
Tanystropheid archosauromorphs in the Lower Triassic of Gondwana
TIANE MACEDO DE OLIVEIRA, DANIEL OLIVEIRA, CESAR L. SCHULTZ, LEONARDO KERBER, and FELIPE L. PINHEIRO
De Oliveira, T.M., Oliveira, D., Schultz, C.L., Kerber, L., and Pinheiro, F.L. 2018. Tanystropheid archosauromorphs in the Lower Triassic of Gondwana. Acta Palaeontologica Polonica 63 (4): 713–723.
Tanystropheidae is a clade of early archosauromorphs with a reported distribution ranging from the Early to the Late Triassic of Asia, Europe, and North America. Although some specimens with possible tanystropheid affinities from the Lower Triassic beds of Brazil have been previously attributed to “Protorosauria”, little is known about the tanystropheid record in Gondwana. Here, two new and one previously reported specimen from the Sanga do Cabral Formation (Induan–Olenekian) of Brazil are described and interpreted as ?Tanystropheidae. These records, together with other tetrapods previously reported for the Sanga do Cabral Formation, increase the knowledge of the biotic diversification during the beginning of the Triassic. This contribution reinforces that the archosauromorph diversification occurred shortly after the Permo-Triassic extinction, making the Sanga do Cabral Formation an important unit for the study of early Mesozoic faunas.
Key words: Diapsida, Archosauromorpha, Tanystropheidae, Triassic, Sanga do Cabral Formation, Brazil.
Tiane Macedo de Oliveira [tiane.m.deoliveira@gmail.com], Programa de Pós-Graduação em Biodiversidade Animal, Universidade Federal de Santa Maria (UFSM), Prédio 13, Av. Roraima, 1000, 97105-900, Bairro Camobi, Santa Maria, Rio Grande do Sul, Brazil.
Felipe L. Pinheiro [felipepinheiro@unipampa.edu.br] and Daniel Oliveira [doliveira.simao@gmail.com], Laboratório de Paleobiologia, Universidade Federal do Pampa (UNIPAMPA), Av. Antônio Trilha, 1847, São Clemente, 97300-000, São Gabriel, Rio Grande do Sul, Brazil.
Cesar Leandro Schultz [cesar.schultz@ufrgs.br], Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, Universidade Federal do Rio Grande do Sul (UFRGS), Av. Bento Gonçalves, 9500, 91540-000, Porto Alegre, Rio Grande do Sul, Brazil.
Leonardo Kerber [leonardokerber@gmail.com], Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia (CAPPA), Universidade Federal de Santa Maria (UFSM), Rua Maximiliano Vizzotto, 598, 97230-000, São João do Polêsine, Rio Grande do Sul, Brazil.
Received 17 April 2018, accepted 8 October 2018, available online 19 November 2018.
Copyright © 2018 T.M. de Oliveira et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Archosauromorphs were one of the first diapsid groups to diversify in body size and morphological disparity in Triassic terrestrial and marine ecosystems from Pangea. The wide radiation of Archosauromorpha resulted in adaptations to different environments and the development of distinct body plans. In sum, these groups include cursorial, semiaquatic, marine and flying forms (e.g., Nesbitt 2011; Ezcurra et al. 2014; Ezcurra 2016; Foth et al. 2016; Pinheiro et al. 2016). Bizarre body plans acquired by archosauromorphs include long-necked presumably aquatic forms (Tanystropheidae), relatively large terrestrial herbivores, sometimes with skull ornaments (Allokotosauria), and highly specialized herbivores (Rhynchosauria) (Nesbitt 2011; Jaquier and Scheier 2017; Sengupta et al. 2017).
Several Permian–Triassic long-necked archosauromorphs of varied sizes have historically been attributed to the “Prolacertiformes” (e.g., Protorosaurus Meyer, 1830, Macrocnemus Nopcsa, 1930, Tanystropheus Wild, 1973, Prolacerta Parrington, 1935) in several taxonomic proposals (e.g., Camp 1945; Evans 1988). However, recent phylogenetic analyses recovered most of the representatives of this group in a widely polyphyletic arrangement, distributed among different lineages of archosauromorphs and early Diapsida (Dilkes 1998; Sues 2003; Modesto and Sues 2004; Senter 2004; Ezcurra et al. 2014; Pritchard et al. 2015; Ezcurra 2016; Pinheiro et al. 2016; Pritchard and Nesbitt 2017). The only classical “prolacertiform” representatives recovered in these analyses as a clade are placed within Tanystropheidae, represented for example by the well-known taxa Tanystropheus longobardicus Bassani, 1886 and Macrocnemus bassanii Nopcsa, 1930, usually used as terminal taxa in phylogenetic analyses.
Tanystropheidae is regarded as a node-based clade, defined as the most recent common ancestor of Macrocnemus, Tanystropheus, Langobardisaurus Renesto, 1994, and all of its descendants (Dilkes 1998), with records in the Early–Late Triassic of Asia, Europe, and North America (Rieppel 1989; Renesto 2005; Dalla Vecchia 2006; Li et al. 2007; Jiang et al. 2011; Pritchard et al. 2015). Fossils with tanystropheid affinities are poorly known in South America and restricted to a few isolated bones attributed to “Protorosauria” (Langer and Schultz 1997; and Dias-da-Silva 1998).
The Middle Triassic taxa Tanystropheus and Macrocnemus have elongated cervical vertebrae, a notably pronounced feature of this group (Ezcurra 2016; Jaquier and Scheier 2017). The body morphology of Macrocnemus is in accordance with a terrestrial habit (Rieppel 1989; Renesto 2005). In contrast, the lifestyle of Tanystropheus is still debatable. However, recent osteological analyses do not support a fully aquatic habit for this animal (Jaquier and Scheier 2017).
The Lower Triassic archosauromorph record is notably scarce, what hinders a satisfactory understanding of one of the most important radiation events of this clade. In this context, the South American Sanga do Cabral, and Buena Vista Formations from Brazil and Uruguay recently contributed with important specimens (Ezcurra et al. 2015; Pinheiro et al. 2016). In this work, we describe isolated archosauromorph vertebrae from the Sanga do Cabral Formation and analyze their phylogenetic relationships. One of the specimens (UFRGS-PV-492-T) was briefly presented by Dias-da-Silva (1998), and attributed to “Protorosauria”. Based on comparative anatomy and phylogenetic protocol, we attribute the vertebrae here presented to ?Tanystropheidae. As most tanystropheids come from North America, Europe, and Asia, the Sanga do Cabral Formation specimens provide valuable information on the geographic distribution of this group.
Institutional abbreviations.—LASEPE, Laboratório de Sedimentologia e Petrologia; PUCRS, Pontifícia Universidade Católica do Rio Grande do Sul, Brazil; UFRGS, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil; UNIPAMPA, Universidade Federal do Pampa, São Gabriel, Brazil; ZAR, Muséum national d’Histoire naturelle (Zarzaitine collection), Paris, France.
