
Cranial variability of the European Middle Triassic sauropterygian Simosaurus gaillardoti
CARLOS DE MIGUEL CHAVES, FRANCISCO ORTEGA, and ADÁN PÉREZ-GARCÍA
Miguel Chaves, C. de, Ortega, F., and Pérez-García, A. 2018. Cranial variability of the European Middle Triassic sauropterygian Simosaurus gaillardoti. Acta Palaeontologica Polonica 63 (2): 315–326.
Simosaurus is a Triassic eosauropterygian genus known from cranial and postcranial elements, found in the Middle and Upper Triassic strata of Europe and the Middle East. Simosaurus gaillardoti is currently recognised as the only valid species of the genus, identified in Ladinian strata of an area that includes northeastern France (Lorraine) and southwestern Germany (Baden-Württemberg). The remains from this area include more than 20 skulls. Although some of them were described in detail and figured by several authors since the discovery of S. gaillardoti, many others remained unpublished. Here we study and figure 25 skulls from this taxon, reviewing the previously known specimens, and presenting and analyzing numerous unpublished skulls. This significant sample of skulls from a single taxon of a European Triassic eosauropterygian allows us to recognise intraspecific variability in characters previously identified as non-variable in this species, including some that are often included in phylogenetic analyses of these reptiles. Therefore, we markedly increase our knowledge of the cranial skeleton in this taxon.
Key words: Sauropterygia, Simosauridae, skulls, Triassic, Ladinian, France, Germany.
Carlos de Miguel Chaves [carlos.miguelchaves@gmail.com], Francisco Ortega [fortega@ccia.uned.es], and Adán Pérez-García [paleontologo@gmail.com], Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, Paseo de la Senda del Rey 9, 28040, Madrid, Spain.
Received 22 February 2018, accepted 2 April 2018, available online 4 June 2018.
Copyright © 2018 C. de Miguel Chaves et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Within the great diversity of Triassic sauropterygians, Simosaurus Meyer, 1842 is a nothosauroid genus of relatively large body size (approximately 3–4 meters), known both by the skull and by the postcranial skeleton (Rieppel 1994a, 2000). The fossil record of this taxon ranges from the Anisian–Ladinian boundary (Middle Triassic) to the Carnian (Late Triassic), and includes remains from Austria, France, Germany, Italy, Spain, Israel, and Saudi Arabia (Haas 1981; Rieppel 1994a, 1996, 2000; Rieppel et al. 1999; Vickers-Rich et al. 1999; Dalla Vecchia 2008; Kear et al. 2010; Miguel Chaves et al. 2015).
Simosaurus gaillardoti Meyer, 1842, from the Ladinian (Middle Triassic) strata of an area corresponding to the northeastern region of France and the southwestern region of Germany (see Appendix 1), is the only valid species of Simosaurus currently recognised (Rieppel 1994a). It was described for the first time by Meyer (1842), based on cranial material from the upper Muschelkalk levels of Lunéville (France). A putative second species, Simosaurus mougeoti Meyer, 1842, was described in the same paper based on a partial lower jaw from the same age and locality, being subsequently recognised as belonging to the genus Nothosaurus Münster, 1834 (Meyer 1847–1855). Some years later, a new simosaur species, Simosaurus guilielmi Meyer, 1852 was described based on a skull (SMNS 16700) from the lower Keuper (upper Ladinian) of Hoheneck, Baden-Württemberg, southwestern Germany (Meyer 1847–1855). Huene (1959) defined Simosaurus guilielmi var. angusticeps by a partial articulated skeleton, including a skull, from the Gipskeuper (upper Ladinian) of Obersontheim (Baden-Württemberg) (i.e., GPIT/RE/1888). The comparison of these two specimens with several skulls of S. gaillardoti from the upper Muschelkalk (Ladinian) of southwestern Germany led Rieppel (1994a) to identify them as also belonging to S. gaillardoti.
Meyer (1842) did not figure any material of S. gaillardoti from Lunéville in the paper where he defined this taxon, but some skulls from the same horizon and locality were subsequently figured with drawings by himself (Meyer 1847–1855: pl. 16: 1, pl. 17, pl. 18: 1, pl. 19), and also by Gervais (1859: pl. 55: 2, pl. 56: 1–3) and Gaudry (1890: fig. 288) (see Discussion). However, most of these historical specimens from Lunéville are currently lost, including the holotype (see Rieppel 1994a). Thus, MNHN.AC. 9028, a preserved skull from Lunéville (Gaudry 1890: fig. 288), was chosen as the neotype of S. gaillardoti by Rieppel (2000). In addition, several skulls of S. gaillardoti have been recovered over the years from Baden-Würtemberg (southwestern Germany), but only some of them having been described and figured (see Appendix 1 and references therein).
Here we present and figure a compilation of most of the skulls of S. gaillardoti found so far (Figs. 1–5); most of them were never figured. The study of this abundant collection of skulls allows us to analyse and compare the morphology of these specimens and the arrangement of the sutures in order to identify intraspecific variability, improving known information on the cranial anatomy of S. gaillardoti.
Institutional abbreviations.—BSP, Bayerische Staatssammlung für Paläontologie und Historische Geologie, Munich, Germany; GPIT/RE, Geologisch-Paläontologisches Institut, Tübingen Universität, Germany; HUJ-Pal., Paleontological Collections, Department of Evolution, Systematics and Ecology, The Hebrew University, Jerusalem, Israel; MB.R., Museum für Naturkunde, Fossil Reptile Collection, Berlin, Germany; MHI, Muschelkalkmuseum, Ingelfingen, Germany; MNHN, Muséum national d’Histoire naturelle, Paris, France; SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany.
