichnial trace fossil cf. Pterichnus isp., middle–late Eocene, Mount Wawel Formation, King George Island, Antarctica, slab ZPAL Tf.8/2007.14. A. Four footprints in the series are visible in the lower left side. B. Less regular form in the lower side and the bilobate median trail of Glaciichnium australis in the upper part, which resembles the trace fossil Diplopodichnus.
Discussion
The described trace fossils co-occur with fragments of plant stems in growth position, around which crescentic marks can be observed. The rocks bearing them are siltstone, very fine sandstone or very fine-grained silty sandstone, which are parallel-laminated. This suggests that their sedimentation took place in very shallow (probably less than 1 m), but flowing water. The sediment transport episodes have been interrupted by still periods in which colonization of the floor by arthropods was possible, but even during the colonization periods, weak flows influenced the animal behaviour recorded by the asymmetry of Glaciichnium, the resting trace and cf. Pterichnus isp. The colonization periods were rather short and infrequent as only some parting surfaces contain trace fossils. Generally, the presence of symmetric (wavy) ripple marks (Fig. 3A), and delicate, well preserved fern-like plants (Fig. 3C) point to short transportation from the adjacent, vegetated land grown by trees as proved by well-preserved leaves of Nothofagus (Fig. 3D). This is in accordance to the lacustrine environment postulated for the Wawel Formation (Birkenmajer 2001), but a more specific sub-environment can be inferred from the studied trace fossils. This was a marginal part of a lake, perhaps a delta, where flows were strong enough to produce parallel lamination and to transport a large amount of plant remains. The presence of mudcracks (Fig. 3B) proves temporary exposure of sediment and suggests changes of the water level in the lake. The problem of the interpretation concerns the absence of muddy sediments typical at greater depths, which are unknown in the context of the Mount Wawel Formation. It is not excluded that this was an evanescent lake in an abandoned fluvial channel, but this requires further investigations.
As proved by the numerous leg imprints and their undertracks, the tracemakers belong to arthropods, especially insects and/or isopods. Their appearance is strongly controlled by subtle changes in substrate consistency, a typical feature of arthropod trackways (Uchman and Pervesler 2006). The trace fossil assemblage resembles the impoverished Mermia ichnofacies, which is typical of permanently submerged lacustrine sediments (Buatois and Mángano 1993, 1998, 2011). However, the absence of grazing trace fossils such as Gordia or Helminthoidichnites, which are common components of this ichnofacies, is striking. These and other ichnotaxa occur in lacustrine sediments of the Fossil Hill Formation in King George Island (Verde et al. 2017), which is lower–middle Eocene in age (Shen 1994; Mansila et al. 2013). Arthropod-dominated trace fossil assemblages, similar to the assemblage from the Mount Wawel Formation, are known from some Pleistocene lacustrine varved sediments of Europe and distinguished as the Glaciichnium ichnocoenosis (Uchman et al. 2009; formerly the cursichnia ichnocoenosis of Walter 1985 and Walter and Suhr 1998), as well as from the late Palaeozoic Gondwanan glaciolacustrine sediments (Netto et al. 2012; Lima et al. 2015, 2017). It is worthwhile noting that the dominant trace fossil from the Mount Wawel Formation also belongs to Glaciichnium and that it is very similar to the Recent trace of Philopotamus montanus, which is a colder climate insect. The occurrence of lacustrine trace fossils dominated by arthropod traces of the Mermia ichnofacies corresponds well to the beginning of climate cooling in Antarctica, which is inferred for the late Eocene on the evidence of declining floral diversity (Birkenmajer and Zastawniak 1989; Birkenmajer 1997) and decreasing marine palaeotemperatures recorded by an increase in δ18O values from molluscan faunas (Zachos et al. 2001; Hunt 2001).
The similarity between described ichnoassemblage, which is related to climatic cooling in the Antarctic, and ichnoassemblages of Pleistocene lacustrine varved sediments in Europe shows that trace fossil assemblages from both ends of the globe, separated by the tropical zone, display some recurrence of features. The same features occurred after a few million year separation as shown by the aforementioned ichnoassemblages from glaciolacustrine deposits related to the Late Palaeozoic Gondwanan glaciation. The recurrence is the fundamental basis for distinguishing of ichnofacies (Buatois and Mángano 2011; MacEachern et al. 2012). This reinforces the ichnofacies concept, especially for continental environments, in which climatic and geographic separation is stronger than in the marine realm.