Material and methods
The specimens described here belong to the paleontological collections of the Universidade Federal do Rio Grande do Sul (UFRGS-PV-492-T and UFRGS-PV-647-T) and Universidade Federal do Pampa (UNIPAMPA 733). To test the phylogenetic affinities of the new specimens, these were scored in the archosauromorph morphological character matrix proposed by Sengupta et al. (2017) which, by its turn, is a modification of Ezcurra (2016) database. After the addition of the new operational taxonomic units using the software Mesquite 3.2 (Maddison and Maddison 2015), the analyses were conducted using the software TNT 1.5 (Goloboff and Catalano 2016). Analyses were made using both reduced and complete matrixes of Sengupta et al. (2017). The reduced matrix of Sengupta et al. (2017), excludes problematic taxa (e.g., taxa with uncertain positioning due to a large amount of missing data), in accordance to Ezcurra (2016)(see also SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app63-Oliveira_etal_SOM.pdf). A first analysis was conducted using the New Technology Algorithms until the best result was hit 100 times and, subsequently, running heuristic searches of 1000 replicates using random addition sequences followed by the TBR (Tree Bisection Reconnection) branch rearrangement algorithm, retaining ten trees per replicate. If some replications overflowed (some of the MPT’s may not have been found) during the first round of analysis, a traditional search was again employed, this time using trees from RAM. The following experiments were performed: (i) analysis of the three cervical vertebrae individually (UNIPAMPA 733, UFRGS-PV-492-T, and UFRGS-PV-647-T); (ii) analysis of the specimens (UNIPAMPA 733 and UFRGS-PV-492-T) together.
Additionally, we used micro-CT scan to analyze the specimens UNIPAMPA 733, UFRGS-PV-492-T, and UFRGS-PV-647-T, and generate tridimensional models. The images were obtained at the Laboratório de Sedimentologia e Petrologia (LASEPE), Pontifícia Universidade Católica do Rio Grande do Sul (PUCRS), Brazil, through a micro-CT scanner Skyscan 1173, using a voltage of 75 kV and amperage of 106 µa. A total of 1723 (1551 slices with information) slices were generated. The image pixel size is 15.166192 µm. 3D Slicer 4.8 (Fedorov et al. 2012) was employed to generate 3D models of this specimens.
Geological setting
The cervical vertebrae UNIPAMPA 733 and UFRGS-PV-492-T come from the Ramal Abandonado locality (municipality of Dilermando de Aguiar). The fossiliferous levels of this outcrop are composed of an intercalation of fine sandstone and intraformational conglomerate, in which argillaceous intraclasts and carbonate nodules are frequent (Zerfass et al. 2003; Da-Rosa et al. 2009; Dias-Da-Silva et al. 2017).
Specimen UFRGS-PV-647-T has an indeterminate provenance, but, according to collection records, was collected from a Sanga do Cabral Formation outcrop.
The Sanga do Cabral Formation (Induan–early Olenekian), was deposited in a vast alluvial plain in southwestern Gondwana (Zerfass et al. 2003). The Brazilian Sanga do Cabral Formation, together with the Buena Vista Formation (Uruguay), constitutes the Sanga do Cabral Supersequence. This unit also includes occasional well-selected fine to medium sandstones with massive cross-bedding stratifications, interpreted as aeolian in origin and indicating constant reworking of the alluvial plain (Faccini 1989; Dias-Da-Silva et al. 2017).
The fossil assemblage of the Sanga do Cabral Formation so far includes procolophonids, temnospondyls, and archosauromorphs (Dias-Da-Silva et al. 2005, 2006; Pinheiro et al. 2016). Vertebrate fossils are often found isolated and disarticulated. This preservation mode suggests extensive exposure and post-mortem transport of bones during the biostratinomic phase, and subsequent reworking after diagenesis. Archosauromorph records are scarce in the Sanga do Cabral Formation, with only a few specimens reported so far (e.g., Langer and Schultz 1997; Dias-Da-Silva 1998; Da-Rosa et al. 2009; Pinheiro et al. 2016; Dias-Da-Silva et al. 2017). In addition to the scarcity of fossils, the lack of anatomical studies focusing on small isolated specimens contributes to the limited knowledge about the diversity of archosauromorphs in this sedimentary unit. Articulated specimens remain almost unknown, except for a few records (e.g., Pinheiro et al. 2016).
An Induan–Olenekian age (251–247Ma) (Cohen et al. 2013) is inferred for this formation based on the presence of the parareptile Procolophon trigoniceps Owen, 1876, and comparisons with the Lystrosaurus Assemblage Zone of the South African Karoo (Dias-Da-Silva et al. 2006; Botha and Smith 2006; Da-Rosa et al. 2009). Currently, no fossil taxa indicate a Permian age for the Sanga do Cabral Supersequence (see Dias-Da-Silva et al. 2017 for a comprehensive revision), and all the biostratigraphically informative fossils so far collected from the Sanga do Cabral and Buena Vista formations reinforce the attribution of a Lower Triassic age for the Sanga do Cabral Supersequence (Dias-Da-Silva et al. 2017). The age of the Sanga do Cabral and Buena Vista formations, lithostratigraphically equivalent, has been the focus of numerous discussions, being alternatively considered as Permian or a broader Permo-Triassic age (Piñeiro et al. 2004, 2007; Ezcurra et al. 2015). According to Piñeiro et al. (2015), at least part of the Sanga do Cabral Formation is Permian in age, but a comprehensive revision provided by Dias-da-Silva et al. (2017) supported an Early Triassic age for the unit.
Systematic palaeontology
Diapsida Osborn, 1903 (sensu Laurin 1991)
Archosauromorpha Huene, 1946 (sensu Gauthier et al. 1988)
Tanystropheidae Gervais, 1859
?Tanystropheidae indet.
Material.—UNIPAMPA 733, UFRGS-PV-492-T, UFRGS-PV-647-T: isolated cervical vertebrae from Sanga do Cabral Formation (Induan–early Olenekian).
Description.—UNIPAMPA 733 (Fig. 1) presents an elongated and amphicoelous centrum. The parapophyses and diapophyses are poorly developed and situated in different processes and near each other in the centrum. In lateral view, the anterior articular surface of UNIPAMPA 733 is raised dorsally relative to the posterior surface. The neural spine is dorsoventrally low and laminar, lacking a transverse expansion (spine-table) and presenting a post-spinal fossa. The neural spine is abraded in its anterior and posterior extremities, preventing the recognition of a possible “overhang” (sensu Ezcurra 2016). A faint longitudinal keel runs the length of the centrum. In ventral view, UNIPAMPA 733 shows a thin anteroposterior keel extending through the whole centrum. The specimen has a total length of 18.45 mm.
