
Synopsis of the terrestrial vertebrate faunas from the Middle Kura Basin (Eastern Georgia and Western Azerbaijan, South Caucasus)
MAIA BUKHSIANIDZE and KAKHABER KOIAVA
Bukhsianidze, M. and Koiava, K. 2018. Synopsis of the terrestrial vertebrate faunas from the Middle Kura Basin (Eastern Georgia and Western Azerbaijan, South Caucasus). Acta Palaeontologica Polonica 63 (3): 441–461.
This paper summarizes knowledge on the Neogene–Quaternary terrestrial fossil record from the Middle Kura Basin accumulated over a century and aims to its integration into the current research. This fossil evidence is essential in understanding the evolution of the Eurasian biome, since this territory is located at the border of Eastern Mediterranean and Central Asian regions. The general biostratigraphic framework suggests existence of two major intervals of the terrestrial fossil record in the area, spanning ca. 10–7 Ma and ca. 3–1 Ma, and points to an important hiatus between the late Miocene and late Pliocene. General aspects of the paleogeographic history and fossil record suggest that the biogeographic role of the Middle Kura Basin has been changing over geological time from a refugium (Khersonian) to a full-fledged part of the Greco-Iranian province (Meotian–Pontian). The dynamic environmental changes during the Quaternary do not depict this territory as a refugium in its general sense. The greatest value of this fossil record is the potential to understand a detailed history of terrestrial life during demise of late Miocene Hominoidea in Eurasia and early Homo dispersal out of Africa. Late Miocene record of the Middle Kura Basin captures the latest stage of the Eastern Paratethys regression, and among other fossils counts the latest and the easternmost occurence of dryopithecine, Udabnopithecus garedziensis, while the almost uninterrupted fossil record of the late Pliocene–Early Pleistocene covers the time interval of the early human occupation of Caucasus and Eurasia.
Key words: Mammalia, Aves, Reptilia, Miocene, Quaternary, South Caucasus, Kura Basin.
Maia Bukhsianidze [maiabukh@gmail.com], Georgian National Museum, 3, Purtseladze street, Tbilisi 0105, Georgia.
Kakhaber Koiava [koiava_ka@yahoo.com], Ivane Javakhishvili Tbilisi State University, Alexandre Janelidze Institute of Geology, A. Politkovskaia str. 31, Tbilisi 0186, Georgia.
Received 29 April 2018, accepted 13 July 2018, available online 22 August 2018.
Copyright © 2018 M. Bukhsianidze and K. Koiava. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The middle part of the Kura Basin (Fig. 1), the territory in the Southern Caucasus between the Mtkvari (Kura) and Alazani rivers, including the vicinities of the Mingachevir Reservoir (eastern Georgia and the western Azerbaijan), is remarkable because of its long stratigraphic sequences preserving an outstandingly rich terrestrial fossil record. Understanding the natural history of this region is integral to understanding the evolution of the Eurasian biome in general and the timing and modalities of faunal exchanges between Europe, Asia, and, to some extent, Africa.
Significant terrestrial fossil evidence has accrued from the Middle Kura Basin since the first mention of fossil vertebrates from the Iori river valley (Riabinin 1913). Terrestrial mammal sites found here cover a time span from the late Miocene through the Quaternary.
The occurrence of a dryopithecine (Udabnopithecus garedziensis Burchak-Abaramovich and Gabashvili, 1945) at the Udabno Site (late Miocene, eastern Georgia; Burchak-Abaramovich and Gabashvili 1945, 1946; Gabunia et al. 2001) makes this place particularly interesting for the evolutionary history of late Miocene hominoids. In addition, the discovery of early Homo at Dmanisi (Early Pleistocene, 1.8 Ma, Southern Georgia; Lordkipanidze et al. 2007; Ferring et al. 2011), although not within the discussed territory, adds to the importance of this Miocene–Quaternary faunal evidence since the Kura Basin record covers the time interval of the early human occupation of Caucasus and Eurasia.
Regrettably, this fossil record is not yet sufficiently integrated into the current knowledge. Several challenging factors have so far inhibited a large-scale integration of this knowledge: the majority of the publications are in Russian and local languages; most of the fauna is known only from preliminary identifications; and the faunal lists are outdated and bear a certain degree of scientific “endemism” due to insufficient comparisons.
Better time resolution is necessary for the accurate interpretation of this evidence, especially since the interior position of this territory within the Kura Basin suggests diachroneity of the Quaternary geological events that occurred in the region (Forte et al. 2015). The existing correlation framework is based mainly on the marine record and lithostratigraphy, while the biostratigraphic research on terrestrial faunas is insufficient; absolute dates are available only for the Akchagylian, Apsheronian, and Bakunian sediments in this area (Early–Middle Pleistocene; Kushkuna, Kvabebi, and Duzdag sites; see Chumakov et al. 1992 and references therein); most of the magnetostratigraphic works/correlations date back to the late 1980s (Nikiforova et al. 1982; Pevzner 1982; Isaeva and Mamedov 1989; Ganzej 1987; Vekua and Trubikhin 1988; Vangengeim et al. 1989; works of Trubikhin in Vekua 1991 and Filippova 1997; Chumakov et al. 1992; Gurarij et al. 1995; and others) and need more detailed research; recent magnetostratigraphic data are published only for one fossil vertebrate site (Kvabebi; Agustí et al. 2009).
This paper discusses all the terrestrial mammal sites from the Middle Kura Basin and provides a general biostratigraphic framework of the record. The European biochronological zonation follows Hilgen et al. (2012); correlations of the East European faunal complexes (Gromov 1948) are according to Vislobokova (2008), Vislobokova and Tesakov (2013). As a narrative review, the paper offers summaries of the existing paleontological literature, while also mentioning for the first time some sites whose collections are housed in the S. Janashia Museum of Georgia, Tbilisi (Georgian National Museum), in order to show the full extent of the terrestrial fossil record. This is not a taxonomic study and the faunal lists for the new sites are preliminary; while the faunal lists for the published sites are based on the literature, preliminary identifications of large mammals of unpublished sites are by M. Bukhsianidze, unless mentioned otherwise. We fully acknowledge the need of taxonomic revisions, which is a long-term future task. This paper is rather a critical summary of what is known about the fossil faunas from this part of Eurasia. As such, it is meant to serve as a starting point for future work.
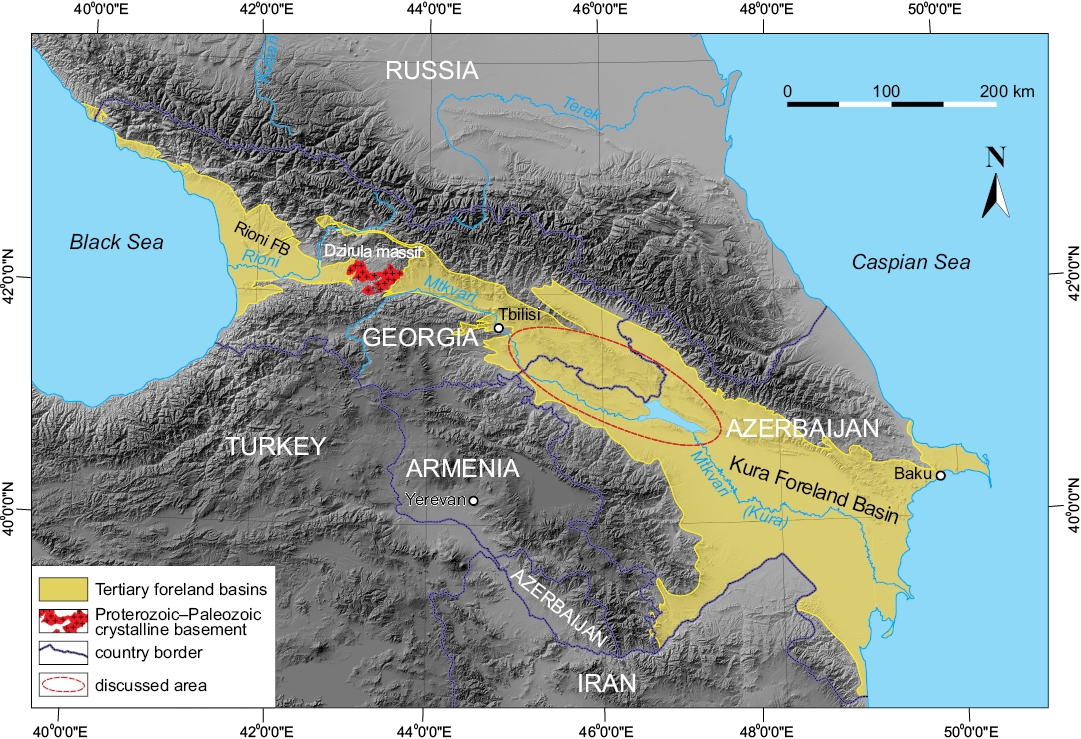
Fig. 1. Map of the Caucasus region showing the Tertiary foreland basins. Modified after Mauvilly et al. (2016); Mauvilly (2017). Topographic map from the United States Geological Survey (USGS) earth explorer web-based platform https://earthexplorer.usgs.gov. FB, foreland basin.
Paleogeographic history
The paleogeography of the Middle Kura Basin has undergone significant transformations through time as it was subject to extensive uplift and subsidence, repeatedly changing from marine to continental settings.
In tectonic terms, this area belongs to the Kura Foreland (Fig. 1), the eastern intermountain depression between Greater and Lesser Caucasus orogene delimited from the west by the Dzirula Crystalline Massif (Adamia et al. 2010; Forte et al. 2010; Nemčok et al. 2013; Gamkrelidze et al. 2015; Alania et al. 2016). The Oligocene–early Miocene Kura Foreland Basin was transformed into a fold and thrust belt as a result of compressional deformation, which started in the middle Miocene and reached its maximum rate at the end of the late Miocene (Adamia et al. 2010; Maisadze 2013; Alania et al. 2016).
During the middle Miocene, the Kura and the Rioni Foreland basins (the western intermountain depression) were covered by the Paratethys Sea and were still connected by the Djava-Tedeletian Strait that passed north of the Dzirula Massif (Buleishvili 1960; Koiava et al. 2012).
During the late Miocene, with the onset of the Attic Orogenic Phase at the end of the middle Sarmatian sensu lato (Bessarabian substage, ca. 10 Ma), the Djava-Tedeletian Strait disappeared as a result of further uplift of the Dzirula Massif (central part of the Likhi, or Surami Mountain Range, a sub-meridional mountain range connecting Greater and Lesser Caucasus) and the Southern Caucasus became irreversibly separated into western and eastern parts. This is an important datum for the natural history of the Southern Caucasus. It marks the onset of the Colchis refugium in the west with humid climate, while relatively arid-continental climatic conditions started to develop in the east (Shatilova et al. 2011). The formation of the Talysh–Hyrcan refugium of ancient mesophilous forests of Tertiary flora along the Alborz Mountain Range (including Talysh Mountains and Lenkoran Lowland) on the southern coast of the Caspian Sea most likely dates back to the same geological epoch (Grossheim 1948; Safarov 1979).
The first continental deposits on the Middle Kura Basin appear in Bessarabian. However, there is no evidence of terrestrial fauna, yet. The Attic Orogenic Phase reached its maximum in the Khersonian. By the end of the Khersonian, this territory became a coastal zone of the Kura Bay (westward extension of the South Caspian Basin) covered with mosaic of isolated and semi-isolated basins, i.e., lakes, lagoons, and large river network forming deltas (Koiava et al. 2012). Intensification of the uplift was accompanied by rapid sea regression; as a result, coastal and shallow marine deposits became gradually dislocated to the east (Buleishvili 1960; Nemčok et al. 2013). This gradual transition from continental to marine facies is observable in the geological sections along the right bank of the Iori river outcrops (Fig. 2): the oldest terrestrial mammal faunas are found along the southern zone of the Kura foreland on the Udabno syncline, and the Akhtatapa (same as Akhtakhtatapa) and Ellyar-Oyugi anticlines.
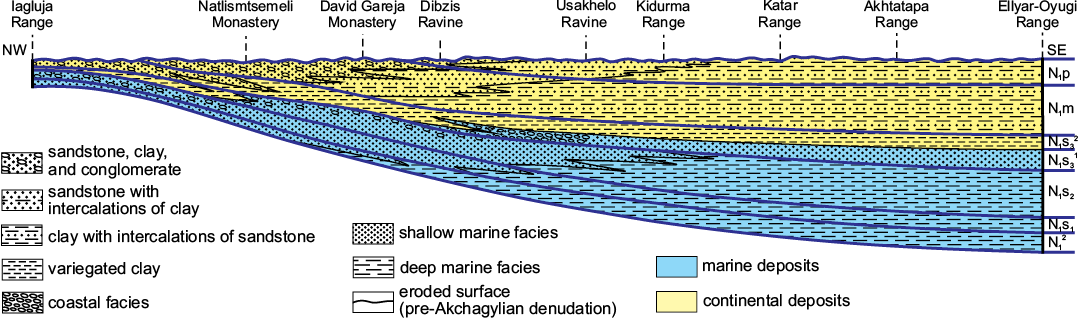
Fig. 2. The scheme of migration of the lithofacies of the Miocene deposits of South Kakheti (Middle Kura Basin), adapted from Buleishvili (1960). The Khersonian continental facies, variegated clay of the Eladri Formation, are the thickest in the south-western part of the area; the transition of this continental formation into marine deposits can be traced starting from the Ravine Dibzis; and the marine deposits are the thickest at the mountain Ellyar-Oyugi. Abbreviations: N1p, Pontian; N1m, Meotian; N1s32, upper Khersonian (upper part of the upper Sarmatian); N1s31, lower Khersonian (lower part of the upper Sarmatian); N1s2, Bessarabian (middle Sarmatian); N1s1, Volhynian (lower Sarmatian) ; N12, Tarkhanian, Chokrakian, Karaganian, Konkian (middle Miocene).
The consequent Meotian and Pontian stages (late Miocene) in this part of the Kura Foreland are represented by the continental Shiraki Formation, a huge succession of clay and sandstone deposited in calm conditions with slow transportation of terrigenous material (Chubinishvili 1982; Gurarij et al. 1995). This territory was subject to important subsidence at those times, especially in the northern part where the Shiraki Formation is the thickest (2900 m; Buleishvili 1960). The variation of thickness of the Shiraki Formation is due not only to the different magnitude of subsidence but also is a result of pre-Akchagylian denudation. The magnitude of erosion increases from east to west, as Akchagylian marine deposits cover increasingly older strata (Meotian–Pontian through Maikopian).
The detachment from the Greater Caucasus and the horizontal displacement of the olistolites of Upper Jurassic limestones in the northern zone of the Kura Basin likely took place at the end of the Miocene during Pontian times (Buleishvili 1960; Dzigrashvili and Sepashvili 1978). The Dedoplistskaro, or Krasnokolodskaya, Formation (Dedoplistskaro is a present day and historical toponym of the place), distinguished by the abundance of Jurassic limestone pebbles, boulders, and larger blocks, is related to this geological phenomenon. It represents the local facies of the uppermost part of the Shiraki Formation developed in the Dedoplistskaro area (Buleishvili 1960; Chubinishvili 1982).
The Shiraki Formation has a wide distribution in the area, yet there are rather few terrestrial mammal faunas found within this formation, most likely due to insufficient explorations.
The presence of continental deposits of early Pliocene age in the Georgian part of the area is uncertain, but such deposits are present in the immediately adjacent territory of Azerbaijan (productive series in Azerbaijan; this formation broadly corresponds to Balakhanian–Kimmerian of the Eastern Paratethys).