Conclusions
The arthropod trackways Glaciichnium australis isp. nov. and cf. Pterichnus isp. and resting traces occur in laminated siltstone and sandstone of the Mount Wawel Formation, together with fragments of plant stems in growth position, delicate ferns and well-preserved leaves of Nothofagus. The sediments were deposited in very shallow, flowing water, probably in the marginal part of a lake, maybe in a delta, which was incidentally exposed. The trace fossils were produced particularly by insects and/or isopods, between episodes of deposition and were influenced by the water flow and subtle changes in substrate consistency.
Glaciichnium australis, with several morphotypes, is similar to those produced nowadays by some caddisflies (Trichoptera) in shallow puddles. The trace fossil assemblage is dominated by arthropod traces and it is similar to the Glaciichnium ichnocoenosis, which occurs in Pleistocene lacustrine varved sediments of Europe. This similarity corresponds well with climatic cooling in Antarctica during the late Eocene, which is recorded by declining floral diversity. This also shows that similar assemblages of trace fossils result from similar close palaeoenvironmental conditions at both ends of the globe, even after long-term separation.
Acknowledgements
We would like to thank Grażyna and Marian Dziewiński for photography of the fossil specimens and Aleksandra Hołda-Michalska for help in preparation of figures (all ZPAL). Stanisław Knutelski (Institute of Zoology, Jagiellonian University, Kraków, Poland) determined caddisfly larvae of the family Leptoceridae. Eileen A. Callaghan (British Geological Survey in Edinburgh, UK) provided photograph of Stiaria intermedia. Mariano Verde (Universidad de la Republica de Uruguay, Montevideo, Uruguay), Nicholas J. Minter (University of Portsmouth, Portsmouth, UK) and Andrew K. Rindsberg (University of West Alabama, Livingston, USA) provided constructive reviews and improved the paper. The fieldwork (BB, AG) in Antarctica was funded by a grant from the Polish Committee for Scientific Research PBZ-KBN-108/PO4/1. AU was supported by the Jagiellonian University (DS funds).
References
Abel, O. 1935. Vorzeitliche Lebensspuren. 644 pp. Gustav Fischer, Jena.
Birkenmajer, K. 1981. Lithostratigraphy of the Point Hennequin Group (Miocene volcanics and sediments) at King George Island, (South Shetland Islands, Antarctica). Studia Geologica Polonica 72: 59–73.
Birkenmajer, K. 1989. A guide to Tertiary geochronology of King George Island, West Antarctica. Polish Polar Research 10: 555–579.
Birkenmajer, K. 1997. Tertiary glacial/interglacial palaeoenvironments and sea-level changes, King George Island, West Antarctica: an overview. Bulletin of the Polish Academy of Sciences. Earth Sciences 44: 157–181.
Birkenmajer, K. 2001. Mesozoic and Cenozoic stratigraphic units in parts of the South Shetland Islands and northern Antarctic Peninsula (as used by the Polish Antarctic Programmes). Studia Geologica Polonica 118: 5–188.
Birkenmajer, K. 2002. Geological Map and Cross-sections (1:50 000). Admiralty Bay, King George Island, South Shetland Islands, West Antarctica. Institute of Geological Sciences, Cracow Research Centre and Department of Antarctic Biology, Warszawa.
Birkenmajer, K. 2003. Admiralty Bay, King George Island (South Shetland Islands, West Antarctica): a geological monograph. In: K. Birkenmajer (ed.), Geological Results of the Polish Antarctic Expeditions. Studia Geologica Polonica 120: 5–73.
Birkenmajer, K., Gaździcki, A., Krajewski, K., Przybycin, A., Solecki, A., Tatur, A., and Yoon, H.I. 2005. First Cenozoic glaciers in West Antarctica. Polish Polar Research 26: 3–12.
Birkenmajer, K. and Zastawniak, E. 1989. Late Cretaceous–early Tertiary floras of King George Island, West Antarctica: their stratigraphic distribution and palaeoclimatic significance. In: J.A. Crame (ed.), Origins and Evolution of the Antarctic Biota. Geological Society, London, Special Publications 47: 227–240.Crossref
Braddy, S.J. 2003. Trackways—arthropod locomotion. In: D.E.G. Briggs and P.R. Crowther (eds.), Palaeobiology II, 389–393. Blackwell, London.
Brady, L.F. 1947. Invertebrate tracks of the Coconino sandstone of Northern Arizona. Journal of Paleontology 21: 466–472.