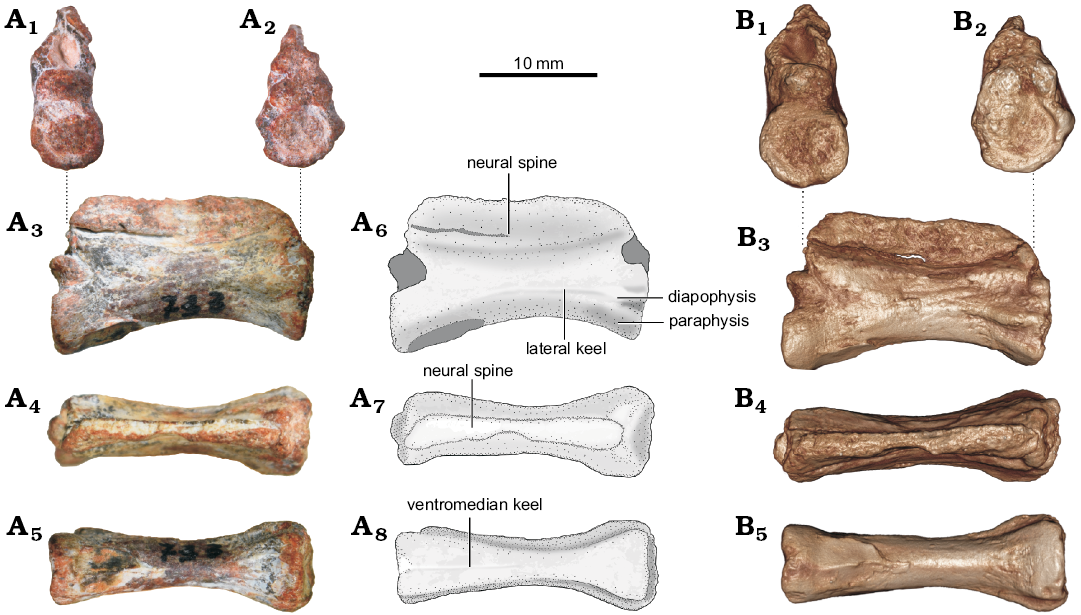
Fig. 1. Cervical vertebra (UNIPAMPA 733) of ?Tanystropheidae indet. from Sanga do Cabral Formation (Induan–early Olenekian). A. Photographs (A1–A5) and explanatory drawings (A6–A8). B. Virtual 3D reconstructions (see SOM 2). In posterior (A1, B1), anterior (A2, B2), right lateral (A3, A6, B3), dorsal (A4, A7, B4), and ventral (A5, A8, B5) views.
Among the vertebrae, UFRGS-PV-492-T (Fig. 2) is better preserved. However, this specimen has a broken neural spine, making it impossible to analyze the shape of this structure. The preserved portion of the neural spine, however, is reminiscent of a thin and low longitudinal lamina, possibly extending for almost the entire anteroposterior length of the centrum. With a total length of 20.26 mm, UFRGS-PV-492-T shows an elongated amphicoelous centrum and a higher anterior articular surface with relation to the posterior one. The diapophyses and parapophyses are close to each other on the anterior surface of the centrum. Notably, this specimen displays a clear longitudinal keel that extends laterally on the centrum. The ventral surface of the centrum shows a keel that extends along its entire length. Although a ventral keel is also present in specimens UNIPAMPA 733 and UFRGS-PV-647-T, they are not as well preserved as in UFRGS-PV-492-T. The ventral surface of the centrum (excluding the keel) is slightly rounded. In lateral view, the postzygapophysis of UFRGS-PV-492-T is slightly elevated dorsally above the level of the prezygapophysis, with short and spaced articular surfaces. Due to the preservation, it is not possible to evaluate the presence of epipophyses or the transpostzygapophyseal lamina.
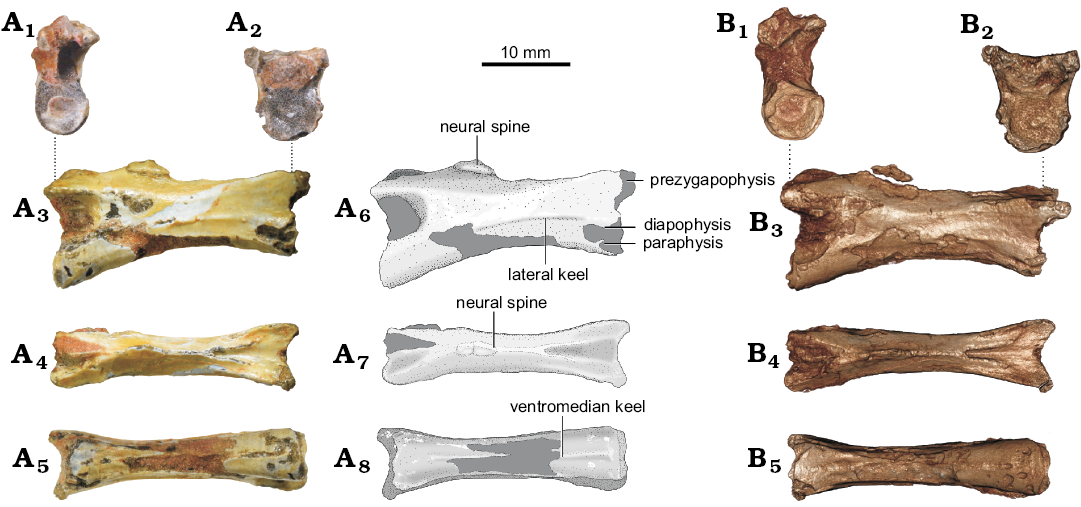
Fig. 2. Cervical vertebra (UFRGS-PV-492-T) of of ?Tanystropheidae indet. from Sanga do Cabral Formation (Induan–early Olenekian). A. Photographs (A1–A5) and explanatory drawings (A6–A8). B. Virtual 3D reconstructions (see SOM 2). In posterior (A1, B1), anterior (A2, B2), right lateral (A3, A6, B3), dorsal (A4, A7, B4), and ventral (A5, A8, B5) views.
The preserved portion of the neural spine of UFRGS-PV-647-T (Fig. 3) indicates a dorsoventrally deep lamina. The vertebral body is elongated, and the centrum is amphicoelous. Due to poor preservation, the parapophyses and diapophyses are not readily distinguishable. However, these structures seem to be poorly developed. It is also not possible to determine if the postzygapophyses are separated or connected to each other. In lateral view, UFRGS-PV-647-T shows the anterior articular surface of the centrum raised dorsally relative to the posterior one. The centrum also possesses a lateral transverse keel. This specimen shows a ventral keel that does not reach the anterior articular margin. Specimen UFRGS-PV-647-T has a total length of 15.40 mm.
In all the vertebrae, it is not possible to determine whether the neural spine ends anteriorly or posteriorly to the postzygapophyses.
