
Systematic revision and phylogenetic assessment of the foraminiferal family Globivalvulinidae
VALERIO GENNARI, MARCO CHERIN, and ROBERTO RETTORI
Gennari, V., Cherin, M, and Rettori, R. 2018. Systematic revision and phylogenetic assessment of the foraminiferal family Globivalvulinidae. Acta Palaeontologica Polonica 63 (4): 807–814.
We present a systematic and phylogenetic revision of the Carboniferous–Permian subfamilies usually included within the family Globivalvulinidae (superfamily Biseriamminoidea), based for the first time on the interrelation between a morphological/structural approach (traditionally used for this fossil group) and a cladistic analysis (rarely applied to Palaeozoic Foraminifera). The systematic review of the globivalvulinid genera defines a new composition of the already existing four subfamilies of Globivalvulinidae (Globivalvulininae, Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae). The phylogenetic analysis of the Globivalvulinidae is aimed at understanding and reconstructing the relationships between its four subfamilies. The resulting phylogenetic tree is obtained by means of fifteen qualitative and one quantitative characters. The analysis shows that the Globivalvulinidae represents a monophyletic clade having the subfamily Globivalvulininae as the first taxon to branch out. The close affinity between the Dagmaritinae and Paradagmaritinae is confirmed.
Key words: Foraminifera, Biseriamminoidea, Globivalvulinidae, cladistics, phylogeny, taxonomy, Palaeozoic.
Valerio Gennari [valeriogennari@tiscali.it], Marco Cherin [marco.cherin@unipg.it], and Roberto Rettori [roberto.rettori@unipg.it], University of Perugia, Dipartimento di Fisica e Geologia, Via Pascoli, 06123 Perugia, Italy.
Received 4 June 2018, accepted 21 September 2018, available online 10 October 2018.
Copyright © 2018 V. Gennari et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Biserially coiled to uncoiled biseriamminoids represent one of the most interesting groups, both from a stratigraphic and an evolutionary point of view, among Palaeozoic smaller foraminifers. According to Vachard et al. (2006), their biostratigraphic value is probably underestimated during the Carboniferous (Perret 1993; Pinard and Mamet 1998), and some genera such as Tenebrosella Villa and Sanchez de Posada, 1986 and Verispira Palmieri, 1988, remain poorly known and are rarely cited in the literature. Within biseriamminoids, some members of the family Globivalvulinidae Reitlinger, 1950 (order Endothyrida) are important biostratigraphical and palaeogeographical markers in the middle–late Permian together with other families such as the Pseudovidalinidae Altıner, 1988 (order Archaediscida) and the Hemigordiopsidae Nikitina, 1969 (order Cornuspirida). The high biostratigraphic significance of these taxa is due to their wide palaeogeographical distribution and rapid evolution (Vachard et al. 2006, 2017; Gaillot and Vachard 2007; Gaillot et al. 2009; Altıner and Özkan-Altıner 2010; Vachard 2016b).
The taxonomic composition and evolution of biseriamminoid foraminifers is a long-standing debate. It started when Schubert (1921) introduced the genus Globivalvulina choosing Valvulina bulloides Brady, 1876 as the type species, and later Chernysheva (1941) erected the genus Biseriammina (type species Biseriammina uralica Chernysheva, 1941) and assigned it to the family Biseriamminidae Chernysheva, 1941. Loeblich and Tappan (1964) recognised the family Biseriamminidae as composed of the only subfamily Biseriammininae Chernysheva, 1941 considering Globivalvulinae (sic!) Reitlinger, 1950 = Globivalvulininae Pokorny, 1958 as synonyms. Zaninetti and Altıner (1981) divided the Biseriamminidae into two subfamilies Biseriammininae and Dagmaritinae Bozorgnia, 1973, to which the third subfamily Louisettitinae was added by Loeblich and Tappan (1984). Marfenkova (1991) and Gaillot and Vachard (2007) included three families into the superfamily Biseriamminoidea Chernysheva, 1941: Biseriamminidae, Koktjubinidae Marfenkova, 1991, and Globivalvulinidae. Subsequently, Hance et al. (2011) introduced the superfamily Globivalvulinoidea Reitlinger, 1950 moving the Biseriamminidae to the superfamily Mstinioidea Lipina, 1989. Cózar and Sommerville (2012) challenged this systematic interpretation and included two families into the superfamily Biseriamminoidea: Biseriamminidae (microgranular wall with agglutinated grains) and Globivalvulinidae (wall microgranular or differentiated with diaphanoteca, sensu Cózar and Sommerville 2012). Finally, Vachard (2016a) revised the Globivalvulinoidea as composed of three families: Globivalvulinidae, ?Biseriamminidae, and ?Koktjubinidae.
Herein, we focus on the family Globivalvulinidae with the aim to review its taxonomic composition and to better define its systematics. The phylogenetic relationships of its subfamilies are tentatively proposed by means of a cladistic approach, which is not commonly applied to Palaeozoic foraminifers.