Material and methods
A total of 25 skulls of Simosaurus gaillardoti have been analysed first-hand and figured in this paper (Figs. 1–5), including the neotype of S. gaillardoti (MNHN.F.AC. 9028), the holotypes of “Simosaurus guilielmi” (SMNS 16700) and “Simosaurus guilielmi var. angusticeps” (GPIT/RE/1888), and a cast of a classic skull from Lunéville, currently lost (MNHN.F.AC. 9026). Considering all this information, an idealised reconstruction of the skull of Simosaurus gaillardoti in dorsal, ventral and occipital views, is provided here (Fig. 6). This reconstruction is based on the character states recognised as most frequent in the samples analysed in this paper. The skulls figured and analysed here are kept in several German institutions (Geologisch-Paläontologisches Institut of the Tübingen Universität, Museum für Naturkunde of Berlin, Muschelkalkmuseum Hagdorn, Staatliches Museum für Naturkunde of Stuttgart) and in the Muséum national d’Histoire naturelle of Paris (Appendix 1). The identification of the morphology and sutures of the skulls, and therefore their inclusion in the figures in the different views, has been realised only when the preservation of the specimens has allowed it, and only when they have been analysed by personal observation.
Thus, most of the skulls of Simosaurus currently preserved in scientific institutions and cited in the literature have been analysed here. The only skulls not figured nor studied here are the skull of S. gaillardoti BSP 1932.1.13 (Ladinian, upper Muschelkalk of Tiefenbach, Germany), and HUJ-Pal. 2086, a fragmentary skull classified as Simosaurus sp. (Anisian–Ladinian boundary, Muschelkalk of Makhtesh Ramon, Israel; see Haas 1981), as a first-hand study could not be undertaken. On the other hand, several unpublished or never figured specimens are included here (see Appendix 1).
Because a first-hand detailed observation of the holotype of “S. guilielmi var. angusticeps” (GPIT/RE/1888) has not been possible due to its location in the exhibition of the GPIT/RE, this specimen is only represented here by dorsal photography, but not by a detailed drawing (Fig. 1U). As MNHN.F.AC. 9026 corresponds to an imperfect cast of the original specimen, now lost, it is also exclusively figured by photographs (Figs. 1C, 3A).
For a detailed description of the cranial anatomy of S. gaillardoti see Rieppel (1994a, b).
In order to test the phylogenetic effect of the identified variability in the skull of S. gaillardoti, a cladistic analysis has been performed here based on a modified version of the data matrix of Cheng et al. (2016), which is based on that from Neenan et al. (2013). The eosauropterygian taxa Chinchenia sungi, Kwangsisaurus orientalis, and Sanchiaosaurus dengi, from the Middle Triassic of China, have been excluded following previous proposals (e.g., Neenan et al. 2013; Cheng et al. 2016). The following characters have been recoded in S. gaillardoti following the study of the skulls: character 16 from 0 to 0 and 1, character 17 from 0 to 0 and 1, and character 42 from 0 to 0 and 1 (see Discussion). The data matrix comprises 48 taxa and 141 characters. It was analysed using TNT 1.0 (Goloboff et al. 2008) in order to find the most parsimonious trees (MPTs). A traditional search was used for the analysis, with 1000 replications of Wagner trees (using random addition sequences), followed by tree bisection recognition (TBR) as a swapping algorithm, saving 100 trees per replication. To test the robustness of this phylogenetic hypothesis, Bremer support and bootstrap frequencies (absolute frequencies based on 1000 replications) values were also calculated using TNT.

Fig. 1. Skulls of simosaurid sauropterygian Simosaurus gaillardoti Meyer, 1842 from the Ladinian, Middle Triassic of southwestern Germany (A, E–Y) and northeastern France (B–D) in dorsal view. A. MB.R. 52. B. MNHN.F.AC. 9028 (neotype of Simosaurus gaillardoti). C. MNHN.F.AC. 9026 (cast). D. MNHN.F.AC. 9025. E. SMNS 18274. F. SMNS 16639. G. SMNS 16767. H. SMNS 18520. I. SMNS 16700 (holotype of “Simosaurus guilielmi”). J. SMNS 56288. K. SMNS 11364. L. SMNS 59366. M. SMNS 18550. N. SMNS 16735a. O. SMSN 18220. P. GPIT/RE/09313. Q. SMNS 10360. R. MHI 1366. S. SMNS 50714. T. MHI 1833. U. GPIT/RE/1888 (holotype of “Simosaurus guilielmi var. angusticeps” Huene, 1959). V. SMNS 16363. W. SMNS 18637. X. SMNS 50715. Y. SMNS 59943.
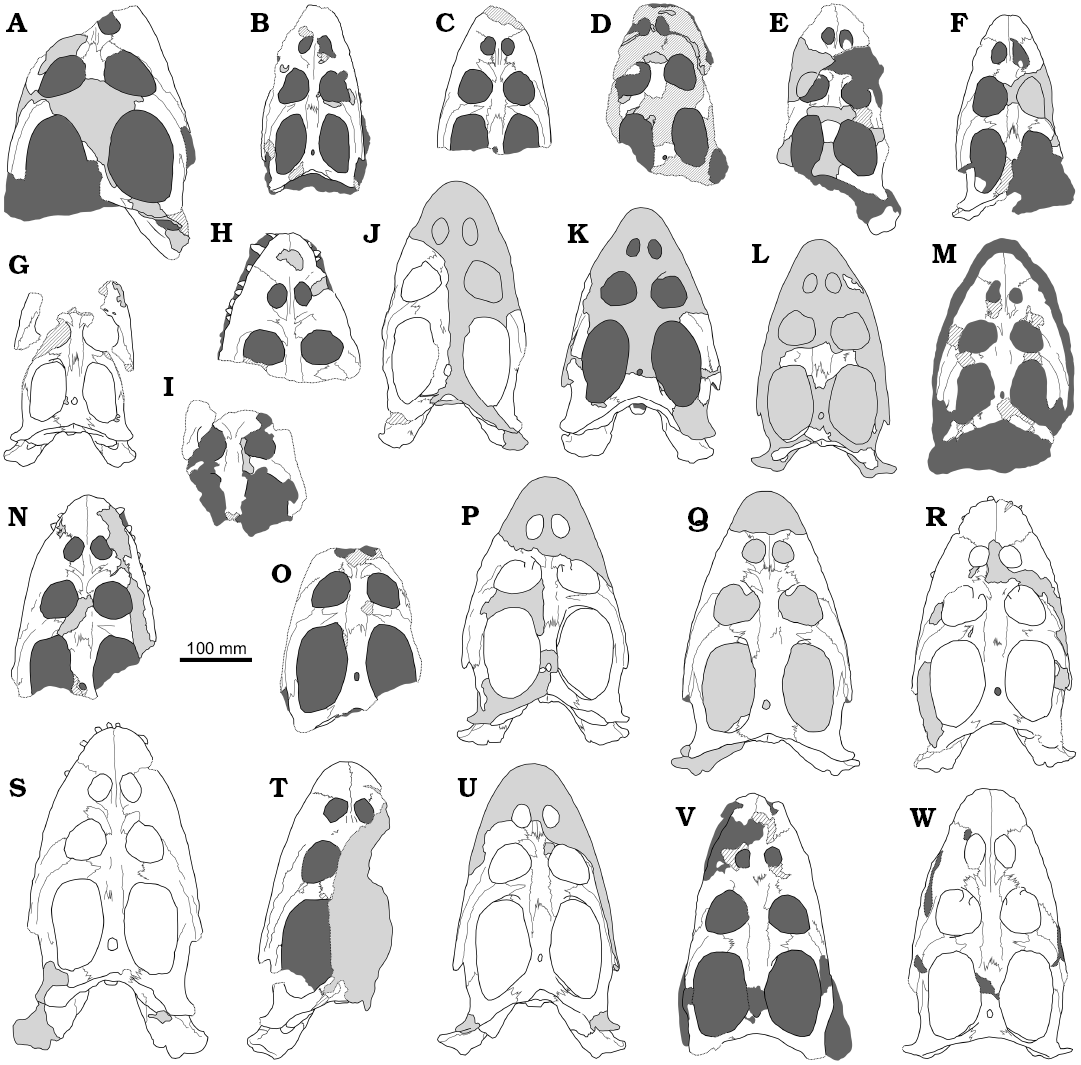