Starting from the late Pliocene, the Caspian Sea periodically extended into the Kura Foreland and the continental settings present here since the late Miocene again changed into marine and coastal environments. The first of these transgressions, the Akchagylian, at the Gauss-Gilbert reversal (date for the western Turkmenistan; Trubikhin 1977) and ca. 3.2 Ma (date for eastern Azerbaijan; Van Baak et al. 2013), was the most powerful. Due to maximum subsidence, the southern and central zones of the Kura Foreland became entirely covered by the sea, including the highest ranges of the Iori uplift (Aladjiri-Chobandag-Akhtatapa), controlling the distribution of facies before the Akchagylian (Buleishvili 1960); simultaneously, continental deposits of significant thickness accumulated in the northern zone (Alazani Formation). Consequent Apsheronian (ca. 2 Ma) and Bakunian (ca. 0.9 Ma) transgressions are evidenced in the Kura Foreland (Buleishvili 1960; Lebedeva 1978). The later major transgressions (Khazarian, Khvalynian, Novocaspian) did not reach the middle part of the Kura Basin. Terrestrial mammal faunas from the late Pliocene and onwards make up the largest group of fossil sites in the area.
In general, the discussed territory has remarkably thick exposures of late Neogene and Quaternary molasse deposits due to the high magnitude of compressional deformation in the Kura Foreland Basin. The total compression of the Kura Foreland Basin from the middle Miocene to date is about 35 km (44%; Forte et al. 2010). But what makes this territory particularly unique is that the compressional movements were especially intense during the Quaternary, which resulted in Quaternary deposits and exposures of unprecedented thickness, represented by intercalations of continental and shallow marine deposits rich with fossil remains of flora and fauna.
As mentioned above, the land on the Middle Kura Basin starts to emerge in the late Miocene (end of Bessarabian onwards); since that time it has always been a lowland with low elevations (now varying between 90–900 m) trapped between the high mountain chains and periodically inundated by the sea. The paleogeographic picture indicates that new elements in the terrestrial fauna were entering the South Caucasus region from the south, south-east, and south-west. This is especially true for the Khersonian terrrestrial faunas; in the later periods a possibility for faunal exchange from the north certainly existed.
Biogeography
From the biogeographic point of view, this lowland is located at the northern margin of the junction of two ancient lands, Aegean–Anatolian, or Eastern Mediterranean, and Central Asian. For example, the present-day distribution patterns of a number of species, especially amphibians and reptiles, in the Alborz Mountains, south of the Caspian Sea, and in the Lesser Caucasus reveal not only climatic and landscape changes, but also continental drift: organisms inhabiting west of the Karabagh Mountains are related to Mediterranean species, and those inhabiting east are related to Asian species (Tarkhnishvili 2014, references therein).
Now, the Middle Kura Basin is characterized by a dry continental climate and is covered by semi-desert, salt and gypsum steppes, xerophitic scrub, arid open deciduous woodlands, and riparian forests (Akhalkatsi and Tarkhnishvili 2012). The vegetation falls broadly into the Irano-Turanian group (Zohary 1973). The composition of the flora is influenced by the arid Eastern Mediterranean, Anatolian, and Irano-Turanian provinces (Gagnidze and Davitadze 2000), and the occurrence of oriental and Ethiopian (or Palaeotropical) taxa is remarkable among the mammals (Indian crested porcupine, leopard, and striped hyaena). This kind of community of plants and animals contrasts with the warm and humid Colchis lowland to the west, and the mountainous biomes of the South Caucasus (well developed mountain forests, grasslands, and rocky habitats; more details in Akhalkatsi and Tarkhnishvili 2012).
In the paleontological narrative, the Middle Kura Basin is usually depicted as an isolated place and refugium of large mammals, especially for the Miocene and late Pliocene epochs (e.g., Vekua 1972; Gabunia et al. 2001; Eronen and Rook 2004; Agustí et al. 2009; Nargolwalla 2009). The paleogeographic situation, however, does not portray the Middle Kura Basin as an isolated place. Here, the Mediterranean realm meets Central Asia and thus, naturally, earlier appearance of Asian mammals than in Europe is expected. Any out of Africa faunal dispersal in Eurasia could have left a trace in the South Caucasus, because the Caucasus, as a part of the Alpide belt, represents a natural climatic shield and this area acts as a natural hub for dispersals of terrestrial fauna.
Overview of fossil terrestrial mammal sites
Excluding the sites with a single taxon (four sites), fourteen terrestrial vertebrate sites are found in the Kura Foreland (Fig. 3; two localities with fossil vertebrates, without mentioning taxa, are also indicated). For better clarity the sites are grouped chronologically within geological formations. (i) The earliest faunas are found in the Khersonian marine and continental (Eldari) formations: Eldari, Chachuna, and Udabno. The Iagluja (or Rustavi) fauna from the Miocene continental deposits is also discussed with this group as it belongs to the same sedimentary basin and it is biostratigraphically close to the aforementioned sites. (ii) The second group of late Miocene sites comes from the Shiraki Formation: Dzedzvtakhevi, Vashlovani, Tetri Udabnos Seri, and Dzhaparidze, with the latter from the Dedoplistskaro Formation. (iii) The third, and largest, group are the late Pliocene to Early and Middle Pleistocene sites from the Akchagylian, Apsheronian, and Bakunian deposits: Kushkuna, Kvabebi, Kotsakhuri, Palantokan, and four sites with single proboscidean finds: Duzdag, Taribana, Godjashen, and Yanikend; a Pliocene site from the Alazani Formation: Zemo Melaani; and a Middle Pleistocene site from the Alazani river terrace: Kvemo Kedi.
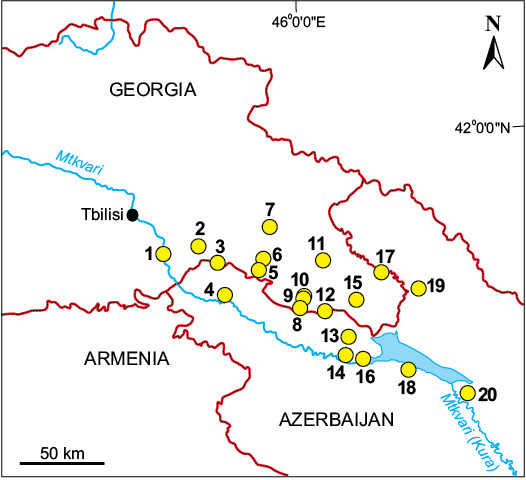
Fig. 3. Fossil terrestrial mammal sites in Middle Kura Foreland Basin. 1, Iagluja; 2, Tetri Udabnos Seri; 3, Udabno; 4, Kushkuna; 5, Dzedzvtakhevi; 6, Kvabebi; 7, Zemo Melaani; 8, Chachuna; 9, Kotsakhuri; 10, Taribana; 11, Dzhaparidze; 12, Eldari; 13, Palantokan; 14, Yenikend; 15, Vashlovani; 16, Qirmizi Samukh; 17, Kvemo Kedi; 18, Duzdag; 19, Almaly; 20, Karadja.
Late Miocene
Sites from the Khersonian marine and continental deposits.—As already mentioned, the late Miocene sites, Iagluja, Udabno, Chachuna, and Eldari (listed from west to east, Fig. 3), are found in the same sedimentary basin along the Kura paleo-bay of the Eastern Paratethys. The continental regime was present in the west earlier than in the east: in the Iagluja Mountain area since late Miocene (Chelidze 1955), in Udabno since the end of Bessarabian onwards (Buleishvili 1960), and in the Chachuna and Eldari continental facies from the upper part of Khersonian (Vangengeim et al. 1989; Fig. 2). The transition between the Eldari and Shiraki Formations is gradual. In Chachuna, Khersonian malacofauna is present in the upper part of the section in brackish water deposits (Bukhsianidze et al. 2018). This defines the upper biostratigraphic limit of this site as Khersonian. Biostratigraphic delimitation of the upper boundary (Khersonian–Meotian boundary) is more problematic for the remaining three sites (Iagluja, Udabno, and Eldari).
Eldari site: Late Miocene, marine Khersonian and Eldari Formation; late Vallesian, early Turolian, MN10–MN11; located on the mountain Ellyar-Oyugi, right bank of the Iori river, Samukh district, Ganja-Qazakh region, Azerbaijan; discovered by Boris S. Dombrovskij in 1913 (Dombrovskij 1914). Composite list of fauna in Table 1.
The site is divided into three lithostratigraphic units, each of them with fossil-bearing horizon/horizons. The order of the numbering of these units/horizons (Table 2) varies from author to author.
Table 1. Composite faunal list of Eldari. The list is mainly based on Gadzhiev 1996; reptiles are from Bakradze and Chkhikvadze 1984; Chkhikvadze and Amiranashvili 1996; birds from Burchak-Abramovich and Gadzhiev 1978; Hipparion garedzicum in the second fossil-bearing horizon from Vangengeim et al. 1989.
|
Reptilia |
|
Testudo eldarica Khozatsky and Alekperov, 1978 Testudo burtschaki Chkhikvadze, 1975 Mauremys sarmatica (Purschke, 1885) |
|
Aves |
|
Struthio sp. Anser eldaricus Burchak-Abramovich, 1978 |
|
Mammalia |
|
Lagomorpha Lepus sp. |
|
Rodentia Dipoides eldaricus Bendukidze and Burchak-Abramovich, 1990 |
|
Carnivora Phoca procaspica Gadzhiev, 1961 Ichtitherium hipparionum Gervais, 1859 Hyaena eldarica Bogachev, 1937 |
|
Proboscidea Deinotheirum giganteum Kaup, 1829 Choerolophodon pentelici (Gaudry and Lertet, 1856) Tetralophodon longirostris (Kaup, 1832) |
|
Perissodactyla Hipparion eldaricum Gabunia, 1959 Hipparion garedzicum Gabunia, 1959 Hipparion sp. (a) Hipparion sp. (b) Eldarotherium burtchaclensis Gadzhiev, 1961 Diceros gabuniai Tsiskarishvili, 1987 Chilotherium (Acerorhinus) aff. zernovi Borissiak, 1914 Chilotherium (Subchilotherium) eldaricum Tsiskarishvili, 1987 Rhinoceros pachygnathus Wagner, 1848 |
|
Artiodactyla Microstonyx major Gervais, 1851 Cervavitus sp. Palaeotragus (Achtiaria) borissiaki Alexeev, 1930 Palaeotragus sp. Helladotherium sp. Mirabilocerus azerbajdjanicus Gadzhiev, 1996 (?= Tragocerus frolovi? Pavlowa, 1914 var. eldaricus Gabashvili, 1956) Tragocerus leskevitszi Borissjak, 1914 Tragocerus sp. Eotragus (?) martinianus (Lartet, 1851) Gazella leili Gadzhiev, 1996 |
Table 2. Numbering of the lithostratigraphic units with fossil-bearing horizons of the Eldari site according to different authors.
|
Formations |
Dombrovskij 1914; Andrianov and Larin 1935; Gadziev 1996 |
|
|
Eldari |
|
Eldari 3 |
|
Eldari 1 |
Eldari 2 |
|
|
Marine Khersonian |
Eldari 2 |
Eldari 1 |
|
Marine Bessarabian |
Eldari 3 |
|
(i) The lowermost fossil-bearing horizon, with remains of marine mammals, is exposed on the foothills of the southern slope of the Ellyar-Oyugi and extends 130 m in an EW direction. This horizon is dated to the Bessarabian by the benthic foraminifera (identifications by Maissuradze in Vangengeim et al. 1989). This lowermost horizon, with pinnipeds and cetacean remains, is not considered to be a part of the Eldari fauna.
(ii) The middle horizon, the major fossil-bearing horizon, is 1.5–2 m thick with lens-like accumulations of vertebrate fossils that are exposed on the southern slope, and extends for 5 km. This horizon is found in the marine Khersonian deposits 70 m above the base of the 30 m thick petroliferous sandstone. The marine Khersonian deposits contain Mactra caspia (Eichwald, 1829) and Solen subfragilis Eichwald, 1830 in the lower part and M. caspia, M. crassicollis Sinzov, 1897 and M. bulgarica (Toula, 1909) in the upper part (mollusc taxa from Vangengeim et al. 1988, 1989).
(iii) The uppermost horizon extends for 3 km on the eastern slope of the Ellyar-Oyugi without any important fossil concentration. Some publications mention that: there are two fossil-bearing horizons in this upper interval of the section (Vangengeim et al. 1989); these horizons are found in the uppermost part of the Eldari Formation; their age is considered to be either terminal Khersonian (Dombrovskij 1914; Kudrjavtsev 1938; Buleishvili 1960; Chubinishvili 1982), or the latest Khersonian–beginning of Meotian (Vangengeim et al. 1989; Gadzhiev 1996).
The Eldari land fauna was traditionally analysed together; many described specimens are without indication of the horizon.
Udabno site: Late Miocene, Eldari and Shiraki formations, late Vallesian, early Turolian, MN10–MN11; located on the right bank of the Iori river near David Gareji monastery, Sagaredjo region, Kakheti, Georgia; discovered in 1931 by Nikolaj A. Gedroiz while mapping the area. Composite list of fauna from Udabno in Table 3.
The Udabno site has a considerable EW extension (ca. 20 km), and fossils are found in several localities: David Gareji (Eldari Formation); Natlismtsemeli (Eldari and Shiraki formations); Adjia, the latter mentioned as Dibsis in Chkhikvadze et al. 2000 (Shiraki Formation), and Dodo (Shiraki Formation). The fossils come from different stratigraphic levels and the entire Udabno site is subdivided into several units. Different authors provide different subdivisions (Table 4). At the base of the Udabno section, middle Sarmatian (Bessarabian) sandy-clayish shallow marine/continental sediments are exposed. Malacofauna from these deposits are represented by the following forms (identification by Muskhelishvili in Vangengeim et al. 1989): Mactra vitalina (D’Orbigny, 1844), Cardium fittoni (D’Orbigny, 1845), Cerastoderma obsoletum ingratum (Kolesnikov, 1929), Paphia vitaliana mediosarmatica (Andrusov, 1917). The Bessarabian sediments are followed by the variegated clays with layers of sandstone and conglomerate of the Eldari Formation, which is conformably overlaid by the Shiraki Formation.
Table 3. Composite faunal list of Udabno. The list is compiled from: Gabunia et al. 2001; Gabunia 1959; Vangengeim et al. 1989; Meladze 1985; Chkhikvadze and Amiranashvili 1996; Chkhikvadze et al. 2000, Tsiskarishvili 1987; Bakradze and Chkhikvadze 1984; Burchak-Abramovich and Gabashvili 1945, 1946, 1980, 1984; Burchak-Abramovich 1957; Gabashvili 1956, 1983, 1987, 2002; Bendukidze et al. 2013; Bendukidze and Burchak-Abramovich 1990; Semenov 1989.
|
Reptilia |
|
Testudo eldarica Khosatzky and Alekperov, 1978 Testudo sp. Megalochelys sp. Trionyx sp. Mauremys sarmatica Purschke, 1885 cf. Centrochelys (= cf. Ergilemys) sp. |
|
Aves |
|
Anser udabnensis Burchak-Abramovich, 1957 Larus udabnensis Burchak-Abramovich, 1984 |
|
Mammalia |
|
Primates Dryopithecus garedziensis (Burchak-Abramovich and Gabashvili, 1945) |
|
Rodentia Palaeomys sp. Steneofiber caucasicus Burchak-Abramovich and Gabashvili, 1980 |
|
Carnivora Simocyon sp. Plesiogulo cf. brachygnathus (Schlosser, 1903) Adcrocuta eximia (Roth and Wagner, 1854) Percrocuta gigantea (Soria, 1980) Miohyaenotherium
bessarabicum (Simonescu, 1937)
Machairodontinae indet. |
|
Proboscidea Deinotherium giganteum Kaup, 1829 Tetralophodon cf. longirrostris Kaup, 1832 |
|
Perissodactyla Hipparion cf. eldaricum Gabunia, 1959 Hipparion garedzicum Gabunia, 1959 Hipparion sp. (aff. garedzicum) Chalicotheriidae (cf. Ancylotherium) Aceratherium cf. incisivum Kaup, 1832 Diceros sp. |
|
Artiodactyla Microstonyx erymanthius (Roth and Wagner, 1854) Cervidae indet. Giraffidae indet. Palaeotragus (Achtiaria) sp. Palaeotragus roueni (Gaudry, 1861) Udabnocerus georgicus Burchak-Abramovich and Gabashvili, 1969 Tragocerus aff. amaltheus var. rugosifrons Schlosser, 1904 Tragocerus sp. Gazella schlosseri Pavlow, 1913 Gazella deperdita (Gervais, 1847) |
Table 4. Numbering of the fossil-bearing horizons of the Udabno site according to different authors.
|
Formations |
|||
|
Shiraki |
Udabno 4 |
|
|
|
Eldari and Shiraki |
|
|
Natlismtsemeli 2 |
|
Udabno 3 |
Udabno 2 |
Udabno 2 |
|
|
Eldari |
Udabno 2, Natlismtsemeli lens |
Udabno 1 |
Udabno 1, Natlismtsemeli 1 |
|
Udabno 1 |
|
|
Regrettably, most of the Udabno fossils lack quality provenance information (but the exact place of finding for Udabnopithecus garedziensis is known); only few fossils were described and illustrated from the site, and it seems that taxa names used to be changed without proper revision from list to list. The lists provided by various authors differ (Tables 5, 6) and, if taken together, they do not include all the taxa found in Udabno site. All these issues make compilation of one comprehensive faunal list of the Udabno site more difficult.