Historical background
Gaillot and Vachard (2007) raised the subfamily Globivalvulininae Reitlinger, 1950 to the family rank (Globivalvulinidae) defining the family as composed of Globivalvulininae, Dagmaritinae, Paradagmaritinae Gaillot and Vachard, 2007, and Paraglobivalvulininae Gaillot and Vachard, 2007. They introduced new genera belonging to three of the above cited four subfamilies: Labioglobivalvulina Gaillot and Vachard, 2007 (= Siphoglobivalvulina Gaillot, 2006) and Retroseptellina Gaillot and Vachard, 2007 to the Globivalvulininae; Labiodagmarita Gaillot and Vachard, 2007, Bidagmarita Gaillot and Vachard in Gaillot et al. (2009), and Siphodagmarita Gaillot and Vachard in Vachard et al. (2006) to the Dagmaritinae; and Paremiratella Gaillot and Vachard, 2007, Paradagmaritella Gaillot and Vachard, 2007, Paradagmacrusta Gaillot and Vachard, 2007, and Paradagmaritopsis Gaillot and Vachard in Gaillot et al. (2009) to the Paradagmaritinae. Partially in agreement with this latter interpretation and with some criticisms about the generic composition of subfamilies, Altıner and Özkan-Altıner (2010) reviewed the evolutionary relationships between the Paradagmaritinae and Dagmaritinae, indicating the former as a junior synonym of the Dagmaritinae. Cózar and Sommerville (2012) considered the superfamily Biseriamminoidea as composed of the family Globivalvulinidae and the family Biseriamminidae, considering the Koktjubinidae as a synonym of the Biseriamminidae. The latter authors synonymised the genera Dzhamansorina Marfenkova, 1991, Admiranda Marfenkova, 1991, and Ulanbela Marfenkova, 1991 with Koktjubina Marfenkova, 1991 on the basis of dubious differences between them. The latest taxonomic rearrangement by Vachard (2016b) maintained the subfamily subdivisions as proposed in Gaillot and Vachard (2007).
The generic revision herein proposed led to suggest a revised taxonomic scheme of the family Globivalvulinidae and its subfamilies (Table 1). In this taxonomic scheme, the superfamily Biseriamminoidea is composed of two families: the Biseriamminidae sensu Cózar and Sommerville (2012) (Biseriammininae) and the Globivalvulinidae (Globivalvulininae, Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae) on the basis of wall structure as stated by Cózar and Sommerville (2012) for Mississippian Biseriamminoidea.
Table 1. New taxonomic arrangement of the family Globivalvulinidae proposed in this paper.
|
Superfamily |
Families |
Subfamilies |
Genera |
|
Biseriamminoidea |
Globivalvulinidae
|
Globivalvulininae |
Globivalvulina, Charliella, Retroseptellina, Labioglobivalvulina, Biseriella, Tenebrosella, Verispira, Paremiratella |
|
Paraglobivalvulininae |
Paraglobivalvulina,
Paraglobivalvulinoides, Urushtenella,
|
||
|
Dagmaritinae |
Dagmarita, Danielita, Louisettita |
||
|
Paradagmaritinae |
Paradagmarita,
Paradagmacrusta, Paradagmaritopsis,
|
||
|
Biseriamminidae (sensu Cózar and Sommerville 2012) |
Biseriammininae |
Biseriammina, Koktjubina, Parabiseriella |
The generic composition of the subfamily Globivalvulininae is modified by moving the genus Septoglobivalvulina Lin, 1978 to the subfamily Paraglobivalvulininae because its last chamber envelops the preceding ones. We also retain Paremiratella as valid, although morphologically it is close to Globivalvulina, because the differences in size of the tests (larger in Paremiratella) and number of chambers (more numerous in Paremiratella) are considered of specific taxonomic value. Furthermore, according to Altıner and Özkan-Altıner (2010), the genus Paremiratella (Paradagmaritinae in Gaillot and Vachard 2007) belongs to the Globivalvulininae because of its globular chambers and dark brown single-layered wall. Additionally, the genus Lateenoglobivalvulina introduced by Filimonova (2016) and placed into the Globivalvulininae, is not discussed herein because the holotype of the type species Globivalvulina spiralis Morozova, 1949 does not clearly show the generic diagnostic features (i.e., the type of wall and aperture) being also figured by Morozova (1949) as a drawing.
The genus Sengoerina Altıner, 1999 (Dagmaritinae in Altıner 1999), was assigned by Nestell and Nestell (2006) to globivalvulinids as later accepted by of Mikhalevich (2014). However, we follow Mohtat-Aghai and Vachard (2003) who evidenced the affinities between Sengoerina and Paradagmarita Lys in Lys and Marcoux, 1978, and include Sengoerina, as well as Crescentia Ciarapica, Cirilli, Martini, and Zaninetti, 1986, in the subfamily Paradagmaritinae. Both genera are characterised by a biserially enrolled initial stage and by chambers that become angular in the uncoiled stage. Based on the type of wall of the test and following Altıner and Özkan-Altıner (2010), we further reduce the generic composition of the Dagmaritinae, keeping outside of this subfamily the genera Bidagmarita and Labiodagmarita, which are probably closer to the family Palaeotextulariidae Galloway, 1933. Due to its coarse granular layer, the genus Paradagmaritella (Paradagmaritinae in Gaillot and Vachard 2007) is not referred to the Globivalvulinidae but in close proximity to the genus Spireitlina Vachard in Vachard and Beckary, 1991, as suggested by Altıner and Özkan-Altıner (2010).
Methods
The phylogenetic relationships within the Globivalvulinidae are herein investigated through a cladistic approach, by means of the analytical technique of maximum parsimony (Kolaczkowski and Thornton 2004; Kannan and Wheeler 2012).
The use of cladistics for Foraminifera is not exempt from some restrictions and problems (Soldan et al. 2011). Some of these obstacles are related to intrinsic characteristics of each taxon. For instance, the relatively simple morphology of the earliest lagenids results in the difficulties to define a significant number of characters to analyse (Groves et al. 2003). In addition, morphological characters are not always recognisable on fossil samples, due to the destructive effects of diagenesis that often affect Palaeozoic sedimentary rocks. Worthy to note are also the limitations related to the two-dimensional microscopic analysis, commonly performed on Foraminifera of this time interval. This implies that some morphological features could not be easily identified, because observable only in 3D view (e.g., the ornamentations). Anyway, we believe that the phylogenetic relationships among Palaeozoic groups of foraminifers can be fruitfully determined, both at the genus and suprageneric level, starting from a correct recognition and definition of characters and character states.
