

Pterosauria of the Great Oolite Group (Bathonian, Middle Jurassic) of Oxfordshire and Gloucestershire, England
MICHAEL O’SULLIVAN and DAVID M. MARTILL
O’Sullivan, M. and Martill, D.M. 2018. Pterosauria of the Great Oolite Group (Bathonian, Middle Jurassic) of Oxfordshire and Gloucestershire, England. Acta Palaeontologica Polonica 63 (4): 617–644.
The current understanding of UK Middle Jurassic pterosaur taxonomy is under-developed, leading to it being previously considered a time of low diversity. This is despite the presence of a productive but under-studied pterosaur-bearing horizon extending over parts of Oxfordshire and Gloucestershire. This unit, informally called the Stonesfield Slate, is part of the Great Oolite Group and it produces the largest number of Middle Jurassic pterosaurs. There are over 200 specimens distributed across museums in the United Kingdom, America, and Australia, almost all of which are accessioned under the genus Rhamphocephalus and referred to three species: the type species Rhamphocephalus prestwichi, Rhamphocephalus bucklandi, and Rhamphocephalus depressirostris. This study reviews the British Middle Jurassic Pterosauria assemblage, evaluating both their systematics and taxonomic diversity. The holotype of Rhamphocephalus, an isolated skull table, is found to be a misidentified crocodylomorph skull and the genus is considered a nomen dubium. The holotype of Rhamphocephalus bucklandi is identified as missing and that of Rhamphocephalus depressirostris has characters diagnostic at a family level, not a generic or specific one. Both species are considered dubious. Detailed examination of the entire assemblage shows that rather than being monogeneric, the assemblage contains at least five pterosaur taxa, representing three families. This diversity includes the potential earliest occurrences of both Monofenestrata and Pterodactyloidea. A new genus, Klobiodon rochei gen. et sp. nov. is described based on a well-preserved mandible. The English Bathonian pterosaur assemblage is shown to be diverse and indicates that, as has been suggested in other studies, the low-diversity signal in the Middle Jurassic is at least partially artificial.
Key words: Pterosauria, Rhamphocephalus, Jurassic, Bathonian, Taynton Limestone Formation, UK, England, Stonesfield Slate.
Michael O’Sullivan [michaelosullivanvp@gmail.com] and David M. Martill [david.martill@port.ac.uk], Palaeobiology Research Group, School of Earth and Environmental Sciences, University of Portsmouth, Burnaby Building, Burnaby Road, Portsmouth, PO1 3QL, UK.
Received 18 April 2018, accepted 25 September 2018, available online 7 November 2018.
Copyright © 2018 M. O’Sullivan and D.M. Martill. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In recent years there has been a renewed interest in Jurassic pterosaur taxonomic diversity (Butler et al. 2009, 2012, 2013; O’Sullivan 2017; O’Sullivan and Martill 2017; O’Sullivan and Rigby 2017). Historically, the understood pattern of pterosaur evolution has been that pterosaur appear in the Late Triassic and serve almost as background fauna, with no radical morphological shifts for much of the Jurassic. The group then underwent an explosion of diversity in the Late Jurassic and remain highly variable for much of the Cretaceous (Romer 1956; Wellnhofer 1991). This was supported by the high level of pterosaur productivity from Late Jurassic formations in Germany and Asia, e.g., the Solnhofen Limestone Formation which currently contains 11 pterosaur genera while almost all Triassic and Early–Middle Jurassic pterosaur units have only one or two species (Butler et al. 2009; Vidovic and Martill 2014). However, several recent studies (e.g., Butler et al. 2009, 2012, 2013; Tutin and Butler 2017) argue that this is an artificial diversity signal, affected by taphonomic biases. As we move into the Late Jurassic there is an increase in Lagerstätten which produce exceptional numbers of exquisite fossils and it is argued this creates a much higher preservation potential for fragile pterosaur remains, resulting in more species being identified. The smaller number of Lagerstätten in the Triassic and Early–Middle Jurassic is said to result in a much lower yield in pterosaur fossils, whose weaker construction makes them more susceptible to destruction under normal preservational conditions (Butler et al. 2012, 2013). The mentioned studies have used data analyses to test Jurassic pterosaur diversity signals but there has not been a practical examination done on a pterosaur assemblage due to the low yield of most Late Triassic and Early Jurassic formations. There are, however, exceptions with several formations which have a fossil assemblage comparable to the Late Jurassic Lagerstätten. Of these the most heavily studied is Middle to Late Jurassic Tiaojishan Formation of China (Czerkas and Ji 2002). This produces large numbers of complete and near complete pterosaurs which identifies not only a high number of taxa but several transitional forms (Lü et al. 2010). There is also another, frequently overlooked, source of information found in the United Kingdom. The Bathonian (Middle Jurassic) Great Oolite Group is an oolitic unit exposed in the Oxfordshire and Gloucestershire regions of the UK. It has produced some 215 isolated pterosaur fossils (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app63-OSullivan_Martill_etal_SOM.pdf). Most of these specimens have been recovered from the Taynton Limestone Formation, colloquially known as the Stonesfield Slate. The material is largely incomplete, isolated and often fragmentary but contains a variety of fossil types ranging from partial mandibles, cervical vertebrae and various appendicular elements (Fig. 1). Despite the large number of specimens, the assemblage contains only one named genus, Rhamphocephalus Seeley, 1880, divided into 3 poorly defined species. Since its inception, the genus Rhamphocephalus has never been systematically evaluated but its validity has been questioned several times (see below). The high productivity and age of the Great Oolite Group makes it a strong candidate for examining whether Jurassic pterosaur diversity really did “explode” in the Late Jurassic or whether the signal has been artificially inflated by the low yield of older formations. What follows is an historical account of Great Oolite Group pterosaur research and the first detailed examination of the assemblage, testing the validity of established taxa, looking for novel animals and gaining an understanding of what the diversity of the Great Oolite Group pterosaur fauna tells us about the nature of pterosaur diversity in the Jurassic.
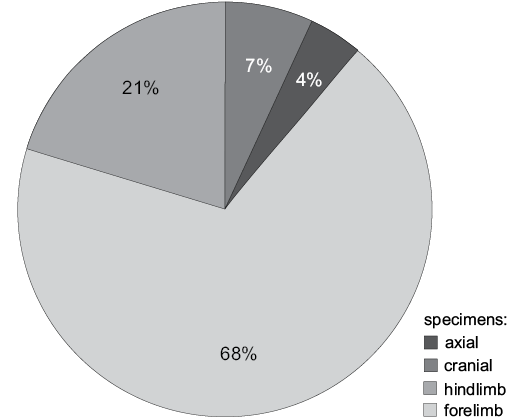
Fig. 1. Pie chart showing the percentage division of the Great Oolite Group pterosaur fossils.
Institutional abbreviations.—GSM, British Geological Survey, Keyworth, UK; MUM, Manchester University Museum, Manchester, UK; NMW, National Museum of Wales, Cardiff, Wales, UK; NHMUK, Natural History Museum, London, UK; NHMW, Natural History Museum, Vienna, Austria; OUM, Oxford University Museum, Oxford, UK; UUPM, University of Uppsala, Sweden.
Other abbreviations.—DPC, deltopectoral crest; MC, metacarpal; WP, wing phalanx.
Nomenclatural acts.—The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank. The LSID for this publication is: urn:lsid:zoobank.org:act:F79D2658-AF38-4F16-87C2-4BAC9F1021E5. The electronic edition of this work was published in a journal with an ISSN and has been archived. It is available from the following digital repositories: PubMed Central, LOCKSS.
Historical background
The earliest known record of Bathonian pterosaurs is an account of “fossil bird bones” from the Taynton Limestone Formation of Stonesfield by an anonymous author A.B., appearing in the March edition of the Gentleman’s Magazine of 1757 (AB 1757). This brief mention was made 27 years before the first scientific description of a pterosaur, Pterodactylus antiquus Sömmerring, 1812, by Alessandro Collini (1784). Delair and Sarjeant (2002) determined that A.B. was a pseudonym for David Stuart Erskine (later Earl of Buchan), a prolific fossil collector of his day. However, Howlett et al. (2017) argued against this identity, pointing out that the pseudonym was not used by Erskine until 1785 and he would have been only 15 years old at the time of publication. Instead, they suggested that the initials stand for “Artium Baccalaureus” or Bachelor of Arts in Oxford University. A pterosaurian identification can be given to this material based on the work of William Buckland (1836), who (on the suggestion of John Samuel Miller an eminent Bristol based naturalist) later proposed that all supposed avian material from the Taynton Limestone should be considered pterosaur; a point reiterated by several others (e.g., Owen 1859a; Weishampel et al 2004). Unfortunately, these original specimens were likely subsumed into various 19th century collections and can no longer be traced.
Over the decades that followed several probable Bathonian pterosaur fossils were described (e.g., Parkinson 1811; Ure 1829) but it wasn’t until 1832 that a specimen from the Taynton Limestone Formation was formally acknowledged. Meyer (1832) erected the taxon Pterodactylus bucklandi for a disarticulated WP3 and WP4 but the specimens were never figured or identified in text. The original description was attributed by Meyer (1832) to the naturalists Spix and Martius, however, there is no record of them describing Stonesfield material. Johann Baptist von Spix (Spix 1817) did publish a description of an associated WPIII and WPIV from the Solnhofen Limestone Formation which he described as a fruit bat, and so it is possible that Meyer (1832) misread this description and mistakenly believed it was from Stonesfield. Regardless, Pterodactylus bucklandi was diagnosed in text on what was perceived as a unique WP ratio, where WPIII was longer than WPIV; and the phalanges being three times the size of the analogous elements in Pterodactylus.
Huxley (1859) discussed several isolated pterosaur elements from the Great Oolite Group, including lower jaws from both Stonesfield and lithologically equivalent strata at Sarsden, also in Oxfordshire (Boneham and Wyatt 1993). Much of this material is now accessioned in the NHMUK, OUM, and GSM collections. The most significant of Huxley’s (1859) mandibles were an unnumbered specimen, hereafter referred to as “Huxley 1859: fig. 2” (Fig. 2A) and GSM 113723 (Fig. 2B). He argued that these jaws represented different species and referred GSM 113723 to the new taxon Rhamphorhynchus depressirostris Huxley, 1859 based on the robust nature of the jaw and shallow anterior rostrum; reduced curvature in the mandible, and 5 teeth in the dentary. The affinities of “Huxley 1859: fig. 2” were less well described with Huxley (1859) figuring the mandible as belonging to either to Rhamphorhynchus depressirostris or Pterodactylus bucklandi. In the text, however, Huxley (1859: 666) noted: “… it is impossible to compare the mandibles I have described with the corresponding parts of the species named Pterodactylus bucklandi, inasmuch as none exist; but since a specific name published without a description has no authority, I shall not hesitate to affix the name of Rhamphorhynchus bucklandi to the nearly perfect mandible from Stonesfield, and provisionally, to the other remains in the same area.”

Fig. 2. A. The holotype mandible (“Huxley 1859: fig. 2”) of “Rhamphocephalus” bucklandi (Meyer, 1832) (nomen nudum) from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK, which has since gone missing and is presumed lost. B. The holotype mandible (GSM 113723) of “Rhamphocephalus” depressirostris Huxley, 1859 from Fuller’s Earth Formation (Bathonian, Middle Jurassic), Smith’s Quarry, Sarsden, Chipping Norton, Oxfordshire, UK. Both after Huxley (1859: figs. 1a/1b and 2 respectively). No scales in the original figures.
Several other specimens were figured within the text including a posterior mandible, first wing phalanx (WPI), a fourth metacarpal (MCIV), a humerus and 2 scapulocoracoids (Huxley 1859: figs. 4–9). These other remains were referred to Rhamphorhynchus bucklandi Huxley, 1859 without a formal diagnosis.
The pterosaur fauna of the Taynton Limestone Formation was discussed in detail by Phillips (1871). He did not revise the taxonomy but figured some of the fossils, even providing a life-restoration of Rhamphorhynchus bucklandi. Shortly after, Owen (1874) erected three new Pterodactylus species for Stonesfield specimens: Pterodactylus kiddi Owen, 1874, Pterodactylus duncani Owen, 1874, and Pterodactylus aclandi Owen, 1874 for three isolated WPIs (Fig. 3) based on their proportions and size. It is likely that size was used as a diagnostic character based on an assumption that fully volant pterosaurs must have been osteologically mature and therefore significant size differences, as in birds, could be taxonomically diagnostic. It has since been argued that pterosaurs may have been precocial flyers (Unwin 2005) and thus overall size may have only limited taxonomic application, although a recent analysis of a pterosaur nesting site suggested that newly hatched pterosaurs were unable to fly (Wang et al 2014, 2017).

Fig. 3. Pterosaur first wing phalanges from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. The holotype of Pterodactylus duncani Owen, 1874 (NHMUK PV R 40126 B), right wing phalanx I in dorsal view. B. The holotype of Pterodactylus kiddi Owen, 1874 (NHMUK PV R 40126 C), left wing phalanx I in ventral view. C. The type specimen of Pterodactylus aclandi Owen, 1874 (NHMUK PV R 40126 A), right wing phalanx I in ventral view. All three species considered to be nomina dubia.
An isolated cranium (OUM J.28266) from the Great Oolite Group was made the holotype of Rhamphocephalus prestwichi Seeley, 1880 (see Systematic palaeontology). This specimen is exposed on a limestone slab in dorsal view and was assigned to Pterosauria based on its perceived thin bone walls. Seeley (1880) noted that the arrangement of bones was more crocodilian than pterosaurian and considered this construction diagnostic of the new taxon. Significantly he (Seeley 1880: 30) stated: “I shall be quite prepared to find that all the ornithosaurians from Stonesfield belong to this or an allied genus which had Rhamphorhynchus for its nearest ally.”
Five years later, Seeley (1885) briefly noted that the Stonesfield Slate pterosaur assemblage included Rhamphorhynchus and a new genus named Dolichorhamphus Seeley, 1885 for which there is no description or referred specimens. Later authors (Romer 1956; Kuhn 1967; Wellnhofer 1978) would treat Dolichorhampus as a junior synonym of Rhamphocephalus. Lydekker (1888) re-evaluated the Stonesfield assemblage and synonymized Rhamphorhynchus bucklandi and Rhamphorhynchus depressirostris with Rhamphocephalus while maintaining the separate species. He also presented a revised diagnosis for Rhamphocephalus depressirostris Lydekker, 1888 that included features from a newly discovered mandible (NHMUK PV OR 47991; Fig. 4) with in-situ teeth. Rhamphocephalus was now defined by the anterior teeth being taller than the posterior and a constriction of the cranium near the orbits. Lydekker (1888) also included the fusion of the scapula and coracoid as a character, a now well-known ontogenetic feature. Lydekker (1888) then provided new definitions for both species: Rhamphocephalus bucklandi Lydekker, 1888 and Rhamphocephalus depressirostris Lydekker, 1888. Rhamphocephalus bucklandi he characterized as possessing more than 5 mandibular tooth pairs, its large size and the inferior border of the mandibular ramus being markedly concave. Rhamphocephalus depressirostris was diagnosed by having 5 mandibular tooth pairs and a prominent ventral dip to the jaw line. Several other specimens were placed in Rhamphocephalus with size determining species assignment. In the process, Lydekker (1888) synonymized Pterodactylus duncani and Pterodactylus kiddi with Rhamphocephalus bucklandi, and Pterodactylus aclandi with Rhamphocephalus depressirostris.

Fig. 4. The holotype of rhamphorhynchid pterosaur Klobiodon rochei gen. et. sp. nov. (NHMUK PV OR 47991) from Stonesfield Slate Member, Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; right mandible in right lateral view.