Table 5. Udabno faunal lists according to Tsikarishvili (1987); reptiles from Chkhikvadze and Amiranashvili (1996).
|
Udabno 1 |
Udabno 2 |
Udabno 3 |
Udabno 4 |
|
Reptilia |
poor fossil material, faunal list has never been published |
||
| |
Testudo cf. eldarica Mauremys cf. sarmatica Megalochelys sp. |
Testudo eldarica Trionyx sp. Mauremys sarmatica |
|
|
Mammalia |
|||
|
Primates |
|||
| |
|
Dryopithecus garedziensis |
|
|
Carnivora |
|||
| |
Percrocuta gigantea |
Adcrocuta eximia |
|
|
Proboscidea |
|||
| |
Deinotherium giganteum |
Deinotherium sp. (cf. giganteum) |
|
|
Perissodactyla |
|||
|
Chalicotheriidae indet. Ceratotherium cf. schleiermacheri |
Hipparion garedzicum Diceros sp. Aceratherium sp.
|
Hipparion sp. Aceratherium cf. incisivum Aceratherium sp. Chilotherium (Subchilotherium) aff. eldaricum |
|
|
Artiodactyla |
|||
|
|
Microstonyx aff. major Palaeotragus roueni Gazella sp. |
Gazella schlosseri
|
|
Table 6. Udabno faunal lists according to Vangengeim et al. (1989) and Gabunia et al. (2001).
|
Vangengeim et al. (1989) |
Gabunia et al. (2001). |
||
|
Udabno 1 |
Natlismtsemeli 1 |
Udabno 2 |
Natlismtsemeli 1 |
|
Primates |
|||
|
Dryopithecus garedziensis |
|
|
|
|
Carnivora |
|||
|
Plesiogulo cf. brachygnathus Adcrocuta cf. eximia Miohyaenotherium bessarabicum |
Percrocuta gigantea
|
Adcrocuta eximia Hyaenotherium magnum |
|
|
Proboscidea |
|||
|
Deinotherium giganteum Tetralophodon cf. longirostris |
|
Deinotherium giganteum |
Chilotherium (Subchilotherium) cf. eldaricum
|
|
Perissodactyla |
|||
|
Hipparion garedzicum Chalicotherium sp. Aceratherium cf. incisuvum |
Hipparion cf. eldaricum
|
Hipparion cf. garedzicum |
Hipparion cf. garedzicum Ictitherium hipparionum cf. garedzicum
|
|
Artiodactyla |
|||
|
Microstonyx major erymanthius Cervidae gen. indet. Giraffidae cf. Palaeotragus sp. Tragocerus sp. Gazella schlosseri |
Microstonix cf. antiquus Diceros sp. Tragocerus sp. Gazella cf. schlosseri
|
Udabnocerus georgicus Palaeotragus sp. |
Tragocerus sp. |
The 30 m thick petroliferous sandstone, containing lower fossil-bearing horizon of the Eldari fauna (Eldari 1 of Vangengeim et al. 1989; Table 1), represents a marker horizon of the southern zone of the Middle Kura Foreland and is easily traceable along 70 km EW from Ellyar-Oyugi to the ravine Dibzis. According to the established point of view the Bessarabian–Khersonian boundary passes underneath this horizon (Buleishvili 1960). The mentioned sandstone falls within the normally magnetised long interval in the Eldari section (Vangengeim et al. 1989; Vangengeim and Tesakov 2008), which corresponds to C4An (9.1–8.8 Ma; Sen 1997) and dates the Eldari 1 fauna. Dating of the Udabno site is more problematic. The above mentioned diachronous appearance of the continental deposits (Eldari Formation) along the Kura Bay theoretically does not exclude contemporaneity of Udabno 1 and Eldari 1 faunas, or even an older age of the former since Bessarabian–Khersonian transition occurs below the mentioned 30 m thick sandstone (Buleishvili 1960). Paleomagnetic study (Vangengeim and Tesakov 2008: fig. 3) indicates that lowermost Khersonian corresponds to C4Ar-1n in the Middle Kura Basin, and thus dates it ca. 9.4 Ma. A major difficulty in the chronostratigraphic interpretations is the highly probable presence of multiple hiatuses in the continental deposits. This aspect was underlined by Gabunia (1959) in his monograph about hipparions and is also highlighted in the paleomagnetic study of Vangengeim et al. (1989), which places Udabno 1 fauna somewhat above Eldari 1 in the section, right at the limit of Chrons 9 and 8 (interpreted as C4An and C4r by Sen 1997). Specifically, their data (uninterrupted normally magnetised long interval in the lower part of the section embracing Bessarabian and Khersonian deposits) indicate that the latest Bessarabian and the lower Khersonian intervals are missing from the Udabno section. The issue of stratigraphic hiatuses is rather acute for the upper, reversely magnetised part of the section as well, and complicates interpretation of paleomagnetic results because the dating of the Meotian lower boundary is problematic in the Eastern Paratethys; according to Vasiliev et al. (2011), the Khersonian–Meotian transition took place at 8.6 or 8.2 Ma (slightly above C4r.1r or C4r.2r-1n), while Radionova et al. (2012) suggest a younger date, ca. 7.6 Ma. Besides, it is challenging to delimit Meotian and Khersonian stages in the continental deposits of Middle Kura Basin; based on biostratigraphy of Udabno vertebrate fauna (Gabunia 1959; Vangengeim et al. 1989), some authors consider that the upper interval of the variegated clay in Udabno might already be Meotian, but others do not adhere to this idea and draw the Khersonian–Meotian boundary under the 5–6 m thick sandstone overlying the variegated clay deposits (Buleishvili 1960). Lithological and paleomagnetic correlations of series of sections along the Kura paleobay need to be done in combination with absolute dating of the existing volcanic ash layers in the Eldari and Shiraki Formations (e.g., in the Eldari Formation at the Udabno section; volcanic ash lense is also mentioned for the Katar section; Buleishvili 1960) in order to obtain to a clear chronological framework for this terrestrial fossil record.
Unfortunately, a detailed biostratigraphic comparison of the Eldari and Udabno faunas is impossible because of insufficient stratigraphic control at these sites, where fossils come from several fossil-bearing horizons and several localities. The absence of typical Vallesian rhinos (e.g., Lartetotherium, Brachypotherium), muntjacs (Euprox, Dicrocerus), and the low diversity of suids (represented only by Microstonyx) points to either a late Vallesian or early Turolian age for these faunas. It is noteworthy that there is a striking difference with the neighbouring early Turolian fauna from Maragheh (Iran, only some 400 km away from these sites). For example, the Lower Maragheh (MN11, 8.9–8.2 Ma; Bernor et al. 1996) bovids are more diverse than the Eldari or Udabno faunas; though rare in the former (represented by Protragelaphus skouzesi and Prostrepsiceros sp.; Kostopoulos and Bernor 2011), spiral-horned antelopes are totally absent in the South Caucasian sites. The bovid community from the Udabno and Eldari sites is represented mainly by boselaphines (Tragocerus leskevitszi, Tragocerus sp., and Mirabilocerus azerbajdjanicus), primitive gazelle species (Gazella leilli in Eldari and Gazella schlosseri in Udabno), Udabnocerus georgicus, a bovid of uncertain phylogenetic affinities (from the locality Adjia found in the Shiraki Formation, Meotian–Pontian), and Eotragus (?) martinianus. The latter could be a relict; however, to substantiale this assumption more research is needed. Giraffids at these two sites are not diverse (Palaeotragus [Achtiaria] borissiaki and Palaeotragus sp. in Eldari; Palaeotragus roueni and Palaeotragus sp. in Udabno). Carnivores are represented by taxa appearing in the Turolian (Adcrocuta eximia, Miohyaenotherium bessarabicum, and Simocyon sp.), by taxa that appear earlier in the Vallesian but persist into Turolian times—Percrocuta gigantea and Plesiogulo cf. brachygnathus, or by taxa not mentioned elsewhere (Hyaenotherium magnum). The primate Udabnopithecus garedziensis, a fragmentary and poorly known late Miocene ape from the Udabno site, is synonymized with Dryopithecus by a majority of researchers (Gabunia et al. 2001; references therein); it is the latest and the easternmost dryopithecine, which makes this fossil one of the key specimens for the evolutionary scenarios of Eurasian primate record (Begun 2001, 2010 and references therein).
In total, this faunal material suggests that the Eldari and Udabno faunas from the lower fossil-bearing horizon most probably belong to the latest late Vallesian (MN10).
Chachuna site: Late Miocene, marine Khersonian and Eldari Formation, early Turolian, MN11; located on the right bank of the Iori river on the Akhtakhta-tapa (or Akhtatapa) and Djighati (or Kanly) ranges, Dedoplistskaro region, Kakheti, Georgia. The site was discovered by Viacheslav M. Chkhikvadze in 2000 (Chkhikvadze and Petrov 2000). The fauna from Chachuna comes from the marine and continental deposits of the Khersonina stage. Remarkable concentrations of terrestrial vertebrate remains were encountered in three points so far (Bukhsianidze et al. 2018). The following vertebrate taxa were found: Pisces indet., Trionychdiae indet., Testudo sp., Falconiformes indet., Castoridae indet., Choerolophodon sp., Ictitheriidae indet., Hipparion sp., Aceratheriinae indet., Microstonyx major Gervais, 1851, Procapreolus sp., Lucentia sp., Muntiacinae gen., Giraffidae indet., Boselaphini indet., and Antilopini indet. (Bukhsianidze et al. 2018).
Cervids from one of the localities of the site, Chachuna-2, are the most dominant and diverse group (Procapreolus sp., Lucentia sp., Muntiacinae gen.), while bovid remains are mostly absent from the locality—an exceptional case for the late Miocene in the South Caucasus. The Chachuna-2 cervid collection represents more than half of the cervid specimens found in the entire late Miocene of Georgia. This assemblage most likely reflects a very brief time interval and a local biotope. The faunal list in combination with the marine biostratigraphy (the presence of the Khersonian malacofauna on top of the fossil-bearing strata) may support its inclusion in the early Turolian (MN11).
Iagluja (Rustavi) site: Late Miocene, early Turolian, MN11; located on the right bank of the river Tavaddere—the right tributary of the Kura river, in the city of Rustavi, on the Iagluja Mountain, Shida Kartli, Georgia; discovered by Karlo Matskhonashvili and David (Devi) Chkheidze in 1962. It represents one of the richest and largest late Miocene sites in South Caucasus. Results of the expeditions of the Institute of Paleobiology, Tbilisi, were published by Meladze (1985). The faunal list of the Iagluja (Rustavi) site counts the following taxa: Testudo sp.; Daboia sp.; Rustaviornis georgicus Burchak-Abramouich and Meladze, 1972; Melinae indet.; Promephitis ex-gr. maeotica Alexeev, 1916; Adcrocuta eximia (Roth and Wagner, 1854); Ictitherium sp.; Machairodontinae indet.; Choerolophodon pentelici (Gaudry and Lartet, 1856); Chalicotheriinae indet.; Dicerorhinus sp.; Aceratherium sp.; Hipparion cf. eldaricum Gabunia, 1959; Microstonyx sp.; Procapreolus sp.; Palaeotragus sp.; Tragicerus sp.; Gazella cf. capricornis (Wagner, 1848); Oioceros atropatenes Rodler and Weithofer, 1890; and Paraoioceros improvisus Meladze, 1985 (= Paraoioceros wagneri [Andrée, 1926]) (Burchak-Abramovich and Meladze 1972; Meladze 1985; Kostopoulos 2014).
The composition of the Iagluja (or Rustavi) fauna is remarkably different from the previously discussed sites due to the diversity of bovids, with Oioceros atropatenes being the dominant taxon. This gives a somewhat younger appearance to this fauna in comparison with Udabno and Eldari. The biostratigraphic distribution of the faunal elements, especially bovids, suggests a late early to middle Turolian age (MN11–MN12). However, because of the presence of a Hipparion species (H. cf. eladricum) that is similar to one from the marine Khersonian of Eldari (H. eldaricum), Iagluja is usually correlated with the lower fossil-bearing horizon of the Eldari site, which is considered to be Vallesian (MN10; Gabunia et al. 1966; Meladze 1985).
In general, local biostratigraphic importance and a low diversity of Hipparion in the late Miocene of the South Caucasian faunas (the genus is represented only by two species H. eldaricum and H. garedzicum) suggest that this group deserves special investigation to integrate this record into a larger picture. It is possible that the two late Miocene species, H. eldaricum and H. garedzicum (Vallesian and Turolian respectively, according to Gabunia 1959), are coeval and their biostratigraphic importance is somewhat exaggerated. In fact, there is evidence from the Udabno site that these taxa were present simultaneously at two localities that are correlated with one another, specifically H. garedzicum in Udabno 1 and H. cf eldaricum in Natlismtsemeli 1.
Sites from the Shiraki Formation.—The continental Shiraki Formation between Khersonian and Akchagylian deposits represents Meotian–Pontian deposits. This biostratigraphic age is based on correlations with the Dusheti Formation of the Kartly Foreland Basin in eastern Georgia, which is dated to the Meotian–Pontian using the biostratigraphy of the large mammal sites, Arkneti and Bazaleti found there (Meotian and Pontian, respectively; Meladze 1967). In the geological literature there is disagreement regarding the magnitude of erosion in the upper part of the Shiraki Formation in the Middle Kura Basin (Khuchua and Shatirishvili 1937; Kudriavtsev 1938; Buleishvili 1960; Chubinishvili 1982).
Four land mammal sites are known from the Shiraki Formation: Dzedzvtakhevi, Vashlovani, Tetri Udabnos Seri, and Dzhaparidze. Brief publications exist only for the Dzhaparidze and Dzedzvtakhevi faunas (Orlov 1941; Vekua and Trubikhin 1988; Vanishvili et al. 2007).
Dzedzvtakhevi site: Late Miocene, Shiraki Formation, middle Turolian, MN12; located on the southern slope of Pirukugma Mountain on the right bank of the Iori river, Sighanghi region, Kakheti, Georgia. The site was discovered by Trubikhin in the 1980s. Faunal list of the Dzedzvtakhevi site: Ergilemys natadzei Chkhikvadze, 1989; Adcrocuta eximia (Roth and Wagner, 1854); Simocyon primigenius Roth and Wagner, 1854; Felis attica Wagner, 1857; Gomphotheriidae gen.; Hipaprion ex. gr. elegans Gromova, 1952; Dicerorhinus sp.; Microstonyx major erymanthius (Roth and Wagner, 1854); Cervidae indet.; Karsimatheirum aff. bazaleticum Meladze, 1961 (the list is based on Vekua and Trubikhin 1988; Vanishvili et al. 2007).