In this study, the cladistic analysis is based on the taxonomic scheme showed in Table 1. The phylogenetic tree has been obtained by means of fifteen qualitative (descriptive) characters and one quantitative (numerical) character. Characters have been selected using original generic descriptions and diagnoses (Reitlinger 1965; Zaninetti and Jenny-Deshusses 1985; Ciarapica et al. 1986; Villa and Sanchez de Posada 1986; Loeblich and Tappan 1987; Palmieri 1988; Altıner 1999; Altıner and Özkan-Altıner 2001; Pronina-Nestell and Nestell 2001; Nestell and Nestell 2006; Gaillot and Vachard 2007; Gaillot et al. 2009; Altıner and Özkan-Altıner 2010; Cózar and Sommerville 2012), without referring to any previous phylogenetic works. The descriptive characters are illustrated in Figs. 1 and 2, whereas the complete list of characters is provided in Appendix 1. The family Biseriamminidae has been designated as outgroup because it is closely related to the Globivalvulinidae due to the type of coiling, the arrangement and the shape of chambers, and the similarity of the aperture (Vachard et al. 2006; Cózar and Sommerville 2012). The ingroup is composed by the four Globivalvulinidae subfamilies (Globivalvulininae, Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae), since one of the aims of the work is to reconstruct the phylogenetic relationships within the family and to evaluate the systematic validity of its subfamilies.
A matrix of five taxa and sixteen characters has been constructed (Appendix 1). All characters have been treated as unordered and unweighted. The data matrix has been processed with the software PAUP* version 4.01 (Swofford 2002). Nodal support has been calculated performing 1000 bootstrap analyses (Felsenstein 1985) using branch and bound searches.

Fig. 1. Graphic explanation of descriptive morphological characters on which the cladistic analysis is based. A. Charliella rossae Altıner and Özkan-Altıner, 2001 (Altıner and Özkan-Altıner 2001: pl. 1: 2). B. Danielita gailloti Altıner and Özkan-Altıner, 2010 (Altıner and Özkan-Altıner 2010: pl. 2: 20). C. Globivalvulina sp. 1 (Nestell and Nestell 2006: pl. 6: 1). D. Paradagmarita flabelliformis Zaninetti, Altıner, and Catal, 1981 (Altıner 1981: pl. 39: 1). E. Tenebrosella asturica Villa and Sanchez de Posada, 1986 (Villa and Sanchez de Posada 1986: holotype, pl. 1: 1). F. Verispira jelli Palmieri, 1988 in spiral (F1) and umbilical (F2) views (modified from Palmieri 1988: fig. 8). G. Dagmarita sp. (Nestell and Pronina 1997: pl. 1: 6). H. Labioglobivalvulina baudi Gaillot and Vachard, 2007 (Gaillot and Vachard 2007: holotype, pl. 12: 10). I. Louisettita elegantissima Altıner and Brönnimann, 1980 (Loeblich and Tappan 1987: holotype, pl. 231: 7). J. Paraglobivalvulinoides septulifer Zaninetti and Altıner, 1981 (Zaninetti and Altıner 1981: holotype, pl. 1: 15). K. Paraglobivalvulinoides septulifer Zaninetti and Altıner, 1981 (Zaninetti and Altıner 1981: pl. 1: 19). Area in gray in C and D indicates the second stage of coiling. Not to scale.

Fig. 2. Graphic explanation of descriptive morphological characters on which the cladistic analysis is based. A. Danielita gailloti Altıner and Özkan-Altıner, 2010 (Altıner and Özkan-Altıner 2010: pl. 1: 3). B. Urushtenella latebrosa Pronina-Nestell, 2001 (Pronina-Nestell and Nestell 2001: pl. 5: 12). C. Koktjubina windsorensis (Mamet, 1970) (Mamet 1970: holotype, pl. 1: 11). D. Retroseptellina decrouezae (Köylüoglu and Altıner, 1989) (modified from Altıner and Özkan-Altıner 2001: fig. 2). E. Charliella rossae Altıner and Özkan-Altıner, 2001 (modified from Altıner and Özkan-Altıner 2001: fig. 2). F. Urushtenella latebrosa Pronina-Nestell, 2001 (Pronina-Nestell and Nestell 2001: pl. 5: 12). G. Globivalvulina graeca Reichel, 1946 (modified from Altıner and Özkan-Altıner 2001: fig. 2). H. Paradagmarita monodi Lys in Lys and Marcoux, 1978 (Loeblich and Tappan 1987: holotype, pl. 230: 6). I. Globivalvulina bulloides (Brady, 1876) (Loeblich and Tappan 1987: pl. 230: 14). J. Paraglobivalvulina mira Reitlinger, 1965 (Loeblich and Tappan 1987: holotype, pl. 230: 10). K. Septoglobivalvulina distensa (Wang in Zhao et al., 1981) (Gaillot and Vachard 2007: pl. 37: 2). L. Paradagmaritopsis kobayashii Gaillot and Vachard, 2009 (Gaillot et al. 2009: holotype, fig. 6: 6). Not to scale.
Systematic palaeontology
Phylum Foraminifera d’Orbigny, 1826
Class Fusulinata Gaillot and Vachard, 2007
Subclass Fusulinana Maslakova, 1990 nom. correct. Vachard et al. (2010)
Order Endothyrida Fursenko, 1958
Superfamily Biseriamminoidea Chernysheva, 1941
Families included: Biseriamminidae Chernysheva, 1941; Globivalvulinidae Reitlinger, 1950.