Buatois, L.A. and Mángano, M.G. 1993. Trace fossils from a Carboniferous turbiditic lake: implications for the recognition of additional nonmarine ichnofacies. Ichnos 2: 237–258. Crossref
Buatois, L.A. and Mángano, G.M. 1998. Trace fossil analysis of lacustrine facies and basins. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 367–382. Crossref
Buatois, L.A. and Mángano, M.G. 2011. Ichnology: Organism-Substrate Interactions in Space and Time. 358 pp. Cambridge University Press, Cambridge. Crossref
Cantrill, D.J. and Poole, I. 2012. The Vegetation of Antarctica through Geological Time. 480 pp. Cambridge University Press, Cambridge. Crossref
Davis, R.B., Minter, N.J., and Braddy, S.J. 2007. The neoichnology of terrestrial arthropods. Palaeogeography, Palaeoclimatology, Palaeoecology, 255: 284–307. Crossref
Davies, B.J., Hambrey, M.J., Smellie, J.L., Carrivick, J.L., and Glasser, N.F. 2012. Antarctic Peninsula ice sheet evolution during the Cenozoic Era. Quaternary Science Reviews 31: 30–66. Crossref
Donovan, E. 1813. The Natural History of British Insects. Vol. 16. 225 pp. Bye & Law, London.
Dutra, T.L. 2001. Paleoflora da ilha 25 de Mayo (King George Island), Peninsula Antarctica: contribuição à paleogeografia, paleoclima e para a evolução de Nothofagus. Asociación Paleontológica Argentina, Publicación Especial 8: 29–37. Crossref
Dutra, T.L. 2004. Paleofloras de Antártica e sua relaçno com os eventos tectonicos e paleoclimáticos nas altas latitudes do sul. Revista Brasileira de Geociencias 34: 401–410.
Farlow, J.O., Robinson, N.J., Kumagai, C.J., Paladino, F.V., Falkingham, P.L., Elsey, R.M., and Martin, A.J. 2018. Trackways of the American crocodile (Crocodylus acutus) in northwestern Costa Rica: Implications for crocodylian ichnology. Ichnos 25: 30–65. Crossref
Fitch, A. 1850. A historical, topographical and agricultural survey of the County of Washington. Parts 2–5. Transactions of the New York Agricultural Society 9: 753–944.
Fontes, D. and Lindner Dutra, T. 2010. Paleogene imbricate-leaved podocarps from King George Island (Antarctica): assessing the geological context and botanical affinities. Revista Brasileira de Paleontologia 13: 189–204. Crossref
Francis, J.E., Ashworth, A., Cantrill, D.J., Crame, J.A., Howe, J., Stephens, R., Tosolini, A.-M., and Thorn, V. 2008. 100 million years of Antarctic climate evolution: evidence from fossil plants. In: A.K. Cooper, P. Barrett, H. Stagg, B. Storey, E. Stump, W. Wise, and the 10th ISAES editorial team (eds.), Antarctica: a Keystone in a Changing World. Proceedings of the 10th International Symposium on Antarctic Earth Sciences, 19–27. National Academies Press, Washington, D.C.
Francis, J.E., Marenssi, S., Levy, R., Hambrey, M. Thorn, V.C., Mohr, B., Brinkhuis, H., Warnaar, J., Zachos, J., Bohaty, S., and DeConto, R. 2009. From greenhouse to icehouse—the Eocene/Oligocene in Antarctica. In: F. Florindo and M. Siegert (eds.), Antarctic Climate Evolution. Development in Earth and Environmental Sciences 8: 309–368.
Gaillard, C., Hantzpergue, P., Vannier, J., Margerard, A.L., and Mazin, J.M. 2005. Isopod trackways from the Crayssac Lagerstätte, Upper Jurassic, France. Palaeontology 48: 947–962. Crossref
Genise, J.F. 2000. The ichnofamily Celliformidae for Celliforma and allied ichnogenera. Ichnos 7: 267–282. Crossref
Genise, J.F. 2017. Ichnoentomology: Insect Traces in Soils and Paleosols. Topics in Geobiology 37. 695 pp. Springer International, Switzerland. Crossref
Getty, P.R., Sproule, R., Stimson, M.R., and Lyons, P.C. 2017. Invertebrate trace fossils from the Pennsylvanian Rhode Island Formation of Massachusetts, USA. Atlantic Geology 53: 185–206 Crossref.