According to palaeomagnetic research (Vekua and Trubikhin 1988; Gurarij et al. 1995), the fossil-bearing horizon at Dzedzvtakhevi is immediately above the reversely magnetized episode Chron 5 (corresponding to the subchron C3An, 6.25–6.44 Ma). Yet, the faunal assemblage, especially the presence of Nisidorcas planicornis (Pilgrim, 1939) (identification of MB), suggests a somewhat older age of the fauna (middle Turolian, MN12, ca. 7.5–7 Ma, possibly Chron C3B). This age supports the idea that the upper part of the Shiraki Formation in the area of the Pirukugma Mountain was removed by the pre-Akchagylian denudation (Khuchua and Shatirishvili 1937) and does not support the alternative point of view (Kudriavtsev 1938; Buleishvili 1960; Chubinishvili 1982; and others) that both the lower and upper parts of the formation are represented in this section.
Tetri Udabnos Seri site: Late Miocene, Shiraki Formation, early–middle Turolian, MN11–MN12, Sagaredjo region, Kakheti, Georgia; no publications. The fauna comes from the sandstone beds of the lower part of the Shiraki Formation. It is rather diverse, though identifications are preliminary, and includes 10 taxa: Testudines indet.; Aves indet.; Machairodontinae indet.; Tetralophodon longirostris (identification by Abesalom Vekua); Rhinocerotidae indet.; Hipparion sp.; Microstonyx cf. major Gervais, 1851; Procapreolus indet.; Giraffidae cf. Palaeotragus; and Gazella cf. capricornis (Wagner, 1848). Current identifications show that this faunal assemblage corresponds to the Turolian in general, but do not allow for further precision. The horizon that yielded the fauna belongs to the lower part of the Shiraki Formation (Buleishvili 1960). This stratigraphic position of the fossil-bearing horizon suggests an early–middle Turolian age (MN11–MN12).
Vashlovani site: Late Miocene, Shiraki Formation, MN12, Vashlovani Reserve, Dedoplistskaro region, Kakheti, Georgia; no publications. This site serves as an attraction point in the Vashlovani Reserve with an erroneous indication of a fossil proboscidean taxon (Mammuthus meridionalis). The collection includes only two taxa, Gomphotheriidae indet. and Microstonyx major Gervais, 1851. The latter taxon delimits the upper age on the fauna to ~7 Ma and, together with geological correlations, suggests the Turolian stage (MN12).
Dzhaparidze site: Late Miocene, Dedoplistskaro–Krasnokolodskaja Formation, upper Meotian?, Pontian? MN12–MN13?; located in the village Samreklo (former Dzhaparidze), Dedoplistskaro district, Kakheti, Georgia; discovered by Nikolaj A. Kudriavtsev in 1935 (Orlov 1941). Faunal list of the Dzhaparidze site according to Orlov (1941) and Gabunia (1959) counts the following forms: Proboscidea gen., Hipparion sp., Rhinocerotidae gen., Suidae indet., Cervidae indet., Giraffidae indet., and Gazella sp.
The Dzhaparidze fauna is considered to be Meotian in the paleontological literature (Meladze 1985). This idea is based on the similarity of the Dzhaparidze Hipparion with H. garedzicum from Udabno (Hipparion sp. in Gabunia 1959). The age of the Dedoplistskaro Formation is debatable, yet the majority of researchers date it to the Pontian (Buleishvili 1960; Dzigrashvili and Sepashvili 1978; Chubinishvili 1982).
Unfortunately, the Dzhaparidze fauna, which offers so far the only biostratigraphic evidence for this geological formation, cannot provide a reliable biostratigraphic date, because none of the taxa mentioned in the faunal list (seven in total) were identified to species (Orlov 1941). The scarce remains of Hipparion (eight highly-worn, isolated teeth) do not invalidate a younger age of the Dzhaparidze site.
Based on this evidence, the late Miocene of the Iori valley demonstrates that the major part of the fossil record fits within the Khersonian–Meotian time-frame, ca 9.4–7 Ma (late Vallesian–Turolian, MN10–MN12). So far, there is no solid evidence for Pontian terrestrial mammal faunas found in the Shiraki Formation, though Dzhaparidze could be one. The earliest faunas at Eldari (Eldari 1) and Udabno (Udabno 1) represent the latest Vallesian (MN10) assemblages. Over time, the faunal composition acquired the typical Turolian aspect.
The Khersonian fossil record of the Iori plateau (Udabno, Chachuna, Eldari sites) captures the very last moment of the long-term regression of the Eastern Paratethys. Sea waters have reached the Middle Kura Basin only ca. 5–4.5 million years later, after the end of the Khersonian. The occurrence of a dryopithecine (Udabnopithecus garedziensis) in the Khersonian of Udabno suggests the presence of humid subtropical coastal forests along the regressing Kura Bay. The Katar flora from the lower part of the Khersonian deposits of the Iori plateau (Fataliev 1964) indeed reflects the presence of gallery and coastal forests with subtropical elements. In western and central Europe, in the Iberian bioprovince and in the Pannonian Basin, the demise of dryopithecines’ habitat, subtropical forests with broadleaved evergreens, occurred earlier (ca. 9.6 Ma) and coincided with the mid-Vallesian crisis (Agustí et al. 2003; Casanovas-Vilar et al. 2005, 2011; Utescher et al. 2017). In this regard, the South Caucasus, and in particular the Kura Basin, can be considered as a refugium for dryopithecines, since the fossil evidence of vegetation cover points to longer survival of humid subtropical elements, althoug better temporal resolution of this fossil record and more paleobotanical evidence are obviously needed. It is difficult to imagine this territory as a refugium of humid biome for the later interval of the Miocene epoch because the west-east moisture gradient (decreasing) was already apparent in the palynological samples from the end of the Bessarabian (Shatilova and Kokolashvili 2011, 2012, 2013; Shatilova et al. 2012), even during the times when the Kura paleo-bay, an important moisture source, was present in the interior of the area. The paleogeography depicts the Middle Kura Basin as part of the Greco-Iranian province during the Meotian–Pontian and most probably the evolution of the land mammal fauna was in line with the faunal evolution of this bio-province driven by the aridisation (Eronen et al. 2009), while neighbouring Northern Caucasus shows warm and forested conditions during Khersonian and lower Meotian (Tesakov et al. 2017).
Plio-Pleistocene
Sites from the Plio-Pleistocene continental formation.—The land mammal sites of Plio-Pleistocene age mainly come from the Akchagylian, Apsheronian, or the strata from transitional Akchagylian–Apsheronian depositional environments.
Zemo Melaani site: Late Pliocene–Early Pleistocene, Alazani Series, early–middle Villafranchian, MN16–MN17; located on the right bank of the Lakbe river, in the village Zemo Melaani, Gurjaani region, Kakheti, Georgia. The following taxa are mentioned in the literature from this site: Nyctereutes megamastoides (Pomel, 1842); Chasmaporthetes lunensis Del Campana, 1914; Dinofelis cf. abeli Zdanski, 1924 (= D. aff. cristata [Falconer, 1836]); Stephanorhinus megarhinus de Christol, 1835; Propotamocheorus sp.; Leptobos sp. (list is based on Gabunia and Vekua 1998; Vekua 1991; Werdelin 2001).
The fossil-bearing lens found in the Alazani Series near Zemo Melaani village (in the northern zone of the Middle Kura Basin) represents the only site found in this continental formation, though undocumented information about finds of terrestrial fossils in this area exists (e.g., Ali-Zade et al. 1972; Abesalom Vekua personal communication 2006).
Among the exceptionally well-preserved fossils in the Zemo Melaani collection, only the Dinofelis cf. abeli skull was studied in detail (Gabunia and Vekua 1998; Werdelin 2001). The presence of Nyctereutes megamastoides implies a late Pliocene–Early Pleistocene age (MN16–MN17) for the site, while other faunal elements identified at the species level (Chasmaporthetes lunensis and Stephanorhinus megarhinus) have longer ranges and do not exclude an earlier age (Pliocene).
Sites from the Akchagylian and Apsheronian deposits.—Marine biostratigraphy provides the main correlation framework for this region during the late Pliocene–Quaternary. Akchagylian, Apsheronian, and Bakunian, the three successive regiostages, embrace the time span from the late Pliocene to the Late Pleistocene. The onset of the Akchagylian transgression is dated to the Gilbert-Gauss reversal in western Turkmenistan, a bit younger in Azerbaijan (Trubikhin 1977; Pevzner 1982; Nikiforova et al. 1982). Recently, the Lokbatan section (South Caspian Basin, near the Caspian Sea coast) was dated to ca. 3.2 Ma (Van Baak et al. 2013). The limit between the Akchagylian and Apsheronian passes at the base of the normal polarity interval corresponding to Gilsa excursion (1.57 Ma) or Gilsa excursion and Olduvai subchron (1.95–1.78 Ma; Trubikhin 1977), or within this interval (Nikiforova et al. 1982; Isaeva and Mamedov 1989), and was recently dated at ca. 2.0 Ma (Van Baak et al. 2013). The onset of the Bakunian transgression at the Brunhes-Matuyama reversal was confirmed by Van Baak et al. (2013), who dated it to 0.85–0.89 Ma.
However, while there is a rather good agreement in dating the limits of the regiostages, the ideas about their subdivisions differ. According to the traditional scheme of Kolesnikov (1940), Akchagylian is divided into lower, middle, and upper sub-stages. This scheme assumes the existence of one transgression-regression cycle during Akchagylian, which was not confirmed by subsequent studies (Ali-Zade 1961; Popov 1969; Trubikhin 1977; Lebedeva 1978; Nevesskaya and Trubikhin 1984; Danukalova 1990; and others). In the Middle Kura Basin, as well as generally in the Caspian Sea Basin, the presence of two major transgression-regression cycles during Akchagylian times is evident (Alizade 1954; Ali-Zade 1961; Trubikhin 1977; Lebedeva 1978). The second transgression cycle starts below the Gauss-Matuyama reversal (in Kushkuna section, western Azerbaijan; Lebedeva 1978) or coincides with the latter (western Turkmenistan; Trubikhin 1977). Researchers differently correlate these two cycles with the sub-stages of the Kolesnikov scheme. There are several schemes of the subdivision of Apsheronian deposits as well (Fig. 4). The sub-stages used in this paper are according to Kolesnikov (1940, 1950) due to largely accepted practice.
Discoveries of fossil vertebrate remains of this geological age are very frequent in the Middle Kura Basin due to the wide distribution of shallow, coastal marine facies and a well developed hydrological network, which are the favouring factors for the accumulation of terrestrial fossils. Impressively long sections of Akchagylian, Apsheronian, and Bakunian deposits with terrestrial mammal remains from a succession of different levels are known from Azerbaijan, while in Georgia the fossil record in terms of number of terrestrial mammal sites is poorer and more dispersed. Most of the sites in Azerbaijan were discovered by Natalija A. Lebedeva, who carried out large-scale correlation works of marine and continental deposits in the Ponto-Caspian region during 1966–1973. For correlation purposes, greater attention was focused on the Quaternary index taxa, such as Mammuthus, and most of the diverse fossils she discovered remain largely unstudied. Consequently, the late Pliocene–Pleistocene biostratigraphy of terrestrial faunas mainly follows the the evolutionary stages of mammuthoid elephants. Although terrestrial faunas from the Georgian territory, especially Kvabebi, are better studied, controversies remain.
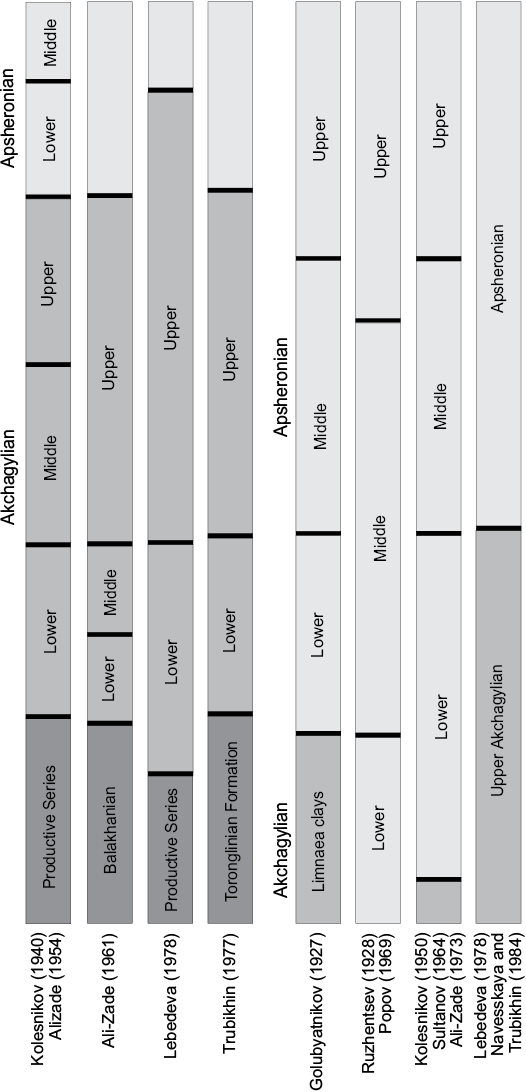
Fig. 4. Schemes of subdivision of Akchagylian and Apsheronian regiostages adapted from Filippova 1997.
Kushkuna site: Late Pliocene–Early Pleistocene; lower, middle, upper Akchagylian and undivided Akchagylian–Apsheronian, early–middle Villafranchian, MN16b–MN17; located on the south-eastern foothills of the mountain Kushkuna, in the Adjidere ravine, Aghstafa region, Azerbaijan, and discovered by Yurij P. Bazhenov in 1961 (Lebedeva 1972). The site contains a succession of land mammal fossil-bearing horizons corresponding to early, middle, and late Villafranchian (European large mammal ages).
Unfortunately, taxonomic identifications from previous authors/works are not sufficient to compile a formal faunal list. Layer by layer succession of terrestrial vertebrates from these horizons is as follows (layers according to Lebedeva in Ali-Zade et al. 1972):
First horizon, lower Akchagylian (layer 14): sporadic, unidentifiable remains of large mammals and tusks of proboscideans.
Second horizon (layers 16, 17, 19), middle Akchagylian: large amounts of vertebrate fossils. Tooth fragments of mastodonts, antler fragments of cervids, gazella tooth and jaw fragments, tusk fragments of Proboscidea, and turtle shell fragments are listed from the layers 16 and 17. Among the identified large mammals, the following taxa are mentioned in Burchak-Abramovich et al. (1980) and Lebedeva (1972, 1974, 1978): Anancus cf. arvernensis, Cervus (Rusa) sp., Euctenoceros (?) sp., and Protobison kushkunensis Burchak-Abramovich, Gajiev, and Vekua, 1980. Mimomys polonicus Kowalski, 1960, and Borsodia ex gr. steklovi–novoasovica (Topachevski and Skorik, 1977) (Tesakov 2004) are coming from the layer 17, and from the layer 19 Mammuthus (Archidiskodon) gromovi Alekseeva and Garutt, 1965 was found.
Third horizon (layers 27–28), upper Akchagylian: large amounts of fragments of proboscidean bones and tusks, camilidae limb bones, cervid antlers, struthio egg shells, and turtle shells. Among them the following taxa were identified by Alekseeva: Leptobos sp., Hipparion (?) sp. (femur), and Gazella sp.