Description.—Test biserial, globular, subglobular or elongated in shape, evolute/semi-evolute or involute/semi-involute, rounded or pinched peripheral outline, from coiled, coiled to uncoiled, to uncoiled. Endoskeletal partitions and supplementary deposits can be present. Test wall microgranular, with agglutinated grains, undifferentiated or poorly layered or microgranular, single layered or plurilayered. Presence of hyaline layer in inner, median or outer position or pseudoalveolar median layer. Perforations of the wall can be present. Aperture narrow, simple or with valvular projection.
Stratigraphic and geographic range.—Late Tournaisian (Mississippian, Carboniferous) to latest Changhsingian (Lopingian, Permian); cosmopolite.
Family Biseriamminidae Chernysheva, 1941
Genera included: Biseriammina Chernysheva, 1941; Koktjubina Marfenkova, 1991; Parabiseriella Cózar and Sommerville, 2012.
Description.—Test biserial, globular, subglobular or elongated in shape, evolute, rounded peripheral outline, low trochospirally or planispirally coiled, with the tendency to uncoil in the terminal part. Test wall microgranular, with “adventitious grains, undifferentiated or poorly layered” (Cózar and Sommerville 2012: 217). Aperture simple, narrow, rarely depressed with a valvular projection (in Parabiseriella).
Stratigraphic and geographic range.—Late Tournaisian (Mississippian, Carboniferous) to ?early Bashkirian (Pennsylvanian, Carboniferous) of the Palaeotethys and Panthalassa (Urals and North America).
Family Globivalvulinidae Reitlinger, 1950
Subfamilies included: Globivalvulininae Reitlinger, 1950; Paraglobivalvulininae Gaillot and Vachard, 2007; Dagmaritinae Bozorgnia, 1973; Paradagmaritinae Gaillot and Vachard, 2007.
Description.—Test biserial, globular, subglobular or elongated in shape, evolute/semi-evolute or involute/semi-involute, rounded or pinched peripheral outline, coiled, coiled to uncoiled, uncoiled. Endoskeletal partitions and supplementary deposits can be present. Test wall microgranular, single layered or plurilayered. Presence of hyaline layer in inner, median or outer position or pseudoalveolar median layer. Perforations of the wall can be present. Aperture interiomarginal depressed and protected by a valvular projection.
Stratigraphic and geographic range.—Late Visean (Mississippian, Carboniferous) to latest Changhsingian (Lopingian, Permian); cosmopolite.
Subfamily Globivalvulininae Reitlinger, 1950
Genera included: Globivalvulina Schubert, 1921; Charliella Altıner and Özkan-Altıner, 2001; Labioglobivalvulina Gaillot and Vachard, 2007; Retroseptellina Gaillot and Vachard, 2007; Tenebrosella Villa and Sanchez de Posada, 1986; Verispira Palmieri, 1988; Biseriella Mamet in Armstrong and Mamet, 1974; Paremiratella Gaillot and Vachard, 2007.
Description.—Test free, globular or subglobular in shape, evolute or semi-evolute, rounded peripheral outline, biserial, coiling planispiral to slightly trochospiral, or trochospiral (helicoidal? in Biseriella). Chambers globular, subglobular to angular. Septal supplementary nodular formations (in Labioglobivalvulina). Test wall dark microgranular, single layered or plurilayered with white inner hyaline layer, sutural fine perforations in Verispira. Aperture interiomarginal depressed and protected by a valvular projection.
Stratigraphic and geographic range.—Late Visean (Mississippian, Carboniferous) to latest Changhsingian (Lopingian, Permian); cosmopolite.
Subfamily Paraglobivalvulininae Gaillot and Vachard, 2007
Genera included: Paraglobivalvulina Reitlinger, 1965; Paraglobivalvulinoides Zaninetti and Jenny-Deshusses, 1985; Urushtenella Pronina-Nestell in Pronina-Nestell and Nestell 2001; Septoglobivalvulina Lin, 1978.
Description.—Test free, globular or subglobular in shape, semi-involute or involute, rounded peripheral outline, biserial, planispirally to trochospirally coiled. Chambers globular to subglobular. Auxiliary interseptal and apertural chamberlets can be present. Test wall dark, microgranular, single layered or plurilayered with pseudoalveolar structure (in Urushtenella). Aperture interiomarginal depressed and protected by a valvular projection.
Stratigraphic and geographic range.—Capitanian (Guadalupian, Permian) to late Changhsingian (Lopingian, Permian) of the Palaeotethys and Neotethys.
Subfamily Dagmaritinae Bozorgnia, 1973
Genera included: Dagmarita Reitlinger, 1965; Louisettita Altıner and Brönnimann, 1980; Danielita Altıner and Özkan-Altıner, 2010.
Description.—Test free, elongated in shape, evolute, rounded peripheral outline, biserial, uncoiled. Subspheric chambers, semi-circular to semi-ellipsoidal in axial section. Presence of outer thornlike projections of the test wall. Endoskeletal septal partitions (peripheral chamberlets) are present in Louisettita. Test wall plurilayered composed of dark microgranular and white median or outer hyaline layer. The microgranular layer described with perforations (in Danielita). Aperture interiomarginal depressed and protected by a valvular projection.
Stratigraphic and geographic range.—Roadian (Zheng 1986) (Guadalupian, Permian) to late Changhsingian (Lopingian, Permian) of the Palaeotethys, the Neotethys, and the Panthalassa (Japan and North America).