Fig. 2. Schematic interpretations of the skulls of simosaurid sauropterygian Simosaurus gaillardoti Meyer, 1842 from the Ladinian, Middle Triassic of northeastern France (B, D) and southwestern Germany (A, C, E–W), in dorsal view. A. MB.R. 52. B. MNHN.F.AC. 9028 (neotype of Simosaurus gaillardoti). C. SMNS 18520. D. MNHN.F.AC. 9025. E. SMNS 16639. F. SMNS 18274. G. SMNS 16767. H. SMSN 18220. I. SMNS 16735a. J. MHI 1366. K. SMNS 59366. L. SMNS 56288. M. SMNS 16700 (holotype of “Simosaurus guilielmi” Meyer, 1852). N. SMNS 18550. O. SMNS 11364. P. SMNS 50714. Q. SMNS 16363. R. GPIT/RE/09313. S. MHI 1833. T. SMNS 18637. U. SMNS 50715. V. SMNS 59943. W. SMNS 10360. Light grey, plaster; dark grey, matrix; grated areas, broken or altered bones; dashed lines, limits of broken bones; thin grey lines, sutures.
Results
The Ladinian levels of an area located in northeastern of France and southwestern Germany have provided an important record for specimens of Simosaurus gaillardoti. More than 25 skulls have been recovered from different localities in this area, including specimens from both the upper Muschelkalk and the lower Keuper. The detailed study of this abundance of cranial material performed here allows us to confirm that, as previously indicated by Rieppel (1994a), variability in the general morphology of the skull of S. gaillardoti cannot be clearly established (e.g., ratio length/width of the skull, straight or curved lateral skull margins, rounded or pointed anterior tip of the snout). This is due not only to the distortion of some skulls, but also due to the fact that several of them are only partially preserved, having been reconstructed with a plaster (see Fig. 2). Rieppel (1994a) recognised intraspecific variability considering the ratios between several elements not affected by the distortion: longitudinal diameter of the upper temporal fossa/longitudinal diameter of the orbit, width of the postorbital arch/distance between the external naris and the orbit, and width of the frontal between the orbits/width of the bony bridge separating the external nares. The study of the large number of skulls analysed here confirms the variability proposed by Rieppel (1994a) for all these characters. However, this study also allows us to recognise many other cranial characters of S. gaillardoti that are subject to variability.
The size, shape and position of the external nares are here identified as relatively variable. For example, SMNS 59943 (Figs. 1Y, 2V) presents relatively small external nares compared to the other skulls, its length being only one third of the length of the orbits, where on the other hand, the length of the external nares of SMNS 10360 (Figs. 1Q, 2W) is three-quarters the length of the orbits. The length of the external nares in other skulls ranges between these two specimens. In addition, the external nares are elongated in SMNS 18274 (Figs. 1E, 2F) and SMNS 10360 (Figs. 1Q, 2W), with a length 70% larger than the width, whereas they are more rounded (i.e., the length of the external nares is equal to their width) in other specimens (e.g., Figs. 1P, 2R). The morphology of the external nares in most specimens presents intermediate states for this character. Finally, the external nares are more anteriorly located relative to the orbits in SMNS 16639 (Figs. 1F, 2E) when compared with those of other skulls. Thus, in SMNS 16639, the distance between the anterior margin of the nares and the preserved tip of the snout is smaller than the distance between the posterior margin of the nares and the anterior border of the orbits, while the opposite condition in present in other skulls (e.g., Figs. 1O and 2H, 1P and 2R, 1Y and 2V). Some external nares are located the same distance from both the tip of the snout and the orbits (e.g., Figs. 1O and 2H, 1Q and 2W).
The morphology of the pineal foramen is also identified as relatively variable, being oval in some specimens (e.g., SMNS 11364, in which it is almost twice as long as it is wide; Figs. 1K, 2O), but subrounded in most of the skulls, with the length equal or subequal to the width (e.g., Fig. 2D, Q, R, S). Although the pineal foramen is always located in a position somewhat behind the middle region of the upper temporal fossae in the skulls of S. gaillardoti (e.g., Fig. 2B, G, Q, S, W), it is located close to the middle length of the parietal skull table in other specimens (e.g., Figs. 1I, K, 2M, O). In addition, the shape of the upper temporal fossae ranges from oval, with a rounded contour in most of the skulls (e.g., Figs. 1A, P, Q, S, T, V, Y, 2A, P–S, V, W), to more angulate, being kidney-shaped in others (Figs. 1B, E, I, 2B, F, M). Intermediate morphologies are also present (e.g., Figs. 1L, R, X, 2J, K, U).