Other investigations into the Bathonian pterosaur assemblage involved several researchers. Owen (1859b) identified two cervical vertebrae as Pterodactylus bucklandi based on their occurrence in the same formation (Andres et al. [2010] noted that these were unusual for non-pterodactyloid cervicals in that they appeared to possess postexapophyses ventrolateral to the posterior condyle). Seeley (1901) attempted to establish the proportions of Rhamphocephalus relative to the Toarcian Campylognathoides Strand, 1928 from the Lower Jurassic of Germany using isolated appendicular elements. He argued that Campylognathoides may be a junior synonym of Rhamphocephalus, based on some perceived crossover of the measurements of the wing and hind limb elements. Arthaber (1922) referred to Rhamphocephalus prestwichi as Rhamphorhynchus prestwichi Arthaber, 1922, perhaps unaware of Lydekker’s work (Lydekker 1888) or simply choosing to ignore it. Between 1926 and 2011, Rhamphocephalus received very little attention with the genus attracting only brief mentions or overviews in the literature (e.g., Kuhn 1967; Wellnhofer 1978; Bennett 1996; Sayão 2003; Andres et al. 2010) and receiving limited evaluation. An exception is Unwin (1996) who provided a slightly expanded diagnosis of Rhamphocephalus, including postcranial characters such as sacral vertebral count. He argued that all Stonesfield material except for the holotype of Rhamphocephalus prestwichi should be synonymized with Rhamphocephalus bucklandi, considering Rhamphocephalus prestwichi highly unusual in that its skull table was massive, texturally coarse and unusually elongate with large temporal openings. He also suggested that Rhamphocephalus prestwichi was not a pterosaur, but subsequent authors continued to treat it as one. Martill (2010) provided a review to the history of pterosaurs in England, including a very brief overview of the Bathonian material currently assigned to the various Rhamphocephalus species, but passed no comment on the validity of Rhamphocephalus. Buffetaut and Jeffery (2012) mentioned the holotype of Rhamphocephalus prestwichi briefly but erroneously referred to it as Rhamphocephalus sedgwicki Buffetaut and Jeffery, 2012.
Geological setting
The Great Oolite Group is a Bathonian (165–166 ma) carbonate dominated stratigraphic unit cropping out widely in central and eastern England, with pterosaur material recovered only from the Oxfordshire and Gloucestershire outcrops (Boneham and Wyatt 1993; Benton and Spencer 1995; Gradstein et al. 2004; Fig. 5). It comprises several oolitic limestone formations with interbedded marly and sandy limestone, silty sandstone, and thin mudstone layers (Arkell 1933; Sellwood et al. 1985; Boneham and Wyatt 1993) which occur throughout the sequence (Arkell 1933; Sellwood et al. 1985). Some of the limestones of the Great Oolite Group split in a “slatey” fashion and thus were commercially mined as a tiling stone between the 17th and early 20th centuries (Plot 1677; Aston 1974; Martill 2010). Mining was concentrated around the village of Stonesfield, Oxfordshire (Fig. 6), where the mines have since been declared the Stonesfield Slate Mines SSSI (Aston 1974).
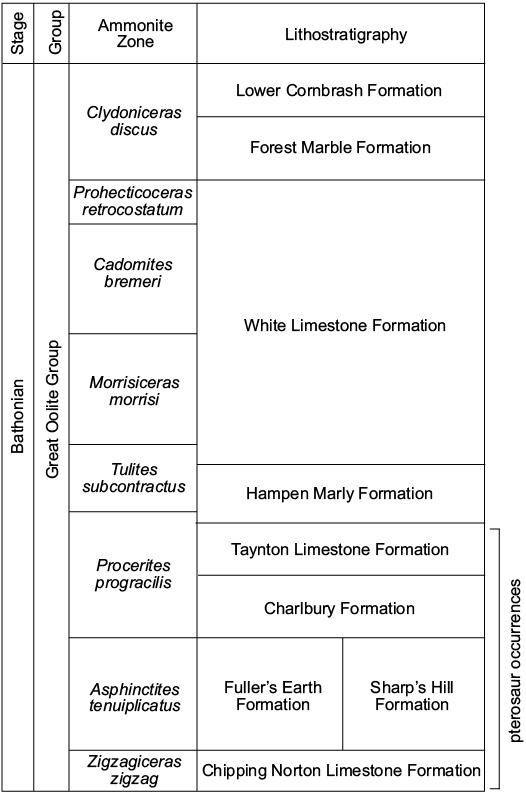
Fig. 5. Simplified stratigraphic column of the Bathonian Great Oolite Group of Oxfordshire and Gloucestershire. An unconformity exists between the White Limestone and Forest Marble formations. Modified from Boneham and Wyatt (1993).
Fig. 6. Maps showing location of the area of the Stonesfield (asterisks) within Oxfordshire, England, UK. Modified from http://www.wikiwand.com/en/Stonesfield.
Most pterosaur specimens from the Great Oolite Group accessioned in British collections are recorded as coming from the Stonesfield Slate (see SOM 1) but as discussed by Boneham and Wyatt (1993), confusion surrounds the source horizons for the tilestones. It has alternately been identified as a sedimentary unit occurring at the top (Hull 1860; Woodward 1894) or bottom (Plot 1677) of the Taynton Limestone Formation; a member within the Sharp’s Hill Formation (Sellwood and McKerrow 1974); or as a unit at the top of the Sharp’s Hill Formation (Arkell 1947; Torrens 1980). Boneham and Wyatt (1993) attempted to identify the source formation of the Stonesfield Slate using cored boreholes drilled from the Stonesfield mines in 1991. They determined that the Stonesfield Slate material was collected from three levels within the Procerites hodsoni Zone (Wyatt 1996) of the Taynton Limestone Formation. The Stonesfield Slate Member was identified as a sequence of interbedded sandstone, siltstone and oolites from the central part of the Taynton Limestone Formation. The Stonesfield Slate Member facies was found to be recurrent throughout the Great Oolite Group in the Stonesfield area and it was concluded that the informal name had been applied to material from other members or even other formations. There is thus doubt as to whether all the material within British collections accessioned under the Stonesfield Slate label is from the Stonesfield Member, from another unit within the Taynton Limestone Formation or even from another formation within the Great Oolite Group. In this study, Middle Bathonian specimens are treated as being from the Taynton Limestone Formation unless explicitly accessioned under another stratigraphic name.
Material and methods
The studied collections house 215 specimens with the majority held at the NHMUK and OUM, 70% of the total assemblage. A complete list of accessioned fossils can be found in the SOM 1.
While most Bathonian pterosaur specimens were freely available for study, two specimens have been excluded from this analysis. NHMUK PV R 464 is a partial pterosaur skull accessioned in the NHMUK. A highly significant fossil, it is currently being studied by Brian Andres. Another specimen, OUM J.01419 is an isolated jaw from the OUM collection described by Buffetaut and Jeffery (2012) as a pterodactyloid that, if correct would make it one of the oldest pterodactyloid pterosaurs. Andres et al. (2014) expressed concern about this identification, suggesting it most likely belonged to a crocodylomorph and consequently it is excluded from this analysis.
This study also does not further evaluate Pterodactylus kiddi, Pterodactylus duncani, and Pterodactylus aclandi. These species were erected for isolated wing phalanges without accompanying diagnoses. For a WPI to be diagnostic of a taxon it must have at least 1 exceptional feature, which these three fossils do not possess, having the typical morphology of Great Oolite Group phalanges (see below; SOM 2). The lack of any distinguishing characteristics leads these three species to be considered nomina dubia and the taxa are not further examined within the text. The phalanges themselves are described as they do have some familial level taxonomic significance.
Systematic palaeontology
See SOM 1 for a complete list of studied specimens. The Bathonian pterosaur assemblage includes numerous high-quality specimens such as a small number of cranial and mandibular specimens, several axial elements and examples of appendicular bones. While a large percentage of the fossils are sufficiently complete to be fully described (examples shown in Fig. 7), reviewing every well-preserved specimen lacking notable diagnostic features is beyond the scope of this paper. Instead, descriptions of the better examples are to be found in SOM 2.

Fig. 7. Examples of Taynoton Limestone Formation limb elements from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. NHMUK PV R 40126, left femur in anterior view. B. NHMUK PV R 11186, left? wing phalanx in dorsal view. These fossils are complete enough for a full description but do not have a great deal of taxonomic significance. They are presented here as examples of the majority of the Stonesfield Slate pterosaur assemblage and are described in SOM 2. Scale bars 10 mm.
Clade Reptilia Laurenti, 1768
Clade Archosauria Cope, 1869
Clade Pterosauria Kaup, 1834
Non-monofenestratan pterosaur indet.
Fig. 8A.
Material.—MUM STR1244b, left MCIV, Stonesfield, Oxfordshire (Bathonian, Middle Jurassic).
Description.—MUM STR1244b (Fig. 8) is a 20 mm long left MCIV exposed in dorsal view with an enlarged large double condyle. It is preserved in anterior view with an expanded ventral condyle. It is short and broad with a length/width ratio of approximately 4:1, falling within the typical range of non-monofenestratan pterosaurs (Wild 1984; Wellnhofer 1991; Martill et al. 2013), and well below the 9:1–20:1 range seen in monofenestratans such as Darwinopterus Lü, Unwin, Jin, Liu, and Ji, 2010.
Remarks.—More precise identification of MUM STR1244b is difficult as pterosaur metacarpals in anteroposterior views are only infrequently figured and their diagnostic features less well documented than those exposed in dorsal, ventral, cranial, and caudal views. We restrict our identification to Non-Monofenestrata indet.
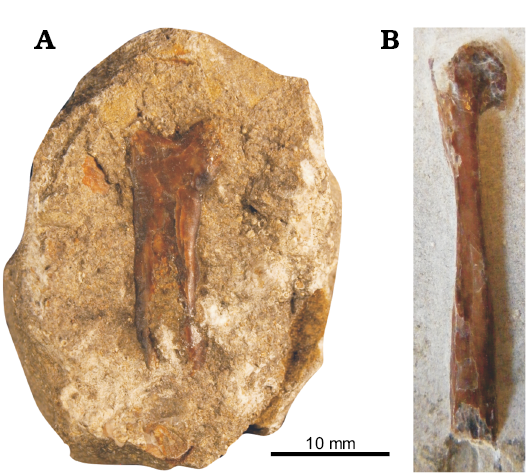
Fig. 8. Metacarpi from the Great Oolite Group from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. A non-monofenestratan pterosaur indet. (MUM STR1244b), left MCIV in anteroposterior view. B. ?Monofenestrata indet. (NHMUK PV R 28160b), right? MCIV in dorsoventral view.
Clade Breviquartossa Unwin, 2003
Family Rhamphorhynchidae Seeley, 1870
Remarks.—Rhamphorhynchidae is one of the best-known Jurassic pterosaur groups. First named by Seeley (1870) for its most famous member Rhamphorhynchus Meyer, 1846 the family is divided into the subfamilies Rhamphorhynchinae and Scaphognathinae. Rhamphorhynchines possess slender skulls with a well-developed elongate anterior prow, procumbent fangs, tall neural spines and some of the longest wing/body ratios of any non-pterodactyloid pterosaur with wingspans reaching 2 m in some species (Witton 2013; O’Sullivan and Martill 2017). Scaphognathines have thicker skulls, more robust cervical vertebrae, shorter prows, more vertically directed teeth and generally shorter wingspans (Unwin 2005; Witton 2013). Rhamphorhynchidae is the most common pterosaur group in the Bathonian of Britain, making up more than 90% of the assemblage and including at least 3 different species, including a novel taxon. Excluded from this categorization is the holotype of Rhamphocephalus prestwichi, which is instead identified as non-pterosaurian material (see below).
Genus Klobiodon nov.
Type species: Klobiodon rochei gen. et sp. nov.; see below.
Etymology: From the Greek klobion, small cage and don, tooth; in reference to the large anterior laniaries that appear to form a fish grab.
Diagnosis.—As for the type and only species (see below).
Klobiodon rochei sp. nov.
Fig. 4.
1878 Pterodactylus raptor; Waterhouse 1878: 34.
1888 Rhamphocephalus depressirostris; Lydekker 1888: 36.
1995 Rhamphocephalus depressirostris (Lydekker, 1888); Benton and Spencer 1995: 144.
2012 Rhamphocephalus depressirostris (Lydekker, 1888); Steel 2012: 1347.
ZooBank LSID: urn:lsid:zoobank.org:act:F79D2658-AF38-4F16-87C2- 4BAC9F1021E5
Etymology: After comic book artist Nick Roche. Comic books are a medium where extinct animals are portrayed in an increasingly scientifically accurate manner, and Roche’s work in the late 2000s was one of the earlier examples in this renaissance in palaeoart.
Holotype: NHMUK PV OR 47991, a right lower jaw in right lateral view.
Type locality: Stonesfield, Oxfordshire, UK.
Type horizon: Stonesfield Slate Member, Taynton Limestone Formation, Bathonian, Middle Jurassic.
Material.—Holotype and OUM 28410, an isolated mandibular symphysis from the type locality.
Diagnosis.—A rhamphorhynchine pterosaur possessing a unique combination of fang-like laniaries and short but robust medial teeth; medial teeth with a vertical height at least 1.3 times the width of the alveolar base; a posterior laniary 1.4–2.4 times the height of the first medial tooth; the longest laniaries are at least 1.5 times the depth of the dentary at its deepest point.
Description.—NHMUK PV OR 47991 (Fig. 4, Table 1) is a 140 mm long jaw preserved in left lateral view with a broken mandibular symphysis and posterior ramus. Using Rhamphorhynchus (NMHUK PV R 37002) and Dorygnathus (MBR 1920.16) as proxies (Padian 2008a; Bonde and Leal 2015) NHMUK PV OR 47991 is estimated to be missing approximately 22% of the total jaw, giving it an estimated original length of ~180 mm. The mandible has a consistent depth of 13 mm posterior to the diverging rami at the mandibular symphysis. From the mandibular symphysis to the anterior break, the jaw depth increases to 17 mm. The ramus curves gently ventrally through an arc of approximately 165°. Mandibular specimen NHMUK PV OR 47991 preserves the second and third anterior teeth as well as the first two medial teeth. Two morphotypes are present; elongate recurved anterior laniaries and shorter, straighter medially positioned teeth. A concave margin on the dorsal surface anterior to the first preserved tooth is identified by Lydekker (1888) as the alveolus for the first tooth. The first preserved laniary is 26 mm long with an 8.6 mm diameter and is directed anteriorly 120° relative to the jaw line. It is elongate and strongly recurved throughout its length. The second preserved is 19 mm long with an 8 mm wide alveolus. It is more robust than its predecessor and less curved distally. Its mesial margin is broader and more sharply recurved relative to the first preserved tooth. It is angled at 110° relative to the jaw line. There is a marked size differentiation between the anterior laniaries and the medial teeth. Even assuming the first medial tooth is a replacement tooth due to its small size, such teeth are always at least 60% erupted (Fastnacht 2008). Thus, the third preserved tooth is at least 1.4–2.4 times the height of the first medial tooth, giving the dental profile a stepped appearance (Fig. 3). The first medial tooth is 7.9 mm tall with a 6.5 mm base. It is squatter and more triangular than the highly recurved laniaries. Like the posterior-most laniary it has a slightly recurved mesial margin and a straight, less expanded distal margin. The second medial tooth is 10.8 mm tall and with a 7.2 mm base. Despite the size difference, the two medial teeth are similarly shaped. Based on a comparison with the jaw of NHMUK PV R 37002, Klobiodon is estimated to have achieved an adult wingspan of ~2 m.
Table 1. Measurements for the teeth of NHMUK PV OR 47991.
|
Tooth |
Crown length (mm) |
Alveolar width (mm) |
Crown width (mm) |
Distance to next tooth (mm) |
Tooth/Jaw depth |
Tooth length |
|
2nd laniary |
26 |
8.6 |
1.7 |
9.3 |
1.7 |
3.3 |
|
3rd laniary |
18.7 |
8 |
1.2 |
7.2 |
1.2 |
2.4 |
|
1st medial |
7.9 |
6.5 |
1.4 |
10.3 |
0.6 |
1 |
|
2nd medial |
10.8 |
7.2 |
?1.6 |
NA |
0.8 |
1.4 |
Besides holotype, only one other specimen can confidently be referred to Klobiodon. OUM J.28410 (Fig. 9) is an isolated mandibular symphysis with three complete alveoli, 63 mm anteroposteriorly and 13 mm dorsoventrally at its deepest point. It is broken posterior to the third alveolus, around the midpoint of the incomplete fourth alveolus and the alveoli are approximately 8 mm wide mesiodistally. The first alveolus is directed dorsoanteriorly, whereas the second and third alveoli are more dorsally oriented. The anterior symphysis is developed into a large sweeping prow which makes up 28% of the preserved jaw. The large alveoli and the relatively deep jaw are similar to the arrangement of the holotype of Klobiodon rochei. However, Klobiodon rochei is primarily defined on dental characters whereas OUM J.28410 lacks teeth. Therefore, it can only tentatively be placed in the genus but is sufficiently similar in its overall morphology to be provisionally identified as Klobiodon cf. rochei.

Fig. 9. An isolated mandibular symphysis of Klobiodon cf. sp. (OUM J.28410), most probably from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; in left lateral view.