The first appearance datum (FAD) of the Dagmaritinae is based on the FAD of the genus Dagmarita. Zheng (1986) recorded the first appearance of this taxon in the sample D40 of the Chihsia (= Qixia) Formation (South China), which has been referred to the Verbeekina grabaui Zone. This biozone corresponds to the Roadian Neoschwagerina simplex Zone (Nestell and Nestell 2006; Angiolini et al. 2015).
Subfamily Paradagmaritinae Gaillot and Vachard, 2007
Genera included: Paradagmarita Lys in Lys and Marcoux, 1978; Paradagmacrusta Gaillot and Vachard, 2007; Paradagmaritopsis Gaillot and Vachard in Gaillot et al., 2009; Crescentia Ciarapica, Cirilli, Martini, and Zaninetti, 1986; Sengoerina Altıner, 1999.
Description.—Test free, elongated in shape, pinched peripheral outline, biserial, involute initial stage, generally slightly trochospiral and later followed by an uncoiled biserial stage. Chambers are globular at the beginning and later angular or inflated. Crustae at the roof of the chambers are present in Paradagmacrusta. Test wall dark, microgranular or plurilayered with white outer hyaline layer. Aperture interiomarginal depressed and protected by a valvular projection.
Stratigraphic and geographic range.—Capitanian (Guadalupian, Permian) to late Changhsingian (Lopingian, Permian) of the Palaeotethys, the Neotethys, and the Panthalassa (Japan and North America).
Results
The phylogenetic analysis produced a single tree of 19 steps (Fig. 3), with a Consistency Index of 0.842 and a Retention Index of 0.571.
The monophyletic clade of Globivalvulinidae has the Globivalvulininae as the first taxon to have branched off, characterised by one autapomorphy: presence of supplementary deposits (nodular structures). Then, a monophyletic clade composed of the Paraglobivalvulininae and Dagmaritinae plus Paradagmaritinae, is recognised. This clade is supported by one synapomorphy: absence of a completely trochospiral coiling mode (although this character is inapplicable for Dagmaritinae; see Appendix 1). The sister-group relationship between Dagmaritinae and Paradagmaritinae is evidenced by three synapomorphies: uncoiled test in the second stage, predominance of plurilayered genera, and elongated shape of the test (Fig. 3; Appendix 1).
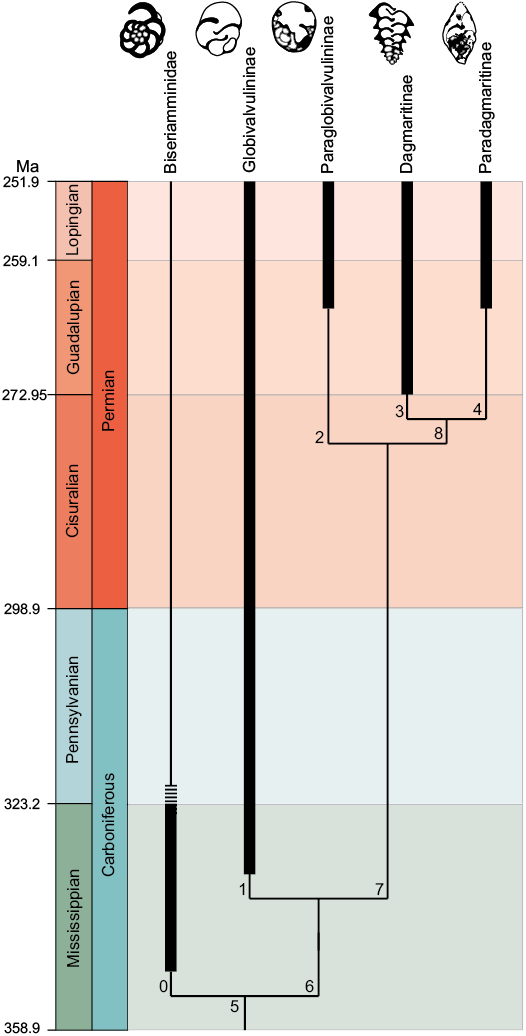
Fig. 3. Phylogenetic tree resulting from the cladistic analysis. Thick lines represent the known stratigraphic ranges of the considered taxa (dashed line indicates uncertain stratigraphic ranges). Nodes are numbered above branches. See text and Appendix 1 for details.
Concluding remarks
The phylogenetic relationships within the Globivalvulinidae have been reconstructed in order to investigate the evolutionary history of this group of Foraminifera. The analysis has to be considered as preliminary because it should be expanded in the future with the inclusion of additional characters, and a larger taxonomic sampling.
The obtained phylogenetic tree (Fig. 3) supports the validity of the four globivalvulinid subfamilies as they all resulted as monophyletic groups.
Within the Globivalvulinidae, the Globivalvulininae are the first group to branch out. This result is in agreement with biostratigraphic data, according to which representatives of this subfamily appear in the fossil record starting from the Mississippian (Gaillot and Vachard 2007; Cózar and Sommerville 2012; Vachard 2016b). However, the alleged relationship between the Globivalvulininae and Paraglobivalvulininae (Altıner 1999; Altıner and Özkan-Altıner 2001; Vachard et al. 2006) is not supported by our results, since the latter subfamily is phylogenetically closer to the Dagmaritinae and Paradagmaritinae than to the Globivalvulininae (i.e., a hypothetical group including the Globivalvulininae and Paraglobivalvulininae would be paraphyletic). Conversely, we firmly confirm the close affinity between the Dagmaritinae and Paradagmaritinae as defined by Altıner (1997, 1999) and Altıner and Özkan-Altıner (2010), in the light of the sister-group relationship between the two subfamilies in our tree, supported by three synapomorphies (Fig. 3). We are aware that the separation between Globivalvulininae and the other three subfamilies (node 7) is relatively weak, being supported by a single synapomorphy (see Appendix 1), and that this weakness could implicitly strengthen the previous morphology-based phylogenetic views (Altıner 1999; Altıner and Özkan-Altıner 2001; Vachard et al. 2006). On the other hand, phylogenetic relationships between Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae are well supported in our tree (see Appendix 1), thus questioning the “traditional” phylogenetic proximity between Globivalvulininae and Paraglobivalvulininae. We hope that further phylogenetic investigations on these groups can provide significant contributions to support our reconstruction.