The shape and contact between some bones of the dorsal area of the skulls of S. gaillardoti, especially those of the preorbital region, are also recognised as subject to variability. Rieppel (1994a) described the nasals of S. gaillardoti as small and triangular bones (Fig. 2H, W), which define the posteromedial margins of the external naris. Our observations show that these bones can also be trapezoidal in some specimens (e.g., Fig. 2M, N, Q, R). In addition, whereas in some skulls the nasals only define the posteromedial margin of the external nares (i.e., less than the half of the posterior width of the nares; Figs. 2C, R, U, W), in others they constitute more than the half of the posterior width of the external nares in others (Fig. 2H, Q).
The nasals may (Fig. 2A, W) or may not (Fig. 2C, H, N, Q–S, U, W) contact the prefrontal, depending on the extension of the anterolateral process of the frontal that stands between both bones. When present, the contact between the nasal and the prefrontal can be recognised as relatively long (i.e., the length of this contact being larger than the length of the contact between the nasal and the maxilla in MB.R.52; Fig. 2A), or as short (i.e., the length of the contact being smaller than the length of the contact between the nasal and the maxilla in the right nasal and prefrontal in SMNS 10360; Fig. 2W). In addition, intraindividual variation was identified in this last specimen by Rieppel (1994a), the left side lacking a nasal-prefrontal contact (Fig. 2W). In addition, in SMNS 18220 and MHI 1833 (Fig. 2H and S, respectively), with poorly developed anterolateral processes of the frontals, the maxillae contribute to the separation between the nasals and the prefrontals, whereas in the other skulls the maxilla does not participate in this separation (e.g., Fig. 2N, Q, U, W).
Rieppel (1994a) indicated that the fused frontals of S. gaillardoti contact with the parietals in a deeply interdigitating suture somewhat behind the anterior margin of the upper temporal fossae, lacking posterolateral processes (Fig. 2B, L, P). However, the presence of posterolateral processes of the frontals is identified in some of the skulls studied here (Fig. 2C, N, U), these processes being clearly separated from these fossae in some of them (Fig. 2N), but in contact or almost in contact with the fossae in others (Fig. 2C, U). Intermediate states considering the development of the posterolateral processes, being slightly marked, are also present (Fig. 2O, W). In addition, variability in the location of the contact between the frontals and the parietals is also recognised here. Thus, in addition to the location previously indicated (Fig. 2P, S, U, W), this suture is located at the same level as the anterior margin of the upper temporal fossae in some specimens (Fig. 2F, L, N, O, R), being slightly anterior to that margin in others (Fig. 2B, G, Q). Some variation can also be recognised relative to the morphology of the contact between the postfrontals and the postorbitals. Most of the specimens of S. gaillardoti present postorbitals with a prominent medial process in the contact with the postfrontals (e.g., Fig. 2C, N, O), but this process is very poorly developed in others (Fig. 2R, T, V).
Rieppel (1994a) indicated that the foramen incisivum of S. gaillardoti was anteriorly and laterally enclosed by the premaxillae, and posteriorly by the vomers (e.g., Fig. 4G, Q, T). However, the foramen incisivum of GPIT/RE/09313 and SMNS 10360 (Fig. 4N, O) is recognised here as exclusively enclosed by the premaxillae. The location of this foramen also displays variation, being located anteriorly in SMNS 18274 with respect to its position in other skulls. Thus, in this specimen, the foramen incisivum is located well in front of the most anterior part of the contact between the premaxillae and the maxillae (Fig. 4B), whereas in other skulls it is located at the same level that the most anterior part of this contact (Fig. 4O, T).
The internal nares are anteriorly limited by the premaxillae, medially by the vomers, posteriorly by palatines and laterally by the maxillae (Rieppel 1994a). However, in some specimens the maxillae expand medially beyond the half of the width of the internal nares, almost reaching the vomers and reducing the contact between the premaxilla and the internal naris (Fig. 4C, E, P). In other skulls, the maxillae do not reach the medial half of the internal nares, the contact between the premaxillae and the anterior margin of the nares being equivalent to one third of the total width of the latter (Fig. 4C, N, O).
The vomers of most specimens of S. gaillardoti are posteriorly separated from other by an anterior process constituted by both pterygoids (e.g., Fig. 4E, O, R). This condition was considered by Rieppel (1994a) for the characterization of S. gaillardoti. However, the anterior processes of the pterygoids are identified as much reduced, being almost absent in one of the specimens studied here (Fig. 4S).
The posterior area of the palatines is wider than the anterior region of the ectopterygoids in most specimens, the width of the posterior area of the palatines being a third larger than the width of the anterior area of the ectopterygoids (e.g., Fig. 4E, M, R, S). However, the width of the posterior part of the palatines and the anterior part of the ectopterygoids are similar in other specimens (Fig. 4G, N). Intermediate states for this character are also present (e.g., Fig. 4B).
Finally, a poorly developed occipital crest is recognized in several skulls of S. gaillardoti (Fig. 5B, C, F, I). This crest is totally absent in other specimens (Fig. 5D, E, G, H, L, M).

Fig. 3. Skulls of simosaurid sauropterygian Simosaurus gaillardoti Meyer, 1842 from the Ladinian, Middle Triassic of northeastern France (A, B) and southwestern Germany (C–V), in ventral view. A. MNHN.F.AC. 9026 (cast). B. MNHN.F.AC. 9025. C. SMNS 18274. D. SMNS 16639. E. SMNS 16767. F. SMNS 18520. G. SMNS 16700 (holotype of “Simosaurus guilielmi” Meyer, 1852). H. SMNS 11364. I. SMNS 18550. J. SMNS 16735a. K. SMNS 56288. L. SMNS 59366. M. MHI 1366. N. SMNS 50714. O. GPIT/RE/09313. P. SMNS 10360. Q. SMNS 16363. R. MHI 1833. S. SMNS 50715. T. SMNS 18637. U. MB.R. 52. V. SMNS 11364b.