Remarks.—Basal pterosaur mandibles are highly variable between taxa (Fig. 10) and often taxonomically informative. The Triassic pterosaurs Austriadactylus Dalla Vecchia, Wild, Hopf, and Reitner, 2002 and Preondactylus Wild, 1984 have similar dental morphologies with tightly packed, sub-triangular serrated teeth (Dalla Vecchia et al. 2002; Dalla Vecchia 2003). Eudimorphodon Zambelli, 1973, Carniadactylus Dalla Vecchia, 2009, and Caviramus Fröbisch and Fröbisch, 2006 share similar complex dentitions, each with tightly spaced heterodont teeth and relatively enlarged anterior laniaries (Wild 1984; Stecher 2008; Dalla Vecchia 2009). Dimorphodon Buckland, 1829 from the lowermost Jurassic of southern England caniform teeth which are tightly packed in the anterior half of the jaws, becoming more widely spaced with the gaps reducing posteriorly (Buckland 1829; Padian 1984a).
Campylognathoides Strand, 1928 from the Toarcian of southern Germany has several anterior enlarged caniform teeth comparable in size to the anterior laniaries seen in several of the more basal pterosaurs (Plieninger 1894). Scaphognathines (sensu Lü et al. 2012) are well known for their jaws whose depth can be as much as 1.4 times the length of the longest dentary tooth crown (Goldfuss 1831; Wellnhofer 1991; Cheng et al. 2012). The tip of the jaw possesses a short prow, less prominent than that of rhamphorhynchines (Wellnhofer 1975; Cheng et al. 2012). The teeth are slightly taller medially, giving the dentition a slightly arched profile (Wellnhofer 1978; Cheng et al. 2012; Bennett 2014) and widely spaced. The curvature and robustness of the teeth in scaphognathines is somewhat variable between taxa and their position in the jaw (Carpenter et al. 2003; Cheng et al. 2012; Bennett 2014).
Rhamphorhynchines (sensu Lü et al. 2012) possess large, procumbent fang-like laniaries (Wellnhofer 1975, 1978; He et al. 1983; Padian 2008b; Hone et al. 2012; Lü et al. 2012) that can mesh together to form a “fish-grab” (Wellnhofer 1991; Kellner and Tomida 2000; Unwin 2003), a cage-like structure at the anterior rostrum associated which some authors associate with an at least partially piscivorous diet (e.g., Wellnhofer 1991). The mandibular symphysis in rhamphorhynchines develops into a hooked anterior prow that may vary in length ontogenetically (Wellnhofer 1975, 1978). The Toarcian Dorygnathus has three enlarged recurved laniaries, which become less procumbent posteriorly. Directly behind the posterior most laniary there is a marked step in tooth height, with the next tooth being 16–33% the height of the last laniary. Posteriorly the successive teeth are of a similar height, are more erect and have a high triangular outline in lateral aspect. The Chinese rhamphorhynchine Angustinaripterus He, Yang, and Su, 1983 has teeth of relatively equal height with the anterior third becoming more strongly procumbent. The Late Jurassic rhamphorhynchines Bellubrunnus Hone, Tischlinger, Frey, and Röper, 2012 from the Late Jurassic of Solnhofen and Qinglongopterus Lü, Unwin, Zhao, Gao, and Shen, 2012 from the Oxfordian of China have a similar dentition to Rhamphorhynchus with the teeth being slightly procumbent and fang-like, and the anteromesial teeth being the most anteriorly inclined. The teeth show a similar profile to those of as Scaphognathus Goldfuss, 1831 with the “arch” created by the elongate third tooth. The Middle Jurassic scaphognathine Jianchangnathus robustus Cheng, Wang, Jiang, and Kellner, 2012 possesses a similar dental arrangement to Scaphognathus but with much larger, more anteriorly oriented laniaries, deep jaw and convex prow. Bennett (2014) argued that these features did not generically separate the two taxa and referred J. robustus to Scaphognathus robustus.
Wukongopteridae (sensu Wang et al. 2009) have numerous slightly recurved, well-spaced teeth (Wang et al. 2009; Lü et al. 2011; Martill and Etches 2012). Ctenochasmatoids (sensu Unwin 2003; also Pterodactylidae sensu Pereda-Suberbiola et al. 2012) possess a wide variety of dental morphologies which fall into two broad categories; well-spaced and slender, or tightly packed and slender (Wellnhofer 1991). Dsungaripteroids have evenly spaced robust teeth situated in a straight jaw with the more derived dsungaripterids have a curving edentulous anterior jaw with very robust medial teeth (Young 1964; Wellnhofer 1991).
Klobiodon rochei can be distinguished from most pterosaurs listed above with little difficulty. Its teeth lack the serrations or multiple cusps of Austriadactylus, Preondactylus, Eudimorphodon, Caviramus, or Carniadactylus. Dimorphodon and Campylognathoides lack laniaries and have more closely spaced alveoli. Wukongopterid teeth have tighter spacing, straighter jaws and do not possess laniaries. The curvature of the jaw and the size of the teeth show that Klobiodon belongs in Rhamphorhynchidae (sensu Lü et al. 2011), the parent clade of Scaphognathinae and Rhamphorhynchinae.
In Klobiodon the longest tooth is much larger relative to the depth of the jaw than is common for scaphognathines (Cheng et al. 2012; Bennett 2014). At least five rhamphorhynchine taxa are known with lower jaws preserved and can be compared directly with Klobiodon. Of these five taxa, the lower jaws of Bellubrunnus and Qinglongopterus are crushed dorsoventrally and difficult to compare to Klobiodon (Hone et al. 2012; Lü et al. 2012). Angustinaripterus lacks complete teeth but the alveoli are of a relatively consistent size (He et al. 1983). This lack of medial/anterior tooth differentiation distinguishes it from Klobiodon. Dorygnathus and Rhamphorhynchus show some size differentiation in the teeth. In Rhamphorhynchus this differentiation is subtle with a slight increase in the tooth size towards the middle of the jaw followed by a gradual decrease posteriorly. This gives it a dental profile not dissimilar to Scaphognathus but distinct from Klobiodon. In contrast, Dorygnathus shares several similarities with Klobiodon. Like Klobiodon the jaws of Dorygnathus can possess a gentle curvature throughout their length, developing into a robust and well-developed prow (Padian 2008b; Fig. 10). Dorygnathus also possesses a well-developed differentiation between small medial teeth and large somewhat recurved laniaries. These similarities do suggest that Klobiodon is more closely allied to Dorygnathus than other rhamphorhynchine pterosaurs, however, Klobiodon does possess a combination of characters that distinguish it from Dorygnathus. While both pterosaurs have small medial teeth, in Dorygnathus they are smaller, more closely spaced and less robust than in Klobiodon (Padian 2008a). In Klobiodon the posterior most laniary is 1.4–2.4 times the size of the first medial tooth. In Dorygnathus (SMNS 55886; Padian 2008b), the posterior most laniary is between 2.6 and 3.7 times the size of the first medial tooth. In the Vienna specimen of Dorygnathus (NHMW 1911/0001/0023 and is close to the size of the Klobiodon holotype [Padian 2008b]), the ratio is 3.3. This difference is a consequence of the medially positioned teeth being much larger and more robust in Klobiodon compared to Dorygnathus, with a tooth length/alveolar width ratio in the anterior medial teeth of at least 1.3. In the above Dorygnathus specimens, the ratio ranges between 2.6 and 6, depending on the mesiodistal width of the tooth. The laniaries of Klobiodon are robust, with broad alveoli while those in Dorygnathus are not only more strongly anteriorly inclined but comparatively thinner mesiodistally. In Klobiodon the longest tooth in the jaw is at least 1.5 times the dorsoventral depth of the jaw at its deepest point. Contrasting this, in NHMW 1911/0001/0023 the tooth/jaw ratio is around 2.6 and in Dorygnathus UUPM R156 the ratio increases to 3. This is due to Klobiodon’s laniaries being shorter relative to the depth of the jaw whereas Dorygnathus has elongate laniaries set in a comparatively narrower mandible (Padian 2008b). Within Scaphognathinae the most comparable animals are the two species of Scaphognathus. Scaphognathus crassirostris Goldfuss, 1831 possess smaller less procumbent teeth and is thus a poor comparison with Klobiodon. Scaphognathus robustus on the other hand has more comparable laniaries, being larger and more procumbent. This may suggest a possible relationship, but the taxa can be distinguished on several characters. Unlike Klobiodon, S. robustus lacks the strong size differentiation between the anterior and medial teeth seen in Klobiodon, with the most posterior tooth being almost 1/1 in scale to the anterior fangs. In S. robustus the longest tooth/jaw depth ratio is 0.8, less than is seen in Klobiodon and is due to the greater depth of its jaw.
While the elongated laniaries, convex anterior prow and deep jaw may indicate a relationship between Klobiodon and S. robustus, the prominent differentiation between the large anterior laniaries and small medial teeth are more like Dorygnathus which also possess a convex prow and anteriorly oriented fangs. The similarities with both taxa, combined with the continued lack of broad consensus in the structure of non-monofenestratan phylogeny (e.g., Lü et al. 2012; Andres and Myers 2013; Vidovic and Martill 2014) recommends caution is assigning the new genus to either Scaphognathinae or Rhamphorhynchinae. Klobiodon rochei is therefore conservatively identified here as a member of the Rhamphorhynchidae rather than assigned to either subgroup.
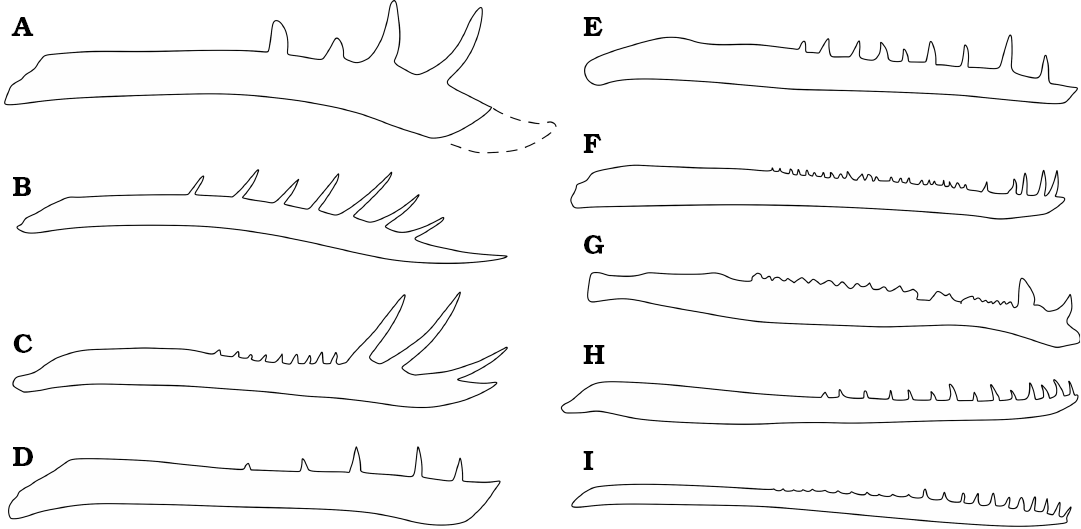
Fig. 10. Simplified diagrams showing the basic morphology of several pterosaur jaws. A. Klobiodon rochei gen. et sp. nov. B. Rhamphorhynchus from Wellnhofer (1975). C. Dorygnathus from Padian (2008b). D. Scaphognathus from Bennett (2014). E. Campylognathoides from Padian (2008a). F. Austriadactylus from Dalla Vecchia et al. 2002. G. Eudimorphodon from Wellnhofer (1991). H. Darwinopterus from Lü et al. (2010). I. Pterodactylus from Wellnhofer (1970). Not to scale.
Remarks.—NHMUK PV OR 47991 was first mentioned by Waterhouse (1878) in a vertebrate acquisitions list for the NHMUK, who confirmed the specimen was donated by Robert Marsham. In the list Waterhouse (1878: 34) can be quoted as listing NHMUK PV OR 47991 as “mandible, right ramus, of Pterodactylus raptor, of Stonesfield Slate, Stonesfield”. The name Pterodactylus raptor was taken from an unpublished manuscript by Richard Owen. No formal description or figure reference has been associated with this name but Ingles and Sawyer’s (1979) compilation of NHMUK illustrations referenced NHMUK PV OR 47991 as being illustrated in Folio 201A of Richard Owen’s unpublished Collection of Drawings under the name Pterodactylus raptor (Michael Hanson, personal communication 2017). Lydekker (1880) placed NHMUK PV OR 47991 in Rhamphocephalus and identified it as a large example of Rhamphocephalus depressirostris. He argued that it possessed two characteristics of the species: five teeth in the dentary and a concave ventral jaw line. It was considered the best example of Rhamphocephalus depressirostris in the NHMUK collections and was noteworthy for its relatively large size. Lydekker’s (1880) description of NHMUK PV OR 47991 was written immediately after his new diagnosis of Rhamphocephalus depressirostris and this may explain some confusion on the part of Benton and Spencer (1995) who mistakenly identified NHMUK PV OR 47991 as the type specimen for Rhamphocephalus depressirostris.
Stratigraphic and geographic range.—NHMUK PV OR is most likely from the Stonesfield Slate Member of the Taynton Limestone Member, collected from the Stonesfield region of Oxfordshire.
Genus Rhamphocephalus Seeley, 1880 (nomen dubium)
Remarks.—The genus Rhamphocephalus is identified in this study as a non-diagnostic thalattosuchian crocodylomorph (see Non-pterosaurian section below). This assignment raises questions as to the taxonomic identification of the holotypes of “Pterodactylus” bucklandi and “Rhamphorhynchus” depressirostris, two isolated jaws described below.
“Pterodactylus” bucklandi Meyer, 1832 (nomen dubium)
(= Rhamphorhynchinae indet.)
Fig. 2A.
1832 Pterodactylus bucklandi; Meyer 1832: 27–30.
1859 Rhamphorhynchus bucklandi (Meyer, 1832); Huxley 1859: 658–670.
1859 Pterodactylus bucklandi Meyer, 1832; Owen 1859b: 169.
1871 Rhamphorhynchus bucklandi (Meyer, 1832); Phillips 1871: 224.
1888 Rhamphocephalus bucklandi (Meyer, 1832); Lydekker 1888: 34–636.
1907 Rhamphocephalus bucklandi (Meyer, 1832); Seitz 1907: 289–6291.
1978 Rhamphocephalus bucklandi (Meyer, 1832), Wellnhofer 1978: 41.
1995 Rhamphocephalus depressirostris (Huxley, 1859); Benton and Spencer 1995: 128.
1996 Rhamphocephalus bucklandi (Meyer, 1832), Unwin 1996: 293.
2000 Rhamphocephalus bucklandi (Meyer, 1832); Riqlés et al. 2000: 351.
2003 Rhamphocephalus bucklandi (Meyer, 1832); Sayão 2003: 335.
2003 Rhamphocephalus bucklandi (Meyer, 1832); Unwin 2003: 177.
2008 Rhamphocephalus bucklandi (Meyer, 1832); Barrett et al. 2008: 68.
2012 Rhamphocephalus bucklandi (Meyer, 1832); Buffetaut and Jeffery 2012: 1.
2012 Rhamphocephalus bucklandi (Meyer, 1832); Steel 2012: 1341.
2013 Rhamphocephalus bucklandi (Meyer, 1832); Witton 2013: 125.
Holotype: “Huxley 1859: fig. 2”, a lower jaw in left lateral view. The whereabouts of the specimen is unknown.
Type locality: Smith’s Quarry, Sarsden, Oxfordshire, UK.
Type horizon: Great Oolite Group, Bathonian, Middle Jurassic.
Material.—NHMUK PV R 28610, 32752, 37765, 38014, 38015, 38016, 38017, 38019, 38020, 38025, 40126, 47994, 47999a, 1028, 1029, 1030, 1824, 2637, 6749, 6750 (Steel 2012); OUM J.28275, J.28537, J.283043 (Huxley 1859), isolated pterosaur limb elements from the Stonesfield region of Oxfordshire with no cross over with the holotype of “Pterodactylus” bucklandi.