Looking at the stratigraphic distribution of the analysed taxa (Fig. 3), it is worth noting that the range of the Dagmaritinae is slightly longer than expected on the basis of their phylogenetic position in the tree. In fact, a shorter stratigraphic range (i.e., coinciding at least with that of their sister taxon, the Paradagmaritinae) would be suggested by the tree topology. Pending on new significant palaeontological discoveries involving these groups (i.e., Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae) and/or new phylogenetic analyses of the Globivalvulinidae and related groups, this may be explained, hypothetically, by the fact that the Paraglobivalvulininae and Paradagmaritinae have ghost ranges, i.e., they appeared earlier than as known in literature (at least in the Roadian), so that their available fossil record is still incomplete. Furthermore, worthy to note is that the first appearance datum of Dagmaritinae is based on a single report of Dagmarita sp. recorded in the upper part of the Chihsia Formation (Zheng 1986), whose reliability has already been questioned by Vachard (2016b).
Despite the relatively low number of considered taxa, our phylogenetic analysis has proven to convincingly solve the phylogenetic relationships between them (i.e., Globivalvulininae, Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae). By providing the first reconstruction of the evolutionary history of the Globivalvulinidae through a cladistic approach, this paper also represents a methodological contribution, which could serve as a starting point for similar studies on other groups of fossil Foraminifera.
Acknowledgements
Thanks are due to Marco Crotti (University of Glasgow, UK) and Giorgio Carnevale (University of Turin, Italy) for the useful suggestions about the phylogenetic analysis. Thanks to the reviewers Rossana Martini (University of Geneva, Switzerland), Katsumi Ueno (University of Fukuoka, Japan), and an anonymous reviewer for their constructive criticisms and useful comments that greatly improved the manuscript.
References
Altıner, D. 1981. Recherches stratigraphiques et micropaléontologiques dans le Taurus Oriental au NW de Pinarbasi (Turquie). 450 pp. Unpublished Ph.D. Thesis, Université de Genève, Genève.
Altıner, D. 1997. Origin, morphologic variation and evolution of dagmaritin-type biseriamminid stock in the Late Permian. In: C.A. Ross, J.R.P. Ross, and P.L. Brenckle (eds.), Late Paleozoic Foraminifera; Their Biostratigraphy, Evolution and Paleoecology; and the Mid-Carboniferous Boundary, Special Publication 36, 1–4. Cushman Foundation for Foraminiferal Research, Washington.
Altıner, D. 1999. Sengoerina argandi, n. gen., n. sp., and its position in the evolution of Late Permian biseriamminid foraminifers. Micropaleontology 45: 215–220. Crossref
Altıner, D. and Özkan-Altıner, S. 2001. Charliella rossae n. gen., n. sp. from the Tethyan Realm: remarks on the evolution of Late Permian biseriamminids. Journal of Foraminiferal Research 31: 309–314. Crossref
Altıner, D. and Özkan-Altıner, S. 2010. Danielita gailloti n. gen., n. sp., within the evolutionary framework of Middle–Late Permian Dagmaritins. Turkish Journal of Earth Sciences 19: 497–512.
Angiolini, L., Zanchi, A., Zanchetta, S., Nicora, A., Vuolo, I., Berra, F., Henderson, C., Malaspina, N., Rettori, R., Vachard, D., and Vezzoli, G. 2015. From rift to drift in South Pamir (Tajikistan): Permian evolution of a Cimmerian terrane. Journal of Asian Earth Sciences 102: 146–169. Crossref
Chernysheva, N. [Černyševa, N.] 1941. New genus of Foraminifera from the Tournaisian deposits of the Urals [in Russian]. Doklady Akademii Nauk SSSR 31: 69–70.
Ciarapica, G., Cirilli, S., Martini, R., and Zaninetti, L. 1986. Une microfaune à petits foraminifères d’âge permien remaniée dans le Trias moyen de l’Apennin méridional (Formation de Monte Facito, Lucanie occidentale): description de Crescentia vertebralis, n. gen., n. sp. Revue de Paléobiologie 5: 207–215.
Cózar, P. and Somerville, J.D. 2012. Mississippian Biseriamminacea and their evolutionary development. Journal of Foraminiferal Research 42: 216–233. Crossref
Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783–791. Crossref
Filimonova, T.V. 2016. A new Pennsylvanian (Late Carboniferous)–Permian foraminiferal genus (Lateenoglobivalvulina nov. gen., Biseriamminoidea) and its paleobiogeographic distribution. Revue de Micropaléontologie 59: 188–199. Crossref
Gaillot, J. and Vachard, D. 2007. The Khuff Formation (Middle East) and time equivalents in Turkey and South China: biostratigraphy from Capitanian to Changhsingian times (Permian), new foraminiferal taxa, and palaeogeographical implications. Coloquios de Paleontología 57: 37−223.