Fig. 4. Schematic interpretations of the skulls of simosaurid sauropterygian Simosaurus gaillardoti Meyer, 1842 from the Ladinian, Middle Triassic of northeastern France (A) and southwestern Germany (B–U), in ventral view. A. MNHN.F.AC. 9025. B. SMNS 18274. C. SMNS 16639. D. SMNS 16767. E. SMNS 18520. F. SMNS 16700 (holotype of “Simosaurus guilielmi” Meyer, 1852). G. SMNS 56288. H. SMNS 11364. I. SMNS 59366. J. SMNS 18550. K. SMNS 16735a. L. MHI 1366. M. SMNS 50714. N. GPIT/RE/09313. O. SMNS 10360. P. SMNS 16363. Q. SMNS 18637. R. MB.R. 52. S. SMNS 50715. T. MHI 1833. U. SMNS 11364b. Light grey, plaster; dark grey, matrix; grated areas, broken or altered bones; dashed lines, limits of broken bones; thin grey lines, sutures.

Fig. 5. Skulls of simosaurid sauropterygian Simosaurus gaillardoti Meyer, 1842 from the Ladinian, Middle Triassic of southwestern Germany, in occipital view. A. SMNS 18274. B. SMNS 16767. C. SMNS 56288. D. SMNS 59366. E. MHI 1366. F. SMNS 50714. G. GPIT/RE/09313. H. SMNS 10360. I. SMNS 16363. J. SMNS 59943. K. SMNS 18637. L. SMNS 50715. M. MHI 1833. Photographs (A1–M1), schematic interpretations (A2–M2). Light grey, plaster; dark grey, matrix; grated areas, broken or altered bones; dashed lines, limits of broken bones; thin grey lines, sutures.
Discussion
Although the fossil record of the genus Simosaurus ranges from the Anisian–Ladinian boundary (Middle Triassic; Rieppel et al. 1999) to the Carnian (Upper Triassic; Dalla Vecchia 2008), only a single species is currently recognised within this genus—Simosaurus gaillardoti. Its presence was confirmed in Ladinian strata of the French-German area from where the 25 skulls analysed here come (Rieppel 19994a). The variability of character states in these skulls is not recognised as defined by any patern (e.g., geographic, temporal, systematic, or ontogenetic), but is observed to be distributed in an uncorrelated way, interpreted as intraspecific variability. Two main areas with recovered skulls of S. gaillardoti have been hitherto identified: Lorraine (France) and Baden-Württemberg (Germany). French specimens (Figs. 2B, D, 4A) lack remarkable differences when compared to the German skulls (Figs. 2A, C, E–W, 4B–U). In fact, variation in two characters is observed when comparing the two French specimens analysed here: the presence of a kidney-shaped upper temporal fenestrae in one of them (Fig. 2B), which is rounded in the other (Fig. 2D), and an oval pineal foramen in the first one (Fig. 2B), which is rounded in the second (Fig. 2D). This variability has been noted in the specimens from Baden-Württemberg.
None of the characters analysed here appear to reflect clear ontogenetic variation. Thus, the size, shape and location of the external nares; the contact between several dorsal and palatal bones; the shape and position of the pineal foramen; the shape of the upper temporal fossae; the nature of the bones constituting the foramen incisivum; and the presence or absence of the occipital crest, are characters that experience some degree of variation in both large and small specimens, with no obvious ontogenetic or size-related trend (Figs. 2, 4, 5).
The only analysed skull from the Keuper, the holotype of “S. guilielmi” (SMNS 16700) presents small and rounded external nares, trapezoidal nasals, kidney-shaped upper temporal fenestrae and postorbitals with poorly developed medial processes (Fig. 2M). These character states are also present in other large and small skulls of Simosaurus gaillardoti found in the upper Muschelkalk.
Therefore, we cannot separate the skulls of S. gaillardoti into groups based on any characters analysed here. Thus, all the specimens analysed in this paper are compatible with the currently available diagnosis of S. gaillardoti (see Rieppel 2000). The variability observed here does not affect any of the characters considered in this diagnosis.
The variability identified here affects the previous scorings of some characters for S. gaillardoti in several previous phylogenetic analyses (e.g., Rieppel 1994a, 1998; Neenan et al. 2013; Cheng et al. 2016). For instance, the posterolateral processes of the frontal (character 16 in Neenan et al. 2013), previously recognised as absent in this taxon (state 0), is also identified as present in some specimens (state 1; e.g., Fig. 2U). The position of the frontal with respect to the upper temporal fossa (character 17 in Neenan et al. 2013), with both elements previously recognised as widely separated (state 0), displays variation; the frontals of some specimens are in fact close to the upper temporal fossae (state 1; e.g., Fig. 2C). The occipital crest (identified as absent in character 42 in Neenan et al. 2013; state 0) is here also identified as in some specimens (state 1; e.g., Fig. 5I). As a result of the rescoring of these three characters in the data matrix considered here, the phylogenetic position of S. gaillardoti does not change with respect to previous studies (e.g., Rieppel 1994a, 1998; Neenan et al. 2013; Cheng et al. 2016). Thus, this taxon is still recognised as a member of Nothosaroidea, and the sister taxon of the clade Nothosauria (Fig. 7).