Fourth horizon (layer 33), undefined Akchagylian–Apsheronian. Equinae indet., Cervus ramosus, Cervus rusa sp., Adjiderebos cantabilis Dubrovo and Burchak-Abramovich, 1986 are listed along with large amounts of fragments of cervid antlers, struthio egg-shells, proboscidean limb-bones and tusks, and turtle shells.
The fauna from the middle Akchagylian layers (the second fossil-bearing horizon), which is found below the Gaus-Matuyama reversal, seems to be rather diverse and is correlated with early Villanyan, early Villafranchian (MN16b), based on the evolutionary stage of small mammals found here (Mimomys polonicus and Borsodia ex gr. steklovi–novoasovica; Tesakov 2004). Above the Gauss-Matuyama reversal, teeth of Mammuthus (Archidiskodon) gromovi are the most remarkable finds as this taxon is an index species of the Khaprovian fauna in Eastern Europe and corresponds to the middle Villafranchian (MN17). The faunas from the upper Akchagylian and the undefined/transitional Akchagylian–Apsheronian sediments are equally diverse and numerous. However, the taxonomic studies are not complete enough for a detailed biostratigraphic discussion. This sequence of faunas does, nevertheless, document the first appearance of a primitive bisontine, Protobison kushkunensis (Burchak-Abramovich et al. 1980) from the middle Akchagylian deposits (early Villafranchian, MN16b), which is the earliest date for a bisontine cattle and it is rather close in time to the Probison dehmi from Northern India (Sahni and Khan 1968; near Chandigarh, Punjab, and Haryana, India; Upper Sivaliks, Tatrot, ca. 2.6 Ma according to Nanda 2002). The Kushkuna section also reveals the appearance of camelids in the region at the regressive phase of the Akchagylian transgression and another bovine of uncertain phylogenetic position (Adjiderebos cantabilis) from the undivided/transitional Akchagyl–Apsheronian freshwater deposits (Dubrovo and Burchak-Abramovich 1986). These are important events for the correlation of faunas at the local scale of the Caspian Basin.
Kvabebi site: Late Pliocene, middle Akchagylian, early Villafranchian MN16b; located on the eastern foothills of the mountain Kvabebi on the right bank of the Iori river, village Iormughanlo, Sighnaghi region, Kakheti, Georgia; discovered in 1962 by Heinrich S. Avakov. For a faunal list see Table 7.
Table 7. List of Kvabebi fauna. The list is based on Vekua 1972; Burchak-Abramovich and Vekua 1981; Sotnikova 1989; Hemmer et al. 2004; Agustí et al. 2009; Pickford 2013; Bendukidze and Vekua 2012; Rook et al. 2017.
|
Reptilia |
|
Testudo cernovi transcaucasica (Chkhikvadze, 1979) |
|
Aves |
|
Struthio transcaucasicus Burchak-Abramovich and Vekua, 1971 Ioriotis gabuniae (Burchak-Abramovich and Vekua, 1981) |
|
Mamalia |
|
Rodentia Hystrix cf. primigenia (Wagner, 1848) |
|
Carnivora Nyctereutes megamastoides (Pomel, 1842) Vulpes cf. alopecoides Major, 1875 Eucyon sp. Ursus minimus Devèze de Chabriol and Bouillet, 1827 Perunium kvabebicus Bendukidze and Vekua, 2012 Chasmaporthetes lunensis Del Campana, 1914 Homotherium davitashvili (Weithofer, 1889) Dinofelis sp. Lynx issiodorensis (Croizet and Jobert, 1828) Puma pardoides (Owen, 1846) |
|
Proboscidea Anancus arvernensis (Croizet and Jobert, 1828) |
|
Hyracoidea Kvabebihyrax kachethicus Gabunia and Vekua, 1966 |
|
Perissodactyla Hipparion rocinantis Villalta, 1948 Stephanorhinus megarhinus (De Christol, 1834) |
|
Artiodactyla Propotamochoerus provincialis (Gervais 1859) (= Dasychoerus sp. according to Pickford 2013) Procapreolus sp. Eucladoceros sp. ?Pseudalces sp. Ioribos aceros Vekua, 1972 Eosyncerus ivericus Vekua, 1972 Parastrepsiceors sokolovi Vekua, 1968 Oryx (Aegoryx) sp. Protoryx heinrichi Vekua, 1972 Gazella postmitilinii Vekua, 1972 |
The Kvabebi site has produced a very rich faunal assemblage, which is rather well studied taxonomically; the site is well dated using absolute age, magneto- and bio-stratigraphy. However, some contradictions among the available results are evident. For example, palaeomagnetic research of this section was conducted by Valerij M. Trubikhin (unpublished material, cited by Vekua 1991) and later by Oriol Oms (in Agustí et al. 2009), but their results differ: according to Trubikhin, the Kvabebi fossil-bearing layers reveal normal polarity, while according to Agustí et al. (2009) fossils are found in two horizons (1 m apart) and the lower (main fossil-bearing horizon) reveals reversed whereas the upper shows normal polarity, capped by a large zone or reversed polarity. Agustí et al. (2009) interpret the paleomagnetic data in combination with the biostratigraphy of vertebrate fauna as Kaena (C2An.1r; 3.032–3.116 Ma). However, this interpretation is not in accordance with the absolute fission-track dates of the ash layers below and above the fossil-bearing horizon (2.53±0.20 Ma and 2.18±0.18 Ma, respectively; Chumakov et al. 1992). The Agustí et al. (2009) interpretation also conflicts with the find of the vertebrate fauna within the middle Akchagylian layers according to Kolesnikov’s (1940) scheme, corresponding to the second transgression-regression cycle of Lebedeva (1978), and with the possible diachronous, delayed appearance of the Akchagylian transgression in the interior of the Kura Foreland Basin (min. of 0.5 Ma later, as argued by Forte et al. 2015); Kvabebi is actually located ~350 km west of the Lokbatan section, where the Akchagylian transgression was dated at ca. 3.2 Ma (Van Baak et al. 2013). However, it must be noted, that the section of Kushkuna does not support a considerable diachrony of the middle Akchagylian transgression either; onset of this transgression cycle at this section is dated to ca. 2.9 Ma (Tesakov 2004).
Another problematic issue at Kvabebi is the biogeographic affinity of some faunal elements, especially bovids, as this group comprises numerous Afrotropical (Ethiopian) taxa (Sincerini, Tragelaphini, and Hippotragini). They were interpreted as relicts of Mio-Pliocene Hipparion faunas and Vekua (1972) suggested that the formation of the main core of the modern fauna of the South African savanna took place during the existence of the Kvabebi-Russilion fauna (Kvabebi was correlated with Russilion by Vekua 1972) and not during the Pikermian fauna as it was previously considered. Alternatively, Agustí et al. (2009) interpret the presence of Ethiopian taxa as the last remnants of the late Miocene Subparatethyan, or Greek-Iranian province, rather than the result of a middle Pliocene migration from Africa. Unusual African affinities of the fauna together with the presence of some Ruscinian elements (e.g., Machairodus davitashvili, later moved to Homotherium davitashvili by Sotnikova 1989) raised a question about the recognition of a special Transcaucasian zone of terrestrial faunas during the middle Akchagylian (Lebedeva 1974, 1978). This author’s preliminary observations on the Kvabebi ruminants suggest many taxonomic changes: among cervids, Arvernoceros sp. (not yet mentioned in the faunal list) and Croizetoceros ramosus (= Eucladoceros sp. in Vekua 1972) are present in addition to Procapreolus sp. and Pseudalces sp., which is more likely a Palaeotragus sp.; among bovids, Eosyncerus ivericus belongs to Caprinae, Parastrepsiceors sokolovi is synonymous with Gazellospira torticornis, Protoryx heinrichi with Gazella borbonica; etc. These faunal elements are typical Early–Middle Villafranchain Eurasian taxa.
The absence of Mammuthus and Equus in the Kvabebi fauna makes a correlation to the second horizon of the Kushkuna section (middle Akchagylian, early Villafranchian, MN16b) plausible. Here, it is relevant to mention that Hipparion (H. apscheronicum Gabunia, 1959 from Shikhov Cape, SW of Baku, Azerbaijan) survived in the South Caucasus after the appearance of Equus in the region (Gabunia 1959). The first record of the latter comes from the Kotsakhuri fauna (late Akchagylian, MN17), where this taxon is recorded together with camelids. In addition, according to Eisenmann and Sondaar (1989), the Kvabebi Hipparion is more derived than the form from the type locality of H. rocinantis in Villaroya, Spain. The presence of some middle Villafranchian taxa, such as Vulpes cf. alopecoides (Rook et al. 2017), as well as Gazellospira torticornis, the dominant antelope in the Kvabebi fauna, might support a younger age (MN17).
Taxonomic revision (especially of artiodactyls) and reconciliation of paleomagnetic data with the existing absolute dates (or new dating) is needed to resolve these controversies.
Kotsakhuri site: Early Pleistocene, undivided Akchagylian–Apsheronian, middle Villafranchian, MN17. The site is located on the southern foothills of the Kotsakhuri range on the left bank of the Iori river, Dedoplistskaro region, Kakheti, Georgia. It was discovered by Valerij M. Trubikhin (Abesalom Vekua, personal communication 2005). The faunal list of the Kotsakhuri site according to Vekua (1991) and Vekua and Lordkipanidze (1998) includes: Emys orbicularis (Linnaeus, 1758); Testudo sp.; Struthio cf. transcaucasicus Burchak-Abramovich and Vekua, 1971; Mammuthus cf. meridionalis (Nesti, 1825); Equus stenonis Cocchi, 1867; Stepahnorhinus cf. etruscus (Falconer, 1868); Paracamelus sp.; Croizetoceros aff. ramosus; Leptobos sp.; and Protoryx sp.
The Kotsakhuri site is mentioned in several publications (Burchak-Abramovich and Vekua 1980; Vekua 1991; Vekua and Lordkipanidze 1998; Vekua et al. 2010), but mentions are limited to general discussions on the biostratigraphy and paleoenvironment, with faunal lists changing from one publication to the other. Unfortunately, the large mammal fossils were never subjected to detailed taxonomic research, nor was the site excavated, although fossil concentration suggests that there is a potential for fruitful excavations there. The mammal fossils are found in freshwater deposits that are correlated to the Domashka Horizon of the scheme of Nikiforova et al. (1982) by Andrej L. Chepaliga (Vekua et al. 2010); Valerij M. Trubikhin (unpublished material mentioned in Vekua 1991 and Vekua et al. 2010) conducted paleomagnetic research indicating that the fossil bearing layer was within a long reversely magnetized interval (interpreted as Matuyama), followed by a normally magnetized strata containing Mammuthus meridionalis taribanensis (interpreted as Jaramillo, see remarks under Taribana site).
The fauna of the Kotsakhuri site can be plausibly correlated with the upper Akchagylian horizon of the Kushkuna section based on the presence of camelids.
Taribana site: Early Pleistocene, undivided Akchagylian–Apsheronian, late Villafranchian, MQ1. The site is located on top of the Kotsakhuri range, Dedoplistskaro region, Kakheti, Georgia. An articulated skeleton of Mammuthus meridionalis taribanensis, the only fossil from this site, was found ~100 m above the Kotsakhuri site. This taxon is considered ancestral to the typical M. meridionalis meridionalis from the Upper Valdarno (1.8 Ma) due to its more primitive morphology (Gabunia and Vekua 1963). In combination with the primitive morphology of the Taribana elephant, the previously mentioned magnetostratigraphic data suggests a context within the Reunion, Olduvai sub-chron or Gilsa excursion. It can be tentatively correlated with late Villafranchian, Psekupsian faunal complex (MQ1).
Palantokan (or Palan-Tyukan, Palantekian) site: Early Pleistocene, undivided Akchagylian–Apsheronian, middle or the earliest late Villafranchian, MNQ17b–MQ1. The site is located in the surroundings of the Palantokan range, Samukh District, Ganja-Qazakh region, Azerbaijan. The site includes several localities, among them Poylu, which is in the north-west foothills of the Palantokan range near Poylu village, and Djeirangel, in the south-west foothills of the Palantokan range, near the Plovdji pass. The first locality is stratigraphically higher than the second. For a faunal list of Palantokan see Table 8.
Table 8. Composite faunal list of Palantokan. Besides the listed taxa numerous fragments of deer antlers, turtle shells, and ostrich egg-shells are also mentioned in literature. The list is based on Lebedeva 1972, 1974, 1978; Sablin 1990; Kuzmina and Sablin 1991; Sotnikova and Sablin 1993.
|
Carnivora Ursus etruscus Cuvier, 1823 Nyctereutes megamastoides (Pomel, 1842) Pachycrocuta perrieri Croizet et Jobert, 1828 Enhydrictis ardea (Bravard, 1828) Meles torali Viret, 1954 Aonyx sp. Megantereon megantereon Croizet and Jobert 1828 ?Homotherium sp. Panthera ex gr. gombaszoegensis Kretzoi, 1938 |
|
Perissodactyla Equus stenonis Cocchi, 1867 Equus sp. |
|
Artiodactyla Sus strozzi Forsyth Major, 1881 Leptobos sp. Gazella (Vetagazella) parasinensis Dmitrieva, 1977 ?Protoryx sp. |
The fossil mammal remains of the Palantokan site document the record of Panthera ex. gr. gombaszoegensis, which is associated with Pliocrocuta perrieri and Nyctereutes megamastoides. This aligns Palantokan with the transitional faunas from the middle to the late or the earliest late Villafranchian, such as: Chilhac, Senèze, and Tegelen (Sotnikova and Sablin 1993). Biostratigraphically, this site immediately precedes the Dmanisi site where Pachycrocuta brevirostris and Canis etruscus are already present together with Panthera gombaszoegensis. However, if the absolute dates are considered, we might be dealing with an interesting biogeographic situation within the South Caucasus. Such indirect evidence as available absolute dates (1.87±0.15 Ma for the volcanic ash layer near the top of the middle Akchagylian in Kushkuna; Chumakov et al. 1992), as well as paleomagnetic data (normally magnetised episode in the upper part of the Kotsakhuri section is attributed to the transitional layers from the Akchagylian to Apsheronian; Lebedeva 1978; Trubikhin in Vekua 1991), suggest a delayed onset of the Apsheronian transgression in the interior of Kura Basin and might indicate a minor time difference or even contemporaneity between the Palantokan and Dmanisi faunas (Dmanisi is dated to 1.85–1.76 Ma, in the upper part of the Olduvai subchron; Ferring et al. 2011). The differences between the faunal compositions could be due to the landscape settings of these two sites: Palantokan is in lowland, lagoon-lake deposits, while Dmanisi is on a high volcanic plateau, in an interior mountainous area. In fact, the Dmanisi carnivores absent in Palantokan, Pachycrocuta brevirostris and Canis etruscus, are already present in the fossil record of Europe and Asia before Dmanisi (Sotnikova and Rook 2010; Palmqvist et al. 2011). The first occurence of the wolf-sized canid of the genus Canis is attested in Diliska (Vekua et al. 2009) in the South Caucasus, on the high volcanic plateau of the Lesser Caucasus, Georgia; fossils in Diliska are found in different horizons of lake sediments sandwiched between two basalts dated at ca. 2.9 and 2 Ma (Tappen et al. 2002). The non-homogenous landscape of the South Caucasus allows such asynchroneity of the faunal composition.
From the remaining Plio-Pleistocene sites in this region (Godjashen, Duzdag, and Yanikend), primarily Mammuthus remains are mentioned and they belong to the following forms (Lebedeva 1972, 1978):
Mammuthus (Archidiskodon) meridioпalis cf. meridionalis (identification of Valerian I. Gromov), a damaged cranium with two well preserved tusks and some other teeth remains. Duzdag 1, lower Apsheronian according to Lebedeva (1972, 1978), correlates with Psekupsian faunal complex, late Villafranchian.