Description.—The holotype (“Huxley 1859: fig. 2”) of Rhamphocephalus bucklandi (Fig. 2A) is an isolated jaw with 5 alveoli and a short anterior prow. The holotype was (and remains) lost but as the other specimens figured by Huxley (1859) are accessioned in the OUM collections, it is likely it may have been held there at one point. Based on Huxley’s (1859) figure 2, it is a rhamphorhynchine jaw due to its anterior prow, its being relatively shallow and its low number of teeth. As we cannot locate the specimen, our only reference point being a drawing (which the holotype of Rhamphocephalus prestwichi highlights can be a poor representation of the actual specimen) and Huxley’s (1859) only identifier being it is morphologically distinct from R. depressirostris, we cannot recommend a further detailed analysis. As there is no crossover between Huxley’s (1859) figure and any of the appendicular material assigned to the genus by Lydekker (1888), we also identify the name as inapplicable to anything but “Huxley 1859: fig. 2” and transfer all remaining material to either Pterosauria indet. or Rhamphorhynchinae indet. (see SOM 1). This unfortunately puts “Pterodactylus” bucklandi in a nebulous position. We can confidently state that the number of teeth, slim jaw and distinct prow would place the taxon within Rhamphorhynchinae. We can also say that the teeth seem less broad and the jaw less dorsoventrally thick than Klobiodon but unless the holotype is one day uncovered, we regrettable cannot test the taxon any further.
Remarks.—Placed within Rhamphorhynchinae indet. due to its prowed mandibular symphysis and alveoli indicating broad, well-spaced teeth. The holotype has been lost.
“Rhamphorhynchus” depressirostris Huxley, 1859 (nomen dubium)
(= ?Scaphognathinae indet.)
Fig. 11.
1859 Rhamphorhynchus depressirostris; Huxley 1859: 658–670.
1888 Rhamphocephalus depressirostris (Huxley, 1859); Lydekker 1888: 34–36.
1978 Rhamphocephalus depressirostris (Huxley, 1859); Wellnhofer 1978: 41.
1994 Rhamphocephalus depressirostris (Huxley, 1859); Evans et al. 1994: 307.
1995 Rhamphocephalus depressirostris (Huxley, 1859); Benton and Spencer 1995: 128.
1996 Rhamphocephalus depressirostris (Huxley, 1859); Unwin 1996: 293.
2003 Rhamphocephalus depressirostris (Huxley, 1859); Unwin 2003: 177.
2008 Rhamphocephalus depressirostris (Huxley, 1859); Barrett et al. 2008: 69.
2012 Rhamphocephalus depressirostris (Huxley, 1859); Buffetaut and Jeffery 2012: 1.
2012 Rhamphocephalus depressirostris (Huxley, 1859); Steel 2012: 1341.
2013 Rhamphocephalus depressirostris (Huxley, 1859); Witton 2013: 125.
Holotype: GSM 113723, a three-dimensional lower jaw symphysis preserved in left lateral view.
Type locality: Smith’s Quarry (51°54’8.62”N 1°34’44.74”W), Sarsden, Oxfordshire, UK.
Type horizon: Fuller’s Earth Formation, Bathonian, Middle Jurassic.
Material.—Holotype and NHMUK PV R 40126, isolated limb material; Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire (Steel 2012).
Description.—GSM 113723 is a three dimensional and nearly complete lower jaw symphysis with partial rami (Fig. 11). It is 87 mm long and contained within a block of oolitic limestone typical of the Stonesfield Slate Member. The medial left and lateral right rami are both obscured by matrix, as is the dorsal aspect of the symphysis. It has been damaged with the right ramus broken off and reattached. The anterior symphysis is missing, exposing its oval cross-section and thin bone walls. The ventral posterior symphysis is bowed indicating the presence of a prow. In ventral view the symphysis has a relatively deep sulcus at its posterior boundary. There are five alveoli preserved on the rami, with the fifth alveolus of both rami bearing an in-situ tooth. There are no alveoli preserved posterior of these teeth. The alveoli are 5–6 mm mesiodistally and spaced 3–8 mm apart, with the spacing increasing posteriorly. The anterior alveoli are slightly splayed, giving the dorsal symphysis an undulose margin and hinting that the teeth projected anterolaterally. The preserved teeth are 15 mm long, thin and peg-like, with teeth directed anteriorly at ~70° relative to the jaw line.
The elongate jaw, thin bone walls, the dental arrangement and smooth bone texture confirm GSM 113723 as a pterosaur. The low tooth count and simple elongate teeth distinguish GSM 113723 from the more complex tooth pattern of basal non-rhamphorhynchid pterosaurs. The height and number of the medially placed teeth also serves to separate it from Jurassic monofenestratans, which tend to have numerous relatively low medial teeth (Wellnhofer 1978, 1991; Wang et al. 2010). GSM 113723 is confidently placed in Rhamphorhynchidae due to the narrow mandibular symphysis, reduced tooth count and wide spacing of the teeth. The apparent lack of teeth posterior to the fifth alveolus may ally it with Scaphognathinae as these pterosaurs have no more than 5–6 teeth in the lower jaw (Unwin 2003). Tall, peg-like teeth, wide medial spacing and relatively short prows are also found in scaphognathines (Cheng et al 2012; Bennet 2014; Zhou 2014), being similar to the jaw of Scaphognathus (= Jianchangnathus) robustus (Bennett 2014), a scaphognathine from the Tiaojishan Formation of China.
GSM 113723 is identified here as ?Scaphognathinae but the question remains: can it be assigned to a genus or species? Neither Huxley’s (1859) or Lydekker’s (1888) definition of Rhamphocephalus depressirostris (robust jaw, shallow anterior rostrum, reduced curvature in the mandible, 5 teeth) are diagnostic by modern taxonomic standards. Unfortunately, the taphonomy of the jaw limits morphological taxonomy. The preserved tooth is clearly distinct from Klobiodon but is still similar to what we see in several Jurassic pterosaur taxa (see above) and lacks the distinct ratios inform the definition of Klobiodon. There is a deep mandibular sulcus which can be seen at the posteroventral prow but this character could be ontological. Overall, morphological definition is limited. We also cannot suggest the erection of a chronotaxon, not only because it runs counter to the taxonomic practices of this research but also because named scaphognathines occur in contemporaneous formations (e.g., Cheng et al. 2012). Therefore, GSM 113723 is not currently considered a strong candidate for a generic or specific name despite representing a distinct mandibular morphotype within the assemblage. Regrettably this puts “Rhamphocephalus” depressirostris in a similar nebulous position as “Pterodactylus” bucklandi, if for different reasons. However, if GSM 113723 can at some point undergo further preparation or perhaps CT-scanning, we may reveal currently obscured diagnostic characters.
Remarks.—Alleged attribution to Scaphognathinae Hooley, 1913 is based on the morphology of the teeth and consistent increase in the alveolar spacing towards the posterior end of the jaw.
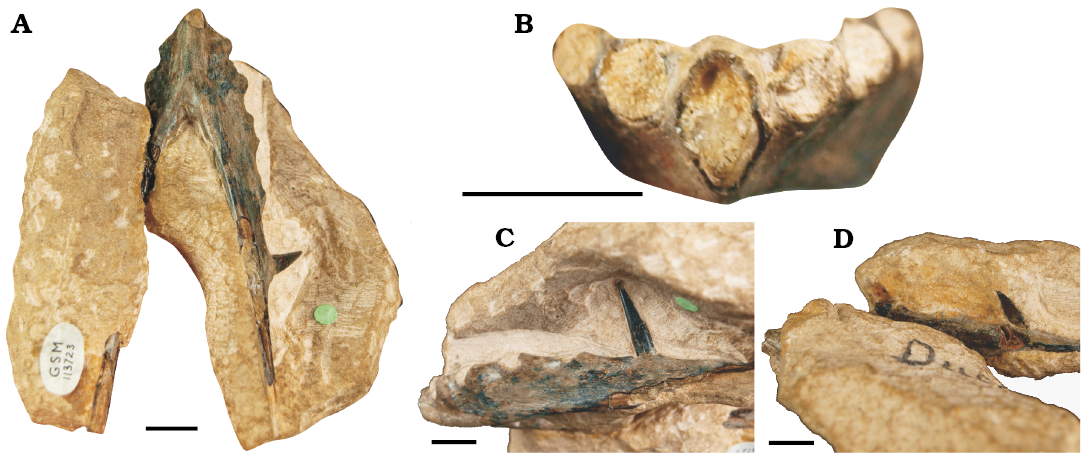
Fig. 11. The holotype specimen of the potential scaphognathine pterosaur “Rhamphocephalus” depressirostris (Huxley, 1859) (GSM 113723) from Fuller’s Earth Formation (Bathonian, Middle Jurassic), Sarsden, Oxfordshire, UK. A. Lower jaw symphysis in ventral view. B. The rostrum in anterior view, showing its splayed alveoli. C. The left ramus in lateral view, displaying the thing and anteriorly directed in-situ 5th tooth. D. The right ramus in medial view with the more stunted in-situ 5th tooth. Scale bars 10 mm.
Other rhamphorhynchid remains
Other identifiable rhamphorhynchid fossils consists of axial and dental remains collected from the Taynton Limestone Formation of Stonesfield and the Chipping Norton Formation of Chipping Norton, both in Oxfordshire (Bathonian, Middle Jurassic). Full locality information is available in SOM 1.
Scapulocoracoids.—OUM J.28294 (Fig. 12B) is a partial left scapulocoracoid preserved in posterolateral view, heavily worn around the glenoidal region. The scapula is 37 mm long and the coracoid is 23 mm long. The coracoid possesses a low biceps tubercle that extends 2 mm ventrally and 6 mm proximodistally. The glenoid is posterolaterally positioned and limited to the scapula. The angle formed between the scapula and coracoid is at 75°. A second complete scapulocoracoid, OUM J.28295 (Fig. 12A) is near complete and preserved in anteroposterior view. It is missing the proximal ends of both the scapula and the coracoid as well as the surface bone of the glenoid. In the case of the glenoid, this exposes the internal trabeculae. The scapula is 28 mm long and the coracoid being 40 mm long. The angle between them is approximately 75°. The restriction of the glenoid to the scapula identifies them as non-pterodactyloid. The proximal coracoid is slenderer than Campylognathoides or Dimorphodon (Buckland 1829; Padian 2008a). The low biceps tubercle is like Dorygnathus, Darwinopterus, and Campylognathoides (Padian 2008a; Lü et al. 2010) but lower than other basal pterosaurs. The angle of 75° falls into the range shared by several basal pterosaurs (Wild 1984; Padian 2008b; Witton 2013). The glenoid limited to the scapula and approximately 10 mm proximodistally. The lateral scapulocoracoid is less inclined than in Rhamphorhynchus but in a similar position to Dorygnathus and Scaphognathus. The coracoid distinguishes it from Darwinopterus where it is wider dorsoventrally and does not taper proximally (Lü et al. 2010). Based on the angle formed by the scapula and coracoid, along with the thin coracoid morphology, the scapulocoracoids are identified as ?Rhamphorhynchidae indet.

Fig. 12. Pterosaur scapulocoracoids from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. A right scapulocoracoid of rhamphorhynchine (OUM J.28295), in right lateral view. B. A partial left scapulocoracoid of non-pterodactyloid pterosaur (OUM J.28294), in left lateral view. Abbreviations: bt, biceps tubercle; co, coracoid; g, glenoid; s, scapula. Scale bars 10 mm.
A complete right scapulocoracoid (OUM J.28297) is preserved in medial view (Fig. 13) and is the most complete pterosaur shoulder element in the assemblage, with a 62 mm long scapula ramus and a 60 mm coracoid ramus. The two elements form an angle of ~70°. The glenoid is restricted to the scapula and is ~16 mm long. The acrocoracoid process is rounded and extends 4.5 mm distal of the glenoid. The coracoid bows slightly behind its scapular articulation. The sternocoracoidal joint is 3 mm wide dorsoventrally and is more rounded on its ventral surface. The biceps tubercle is very low, extending 1 mm ventrally and 7–9 mm proximodistally. OUM J.28297 is identified as a non-pterodactyloid by the glenoid restriction and the coracoid not exceeding the scapula in length. However, the scapula is of sub-equal length to the coracoid whereas in most basal pterosaurs, the scapula is between 124–60% the length of the coracoid (Buckland 1829; Wild 1984; Padian 2008a, b). In Sericipterus (Andres et al. 2010) and Rhamphorhynchus the scapula and coracoid are of sub-equal lengths. Although this suggests a rhamphorhynchine affinity, the same situation occurs in Darwinopterus (Lü et al. 2011). However, Darwinopterus has a larger biceps tubercle than OUM J.28297 while in rhamphorhynchines the size of the biceps tubercles is variable (Padian 2008b; Andres et al. 2010; O’Sullivan and Martill 2015). Based on the ratio of the scapula to the coracoid and the profile of the biceps tubercle, OUM J.28297 is tentatively identified as ?Rhamphorhynchinae indet.

Fig. 13. An isolated left scapulocoracoid of ?Rhamphorhynchine indet. (OUM J.28297) from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; in left lateral view.
Humeri.—OUM J.23043 (Fig. 14A) is a complete left humerus exposed in dorsal view, 90 mm long proximodistally with a diaphysis 10 mm wide anteroposteriorly at its medial point with a length/width ratio of 9/1. The medial process and distal articulation are preserved primarily as an external mould on the rock surface, but with a clearly distinct outline. The diaphysis develops a gentle anterior bowing distally, angled at approximately 165° and distally has a round anterior articulation which extends 8 mm anterior to the shaft. The medial process has a triangular outline and is 10 mm long anteroposteriorly. The DPC is 13 mm anteroposteriorly and 19 mm proximodistally. The anterior margin is rounded, giving the DPC a short tongue-like shape (Fig. 14A). OUM J.23043 is one of the largest Jurassic pterosaur humeri known, 7% larger than the largest Dorygnathus humerus and 13% that of the largest Rhamphorhynchus (Wellnhofer 1975; Padian 2008b). It can be distinguished from via the morphology of the DPC and medial process. Eudimorphodon (Wild 1984), Caviramus (Stecher 2008) and Campylognathoides (Padian 2008a) have enlarged quadrangular DPC. Wukongopterids have similar morphologies to OUM J.23043 but the DPC is less robust (Lü et al. 2011). The medial process of aurorazhdarchids and basal ctenochasmatoids are distally deflected away from the proximal articulation (Wellnhofer 1978; Vidovic and Martill 2014). OUM J.23043 lacks the medial pinching of the DPC seen in Nesodactylus Colbert, 1969 and some examples of Rhamphorhynchus. The DPC of Scaphognathus is a like OUM J.23043 with a comparable placement of the medial process. However, the proximal margin of the DPC is straighter and the DPC is less strongly deflected (Bennett 2014). Sericipterus has a higher length/width ratio than OUM J.23043 (O’Sullivan et al. 2013) and a more elongate DPC (Andres et al. 2010). The humerus of Dorygnathus is like OUM J.23043 (Padian 2008b) but the DPC is generally more elongate. Ultimately OUM J.23043 does not perfectly correlate with any described pterosaur humerus but the DPC, medial process and the gentle curvature of the diaphysis is most like Scaphognathus (Cheng et al. 2012; Bennett 2014), and thus it is assigned to ?Scaphognathinae indet. Scaphognathine humeri make up at least 6% of the total wingspan (Bennett 2014), suggesting a total wingspan of 1.5 m.
NHMUK PV R 40126b (Fig. 14B) is a small 15 mm long humerus, and 1.5 mm wide medially. It is almost complete, missing only the proximal humeral head, medial process and some of the distal articulation. The DPC is 2.4 mm anteriorly, semi-tongue shaped with a broad sub-rectangular anterior margin and a slight proximodistal pinching towards the extremity of the DPC. NHMUK PV R 40126b is identified as ?Rhamphorhynchinae indet based on the slight elongation and minor pinching of the DPC. At 15 mm long NHMUK PV R 40126b is one of the smallest pterosaur humeri known and based on similarly sized examples of Rhamphorhynchus from the Solnhofen Limestone (Wellnhofer 1975), is most likely a juvenile with a 290–340 mm wingspan. NHMUK PV R 40126c (Fig. 14C) is a right humerus preserved in dorsal view. It is 8 mm long proximodistally with a 6 mm wide diaphysis, giving it a length/width ratio of 13:1. It is missing a large section of its humeral head and distal diaphysis. The distal articulation is not preserved. The diaphysis curves anteriorly at 170–175°. NHMUK PV R 40126c does preserve the medial process. The DPC has a rounded anterior margin. The high length/width ratio in combination with the anteriorly bowed shaft of NHMUK PV R 40126c is indicative of it being a possible rhamphorhynchine identification. NHMUK PV R 40126c is identified here as Rhamphorhynchidae, most likely ?Rhamphorhynchinae. The possibility that NHMUK PV R 40126c represents an adult example of NHMUK PV R 40126b and that the straighter shaft in the latter specimen is an ontogenetic feature cannot be ruled out. However, without any intermediaries the two morphologies are considered sufficiently different to rule out an ontogenetic transition. Thus, NHMUK PV R 40126a–c confirms the presence of a second large humeral morphotype that can be assigned to Rhamphorhynchinae rather than Scaphognathinae.