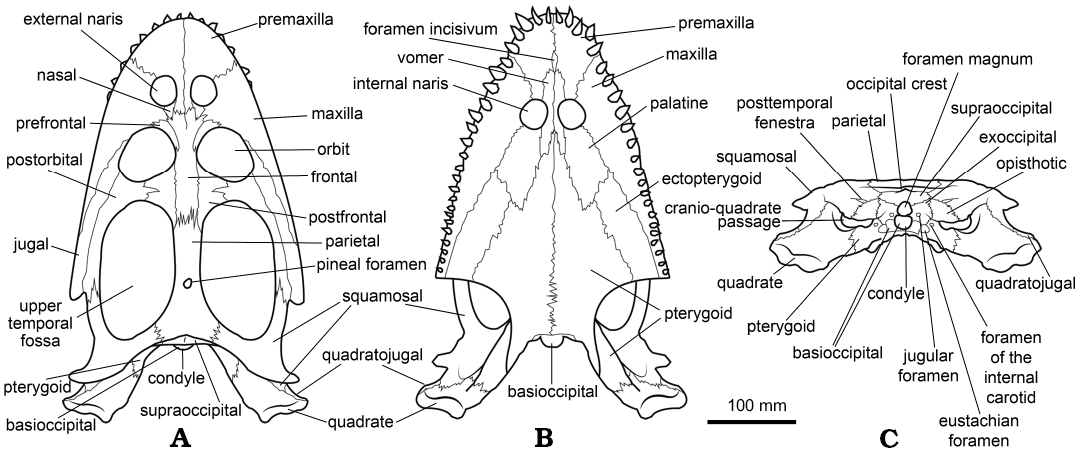
Fig. 6. Schematic reconstruction of an idealized skull of Simosaurus gaillardoti Meyer, 1842, in dorsal (A), ventral (B), and occipital (C) views.
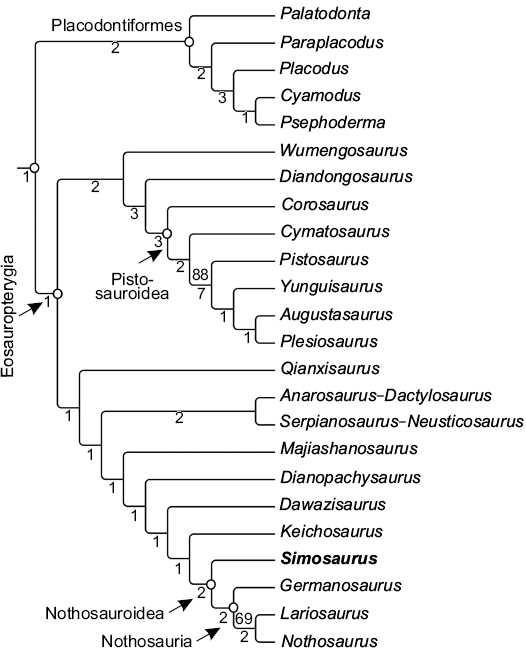
Fig. 7. Phylogeny of Sauropterygia from the strict consensus tree obtained from our phylogenetic analysis based on the modified data matrix of Cheng et al. (2016). The phylogenetic analysis resulted in four most parsimonious trees, with a length of 615 steps (CI = 0.311; RI = 0.663; RC = 0.206). Bootstrap frequencies that exceed 50 per cent (top) and Bremer support values (bottom) are indicated.
Conclusions
Simosaurus gaillardoti is a nothosauroid eosauropterygian known from the Ladinian (Middle Triassic) levels of an area that includes northeastern France (Lorraine) and southwestern Germany (Baden-Würtemberg). The study of 25 skulls from this region, corresponding to different ontogenetic stages, allows us to recognise variation in several characters. This intraspecific variation cannot be easily explained by ontogenetic, geographic, temporal or sexual causes. This study also allows us to recognise the most frequent state for these variable characters in S. gaillardoti, as well as to rescore some of them in the data matrices.
A revision of the postcranial skeleton of S. gaillardoti is currently in progress. That study, the information provided here relative to cranial variability, and the description of a new sauropterygian taxon under study, also attributable to Simosauridae, will allow us to review the complete phylogenetic scoring of S. gaillardoti and improve our knowledge of the phylogenetic position of this clade.
Acknowledgements
We thank Rainer Schoch (SMNS), Nour-Eddine Jalil (MNHN), Hans Hagdorn (MHI), Ingmar Werneburg (GPIT/RE), and Thomas Schossleitner and Daniela Schwarz (both MB.R) for the access to the specimens studied here. We thank Sven Sachs (Naturkunde Museum Bielefeld, Germany) for providing abundant bibliography and information. We thank the editor Stephen Brusatte (University of Edinburgh, UK), the reviewer Olivier Rieppel (Field Museum of Natural History, Chicago, USA) and an anonymous reviewer for their kind comments, which have helped to improve this paper. This research has been funded by a FPI UNED Grant (Ref. 0271864713 Y0SC001170) and the Ayudas para Estancias breves en España y en el extranjero 2017 of the UNED, and received support from the SYNTHESYS Project (http://www.synthesys.info/) which was financed by European Community Research Infrastructure Action under the FP7 ‘‘Capacities’’ Program (FR-TAF-5710 and DE-TAF-5955).
References
Arthaber, G. 1924. Die Phylogenie der Nothosaurier. Acta Zoologica, Stockholm 5: 439–516. Crossref
Cheng, Y., Wu, X., Sato, T., and Shan, H. 2016. Dawazisaurus brevis, a new eosauropterygian from the Middle Triassic of Yunnan. China. Acta Geologica Sinica (English Edition) 90 (2): 401–424. Crossref
Dalla Vechia, F.M. 2008. First record of Simosaurus (Sauropterygia, Nothosauroidea) from the Carnian (Late Triassic) of Italy. Rivista Italiana di Paleontologia e Stratigrafia 114: 273–285.
Fraas, E. 1896. Die Schwäbischen Trias-Saurier. 18 pp. E. Schweizerbart, Stuttgart.
Gaudry, A. 1890. Les Enchainements du Monde Animal dans les Temps Geologiques. Fossiles Secondaires. 322 pp. S. Savy, Paris.
Gervais, P. 1859. Zoologie et Paléontologie Françaises, 2ième édition. 544 pp. Arthus Bertrand, Paris.
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics 24: 774–786. Crossref
Haas, G. 1981. A fragmentary skull of Simosaurus (Reptilia: Sauropterygia) from the Middle Triassic of the Makhtesh Ramon, Israel. Israel Journal of Zoology 30: 30–34.
Huene, F. von 1921. Neue Beobachtungen an Simosaurus. Acta Zoologica, Stockholm 1921: 201–239.
Huene, F. von 1948. Simosaurus and Corosaurus. American Journal of Science 246: 41–43. Crossref
Huene, F. von 1956. Paläontologie und Phylogenie der Niederen Tetrapoden. 716 pp. Gustav Fischer, Jena.