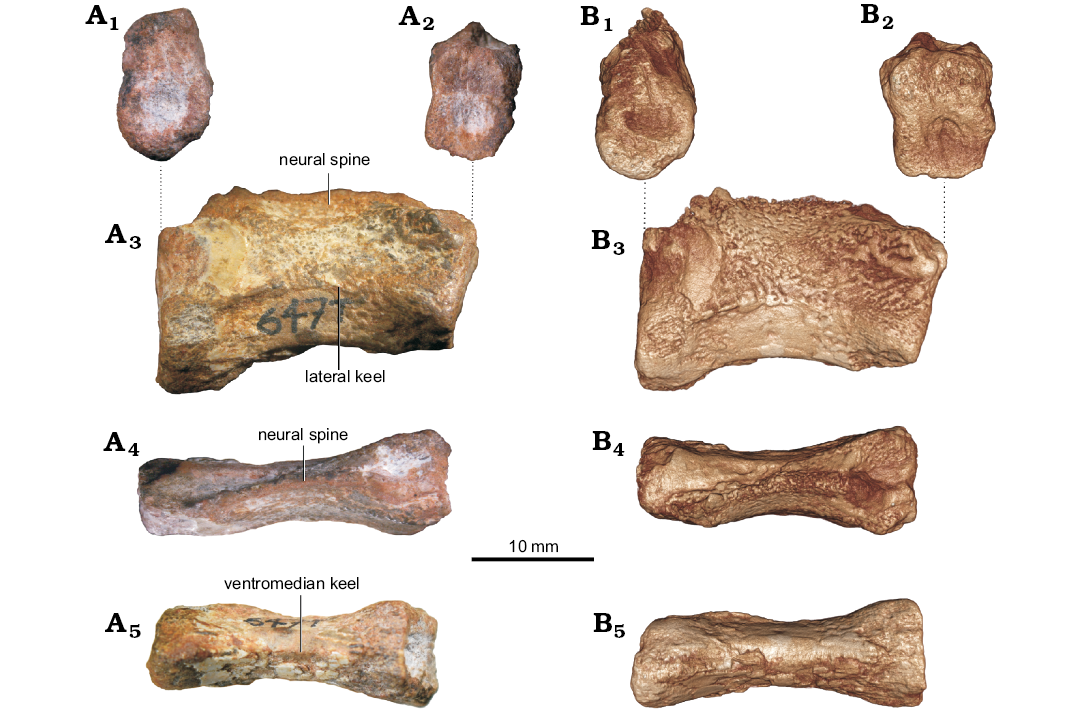
Fig. 3. Cervical vertebra (UFRGS-PV-647-T) of ?Tanystropheidae indet. from Sanga do Cabral Formation (Induan–early Olenekian). A. Photographs. B. Virtual 3D reconstructions (see SOM 2). In posterior (A1, B1), anterior (A2, B2), right lateral (A3, B3), dorsal (A4, B4), and ventral (A5, B5) views.
Phylogenetic analyses
First of all, the cervical vertebrae were evaluated individually using the reduced matrix of Sengupta et al. (2017). The analysis including UNIPAMPA 733 resulted in this specimen as the sister taxon to Tanystropheidae (Macrocnemus bassanii and (Tanystropheus longobardicus + Amotosaurus rotfeldensis Fraser and Rieppel, 2006)) (SOM 3: fig. 1). The clade formed by UNIPAMPA 733 and Tanystropheidae has Jesairosaurus lehmani Jalil, 1997 as sister taxon.
In the analysis including UFRGS-PV-492-T, this specimen occupies the same position as UNIPAMPA 733 in the previous analysis (SOM 3: fig. 2). These two analyses generated one tree with 2783 steps, consistency index 0.291 and retention index 0.633. Phylogenetic analysis including UFRGS-PV-647-T resulted in a strict consensus with low resolution and a large number of polytomies. This analysis generated six trees, with 2783 steps, retention index 0.633 and consistency index 0.291. The low resolution presented in the strict consensus can be justified by the lack of informative morphological characters, reflecting poor preservation of relevant structures. In the analysis including both UFRGS-PV-492-T and UNIPAMPA 733, these specimens were recovered into a polytomy together with Jesairosaurus lehmani, Macrocnemus bassanii and (Tanystropheus longobardicus + Amotosaurus rotfeldensis) (Fig. 4). This analysis generated three trees with 2783 steps, consistency index 0.291 and retention index 0.633. This last analysis did not include specimen UFRGS-PV-647-T due to the lack of informative character-states.

Fig. 4. Strict consensus of the analysis including UNIPAMPA 733 and UFRGS-PV-492-T (analysis ii) based on the reduced matrix of Sengupta et al. (2017).
Discussion
Anatomical and taxonomic remarks.—Most of the Tanystropheidae fossil record is concentrated in Europe, Asia, and North America. Isolated bones assigned to “Protorosauria” were briefly presented for Lower Triassic deposits of Brazil (Langer and Schultz 1997; Dias-Da-Silva 1998) and were so far the only records of the group in South America. The specimen UFRGS-PV-492-T was originally described as a “protorosaurid” cervical vertebra (Dias-Da-Silva 1998), a taxonomical attribution mostly based on its elongated amphicoelous centrum and a laminar neural spine.
The three vertebrae here described, UFRGS-PV-492-T, UFRGS-PV-647-T, and UNIPAMPA 733 are morphologically compatible with the Tanystropheidae. These materials resemble this group in displaying a dorsoventrally short and laminar neural spine (Ezcurra 2016). Also, it is possible to distinguish a laterodorsal keel directed to the pre- and postzygapophysis throughout the length of the centra of all specimens, which, according to Wild (1973) and Nosotti (2007), is diagnostic for Tanystropheidae.
The cervical vertebrae have the anterior articular surface of the centrum elevated dorsally relative to the posterior one. In contrast, the anterior and posterior surfaces of non-archosauromorph diapsids and some archosauriforms (Proterochampsa barrionuevoi Dilkes and Arcucci, 2012; Riojasuchus tenuisceps Bonaparte, 1967) are located at the same dorsoventral level in lateral view. As a result, the neck is mainly straight in these latter taxa, instead of sigmoidal as in most archosauromorphs (Ezcurra 2016).
According to Jalil (1997) and Ezcurra (2016) anteroposteriorly long and dorsoventrally low cervical neural spines are present in putative “prolacertiforms” (e.g., Protorosaurus speneri, Amotosaurus rotfeldensis, Macrocnemus bassanii, Tanystropheus longobardicus, Prolacerta broomi, Boreopricea funerea), being also a present in other archosauromorphs, such as Trilophosaurus (Gregory, 1945) and Azendohsaurus (Flynn, Nesbitt, Parrish, Ranivoharimanana, and Wyss, 2010), whereas most non-archosaurian archosauriforms possess dorsoventrally taller cervical neural spines (e.g., Proterosuchus fergusi Broom, 1903, Proterosuchus alexanderi Hoffman, 1965), Garjainia prima Ochev, 1958, Euparkeria capensis Broom, 1913).