M. (A.) m. tamanensis (identification of Valerian I. Gromov and Vadim E. Garutt)—a cranium with tusks and other teeth. Duzdag 2, middle Apsheronian, Tamanian faunal complex.
M. (A.) m. tamanensis (identification of Valerian I. Gromov), some skull and teeth fragments. Godjashen, marine Apsheronian deposits, presumably middle Apsheronian, Tamanian faunal complex.
A transitional form from M. (A.) meridionalis tamanensis to M trogontherii wusti (identification of Valerian I. Gromov), in situ remains of a skeleton in the village Yanikend, upper Apsheronian.
Kvemo Kedi site: Dedoplistskaro region, Kakheti, Georgia, Middle Pleistocene, NQ2; no publications. A very small fossil collection from the village of Kvemo Kedi comes from a lens in the Alazani river terrace in the northern foothills on the eastern edge of the Gombori Range. A tooth of Mammuthus trogontherii and some limb bones and an antler fragment of a large deer were found here. This is the only find of M. trogontherii in Georgia, and its presence indicates a Middle Pleistocene age.
Among other fossil localities are the Karadja Mountain Range, upper Apsheronian, Qirmizi Samukh village (Lebedeva 1978) and Almaly village, on the left bank of the Alazani river, Late Pleistocene, Gyurgyan Horizon (Bos primigenius; Burchak-Abramovich and Vekua 1980).
Thus, in the Kura foreland there is an almost uninterrupted terrestrial fossil record of the earliest phase of the Quaternary period (Fig. 5). The evidence for Pliocene faunas is controversial. The fossil record of a younger interval from the Early Pleistocene in general reveals the enormous paleontological potential of this area, which is very poorly explored and relatively poorly studied. The greatest value of this Early Pleistocene fossil record is the potential to understand a detailed history of terrestrial life during one of the most intriguing episodes of early Homo dispersal into Eurasia, in the Caucasus—an area where evidence for a rather continuous early Homo occupation is just emerging: Dmanisi attests to the continuous presence of H. erectus in the area for several tens of thousand years (Ferring et al. 2011); there are new archeological discoveries in Armenia (South Caucasus) and Daghestan and Taman peninsula (North Caucasus) that are older than roughly 1.5 Ma (Shchelinsky 2014 and references therein; Shchelinsky et al. 2016).
The correlation of changes in the terrestrial biome with glaciation cycles is a challenge for the South Caucasus due to complicated orography, active tectonics, and different precipitation histories. Complex and confused stratigraphy, especially in the periphery of the Caspian Sea, such as in the Middle Kura Basin, brings further uncertainties. The general correlation framework of the environmental changes with glacial cycles in the comprehensive work done by Filippova (1997) suggests that precipitation and heat supply curves (the scheme proposed by Grichuk 1969) are offset from each other, so that the last ¾ of pluvial and the first ¼ of arid phases correspond to glaciations and the last ¾ of arid and the first ¼ of pluvial phases to interglacials (Filippova 1997: fig. 35). This means that the observed alternation of forest versus open vegetation in the Middle Kura Basin occurred in a delayed, almost inversed mode (forests during most of the glacials and open vegetation during most of the interglacials) compared to western and central Europe (Birks and Tinner 2016). Obviously, this model should have important biostratigraphic consequences for the terrestrial fauna. The discussed fossil record reflects some changes, e.g., animals of open woodland, meadows, swampy biotopes, as well as grasslands were present during transgression in the Kvabebi fauna, while in the Kushkuna (Kushkuna 3) and Kotsakuri sites from the regressive phase of the Akchagylian arid and open habitat dwellers (camelids, Equus, Struthio) are found. Yet, the present state of our knowledge about the terrestrial faunas does not allow for making far reaching conclusions. The reality might not be as straightforward as suggested by this model; neither the hypothesis linking transgressions with pluvial phases and regressions with arid phases, nor the direct association between glacial-interglacial periods with highstands and lowstands of the Caspian Basin finds support in Holocene studies (Leroy et al. 2013). Besides global climatic oscillations, other factors such as changes in the drainage network (for the water level), lake effect, distance from the glacial refugia, altitude, and insolation influence the terrestrial biome of the Caspian Sea region, causing asynchronous changes of vegetation cover (Leroy et al. 2013 and references therein). Deciphering of their effects for the older periods covered by the fossil record in the Middle Kura Basin greatly depends on the precision of the time-scale and extent of the fossil evidence.
Filippova (1997) depicts a highly dynamic, oscillating picture of the environment in the Middle Kura Basin. The vegetation cover was changing from semi-desert xerophylous to mesophilous forested landscape. Presence of the mesophilous biome here was a result of range expansion of the forests developed on the southern slopes of the Great and north-eastern slopes of the Lesser Caucasus during favorable conditions. This kind of alternation does not fit the hypothesis of a humid, forest biome refugium in the Middle Kura Basin.

Fig. 5. Tentative correlations of the Middle Kura Basin fossil vertebrate sites. Geomagnetic Polarity Time scale (GPTS) according to Ogg (2012). Regional stages: Vasiliev et al. (2011), Radionova et al. (2012), Van Baak et al. (2012). Boundaries between geological formations: Buleishvili (1960), Vangengeim and Tesakov (2008). European Land Mammal Ages (ELMA): Hilgen et al. (2012), Nomade et al. (2014). Eastern European Faunal Complexes: Vislobokova (2008), Vislobokova and Tesakov (2013). Biozones: Hilgen et al. (2012).
Conclusions
This critical summary of the terrestrial fossil record of the middle part of the Kura Basin accumulated during the last 100 years shows that:
(i) Two major intervals of terrestrial fossil record are present, spanning ca. 10–7 Ma and ca. 3–1 Ma (Fig. 5). Both intervals coincide with decisive episodes in the evolution of primates, the demise of late Miocene Hominoidea in Eurasia and the first dispersal of early Homo out of Africa.
(ii) The present fossil record supports the existence of an important hiatus in conjunction with the pre-Akchagylian denudation, and even suggests that this gap may be longer than previously thought, since it does not provide any solid evidence for the existence of Pontian fauna (possibly Dzhaparidze site). The Pliocene terrestrial fossil record is extremely scarce (e.g., the lowermost fossil-bearing horizons of Kushkuna) and controversial (e.g., Kvabebi).
(iii) The biogeographic role of the Middle Kura Basin was changing over the geological time span from a refugium (Khersonian) to a full-fledged part of the Greco-Iranian province (Meotian–Pontian). During the Quaternary period the highly dynamic changes of the environment do not depict this area as refugium in its general sense.
(iv) Much field, chronostratigraphic, and taxonomic work remains to be done to uncover the detailed picture of faunal evolution in this part of the South Caucasus. There is enormous paleontological potential in this area. The impressive successions of alternating marine and continental deposits with intercalations of volcanic ash can potentially provide a direct correlation of terrestrial and marine records through absolute dating, making this place promising for complex, interdisciplinary research on the evolutionary history of life from the late Miocene through Quaternary.
Acknowledgements
The idea to write this article comes from the meeting “Late Miocene to early Pleistocene paleobiology of eastern Caucasus” in Frankfurt, October 9–10 2015, organised by Angela Bruch (Senckenberg, Frankfurt, Germany), David Lordkipanidze (Georgian National Museum, Tbilisi, Georigia), and Friedemann Schrenk (Senckenberg, Frankfurt, Germany). The basis for writing this article are the curating works in the Simon Janashia Museum of Georgia, Georgian National Museum, and introductory field works aiming to rediscover, and further explore the Georgian part of the discussed territory. Authors are grateful to Reed Coil (Nazarbayev University, Astana, Kazakhstan) for editing the text and critical remarks, Angela Bruch for critical remarks, Ira Shatilova and Lamara Maissuradze (both Georgian National Museum, Tbilisi Georgia) for advices. We are grateful to Alexey S. Tesakov (Geological Institute of the Russian Academy of Sciences, Moscow, Russia) and Vadim V. Titov (Institute of Arid Zones, Russian Academy of Sciences, Rostov-on-Don, Russia) and one anonymous reviewer for their insightful comments and suggestions. These works were conducted in the framework of the following projects: NSF RHOI project upper Miocene of Georgia (2005–2007); Pleistocene NSF project #BCS-1019408 (2011–2013); Volkswagen project #85 820 2 (2010–2014); SNF IZ73Z0 152380 (2012–2014); SNF IZ73Z0 127940 (2014–2016); Paleobiomics Project http://www.paleobiomics.org; Rustaveli foundation projects: 1-5/23 (2010–2012), #11/05 (2012–2015), #217626 (2016–2019).
References
Adamia, S., Alania, V., Chabukiani, A., Chichua, G., Enukidze, O., and Sadradze, N. 2010. Evolution of the Late Cenozoic basins of Georgia (SW Caucasus): a review. Geological Society, London, Special Publications 340: 239–259. Crossref
Agustí, J., Sanz de Siria, A., and Garcés, M. 2003. Explaining the end of hominoid experiment in Europe. Journal of Human Evolution 45: 145–153. Crossref
Agustí, J., Vekua, A., Oms, O., Lordkipanidze, D., Bukhsianidze, M., Kiladze, G., and Rook, L. 2009. The Pliocene–Pleistocene succession of Kvabebi (Georgia) and the background to the early human occupation of South Caucasus. Quaternary Science Reviews 28: 3275–3280. Crossref
Akhalkatsi, M. and Tarkhnishvili, D. 2012. Habitats of Georgia. 118 pp. WWF, Tbilisi.
Alania, V., Chabukiani, A., Chagelishvili, R., Enukidze, O., Gogrichiani, K., Razmadze, A., and Tsereteli, N. 2016. Growth structures, piggyback basins and growth strata of Georgian part of Kura foreland fold and thrust belt: implication for Late Alpine kinematic evolution. Geological Society, London, Special Publications 428: 171–185. Crossref
Ali-Zade, A.A. 1961. Akčagil Turkmenistana. 300 pp. Gosgeoltehizdat, Moskva.
Ali-Zade, A.A. 1973. Apšeron Azerbajdžana. 228 pp. Nedra, Moskva.
Ali-Zade, A.A., Alizade, K.A., Aleskerov, D.A., Buleishvili, D.A., Vekua, A.K., Konstantinova, N.A., Lebedeva, N.A., Negadaev-Nikonov, K.N., Bikiforova, K.V., Pevzner, M.A., Khubka, A.N, Chepalyga, A.L., and Chernyakhovsky, A.G. 1972. Guidbook Excursions in Moldavia, Georgia, Azerbaijan May–June, 1972. 104 pp. INQUA Subcommission on the Pliocene/Quaternary Boundary, IUGS Subcommission on Neogene Stratigraphy, Geological Institute of the Academy of Sciences of the USSR, Department of Paleontology and Stratigrapby of the Academy of Sciences of Moldavian SSR, Institute of Paleobiology of the Academy of Sciences of Georgian SSR, Institute of Geology of the Academy of Sciences of Azerbaijan SSR, Azerbaijan Research Design Institute of Oil Prospecting.
Alizade, K.A. 1954. Akčagil’skij Ârus Azerbajdžana. 336 pp. AS AzSSR, Baku.
Andrianov, K.C. and Larin, N.I. 1935. Conditions of fossilization of bones of the upper Sarmatian vertebrates in the step El’dar [in Russian]. Bûlleten’ Moskovskogo Obŝestva Ispytatelej Prirody, Otdel Geologičeskij, Novaâ Seriâ 43 (4): 559–569.
Bakradze, M.A. and Chkhikvadze, V.M. [Čhikvadze, V.M.] 1984. Fossil terrestrial turtles of the genus Testudo in the USSR [in Russian]. Vestnik Gosudarstvennogo Muzeâ Gruzii 33A: 163–174.
Begun, D.R. 2001. African and Eurasian Miocene hominoids and the origins of the Hominidae. In: L. de Bonis, G.D. Koufos, and P. Andrews (eds.), Hominoid Evolution and Climate Change in Europe, Vol. 2: Phylogeny of the Neogene Hominoid Primates of Eurasia, 231–253. Cambridge University Press, Cambridge. Crossref
Begun, D.R. 2010. Miocene Hominids and the Origins of the African Apes and Humans. The Annual Review of Anthropology 39: 67–84. Crossref
Bendukidze, O.G. and Burchak-Abramovich, N.O. [Burčak-Abramovič, N.I.] 1990. Fossil beavers from the Miocene deposits of eastern Georgia [in Russian]. In: I.G. Taktakišvili (ed.), Flora i Fauna Mezo-Kainozoâ Gruzii, 145–151. Mecniereba, Tbilisi.
Bendukidze, O.G. and Vekua, A.K. 2012. The fossil marten from Kvabebi (eastern Georgia, Kakheti) [in Russian with abstract in English]. Sak’art’velos Erovnuli Muzeumis Mac’ne Sabunebismetqvelo da Preistoriuli Sek’c’ia 4: 64–70.
Bendukidze, O.G., Vanishvili, N. [Vanišvili, N.], and Vekua, A.K. 2013. Simocyon—a new element in the Udabno mammal fauna [in Russian with abstract in English]. Sak’art’velos Erovnuli Muzeumis Mac’ne Sabunebismetqvelo da Preistoriuli Sek’c’ia 5: 97–104.
Bernor, R.L., Solounias, N., Swisher III, C.C., and Van Couvering, J.A. 1996. The correlation of three classical “Pikermian” mammal faunas—Maragheh, Samos, Pikermi—with the European MN Unit System. In: R.L. Bernor, V. Fahlbusch, and H.-W. Mittman (eds.), The Evolution of Western Eurasian Neogene Mammal Faunas, 137–154. Columbia University Press, New York.
Birks, H.J.B. and Tinner, W. 2016. Past forests of Europe. In: J. San-Miguel-Ayanz, D. de Rigo, G. Caudullo, T. Houston Durrant, and A. Mauri (eds.), European Atlas of Forest Tree Species, 36–39. Publications Office of the European Union, Luxembourg.
Bukhsianidze, M., Chagelishvili, R., and Lordkipanidze, D. 2018. Late Miocene vertebrate site of Chachuna (Iori valley, Georgia, Southern Caucasus). Bulletin of the Georgian National Academy of Sciences 12 (2): 70–75.
Buleishvili, D.A. [Buleišvili, D.A.] 1960. Geologiâ i Neftegazenosnost’ Mežgorni Vpadiny Vostočnoj Gruzii. 238 pp. Gostoptehizdat, Leningrad.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] 1957. Fossil goose from the Udabno hipparion fauna [in Russian]. Doklady Akademii Nauk Azerbajdžanskoj SSR 13: 655–659.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] and Gabashvili, E.G. [Gabašvili, E.G.] 1945. A superior anthropoid ape from the Late Tertiary deposits of east Georgia [in Georgian and Russian with English abstract]. Soobŝeniâ Akademii Nauk Gruzinskoj SSR 6 (6): 451–464.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] and Gabashvili, E.G. [Gabašvili, E.G.] 1946. A superior antropoid from the Upper Tertiar deposits of the eastern Georgia [in Georgian and Russian with abstract in English]. Vestnik Gosudarstvennogo Muzeâ Gruzii 13A: 235–252.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] and Gabashvili, E.G. [Gabašvili, E.G.] 1980. Fossil beavers in the Caucasus [in Russian]. Izvestiâ Geologičeskogo Obŝestva Gruzii 6: 20–34.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] and Gabashvili, E.G. [Gabašvili, E.G.] 1984. Fossil gull in the hipparion fauna Garedjis Udabno [in Russian]. Vestnik Gosudarstvennogo Muzeâ Gruzii 33A: 3–13.
Burchak-Abramovich, N.I. and Gadzyev, D.V. 1978. Anser eldaricus sp. nov. from upper Sarmatian hipparion fauna of Eldar. Acta Zoologica Cracoviensia 23: 67–68.