Huene, F. von 1959. Simosaurus guilielmi aus dem unteren Mittelkeuper von Obersontheim. Palaeontographica A 113: 180–184.
Jaekel, O. 1905. Über den Schädelbau der Nothosauridae. Sitzungsberichte der Gesellschaft Naturforschender Freunde in Berlin 1905: 60–84.
Jaekel, O. 1910. Über das System der Reptilien. Zoologischer Anzeiger 35: 324–341.
Kear, B.P., Rich, T.H., Vickers-Rich, P., Ali, M.A., Al-Mufarrih, Y.A., Matari, A.H., Masary, A.M., and Halawani, M.A. 2010. A review of aquatic vertebrate remains from the Middle–Upper Triassic Jilh Formation of Saudi Arabia. Proceedings of the Royal Society of Victoria 122: 1–8. Crossref
Kuhn-Schnyder, E. 1961. Der Schädel von Simosaurus. Paläontologische Zeitschrift 35: 95–113. Crossref
Kuhn-Schnyder, E. 1962. La position des nothosauroidés dans le système des reptiles. Colloques Internationaux du Centre National de la Recherche Scientifique 104: 135–144.
Kuhn-Schnyder, E. 1963. Wege der Reptiliensystematik. Paläontologische Zeitschrift 37: 61–87. Crossref
Meyer, H. von 1842. Simosaurus, die Stumpfschnauze, ein Saurier aus dem Muschelkalke von Luneville. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde 1842: 184–197.
Meyer, H. von 1847–1855. Zur Fauna der Vorwelt. Die Saurier des Muschelkalkes mit Rücksicht auf die Saurier aus buntem Sandstein und Keuper. 167 pp. Heinrich Keller, Frankfurt a.M.
Meyer, H. von and Plieninger, T. 1844. Beiträge zur Paläontologie Württembergs. 132 pp. E. Schweizerbart, Stuttgart.
Miguel Chaves, C. de, Pérez-García, A., Cobos, A., Royo-Torres, R., Ortega, F., and Alcalá, L. 2015. A diverse Late Triassic tetrapod fauna from Manzanera (Teruel, Spain). Geobios 48: 479–490. Crossref
Münster, G. 1834. Vorläufige Nachricht über einige neue Reptilien im Muschelkalke von Baiern. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1834: 521–527.
Neenan, J.M., Klein, N., and Scheyer, T.M. 2013. European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia. Nature Communications 4 (1621): 1–7.
Rieppel, O. 1994a. Osteology of Simosaurus gaillardoti and the relationships of stem-group Sauropterygia. Fieldiana: Geology 28: 1–85.
Rieppel, O. 1994b. The braincases of Simosaurus and Nothosaurus: Monophyly of the Nothosauridae (Reptilia: Sauropterygia). Journal of Vertebrate Paleontology 14: 9–23. Crossref
Rieppel, O. 1996. The status of the sauropterygian reptile Partanosaurus zitteli Skuphos from the Middle Triassic of the Austrian Alps, with comments on Microleptosaurus schlosseri Skuphos. Paläontologische Zeitschrift 70: 567–577. Crossref
Rieppel, O. 1998. Corosaurus alcovensis Case and the phylogenetic interrelationships of Triassic stern-group Sauropterygia. Zoological Journal of the Linnean Society 124: 1–41. Crossref
Rieppel, O. 2000. Sauropterygia I. In: P. Wellnhofer (ed.), Handbuch der Paläoherpetologie, 1–134. Verlag Friedrich Pfeil, München.
Rieppel, O., Mazin, J.M., and Tchernov, E. 1999. Sauropterygia from the Middle Triassic of Makhtesh Ramon, Negev, Israel. Fieldiana: Geology 40: 1–85.
Schmidt, M. 1928. Die Lebewelt unserer Trias. 461 pp. F. Rau, Öhringen.
Schmidt, S. 1986. Lokomotion und Lebensweise der Nothosaurier (Diapsida, Sauropterygia, Trias). 102 pp. Ph.D. Dissertation, Eberhard-Karls-University, Tübingen.
Schrammen, A. 1899. Beitrag zur Kenntnis der Nothosauriden des unteren Muschelkalkes in Oberschlesien. Zeitschrift der Deutschen Geologischen Gesellschaft 51: 388–408.
Schultze, H.P. 1970. Über Nothosaurus. Neubeschreibung eines Schädels aus dem Keuper. Senckenbergiana Lethaea 51: 211–237.
Vickers-Rich, P., Rich, T.H., Rieppel, O., Thulborn, R.A., and McClure, H.A. 1999. A Middle Triassic vertebrate fauna from the Jilh Formation, Saudi Arabia. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 213: 201–232. Crossref
Acta Palaeontol. Pol. 63 (2): 315–326.,
2018 https://doi.org/10.4202/app.00471.2018
Appendix 1
Skulls of Simosaurus gaillardoti Meyer, 1842, from the Ladinian (Middle Triassic) of an area corresponding to northeastern France and southwestern Germany, analyzed and figured in this paper. Abbreviations: D, drawing; d, dorsal view; l, lateral view; o, occipital view; P, photograph; v, ventral view.