Following recent phylogenetic proposals (Ezcurra 2016), the sister-taxon to Tanystropheidae is Jesairosaurus lehmani, described based on cranial and postcranial remains from the Zarzaitine Series (Olenekian–Anisian) of Algeria. As described above, our phylogenetic analysis with the inclusion of both UFRGS-PV-492-T and UNIPAMPA 733 in the data matrix of Sengupta et al. (2017) resulted in a polytomy including the new specimens, J. lehmani, and tanystropheid taxa. Although the cervical vertebrae of the holotype of J. lehmani (ZAR 06) are well preserved, they are still partially covered by matrix. However, is possible to see that they differ from the material described herein in several characters. In ZAR 06 all preserved cervical neural spines are anterodorsally oriented, being the posteriormost cervical neural arches low, narrow, and slightly concave laterally (Jalil 1997; Ezcurra 2016). The vertebrae described herein are significantly more elongated than those of ZAR 06. In ZAR 06, the presence of lateral keels in the cervical centra cannot be determined. In addition, our material shows dorsoventrally lower neural spines than those of J. lehmani.
Comparisons with Macrocnemus and Tanystropheus demonstrate that the pronounced elongation of the vertebral body, as well as the presence of lateral and ventral keels and a low neural spine present in the referred Sanga do Cabral Formation specimens, are, together, typical features of tanystropheid cervicals. This same combination of features is present not only in the well-known Macrocnemus bassanii and Tanystropheus longobardicus but in all known Tanystropheidae (e.g., Nosotti 2007; Jaquier and Scheier 2017).
Phylogenetic remarks.—In recent studies, Tanystropheidae and Jesairosaurus lehmani (previously identified as belonging to “Protorosauria”/”Prolacertiformes”) were recovered as more related to each other than to other archosauromorphs and as the sister-taxa to Crocopoda (Ezcurra 2016). The unnamed clade formed by Jesairosaurus + Tanystropheidae has a distribution from the Early Triassic (e.g., Augustaburiania vatagini Sennikov, 2011) to the Norian (Late Triassic) (e.g., Tanytrachelos ahynis Fraser, Grimaldi, and Olsen, 1996) (Ezcurra 2016).
Based on the current dataset, the results of our analyses including both UNIPAMPA 733 and UFRGS-PV-492-T provide phylogenetic support for the attribution of the new material to Archosauromorpha more closely related to Tanystropheidae than to other archosauromorphs. The individual analysis of UNIPAMPA 733 and UFRGS-PV-492-T recovered these specimens as a sister taxon of Tanystropheidae (Macrocnemus and [Tanystropheus + Amotosaurus]). However, the specimens share some characters with tanystropheids that are not present in Jesairosaurus (see above). Both UNIPAMPA 733 and UFRGS-PV-492-T have morphologies similar to what is recognized as the standard tanystropheid pattern. For instance, these specimens exhibit elongated centra, neural spine consisting of a thin lamina and the presence of a lateral keel, while Jesairosaurus lehmani presents the posteriormost cervical neural arches low, narrow, slightly concave laterally and the neural spine substantially higher than what is displayed by the material we describe. Hence, although the specimens are not placed within the node-based clade Tanystropheidae, they are more similar to tanystropheids than to J. lehmani, and the placement of UNIPAMPA 733 and UFRGS-PV-492-T outside Tanystropheidae may reflect the low number of phylogenetic characters present in a single bone element.
Paleobiogeographic relevance.—As a consequence of the mass extinction that marks the Permian/Triassic boundary, the continental communities at the beginning of the Triassic were remarkably depleted, and the diversity was represented only by few, mainly unspecialized tetrapod taxa (Benton 2003, 2016; Sennikov 2011; Irmis and Whiteside 2011). In archosauriforms, for example, the morphological disparity remained low and did not expand much until the late Olenekian (late Early Triassic) to early Anisian (early Middle Triassic), when erythrosuchids, Euparkeria and poposauroid pseudosuchians are recorded for the first time (Gower and Sennikov 2000; Butler et al. 2011; Ezcurra and Butler 2015). During the Early Triassic, the diversity of the terrestrial biota was gradually restored. The tetrapod fauna already documented in the Sanga do Cabral Formation is composed of procolophonoids, temnospondyls, and archosauromorphs (including the tanystropheids described herein). Like most tetrapods in the Early Triassic, ?Tanystropheidae representatives were smaller in size than their younger representatives. The Sanga do Cabral fauna provide evidence for a relatively complex community that lived in western Gondwana during the Early Triassic.
The oldest archosauromorph from South America is represented by a distal portion of a left humerus (UFRGS-PV-0546-P) from the Guadalupian (mid-Permian) Rio do Rasto Formation (Paraná Basin) (Martinelli et al. 2016). Although the humerus does not possess any particular apomorphy of any archosauromorph group, the overall pattern of this specimen more closely resembles tanystropheids than other archosauromorphs (Martinelli et al. 2016). Although the morphology of the humerus described by Martinelli et al. (2016) indeed resembles Tanystropheidae, the fragmentary nature of this specimen precludes a categorical attribution.
Before the end of the Early Triassic, the archosauromorphs were already anatomically diversified, including semi-aquatic or entirely aquatic forms such as the Tanystropheidae (Renesto 2005; Benton 2016; Ezcurra 2016). The oldest representatives of the group traditionally called “Protorosauria” were probably terrestrials and include the enigmatic Aenigmastropheus parringtoni Ezcurra, Scheyer, and Butler, 2014 from Permian of Tanzania and the well-known Protorosaurus speneri Meyer, 1832 from middle–late Permian of Germany and England (Ezcurra 2016). According to recent contributions (Nesbitt et al. 2015; Pritchard et al. 2015; Ezcurra 2016), Tanystropheidae include Macrocnemus bassanii and Tanystropheus longobardicus from the Middle Triassic of Switzerland and Italy, Macrocnemus fuyuanensis Li, Zhao, and Wang, 2007 from the Middle Triassic of China (Li et al. 2007; Jiang et al. 2011), Langobardisaurus pandolfii from the Late Triassic of Italy and Austria (Renesto and Dalla Vecchia 2007; Saller et al. 2013), Tanytrachelos ahynis Olsen, 1979 from the Late Triassic of the USA, Amotosaurus rotfeldensis Fraser and Rieppel, 2006 from the Middle Triassic of Germany and Augustaburiania from the Early Triassic of Russia (Sennikov 2011) (Fig. 5). The earliest fossils of known Tanystropheidae (e.g., Amotosaurus Fraser and Rieppel, 2006) have been recovered from the northwest margin of Tethys Province, while Jesairosaurus lehmani, the sister-taxon to the Tanystropheidae (Ezcurra, 2016), comes from Algeria with distribution from the Olenekian to the Anisian (Jalil 1997).