NHMUK PV R 28160a is a left pterosaur humerus preserved in dorsoposterior view (Fig. 14D). It is 14 mm proximodistally with a 1–2 mm medial diaphysis. It is anteriorly bowed through an arc of ~160°. The medial process and DPC both project into and are partially overlain by the matrix. The diaphysis has several “dimples” across the posterior margin corresponding with an immature bone texture as illustrated by Tumarkin-Deratzian et al. (2007). This, combined with its small size, indicates that NHMUK PV R 28160a is a juvenile. The DPC proximally deflected ventral margin. It is similar in overall morphology to OUM J.23043 and is thus also identified as ?Scaphognathinae indet. It should be noted that Unwin (2015) considered NHMUK PV R 28160a as a possible monofenestratan humerus but did not explain his reasoning in detail.
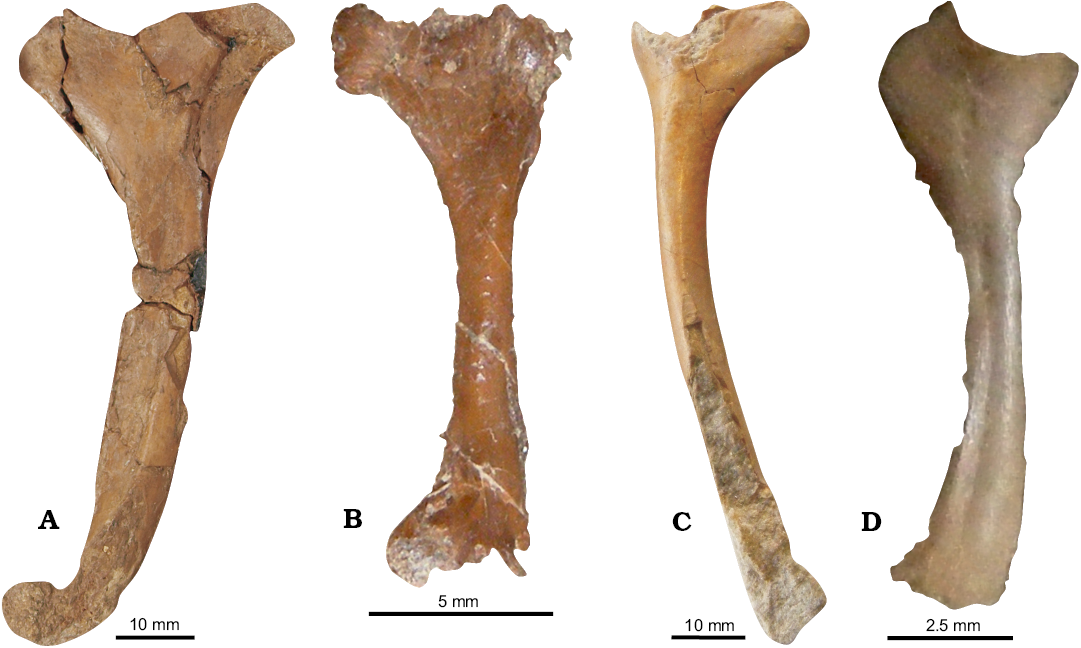
Fig. 14. Pterosaur humeri from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. A left humerus of ?Scaphognathinae indet. (OUM J.23043), in dorsal view (matrix digitally removed). B. A small humerus of rhamphorhynchine (NHMUK PV R 40126b), in dorsoventral view. C. A humerus of rhamphorhynchid (NHMUK PV R 40126c), in dorsal view (matrix digitally removed). D. A small humerus of ?Scaphognathinae indet. (NHMUK PV R 28160a), in dorsal view.
Wing phalanges.—There are several WPIs within the Bathonian pterosaur assemblage (Fig. 2; see SOM 1), although their preservation is variable. The WPIs are of a seemingly uniform morphology, being relatively large and robust elements between 101–142 mm long. Each WPI diaphysis is bowed medially such that the proximal and distal termini are angled at 170–175° relative to the midpoint. The presence or absence of a posterior longitudinal groove is indeterminate in all specimens given their preservation, however, NHMUK PV R 40126 C (Fig. 2B) is not only free of surrounding sediment but broken medially, revealing a triangular cross-section with a relatively thick bone wall and a more rounded anterior surface. Taxonomic identification of these phalanges is problematic given their isolated nature. The consistent robustness and slightly bowed diaphysis suggest they are most likely from a single taxon with distinctive anterior bowing. This is unusual as most pterosaurs have straight proximal phalanges (Wellnhofer 1991; Witton 2013). Unwin (2003) suggests that a bowed phalanx may be a dsungaripteroid apomorphy but notes that a dsungaripteroid presence in the Taynton Limestone is problematic as presently there is no substantive evidence for Dsugaripteroidea in strata older than Kimmeridgian (Fastnacht 2008). Anterior bowing is, however, found in several Dorygnathus specimens (Padian 2008b) and it is therefore not a dsungaripteroid apomorphy. The phalanges of dsungaripteroids are more elongate (Young 1964) than those of the British Bathonian pterosaurs while those of Dorygnathus (e.g., SMNS 56255; Padian 2008b) are shorter and somewhat broader, as in NHMUK PV R 40216. He we identify the phalanges as Rhamphorhynchidae indet. Based on their similarity to the phalanges of Dorygnathus the wingspans of these pterosaurs are estimated to be between 1.4 m and 2 m.
Mandibles.—NHMUK PV R 1824 (Fig. 15), is broken transversely and divided into part and counterpart. The jaw is missing most of the anterior symphysis. It is 100 mm long with rami approximately 13 mm deep dorsoventrally. Both rami preserve five alveoli but with no teeth. The alveolar sections of the rami comprise approximately 49% of the total ramus length. The posterior alveoli are dorsally placed with the more anterior alveoli becoming more laterally placed. The alveoli range between 4–5 mm mesiodistally and are set 6–8 mm apart. The taxonomic identity of NHMUK PV R 1824 is difficult to determine. The number of medial teeth posterior to the symphysis, their relatively uniform size and spacing suggest it is a rhamphorhynchid but it is unclear if it is distinct from or synonymous with Klobiodon rochei. While the holotype of Klobiodon rochei has similar anterior alveolar spacing, the rami of NHMUK PV R 1824 appears more gracile than Klobiodon rochei, and the rami lack the gentle curve seen in Klobiodon. The rami remain relatively straight but curve ventrally proximal to the posterior articulation. It is tentatively identified as Rhamphorhynchidae indet.

Fig. 15. A mandible (split transversely) of ?Rhamphorhynchidae indet. (NHMUK PV R 1824) from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. Two halves of the specimen together, in dorsal view. B. The dorsal slab seen in ventral view. C. Composite image in left lateral view.
Cranial material.—UMZC T.718 (Fig. 16) is a semi-3D posterior skull, 52 mm anteroposteriorly and 37 mm dorsoventrally. It is catalogued under the genus “Rhamphinion jenkensi” Padian, 1984b, an Early Jurassic American genus from the Kayenta Formation of north-eastern Arizona. While the holotype of Rhamphinion is from the same region of the skull, there is very little proportional similarity between the two fossils (see Padian 1984: fig. 1 for comparison) and as the identification has no accompanying descriptive text, this identification is considered here to be unsupported. It preserves the quadrate, quadratojugal, jugal, postorbital and squamosopostorbital bar. The quadrate is 32 mm dorsoventrally and 1.6 mm medially in lateral view. It is elongated, strap-like in posterior view and angled posteriorly at 118°. Ventrally it has a well-developed and rounded condyle for articulating with the mandibular glenoid. The condyle is ~3.8 mm wide. Only the postorbital and quadrate processes of the squamosal are preserved but their full extent is obscured by fractures and some fusion of the skull. The postorbital is tri-radiate, 8 mm anteroposteriorly and 11 mm dorsoventrally. The squamosoparietal bar is elongate and sub-rectangular, 11 mm long anteroposteriorly and 4 mm wide dorsoventrally. The quadratojugal is sub-triangular, with slightly elongated jugal and quadrate processes. The quadratojugal is well developed, 2.3 mm dorsoventrally and 6.7 mm anteroposteriorly, with the jugal being the largest element in the specimen. Including its processes, it is 34 mm anteroposteriorly and ~6 mm dorsoventrally. It is a sub-quadrangular bone, although here the maxillary process is more of a flange, being large and sub-rounded where it borders the antorbital fenestra. The postorbital process is the longest of the four but the point of contact with the postorbital is one of the most damaged regions of bone, obscuring its true length. It is estimated to be 8–16 mm long. The postorbital and lacrimal processes form an angle of ~70° around the ventral margin of the orbit. UMZC T.718 preserves three fenestrae in varying degrees of completeness. The superior temporal fenestra is missing its dorsal half. It is bounded by the squamosal and postorbital and is 14 mm anteroposteriorly. The ventral margin is smooth and sub-oval. The anterior border is more vertical than the posterior, and the anteroventral margin is shallower than the posterior. The inferior temporal fenestra is the only complete fenestra in the specimen. It is 28 mm dorsoventrally and 14 mm anteroposteriorly. The ventral margin is smooth and sub-oval. The anterior border is more vertical than the posterior, and the anteroventral margin is shallower than the posterior. Its boundary includes the quadrate, quadratojugal, jugal, postorbital and squamosal. It possesses an irregular piriform morphology outline, with the dorsal half being wider with a relatively straight dorsal boundary in comparison to the ventral half’s thinner, more angular appearance. The orbit is missing its dorsal half but is clearly the largest of the three fenestrae. It is bounded by the jugal ventrally and the postorbital posteriorly. Like the inferior temporal fenestra, the orbit is piriform with the ventral boundary approximately 10 mm wide in comparison to the 24 mm dorsally positioned widest point. UMZC T.718 is identified as a non-monofenestratan based on its low-lying antorbital fenestra, the relatively shallow angle of the quadrate and its tetraradiate jugal. Generic identification is problematic given the limited material: the angle of the quadrate at 118° distinguishes it from Dimorphodon (Buckland 1829), Campylognathoides (Padian 2008a) and Anurognathus (Bennett 2007) but is comparable to several other basal pterosaurs, including Eudimorphodon, Austriadactylus, and Dorygnathus (Wild 1984; Padian 2008a, b). The angle formed by the dorsal processes of the jugal is typical of basal pterosaurs more derived than Campylognathoides or Eudimorphodon (Wellnhofer 1975; He et al 1983; Wild 1984; Padian 2008b; Bennett 2014). The broad sub-rounded inferior temporal fenestra is distinct from most basal pterosaurs (Wild 1984; Padian 2008a) but a similar condition is seen in several rhamphorhynchids (He et al. 1983; Padian 2008b; Cheng et al. 2012; Bennett 2014). Based on the angle of the quadrate combined with the ventral orbital and inferior temporal fenestra morphology, UMZC T.718 is identified here as ?Rhamphorhynchidae.
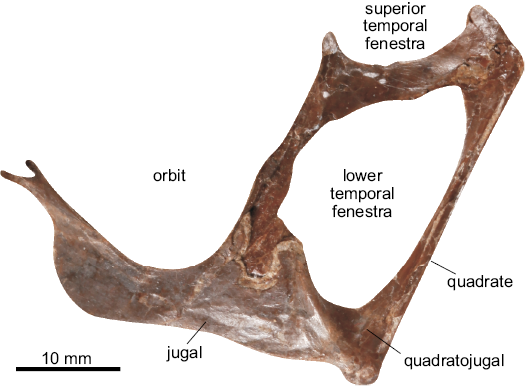
Fig. 16. Partial skull of “Rhamphinion jenkensi” Padian, 1984b (UMZC T.718) from the Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; in left lateral view (matrix digitally removed).
OUM J.28409 (Fig. 17) is an 83 mm long isolated and near-complete left maxilla and a fragment of the posterior premaxilla. Much of the premaxilla and the posterior maxilla are absent. The surface texture appears irregular, but this is a taphonomic artefact as most of the external bone wall has been eroded away anteriorly, revealing the internal trabeculae. The centrally positioned nasal process is angled posteriorly at 70–90°. While the posterior margin of the antorbital fenestra is not present in the maxilla, the preserved section of the premaxilla defines the anterior border of the nares. The nares is approximately 23 mm anterioposteriorly. The antorbital fenestra is at least 27 mm along the same plane. It has five maxillary alveoli. The smallest is 4 mm mesiodistally, with the subsequent alveoli approximately equidimensional at around 7 mm. OUM J.28409 is identified as pterosaur based on a combination of the nares and antorbital fenestra being proximal to each other and the thin bone walls. It is further be identified as a non-monofenestratan as it possesses differentiated nares and antorbital fenestra. The possession of five relatively widely spaced maxillary tooth pairs is a rhamphorhynchid characteristic and OUM J.28409 is identified as Rhamphorhynchidae indet.

Fig. 17. An isolated partial maxilla (OUM J.28409) of Rhamphorhynchidae indet. from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; in lateral view.
Teeth.—Numerus isolated teeth (Fig. 18) have been collected from the Taynton Limestone Formation and accessioned within pterosaur collections. However, due to the difficulty in identifying ex-situ teeth with a simple slender conical, gently curved morphology, many of these have been misidentified. Gently recurved teeth with a slightly sigmoidal shape accessioned as pterosaur might more correctly be assigned to Teleosauridae (Massare 1987). Pterosaur teeth can be identified by the restriction of the enamel to the higher parts of the tooth crown while the base of the crown is exposed dentine. The exposed dentine sometimes extends laterally to a median point of the crown (Witton 2013). Six teeth thought to be pterosaurian, lacking carinae and seemingly possessing restricted enamel are illustrated here (Fig. 18). They share a similar morphology of being relatively elongate and distally recurved, making them likely to be rhamphorhynchine.

Fig. 18. Teeth of probable pterosaur from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; in labiolingual view. A. OUM J.28489. B. OUM J.28495. C. OUM J.28490. D. OUM J.28491. Not to scale.
Monofenestrata remains
Monofenestrata is a recently erected group (Lü et al. 2010) including the transitional Wukongopteridae and the derived Pterodactyloidea. Wukongopteridae includes several taxa showing mosaic evolution with animals possessing derived skull morphology but plesiomorphic bodies. The recognition of this group has not only revolutionised our understanding of pterosaur evolution but complicates Middle Jurassic taxonomy. With few uniquely diagnostic features (Witton et al. 2015) found within the group, isolated remains are easily assigned to either basal or pterodactyloid pterosaurs, and consequently, this has been a major contributor to the conservative approach adopted in this study. Pterodactyloidea is a highly diverse group including at least 11 families. Distinguished from basal pterosaurs by features such as metacarpal length, rostral index, dentition, cervical structure, pelvic morphology and numerous other features, the group includes the most well-known pterosaurs (Wellnhofer 1991; Unwin 2005). The Middle Jurassic has gained increasing relevance in their evolution as several discoveries push the group further and further back (Andres et al. 2014). Establishing if any monofenstratans are present in the Bathonian of Britain has considerable significance to Jurassic pterosaur diversity.
Vertebrae.—Whereas several examples of Bathonian pterosaur vertebrae are known (see SOM 1), the most diagnostic specimens are those accessioned under NHMUK PV R 40126a. These are two isolated cervical vertebrae referred to here as CVA and CVB (Fig. 19). CVA is an isolated cervical vertebra within a slab of oolitic limestone while CVB is a compressed, but three dimensional isolated cervical free of matrix. Both were described by Owen (1859b) as coming from the “Stonesfield oolite” but no locality data was provided. CVA is identified as a cervical based on its robustness, the lack of strongly developed horizontal transverse processes and its relatively elongate centrum (Howse 1986). It is 16 mm anteroposteriorly and 12 wide mm transversely. Based on its quadrangular appearance it is identified as a cervical 3–7 (Wellnhofer 1991). In comparison to CVB, CVA is the more poorly preserved. As well as being dorsoventrally compressed, it is heavily fractured along its lateral margins. Both the pre- and postzygapophyses are preserved but are incomplete. The neural spine has been broken, most likely during compaction. CVB is morphologically similar CVA and considered a cervical 3–7. It is 25 mm anteroposteriorly (excluding the posterior condyle) and 15 wide mm medially. It has undergone dorsoventral compression, the loss of the neural spine and the right prezygapophysis. CVB has undergone some repair or preparation as an adhesive fills several fractures in the surface. Despite this damage, CVB is well preserved and provides a clear view of the centrum in ventral view. Ventrolateral to the posterior condyle are two enlarged projections that in dorsal view are slightly dished. Andres et al. (2010) provided a brief description of NHMUK PV R 40126e, identifying these structures as postexapophyses. In the anteroventral centrum a prominent hypapophysis extends posteriorly towards the midpoint of the cervical. On either side of the neural arch are oval structures which are most likely lateral pneumatic foramina (Wellnhofer 1991; Andres and Norell 2005). On either side of the vertebra are transverse processes, broken proximal to the centrum, corresponding to the dorsal articulation loci for the cervical ribs. NHMUK PV R 40126e have been discussed previously by Owen (1859b) and Andres et al. (2010). Owen figured CVB and identified it as “Pterodactylus” bucklandi but did not provide a description. Andres et al. (2010) discussed the vertebrae in comparison with the cervicals of the rhamphorhynchine Sericipterus from the Shishugou Formation in Xinjiang, China. They noted that NHMUK PV R 40126e cervicals were of a similar size to those of Sericipterus Andres et al. (2010), arguing that they were non-pterodactyloid with the unusual feature of possessing postexapophyses. They suggested that development of postexapophyses may correlate with increasing size of cervicals.