Getty, P.R., Sproule, R., Wagner, D.L., and Bush, A.M. 2013. Variation in wingless insect trace fossils: Insights from neoichnology and the Pennsylvanian of Massachusetts. Palaios 28: 243–258. Crossref
Gibbard, P.L. and Dreimanis, A. 1978. Trace fossils from late Pleistocene glacial lake sediments in southwestern Ontario, Canada. Canadian Journal of Earth Sciences 15: 1967–1976. Crossref
Gibbard, P.L. and Stuart, A.J. 1974. Trace fossils from proglacial lake sediments. Boreas 3: 69–74. Crossref
Gilmore, C.W. 1926. Fossil footprints from the Grand Canyon. Smithsonian Miscellaneous Collections 77: 1–41.
Gilmore, C.W. 1927. Fossil footprints from the Grand Canyon, 2nd Contribution. Smithsonian Miscellaneous Collections 80: 1–78.
Hanken, N. and Strømer, L. 1975. The trail of large Silurian eurypterid. Fossil and Strata 4: 225–270.
Häntzschel, W. 1975. Trace fossils and problematica. In: R.C. Moore and C. Teichert (eds.), Treatise on Invertebrate Paleontology. Part W. Miscellanea, W1–W269. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
Hitchcock, E. 1858. Ichnology of New England. A Report on the Sandstone of the Connecticut Valley, Especially its Footprints. 220 pp. W. White, Boston.
Hitchcock, E. 1865. Supplement of the Ichnology of New England. 96 pp. Wright & Porter, Boston.
Hunt, R.J. 2001. Biodiversity and Palaeoecological Significance of Tertiary Fossil Floras from King George Island, West Antarctica. 339 pp. PhD Thesis, University of Leeds, Leeds.
Hunt, R.J. and Poole, I. 2003. Revising Palaeogene West Antarctic climate and vegetation history in light of new data from King George Island. In: S.L. Wing, P.D. Gingerich, B. Schmitz, and E. Thomas (eds.), Causes and Consequences of Globally Warm Climates in the Early Paleogene. Geological Society of America Special Paper 369: 395–412. Crossref
Keighley, D.G. and Pickerill, R.K. 1998. Systematic ichnology of the Mabou and Cumberland groups (Carboniferous) of western Cape Breton Island, Eastern Canada: 2. Surface markings. Atlantic Geology 34: 83–112. Crossref
Knaust, D. 2015. Siphonichnidae (new ichnofamily) attributed to the burrowing activity of bivalves: Ichnotaxonomy, behaviour and palaeoenvironmental implications. Earth-Science Reviews 150: 497–519. Crossref
Lima, J.H.D., Minter, N.J., and Netto, R.G. 2017. Insights from functional morphology and neoichnology for determining tracemakers: a case study of the reconstruction of an ancient glacial arthropod-dominated fauna. Lethaia 50: 576–590. Crossref
Lima, J.H.D., Netto, R.G., Corrêa, C.G., and Lavina, E.L.C. 2015. Ichnology of deglaciation deposits from the Upper Carboniferous Rio do Sul Formation (Itararé Group, Paraná Basin) at central-east Santa Catarina State (southern Brazil). Journal of South American Earth Sciences 63: 137–148. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Tomus 1. 824 pp. Laurentii Salvii, Holmiae.
MacEachern, J.A., Bann, K.L., Gingras, M.K., Zonneveld, J.-P., Dashtgard, S.E., and Pemberton, S.G. 2012. The ichnofacies paradigm. In: D. Knaust and R.G. Bromley (eds.), Trace fossils as indicators of sedimentary environment. Developments in Sedimentology 64: 103–138. Crossref
Mansilla, H., Stinnesbeck, W., Varela, N., and Leppe, M. 2013. Eocene fossil feather from King George Island, South Shetland Islands, Antarctica. Antarctic Science 26: 384–388. Crossref
Minter, N.J., Braddy, S.J., and Davies, R.B. 2007. Between a rock and a hard place: arthropod trackways and ichnotaxonomy. Lethaia 40: 365–375. Crossref
Mozer, A. 2013. Eocene sedimentary facies in volcanogenic succession on King George Island, South Shetland Islands: a record of pre-ice sheet terrestrial environments in West Antarctica. Geological Quarterly 57: 385–394.
Nawrocki, J., Pańczyk, M., and Williams, I. 2011. Isotopic ages of selected magmatic rocks from King George Island (West Antarctica) controlled by magnetostratigraphy. Geological Quarterly 55: 323–334.