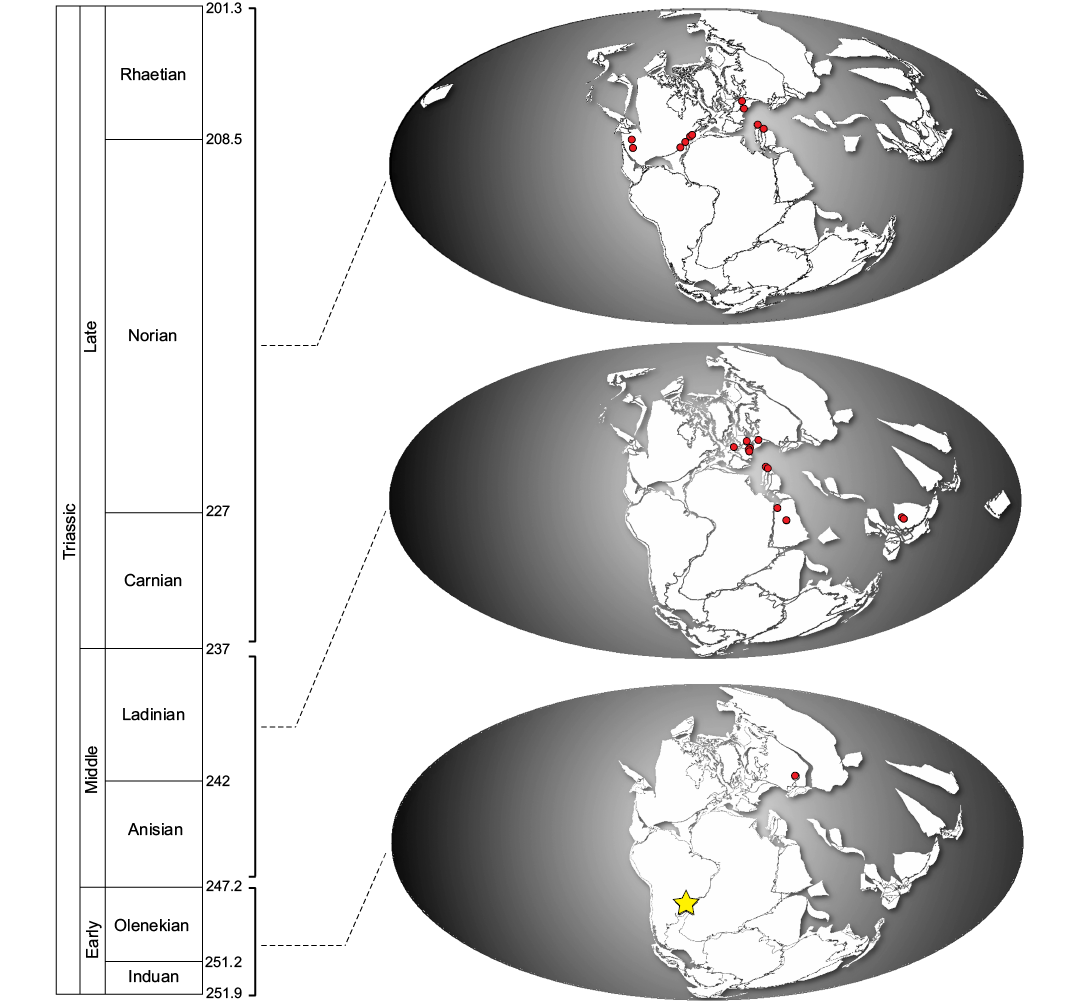
Fig. 5. Geographic distribution maps for Tanystropheidae through time (circles) and the brazilian fossil record (star) (data from the Paleobiology Database, https://paleobiodb.org/#/) and timescale from Gradstein et al. (2012).
According to previous paleobiogeographic scenarios (e.g., Pritchard et al. 2015; Jaquier and Scheier 2017), it is proposed that tanystropheids diversified from their Permian and Early Triassic ancestors during the Early Triassic, with an initial dispersion along the western margin of the Tethys ocean, while the dispersion to what is now North America began only sometime later, during the Middle Triassic. The record of ?Tanystropheidae indet. in South America indicates an initial dispersion in a central region of Pangaea, following to the west of Gondwana during the Early Triassic and, later, to the region that is currently North America, becoming abundant during the Middle Triassic.
The first and sparse records of Tanystropheidae dates back to the Early Triassic, and the most significant diversity peak of this clade occurs only during the Middle/Late Triassic. The presence of fossils related to this clade in Lower Triassic strata suggests that it had a substantial evolutionary history right after the mass extinction at the end of the Permian, with an initial diversification that could date to even earlier than the Triassic.
The fossils here studied significantly expands the geographical range of these organisms to the West Gondwana but is in accordance with the Early Triassic paleogeography. The arrangement of continents during that time allowed the dispersion of terrestrial animals (e.g., Ezcurra 2010), making possible a cosmopolitan distribution of several groups (Button et al. 2017).
The Sanga do Cabral Formation was deposited in a system of ephemeral, high-energy river channels in broad and extensive alluvial plains, containing a rich assemblage of terrestrial and aquatic tetrapods (Holz and Souto-Ribeiro 2000; Da-Rosa et al. 2009). The presence of amphibians indicates shorter and more humid phases and the associated growth of lakes and perennial ponds in the floodplains (Zerfass et al. 2003). Most tanystropheids were probably specialized to live in coastal environments and margins of aquatic ponds, although the considerable continental record of the clade indicate that some taxa were terrestrial. This is probably the reason why they reached a diversity peak in the Middle Triassic epicontinental marine basin of Central Europe, with shallow banks, indented coastline, many islands, gulfs, and lagoons, as well as abundant marine biota (Sennikov 2011). It should, however, be also taken into account that the best-preserved fossils come from marine deposits of Central Europe and Eastern China, and the fragmentary record from other localities may be hiding a considerable diversity in terrestrial environments.
Conclusions
The morphology of the specimens here studied resembles more Tanystropheidae than Jesairosaurus and other archosauromorphs (see Discussion). On the other hand, phylogenetic analyses of these specimens were not able to unambiguously recover them inside Tanystropheidae (SOM 3: figs. 1, 2), what is possibly related to the scarcity of phylogenetic characters present in isolated bones. Hence, although the specimens display a combination of features typical of Tanystropheidae, we considered then as ?Tanystropheidae indet., depending on further findings to solve this question.
This study contributes to the understanding of the diversity of archosauromorphs after the Permo-Triassic extinction in western Gondwana. Considering that most representatives of Tanystropheidae are known from Middle and Upper Triassic strata, the material here described represents some of the oldest fossils that can be related to Tanystropheidae, suggesting a wide distribution of such forms during the Early Triassic. The fossil record found in the Sanga do Cabral Formation is mostly represented by isolated and fragmented bones. The lack of anatomical and phylogenetic analysis of these fossils contributes to limited knowledge on the archosauromorph diversity of this sedimentary unit and consequently about the diversification of these animals after the greatest extinction ever recorded.