Whereas CVA is more limited in its taxonomic information due to taphonomy, the three-dimensional preservation of CVB allows for a more detailed comparison. Howse (1986) considered the possession of cervical ribs as a characteristic of basal pterosaurs, but reduced cervical ribs are known from several pterodactyloid clades (Zhou and Schoch 2011), including the highly derived Azhdarchidae (Witton and Naish 2008). Cervical ribs are not preserved in the holotype of the non-pterodactyloid monofenestratan Darwinopterus modularis Lü, Unwin, Jin, Liu, and Ji, 2010, but there appear to be anterolateral projections on several vertebrae which may correspond to cervical rib facets. Therefore, the presence of these facets is an unreliable character for distinguishing between pterodactyloid and non-pterodactyloid vertebrae. The presence of hypapophyses in combination with the postexapophyses corresponds with Howse’s (1986) descriptions of pterodactyloid cervical vertebrae however, as Andres et al. (2010) note, in most respects these vertebrae appear typically non-pterodactyloid. Most likely these vertebrae are from a non-pterodactyloid monofenestratan, identified here as ?Monofenestrata indet., which have garnered considerable attention for possessing a mosaic of features of both pterodactyloids and non-pterodactyloids (Lü et al. 2010), as seen in NHMUK PV R 40126a.
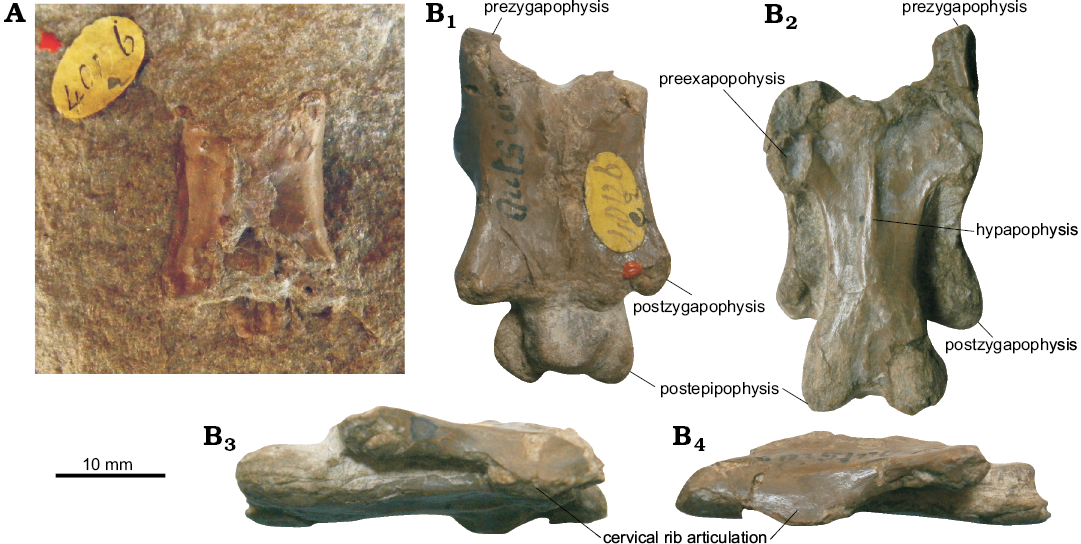
Fig. 19. Isolated vertebrae of monofenestratan pterosaurs from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK. A. NHMUK PV R 40126a (CVA). B. NHMUK PV R 40126a (CVB). In dorsal (A, B1), ventral (B2), right lateral (B3), and left lateral (B4) views.
Metacarpals.—NHMUK PV R 28160b (Fig. 8B) is a 41 mm MCIV with a preserved length/width ratio of 8/1. It has a prominent distal condyle above which is a piece interpreted as either a broken sliver of the shaft of MCIV or the remnants of one of the associated metacarpals I–III. Basal pterosaurs have MCIVs with a maximum length/width ratio of approximately 5:1 whereas pterodactyloids have a length/width ratio of 9:1–20:1 (Wild 1984; Wellnhofer 1991; Martill et al. 2013). Although a ratio of 8:1 in NHMUK PV R 28160b is closer to the pterodactyloid condition, the proximal and distal epiphyses are relatively broader and more like those of basal pterosaurs. The non-pterodactyloid monofenestratan Darwinopterus has a MCIV with a ratio of ~10:1, with a similar constricted medial shaft along with wider proximal and distal epiphyses (Lü et al. 2010). NHMUK PV R 28160b is therefore identified here as ?Monofenestrata indet.
Suborder Pterodactyloidea Plieninger, 1901
?Pterodactyloidea indet.
Fig. 20.
Material.—NHMUK PV R 2637 (Fig. 20) from the Taynton Limestone Formation (Bathonian, Middle Jurassic) of Stonesfield, Oxfordshire, is a synsacrum preserved in dorsal view, 21 mm anteroposteriorly and 14 mm laterally. It is poorly preserved, having been crushed flat and heavily worn. There are at least 4 sacral vertebrae with another preserved anterior to these, although it is uncertain if this is a sacral or a dorsal vertebra. The neural spines have broken and while the transverse processes are all present, they are significantly damaged anteriorly. The transverse processes are approximately 2–3 mm anteroposteriorly with the posterior pairs horizontally oriented while the anterior transverse processes are angled posteriorly at ~160°. The last vertebra is 3 mm anteroposteriorly and its broken transverse processes appear to connect with the illia. They form the anterior margin of a fenestra twice as broad anteroposteriorly as the previous fenestrae. Synsacra are unique to dinosaurs and pterosaurs among Taynton Limestone vertebrates (Unwin 2003; Norman 1988). NHMUK PV R 2637 is identified as pterosaurian based on its smooth and ultra-thin bone walls, and its relatively broad transverse processes. In basal pterosaurs, the sacrum comprises 4 vertebrae (Unwin 2003, 2005). Wukongopterids have at least 5 sacrals (Lü et al. 2011) and the sacrum tends to be broad medially. In pterodactyloids, there are at least 5 sacral vertebrae (Wellnhofer 1978) and several have a large fenestra around the first sacral vertebra delineating the anterior and posterior synsacrum. The pterodactyloid synsacrum is elongate, thinning posteriorly throughout its length (Wellnhofer 1991; Witton 2013). The broad anterior fenestra and the elongate sacrum suggests that NHMUK PV R 2637 may be either a basal monofenestratan with a modified synsacrum (Lü et al. 2010) or an indeterminate pterodactyloid. If the former it would represent the first evidence of a monofenestratan modifying a module other than the skull and neck. If the latter it represents the earliest occurrence of a pterodactyloid extending the temporal range of pterodactyloids down into the Middle Jurassic, before the Callovian–Oxfordian Kryptodrakon progenitor Andres, Clark, and Xu, 2014.

Fig. 20. A synsacrum of ?Pterodactyloidea indet. (NHMUK PV R 2637) from Taynton Limestone Formation (Bathonian, Middle Jurassic), Stonesfield, Oxfordshire, UK; in dorsal view. Photograph (A) and accompanying diagram (B), black regions represent the sacral fenestration.
Non-pterosaurian material
The fragmentary nature of the Bathonian pterosaur assemblage has led to several specimens being misidentified as pterosaur, although most are small isolated specimens of little note. Buffetaut and Jeffreys (2012) described a supposed pterosaur jaw that Andres et al. (2014) later described as thalattosuchian, an identification with which we concur. The most significant misidentified non-pterosaurian fossil is also the most well-known, the holotype of Rhamphocephalus prestwichi.
Thalattosuchia incertae sedis
Genus Rhamphocephalus Seeley, 1880
Type species: Rhamphocephalus prestwichi Seeley, 1880; unknown horizon, Great Oolite Group, Kineton Thorns Quarry, Stow-On-Wold, Gloucestershire.
Included species: Rhamphocephalus prestwichi Seeley, 1880, “Rhamphocephalus” bucklandi (Meyer, 1832) and “Rhamphocephalus” depressirostris (Huxley, 1859).
Rhamphocephalus prestwichi Seeley, 1880
Fig. 21.
1880 Rhamphocephalus prestwichi; Seeley1880: 27–30.
1888 Rhamphocephalus prestwichi Seeley, 1880; Newton 1888: 534.
1888 Rhamphocephalus prestwichi Seeley, 1880; Lydekker 1888: 33.
1901 Rhamphocephalus prestwichi Seeley, 1880; Seeley 1901: 153–155.
1922 Rhamphorhynchus prestwichi (Seeley, 1880); Arthaber 1922: 9.
1978 Rhamphocephalus prestwichi Seeley, 1880; Wellnhofer 1978: 41.
1995 Rhamphocephalus prestwichi Seeley, 1880; Benton and Spencer 1995: 149.
1996 Rhamphocephalus prestwichi Seeley, 1880; Unwin 1996: 293.
2003 Rhamphocephalus prestwichi Seeley, 1880; Unwin 2003: 177.
2012 Rhamphocephalus sedgwicki (sic); Buffetaut and Jeffery 2012: 1.
2013 Rhamphocephalus prestwichi Seeley, 1880; Witton 2013: 125–127.
Holotype: Rhamphocephalus prestwichi (OUM J.28266) (Fig. 21A) poorly preserved cranium presenting a dorsal view of the palate.
Type locality: Kineton Thorns Quarry, Gloucestershire, UK.
Type horizon: Bathonian, Middle Jurassic.
Remarks.—Rhamphocephalus is one of the oldest British “pterosaur” taxa re-interpreted here as a thallatosuchian based on its crocodylomorph skull morphology. The type specimen of Rhamphocephalus prestwichi OUM J.28266 (Fig. 21A) from an unrecorded horizon in the Kineton Thorns Quarry, Stow-On-Wold, Gloucestershire (Fig. 22) is a poorly preserved cranium presenting a dorsal view of the palate. It has been a contested taxon for several years due to the lack of well-supported diagnostic characters (see Introduction). Lydekker’s (1888) characters occur plesiomorphically in several other pterosaur genera (Wellnhofer 1991) and the skull arrangement is, as noted by Seeley (1880), more crocodilian than pterosaurian. When a simplified reconstruction of OUM J.28266 is presented next to the skull of Rhamphorhynchus (Fig. 21B), the differences become apparent. Pterosaur skulls possess large, sometimes medially positioned orbits which are like the larger fenestrae of OUM J.28266 but the nares are positioned more anteriorly along the rostrum. The antorbital fenestrae are proximal to the orbits but are laterally placed and do not significantly invade the dorsal profile of the skull. The rugose striations exposed on OUM J.28266 are absent in pterosaur bone which has a smoother internal texture (Bennett 1993). Given the lack of morphological convergence between OUM J.28266 and pterosaur skulls, it can be confidently identified as non-pterosaurian. Its cranial morphology is instead more like choristodires and thalattosuchians. Figure 23 reconstructs OUM J.28266 in two orientations, one presenting it as a near complete medial skull and the other as an isolated squamosoparietal region. Choristodires usually have extremely broad superior temporal fenestrae and orbits more anteriorly positioned than OUM J.28266 and are therefore not a likely candidate. Thallatosuchian crocodylomorphs such as Steneosaurus Geoffroy, 1825 and Teleosaurus Geoffroy, 1825 more strongly matches OUM J.28266 in either orientation with their large superior temporal fenestra separated by a squamosoparietal bar with a broad ventral base (Walkden et al. 1987), small orbits proximal to the temporal fenestrae and medially position choanae corresponding with the elevated domes of OUM J.28266 (Pierce and Benton 2006). Thus OUM J.28266 is identified as thalattosuchian but the incompleteness of the fossil limits the taxonomic resolution. With no identifiable autapomorphies, the genus Rhamphocephalus cannot be transferred to Thalattosuchia and is therefore regarded as a nomen dubium.
Stratigraphic and geographic range.—The holotype of was collected from the Stonesfield Slate of Kineton, near Stow-on-Wold prior to 1880 (Seeley 1880). Kineton is approximately 7 miles from Stow-on-the-Wold, in Gloucestershire, near the River Windrush (Fig. 22). Fossils were collected from the Kineton Thorns Quarry where exposures of the Cotswold Slate Formation and the Eyeford Member of the Fuller’s Earth Formation underlie the Taynton Limestone Formation (Boneham and Wyatt 1993; Sumbler et al. 2002). The exact origin of OUM J.28266 is unrecorded, however, it is older than the Taynton Limestone Formation pterosaurs (~167 million years old).

Fig. 21. A. The holotype of rhamphorhynchid pterosaur Rhamphocephalus prestwichi Seeley, 1880 (OUM J.28266), from Cotswold Slate Formation Bathonian, Middle Jurassic), Kineton Thorns Quarry near Stow-on-Wold, Gloucestershire, UK; A1, photograph in dorsal view; A2, an engraving as figured by Seeley (1880); A3, drawing of outline. B. Skull of Rhamphorhynchus sp. in dorsal view (modified from Wellnhofer 1975: fig. 3); included to demonstrate the rhamphorhynchid skull arrangemnt.
Fig. 22. Maps showing location of the area of the Kineton Thorns Quarry (asterisks) within the Stow-On-Wold region, Gloucestershire, England, UK. From http://www.cotswoldphotolibrary.com.
Fig. 23. Explanatory drawings of OUM J.28266 along side the skulls of a choristodire and a thallatosuchian. A. The possible arrangement of the Rhamphocephalus prestwichi Seeley, 1880 (OUM J.28266), holotype reconstructed as a complete skull (A1) and the frontoparietal region (A2). B. Simoedosaurus Gervais, 1877, a choristodire from the Early Palaeocene of Saskatchewan, Canada; in dorsal view (from Sigogneau-Russell and Russell 1978). C. Pelagosaurus Bronn, 1841, a thallatosuchian from the Lower Jurassic of Somerset, England; in dorsal view. B, C not to scale.
Discussion
Prior to this study the most diverse Middle–Upper Jurassic pterosaur bearing unit was the Bathonian–Oxfordian Tiaojishan Formation of Liaoning, China. It is a highly lithologically variable unit with 13 named genera (Lü et al. 2010, 2012; Bennett 2014). The assemblage includes groups present in the Great Oolite Group like rhamphorhynchids (3 taxa) and monofenestratans (potentially 7 taxa) as well as 2 anurognathids, a group of highly derived small pterosaurs known for their short robust skulls (Bennett 2007). Unlike the British material, the Tiaojishan pterosaur assemblage is dominated by exceptionally preserved articulated skeletons. With at most 5 potential taxa in the Great Oolite Group the Tiaojishan Formation is clearly the most specifically diverse, but this does not diminish the importance of the British fossils. There is clear continuity between the pterosaur fauna of Europe and Asia, therefore each unit can suggest what taxa are notable by their absence. The presence of anurognathids in the Tiaojishan Formation and the Late Jurassic of Germany and Kazakhstan (Wellnhofer 1991; Witton 2015) suggests the group may have been present in the Bathonian of Britain. Perhaps their absence from the British assemblage can be attributed to their small size reducing their preservation potential.