Netto, R.G., Benner, J.S., Buatois, L.A., Uchman, A., Mángano, G., Riddge, J.C., Kazakauskas, V., and Gaigalas, A. 2012. Glacial environments. In: R.G. Bromley and D. Knaust (eds.), Trace Fossils as Indicators of Sedimentary Environments. Developments in Sedimentology 64: 299–327. Crossref
Owen, R. 1852. Description of the impressions and footprints of the Protichnites from the Potsdam sandstone of Canada. Quarterly Journal of the Geological Society of London 8: 214–225. Crossref
Pańczyk, M. 2013. Geochronology of the volcanic rocks from the central and south part of the King George Island (South Shetland Islands, West Antarctica). Przegląd Geologiczny 61: 117–119.
Pollard, J.E. and Walker, E.F. 1984. Reassessment of sediments and trace fossils from Old Red Sandstone (Lower Devonian) of Dunure, Scotland, described by John Smith (1909). Geobios 17: 567–576. Crossref
Rindsberg, A.K. 2012. Ichnotaxonomy: finding patterns in a welter of information. In: D. Knaust and R.G. Bromley (eds.), trace fossils as indicators of sedimentary environments. Developments in Sedimentology 64: 45–78. Crossref
Schwarzbach, M. 1938. Tierfährten aus eiszeitlichen Bändertonen. Zeitschrift für Geschiebeforschung und Flachlandsgeologie 14: 143–152.
Shen, Y. 1994. Subdivision and correlation of Cretaceous to Paleogene volcano-sedimentary sequence from Fildes Peninsula, King George Island, Antarctica. In: Y. Shen (ed.), Stratigraphy and palaeontology of Fildes Peninsula, King George Island, Antarctica. State Antarctic Committee, Science Press, Monograph 3: 1–36.
Smellie, J.L., Pankhurst, R.J. Thomson, M.R.A., and Davies R.E.S. 1984. The geology of the South Shetland Islands: VI. Stratigraphy, Geochemistry and Evolution. British Antarctic Survey Scientific Reports 87: 1–85.
Smith, J. 1909. Upland Fauna of the Old Red Sandstone Formation of Carrick, Ayrshire. 60 pp. Cross, Kilwinning.
Trewin, N.H. 1994. A draft system for the identification and description of arthropod trackways. Palaeontology 37: 811–823.
Uchman, A. and Pervesler, P. 2006. Surface lebensspuren produced by amphipods and isopods (crustaceans) from the Isonzo Delta tidal flat, Italy. Palaios 21: 384–390. Crossref
Uchman, A., Kazakauskas, V., and Gaigalas, A. 2009. Trace fossils from Late Pleistocene lacustrine varve sediments in eastern Lithuania. Palaeogeography, Palaeoclimatology, Palaeoecology 272: 199–211. Crossref
Uchman, A., Hu, B., Wang, Y., and Song, H. 2011. The trace fossil Diplopodichnus from the Lower Jurassic lacustrine sediments of central China and the isopod Armadillidium vulgare (pillbug) lebensspuren as its recent analogue. Ichnos 17: 147–155. Crossref
Verde, M., Corona, A., Cabrera, F., Montenegro, F., Batista, A., Perea, D., Morosi, E., Toriño, P., and Manzuetti, A. 2017. Mermia to Scoyenia-shorebird ichnofacies in lacustrine deposits from the Eocene of Antarctica. In: E.M. Bordy (ed.), 2nd International Conference of Continental Ichnology, Nuy Valley, West Cape, South Africa, 1st–4th October 2017, Abstract Book, 91–92. University of Cape Town, Cape Town.
Walker, E.F. 1985. Arthropod ichnofauna of the Old Red Sandstone at Dunure and Montrose, Scotland. Transactions of the Royal Society of Edinburgh, Earth Sciences 76: 287–297. Crossref
Walter, H. 1985. Zur Ichnologie des Pleistozäns von Liebegast. Freiberger Forschungshefte C400: 101–116.
Walter, H. and Suhr, P. 1998. Lebensspuren aus kalzeitlichen Bändersedimenten des Quartärs. Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden 43/44: 311–328.
Wisshak, M. 2017. Taming an ichnotaxonomical Pandora’s box: revision of dendritic and rosetted microborings (ichnofamily: Dendrinidae). European Journal of Taxonomy 390: 1–99. Crossref
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292: 686–693. Crossref
Zastawniak, E., Wrona, R., Gaździcki, A., and Birkenmajer, K. 1985. Plant remains from the top part of the Point Hennequin Group (Upper Oligocene), King George Island (South Shetland Islands, Antarctica). Studia Geologica Polonica 81: 143–164.
Acta Palaeontol. Pol. 63 (2): 383–396, 2018
https://doi.org/10.4202/app.00467.2018