Acknowledgements
For granting access to relevant fossil collections, the authors are indebted to Christian Klug (Paläontologisches Institut und Museum, Universität Zürich, Germany); Rainer Schoch (Naturkunde Museum Stuttgart, Germany); Oliver Rauhut and Markus Moser (both Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany); Mark Norell and Carl Mehling (both American Museum of Natural History, New York, USA); Sandra Chapman and Lorna Steel (both Natural History Museum, London, UK). We also thank Jaqueline Figueiredo (UNIPAMPA) for recovering one of the specimens in the field and Voltaire Neto (UFRGS) for critical reading an early version of this manuscript. Flavio Pretto, Eduardo Neves, and Rodrigo Müller (all Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia, Rio Grande do Sul, Brazil) made useful comments on figures and in the methodology employed herein. Reviews by Adam Pritchard (Smithsonian Institution, National Museum of Natural History, Washington, DC) and Martin Ezcurra (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) greatly improved the final paper. TMO and DO are supported by grants from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). LK is supported by a Fundação de Amparo à Pesquisa do Estado do RS (FAPERGS) grant (17/2551-0000816-2). FLP is supported by grants from CNPQ and FAPERGS (305758/2017-9, 407969/2016-0, 16/2551-0000271-1). Visits to most European collections were supported by a Deutscher Akademischer Austausch Dienst (DAAD) scholarship granted to FLP, whereas a visit to the AMNH was funded by a Collection Study Grant (Richard Gilder Graduate School), also to FLP.
References
Benton, M.J. 2003. When Life Nearly Died. The Greatest Mass Extinction of All Time. 336 pp. Cambridge University Press, London.
Benton, M.J. 2016. The Triassic. Current Biology 26: R1214–R1218. Crossref
Botha, J.A. and Smith, R.M.H. 2006. Rapid vertebrate recuperation in the Karoo Basin of South Africa following the end-Permian extinction. Journal of African Earth Sciences 45: 502–514. Crossref
Butler, R.J., Brusatte, S.L., Reich, M., Nesbitt, S.J., Schoch, R.R., and Hornung, J.J. 2011. The sail-backed reptile Ctenosauriscus from the latest Early Triassic of Germany and the timing and biogeography of the early archosaur radiation. PLoS One 6: e25693. Crossref
Button, D., Lloyd, G.T., Ezcurra, M.D., and Butler, R.J. 2017. Mass extinctions drove increased global faunal cosmopolitanism on the supercontinent Pangaea. Nature Communications 8: 733. Crossref
Camp, C.L. 1945. Prolacerta and the protosaurian reptiles; Part II. American Journal of Science 243: 84–101. Crossref
Cohen, K.M., Finney, S.C., Gibbard, P.L., and Fan, J.-X. 2013. The ICS International Chronostratigraphic Chart. Episodes 36: 199–204.
Dalla Vecchia, F.M. 2006. Remains of Tanystropheus, Sauropterygians and “Rauisuchians”(Reptilia) in the Middle Triassic of Aupa Valley (Udine, Friuli Venezia Giulia, Italy). Gortania Atti del Museo Friulano di Storia Naturale 27: 25–48.
Da-Rosa, A.A.S., Piñeiro, G., Dias-Da-Silva, S., Cisneros, J.C., Feltrin, F.F., and Neto, L.W. 2009. Bica São Tomé, um novo sítio fossilífero para o Triássico Inferior do sul do Brasil. Revista Brasileira Paleontologia 12: 67–76. Crossref
Dias-Da-Silva, S. 1998; Novos achados de vertebrados fosseis na formação Sanga do Cabral Eotriassico da Bacia do Parana. Acta Geológica Leopoldensis 46–47: 101–108.
Dias-Da-Silva, S., Marsicano, C., and Schultz, C.L. 2005. Early Triassic temnospondyl skull fragments from Southern South America (Paraná Basin, Brazil). Revista Brasileira de Paleontologia 8: 165–172.
Dias-Da-Silva, S., Modesto, S.P., and Schultz, C.L. 2006. New material of Procolophon (Parareptilia: Procolophonoidea) from the Lower Triassic of Brazil, with remarks on the ages of the Sanga do Cabral and Buena Vista Formations of South America. Canadian Journal of Earth Sciences 43: 1685–1693. Crossref
Dias-Da-Silva, S., Pinheiro, F.L., Da-Rosa, A.A.S., Martinelli, A.G., Schultz, C.L., Silva-Neves, E., and Modesto, S.P. 2017. Biostratigraphic reappraisal of the Lower Triassic Sanga do Cabral Supersequence from South America, with a description of new material attributable to the parareptile genus Procolophon. Journal of South American Earth Sciences 79: 281–296.
Dilkes, D.W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B 353: 501–541. Crossref
Evans, S.E. 1988. The early history and relationships of the Diapsida. In: M.J. Benton (ed.), The Phylogeny and Classification of the Tetrapoda, Volume 1, 221–260. Clarendon Press, Oxford.
Ezcurra, M.D. 2010. Biogeography of Triassic tetrapods: evidence for provincialism and driven sympatric cladogenesis in the early evolution of modern tetrapod lineages. Proceedings of the Royal Society B 277: 2547–2552. Crossref
Ezcurra, M.D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematic of proterosuchian archosauriforms. PeerJ 4: e1778. Crossref
Ezcurra, M.D. and Butler, R.J. 2015. Taxonomy of the proterosuchid archosauriforms (Diapsida: Archosauromorpha) from the earliest Triassic of South Africa, and implications for the early archosauriform radiation. Palaeontology 58: 141–170. Crossref
Ezcurra, M.D., Scheyer, T.M., and Butler, R.J. 2014. The origin and early evolution of Sauria: reassessing the Permian saurian fossil record and the timing of the crocodile-lizard divergence. Plos One 9(2): e89165. Crossref
Ezcurra, M.D., Velozo, P., Meneghel, M., and Piñeiro, G. 2015. Early archosauromorph remains from the Permo-Triassic Buena Vista Formation of north-eastern Uruguay. PeerJ 3: e776. Crossref
Faccini, U.F. 1989. O Permo-Triássico do Rio Grande do Sul - uma análise sob o ponto de vista das seqüências deposicionais. Porto Alegre. 121 pp. Dissertação de Mestrado em Geociências, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre.
Fedorov, A., Beichel, R., Kalpathy-Cramer, J., Finet, M.S., Fillion-Robin, J.-C., Pujol, S., Bauer, C., Jennings, D., Fennessy, F., Sonka, M., Buatti, J., Aylward, S., Miller, J.V., Pieper, S., and Kikinis, R. 2012. 3D Slicer as an image computing platform for the quantitative imaging network. Magnetic Resonance Imaging 30: 1323–1341. Crossref
Foth, C., Ezcurra, M.D., Sookias, R.B., Brusatte, S.L., and Butler, R.J. 2016. Unappreciated diversification of stem archosaurs during the Middle Triassic predated the dominance of dinosaurs. BMC Evolutionary Biology 16: 1. Crossref
Goloboff, P.A. and Catalano, S.A. 2016. TNT: tree analysis using new technology, vers. 1.5. Willi Hennig Society edition. http://Www.Zmuc.Dk/Public/Phylogeny/Tnt.
Gower, D.J. and Sennikov, A.G. 2000. Early archosaurs from Russia. In: M.J. Benton, E.N. Kurochkin, M.A. Shishkin, and D.M. Unwin (eds.), The Age of Dinosaurs in Russia and Mongolia, 140–159. Cambridge University Press, Cambridge.