Gaillot, J., Vachard, D., Galfetti, T., and Martini, R. 2009. New latest Permian foraminifers from Laren (Guangxi Province, South China): palaeobiogeographic implications. Geobios 42: 141–168. Crossref
Groves, J.R., Altıner, D., and Rettori, R. 2003. Origin and early evolutionary radiation of the order Lagenida (Foraminifera). Journal of Paleontology 77: 831–843. Crossref
Hance, L., Hou, H., and Vachard, D. (with the collaboration of Devuyst, F.X., Kalvoda, J., Poty, E., and Wu, X.) 2011. Upper Famennian to Visean Foraminifers and Some Carbonate Microproblematica from South China-Hunan, Guangxi and Guizhou. 359 pp. Geological Publishing House, Beijing.
Kannan, L. and Wheeler, W.C. 2012. Maximum parsimony on phylogenetic networks. Algorithms for Molecular Biology 7: 9. Crossref
Kolaczkowski, B. and Thornton, J.W. 2004. Performance of maximum parsimony and likelihood phylogenetics when evolution is heterogeneous. Nature 431: 980–984. Crossref
Loeblich, A.R., Jr. and Tappan, H. 1964. Sarcodina, chiefly ‘‘Thecamoebians’’ and Foraminiferida. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part C, Protista 2, 1–900. Geological Society of America, New York and Kansas University Press, Lawrence.
Loeblich, A.R., Jr. and Tappan, H. 1984. Suprageneric classification of the Foraminiferida (Protozoa). Micropaleontology 30: 1–70. Crossref
Loeblich, A.R., Jr. and Tappan, H. 1987. Foraminiferal Genera and Their Classification. 970 pp. Van Nostrand Rheinhold Company, New York.
Lys, M. and Marcoux, J. 1978. Les niveaux du Permien supérieur des Nappes d’Antalaya (Taurides occidentales, Turquie). Comptes Rendus Académie Sciences 286 (série D): 1417–1420.
Mamet, B.L. 1970. Carbonate Microfacies of the Windsor Group (Carboniferous) of Nova Scotia and New Brunswick. Geological Survey of Canada 21: 1–121.
Marfenkova, M.M. 1991. Marine Carboniferous of Kazakhstan {provide original tilte in transliteration}. 176 pp. Akademiâ Nauk Kazahskoj SSR, Institut Geologičeskih Nauk, “Gylym”, Alma-Ata.
Mikhalevich, V.I. 2014. Post Cambrian Testate Foraminifera as a System in Its Evolution. 444 pp. Nova Science Publishers, Inc., New York.
Mohtat-Aghai, P. and Vachard, D. 2003. Dagmarita shahrezaensis n. sp. Globivalulinid foraminifer (Wuchiapingian, Late Permian, Central Iran). Rivista Italiana di Paleontologia e Stratigrafia 109: 37–44.
Morozova, V.G. 1949. Members of families Lituolidae and Textulariidae from late Carboniferous and Artinskian from the Bashkir Pre-Urals [in Russian]. In: N.S. Šatskij and D.M. Rauzer-Černousova (eds.), Foraminifery verh kamennougol’nyh i artinskih otloženij Baškirskogo Priural’â. Trudy Instituta Geologičeskih Nauk 105 (35): 244–275.
Nestell, G.P. and Nestell, M.K. 2006. Middle Permian (Late Guadalupian) foraminifers from Dark Canyon, Guadalupe Mountains, New Mexico. Micropaleontology 52: 1–50. Crossref
Nestell, M.K. and Pronina, G.P. 1997. The distribution and age of the genus Hemigordiopsis. In: C.A. Ross, J.R.P. Ross, and P.L. Brenckle (eds.), Late Paleozoic Foraminifera; Their Biostratigraphy, Evolution and Paleoecology; and the Mid-Carboniferous Boundary, Special Publication 36, 105−110. Cushman Foundation for Foraminiferal Research, Washington.
Palmieri, V. 1988. Globivalvulinid Foraminifera from the Permian of Queensland. Alcheringa 12: 27–47. Crossref
Perret, M.F. 1993. Recherches micropaléontologiques et biostratigraphiques (conodontes–foraminifères) dans le Carbonifère pyrénéen. Strata, série 2, Mémoires 21: 1–597.
Pinard, S. and Mamet, B. 1998. Taxonomie des petits foraminifères du Carbonifère supérieur – Permien inférieur du bassin de Sverdrup, Arctique Canadien. Paleontographica Canadiana 15: 1–253.
Pronina-Nestell, G.P. and Nestell, M.K. 2001. Late Changhsingian foraminifers of the Northwestern Caucasus. Micropaleontology 47: 205–234. Crossref
Reitlinger, E.A. 1965. On the development of the foraminifera of the Late Permian and Early Triassic in Transcaucasia [in Russian]. Voprosy Mikropaleontologii 9: 45–70.
Schubert, R.J. 1921. Paleontological data about the phylogenetic history of Protozoa. Paläontologische Zeitschrift 3: 129–188. Crossref
Soldan, D.M., Petrizzo, M.R., Premoli Silva, I., and Cau, A. 2011. Phylogenetic relationships and evolutionary history of the Paleogene genus Igorina through parsimony analysis. Journal of Foraminiferal Research 41: 260–284. Crossref
Swofford, D.L. 2002. PAUP*: Phylogenetic Analysis Using Parsimony (* and other methods), Version 4.0 Beta 10. Sinauer Associates, Sunderland.
Vachard, D. 2016a. Macroevolution and biostratigraphy of Paleozoic foraminifers. In: M. Montenari (ed.), Stratigraphy and Timescales, 257–323. Elsevier, Amsterdam. Crossref
Vachard, D. 2016b. Permian Smaller Foraminifers: Taxonomy, Biostratigraphy and Biogeography. 450 pp. Geological Society of London, Special Publications, London.