Burchak-Abramovich, N.I. and Meladze, G.K. 1972. Rustaviornis georgicus gen. et sp. n., a new fossil bird from the Hipparion fauna of Georgia. Acta Zoologica Cracoviensia 17: 373–388.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] and Vekua, A.K. 1980. Paleobiologičeskaâ istoriâ pozdnekajnozoiskih bykov Kavkaza. 92 pp. Mecniereba, Tbilisi.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.] and Vekua, A.K. 1981. Fossil bustard-runner of East Georgian Akchagil [in Russian with abstract in English]. Soobŝeniâ Akademii Nauk Gruzinskoj SSR, Seriâ Biologičeskaâ 7 (1): 53–59.
Burchak-Abramovich, N.I. [Burčak-Abramovič, N.I.], Gadzhiev, D.V. [Gadžiev, D.V.], and Vekua, A.K. 1980. Concerning ancestral form of bisons from the Akchagyl of South Caucasus [in Russian]. Soobŝeniâ Akademii Nauk Gruzinskoj SSR 97 (2): 485–488.
Casanovas-Vilar, I., Alba, D.M., Garcés, M., Roblesa, J.M., and Moyà-Solà, S. 2011. Updated chronology for the Miocene hominoid radiation in Western Eurasia. Proceedings of the National Academy of Sciences 108 (14): 5554–5559. Crossref
Casanovas-Vilar I., Moyá-Solá S., Agustí J., and Köhler M. 2005. The geography of a faunal turnover: tracking the Vallesian crisis. In: A.M.T. Elewa (eds.), Migration of Organisms, 247–300. Springer, Berlin. Crossref
Chelidze, G. 1955. [Čelidze, G.] The geological structure of the Kura Gorge between city Rustavi and the Krasnyj bridge [in Georgian, with summary in Russian]. Trudy Geologičeskogo Instituta Akademii Nauk Gruzinskoj SSR, Seriâ Geologičeskaâ 7 (13): 186–215.
Chkhikvadze, V.M. and Amiranashvili, N.G. 1996. History of the fossil turtles studies in the Caucasus. Bulletin of the Georgian Academy of Sciences 154 (3): 470–474.
Chkhikvadze, V.M. [Čhikvadze, V.M.] and Petrov, V.A. 2000. New late Neogene turtle localities in the Iori Valley (eastern Georgia) [in Russian]. In: N.B. Ananjeva, I. Darevskij, N.N. Dunaev, N.N. Iordanskij, S.L. Kuzmin, and N.L. Orlov (eds.), Voprosy Gerpetologii, Materialy Pervogo S”ezda Gerpetologičeskogo Obŝestva Imeni A.M. Nikol’skogo, 4–7 Dekabrâ 2000, 332–334. Puŝino, Moskva.
Chkhikvadze, V.M. [Čhikvadze, V.M.], Mchedlidze, G.A. [Mčedlidze, G.A.], Burchak-Abramovich, N.O. [Burčak-Abramovič, N.O.], Bendukidze, O.G., Burchak, D.N. [Burčak, D.N.], Gabelaia, T.D., Amiranashvili, N.G. [Amiranašvili, N.G.], Meladze, G.K., and Kharabadze, E.S. [Harabadze, E.S.] 2000. Review of the localities of tertiary vertebrates of Georgia [in Georgian, with abstract in Russian]. Trudy Geologičeskogo Instituta Imeni A.I. Džanelidze Akademii Nauk Gruzinskoj SSR Novaâ Seriâ 115: 153–162.
Chubinishvili, K.G. [Čubinišvili, K.G.] 1982. Neogenovye Kontinental’nye Molassy Vostočnoj Gruzii. 314 pp. Mecniereba, Tbilisi.
Chumakov, I.S. [Čumakov, I.S.], Byzova, S.L., and Ganzei, S.S. 1992. Geohronologiâ i Korrelâtsiâ Pozdnego Kainozoâ Paratetisa. 97 pp. Nauka, Moskva.
Danukalova, G.A. 1990. Dvustvorčatye Mollûski iz Stratigrafiâ Akčagila. 367 pp. Ph.D. Thesis, Paleontologičeskij Institut, Akademiâ Nauk SSSR, Moskva.
Dombrovskij, V.S. 1914. A note on the geological conditions of the location of the bones of fossil mammals in El’dar (Tiflis province) [in Russian]. Trudy Geologičeskogo Muzeâ Imeni Petra Veliokogo Imperatorskoj Akademii Nauk 8 (14): 231—238.
Dubrovo, I.A. and Burchak-Abramovich, N.I. 1986. New data on the evolution of Bovine of the tribe Bovini. Quartärpälaontologie 6: 13–21.
Dzigrashvili, A.T. [Dzigrašvili, A.T.] and Sepashvili, O.A. [Sepašvili, O.A.] 1978. The conditions of deposition of the Tsteltskaro limestones [in Russian]. Trudy Vsesoûznogo Neftânogo Naučno-Issledovatel’skogo Geologorazvedočnogo Instituta 115 (14): 201–209.
Eisenmann, V. and Sondaar, P. 1989. Hipparions and the Mio-Pliocene boundary. Bulletino della Società Paleontologica Italiana 28 (2–3): 217–226.
Eronen, J.T. and Rook, L. 2004. The Mio-Pliocene European primate fossil record: dynamics and habitat tracking. Journal of Human Evolution 47: 323–341. Crossref
Eronen, J.T., Ataabadi, M.M., Micheels, A., Karmea, A., Bernor, R.L., and Fortelius, M. 2009. Distribution history and climatic controls of the Late Miocene Pikermian chronofauna. Proceedings of the National Academy of Sciences 106 (29): 11867–11871. Crossref
Fataliev, R.A. 1964. Verhnesarmatskaâ Flora Gory Katar v Meždureč’e Kury i Iori. 17 pp. Synopsis of the Ph.D. Thesis, Botaničeskij institut imeni V.L. Komarova, Akademiâ Nauk SSSR, Leningrad.
Ferring, R., Oms, O., Agustí, J., Berna, F., Nioradze, M., Shelia, T., Tappen, M., Vekua, A., Zhvania, D., and Lordkipanidze, D. 2011. Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85–1.78 Ma. Proceedings of the National Academy of Sciences 108 (26): 10432–10436. Crossref
Filippova, N.Y. 1997. Palinologiâ Verhnego Pliotsena–Srednego Plejstotsena Ûga Kaspiiskoj Oblasti. 164 pp. GEOS, Moskva.
Forte, A., Cowgill, E., Bernardin, T., Kreylos, O., and Hamann, B. 2010. Late Cenozoic deformation of Kura fold-thrust belt, southern Greater Caucasus. Geological Society of America Bulletin 122: 465–486. Crossref
Forte, A., M., Sumner, D. Y., Cowgill, E., Stoica, M., Murtuzayev, I., Kangarli, T., Elashvili, M., Godoladze, T., and Javakhishvili, Z. 2015. Late Miocene to Pliocene stratigraphy of the Kura Basin, a subbasin of the South Caspian Basin: implications for the diachroneity of stage boundaries. Basin Research 27 (3): 247–271. Crossref
Gabashvili, E.G. [Gabašvili, E.G.] 1956. Miocene antelopes hold in the collections of the Georgian State Museum [in Georgian with extended summary in Russian]. Vestnik Gosudarstvennogo Muzeâ Gruzii 17A: 15–26.
Gabashvili, E.G. [Gabašvili, E.G.] 1983. Late Miocene proboscideans of Udabno [in Russian]. Vestnik Gosudarstvennogo Muzeâ Gruzii 32A: 29–56.
Gabashvili, E.G. [Gabašvili, E.G.] 1987. History of development of the Late Miocene bovids from Udabno [in Russian]. Vestnik Gosudarstvennogo Muzeâ Gruzii 34A: 3–44.
Gabashvili, E.G. [Gabašvili, E.G.] 2002. Gareǰis Xerxemliant’a P’auna. 86 pp. Georgian centre for prehistory researches, Tbilisi.
Gabunia, L.K. [Gabuniâ, L.K.] 1959. K Istorii Gipparionov. 570 pp. Akademiâ Nauk SSSR, Moskva.
Gabunia, L.K. [Gabuniâ, L.K.] and Vekua, A.K. 1963. Taribanis Namarxi Spilo. 67 pp. Akademiâ Nauk Gruzinskoj SSR, Tbilisi.
Gabunia, L.K. and Vekua, A.K. 1998. The find of Dinopelis in the Pliocene of Georgia. Bulletin of the Georgian Academy of Sciences 157 (2): 335–338.
Gabunia, L.K., Gabashvili, E.G., Vekua, A.K., and Lordkipanidze, D. 2001. The Late Miocene hominoid from Georgia. In: L. de Bonis, G.D. Koufos, and P.J. Andrews (eds.), Hominoid Evolution and Climate Change in Europe. Vol. 2: Phylogeny of the Neogene Hominoid Primates in Eurasia, 316–325. Cambridge University Press, Cambridge. Crossref
Gabunia, L.K. [Gabuniâ, L.K.], Matskhonashvili, K.G. [Matshonašvili, K.G.], and Chkheidze, D.V. [Čheidze, D.V.] 1966. On the age of continental deposits of the Iagluja Mountain [in Georgian, with abstract in Russian]. Soobŝeniâ Akademii Nauk Gruzinskoj SSR 43 (1): 129–132.
Gadzhiev, D.V. [Gadžiev, D.V.] 1996. El’darskaâ Verhnesarmatskaâ Gipparionovaâ Fauna. 344 pp. ELM, Baku.
Gagnidze, R. and Davitadze, M. 2000. Adgilobrivi P’lora (Sak’art’velos Mc’enaret’a Samqaro). 274 pp. Ač’ara, Batumi.
Gamkrelidze, I., Gamkrelidze, M., Loladze, M., and Tsamalashvili, T. 2015. New tectonic map of Georgia (explanatory note). Bulletin of the Georgian National Academy of Sciences 9 (1): 211–216.
Ganzej, S.S. 1987. Late Caenosoic deposits of Ponto-Caspian and fission-treck dating of ash layers [in Russian]. In: A.M. Korotkij (ed.), Metod Trekov v Geologii i Geografii, 34–45. Dal’nevostočnyj Naučnyj Centr, Tihookeanskij Institut Geografii, Akademiâ Nauk SSSR, Vladivostok.
Golubjatnikov, D.V. [Golubâtnikov, D.V. ] 1927. Detailed geological map of Absheron peninsula. Atashan district [in Russian]. Trudy Geologičeskoj Komissii, Novaâ Seriâ 130: 1–314.
Grichuk, V.P. [Gričuk, V.P.] 1969. Glacial floras and their classification [in Russian]. In: I.P. Gerasimov (ed.), Poslednij Lednikovyj Pokrov Na Severo-Zapade Evropeiskoj Časti SSSR, k VIII Kongressu INQUA Pariž 1969, 57–70. Nauka, Moskva.
Gromov, V.I. 1948. Palaeontological and archaeological background for the stratigraphy of the continental Quaternary deposits on USSR territory [in Russian]. Trudy Instituta Geologičeskih Nauk AN SSSR, Seriâ Geologičeskaâ 64 (17): 1–521.
Grossheim, A.A. 1948. Rastitel’nyj Pokrov Kavkaza. 267 pp. Akademiâ Nauk SSSR, Moskva.
Gurarij, G.Z., Kudasheva, I.A., Trubikhin, V.M., Fein, A.G., and Shipunov, S.V. 1995. Age of the Natskhor and Shiraki Formations of Eastern and Central Georgia and the problem of the Miocene–Pliocene boundary—paleomagnetism and stratigraphy. Stratigraphy and Geological Correlation 3: 156–166.
Hemmer, H., Kahlke, R.-D., and Vekua, A.K. 2004. The Old World puma – Puma pardoides (Owen, 1846) (Carnivora: Felidae)—in the lower Villafranchian (upper Pliocene) of Kvabebi (East Georgia, Transcaucasia) and its evolutionary and biogeographical significance. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 233: 197–231.
Hilgen, F., Lourens, L., Van Dam, J.A., Beu, A., Boyes, A.F., Cooper, R., Krijgsman, W., Ogg, J., Piller, W., and Wilson, D. 2012. The Neogene Period. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), The Geologic Time Scale, 923–978. Elsevier, Boston. Crossref
Isaeva, M.I. and Mamedova, N.R. 1989. Paleomagnetic studies of the major sections of the Middle Kura depression (Pliocene–Pleistocene) [in Russian]. Izvestiâ Akademii Nauk Azerbajdžanskoj SSR, Seriâ Nauka o Zemle 3: 123–127.
Khuchua, M.F. [Hučua, M.F.] and Shatirishvili, T.M. [Šatirišvili, T.M.] 1937. Mineralogical composition of sandy deposits of the upper Sarmatian and Shiraki Formation in the sections of Akhtatapa, Kotsakhuriskedi [in Russian]. Azerbajdžanskoje Neftânoe Hozâistvo 2: 9–17.
Koiava, K., Maissuradze, L., Strasser, A., Shatilova, I., Kvaliashvili, L., and Glonti, V. 2012. Palaeogeography of the Sarmatian of eastern Georgia. Bulletin of the Georgian National Academy of Sciences 6 (3): 91–98.
Kolesnikov, V.P. 1940. Scheme of stratigraphic subdivision of Neogene deposits of the Ponto-Caspian region [in Russian]. In: A.D. Arhangel’skij (ed.), Neogen. Stratigrafiâ SSSR 12: 567–574.
Kolesnikov, V.P. 1950. Akčagil’skie i Apšeronskie Mollûski. Paleontologiâ SSSR, Vol. 10, Part 3 (12). 259 pp. Akademiâ Nauk SSSR, Moskva.
Kostopoulos, D.S. 2014. Taxonomic reassessment and phylogenetic relationships of Miocene homonymously spiral-horned antelopes. Acta Palaeontologica Polonica 59 (1): 9–29.
Kostopoulos, D.S. and Bernor, R.L. 2011. The Maragheh bovids (Mammalia, Artiodactyla): systematic revision and biostratigraphiczoogeographic interpretation. Geodiversitas 33 (4): 649–708. Crossref
Kudrjavtsev, N.A. [Kudrâvtsev, N.A.] 1938. Geological research in the Alazani and Kura region (brief preliminary report for the years 1930–1931) [in Russian]. Trudy Neftânogo Geologo-Razvedovočnogo Instituta Seriâ B 32:1–58.
Kuzmina, I.E. and Sablin, M.V. 1991. On new finds of fossil remains of Equus stenonis Cocchi in the South Caucasus [in Russian]. Trudy Zoologičeskogo Instituta Akademii Nauk SSSR 238: 61–67.
Lebedeva, N.A. 1972. Geological position of the remains of terrestrial mammals of Khapry, Taman’ and Tiraspol faunistical complexes in the cut of marine layers of Akchagyl and Apsheron of eastern Transcaucasia [in Russian]. Byulleten’ Komissii po Izučeniû Četvertičnogo Perioda 38: 99–117.
Lebedeva, N.A. 1974. Korrelâtsiâ Morskih i Kontinental’nyh Otloženij Eopleistotsena i Nižnego Pleistotsena Ponto-Kaspijskoj Oblasti. 53 pp. Synopsis of the Ph.D. Thesis, Geologičeskij institut, Akademiâ Nauk SSSR, Moskva.
Lebedeva, N.A. 1978. Korrelâciâ Antropogenovyh Tolŝ Ponto-Kaspiâ. 135 pp. Nauka, Moskva.