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G. 2012. The geologic time scale. Elsevier 2: 1176.
Holz, M. and Souto-Ribeiro, A. 2000. Taphonomy of the south-Brazilian Triassic vertebrates. Revista Brasileira de Geociências 30: 487–490. Crossref
Irmis, R.B. and Whiteside, J.H. 2011. Delayed recovery of non-marine tetrapods after the end-Permian mass extinction tracks global carbon cycle. Proceedings of the Royal Society B 279: 1732.
Jalil, N-E. 1997. A new prolacertiform diapsid from the Triassic of North Africa and the interrelationships of the prolacertiformes. Journal of Vertebrate Paleontology 17: 506–525. Crossref
Jaquier, V.P. and Scheier, T.M. 2017. Bone histology of the Middle Triassic long-necked reptiles Tanystropheus and Macrocnemus (Archosauromorpha, Protorosauria). Journal of Vertebrate Paleontology 37: 2. Crossref
Jiang, D.-Y., Rieppel, O., Fraser, N.C., Motani, R., Hao, W.-C., Tintori, A., Sun, Y.-L., and Sun, Z.-Y. 2011. New information on the protorosaurian reptile Macrocnemus fuyuanensis Li et al., 2007, from the Middle/Upper Triassic of Yunnan, China. Journal of Vertebrate Paleontology 31: 1230–1237. Crossref
Langer, M.C. and Schultz, C.L. 1997. Further comments on the Brazilian protorosaurs. Ameghiniana 34: 537.
Li, C., Zhao, L., and Wang, L.-T. 2007. A new species of Macrocnemus (Reptilia: Protorosauria) from the Middle Triassic of southwestern China and its paleogeographical implication. Science in China Series D: Earth Sciences 50: 1601–1605. Crossref
Maddison, W.P. and Maddison, D.R. 2015. Mesquite: A modular system for evolutionary analysis. Version 3.2. http://Mesquiteproject.org.
Martinelli, A.G., Francischini, H., Dentzien-Dias, P.C., Soares, M.B., and Schultz, C.L. 2016. The oldest archosauromorph from South America: postcranial remains from the Guadalupian (mid-Permian) Rio do Rasto Formation (Paraná Basin), southern Brazil. Historical Biology 29: 76–84. Crossref
Modesto, S.P. and Sues, H.-D. 2004. The skull of the Early Triassic archosauromorph reptile Prolacerta broomi and its phylogenetic significance. Zoological Journal of the Linnean 140: 335–351. Crossref
Nesbitt, S.J. 2011. The Early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Nesbitt, S.J., Flynn, J.J., Pritchard, A.C., and Parrish, J.M. 2015. Postcranial osteology of Azendohsaurus madagaskarensis (?Middle to Upper Triassic, Isalo Group, Madagascar) and its systematic position among stem archosaur reptiles. Bulletin of the American Museum of Natural History 398: 1–126. Crossref
Nosotti, S. 2007. Tanystropheus longobardicus (Reptilia, Protorosauria): Re-interpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, Northern Italy). Memorie Della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 35: 94.
Piñeiro, G., Ferigolo, J., Ribeiro, A.M., and Velozo, P. 2015. Reassessing the affinities of vertebral remains from Permo-Triassic beds of Gondwana. Comptes Rendus Palevol 14: 387–401. Crossref
Piñeiro, G., Marsicano, C., Goso, C., and Morosi, E. 2007. Temnospondyl diversity of the Permian–Triassic Colônia Orozco local fauna (Buena Vista formation) of Uruguay. Revista. Brasileira de Paleontologia 10: 169–180. Crossref
Piñeiro, G., Rojas, A., and Ubilla, M. 2004. A new procolophonoid (Reptilia, Parareptilia) from the upper permian of Uruguay. Journal of Vertebrate Paleontology 24: 814–821. Crossref
Pinheiro, F.L., França, M.A.G., Lacerda, M.B., Butler, R.J., and Schultz, C.L. 2016. An exceptional fossil skull from South America and the origins of the archosauriform radiation. Scientific Reports 6: 22817. Crossref
Pritchard, A.C, Turner A.H., Nesbitt, S.J., Irmis, R.B., and Smith, N.D. 2015. Late Triassic tanystropheids (Reptilia, Archosauromorpha) from northern New Mexico (Petrified Forest Member, Chinle Formation) and the biogeography, functional morphology, and evolution of Tanystropheidae. Journal of Vertebrate Paleontology 35: 911186. Crossref
Pritchard, A.C. and Nesbitt, S.J. 2017. A bird-like skull in a Triassic diapsid reptile increases heterogeneity of the morphological and phylogenetic radiation of Diapsida. Royal Society Open Science 4: 170499. Crossref
Renesto, S. 2005. A new specimen of Tanystropheus (Reptilia Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus. Rivista Italiaa di Paleontologia e Sratigrafia 11: 377–394.
Renesto, S. and Dalla Vecchia, F.M. 2007. A revision of Langobardisaurus rossii Bizzarrini and Muscio, 1995 from the Late Triassic of Friuli. Revista Italiana di Paleontologia e Stratigrafia 113: 191–201.
Rieppel, O. 1989. The hind limb of Macrocnemus bassani (Nopcsa) (Reptilia Diapsida): development and functional anatomy. Journal of Vertebrate Paleontology 9: 373–387. Crossref
Saller, F., Renesto, S., and Dalla Vecchia, F.M. 2013. First record of Langobardisaurus (Diapsida, Protorosauria) from the Norian (Late Triassic) Austria, and a revision of the genus. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 268: 83–95. Crossref
Sengupta, S., Ezcurra, M.D., and Bandyopadhyay, S. 2017. A new horned and long-necked herbivorous stem-archosaur from the Middle Triassic of India. Scientific Reports 7: 8366. Crossref
Sennikov, A.G. 2011. New tanystropheids (Reptilia: Archosauromorpha) from the Triassic of Europe. Paleontological Journal 45: 90–104. Crossref
Senter, P. 2004. Phylogeny of Drepanosauridae (Reptilia: Diapsida). Journal of Systematic Palaeontology 2: 257–268. Crossref
Sues, H-D. 2003. An unusual new archosauromorph reptile from the Upper Triassic Wolfville Formation of Nova Scotia. Canadian Journal of Earth Sciences 40: 635–649. Crossref
Wild, R. 1973. Die Triasfauna dr Tessiner Kalkalpen. XXIII Tanystropheus longobardicus (Bassani) (Neue Ergebnisse). Abhandlungen der Schweizerischen Paläontologischen Gesellschaft 95: 1–162.
Zerfass, H., Lavina, E.L., Schultz, C.L., Garcia, A.J.V., Faccini, U.F., and Chemale, F., Jr. 2003. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: a contribution to southwestern Gondwana paleogeography and paleoclimate. Sedimentary Geology 161: 85–105. Crossref
Acta Palaeontol. Pol. 63 (4): 713–723, 2018
https://doi.org/10.4202/app.00489.2018