While not the most diverse Middle Jurassic pterosaur assemblage, the Great Oolite Group presents several important points regarding pterosaur evolution. The monofenestratans of the Tiaojishan Formation are regarded as the earliest occurrence of the group, dating to the Callovian (Witton 2013). The Great Oolite monofenestratan fossils are reliably dated to the Bathonian, ~2 million years older, and thus represent an even earlier appearance for the clade. It is likely that the origins of Monofenestrata are older still with the Early Jurassic Allkaruen Codorniú, et al. 2016 recovered as the sister taxon to the group (Codorniú et al. 2016). Furthermore, the identification of Cuspicephalus Martill and Etches, 2013, as a late surviving wukongopterid (Witton et al. 2015) suggests the basal monofenestratan lineage was well established in Britain throughout the Jurassic. With regards to the potential for pterodactyloids in the assemblage, if NHMUK PV R 2637 can be placed within the group it would place the evolution of Pterodactyloidea several of million years earlier than previously recorded. Presently the oldest pterodactyloid is Liaodactylus Zhou, Gao, Yi, Xue, Li, and Fox, 2017, a ctenochasmatoid from the Oxfordian of the Tiaojishan Formation and Kryptodrakon from the Callovian–Oxfordian boundary in the Shishugou Formation of Xinjiang, China, dating to 160 and 162 ma respectively. This is around 4–6 million years younger than the Great Oolite Group pterosaurs, making NHMUK PV R 2637 the oldest pterodactyloid and strongly suggesting the group has an early Middle or perhaps even Lower Jurassic origin. Unfortunately, NHMUK PV R 2637’s identification is tenuous but further examination will hopefully answer if this is an important fossil in pterosaur evolution.
With regards to the Great Oolite Group and Jurassic pterosaur diversity, it appears to support the findings of previous studies (Butler et al. 2009, 2012, 2013) that the appearance of a sudden radiation in the Late Jurassic (Kimmeridgian–Tithonian) has been somewhat inflated by the poor yield of most old deposits. The unit may possess up to 5 taxa from 3 groups, including the earliest recorded appearance of the monofenestratans. While not directly comparable to the Solnhofen Limestone’s assemblage of 14 species, it is notably higher than other Lower or Middle Jurassic units (e.g., the Whitby Mudstone Formation which has 2 families [O’Sullivan 2017; O’Sullivan and Martill 2017] or the Posidonia Shale Formation which despite being a pterosaur Lagerstätte includes only 2 genera [Padian 2008a, b]). The Bathonian Great Oolite Group strata are separated from the Kimmeridgian “radiation” by more than 10 million years, yet there’s little suggestion that this is a period of low diversity. Further evidence for pterosaurs remaining diverse for much of the Jurassic, as well as the at least partially artificial nature of the Late Jurassic pterosaur radiation, can be found by comparing U.K. formations with the Tiaojishan Formation of Hebel and Liaoning, China (Lü et al. 2010, 2012; Bennet 2014). As discussed above it contains 13 species, at least 9 of occur in Bathonian–Oxfordian aged strata. Clearly this formation had diverse biota comparable with the Great Oolite Group which it maintained right up until the Late Jurassic transition. Yet if we look at the contemptuous Oxford Clay Formation of Britain, a Callovian–Oxfordian unit, we find a signal identifying a period of very low diversity with only a small number of fossils preserved and no well supported taxa (O’Sullivan 2017). The Tiaojishan Formation is a Lagerstätte laid down in a terrestrial environment with a diverse water system, resulting in better preservation and a reasonably high population density (Tan et al. 2006) nature of the Tiaojishan Formation whereas the Oxford Clay Formation is laid down in a marine environment where pterosaurs have a much lower preservation potential and a much lower population density (O’Sullivan 2017). Therefore, it seems likely that as suggested by Butler et al. (2009, 2012, 2013) pterosaur radiation at the end of the Jurassic has been artificially inflated by at least preservational biases. That does not mean that there is no evidence for greater pterosaur taxic diversification in the Late Jurassic. While the Tiaojishan Formation contains 13 species from the Bathonian to the Oxfordian, the Solnhofen Limestone Formation contains that many species within the Kimmmeridgian and Tithonian, a notably shorter time frame. This may still be a result of the Lagerstätten effect, but we should not assume that while pterosaurs were much more diverse in the Middle Jurassic than traditional models may indicate that there wasn’t an increase of some kind in diversity, possibly associated with a transition from a non-pterodactyloid to a pterodactyloid dominated assemblage.
Conclusions
The Middle Jurassic pterosaur assemblage of the UK is not the most diverse assemblage of its time, but it is still important as it highlights the strong phenotypic and specific variance found in the Middle Jurassic. Lacking any truly exceptional fossils, we have identified multiple lineages and morphotypes and the presence of a new genus. With two diverse assemblages on opposite sides of the world, there is now stronger evidence that the Mid Jurassic Pterosaur Gap is an artefact of low yield and poor preservation. Far from their being an evolutionary bottleneck, pterosaurs appear to have been undergoing diversification from at least the Middle and likely the late Lower Jurassic. We would recommend all early to mid-Jurassic pterosaur material would benefit from further examination and that future research take care to recognise the misleading signals of previous diversity studies.
Acknowledgements
David Loydell (University of Portsmouth, UK) is thanked for his support of this project. Lorna Steel (NHMUK), Paul Jeffery (OUM), Louise Neep (GSM), Eliza Howlett (OUM), Matt Lowe (Sedgwick Museum of Earth Sciences, Cambridge, UK), Cindy Howells (NMW), Dino Frey (State Museum of Natural History, Stuttgart, Germany), and Kate Sherburn (MUM) are thanked for collection access. Michael Hanson (Loyola University, Chicago, USA) is thanked for providing information about Waterhouse’s account of NHMUK PV OR 47991 as Pterodactylus raptor, and advice as to the correct etymology for Klobiodon. Steve Vidovic, Mark Witton, David Unwin (all University of Portsmouth, UK), and Liz Martin-Silverstone (University of Bristol, UK) are thanked for useful discussion. The Palaeontographical Society and the Palaeontological Association are both thanked for providing research funding.
References
A.B. 1757. A further account of fossils. Gentleman’s Magazine 27: 122–123.
Andres, B. and Myers, 2013. Lone Star pterosaurs. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 103: 383–398. Crossref
Andres, B. and Norell, M.A. 2005. The first record of a pterosaur from the Early Cretaceous strata of Öösh (Övörkhangai; Mongolia). American Museum Novitates 3472: 1–6. Crossref
Andres, B., Clark, J.M., and Xing, X. 2010. A new rhamphorhynchid pterosaur from the Upper Jurassic of Xinjiang, China, and the phylogenetic relationships of basal pterosaurs. Journal of Vertebrate Paleontology 30: 163–187. Crossref
Andres, B., Clark, J., and Xu, X. 2014 The earliest pterodactyloid and the origin of the group. Current Biology 24: 1011–1016. Crossref
Arkell, W.J. 1933. The Jurassic System in Great Britain. 681 pp. Clarendon Press, Oxford.
Arkell, W.J. 1947. The Geology of Oxford. 267 pp. Clarendon Press, Oxford.
Arthaber, G. 1922. Über Entwicklung, Ausbildung und Austerben der Flugsaurier. Paläontologische Zeitschrift 4: 1–47. Crossref
Aston, M. 1974. Stonesfield Slate. Oxfordshire County Council Department of Museum Services, Publication 5: 1–86.
Barrett, P.M., Butler, R.J., Edwards, N.P., and Milner, A.R. 2008. Pterosaur distribution in time and space: an atlas. Zitteliana B 28: 61–107.
Benton, M.J. and Spencer, P.S. 1995. Fossil Reptiles of Great Britain. Geological Review Series. Vol. 10. 386 pp. Chapman and Hall, London. Crossref
Bennett, S.C. 1993. The ontogeny of Pteranodon and other pterosaurs. Paleobiology 19: 92–106. Crossref
Bennett, S.C. 1996. Year-classes of pterosaurs from the Solnhofen Limestone of Germany: taxonomic and systematic implications. Journal of Vertebrate Paleontology 16: 432–444. Crossref
Bennett, S.C. 2007. A second specimen of the pterosaur Anurognathus ammoni. Paläontologische Zeitschrift 81: 376–398. Crossref
Bennett, S.C. 2014. A new specimen of the pterosaur Scaphognathus crassirostris, with comments on constraint of cervical vertebrae number in pterosaurs. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 271: 327–348. Crossref
Bonde, N. and Leal, M.E.C. 2015. The detailed anatomy of Rhamphorhynchus II: braincase, pneumatics and jaws. Historical Biology 27: 755–770. Crossref
Boneham, B.F.W. and Wyatt, R.J. 1993. The stratigraphical position of the Middle Jurassic (Bathonian) Stonesfield Slate of Stonesfield, Oxfordshire, UK. Proceedings of the Geologists’ Association 1041: 123–136. Crossref
Bronn, H.G. 1841. Über die fossilen Gaviale der Lias-Formation und der Oolithe. Archiv für Naturgeschichte Berlin 8:77–82.
Buckland, W. 1829. On the discovery of a new species of pterodactyle in the Lias at Lyme Regis. Transactions of the Geological Society of London 3: 217–222. Crossref
Buckland, W. 1836. Geology and Mineralogy Considered with Reference to Natural Theology Vol. 1. 618 pp. William Pickering, London. Crossref
Buffetaut, E. and Jeffery, P. 2012. A ctenochasmatid pterosaur from the Stonesfield Slate (Bathonian, Middle Jurassic) of Oxfordshire, England. Geological Magazine 149: 552–556. Crossref
Butler, R.J., Barrett, P.M., Nowbath, S., and Upchurch, P. 2009. Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement. Paleobiology 35: 432–446. Crossref
Butler, R.J., Benson, R.B.J., and Barrett, P.M. 2013. Pterosaur diversity: untangling the influence of sampling biases, Lagerstätten, and genuine biodiversity signals. Palaeogeography, Palaeoclimatology, Palaeoecology 372: 78–87. Crossref
Butler, R.J., Brusatte, S.L., Andres, B., and Benson, R.B.J. 2012. How do geological sampling biases affect studies of morphological evolution on deep time? A case study of pterosaur (Reptilia: Archosauria) disparity. Evolution 66: 147–162. Crossref
Carpenter, K., Unwin, D., Cloward, K., and Miles, C. 2003 A new scaphognathine pterosaur from the Upper Jurassic Morrison Formation of Wyoming, USA. Geological Society, London, Special Publications 217: 45–54. Crossref
Cheng, X., Wang, X., Jiang, S., and Kellner, A.W. 2012. A new scaphognathid pterosaur from western Liaoning, China. Historical Biology 24: 101–111. Crossref
Codorniú, L., Carabajal, A.P., Pol, D., Unwin, D., and Rauhut, O.W.M. 2016. A Jurassic pterosaur from Patagonia and the origin of the pterodactyloid neurocranium. PeerJ 4: e2311. Crossref
Colbert, E.H. 1969 A Jurassic pterosaur from Cuba. American Museum Novitates 2370: 1–26.
Collini, C.A. 1784. Sur quelques Zoolithes du Cabinet d’Histoire naturelle de S.A.S.E. In: Acta Theodoro-Palatinae Mannheim 5 Pars Physica, 58–103. Palatine & de Bavière, Mannheim.
Cope, E.D. 1869. Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Transactions of the America Philosophical Society 14: 1–252. Crossref
Czerkas, S.A. and Ji, Q. 2002. A new rhamphorhynchoid with a headcrest and complex integumentary structures. Feathered Dinosaurs and the Origin of Flight 1: 15–41.
Dalla Vecchia, F.M. 2003. New morphological observations on Triassic pterosaurs. Geological Society London, Special Publications 217: 23–44. Crossref
Dalla Vecchia, F.M. 2009. Anatomy and systematics of the pterosaur Carniadactylus gen. n. rosenfeldi (Dalla Vecchia, 1995). Rivista Italiana di Paleontologia e Stratigrafia 115: 159–188.
Dalla Vecchia, F.M., Wild, R., Hopf, H., and Reitner, J. 2002. A crested rhamphorhynchoid pterosaur from the Late Triassic of Austria. Journal of Vertebrate Paleontology 22: 196–199. Crossref
Delair, J.B. and Sarjeant, W.A. 2002. The earliest discoveries of dinosaurs: the records re-examined. Proceedings of the Geologists’ Association 113: 185–197. Crossref
Evans, S.E., Milner, A.R., Fraser, N.C., and Sues, H.D. 1994. Middle Jurassic microvertebrate assemblages from the British Isles. In: N.C. Fraser and H.D. Sues (eds.), In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods, 303–321. Cambridge University Press, Cambridge.
Fastnacht, M. 2008. Tooth replacement pattern of Coloborhynchus (Pterosauria) from the Lower Cretaceous of Brazil. Journal of Morphology 269: 332–348. Crossref
Fröbisch, N.B. and Fröbisch, J. 2006. A new basal pterosaur genus from the Upper Triassic of the Northern Calcareous Alps of Switzerland. Palaeontology 49: 1081–1090. Crossref
Geoffroy, S. 1825. Recherches sur l’organisation des Gavials, sur leurs affinite’s naturelles desquelles re’sulte la necessite’ d’une autre distribution géneérique: Gavialis, Teleosaurus, Steneosaurus. Mémoires du Muséum National d’histoire 12: 97–155.
Gervais, P. 1877. Enumeration de quelques ossements d’animaus vértebrés recuillis aux environs de Reims par M. Leoine. Journal of Zoology 6: 74–79.
Goldfuss, G. 1831. Beiträge zur Kenntnis verschiedener Reptilien der Vorwelt. Nova Acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum 15: 61–128.
Gradstein, F.M., Ogg, J.G., and Smith, A.G. 2004. A Geologic Time Scale 2004 Vol. 86. 610 pp. Cambridge University Press, Cambridge. Crossref
He, X., Yan, D., and Su, D. 1983. A new pterosaur from the middle Jurassic of Dashanpu, Zigong, Sichuan. Journal of Chengdu College of Geology Supplemental 1: 27–33.
Hone, D.W., Tischlinger, H., Frey, E., and Röper, M. 2012. A new non-pterodactyloid pterosaur from the Late Jurassic of Southern Germany. PloS ONE 7: e39312. Crossref
Hooley, R.W. 1913. On the skeleton of Ornithodesmus latidens; an ornithosaur from the Wealden Shales of Atherfield (Isle of Wight). Quarterly Journal of the Geological Society 96: 372–422. Crossref
Howlett, E.A., Kennedy, K.J., Powell, H.P., and Torrens, H.S. 2017. New light on the history of Megalosaurus, the great lizard of Stonesfield. Archives of Natural History 44: 82–102. Crossref
Howse, S.C.B. 1986. On the cervical vertebrae of the Pterodactyloidea (Reptilia: Archosauria). Zoological Journal of the Linnean Society 88: 307–328. Crossref
Hull, E. 1860. On the Blenheim Iron Ore and the thickness of the Formations below the Great Oolite at Stonesfield, Oxfordshire. Geologist 3: 303–305.
Huxley, T.H. 1859. On Rhamphorhynchus bucklandi a new pterosaurian from the Stonesfield Slate. Quarterly Journal of the Geological Society of London 15: 658–670. Crossref
Ingles, J.M. and Sawyer, F.C. 1979. A catalogue of the Richard Owen collection of palaeontological and zoological drawings in the British Museum (Natural History). Bulletin of the British Museum (Natural History). Historical Series 6: 109–197.
Kaup, J.J. 1834. Versuch einer Eintheilung der Saugethiere in 6 Stämme und der Amphibien in 6 Ordnungen. Isis 3: 311–315.
Kellner, A.W. and Tomida, Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian–Albian), Northeastern Brazil. National Science Museum Monographs 17: 1–137.
Kuhn, O. 1967. Die fossile Wirbeltierklasse Pterosauria. 52 pp. Oben, Krailling.
Laurenti, J.N. 1768. Specimen medicum, exhibens synopsin reptilium emendatam cum experimentis circa venena et antidota reptilium austriacorum. 214 pp. Johann Thomas von Trattner, Vienna. Crossref
Lü, J.C., Unwin, D.M., Jin, X., Liu, Y., and Ji, Q. 2010. Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. Proceedings of the Royal Society of London B: Biological Science 277: 383–389. Crossref
Lü, J.C., Unwin, D.M., Zhao, B., Gao, C., and Shen, C. 2012. A new rhamphorhynchid (Pterosauria: Rhamphorhynchidae) from the Middle/Upper Jurassic of Qinglong, Hebei Province, China. Zootaxa 3158: 1–19.
Lü, J.C., Xu, L., Chang, H., and Zhang, X. 2011. A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, northeastern China and its ecological implications. Acta Geologica Sinica (English Edition) 85: 507–514. Crossref
Lydekker, R. 1888. Catalogue of the fossil reptilia and amphibia in the British Museum (Natural History) Part I: Containing the Orders Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria. 339 pp. Order of the Trustees, London.
Martill, D.M. 2010. The early history of pterosaur discovery in Great Britain. Geological Society, London, Special Publications 343: 287–311. Crossref
Martill, D.M. and Etches, S. 2012. A new monofenestratan pterosaur from the Kimmeridge Clay Formation (Kimmeridgian, Upper Jurassic) of Dorset, England. Acta Palaeontologica Polonica 58: 285–294.