Vachard, D. and Beckary, S. 1991. Algues et Foraminifères bachkiriens des coal balls de la Mine Rosario (Truebano, Léon, Espagne). Revue de Paléobiologie 10: 315–357.
Vachard, D., Gaillot, J., Pille, L., and Blażejowski, B. 2006. Problems on Biseriamminoidea, Mississippian–Permian biserially coiled foraminifera. A reappraisal with proposals. Revista Española Micropaleontología 38: 453–492.
Vachard, D., Pille, L., and Gaillot, J. 2010. Palaeozoic Foraminifera: Systematics, palaeoecology and responses to global changes. Revue de Micropaléontologie 53: 209–254. Crossref
Vachard, D., Rettori, R., Altıner, D., and Gennari, V. 2017. The Permian foraminiferal family Pseudovidalinidae and the genus Altineria emend. herein. Journal of Foraminiferal Research 47: 279–283. Crossref
Villa, E. and Sanchez de Posada, L.C. 1986. Tenebrosella (Foraminiferida, Biseriamminidae), nuevo género del Carbonífero cantábrico. Revista Española de Paleontología 1: 85–92.
Zaninetti, L. and Altıner, D. 1981. Les Biseriamminidae (Foraminifères) dans le Permien supérieur mésogéen: évolution et biostratigraphie. Notes du Laboratoire de Paléontologie de l’Université de Genève 7: 39–46.
Zaninetti, L. and Jenny-Deshusses, C. 1985. Les Paraglobivalvulines (foraminifères) dans le Permien supérieur téthysien; répartition géographique et description de Paraglobivalvulinoides n. gen. Revue de Paléobiologie 4: 343–346.
Zheng, H. 1986. The smaller foraminifer faunas in Qixia stage (Early Permian) of Daxiakou, Xingshan County, Hubei Province. Earth Science-Journal of Wuhan College of Geology 11: 489−497.
Supplementary data regarding the phylogenetic analysis.
Character list
Unordered and unweighted states for all characters.
0. Coiling mode: uncoiled (0); coiled (1).
1. Coiling mode, second stage: uncoiled (0); coiled (1).
2. Coiling mode, trochospiral: present (0); absent (1).
Remarks: We assigned state 0 to Biseriamminidae, Globivalvulininae, Paraglobivalvulininae, and Paradagmaritinae which are at least composed by one genus having a test made by a trochospirally coiled stage; we assigned state 1 to subfamily Dagmaritinae, whose genera are never characterised by trochospirally coiled stage.
3. Coiling mode, completely trochospiral: present (0); absent (1).
Remarks: We coded this character as inapplicable (–) for Dagmaritinae because this group never shows a trochospirally coiled stage (see character 2).
4. Involute test: present (0); absent (1).
5. Wall microgranular with agglutinated grains: present (0); absent (1).
6. Wall, number of layers: one (0); more than one (1).
Remarks: The taxa showing state 0 include genera with predominantly single layered wall; the taxa showing state 1 include genera with predominantly plurilayered wall.
7. Wall, hyaline or pseudoalveolar layer: present (0); absent (1).
8. Pseudoalveoli or pores: present (0); absent (1).
9. Internal partitions, interseptal or peripheral chamberlets: present (0); absent (1).
10. Supplementary deposits: present (0); absent (1).
11. Thornlike projections: present (0); absent (1).
12. Shape of the test: globular/subglobular (0); elongated (1).
13. Peripheral outline: rounded (0); pinched (1).
14. Valvular projection: absent (0); present (1).
Remarks: We assigned state 0 to Biseriamminidae which are at least composed by one genus having a simple aperture without a valvular projection; we assigned state 1 to Globivalvulininae, Paraglobivalvulininae, Dagmaritinae, and Paradagmaritinae, whose genera are characterised by an aperture protected by a valvular projection.
15. Apertural chamberlets: present (0); absent (1).
Character-taxon matrix
| |
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
|
Biseriamminidae |
1 |
0/1 |
0 |
0 |
1 |
0 |
0 |
1 |
1 |
1 |
1 |
1 |
0/1 |
0 |
0 |
1 |
|
Globivalvulininae |
1 |
1 |
0 |
0 |
1 |
1 |
0 |
0 |
0 |
1 |
0 |
1 |
0 |
0 |
1 |
1 |
|
Paraglobivalvulininae |
1 |
1 |
0 |
1 |
0 |
1 |
0 |
0 |
0 |
0 |
1 |
1 |
0 |
0 |
1 |
0 |
|
Dagmaritinae |
0 |
0 |
1 |
– |
1 |
1 |
1 |
0 |
0 |
0 |
1 |
0 |
1 |
0 |
1 |
1 |
|
Paradagmaritinae |
1 |
0 |
0 |
1 |
1 |
1 |
1 |
0 |
1 |
1 |
0 |
1 |
1 |
1 |
1 |
1 |
Character state transformations at nodes on the most parsimonious tree
Node 1: Char. 10: 1 > 0.
Node 2: Char. 4: 1 > 0; Char. 15: 1 > 0.
Node 3: Char. 0: 1 > 0; Char. 2: 0 > 1; Char. 11: 1 > 0.
Node 4: Char. 8: 0 > 1; Char. 10: 1 > 0; Char. 13: 0 > 1.
Node 7: Char. 3: 0 > 1.
Node 8: Char. 1: 1 > 0; Char. 6: 0 > 1; Char. 12: 0 > 1.
Acta Palaeontol. Pol. 63 (4): 807–814, 2018
https://doi.org/10.4202/app.00511.2018