|
Collection number |
Facies |
Locality |
Comments |
Previous figures |
Figures in this paper |
|
GPIT/RE/1888 |
Lower Keuper |
Obersontheim (Baden-Württemberg, Germany) |
holotype of “Simosaurus guilielmi var. agusticeps” |
Huene 1959 (fig. 1: d, D; pl. 19: d, P); Rieppel 1994a (fig. 6: d, P) |
1U (d, P) |
|
GPIT/RE/09313 |
Upper |
Tiefenbach (Baden-Württemberg) |
|
Huene
1921 (pl. 1: d, D; pl. 2: v, D; pl. 3: 1: l, D; pl. 3:
2: |
1P (d, P), 2R (d, D), 3O (v, P), 4N (v, D), 5G1 (o, P), 5G2 (o, D) |
|
MB.R. 52 |
Upper |
Tiefenbach (Baden-Württemberg) |
|
|
1A (d, P), 2A (d, D), 3U (v, P), 4R (v, D) |
|
MHI 1366 |
Upper |
Künzelsau-Garnberg (Baden-Württemberg) |
|
|
1R (d, P), 2J (d, D), 3M (v, P), 4L (v, D), 5E1 (o, P), 5E2 (o, D) |
|
MHI 1833 |
Upper |
Wilhelmsglück (Baden-Württemberg) |
|
|
1T (d, P), 2S (d, D), 3R (v, P), 4T (v, D), 5M1 (o, P), 5M2 (o, D) |
|
MNHN.F.AC. 9025 |
Upper |
Lunéville |
|
|
1D (d, P), 2D (d, D), 3B (v, P), 4A (v, D) |
|
MNHN.F.AC. 9026 |
Upper |
Lunéville |
cast of a lost |
Gervais 1859 (pl. 56: 1, l, D; fig. 2: o, D; fig. 3: d, D; fig. 3a: v, D); Schrammen 1899 (pl. 24: 2a, b: v, d, D); Kuhn-Schnyder 1961 (fig. 1: v, d, D) |
1C (d, P), 3A (v, P) |
|
MNHN.F.AC. 9028 |
Upper |
Lunéville |
neotype of Simosaurus gaillardoti |
Gaudry 1890 (fig. 288: d, D); Rieppel 2000 (fig. 50: d, P) |
1B (d, P), 2B (d, D) |
|
SMNS 10360 |
Upper |
Neidenfels (Baden-Württemberg) |
|
Jaekel
1905 (fig. 5: d, P); Kuhn-Schnyder 1961
(fig. 2: d, D; fig. 3: o, D; fig. 5: o, P-D; fig. 6: d, D; pl.
9: d, P; pl. 10: 1: o, P); Kuhn-Schnyder 1962
(fig. 1: d, v, D; fig. 2: o, D); Kuhn-Schnyder
1963 (fig. 1a: d, D; fig. 2a: v, D; |
1Q (d, P), 2W (d, D), 3P (v, P),
4O (v, D), |
|
SMNS 11364 |
Upper Muschelkalk |
Neidenfels (Baden-Württemberg) |
currently broken relative to previous figs. in Jaekel (1905, 1910) and Schmidt (1928) |
Jaekel 1905
(fig. 4: d, D; fig. 6: v, D); Jaekel 1910
(fig. 3: v, D); Huene 1921 (fig. 10: d, D;
fig. 13: v, P); Schmidt 1928 |
1K (d, P), 2O (d, D), 3H (v, P), 4H (v, D) |
|
SMNS 11364b |
Upper |
Neidenfels (Baden-Württemberg) |
counterplate of SMNS 11364 |
Huene 1921 (fig. 11: v, D; fig. 12: v, P) |
3V (v, P), 4U (v, D) |
|
SMNS 16363 |
Upper |
Murr |
|
|
1V (d, P), 2Q (d, D), 3Q (v, P),
4P (v, D), |
|
SMNS 16639 |
Upper |
Tiefenbach (Baden-Württemberg) |
|
|
1F (d, P), 2E (d, D), 3D (v, P), 4C (v, D) |
|
SMNS 16700 |
Lower Keuper |
Hoheneck (Baden-Württemberg) |
holotype of “Simosaurus guilielmi” |
Meyer and Plieninger 1844 (pl. 11: 1: d, D); Meyer 1847–1855 (pl. 20: 1: d, D); Huene 1921 (fig. 14: d, D); Schmidt 1928 (fig. 1135: d, D); Rieppel 1994a (fig. 4: d, P; fig. 5: d, D) |
1I (d, P), 2M (d, D), 3G (v, P), 4F (v, D) |
|
SMNS 16735a |
Upper |
Tiefenbach (Baden-Württemberg) |
|
|
1N (d, P), 2I (d, D), 3J (v, P), 4K (v, D) |
|
SMNS 16767 |
Upper |
Tiefenbach (Baden-Württemberg) |
|
|
1G (d, P), 2G (d, D), 3E (v, P),
4D (v, D), |
|
SMNS 18220 |
Upper |
Heldenmühle (Baden-Württemberg) |
|
|
1O (d, P), 2H (d, D) |
|
SMNS 18274 |
Upper |
Heldenmühle (Baden-Württemberg) |
|
|
1E (d, P), 2F (d, D), 3C (v, P),
4B (v, D), |
|
SMNS 18520 |
Upper |
Heldenmühle (Baden-Württemberg) |
|
|
1H (d, P), 2C (d, D), 3F (v, P), 4E (v, D) |
|
SMNS 18550 |
Upper |
Heldenmühle (Baden-Württemberg) |
|
|
1M (d, P), 2N (d, D), 3I (v, P), 4J (v, D) |
|
SMNS 18637 |
Upper |
Heldenmühle (Baden-Württemberg) |
|
Huene 1921 (fig. 1: d, D; fig. 3: v, D; fig. 4: o, D) |
1W (d, P), 2T (d, D), 3T (v, P),
4Q (v, D), |
|
SMNS 50714 |
Upper |
Schmalfelden (Baden-Württemberg) |
|
Schmidt 1986
(fig. 51: l, d, v, D); Rieppel 1994b |
1S (d, P), 2P (d, D), 3N (v, P),
4M (v, D), |
|
SMNS 56288 |
Upper |
Markgröningen (Baden-Württemberg) |
|
|
1J (d, P), 2L (d, D), 3K (v, P),
4G (v, D), |
|
SMNS 59366 |
Upper |
Rüblingen (Baden-Württemberg) |
|
|
1L (d, P), 2K (d, D), 3L (v, P),
4I (v, D), |
|
SMNS 59943 |
Upper |
Crailsheim (Baden-Württemberg) |
|
Fraas 1896 (pl. 3: 2: d, P) |
1Y (d, P), 2V (d, D), 5J1 (o, P), 5J2 (o, D) |
|
SMNS 50715 |
Upper |
Rüblingen (Baden-Württemberg) |
|
|
1X (d, P), 2U (d, D), 3S (v, P),
4S (v, D), |