Leroy, S.A.G., Tudryn, A., Chalié, F., and López-Merino, L. 2013. From the Allerød to the mid-Holocene: palynological evidence from the South Basin of the Caspian Sea. Quaternary Science Reviews 78: 77–97. Crossref
Lordkipanidze, D., Jashashvili, T., Vekua, A., Ponce de León, M.S., Zollikofer, C.P.E., Rightmire, G.P., Pontzer, H., Ferring, R., Oms, O., Tappen, M., Bukhsianidze, M., Agusti, J., Kahlke, R.-D., Kiladze, G., Martinez-Navarro, B., Mouskhelishvili, A., Nioradze, M., and Rook, L. 2007. Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 449: 305–310. Crossref
Maisadze, F. 2013. Duration of Cenozoic orogenies (on the example of Georgia). Bulletin of the Georgian National Academy of Sciences 7 (2): 97–104.
Meladze, G.K. 1967. Gipparionovaâ Fauna Arkneti i Bazaleti. 168 pp. Mecsniereba, Tbilisi.
Meladze, G.K. 1985. Obzor Gipparionovyh Faun Kavkaza. 75 pp. Mecniereba, Tbilisi.
Mauvilly, J., Koiava, K., Gamkrelidze, I., and Mosar, J. 2016. Tectonics in the Georgian Greater Caucasus: a structural cross-section in an inverted rifted basin setting. In: S. Castelltort, S. Girardclos, E. Gnos, E. Samankassou, S. Scmits, M. Lupi, C. Frischkencht, M. Chiradia, G. Simpson , R. Martini, U. Schaltegger, and D. Ariztegui (eds.), 14th Swiss Geoscience Meeting, Geneva, Switzerland, November 18–19, 33–34. Swiss Academy of Sciences, Geneva. [published online, doi: 10.13140/RG.2.2.26540.56963]
Mauvilly, J. 2017. Tectonics in the Georgian Greater Caucasus: a Structural Cross Section in an Inverted Rifted Basin Setting. 127 pp. Unpublished M.Sc. Thesis, University of Fribourg, Fribourg.
Nanda, A.C. 2002. Upper Siwalik mammalian faunas of India and associated events. Journal of Asian Earth Sciences 21: 47–58. Crossref
Nargolwalla, M.C. 2009. Eurasian Middle and Late Miocene Hominoid Paleobiogeography and the Geographic Origins of the Homininae. 534 pp. Ph.D. Thesis, University of Toronto Library and Archives Canada, Toronto.
Nemčok, M., Glonti, B., Yukler, A., and Marton, B. 2013. Development history of the foreland plate trapped between two converging orodens; Kura valley, Georgia, case study. Geological Society, London, Special Publications 377: 159–188. Crossref
Nevesskaya, L.A. [Nevesskaâ, L.A.] and Trubikhin, V.M. [Trubihin, V.M.] 1984. The history of the Caspian Basin and its fauna of mollusks in the Late Pliocene and Early Pleistocene [in Russian]. In: V.L. Jahimovič (ed.), Antropogen Evrazij, 19–27. Nauka, Moskva.
Nikiforova, K.V., Krasnov, I.I., Aleksandrova, L.P., Vasiliev, Y.M., Konstantinova, N.A., and Tshepalyga, A.L. [Tšepalyga, A.L.] 1982. Chronostratigraphic scheme of the Late Pliocene–Antropogene deposits of the European part of the USSR [in Russian]. In: D.V. Nalivkin and B.S. Sokolov (eds.), Stratigrafiâ SSSR, Četvertičnaâ Sistema 1, unnumbered insert. Nedra, Moskva.
Nomade, S., Pastre, J.F., Guillou, H., Faure, M., Guérin, C., Delson, E., Debard, E., Voinchet, P., and Messager, E. 2014. 40Ar/39Ar constraints on some French landmark Late Pliocene to Early Pleistocene large mammalian paleofaunas: Paleoenvironmental and paleoecological implications. Quaternary Geochronology 21: 2–15 Crossref.
Ogg, J.G. 2012. Geomagnetic Polarity Time Scale. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), The Geologic Time Scale, 85–114. Elsevier, Boston. Crossref
Orlov, Y.A. 1941. A new locality of hipparion-fauna at Djaparidze, east Georgia [in Russian with abstract in English]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 8 (3): 99–101.
Palmqvist, P., Martínez-Navarro, M., Pérez-Claros, J.A., Torregrosa, V., Figueirido, B., Jiménez-Arenas, J.M., Espigares, M.-P., Ros-Montoya, S., De Renzi, M. 2011. The giant hyena Pachycrocuta brevirostris: Modelling the bone-cracking behavior of an extinct carnivore. Quaternary International 243 (1): 61–79. Crossref
Pevzner, M.A. 1982. Application of paleomagnetic method for the stratigraphy of Quaternary deposits [in Russian]. In: D.V. Nalivkin and B.S. Sokolov (eds.), Stratigrafiâ SSSR, Četvertičnaâ Sistema 1, 149–154. Nedra, Moskva.
Pickford, M. 2013. Re-assessment of the suids from the Sables marins de Montpellier and selection of a lectotype for Sus provincialis Blainville, 1847. Geodiversitas 35 (3): 655–689. Crossref
Popov, G.I. 1969. Stratigraphic scheme of the Akchagylian stage [in Russian]. Geologiâ i Poleznie Iskopaemye Turkmenii 2: 179–224.
Radionova, E.P., Golovina, L.A., Filippova N.Y., Trubikhin, V.M., Popov, S.V., Goncharova, I.A., Vernigorova, Y.V., and Pinchuk, T.N. 2012. Middle–Upper Miocene stratigraphy of the Taman Peninsula, Eastern Paratethys. Central European Journal of Geosciences 4 (1): 188–204. Crossref
Riabinin, A. 1913. Recherches géologiques dans la region de la steppe de Chiraki et de ses alentours [in Russian with summary in French]. Trudy Geologičeskogo Komiteta, Novaâ Seriâ 93: 1–73.
Rook, L., Bartolini Lucenti, S., Bukhsianidze, M., and Lordkipanidze, D. 2017. The Kvabebi Canidae record revisited (Late Pliocene, Sighnaghi, eastern Georgia). Journal of Paleontology 91 (6): 1258–1271. Crossref
Ruzhentsev, V.E. [Ružentsev, V.E.] 1928. The division of the Apsheronian stage based on the study of the Apsheronian deposits of the Lesser Kharami Range [in Russian]. Bûlleten’ Moskovskogo Obŝestva Ispytatelej Prirody, Otdel Geologičeskij, Novaâ Seriâ 36 (6): 241–266.
Sablin, M.V. 1990. Remains of carnivores and ungulates from the Lower Absheronian deposits of Azerbaidjan [in Russian]. Trudy Zoologičeskogo Instituta Akademii Nauk SSSR 213: 138–142.
Safarov, I.S. 1979. Subtropičeskie Lesa Talyša. 157 pp. ELM, Baku.
Sahni, M.R. and Khan, E. 1968. Probison dehmi n. g. n. sp. a recent find of an upper Sivalik bovid. Bayerische Staatssammlung für Paläontologie und Historische Geologie 8: 247–251.
Semenov, Y.A. 1989. Iktiterii i Morfologičeski Shodnye Geny Neogena SSSR. 178 pp. Naukova Dumka, Kiev.
Sen, S. 1997. Magnetostratigraphic calibration of the European Neogene mammal chronology. Palaeogeography, Palaeoclimatology, Palaeoecology 133: 181–204. Crossref
Shatilova, I. and Kokolashvili, I. 2011. The flora and vegetation of eastern Georgia in the Sarmatian. Proceedings of the Georgian Academy of Sciences, Biological Series B 9 (1–4): 65–70.
Shatilova, I. and Kokolashvili, I. 2012. The comparison of eastern and western Georgia’s Sarmatian vegetation and climate. Proceedings of the Georgian National Museum, Natural Sciances and Prehistory Section 4: 31–36.
Shatilova, I. [Šatilova, I.] and Kokolashvili, I. [Kokolašvili, I.] 2013. Palinologičeskaâ Harakteristika Sarmatskih Otloženii Vostočnoj Gruzii [in Russian with English summary]. 110 pp. Universal, Tbilisi.
Shatilova, I., Maissuradze, L., Kokolashvili, I., and Koiava, K. 2012. The comparison of the results of palynological and microfaunistical investigations of the Sarmatian deposits of eastern Georgia. Bulletin of the Georgian National Academy of Sciences 6 (2): 139–144.
Shatilova, I., Mchedlishvili, N., Rukhadze, L., and Kvavadze, E. 2011. The History of the Flora and Vegetation of Georgia (South Caucasus). 200 pp. Universal, Tbilisi.
Shchelinsky, V.E. [Ŝelinskij, V.E.] 2014. Eopleistotsenovaâ Rannepaleolitičeskaâ Stoânka Rodniki 1 v Zapodnom Predkavkaz’e. 168 pp. IIMC RAS, Saint-Petersburg.
Shchelinsky, V.E., Gurova, M., Tesakov, A.S., Titov, V.V., Frolov, P.D., and Simakova, A.N. 2016. The Early Pleistocene site of Kermek in western Ciscaucasia (southern Russia): Stratigraphy, biotic record and lithic industry (preliminary results). Quaternary International 393: 51–69. Crossref
Sotnikova, M.V. 1989. Hiŝnye Mlekopitaûŝie Pliotsena—Rannego Plejstotsena. 123 pp. Nauka, Moskva.
Sotnikova, M.V. and Rook, L. 2010. Dispersal of the Canini (Mammalia, Canidae: Caninae) across Eurasia during the Late Miocene to Early Pleistocene. Quaternary International 212: 86–97. Crossref
Sotnikova, M.V. and Sablin, M.V. 1993. The late Villafranchian association of carnivorous mammals from the locality Palan-Tyukan [in Russian with abstract in English]. Trudy Zoologičeskogo Instituta Akademii Nauk SSSR 249: 134–145.
Sultanov, K.M. 1964. Apšeronskij Ârus Azerbajdžana. 233 pp. Azernašr, Baku.
Tappen, M., Adler, D.S., Ferring, C.R., Gabunia, M., Vekua, A., and Swisher III, C.C. 2002. Akhalkalaki: The taphonomy of an Early Pleistocene locality in the Republic of Georgia. Journal of Archaeological Science 29: 1367–1391. Crossref
Tarkhnishvili, D. 2014. Historical Biogeography of the Caucasus. 180 pp. Nova Science Publishers Inc., New York.
Tesakov, A.S. 2004. Biostratigraphy of middle Pliocene–Eopleistocene of Eastern Europe (based on small mammals) [in Russian]. Trudy Geologičeskogo Instituta 554: 1–247.
Tesakov, A.S., Titov, V.V., Simakova, A.N., Florov, P.D., Syromyatnikova, E.V., Kurshakov, S.V., Volkova, N.V., Trikhunkov, Y.I., Sotnikova, M.V., Kruskop, S.V., Zelenkov, N.V., Tesakova, E.M., and Palatov, D.M. 2017. Late Miocene (Early Turolian) vertebrate faunas and associated biotic record of the Northern Caucasus: geology, taxonomy, palaeoenvironment, biochronology. Fossil Imprint 73 (3–4): 383–444.
Trubikhin, V.M. [Trubihin, V.M.] 1977. Paleomagnetizm i stratigrafiâ akčagil’skih otloženii Zapodnoj Turkmenii. 77 pp. Nauka, Moskva.
Tsiskarishvili, G.V. [Tsiskarišvili, G.V.] 1987. Pozdnetretičnye Nosorogi (Rhinocerotidae) Kavkaza. 142 pp. Mecniereba, Tbilisi.
Utescher, T., Erdei, B., Hably, L., and Mosbrugger, V. 2017. Late Miocene vegetation of the Pannonian Basin. Palaeogeography, Palaeoclimatology, Palaeoecology 467: 131–148. Crossref
Van Baak, C.G.C., Vasiliev, I., Stoica, M., Kuiper, K.F., Forte A.M., Aliyeva, E., and Krijgsman, W. 2013. A magnetostratigraphic time frame for Plio-Pleistocene transgressions in the South Caspian Basin, Azerbaijan. Global and Planetary Change 103: 119–134. Crossref
Vangengeim, E.A. and Tesakov, A.S. 2008. Late Sarmatian mammal localities of the Eastern Paratethys: Stratigraphic position, magnetochronology, and correlation with the European continental scale. Stratigraphy and Geological Correlation 16 (1): 92–103.
Vangengeim, E.A., Muskhelishvili, L.V. [Mušelišvili, L.V.], Pevzner, M.A., and Tsiskarishvili, G.V. [Tsiskarišvili, G.V.] 1988. Place of the upper boundary of Vallesian on the stratigraphic scheme of the Eastern Paratethys [in Russian]. Izvestiâ Akademii Nauk SSSR, Seriâ Geologičeskaâ 6: 67–74.
Vangengeim, E.A., Gabunia, L.K. [Gabuniâ, L.K.], Pevzner, M.A., and Tsiskarishvili, G.V. [Tsiskarišvili, G.V.] 1989. Stratigraphic position of the sites with hipparion faunas from the South Caucasus in the light of the data of magnetostratigraphy [in Russian]. Izvestiâ Akademii Nauk SSSR, Seriâ Geologičeskaâ 8: 70–77.
Vanishvili, N., Vekua, A.K., and Kiladze, G. 2007. Simocyon from the Late Miocene of Georgia. Problems of Paleobiology 2: 142–146.
Vasiliev, I., Iosifidi, A.G., Khramov, A.N., Krijgsman, W., Kuiper, K., Langereis, C.G., Popov, V.V., Stoica, M., Tomsha, V.A., and Yudin, S.V. 2011. Magnetostratigraphy and radio-isotope dating of Upper Miocene–Lower Pliocene sedimentary successions of the Black Sea Basin (Taman Peninsula, Russia). Palaeogeography, Palaeoclimatology, Palaeoecology 310 (3): 163–175. Crossref
Vekua, A.K. 1972. Kvabebskaâ Fauna Akčagil’skih Pozvonočnyh. 351 pp. Nauka, Moskva.
Vekua, A.K. 1991. History of the animal world—vertebrates [in Russian]. In: L.I. Maruašvili (ed.), Gruziâ v Antropogene, 308–339. Sak’art’velo, Tbilisi.
Vekua, A.K. and Lordkipanidze, D. 1998. The Pleistocene paleoenvironment of the Transcaucasus. Quaternaire 9 (4): 261–266. Crossref
Vekua, A.K., Lordkipanidze, D., and Bukhsianidze, M. 2009. Importance of Diliska Vertebrate Fauna for the Biostratigraphy of South Caucasus. Bulletin of the Georgian National Academy of Sciences 3 (2): 130–135.
Vekua, A.K., Lordkipanidze, D., Bukhsianidze, M., and Chagelishvili, R. 2010. Kotsakhuri (eastern Georgia) Pliocene fauna of vertebrates and Taribana elephant. Proceedings of the Georgian Academy of Sciences, Biological Series B 8 (1–2): 79–84.
Vekua, A.K. and Trubikhin, V.M. [Trubihin, V.M.] 1988. New site of fossil mammals in eastern Georgia [in Russian with Georgian and English abstract]. Soobŝeniâ Akademii Nauk Gruzinskoj SSR 132 (1): 197–200.
Vislobokova, I.A. 2008. The major stages in the evolution of artiodactyl communities from the Pliocene–early Middle Pleistocene of northern Eurasia: Part 1. Paleontological Journal 42: 297–312. Crossref
Vislobokova, I.A. and Tesakov, A.S. 2013. Early and Middle Pleistocene of Northern Eurasia. In: S.A. Elias (ed.), The Encyclopedia of Quaternary Sciences, 605–614. Elsevier, Amsterdam.
Werdelin, L. 2001. A revision of the genus Dinofelis (Mammalia, Felidae). Zoological Journal of the Linnean Society 132: 147–258. Crossref
Zohary, M. 1973. Geobotanical Foundations of the Middle East. 739 pp. Verlag, Stuttgart.
Acta Palaeontol. Pol. 63 (3): 441–461, 2018
https://doi.org/10.4202/app.00499.2018