Martill, D.M., O’Sullivan, M., and Newman, C. 2013. A possible azhdarchid pterosaur (Pterosauria, Azhdarchidae) in the Durlston Formation (Early Cretaceous, Berriasian) of southern England. Cretaceous Research 43: 26–39. Crossref
Massare, J.A. 1987. Tooth morphology and prey preference of Mesozoic marine reptiles. Journal of Vertebrate Paleontology 7: 121–137. Crossref
Meyer, H. 1832. Palaeologica zur Geschichte der Erde und ihrer Geschopfe. 560 pp. Verlag von Siegmund Schmerber, Frankfurt am Main.
Meyer, H. 1846. Homoeosaurus maximiliani und Rhamphorhynchus (Pterodactylus) longicaudus: zwei fossile reptilien aus dem kalkschiefer von Solenhofen. 22 pp. Schmerber’schen buchhandlung, Frankfurt.
Newton, E.T. 1888. On the skull, brain and auditory organ of a new species of pterosaurian (Scaphognathus purdoni), from the Upper Lias near Whitby, Yorkshire. Philosophical Transactions of the Royal Society B: Biological Sciences 179: 503–537. Crossref
Norman, D. 1988. The Illustrated Encyclopedia of Dinosaurs. 208 pp. Salamander Books Ltd., London.
O’Sullivan, M. 2017. The pterosaur assemblage of the Oxford Clay Formation (Jurassic, Callovian–Oxfordian) from the U.K. Geological Society, London, Special Publications 455: 171–180. Crossref
O’Sullivan, M. and Martill, D.M. 2015. Evidence for the presence of Rhamphorhynchus (Pterosauria: Rhamphorhynchinae) in the Kimmeridge Clay of the UK. Proceedings of the Geologist’s Association 126: 390–401. Crossref
O’Sullivan, M. and Martill, D.M. 2017. The taxonomy and systematics of Parapsicephalus purdoni (Reptilia: Pterosauria) from the Lower Jurassic Whitby Mudstone Formation, Whitby, U.K. Historical Biology 29: 1–10. Crossref
O’Sullivan, M. and Rigby, M. 2017. The first evidence of a Campylognathoides-like pterosaur in the Toarcian (Lower Jurassic) Whitby Mudstone Formation of Lincolnshire, England. Proceedings of the Geologists’ Association 128: 287–291. Crossref
O’Sullivan, M., Martill, D.M., and Groodcock, D. 2013. A pterosaur humerus and scapulocoracoid from the Jurassic Whitby Mudstone Formation, and the evolution of large body size in early pterosaurs. Proceedings of the Geological Association 124: 973–981. Crossref
Owen, R. 1859a. On a new genus (Dimorphodon) of pterodactyle, with remarks on the geological distribution of flying reptiles. Report for the British Association for the Advancement of Science 28: 97–103.
Owen, R. 1859b. On the vertebral characters of the order Pterosauria, as exemplified in the genera Pterodactylus (Cuvier) and Dimorphodon (Owen). Philosophical Transactions of the Royal Society of London 149: 161–169. Crossref
Owen, R. 1874. Monograph of the fossil Reptilia of the Mesozoic formations. I. Pterosauria. Palaeontographical Society Monograph 1874: 1–14.
Padian, K. 1984a. Osteology and Functional Morphology of Dimorphodon macronyx (Buckland) (Pterosauria: Rhamphorhynchoidea) Based on New Material in the Yale Peabody Museum. 44 pp. Peabody Museum of Natural History, New Haven.
Padian, K. 1984b. Pterosaur remains from the Kayenta Formation (?early Jurassic) of Arizona. Palaeontology 27: 407–413.
Padian, K. 2008a. The Early Jurassic pterosaur Campylognathoides Strand, 1928. Special Papers in Palaeontology 80: 65–107.
Padian, K. 2008b. The Toarcian (Early Jurassic) pterosaur Dorygnathus Wagner, 1860. Palaeontology 80: 1–64.
Parkinson, J. 1811. Organic Remains of a Former World. An Examination of the Mineralized Remains of the Vegetables and Animals of the Antediluvian World; Generally Termed Extraneous Fossil. 508 pp. J. Robson, London. Crossref
Pereda-Suberbiola, X., Knoll, F., Ruiz-Omeñaca, J.I., Company, J., and Torcida Fernández-Baldor, F. 2012. Reassessment of Prejanopterus curvirostris, a basal pterodactyloid pterosaur from the Early Cretaceous of Spain. Acta Geologica Sinica 86: 1389–1401. Crossref
Phillips, J. 1871. Geology of Oxford and the Valley of the Thames. 574 pp. Clarendon Press, London.
Pierce, S.E. and Benton, M.J. 2006. Pelagosaurus typus Bronn, 1841 (Mesoeucrocodylia: Thalattosuchia) from the Upper Lias (Toarcian, Lower Jurassic) of Somerset, England. Journal of Vertebrate Paleontology 26: 621–635. Crossref
Plieninger, F. 1894. Campylognathus Zittelli. Ein neuer Flugsaurier aus dem Oberen Lias Schwabens. Palaeontographica 41: 193–222.
Plot, R. 1677. The Natural History of Oxfordshire, Being an Essay Towards the Natural History of England. 358 pp. The Theater, Oxford.
Ricqlès, A.J., Padian, K., Horner, J.R., and Francillon-Vieillot, H. 2000. Palaeohistology of the bones of pterosaurs (Reptilia: Archosauria): anatomy, ontogeny, and biomechanical implications. Zoological Journal of the Linnean Society 129: 349–385. Crossref
Romer, A.S. 1956. Osteology of the Reptiles. 772 pp. University of Chicago Press, Chicago.
Sayão, J.M. 2003. Histovariability in bones of two pterodactyloid pterosaurs from the Santana Formation, Araripe Basin, Brazil: preliminary results. Geological Society, London, Special Publications 217: 335–342. Crossref
Seeley, H.G. 1870. The Ornithosauria. An Elementary Study of the Bones of Pterodactyls, Made From Fossil Remains Found in the Cambridge Upper Greensand, And Arranged in the Woodwardian Museum of the University of Cambridge. 135 pp. Deighton, Bell & Co, Cambridge.
Seeley, H.G. 1880. On Rhamphocephalus prestwichi, Seeley, an ornithosaurian from the Stonesfield Slate of Kineton. Quarterly Journal of the Geological Society 36: 27–30. Crossref
Seeley, H.G. 1885. Phillips’ Manual of Geology 2nd edition. 571 pp. Charles Griffin & Company, London.
Seeley, H.G. 1901. Dragons of the Air: An Account of Extinct Flying Reptiles. 239 pp. Methuen & Company, London.
Seitz, A.L. 1907. Vergleichende Studien über den mikroskopischen Knochenbau fossiler und rezenter Reptilien. Nova Acta Akademia Caesar Leopold Naturforschung 87: 230–370.
Sellwood, B.W. and McKerrow, W.S. 1974. Depositional environments in the lower part of the Great Oolite Group of Oxfordshire and North Gloucestershire. Proceedings of the Geologists Association 85: 189–210. Crossref
Sellwood, B.W., Scott, J., Mikkelsen, P., and Akroyd, P. 1985. Stratigraphy and sedimentology of the Great Oolite Group in the Humbly Grove oilfield, Hampshire. Marine and Petroleum Geology 2: 44–55. Crossref
Sigogneau-Russell, D. and Russell, D.E. 1978. Etude osteologique du reptile Simoedosaurus (Choristodera). Annales de Paleontologie (Vertebres) 64: 1–84.
Sömmerring, S.T. von 1812. Über einen Ornithocephalus oder über das unbekannten Thier der Vorwelt, dessen Fossiles Gerippe Collini im 5. Bande der Actorum Academiae Theodoro-Palatinae nebst einer Abbildung in natürlicher Grösse im Jahre 1784 beschrieb, und welches Gerippe sich gegenwärtig in der Naturalien-Sammlung der königlichen Akademie der Wissenschaften zu München befindet. Denkschriften der königlichen bayerischen Akademie der Wissenschaften, München: mathematisch-physikalische Classe 3: 89–158
Spix, J. 1817. Ueber ein neues, vermutlich dem Pteropus Vampyrus Linn. zugehöriges Petrifikat aus dem Solenhofer Kalkbruch in Baiern. Denkschriften der Königlichen Akademie der Wissenschaften zu München 6: 60–68.
Stecher, R. 2008. A new Triassic pterosaur from Switzerland (Central Austroalpine, Grisons), Raeticodactylus filisurensis gen. et sp. nov. Swiss Journal of Geosciences 101: 185–201. Crossref
Steel, L. 2012. The pterosaur collection at the Natural History Museum, London, UK: an overview and list of specimens, with description of recent curatorial developments. Acta Geologica Sinica (English Edition) 86: 1340–1355. Crossref
Strand, E. 1928. Miscellanea nomenclatoria zoologica et palaeontologica I–II. Archiv für Naturgeschichte 92A: 40–41.
Sumbler, M.G., Cox, B.M., Wyatt, R.J., and Page, K.N. 2002. The Middle Jurassic stratigraphy of the Cotswolds. Geological Conservation Review Series 26: 113–218.
Tan, J., Ren, D., and Shih, C. 2006. New cupedids from the Middle Jurassic of Inner Mongolia, China (Coleoptera: Archostemata). Annales Zoologici 56: 1–6.
Torrens, H.S. 1980. Bathonian correlation chart. In: J.C.W. Coper (ed.), A Correlation of Jurassic Rocks in the British Isles. Part 2, 21–45. Geological Society, London.
Tumarkin-Deratzian, A.R., Vann, D.R., and Dodson, P. 2007. Growth and textural ageing in long bones of the American alligator Alligator mississippiensis (Crocodylia: Alligatoridae). Zoological Journal of the Linnean Society 150: 1–39. Crossref
Tutin, S.L. and Butler, R.J. 2017. The completeness of the fossil record of plesiosaurs, marine reptiles from the Mesozoic. Acta Palaeontologica Polonica 62 : 563–573. Crossref
Unwin, D.M. 1996 The fossil record of Middle Jurassic pterosaurs. Museum of Northern Arizona Bulletin 60: 291–304.
Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. Geological Society, London, Special Publications 217: 139–190.Unwin, D.M. 2005. The Pterosaurs From Deep Time. 347 pp. Pi Press, New York.
Unwin, D.M. 2015. Non-pterodactyloid monofenestratans—rewriting the evolutionary history of pterosaurs. In: M.P. Witton, D.M. Martill, and D.W. Hone, (eds.), Flugsaurier 2015, 50–52. University of Portsmouth Press, Portsmouth.
Ure, A. 1829. A New System of Geology, in which the Great Revolutions of the Earth and Animated Nature are Reconciled to Modern Science and Sacred History. 702 pp. Longman, London.
Vidovic, S.U. and Martill, D.M. 2014. Pterodactylus scolopaciceps Meyer, 1860 (Pterosauria, Pterodactyloidea) from the Upper Jurassic of Bavaria, Germany: The Problem of Cryptic Pterosaur Taxa in Early Ontogeny. PLoS ONE 9: e110646. Crossref
Wagner, A. 1860. Bemerkungen über die Arten des Flugsaurier. Sitzungs-Berichte der Bayerischen Akademie der Wissenschaften 3: 36–52.
Walkden, G.M., Fraser, N.C., and Muir, J. 1987. A new specimen of Steneosaurus (Mesosuchia, Crocodilia) from the Toarcian of the Yorkshire coast. Proceedings of the Yorkshire Geological Society 46: 279–287. Crossref
Wang, X., Kellner, A.W., Jiang, S., and Meng, X. 2009. An unusual long-tailed pterosaur with elongated neck from western Liaoning of China. Anais da Academia Brasileira de Ciências 81: 793–812. Crossref
Wang, X., Kellner, A.W., Jiang, S., Cheng, X., Meng, X., and Rodrigues, T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. Anais da Academia Brasileira de Ciências 82: 1045–1062. Crossref
Wang, X., Kellner, A.W.A. Jiang, S., Cheng, X., Wang, Q., Ma, Y., Paidoula, Y., Rodrigues, T., Chen, H., Sayão, J.M., Li, N., Zhang, J., Bantim, R.A.M., Meng, X., Zhang, X., Qiu, R., and Zhou, Z. 2017. Egg accumulation with 3D embryos provides insight into the life history of a pterosaur. Science 358: 1197–1201. Crossref
Wang, X., Kellner, A.W.A., Jiang, S., Wang, Q., Ma, Y., Paidoula, Y., Cheng, X., Rodrigues, T., Meng, X., Zhang, J., Li, N., and Zhou, Z. 2014. Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China. Current Biology 24: 1323–1330. Crossref
Waterhouse, G. 1878. Department of Geology. Acquisitions. I. By Donation A. Vertebrata. (3.) Reptilia. In: C.T. Newton (ed.), Account of the Income and Expenditure of the British Museum (Special Trust Funds), for the Financial Year Ended the 31st Day of March 1878: and, Return of the Number of Persons Admitted to Visit the Museum in Each Year From 1872 to 1877, Both Years Inclusive; Together With a Statement of the Progress Made in the Arrangement of the Collections; and an Account of Objects Added to Them in the Year 1877. p. 593–636 (1–44) Great Britain. Parliament. House of Commons. 1878. Accounts and Papers: 1878. Vol. LX. Volume 15. 626 pp. The Queen’s Printer, London.
Weishampel, D.B., Dodson, P., and Osmólska, H. 2004. Introduction. In: D.B. Weishampel, P. Dodson, and H. Osmólska (eds.), The Dinosauria, Second Edition, 1–13. University of California Press, Berkeley. Crossref
Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Suddeutschlands. Bayerische Akademie der Wissenschaften, Mathematisch Wissenschaftlichen Klasse, Abhandlungen 141: 1–133.
Wellnhofer, P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Teil I: Allgemeine Skelettmorphologie. Paläontographica A 148:1–33.
Wellnhofer, P. 1978. Pterosauria. In: G. Fischer (ed.), Handbuch Der Paläoherpetologie. 82 pp. Fischer, Stuttgart.
Wellnhofer, P. 1991. The Illustrated Encyclopedia of Pterosaurs. 192 pp. Salamander Books Ltd., London.
Wild, R. 1984. A new pterosaur (Reptilia, Pterosauria) from the Upper Triassic (Norian) of Friuli, Italy. Gortiana Atti Museo Friuliano di Storia Naturale 5: 45–62.
Witton, M.P. 2013. Pterosaurs: Natural History, Evolution, Anatomy. 291 pp. Princeton University Press, Princeton. Crossref
Witton, M.P. and Naish, D. 2008 A reappraisal of azhdarchid pterosaur functional morphology and paleoecology. PLoS ONE 3: e2271. Crossref
Witton, M.P., O’Sullivan, M., and Martil, D.M. 2015. The relationships of Cuspicephalus scarfi Martill and Etches, 2013 and Normannognathus wellnhoferi Buffetaut et al. 1998 to other monofenestratan pterosaurs. Contributions to Zoology 84: 115–127.
Woodward, H.R. 1894. The Jurassic Rocks of Britain Vol. 4. The Lower Oolitic Rocks. 628 pp. HMSO, London.
Wyatt, R.J. 1996. A correlation of the Bathonian (Middle Jurassic) succession between Bath and Burford, and its relation to that near Oxford. Proceedings of the Geologists’ Association 107: 299–322.
Young, G.C. 1964. On a new pterosaurian from Sinkiang, China. Vertebrata PalAsiatica 8: 221–255.
Zambelli, R. 1973. Eudimorphodon ranzii gen. nov., sp. nov. Uno Pterosauro Triassico. Rendiconti Instituto Lombardo Accademia 107: 27–32.
Zhou, C.F. 2014. Cranial morphology of a Scaphognathus-like pterosaur, Jianchangnathus , based on a new fossil from the Tiaojishan Formation of western Liaoning, China. Journal of Vertebrate Paleontology 34: 597–605. Crossref
Zhou, C.F. and Schoch, R.R. 2011. New material of the non-pterodactyloid pterosaur Changchengopterus pani Lü, 2009 from the Late Jurassic Tiaojishan Formation of western Liaoning. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 260: 265–275. Crossref
Zhou, C.F., Gao K.Q., Yi,H., Xue, J., Li, Q., and Fox R.C. 2017 Earliest filter-feeding pterosaur from the Jurassic of China and ecological evolution of Pterodactyloidea. Royal. Society Open Science 4: 160672. Crossref
Acta Palaeontol. Pol. 63 (4): 617–644, 2018
https://doi.org/10.4202/app.00490.2018